1 This is the post-print version (post-refereeing, i.e. the final version of the article that the authors have seen before it was published) of the following article: Linstädter, A., Kemmerling, B. (equally contributing), Baumann, G., Kirscht, H., 2013. The importance of being reliable – Local ecological knowledge and management of forage plants in a dryland pastoral system (Morocco), Journal of Arid Environments 95: 30-40, which has been published in final form at http://dx.doi.org/10.1016/j.jaridenv.2013.03.008 The importance of being reliable - valuation and management of forage plants in a dryland pastoral system Anja Linstädter a *, Birgit Kemmerling b , Gisela Baumann a,c , Holger Kirscht d,e a Range Ecology and Range Management Group Botanical Institute University of Cologne Zülpicher Straße 47b D-50674 Cologne, Germany b Institute of Oriental Studies University of Leipzig Schillerstraße 6 D-04109 Leipzig, Germany E-mail address: [email protected] c Present address: Botanic Garden and Botanical Museum Berlin-Dahlem Freie Universität Berlin Königin-Luise-Straße 6-8 D-14195 Berlin, Germany E-mail address: [email protected] d Institute of Social and Cultural Anthropology University of Cologne Albertus-Magnus-Platz D-50923 Cologne, Germany e Present address: International Institute of Tropical Agriculture Cameroon 1 Main Road IRAD Nkolbisson P.O. Box 2008 (Messa) Yaoundé, Cameroon E-mail address: [email protected] * Corresponding author Dr. Anja Linstädter Tel. +49-(0)221-470 7905 Fax +49-(0)221-470 7908 E-mail address: [email protected] A. Linstädter and B. Kemmerling contributed equally to this study.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
This is the post-print version (post-refereeing, i.e. the final version of the article that the
authors have seen before it was published) of the following article: Linstädter, A.,
Kemmerling, B. (equally contributing), Baumann, G., Kirscht, H., 2013. The importance of
being reliable – Local ecological knowledge and management of forage plants in a dryland
pastoral system (Morocco), Journal of Arid Environments 95: 30-40, which has been
published in final form at http://dx.doi.org/10.1016/j.jaridenv.2013.03.008
The importance of being reliable - valuation and management of forage
plants in a dryland pastoral system
Anja Linstädtera*, Birgit Kemmerling
b, Gisela Baumann
a,c, Holger Kirscht
d,e
a Range Ecology and Range Management Group
Botanical Institute
University of Cologne
Zülpicher Straße 47b
D-50674 Cologne, Germany
b
Institute of Oriental Studies
University of Leipzig
Schillerstraße 6
D-04109 Leipzig, Germany
E-mail address: [email protected]
c Present address:
Botanic Garden and Botanical Museum Berlin-Dahlem
Freie Universität Berlin
Königin-Luise-Straße 6-8
D-14195 Berlin, Germany
E-mail address: [email protected]
d
Institute of Social and Cultural Anthropology
University of Cologne
Albertus-Magnus-Platz
D-50923 Cologne, Germany
e Present address:
International Institute of Tropical Agriculture Cameroon
1 Main Road IRAD Nkolbisson
P.O. Box 2008 (Messa)
Yaoundé, Cameroon
E-mail address: [email protected]
* Corresponding author
Dr. Anja Linstädter
Tel. +49-(0)221-470 7905
Fax +49-(0)221-470 7908
E-mail address: [email protected]
A. Linstädter and B. Kemmerling contributed equally to this study.

2
Abstract
In dryland rangelands with their high environmental variability, local knowledge of forage
plants is essential for management decisions. Ecological apparency hypothesis (EAH) predicts
plants’ availability and visibility to be important criteria for local valuation. However, EAH
has mainly been tested in low-variability systems. We ask whether EAH is valid for forage
plants in drylands; which other local criteria exist; and how criteria are connected to
management decisions.
In a Moroccan pastoral system, we applied a novel ethnobotanical method by calculating the
Cognitive Salience Index (CSI) both for plants’ valuation (CSIantro) and availability (CSIeco).
To evaluate explicit criteria, we correlated palatability and nutritive value to CSIanthro.
ANCOVAs related CSIanthro to EAH criteria (CSIeco and lifetime) and to plant occurrence on
pasture types. We found EAH criteria to better predict CSIantro than explicit criteria. Apparent
plants from semi-arid pastures were more valued than those from arid pastures (HSD; p <
0.05). We introduce reliability as one criterion into EAH to explain this, and demonstrate how
pastoralists adjust management decisions to resource reliability. Linking resource valuation to
management decisions can thus improve our understanding of resilience mechanisms. Our
study also confirms the validity of EAH for forage species and dryland environments.
Keywords: Arid and semi-arid rangelands; Cognitive salience index; Ecological apparency
theory; Local ecological knowledge; Pastoralism

3
1. Introduction
In arid and semi-arid environments, a sustainable land management requires a profound local
ecological knowledge (LEK) of natural resources and their dynamics (McAllister et al., 2006).
For this reason, LEK and its integration in scientific knowledge has received increasing
attention in dryland science (e.g. Fernandez-Gimenez, 2000; Thomas et al., 2007; Whitfield
and Reed, 2012). In the case of pastoralism, LEK is particularly required on the spatio-
temporal variability of forage resources (McAllister et al., 2006; Thomas and Twyman, 2004).
Based on their herding experiences, cognitive abilities, and world views, pastoralists gain an
extensive body of LEK on forage resources. This includes the environmental conditions in
which a plant grows, its palatability depending on livestock species, its phenology and life
history, and its availability on local pastures (Fernandez-Gimenez, 2000; Thomas and
Twyman, 2004). All parameters may be relevant for the perception of a species’ value as
forage plant. However, quality judgements are often not explicit, but part of implicit or even
tacit knowledge (Fazey et al., 2006), which makes them empirically difficult to extract
(Wesuls and Lang, 2010). To explain the valuation and use of plant resources in local
communities, the ecological apparency hypothesis (EAH) has been among the most promising
approaches (Lucena et al., 2007; Lucena et al., 2012). It was initially proposed for herbivore
behaviour and has been later adapted to ethnobotanical research questions (Phillips and
Gentry, 1993a, b). The hypothesis claims that apparent plants i.e., the most visible, most
dominant, and most frequent plants will have a higher cultural importance than less apparent
plants, not because they are necessarily more useful, but simply because they are more
available or visible to human communities (Lucena et al., 2007). However, EAH has mainly
been tested in tropical systems with a comparatively low spatial and temporal variability, such
as humid forests (Albuquerque and Lucena, 2005; Phillips and Gentry, 1993a, b), and dry
forests (Lucena et al., 2007; Lucena et al., 2012). It thus remains unclear which implications
environmental variability has for the validity of EAH.
The perception of natural resources and their dynamics is the basis for pastoral management
decisions (Fernandez-Gimenez, 2000; Steele and Shackleton, 2010). In a given situation, a
particular action is chosen from a set of strategies. Herd mobility is the most important
pastoral strategy, as it enables herders to cope with effects of rainfall variability on the natural
resource base (Butt, 2010; Dwyer and Istomin, 2008). Pastoral-nomadic range management
may have negative effects on the sustainability of natural resources (Bollig and Schulte, 1999)
but it often has positive effects (Müller et al., 2007b) and increases the resilience of the
pastoral system to external shocks such as meteorological droughts (Dougill et al., 2010;
Linstädter et al., 2010). In this paper we focus on a dryland pastoral system along a steep
gradient of environmental variability to test the validity of EAH for forage plants in highly
variable environments. We apply a novel, quantitative methodology to analyse the local
valuation of forage plants and to ask whether it is based on explicit local knowledge or
ecological criteria. Explicit LEK is analysed as the ‘palatability for goats’, ‘nutritive value for
goats’, ‘palatability for sheep’ and ‘nutritive value for sheep’. As ecological criteria related to
EAH we test a plant’s availability on local pastures, its life form and its lifetime. We then
relate management decisions to the local valuation of pasture quality, i.e. to the availability
and quality of forage resources on pasture types. Besides mobility decisions, we evaluate
alternative strategies of herd management which may be connected to the local valuation of
forage resources. Our key questions are:
- Does EAH explain the local valuation of forage plants in a highly variable environment?
- Do pastoralists have other explicit or non-explicit criteria for the valuation of forage plants?
- How are these criteria connected to herd management decisions?

4
2. Methods
2.1. Regional context and study area
Data were collected on Morocco’s southern slopes of the High Atlas Mountains among the
pastoral-nomadic group of the Ait Toumert. The Ait Toumert who settle in the Mgoun district
speak Tachelhit, one of the three Berber languages of Morocco. So far, little research has been
conducted in that area and no recent official census data were available on household level.
Through interviews with local leaders, 29 households of the Ait Toumert have been identified,
some of them sedentary and not applying transhumance anymore, others sedentary, but
sending their herds with other herdsmen, while some are still pastoral-nomadic. We restricted
our sample to pastoral-nomadic households. These use sheep and goats for livestock
production, while donkeys, mules and dromedaries are used for transportation (Akasbi et al.,
2012). The normative transhumance cycle (see Linstädter et al., 2010) runs along a steep
altitudinal gradient, from the mountainous summer pastures at 3,000 m asl (used from mid-
May to September) to the lowland pastures at 1,300 m asl used during the winter months from
November to February. In October and again from March to mid-May nomads pass the
intermediate pastures in between (Fig. 1).
Fig. 1. Grazing area of the Ait Toumert (High Atlas, Morocco), with four pasture types arranged along
a gradient of increasing aridity (from the summer pasture in the northern highlands to the far winter
pasture in the Basin of Ouarzazate). Pasture types were identified by means of expert interviews and
mental maps. Study sites are Trab Labied (TRB) for far winter pastures, Taoujgalt (TAO) for near
winter pastures, Imeskar (IMS) for intermediate pastures, and Tizi n’Tounza (TZT) for summer
pastures.
Summer and intermediate pastures are exclusively used by Ait Toumert households and are
subject to strong institutional arrangements. Summer pastures are temporarily protected by
means of the agdal institution. The agdal is a communal pasture; opening and closing dates
are fixed by the community of users (Genin and Simenel, 2011). Access to the summer
pasture is closed from mid-March to mid-May, and generally restricted to members of the

5
tribal fraction with access rights. The comparatively vast winter pastures have an open-access
tenure regime and are shared with at least two neighboring fractions. The schedule of
transhumance may deviate from this normative cycle, depending on rainfall, herd dynamics,
and on socio-cultural factors (Akasbi et al., 2012).
Table 1. Location, climate, and rangeland characteristics of the four study sites representing
the Ait Toumert pasture types. The map codes of study sites refer to Fig. 1. MAP is the mean
annual precipitation. Tmin is the mean daily minimum temperature of the coldest month, Tmax is
the mean daily maximum temperature of the warmest month. The UNEP aridity index AIU is
the ratio of MAP to annual potential evapotranspiration (Middelton and Thomas, 1997).
Bioclimatic unit follows Oldeland et al. (2008), the vegetation type is taken from the
vegetation map by Finckh and Poete (2008). ANPP is the above-ground primary production,
measured (like standing crop) for the growth period 2007/08 within temporal exclosures.
Grazing offtake is the percentage of ANPP consumed by livestock during that period; it
indicates recent grazing pressure. For details of standing crop and ANPP measurements refer
to Linstädter and Baumann (2013).
Ait Toumert Pasture/ Far Winter/ Near Winter/ Intermediate/ Summer/
Study site (map code) TRB TAO AMS TZT
Location
Latitude 31°10’ N 31°23’ N 31°29’ N 31°34’ N
Longitude 6°34’ W 6°19’ W 6°14’ W 6°17’ W
Altitude (m asl) 1380 1870 2250 2960
Abiotic site conditions
Bioclimatic unit Arid cool Arid cold Semiarid cold Subhumid cold
MAP (mm a-1
)* 124 170 285 363
Tmin coldest month (°C)** 9.2 3.2 3.7 -2.2
Tmax hottest month (°C)** 30.7 25.5 23.4 16.5
UNEP aridity index AIU* 0.06 0.12 0.16 0.22
Rangeland characteristics
Vegetation type Saharan
Hammada
semidesert
Ibero-Maurita-
nian Artemisia
steppe
Juniperus
woodsteppe
Oromediterra-
nean shrubland
Total standing crop (kg ha-1
) 331 889 565 8786
Woody perennials (% biomass) 59.8 80.5 81.8 98.0
Herbaceous peren. (% biomass) 1.6 10.0 7.8 0.6
ANPP (kg ha-1
yr-1
) 244 560 224 419
Grazing offtake (% of ANPP) 57.5 91.4 71.5 56.3
Abiotic site conditions: * = calculated for the years 2000 – 2006 (01.01. to 31.12.), ** =
calculated for the hydrological years 2001/02 – 2007/08.
Sheep and goats share the same pasture areas and are jointly herded. Herdsmen do not specify
their herd management for livestock species; they merely adjust their micro-mobility on local
pastures to the proportion of goats within a herd to account for the different feeding needs of
the two ruminant species. While herds dominated by goats easily access rocks and scarps, and
eat more woody plants such as cushion-like xerophytes and dwarf shrubs, herds dominated by
sheep are not able to exploit a pasture in that way. In consequence herdsmen have to be more
mobile to meet the feeding needs of sheep.
The altitudinal gradient from the central High Atlas Mountains to the basin of Ouarzazate is
also a steep gradient of environmental variability from a semi-arid to an arid environment

6
(Table 1). Similar to other altitudinal gradients of decreasing aridity (e.g. Zemmrich et al.,
2010), typical vegetation belts are distinguishable. In the High Atlas Mountains, they are
roughly congruent to the altitudinal levels of the four pasture types. The mountainous summer
pastures from 3,000 to 2,500 m asl are covered by an Oromediterranean shrubland dominated
by persistent, cushion-forming shrubs. Intermediate pastures running from 2,500 to 2,000 m
asl are mainly covered by a Juniperus woodsteppe. The near winter pastures cover the
altitudinal belt from 2,000 to 1,500 m asl. This is the zone of an Artemisia steppe, dominated
by dwarf shrubs and perennial grasses. The Saharan Hammada semidesert of the far winter
pastures (from 1,500 to 1,000 m asl) is dominated by annual species, interspersed with shrubs
such as Hammada scoparia (Benabid and Fennane, 1994; Finckh and Poete, 2008; Linstädter
and Baumann, 2013).
2.2. Implicit and tacit criteria for the valuation of forage plants
Within the context of testing EAH, the use-value index has been proposed as a quantitative
measure for the relative importance of species to a human population (Phillips and Gentry,
1993a, b) and frequently been applied (Albuquerque and Lucena, 2005; de Almeida et al.,
2005; Lawrence et al., 2005; Lucena et al., 2007; Lucena et al., 2012). However, this index
was not applicable in our case as we were not interested in multiple uses of plants, but in
species’ valuation as forage plants. Moreover, we aimed to record quality criteria including
those which are implicit or tacit. For this purpose, tools of Cultural Domain Analysis such as
scoring and ranking methods have been proposed (Bernard, 1994; Wesuls and Lang, 2010).
We chose the free-list technique (Borgatti, 1999) to gain quantitative information on the
valuation of forage species. While free-lists have mostly been used to elicit the items within a
cultural domain, they can also be analysed in terms of an item’s salience (Borgatti, 1999).
Salience analysis assumes that informants will arrange items in terms of individual saliency,
and that tacit and/or implicit criteria will determine saliency (valuation). The free-list method
has also been recommended for ethnobotanical purposes, specifically to find out the most
culturally salient plants of a particular use category (Quinlan, 2005).
We separately questioned 17 informants from 15 households to name all forage plants they
knew to be eaten by sheep and goats. Informants, normally the male head of the household,
represented the complete sample of pastoral-nomadic households that could be met on
summer and intermediate pastures during fieldwork. In three cases, women were interviewed
and in one case the son of the household-head. All households owned the animals they were
herding, and all informants were or had been engaged in tending herds. The average age of
interviewees could not be identified, as indications of birth were based on estimations due to
the lack of birth certificates. It can be assumed that the majority of informants were between
35 and 55, with one informant significantly younger and one significantly older. Some of the
informants had gone to Quran School, but none had a formal education. Those who could
afford it had started to send some of their children to school. Two translators from the region,
familiar with the socio-cultural context and with interviewing techniques assisted in
interviews which were conducted in Tachelhit. To maximize output in free-listing tasks, we
used two interviewing techniques suggested by Brewer (2002), i.e. a nonspecific prompting,
and reading back the list of free-listed items.
Although more than 20 interviews had been planned, only 17 could be realized as we were
constrained by the great difficulty to track nomadic households by foot in their pasture areas.
The sample thus misses the recommended minimum size of 20 to 30 interviews (see Sutrop,
2001 and references therein). However, this recommendation refers to the situation when only
term frequency is evaluated to establish a cultural domain. If the mean position of an item is

7
additionally considered (which we did with Sutrop’s cognitive salience index; see below),
smaller samples are sufficient (see Sutrop 2001:272).
We aimed to minimize potential bias by conducting interviews at different times and in
different localities. Most interviews were conducted by visiting the pastoral-nomads at their
current housing or on pastures (intermediate or summer pastures) in September and October
2007. At this time of the year pastoral-nomads were moving from the summer pastures to the
lower intermediate pastures, two of the smaller pasture areas where informants could be
tracked within walking distance. Upland pastures of the Ait Toumert fraction were hardly
accessible by car. Conducting interviews at two different pasture areas also served to
minimize bias by the fact that informants could tend to name species first growing on these
pastures. 45% of the informants were met on summer pastures while 55% had already moved
to intermediate pastures. As informants did not stay on winter pastures during the field study,
interviews could not be conducted there. It would also have been hardly possible to track Ait
Toumert households during winter months, when herders mix up with those of other fractions
throughout the vast areas of winter and intermediate pastures.
The results of individual interviews were later (in April 2008) supplemented and triangulated
by expert interviews with herdsmen from different households. The expert interviews were
carried out as group interviews with male herders, using a general interview guide approach.
In addition the responsible person (sheikh) for formally opening the agdal was questioned in
an individual interview. The interviews took place at the weekly market near Ait Khalifa.
During that time, households stayed on intermediate pastures which allowed an easy and
regular access to the market and the gathering of the assembly of Ait Toumert representatives
to discuss the opening of the agdal.
2.3. Explicit criteria for the valuation of forage plants
To extract explicit LEK on the valuation of forage plants, we conducted semi-structured
interviews with Ait Toumert herdsmen directly after free-list tasks. We asked what
characterizes a species as valuable forage plant for sheep and goats. Interviews rendered two
criteria roughly similar to the scientific concepts of palatability and nutritive value.
Palatability relates to plant characteristics or conditions stimulating a selective response by
animals, with palatable species being preferentially eaten (Baumont et al., 2000). The
nutritive value refers to whether a plant species will promote rapid weight gain and animal
health (Komwihangilo et al., 2001).
In a later step, we recorded local estimations of palatability and nutritive value for 62
abundant species representing different growth forms on local pasture types. This was done
during a group interview with three knowledgeable herdsmen in October 2007. Informants
were asked to classify species into ‘refused’ (0), ‘little or occasionally eaten’ (1), ‘regularly
eaten’ (2) or ‘highly favoured’ (3). Informants also classified species’ nutritive value as ‘bad’
(0), ‘medium’ (1) or ‘good’ (2). These categories resulted from the preceding interviews on
explicit criteria for the valuation of forage plants (see above). Preference ranking was done
separately for sheep and goats.
2.4. Vegetation sampling
We recorded vegetation parameters reflecting plants’ occurrence on pasture types, and their
ecological apparency (visibility and availability). Based on vegetation studies (Benabid and
Fennane, 1994; Linstädter and Baumann, 2013) and own observations, plants were assigned
to the pasture type where they mainly occurred. To record the availability components
abundance and frequency, we selected four sites representing the four vegetation belts along
the altitudinal gradient (Fig. 1). Sites were located close to weather stations established by the
IMPETUS research project in 2000. Hence, we were able to record specific climatic

8
conditions of study sites (Table 1). Even though sites were mostly positioned at the border of
or adjacent to the Ait Toumert pasture area, they were representative for the vegetation
composition the used pasture types, as the altitudinal belts of pasture types are roughly
congruent to vegetation zones. Species cover values were visually estimated on 25 m²
vegetation plots. We sampled 24 plots in each of the four pasture types (total: 96 plots),
keeping a minimum distance of 20 m between them. On each site, data were collected at the
end of the local growth period (i.e. for the time of peak biomass), which was between March
2008 (winter pastures) and June 2008 (summer pastures).
Visibility was recorded as life form and life history (lifetime). We distinguished three life form
classes, i.e. therophytes (herbaceous annuals), hemicryptophytes (herbaceous perennials) and
woody species, subsuming chamaephytes, nano-phanerophytes and phanerophytes (Table 2).
For lifetime, we ranked plant growth forms (see Table A.2 for data) according to life duration,
from short-lived annual plants (1) over perennial forbs (2), perennial grasses (3) and short-
lived woody plants (dwarf shrubs; 4) to shrubs (5) and persistent woody plants, i.e. cushion-
like xerophytes and trees (6).
2.5. Management decisions
We conducted semi-structured interviews to evaluate how the local valuation of forage
resources was linked to management decisions. To consider environmental variability and its
influence on resource availability, 16 of the 17 informants interviewed with the free-list
technique were asked to classify forage availability in past years as being very good, good,
average, bad, or very bad. We did not suggest any standard with respect to an ‘average’ year,
and interviewees were asked to go back in their memory as far as possible.
To match herders’ reports on available forage to precipitation records, we used data for two
subregions of the broader study area with similar rainfall characteristics. The classification
was based on the Standardized Precipitation Index (SPI) of hydrological years (August of the
preceding year to July of the following year), calculated for all available Moroccan weather
stations at a given time (Born et al., 2008).
As a local classification of years may, besides the availability of forage resources, reflect
other aspects such as the individual economic situation, qualitative information was used to
distinguish ecological from economic reasons. Informants were asked to recall individual
mobility decisions and alternative strategies of herd management in four hydrological years.
As neither precipitation records nor informants’ reports indicated a year with ‘very good’
resources in the close past (Table 5), we concentrated on years named as either ‘good’,
‘average’, ‘bad’ or ‘very bad’ for resource availability. With respect to management
decisions, we focused on the winter months (October to March), because this is the time of
resource scarcity in the annual cycle (Linstädter and Baumann, 2013) when local knowledge
on quality and availability of forage resources should be most important for management
decisions.
2.6. Data analysis
2.6.1. Match of local and scientific plant names
For free-listed plants only known by their vernacular, informants were asked to collect
samples for a taxonomic identification. Missing vernacular names were obtained in interviews
with local informants. Additionally, ethnobotanical publications on Moroccan plant species
were consulted (e.g. Iav, 2002; Trabut, 2006). The reconciliation of vernacular and taxonomic

9
names is prone to confusion (Szabó, 2004). We distinguished five cases (see Table A.1): (i)
plants with a clear match, (ii) synonymy, i.e. more than one vernacular for one scientific
name, (iii) homonymy, i.e. one vernacular for several scientific names, (iv) more than one
vernacular for several scientific names, and (v) no scientific information for a vernacular. In
case of synonymy, the most salient vernacular was used for further analysis. When homonyms
occurred, ground cover values of all homonymous species on a certain plot were accumulated.
If several vernaculars were related to several scientific names, plant groups were formed.
Here, 33 vernaculars and 84 scientific names were summarized into eleven plant groups. If no
scientific information was available for a vernacular, it was excluded from further analysis.
We also omitted all items that were mentioned only by one informant to improve the
robustness of the list task parameter ‘term frequency’ (see Sutrop 2001). The final
anthropological data set consisted of 79 identifiable items (out of 109 occurring on two or
more free-list, see Table A.1). In the ecological data set, 103 out of 133 items were identified
(see Table A.2) and occurred more than once on plots.
2.6.2. Match of anthropological and ecological data sets
We used a novel ethnobotanical method to match free-list (anthropological) data and
ecological data. Both data sets were parameterized in the same way with a weighted rank
index, i.e. the Cognitive Salience Index (CSI) proposed by Sutrop (2001) for free-lists. CSI
calculates the salience (S) of an item based on its frequency (F) and mean position (mP) on
free-lists, and the number (N) of informants, where S = F/(N mP). Values range between zero
and one. Plants with high CSI values are listed most commonly by informants, and are
recalled more immediately than other plants (Thompson and Juan, 2006). To calculate CSI
values for the ecological data set (CSIeco), we set CSI parameters defined for free-lists as
analogous to parameters of vegetation plots:
1a. Individual free-lists of N interview partners on forage species to
1b. Plant species lists of N vegetation plots (including non-forage species)
2a. Mean rank of a species in each free-list (mP) to
2b. Mean rank of a species on each plot (mP) (abundance, measured as ground cover)
3a. Species’ frequency in all free-lists (F>2) to
3b. Species’ frequency on all plots (F>2).
In analogy to CSIanthro which quantifies a forage plant’s salience (local valuation), its mean
abundance (mP) on vegetation plots together with its frequency (F) determines its availability
(CSIeco). We used the statistical software ANTHROPAC 4.0 (Borgatti, 1996) for all CSI
calculations (see Table A.2 for CSI values).
2.6.3. Statistical analysis of explicit LEK
We treated CSI data and the index of lifetime as continuous. To improve normality and
homogeneity of variances, CSIanthro and CSIeco data were log-transformed, and data on plant
lifetime were z-transformed before analysis. As ‘palatability for sheep’, ‘nutritive value for
sheep’, ‘palatability for goats’, and ‘nutritive value for goats’ were on an ordinal scale, we
used Spearman’s rank correlation coefficient to correlate them to each other and to CSIanthro.
2.6.4. Statistical analysis of implicit LEK
For the intersecting data set of plants with non-zero CSI values in both the anthropological
and the ecological data set, we used general linear models (ANCOVAs) to test the effects of
the factors ‘pasture type’ and ‘life form’ and the linear predictors ‘lifetime’ and ‘CSIeco’ and
their interactions on the response variable CSIanthro (Table 2). We first fitted a full model,

10
including all variables and two-way interactions. Model simplification involved a stepwise
backward selection based on the Akaike Information Criterion (AIC). This goodness-of-fit
statistic considers both fit and model complexity; it penalizes models for each explanatory
variable, i.e. for greater complexity (Johnson and Omland, 2004). Analyses were performed
using the software R 2.13.1 (R Development Core Team 2011).
Table 2. Parameters tested as predictors for the local valuation (Cognitive Salience Index,
CSIanthro) of forage plants, and their connection to criteria of Ecological Apparency
Hypothesis (EAH).
Parameter Acronym Description Range EAH criterion
Ecological
performance
CSIeco Weighted rank based on
a plant’s abundance and
frequency on plots
0.01-0.27 (potential: 0-1) Availability
Life form LF Raunkiaer life form,
chamaephytes and
phanerophytes grouped
as ‘woody’
Therophytes (annuals)/
hemicryptophytes (perennial
forbs and grasses)/ woody
plants (shrubs and trees)
Visibility
Plant
lifetime
Lifetime Index of plant life
duration
1 (annuals) -6 (trees) Visibility
Occurrence
on pasture
types
Pasture Pasture type where
plant mainly occurs
Summer pasture (SU)/
intermediate pasture (TR)/
near winter pasture (NW)/
far winter pasture (FW)
Not related to
EAH
3. Results
3.1. Explicit LEK on forage plants
The four explicit local criteria for plant quality (‘palatability for sheep’, ‘nutritive value for
sheep’, ‘palatability for goats’, and ‘nutritive value for goats’) were not significantly
correlated to CSIanthro values (Table 3). In contrast, all local criteria were correlated, with the
highest correlation between a plant’s nutritive value for sheep and its nutritive value for goats.
Pastoral nomads did thus not clearly distinguish between a plant’s forage value for sheep and
goats.
Table 3. Correlation (Spearman's rank correlation coefficient) between explicit local criteria
for forage plant quality (palatability and nutritive value for sheep and goats) and the local
valuation of 62 fodder plants, quantified with the Cognitive Salience Index (CSIanthro).
Significant correlations (p < 0.01) are marked with an asterisk.
CSIanthro Palatability
for sheep
Nutritive value
for sheep
Palatability
for goats
Nutritive
value for goats
Palatability for sheep 0.054 1.000
Nutritive value for
sheep
0.282 0.667* 1.000
Palatability for goats 0.167 0.536* 0.457* 1.000
Nutritive value for
goats
0.151 0.454* 0.681* 0.642* 1.000
11
3.2. Implicit and tacit LEK on forage plants
From the two complementary data sets on forage plants, we identified an intersecting set of 49
forage plants that had two non-zero CSI values. In the minimal adequate model (Table 4),
CSIanthro values of these plants depended on three out of four explanatory variables included in
the full model.
Table 4. Minimal adequate model for the local valuation of forage plants (quantified with the
Cognitive Salience Index, CSIanthro) as a function of the predictor variables ‘pasture’ (presence
of plants on local pasture types), ‘lifetime’, ‘CSIeco’ (plants’ ecological performance
quantified with the Cognitive Salience Index) and ‘life form’ (not included in the minimal
adequate model). Effect sizes are given as Eta-squared ( ). For details on predictor variables
refer to Table 2.
Parameter SS Df MS F P
Pasture 8.29 3 2.76 5.34 0.003 0.17
Lifetime 7.99 1 7.99 15.48 < 0.001 0.16
CSIeco 6.70 1 6.70 12.99 < 0.001 0.14
Pasture : Lifetime 4.86 3 1.62 3.14 0.035 0.10
Residuals 21.16 41 0.52
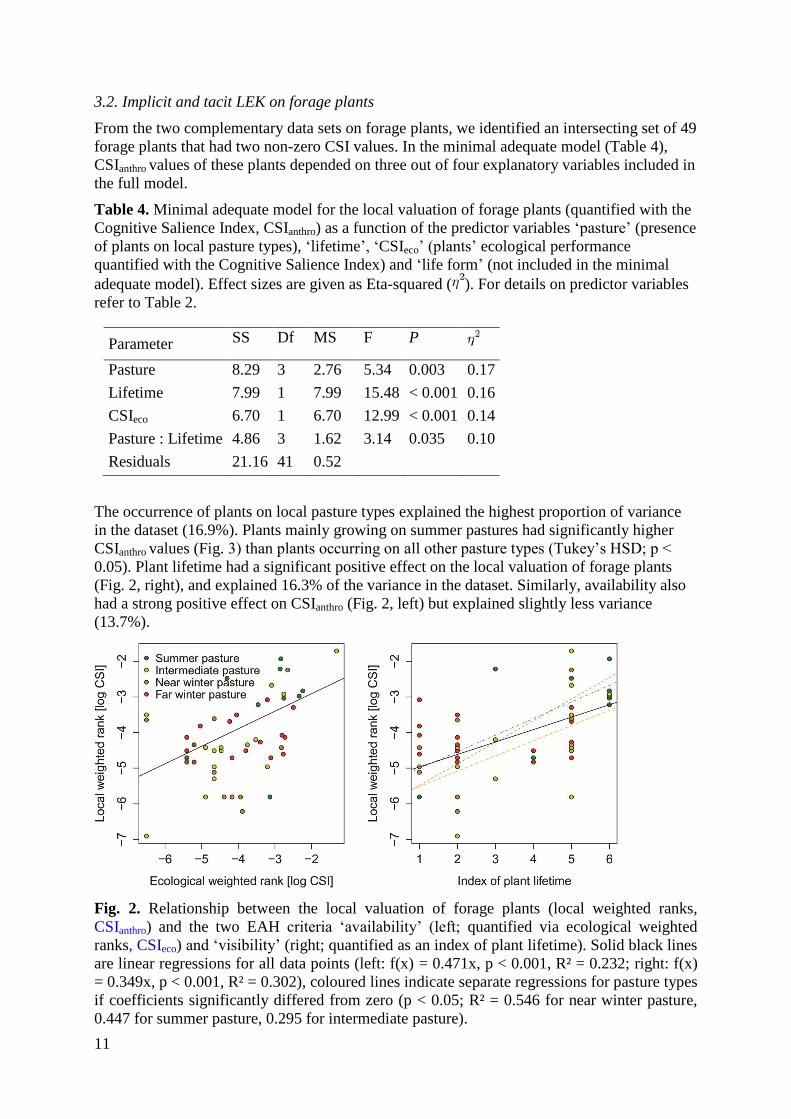
The occurrence of plants on local pasture types explained the highest proportion of variance
in the dataset (16.9%). Plants mainly growing on summer pastures had significantly higher
CSIanthro values (Fig. 3) than plants occurring on all other pasture types (Tukey’s HSD; p <
0.05). Plant lifetime had a significant positive effect on the local valuation of forage plants
(Fig. 2, right), and explained 16.3% of the variance in the dataset. Similarly, availability also
had a strong positive effect on CSIanthro (Fig. 2, left) but explained slightly less variance
(13.7%).
Fig. 2. Relationship between the local valuation of forage plants (local weighted ranks,
CSIanthro) and the two EAH criteria ‘availability’ (left; quantified via ecological weighted
ranks, CSIeco) and ‘visibility’ (right; quantified as an index of plant lifetime). Solid black lines
are linear regressions for all data points (left: f(x) = 0.471x, p < 0.001, R² = 0.232; right: f(x)
= 0.349x, p < 0.001, R² = 0.302), coloured lines indicate separate regressions for pasture types
if coefficients significantly differed from zero (p < 0.05; R² = 0.546 for near winter pasture,
0.447 for summer pasture, 0.295 for intermediate pasture).

12
We found that the factor life form and all interactions except the two-way interaction between
pasture type and lifetime did not influence local valuation of forage plants (p > 0.05). The
significant interaction (pasture type: lifetime) confirms that regression coefficients differed
between pasture types (p < 0.05; see Fig. 2 for linear regressions between CSIanthro values and
lifetime indices specified for pasture types).
Fig. 3. Local valuation (log CSIanthro) of forage plants on the four pasture types of the Ait
Toumert, arranged (from left to right) along a gradient of decreasing environmental
variability. FW = far winter pasture, NW = near winter pasture, TR = intermediate pasture,
SU = summer pasture. Boxes show medians and 25th
to 75th
percentiles, whiskers stand for the
non-outlier ranges of the data. Different superscripts (a, b) indicate significant differences
(Tukey’s HSD; p < 0.05).
3.3. Forage availability and management decisions
While some herdsmen were able to recall the quality of years back to the mid 1970ies, most
herdsmen only remembered the past four to six years. Thus, the frequency of mentions
declined for more distant years, with 51 % of all statements referring to the past two years.
Although herdsmen referred to the entire hydrological year, it became clear throughout the
interviews that the winter months with the erratic precipitation patterns on winter pastures
were crucial for the classification of years. The local perception of available resources showed
little variation for the past four years (2003/04 to 2006/07) (coefficients of variation < 0.4; see
Table 5) but variation increased for the years 2002/03 and 2001/02. Reports for prior years
generally concerned ‘very bad’ years (results not shown). We matched precipitation records
from two subregions of the Ait Toumert pasture area to herder reports (Table 5) and found
that reports tended to be quite accurate, although very sensitive to low precipitation in any of
the two subregions. Individual mobility decisions during winter considerably deviated from
the normative transhumance cycle (Fig. 4), which would be to move to near winter pastures in
November, and then – in years with good resource availability – to far winter pastures in
December. In years remembered as average and bad, households often remained on
intermediate pastures and on far winter pastures. The near winter pastures were preferably
used in good, but also in very bad (drought) years. Households exhibited different patterns of
intra-seasonal movements on pastures. Whereas transhumant mobility between different
pastures was practised in average and bad years, a constrained mobility occurred during good
and very bad years: households tended to stay on one pasture type. Decisions for a
constrained mobility were often combined with new strategies of herd management, such as
feed supplementation, and truck transport, or a mass selling of animals.

13
Fig. 4. Macro-mobility of Ait Toumert households (n = 16) during the winter months of four
years which were recalled as either ‘good’, ‘average’, ‘bad’ or ‘very bad’ with respect to
available forage resources (A-D; Very Bad Year = drought year; see Table 5).
Table 5
Herders’ assessment of forage availability on local pastures for the six most recent growth
periods (LEK-based classification), compared to a record-based classification of precipitation
which uses the Standardized Precipitation Index (SPI) of hydrological years for two
subregions of the study area with similar rainfall characteristics (see Born et al., 2008), i.e.
SOA = the semi-arid region south of the Atlas Mountains, and ATL = the Atlantic region,
including high elevations of the Atlas Mountains. SPI classes follow the categories proposed
by McKee et al. (1993), with an additional indication of above-average (+) and below-average
(-) precipitation for near-normal years. LEK classes are based on the median of individual
assessments (‘very bad’ years scoring as 0, and ‘very good’ years as 4). Coefficients of
variation (CV; the ratio of the standard deviation to the mean) are given as a measure of the
relative variability of assessments.
Year LEK-based classification Record-based classification
N Mean CV Median LEK class N
(SOA)
SPI class SOA N
(ATL)
SPI class ATL
2006/07
13 2.46 0.356 3.00 Good
10 Near normal (+) 10 Near normal (+)
2005/06
10 1.40 0.369 1.00 Bad
10 Near normal (-) 12 Severely dry
2004/05
5 2.20 0.203 2.00 Bad
10 Moderately wet 13 Near normal (+)
2003/04
4 1.50 0.385 1.50 Average to bad
11 Near normal (-) 13 Near normal (+)
2002/03
4 1.75 0.719 2.00 Average
11 Near normal (-) 16 Near normal (+)
2001/02 4 0.75 2.000 0.00 Very bad 11 Severely dry 15 Moderately dry

14
During the winter months, constrained mobility was a common alternative to transhumance in
all types of years (Fig. 5). Some informants stated that they did not move to far winter
pastures in spite of good forage availability because they had already bought supplementary
feed for the winter. Feed supplementation had been introduced by the Moroccan government
during a drought in the 1980s. Today pastoralists have adopted it as a regular strategy for the
unpredictable winter months: About two-thirds of informants stated that they provided
supplementary feed during winter months, irrespective of the availability of natural resources.
As households could buy supplementary feed on a market close to the intermediate or near
winter pastures, they tended to stay there in average, bad or very bad years. The truck
transport constitutes another new strategy to react more flexibly in drought years (Fig. 5).
While the majority of households remained on the near winter pastures of the Ait Toumert
territory, some households with access to alternative income sources could afford to rent a
truck, and transported their herds to far-off pastures. The remaining households often sold
their animals.
Range management decisions were clearly based on the set of strategies available to
households, which has been constantly changing and included new strategies introduced
during the last decades. While all households moved to summer pastures, mobility decisions
during winter months were taken based on the perception of forage availability (see above)
and on a household’s resource portfolio such as income, manpower and social networks. For
example, the access of winter pastures required manpower and transport animals, while a
constraint transhumance during winter months required feed supplementation which needed
monetary resources (see Breuer (2007) for more detailed information on monetary,
incorporated and socially institutionalized resources in pastoral-nomadic groups of the High
Atlas Mountains).
Fig. 5. Proportion of Ait Toumert households (n = 16) which had applied alternative strategies
during the winter months of four years which were recalled as either ‘good’, ‘average’, ‘bad’
or ‘very bad’ with respect to available forage resources. As households could apply several
strategies during the same year, proportions may sum up to > 100%.
4. Discussion
4.1. Explicit and implicit LEK on forage plants
We tested explicit criteria for the local valuation of plants as forage, as well as non-explicit
criteria reflecting a plant’s ecological apparency and its occurrence on pasture types. Our
results show that the explicit local criteria ‘nutritive value’ and ‘palatability’ did not
determine if a plant was immediately recalled and frequently named as a valuable forage

15
plant. By relating weighted ranks to criteria recorded with ecological methods, we were able
to identify and rank their relative importance. The attribution of a plant to a specific pasture
type was the most important non-explicit criterion for its valuation, followed by the plant’s
lifetime and its availability. Plants mainly growing on summer pastures had significantly
higher CSIanthro values than plants on all other pasture types. This was partly determined by
the higher average lifetime of plants growing on summer pastures, and by their higher
abundance and frequency (i.e. availability). However, even if the effects of lifetime and
availability on CSIanthro were statistically removed, the plant’s attribution to a certain pasture
type remained an important criterion for its local valuation. Plants growing on summer
pastures are much more valued than other plants.
Our data indicate that herdsmen used – besides the pasture type where a plant mainly occurs –
the lifetime and the availability of plants as indicators of their quality. The high valuation of
persistent species is in congruence with the ecological apparency hypothesis (EAH) and
corresponds to studies from elsewhere, showing that woody plant species have generally
higher use values than herbaceous plants (e.g. Ayantunde et al., 2009; Lucena et al., 2007). In
contrast to our results, though, earlier ethnobotanical studies found no positive correlation
between the relative importance and the availability of forage species (Lucena et al., 2007;
Lucena et al., 2012). Hence, the present study is to our knowledge the first to confirm the
validity of EAH within the use category of forage species. For Ait Toumert herdsmen,
visibility (expressed as lifetime) and availability (expressed as CSIeco values) were almost
equally important in determining a forage plant’s apparency.
However, herders had (besides the EAH criteria ‘visibility’ and ‘availability’) another implicit
criterion for the quality of a forage plant, i.e. its occurrence on upland pastures. We assume
that the need to cope with the high spatio-temporal variability of natural resources which is
typical for dryland systems (Thomas et al., 2007) may explain why apparent forage plants
from pasture types with a relatively constant environment were higher valued than those from
more arid environments. While persistent plants from upland pastures were hardly more
productive than those of lowland pastures (Linstädter and Baumann, 2013), they offered
reliable forage resources. We thus identify the reliability of dryland plants and pastures as a
key element for local valuation. Moroccan pastoral-nomads benefit from a cultural domain of
forage resources including reliability as a specific value in a highly variable environment.
Hence, reliability is - besides the two EAH criteria visibility and availability - another
important criterion to explain the use of plant resources in local communities. It also gives a
more functional explanation than EAH criteria why a woody forage plant should be more
valued than a herbaceous plant: It is not only more visible for livestock and/or for herdsmen,
but also offers more predictable forage resources.
As part of the interviews was carried out on summer pastures, our result that plants mainly
growing on summer pastures had significantly higher CSIanthro values could also be related to
informants’ bias (i.e. informants could have tended to name first species occurring on the
pasture type where they stayed during the time of interview). For logistical reasons, we were
not able to spread interviews over the whole annual cycle, which would have been necessary
to completely avoid possible bias. However, we assume any bias to be small, as ca. 50% of
interviews were carried out when households stayed on intermediate pastures. Another
methodological issue is the comparatively small sample size of free-lists (17 informants
representing more than 50 % of all Ait Toumert households) used for CSIanthro calculation. It
lies below the minimum number of 20-30 which is accepted as the norm to attain a robust
frequency parameter from free-list tasks (Sutrop, 2001). However, as mentioned in section
2.2, smaller samples may be sufficient if not only item frequency but also the mean position
of items is considered (as done in our study via CSI calculation). Sutrop (2001:272) explicitly
mentions this as an advantage of the CSI. Moreover, a methodological study on free-list tasks

16
– which evaluated the relative importance of sample size and the deletion of low-frequency
terms for robust frequency-based results – stressed the crucial importance of the latter
procedure only (Schrauf and Sanchez, 2010). Due to the fact that we omitted low-frequency
items prior to CSI calculation, and due the robustness of CSI with respect to a small sample
size, we assume that our conclusions based on CSIanthro are sound.
We triangulated our conclusions in April 2008 with an expert group interview with herdsmen
from different households. Herdsmen agreed that a plant with a higher lifetime would be a
more valuable forage resource, because it had a higher ability to survive drought times, and to
provide forage in bad and very bad years. They argued that plants growing on summer and
intermediate pastures were generally more valuable because they were a forage resource
which could be exploited annually.
We assume that plant valuation is amplified by the high cultural importance of reliable upland
pastures, which are under an exclusive tenure regime with strong institutions. In contrast,
winter pastures are an unreliable, low-quality resource type (following Scoones, 1999). They
are less contested and only protected by weak institutions. Hence, resource valuation may be a
self-enforcing process, with reliable, contested resources protected by strong institutions
becoming even more valuable in local perception. Incidentally, in the case of forage plants
EAH does not allow distinction whether a plant’s cultural importance is influenced by an
observation of livestock behaviour, and/or by herdsmen demonstrating the same behaviour as
herbivores (as assumed by Phillips and Gentry, 1993a, b). However, this conceptual
vagueness does not undermine the general applicability of EAH to forage plants. Instead of
use values, which were originally proposed within the context of EAH (Phillips and Gentry,
1993a, b), we have applied a combination of free-list rankings and weighted rank calculations
to obtain a quantitative measure of the relative importance of species. This approach has the
advantage that it can evaluate the relative importance within a single use category, and has for
this reason been recommended ‘if one wants to find the most culturally salient plants of a
particular sort’ (Quinlan, 2005). If one was more interested in explicit criteria for the
valuation of species, one could combine weighted rank calculations with explicit rankings
obtained in individual or group interviews (see e.g. Oba et al., 2008).
4.2. Linking the availability and valuation of forage resources to management decisions
Herdsmen’s reports on available resources were reasonably consistent for the past 3-4 years
but less so for years further in the past; for years more a decade away herdsmen tended to
recall only drought years. This appears to be common for pastoral and agro-pastoral societies
without written historical records or documentation about resources use (Meze-Hausken,
2004, Mogotsi et al., 2013). Precipitation records from the past six years corroborate the
accuracy of local reports for the six years, and suggest that the yardstick for an ‘average’ year
roughly corresponds to a ‘near-normal’ annual precipitation. Interestingly, the uncertainty
and risk experienced during winter months dominated the perception of past years.
Pastoral perception of forage resources on local pastures was an important component of
management decisions (Wesuls and Lang, 2010). In our case, Ait Toumert decision-making
can both be linked to the resource availability in a particular year and to the reliability of
forage plants on pasture types, i.e. to a non-explicit criterion for plant valuation. Pastures with
a high abundance of perennial plants were more reliable and thus more valuable. The reliable
upland pastures were regularly grazed during spring, summer and autumn. Herdsmen were
conscious about the unreliability of winter pastures where annual plant species are dominant.
Their decision to move there largely depended on the resource availability in a particular year.
An analysis of individual herd mobility patterns, based on mobility decision in four years
from the near past differing in resource availability, showed that considerable between-year
differences occurred in mobility decisions during winter time. Only in a year with good

17
rainfall, most of the households decided to move to winter pastures, because herdsmen knew
that they would then find an abundance of herbaceous biomass for their livestock. In other
types of years, only a certain proportion of pastoral-nomadic households moved to winter
pastures. Instead, households stayed on intermediate pastures where perennial species are
more abundant. Thus, mobility decisions depended on the availability of forage resources in
general (which is related to the amount of rainfall in a given year; Ruppert et al., 2012), and
specifically on the availability of forage resources provided from perennial species: The less
perennial species occurred on a certain pasture, the more the movement to this pasture
depended on the type of year.
The mobility patterns of the Ait Toumert allow an exploitation of the diversity of vegetation
types along a steep aridity gradient, securing the access to natural resources which were
highly variable in space and time. This opportunistic mobility, which implies ‘to be at the
right place at the right moment’ has been described as analogous to the movements of large
wild herbivores in their arid environments and would then be a mere coping or resource
exploitation strategy on a short-term level (Fabricius et al., 2007), i.e. a fall-back mechanism.
However, several aspects of Ait Toumert mobility patters may also be interpreted as a
resilient form of natural resource management. First, the regular seasonal movement between
pastures leaves enough forage on certain pasture types for the scarce times of the year or for
drought times (Linstädter et al., 2010), increasing forage resource reliability. Another
mechanism to promote resource reliability is through resting highly utilised pastures in years
with abundant rainfall (Müller et al., 2007a). In our case study, herds were frequently moved
to near and far winter pastures in years with good resource availability (Fig. 5), allowing
perennial vegetation on intermediate pastures to recover (Linstädter and Baumann, 2013). On
upland pastures, resource reliability is also maintained by means of mobility patterns
regulated by the agdal institution. The agdal protects perennial species on summer pastures
from grazing during the onset of the vegetation period (Genin and Simenel, 2011), when plant
individuals are highly sensitive to defoliation. This mechanism is clearly articulated by Ait
Toumert herders: ‘Forage plants need this time to flourish and to provide sufficient forage
during the summer months. If pastoral-nomads would move to the Awjgal [summer pasture]
in spring time, there would be less forage for livestock’. (Informant 14, October 6th
2007). The
agdal thus allows a recovery of palatable perennial plants, and minimizes resource
degradation via a deferred mobility. The crucial importance of protecting or improving
pasture reliability through resting in times when forage plants are particularly sensitive to
grazing has also been reported to build resilience in other dryland systems (Müller et al.,
2007b).
In arid and semi-arid environments with their high spatial and temporal variability, ecological
reliability is thus both sought after and protected via mobility patterns. Perceiving reliability,
and adjusting management decisions to it, are thus important mechanisms of resilience in
dryland pastoral systems (Fig. 6): They increase the system’s ability to cope with and
regenerate from external shocks such as droughts (Dougill et al., 2010).
Linking the local valuation of forage plants to mobility decisions has certain limitations as
individual mobility strategies are also influenced by socio-economic factors like individual
access to incorporated, monetary and socially institutionalized resources (Akasbi et al., 2012;
Breuer, 2007). Hence, the capacity of different households to search and protect ecological
reliability can vary (Kuhn et al., 2010).
4.3. New management strategies of the Ait Toumert: Implications for resilience
Today, Ait Toumert households are not as dependent on forage resources and their reliability
as they used to be. Feed supplementation became a common practice (Akasbi et al., 2012).

18
Transporting livestock to other grazing areas, and mass selling of animals are also new
management strategies. As typical for recent changes in pastoral societies of the High Atlas
region (Breuer, 2007) and elsewhere (Bollig, 2006; Greiner, 2011), new strategies such as
reciprocal grazing agreements with neighboring fractions, alternative monetary income from
wage labor, and socio-economic reciprocity have been introduced, and the social, economic,
or personal situation of a household influences its portfolio of old and new strategies, and its
selection of a herd management strategy in a specific situation. Alternative strategies also
seek or create reliability within the social-ecological system by buffering resource variability
(McAllister et al., 2006). However, the Ait Toumert increasingly rely not on ecological, but
on economic buffers: A resilience mechanism closely connected to the ecological system is
increasingly replaced by other mechanisms. Due to a weakening of environmental feedback
(Lorent et al., 2009; McAllister et al., 2006), the recent tendency of the Ait Toumert to
become decoupled from their natural resource base may accelerate resource degradation, and
decrease the resilience of the pastoral system. To evaluate the resilience of changing dryland
social-ecological systems, an integrated view of the social and natural sciences will be
necessary (Easdale and Aguiar, 2012).
Fig. 6. Interactions in a dryland pastoral system. Between the ecological subsystem (right-
hand side) and the social subsystem (left-hand side) of the social-ecological system, dynamic
interactions are the perception of natural resources, a decision-making-process leading to a
selection of a particular management strategy, and the impact of land management on
resources. Local perception of available natural resources is related to local ecological
knowledge on their spatio-temporal patterns and their quality.
5. Conclusion
Our study confirms the validity of EAH for the use category of forage species and for dryland
environments. EAH criteria ‘visibility’ and ‘availability’ explained the valuation of forage
plants even better than herdsmen’s explicit criteria. Our results also imply that EAH should be
modified for dryland environments with their high environmental variability. Apparent forage
plants from pastures within a relatively constant environment were higher valued than those
growing under more variable environmental conditions. We have introduced the concept of
reliability into EAH to explain the disproportionately high valuation of plants from less arid

19
environments. We could also show that reliability is a useful concept to link the perception of
natural resources in arid and semi-arid environments to mobility decisions. Finally, our results
underline that exploring local criteria for the valuation of natural resources can improve our
understanding of management decisions and resilience mechanisms in dryland systems.
Acknowledgements
We are deeply indebted to the Ait Toumert families for their hospitality and willingness to
share their knowledge. Our Moroccan partners, particularly Sabah Benchaaboune, Aziz
Rahou, and Ahmet Ramedane (CBTHA) are thanked for fruitful discussions on local
knowledge and rangeland management. Special thanks to Redouane Oumouch and Aziz
Labdi for translation and field assistance. We thank Jenny Eisold, Pierre Fritzsche and Silke
Tönsjost for technical and methodological support. Michael Bollig, Christoph Zielhofer, the
editor and two anonymous referees provided insightful comments on earlier versions of the
manuscript.
Disclosure statement
No actual or potential conflicts of interest have influenced the work of the authors. Authors
have closely collaborated in all stages of the research, and have approved the final version of
this manuscript.
Role of the funding source
This work was supported by the Federal German Ministry of Education and Research
(BMBF) under grant No. 01 LW 06001B, and by the Ministry of Innovation, Science,
Research and Technology (MIWFT) of the federal state of Northrhine-Westfalia under grant
No. 313-21200200. A. L. also acknowledges funding by the German Science Foundation
(DFG) through the collaborative research project FOR 1501. Funding sources have not
participated in, or conditioned any stage of the work here presented.
References
Akasbi, Z., Oldeland, J., Dengler, J., Finckh, M., 2012. Social and ecological constraints on
decision making by transhumant pastoralists: a case study from the Moroccan Atlas
Mountains. Journal of Mountain Science 9, 307-321.
Albuquerque, U.P., Lucena, R.F.P., 2005. Can apparency affect the use of plants by local
people in tropical forests? Interciencia 30, 506-510.
Ayantunde, A.A., Hiernaux, P., Briejer, M., Udo, H., Tabo, R., 2009. Uses of local plant
species by agropastoralists in south-western Niger. Ethnobotany Research & Applications 7,
53-66.
Baumont, R., Prache, S., Meuret, M., Morand-Fehr, P., 2000. How forage characteristics
influence behaviour and intake in small ruminants: a review. Livestock Production Science
64, 15-28.
Benabid, A., Fennane, M., 1994. Connaissances sur la végétation du Maroc: Phytogéographie,
phytosociologie et séries de végétation. Lazaroa 14, 21-97.
Bernard, H.R., 1994. Research Methods in Anthropology: Qualitative and Quantitative
Approaches. Sage Publications, Thousand Oaks, Canada.
Bollig, M., 2006. Risk Management in a Hazardous Environment - a Comparative Study of
two Pastoral Societies. Springer, New York.

20
Bollig, M., Schulte, A., 1999. Environmental change and pastoral perceptions: degradation
and indigenous knowledge in two African pastoral communities. Human Ecology 27, 493-
514.
Borgatti, S., 1996. Anthropac, 4.0 ed. Analytic Technologies, Lexington, USA.
Borgatti, S., 1999. Elicitation techniques for cultural domain analysis, in: Schensul, J.,
LeCompte, M., Nastasi, B.K., Borgatti, S. (Eds.), Enhanced Ethnographic Methods:
Audiovisual Techniques, Focused Group Interviews, and Elicitation Techniques. AltaMira
Press, Walnut Creek, USA, pp. 115-151.
Born, K., Fink, A.H., Paeth, H., 2008. Dry and wet periods in the northwestern Maghreb for
present day and future climate conditions. Meteorologische Zeitschrift 17, 533-551.
Breuer, I., 2007. Livelihood security and mobility in the High Atlas Mountains, in: Gertel, J.,
Breuer, I. (Eds.), Pastoral Morocco: Globalizing Scapes of Mobility and Insecurity. Reichert,
Wiesbaden, Germany, pp. 165-179.
Brewer, D.D., 2002. Supplementary interviewing techniques to maximize output in free
listing tasks. Field Methods 14, 108-118.
Butt, B., 2010. Seasonal space-time dynamics of cattle behavior and mobility among Maasai
pastoralists in semi-arid Kenya. Journal of Arid Environments 74, 403-413.
de Almeida, C.F.C.B., de Lima e, S., de Amorim, E.L.C., Maia, M.B., de Albuquerque, U.P.,
2005. Life strategy and chemical composition as predictors of the selection of medicinal
plants from the caatinga (Northeast Brazil). Journal of Arid Environments 62, 127-142.
Dougill, A.J., Fraser, E.D.G., Reed, M.S., 2010. Anticipating vulnerability to climate change
in dryland pastoral systems: using dynamic systems models for the Kalahari. 15(2). Ecology
and Society 15 (2), 17. [online] URL: http://www.ecologyandsociety.org/vol15/iss12/art17/.
Dwyer, M., Istomin, K., 2008. Theories of nomadic movement: A new theoretical approach
for understanding the movement decisions of Nenets and Komi reindeer herders. Human
Ecology 36, 521-533.
Easdale, M.H., Aguiar, M.R., 2012. Regional forage production assessment in arid and semi-
arid rangelands – A step towards social–ecological analysis. Journal of Arid Environments 83,
35-44.
Fabricius, C., Folke, C., Cundill, G., Schultz, L., 2007. Powerless spectators, coping actors,
and adaptive co-managers: a synthesis of the role of communities in ecosystem management.
Ecology and Society 12(1), 29. [online] URL:
http://www.ecologyandsociety.org/vol12/iss21/art29/.
Fazey, I., Proust, K., Johnson, B., Fazey, J.A., 2006. Eliciting the implicit knowledge and
perceptions of on-ground conservation managers of the Macquarie Marshes. Ecology and
Society 11, 25. [online] URL: http://www.ecologyandsociety.org/vol11/iss21/art25/.
Fernandez-Gimenez, M.E., 2000. The role of Mongolian nomadic pastoralists' ecological
knowledge in rangeland management. Ecological Applications 10, 1318-1326.
Finckh, M., Poete, P., 2008. Vegetation map of the Drâa basin. In: Schulz, O., Judex, M.
(Eds.), IMPETUS Atlas Morocco. Research Results 2000-2007. Department of Geography,
University of Bonn, Bonn, pp. 31–32.
Genin, D., Simenel, R., 2011. Endogenous Berber forest management and the functional
shaping of rural forests in Southern Morocco: implications for shared forest management
options. Human Ecology 39, 257-269.
Greiner, C., 2011. Migration, translocal networks and socio-economic stratification in
Namibia. Africa 81, 606-627.

21
Iav, 2002. Étude relative à l'inventaire participatif de la biodiversité dans le versant sud du
Haut Atlas, in: Institut Agronomique et Vétérinaire, H., II (Ed.). Conservation de la
Biodiversité par la Transhumance dans le Versant sud du Haut Atlas (CBTHA), Ouarzazate,
Morocco, pp. 1-78.
Johnson, J.B., Omland, K.S., 2004. Model selection in ecology and evolution. Trends in
Ecology & Evolution 19, 101-108.
Komwihangilo, D.M., Sendalo, D.S.C., Lekule, F.P., Mtenga, L.A., Temu, V.K., 2001.
Farmers' knowledge in the utilisation of indigenous browse species for feeding of goats in
semi arid central Tanzania. Livestock Research for Rural Development 13, 1-7.
Kuhn, A., Heidecke, C., Roth, A., Goldbach, H., Burkhardt, J., Linstädter, A., Kemmerling,
B., Gaiser, T., 2010. The importance of resource management for livelihood security under
climate change, in: Speth, P., Christoph, M., Diekkrüger, B. (Eds.), Impacts of Global Change
on the Hydrological Cycle in West and Northwest Africa. Springer, Berlin, pp. 565-590.
Lawrence, A., Phillips, O.L., Ismodes, A.R., Lopez, M., Rose, S., Wood, D., Farfan, A.J.,
2005. Local values for harvested forest plants in Madre de Dios, Peru: Towards a more
contextualised interpretation of quantitative ethnobotanical data. Biodiversity and
Conservation 14, 45-79.
Linstädter, A., Baumann, G., 2013. Abiotic and biotic recovery pathways of arid rangelands:
Lessons from the High Atlas Mountains, Morocco. CATENA 103, 3-15.
Linstädter, A., Baumann, G., Born, K., Diekkrüger, B., Fritzsche, P., Kirscht, H., Klose, A.,
2010. Land use and land cover in Southern Morocco: managing unpredictable resources and
extreme events, in: Speth, P., Christoph, M., Diekkrüger, B. (Eds.), Impacts of Global Change
on the Hydrological Cycle in West and Northwest Africa. Springer, Berlin, pp. 611-632.
Lorent, H., Sonnenschein, R., Tsiourlis, G.M., Hostert, P., Lambin, E., 2009. Livestock
subsidies and rangeland degradation in central Crete. Ecology and Society 14(2), 41. [online]
URL: http://www.ecologyandsociety.org/vol14/iss42/art41/.
Lucena, R.F.P., Araújo, E.L., Albuquerque, U.P., 2007. Does the local availability of woody
Caatinga plants (Northeastern Brazil) explain their use value? Economic Botany 61, 347-361.
Lucena, R.F.P., Medeiros, P.M., Araújo, E.L., Alves, A.G.C., Albuquerque, U.P., 2012. The
ecological apparency hypothesis and the importance of useful plants in rural communities
from Northeastern Brazil: An assessment based on use value. Journal of Environmental
Management 96, 106-115.
McAllister, R.R.J., Gordon, I.J., Janssen, M.A., Abel, N., 2006. Pastoralists' responses to
variation of rangeland resources in time and space. Ecological Applications 16, 572-583.
McKee, T.B., Doesken, N.J., Kleist, J., 1993. The relationship of drought frequency and
duration to time scales. 8th Conference on Applied Climatology, January 17-22, 1993,
Anaheim, California, pp. 179-184.
Meze-Hausken, E., 2004. Contrasting climate variability and meteorological drought with
perceived drought and climate change in northern Ethiopia. Climate Research 27, 19-31.
Middelton, N.J., Thomas, D.S.G., 1997. World Atlas of Desertification. United Nations
Environmental Programme (UNEP), London, UK.
Mogotsi, K., Nyangito, M.M., Nyariki, D.M., 2013. The role of drought among agro-pastoral
communities in a semi-arid environment: The case of Botswana. Journal of Arid
Environments 91, 38-44.
Müller, B., Frank, K., Wissel, C., 2007a. Relevance of rest periods in non-equilibrium
rangeland systems - a modelling analysis. Agricultural Systems 92, 295-317.

22
Müller, B., Linstädter, A., Frank, K., Bollig, M., Wissel, C., 2007b. Learning from local
knowledge: modeling the pastoral-nomadic range management of the Himba, Namibia.
Ecological Applications 17, 1857-1875.
Oba, G., Byakagaba, P., Angassa, A., 2008. Participatory monitoring of biodiversity in East
African grazing lands. Land Degradation & Development 19, 636-648.
Oldeland, J., Finckh, M., Born, K., 2008. A bioclimatic map for southern Morocco. In:
Schulz, O., Judex, M. (Eds.), IMPETUS Atlas Morocco. Research Results 2000–2007.
Department of Geography, University of Bonn, Bonn, pp. 21–22.
Phillips, O., Gentry, A., 1993a. The useful plants of Tambopata, Peru: I. Statistical hypotheses
tests with a new quantitative technique. Economic Botany 47, 15-32.
Phillips, O., Gentry, A., 1993b. The useful plants of Tambopata, Peru: II. Additional
hypothesis testing in quantitative ethnobotany. Economic Botany 47, 33-43.
Quinlan, M., 2005. Considerations for collecting freelists in the field: examples from
ethnobotany. Field Methods 17, 219-234.
Ruppert, J.C., Holm, A.M., Miehe, S., Muldavin, E., Snyman, H.A., Wesche, K., Linstädter,
A., 2012. Meta-analysis of rain-use efficiency confirms indicative value for degradation and
supports non-linear response along precipitation gradients in drylands. Journal of Vegetation
Science 23, 1035-1050.
Schrauf, R.W., Sanchez, J., 2010. Age effects and sample size in free listing. Field Methods
22, 70-87.
Scoones, I., 1999. Ecological dynamics and grazing-resource tenure: a case study from
Zimbabwe, in: Niamir-Fuller, M. (Ed.), Managing Mobility in African Rangelands. The
Legitimization of Transhumance. Intermediate Technology Publications Ltd, London, pp.
217-235.
Steele, M.Z., Shackleton, C.M., 2010. Using local experts as benchmarks for household local
ecological knowledge: Scoring in South African savannas. Journal of Environmental
Management 91, 1641-1646.
Sutrop, U., 2001. List task and a cognitive salience index. Field Methods 13, 263-276.
Szabó, A.T., 2004. Definitions, concepts and methods in the ethnobotany of food plants, in:
Macbeth, H., MacClancy, J. (Eds.), Researching Food Habits: Methods and Problems.
Berghahn, Oxford, UK, pp. 29-40.
Thomas, D.S.G., Twyman, C., 2004. Good or bad rangeland? Hybrid knowledge, science, and
local understandings of vegetation dynamics in the Kalahari. Land Degradation &
Development 15, 215-231.
Thomas, D.S.G., Twyman, C., Osbar, H., Hewitson, B., 2007. Adaptation to climate change
and variability: farmer responses to intra-seasonal precipitation trends in South Africa.
Climatic Change 83, 301-322.
Thompson, E.C., Juan, Z., 2006. Comparative cultural salience: Measures using free-list data.
Field Methods 18, 398-412.
Trabut, L., 2006. Noms Indigènes des Plantes d'Afrique du Nord. Ibis Press, Paris, France.
Wesuls, D., Lang, H., 2010. Perceptions and measurements: the assessment of pasture states
in a semi-arid area of Namibia. Human Ecology 38, 305-312.
Whitfield, S., Reed, M.S., 2012. Participatory environmental assessment in drylands:
Introducing a new approach. Journal of Arid Environments 77, 1-10.
Zemmrich, A., Manthey, M., Zerbe, S., Oyunchimeg, D., 2010. Driving environmental factors
and the role of grazing in grassland communities: A comparative study along an altitudinal
gradient in Western Mongolia. Journal of Arid Environments 74, 1271-1280.
Related Documents











