This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The identity of the cell adhesive protein substrate affects the efficiencyof adeno-associated virus reverse transduction
Kellie I. McConnell 1, Eric J. Gomez, Junghae Suh ⇑Department of Bioengineering, 6100 Main Street, MS-142, Houston, TX 77005, USA
a r t i c l e i n f o
Article history:Received 14 February 2012Received in revised form 25 June 2012Accepted 27 June 2012Available online 6 July 2012
Keywords:Adeno-associated virusReverse transductionGene deliveryProtein adsorptionMicropatterning
a b s t r a c t
Delivering genes from surfaces, called substrate-mediated gene delivery or reverse transduction, is a use-ful method to achieve spatial localization of gene delivery. We tested the compatibility of adeno-associ-ated virus (AAV) vectors with various cell adhesive proteins to mediate gene delivery from surfaces. Ourstudies demonstrate that AAV vectors can be successfully adsorbed on collagen I, elastin, and lamininsubstrates leading to robust gene delivery to overlying cells. Notably, AAV immobilization on lamininyields the highest efficiency of gene expression. This increased gene expression cannot be explained byincreases in the levels of virus deposition, transcriptional activity of cells, or virus vector uptake into cells.Further refinement of our knowledge of AAV interactions with extracellular matrix proteins may haveimportant implications in a variety of applications ranging from tissue engineering to in vivo genetherapy.
� 2012 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Incorporation of genetic factors into tissue engineering scaffoldsis a promising approach to developing functional, complex tissues[1]. The genetic factors, in the form of plasmids [2] or gene deliveryvectors [3], will affect gene expression profiles in the developingconstruct and may help guide the formation of appropriate cellphenotypes as well as the cell–cell contacts necessary to achieveproper tissue function [1]. Substrate-mediated gene delivery, alsoreferred to as reverse transfection (using plasmid DNA-based vec-tors) or reverse transduction (using virus vectors), is a method bywhich genetic factors are immobilized on the surface of a scaffoldfollowed by cell seeding on top of the factors [4,5]. The vectors re-lease from the substrate surface, resulting in gene delivery into theoverlying cells. Substrate-mediated gene delivery provides severalpotentially useful opportunities over conventional bulk gene deliv-ery, including: (i) the spatially constrained presentation of genes(leading to spatially defined gene expression profiles and mini-mized off-target effects); (ii) an increased concentration of genedelivery vectors in the cellular microenvironment (leading to in-creased transfection efficiencies with lower absolute amounts ofdelivery vectors, thereby decreasing the vector dose required);(iii) the ability to achieve sustained release of gene delivery vectors
over time (resulting in long-term gene expression) (reviewed inPannier and Shea [6]).
We are interested in investigating the use of adeno-associatedviruses (AAV) for substrate-mediated gene delivery applicationsfor a number of reasons. AAV is a non-pathogenic virus presentin humans and a number of other species [7]. Approximately 80%of the human population is estimated to have been exposed toAAV serotype 2 (AAV2) with no adverse effects [8]. Wild-typeAAV integrates site-specifically into chromosome 19 [9] suggestingthat this delivery vector can be used to integrate transgenes in ahighly controlled fashion [10] and minimize the insertional onco-genesis observed with other vectors, such as retroviruses [11]. Re-combinant AAV vectors, on the other hand, do not normally carrythe rep gene and therefore exhibit low integration efficiencies[12]. The AAV capsid, composed of 60 protein subunits and encaps-idating a 4.7 kb single-stranded DNA genome, is 25 nm in diameter[13] and is highly stable, being able to withstand a range of pH val-ues and with increased thermostability compared with otherviruses, such as adenovirus [14,15]. While AAV2 is known to trans-duce a number of cell types [7], additional rational and combinato-rial approaches can be used to increase transduction efficiency in adesired cell type [16–19].
AAV has already been tested in the context of reverse transduc-tion for several applications. AAV encoding VEGF or RANKL havebeen lyophilized directly onto bone allografts, resulting in positivein vivo murine bone repair and remodeling [20]. AAV2 capsids withgenetically inserted histidine tags have been attached to nickelsurfaces, allowing substrate-mediated gene delivery [21]. AAVhas also been successfully immobilized on elastin-like peptide
1742-7061/$ - see front matter � 2012 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.actbio.2012.06.038
⇑ Corresponding author. Tel.: +1 713 348 2853; fax: +1 713 348 5877.E-mail address: [email protected] (J. Suh).
1 Present address: Department of Nanomedicine, The Methodist Hospital ResearchInstitute, Houston, TX 77030, USA.
Acta Biomaterialia 8 (2012) 4073–4079
Contents lists available at SciVerse ScienceDirect
Acta Biomaterialia
journal homepage: www.elsevier .com/locate /actabiomat

Author's personal copy
matrices for increased gene delivery efficiency in fibroblasts andneural stem cells [22]. For high throughput microarray type appli-cations AAV has been immobilized on tissue culture plastic [23] oron nitrocellulose substrates [24], yielding robust reverse transduc-tion. Overall, AAV appears highly compatible with a number of dif-ferent experimental configurations for substrate-mediated genedelivery.
We have previously found that human fibronectin (HFN) is apromising substrate from which AAV-mediated reverse transduc-tion can occur [25]. An HFN substrate was shown to improve virusdeposition, resulting in enhanced gene delivery. In particular, thepresence of an HFN mediating layer led to a nearly 300% increasein virus deposition on –NH2 functionalized surfaces and a 7-foldincrease in gene delivery compared to virus adsorbed on uncoated–CH3 surfaces. HFN is an extracellular matrix protein containingthe cell adhesive sequence RGD (Arg–Gly–Asp). Fibronectin hasbeen used for a variety of tissue engineering applications, includingthe support of neurite out-growth [26] and vascular engineering[27]. This led us to question whether other cell adhesive proteinscan be used as substrates for AAV-mediated reverse transduction.The interaction of AAV with HFN and other types of extracellularmatrix (ECM) proteins has yet to be fully investigated. AAV isknown to use extracellular sugar moieties as primary cellularreceptors. For example, AAV2 binds heparan sulfate proteoglycans,[28] and AAV1 and AAV6 bind sialic acid [29]. The interaction ofAAV with other components in the ECM has so far received littleattention so it is unclear if any other ECM proteins can effectivelyimmobilize AAV vectors for reverse transduction.
In this study we investigated the ability of additional cell adhe-sive proteins to serve as the substrate in our approach. We ex-plored the use of collagen I, laminin, elastin, and poly-L-lysine fortheir ability to immobilize AAV vectors and allow for reverse trans-duction. We first adsorbed the proteins on gold surfaces using de-fined alkanethiol chemistries in case the way in which the proteinwas adsorbed affected the peptide domains displayed to AAV [30].After incubating the protein surfaces with AAV vectors we exam-ined virus deposition on the substrate and then seeded cells ontop to assay for reverse transduction. Interestingly, we found thatlaminin significantly increases the efficiency of AAV2-mediated re-verse transduction.
2. Materials and methods
2.1. Virus preparation
AAV2–GFP was prepared as described previously [25]. Virus ti-ters were determined using quantitative polymerase chain reac-tion (QPCR) with primers for the CMV promoter in the cassette.The virus used in this study was determined to have a concentra-tion of 1 � 1012 vector genomes ml�1 in 40% iodixanol.
2.2. PDMS stamp preparation and surface functionalization
PDMS stamps were prepared as described previously [25].PDMS stamps of 500 lm diameter circles with a 1.2 mm pitch werecleaned by sonication in 100% ethanol. Gold-coated glass coverslips(35 mm diameter, 100 Å gold over 20 Å titanium, Platypus, Madi-son, WI) were cleaned in dilute TL1 solution (6:1:1 H2O:NH4OH:-H2O2) for 1 min at 80 �C, rinsed twice in Millipore purified water(MPH2O), and dried with ultrapure nitrogen (N2). Alkanethiolswere diluted to 2 mM in 100% ethanol. Stamps were inked with–CH3 (1-hexadecanethiol), –COOH (16-mercaptohexadecanoicacid), or –NH2 (11-amino-1-undecanethiol hydrochloride) termi-nated alkanethiol, all from Sigma Aldrich (St Louis, MO). Patternswere created by inking the PDMS stamps, placing them gently
on the substrate for 20 s, and then carefully removing them.Coverslips were then incubated with 2 mM oligo(ethylene glycol)(–OEG) terminated alkanethiol (HSC11-EG6, Prochimia, Poland) for1 h to passivate the background. Samples were rinsed with 100%ethanol and dried with N2.
2.3. Protein attachment
Functionalized coverslips were incubated with 1 ml of proteinwarmed to 37 �C diluted in phosphate-buffered saline (PBS)pH 7.0 for 30 min. HFN (Sigma Aldrich, Franklin Lakes, NJ) wasused at a concentration of 25 lg ml�1. Bovine collagen I (BectonDickinson) was used at a concentration of 50 lg ml�1. Human lam-inin (Sigma Aldrich) was used at a concentration of 25 lg ml�1.Soluble elastin was obtained from Spectrum Chemical (Gardena,CA) and was used at a concentration of 100 lg ml�1. Poly-L-lysine(PLL) from Sigma Aldrich was used at a concentration of50 lg ml�1. Coverslips were rinsed twice with 5 ml of PBS usinga platform rocker. Virus was incubated with protein pre-adsorbedcoverslips for 30 min at room temperature. Samples were againrinsed twice with PBS.
2.4. Immunostaining
Samples were blocked with 1% bovine serum albumin (BSA) inPBS for 1 h and rinsed twice with PBS. Patterns were incubatedwith A20 antibody (mouse anti-AAV2 capsid, American ResearchProducts, Belmont, MA) diluted 1:200 in PBS overnight at 4 �C.Samples were rinsed with PBS and incubated with goat anti-mouseAlexaFluor532-conjugated antibody (Invitrogen) diluted 1:100 inPBS for 1 h at room temperature. Coverslips were rinsed withPBS and mounted using Fluromount (SouthernBiotech, Birming-ham, AL).
2.5. Cell studies
Coverslips were placed in 60 mm tissue culture dishes and HeLacells were seeded at a density of 100 cells mm�2. Cells were seededin Dulbecco’s modified Eagle medium (Invitrogen) supplementedwith 1% penicillin/streptomycin (Invitrogen). After 8 h the mediumwas exchanged for Dulbecco’s modified Eagle medium supple-mented with 1% penicillin/streptomycin and 10% fetal bovine ser-um (Invitrogen). Cells were maintained at 37 �C with 5% CO2. After72 h the cells were fixed in 4% paraformaldehyde in PBS andstained with 2 lg ml�1 Hoechst 33342 stain (Invitrogen). Cover-slips were mounted using Fluoromount. For Bromouridine (BrU)studies cells were incubated with 2 mM BrU (Sigma Aldrich) for1 h and permeabilized with 0.25% Triton X-100 in PBS. Cells wereblocked with bovine serum albumin BSA and rinsed with PBS. Pri-mary anti-BrU antibody (Sigma Aldrich) and secondary antibody(goat anti-mouse AF532-conjugated, Invitrogen) were incubatedfor 1 h each at 1:200 dilution in PBS. Samples were mounted usingFluoromount.
For virus uptake studies, -CH3-coated surfaces were prepared byincubating 2 mM alkanethiol solutions with cleaned surfaces for1 h. Samples were rinsed with ethanol and dried with ultrapurenitrogen. Samples were incubated with HFN, collagen I, laminin,or elastin for 30 min at room temperature, rinsed with PBS, andthen incubated with AAV2–GFP for 30 min. Substrates were thenrinsed with PBS. Samples were seeded with HeLa cells and har-vested 24 h after seeding. Cells were lysed and the internalizedgenomes were quantified using QPCR. Internalized genomes werenormalized to the total protein content of the well, as determinedby the DC protein assay (Bio-Rad, Hercules, CA).
4074 K.I. McConnell et al. / Acta Biomaterialia 8 (2012) 4073–4079
Author's personal copy
2.6. Imaging and image analysis
Images were acquired using a Zeiss LSM 5 LIVE confocal micro-scope (Carl Zeiss, Munich, Germany). Images within a given sampleset (immunostaining, GFP expression, Hoechst staining, BrU analy-sis) were acquired under identical conditions. Virus immunostain-ing images were collected using 532 nm excitation at 50% powerand data was collected for a 2 ms pixel dwell time using a band-pass filter between 550 and 600 nm. GFP images were obtainedusing 489 nm excitation at 1.8% power and data was collected fora 2 ms pixel dwell time with a bandpass filter between 500 and525 nm. BrU immunostaining images were collected using532 nm excitation at 3% power and data was collected for a 2 mspixel dwell time using a bandpass filter between 550 and600 nm. To quantify virus attachment 10 images were taken andintegrated densities were measured with ImageJ (NIH, Bethesda,MD). For cellular studies, 10 fluorescence and DIC images were ta-ken of each sample (five images from two independent replicates).For BrU analysis the integrated densities of 20 cell nuclei per sam-ple were determined. Data are given as means and standard errorsof the mean. Statistical testing was performed using JMP 9. Signif-icance was determined using ANOVA and post-hoc testing usingTukey’s HSD with p < 0.05.
3. Results
3.1. AAV2 can be immobilized on a variety of cell adhesive proteins
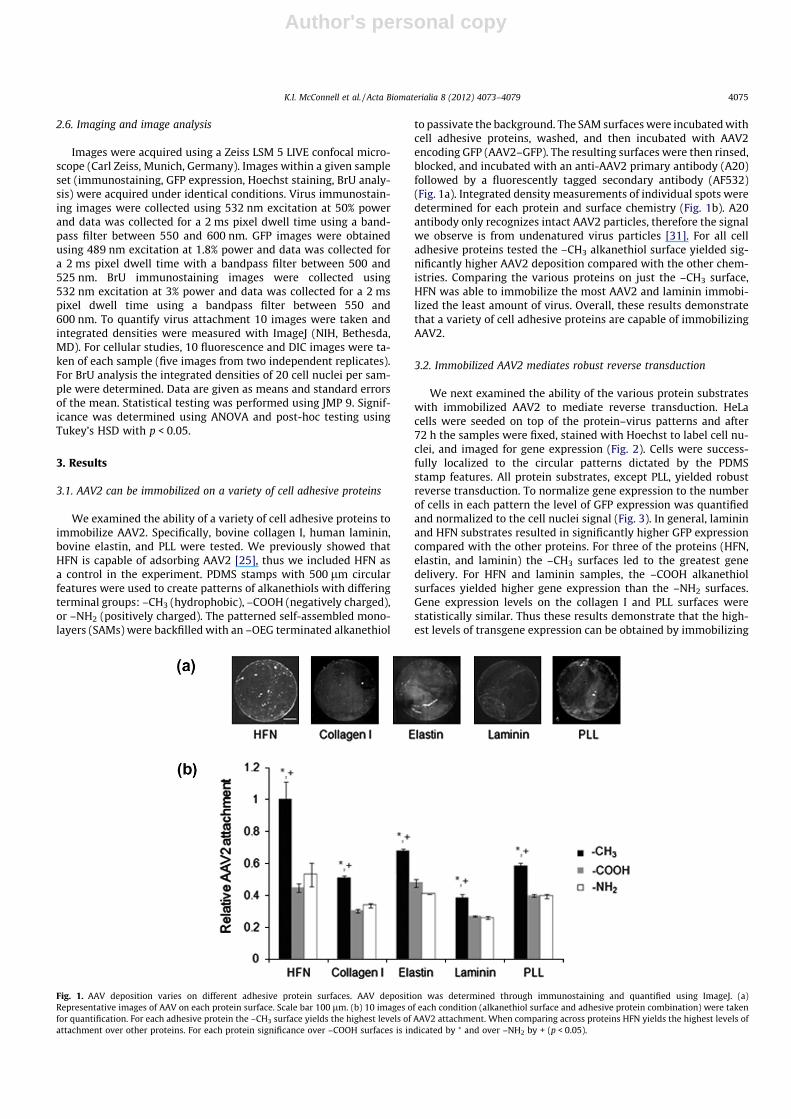
We examined the ability of a variety of cell adhesive proteins toimmobilize AAV2. Specifically, bovine collagen I, human laminin,bovine elastin, and PLL were tested. We previously showed thatHFN is capable of adsorbing AAV2 [25], thus we included HFN asa control in the experiment. PDMS stamps with 500 lm circularfeatures were used to create patterns of alkanethiols with differingterminal groups: –CH3 (hydrophobic), –COOH (negatively charged),or –NH2 (positively charged). The patterned self-assembled mono-layers (SAMs) were backfilled with an –OEG terminated alkanethiol
to passivate the background. The SAM surfaces were incubated withcell adhesive proteins, washed, and then incubated with AAV2encoding GFP (AAV2–GFP). The resulting surfaces were then rinsed,blocked, and incubated with an anti-AAV2 primary antibody (A20)followed by a fluorescently tagged secondary antibody (AF532)(Fig. 1a). Integrated density measurements of individual spots weredetermined for each protein and surface chemistry (Fig. 1b). A20antibody only recognizes intact AAV2 particles, therefore the signalwe observe is from undenatured virus particles [31]. For all celladhesive proteins tested the –CH3 alkanethiol surface yielded sig-nificantly higher AAV2 deposition compared with the other chem-istries. Comparing the various proteins on just the –CH3 surface,HFN was able to immobilize the most AAV2 and laminin immobi-lized the least amount of virus. Overall, these results demonstratethat a variety of cell adhesive proteins are capable of immobilizingAAV2.
3.2. Immobilized AAV2 mediates robust reverse transduction
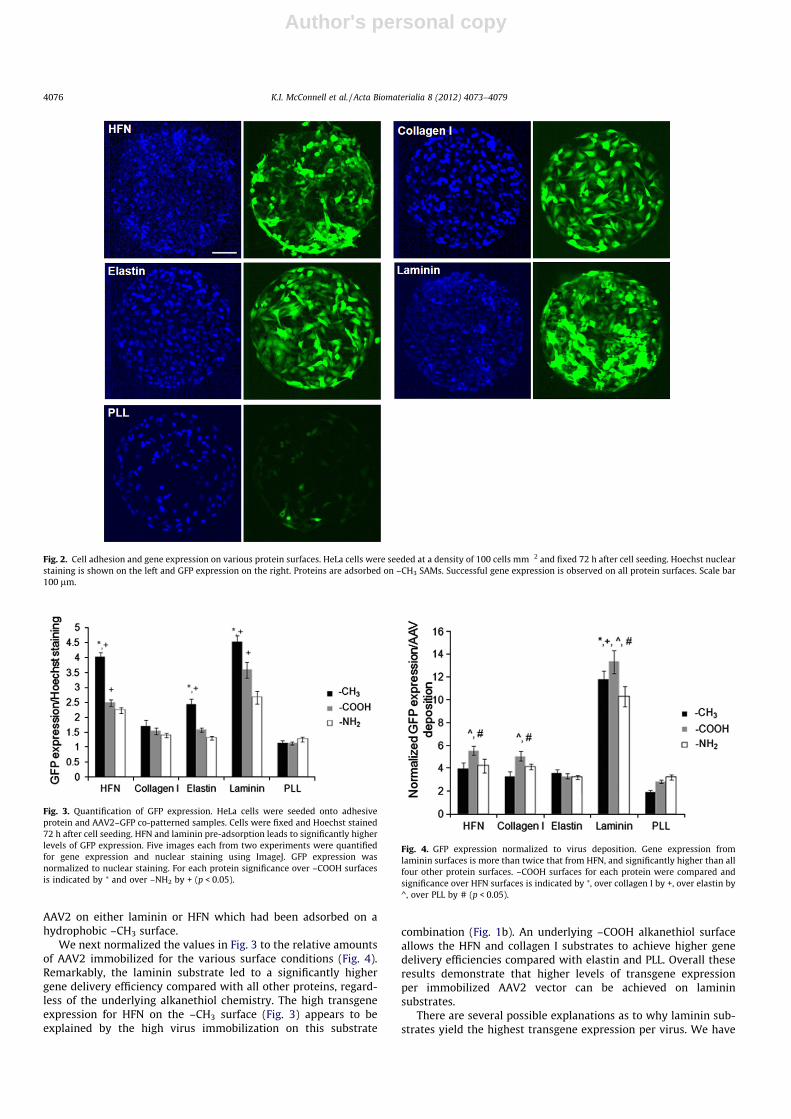
We next examined the ability of the various protein substrateswith immobilized AAV2 to mediate reverse transduction. HeLacells were seeded on top of the protein–virus patterns and after72 h the samples were fixed, stained with Hoechst to label cell nu-clei, and imaged for gene expression (Fig. 2). Cells were success-fully localized to the circular patterns dictated by the PDMSstamp features. All protein substrates, except PLL, yielded robustreverse transduction. To normalize gene expression to the numberof cells in each pattern the level of GFP expression was quantifiedand normalized to the cell nuclei signal (Fig. 3). In general, lamininand HFN substrates resulted in significantly higher GFP expressioncompared with the other proteins. For three of the proteins (HFN,elastin, and laminin) the –CH3 surfaces led to the greatest genedelivery. For HFN and laminin samples, the –COOH alkanethiolsurfaces yielded higher gene expression than the –NH2 surfaces.Gene expression levels on the collagen I and PLL surfaces werestatistically similar. Thus these results demonstrate that the high-est levels of transgene expression can be obtained by immobilizing
Fig. 1. AAV deposition varies on different adhesive protein surfaces. AAV deposition was determined through immunostaining and quantified using ImageJ. (a)Representative images of AAV on each protein surface. Scale bar 100 lm. (b) 10 images of each condition (alkanethiol surface and adhesive protein combination) were takenfor quantification. For each adhesive protein the –CH3 surface yields the highest levels of AAV2 attachment. When comparing across proteins HFN yields the highest levels ofattachment over other proteins. For each protein significance over –COOH surfaces is indicated by ⁄ and over –NH2 by + (p < 0.05).
K.I. McConnell et al. / Acta Biomaterialia 8 (2012) 4073–4079 4075
Author's personal copy
AAV2 on either laminin or HFN which had been adsorbed on ahydrophobic –CH3 surface.
We next normalized the values in Fig. 3 to the relative amountsof AAV2 immobilized for the various surface conditions (Fig. 4).Remarkably, the laminin substrate led to a significantly highergene delivery efficiency compared with all other proteins, regard-less of the underlying alkanethiol chemistry. The high transgeneexpression for HFN on the –CH3 surface (Fig. 3) appears to beexplained by the high virus immobilization on this substrate
combination (Fig. 1b). An underlying –COOH alkanethiol surfaceallows the HFN and collagen I substrates to achieve higher genedelivery efficiencies compared with elastin and PLL. Overall theseresults demonstrate that higher levels of transgene expressionper immobilized AAV2 vector can be achieved on lamininsubstrates.
There are several possible explanations as to why laminin sub-strates yield the highest transgene expression per virus. We have
Fig. 2. Cell adhesion and gene expression on various protein surfaces. HeLa cells were seeded at a density of 100 cells mm�2 and fixed 72 h after cell seeding. Hoechst nuclearstaining is shown on the left and GFP expression on the right. Proteins are adsorbed on –CH3 SAMs. Successful gene expression is observed on all protein surfaces. Scale bar100 lm.
Fig. 3. Quantification of GFP expression. HeLa cells were seeded onto adhesiveprotein and AAV2–GFP co-patterned samples. Cells were fixed and Hoechst stained72 h after cell seeding. HFN and laminin pre-adsorption leads to significantly higherlevels of GFP expression. Five images each from two experiments were quantifiedfor gene expression and nuclear staining using ImageJ. GFP expression wasnormalized to nuclear staining. For each protein significance over –COOH surfacesis indicated by ⁄ and over –NH2 by + (p < 0.05).
Fig. 4. GFP expression normalized to virus deposition. Gene expression fromlaminin surfaces is more than twice that from HFN, and significantly higher than allfour other protein surfaces. –COOH surfaces for each protein were compared andsignificance over HFN surfaces is indicated by ⁄, over collagen I by +, over elastin by^, over PLL by # (p < 0.05).
4076 K.I. McConnell et al. / Acta Biomaterialia 8 (2012) 4073–4079

Author's personal copy
investigated two potential reasons: (1) cells may be more trans-criptionally active when seeded on laminin leading to greaterexpression of all genes, including the transgene; (2) AAV2 maybe more efficiently internalized from laminin, making this sub-strate optimal for reverse transduction.
3.3. Variabilities in overall transcriptional activity do not explain genedelivery differences
We compared the relative transcriptional activities of cellsseeded on the different proteins by measuring BrU incorporation(Fig. 5). We observed differences in transcriptional activity on dif-ferent protein substrates, with cells being nearly twice as active onPLL–COOH surfaces compared with HFN–CH3 surfaces. Notably,transcriptional activity on laminin–CH3 surfaces is not significantly
higher than the activity on any of the other protein–CH3 surfaces.Overall, this result suggests that the increase in gene delivery effi-ciency observed on laminin is not due to increased transcriptionalactivity of cells seeded on this protein substrate. Other AAV2–lam-inin and/or more complex cell–laminin interactions may explainthe observation.
3.4. Variabilities in vector internalization do not explain gene deliverydifferences
We also investigated the uptake of virus capsids into HeLa cells.Gold substrates were –CH3 functionalized and sequentially incu-bated with protein and AAV2–GFP. 24 h after seeding the cellswere collected and lysed, and internalized genomes were quanti-fied using QPCR. The number of genomes was normalized to the to-tal protein content of the wells using the DC protein assay (Fig. 6).Interestingly, we did not observe a correlation between the num-ber of internalized genomes (Fig. 6) and transduction efficiency(Figs. 3 and 4), as the highest levels of internalization were ob-served on elastin and collagen I. Overall, internalization of highnumbers of vector alone does not appear to yield increased levelsof transduction, indicating that additional more complex mecha-nisms may explain the observed transduction results.
4. Discussion
We have investigated the immobilization of AAV2 gene deliveryvectors on various ECM proteins for possible use in substrate-med-iated gene delivery applications. Our results indicate that AAV2 iscapable of interacting with a variety of cell adhesive proteins,including collagen I, elastin, and laminin (Fig. 1). We have previ-ously reported on the ability of HFN to immobilize AAV2 [25]. Alltested protein surfaces were able to mediate cell adhesion in ser-um-free medium (which was replaced with complete medium8 h after cell seeding), as well as reverse transduction with AAV2
Fig. 5. Relative transcriptional activity of HeLa cells seeded on various adsorbed proteins. HeLa cells were seeded onto adhesive protein adsorbed onto SAMs. (a) Cells weretreated with BrU 24 h later and immunostained to observe BrU incorporation. Scale bar 100 lm. (b) Integrated densities of 20 cell nuclei per condition were quantified. Valueswere normalized to the HFN–CH3 surface. The cases were significantly different by ANOVA (p < 0.05).
Fig. 6. Cellular uptake of virus vectors from adhesive protein surfaces. Virus wasadsorbed to protein (HFN, collagen, elastin, or laminin) on a –CH3 functionalizedsurface. Substrates were then seeded with HeLa cells. Cells were collected and lysed24 h after cell seeding. Virus genomes contained within cell lysates were quantifiedusing QPCR and normalized to total protein content. There was no statisticallysignificant difference as determined by ANOVA.
K.I. McConnell et al. / Acta Biomaterialia 8 (2012) 4073–4079 4077

Author's personal copy
(Fig. 2), suggesting that this virus vector may be compatible with arange of experimental configurations involving different types ofcell adhesive proteins. Interestingly, laminin surfaces are most effi-cient for AAV2 reverse transduction of HeLa cells (Fig. 4). However,studies on COS-7 cells did not yield the same results (Supplemen-tary Fig. 1), suggesting that the observed phenomenon is cell-typedependent.
We investigated two possible reasons for laminin surfacesyielding the highest reverse transduction. First, we postulated thatthe dramatic increase in transduction efficiency could be due to in-creased transcriptional activity of HeLa cells on laminin. ECM sub-strates have been shown to affect cellular behavior; for example, ithas been reported that seeding endothelial cells on collagen Irather than laminin leads to changes in gene activation that affectcell morphogenesis [32]. In our case, BrU studies indicated that thecells under our experimental conditions were no more transcrip-tionally active on laminin surfaces as compared with the other pro-teins (Fig. 5), suggesting that other mechanisms must lead to theobserved results. Thus we next hypothesized that favorable AAV2uptake profiles from laminin surfaces yield increased reversetransduction. However, increased vector internalization alone doesnot appear to yield higher transduction profiles (Fig. 6). Futurestudies utilizing more quantitative analytical techniques willreveal more in-depth information about the on–off bindingkinetics of AAV with the various proteins. One hypothesis we havenot yet investigated is that the intracellular processing of AAV2may be optimal in cells seeded on laminin, which may lead togreater numbers of transgenes being delivered to the cell nucleus.Additional experiments to test this hypothesis may reveal newinsights.
By testing different surface chemistries provided by the pat-terned SAMs we endeavored to determine how different experi-mental configurations affected reverse transduction efficiencies.The rationale behind the differently terminated alkanethiols wasthat certain chemical surfaces may lead to more optimal presenta-tion of virus-attractive peptide motifs present on the ECM proteins,leading to higher virus immobilization and improved gene deliv-ery. Others have reported that, depending on the underlyingSAM, different peptide motifs are displayed by HFN [33]. For lam-inin, HFN, and collagen I an underlying –COOH SAM yields slightlyimproved transgene expression per immobilized virus vector(Fig. 4). Further studies on protein configurations on these variousSAMs are needed to better understand these observations.
Other viruses have been reported to interact with ECM proteins.Specifically, retrovirus is capable of associating with a heparin-binding domain on fibronectin [34], an interaction that can beblocked by the addition of heparin [35]. This retrovirus–fibronectininteraction has been harnessed to create an efficient commerciallyavailable reverse transduction platform called RetroNectin (TakaraBio Inc.). Plates coated with RetroNectin can be incubated with ret-rovirus or lentivirus and then seeded with cells to increase virus–cell interaction and increase gene delivery into cells [36,37].
The interaction of AAV gene delivery vectors with ECM proteinsis an important property that can potentially be harnessed for sub-strate-mediated gene delivery applications in tissue engineering,regenerative medicine, and biomedical implants. For example,ECM proteins are used for wound healing applications [38]. Incor-poration of AAV vectors into such technologies may allow geneticcues to aid in the proper orchestration of healing factors. ECM com-ponents have been used for a variety of applications in tissue engi-neering, including the creation of scaffolds for cardiovasculartissue engineering [39], the development of bone microenviron-ments [40], and as coatings for substrate-mediated gene deliveryin spinal cord therapies [41]. Ultimately, by choosing the rightcombination of AAV serotype, transgene, and ECM componentsthe desired therapeutic outcome may be achieved.
Furthermore, our studies have important implications forin vivo gene therapy. As AAV vectors move to the target tissuesand cells, they must negotiate the extracellular environment com-posed of different ECM proteins. Adverse binding of vectors to cer-tain extracellular components may critically decrease the overallgene delivery efficiency. Currently, little is known about the inter-actions between AAV vectors and ECM proteins, although thein vivo importance of these interactions has been suggested previ-ously in a study in which co-injection of heparin with AAV2 vectorsinto the brain allowed the vectors to travel greater distances [42].It has also been shown that retinal transduction is increased ondigestion of the inner limiting membrane (ILM) by protease [43].The ILM contains a variety of ECM proteins, including laminin-1,collagen 4, collagen 18, and perlecan [44]. AAV5 is unable to deli-ver genes efficiently to the inner retinal cells unless this membraneis digested [43]. Notably, the authors suggested that this was dueto a lack of AAV5 binding to the ILM, causing AAV5 to remain dif-fuse within the surrounding area. An improved understanding ofAAV–ECM protein interactions will provide us with a more ‘‘sys-tems approach’’ picture of how AAV vectors are transportedthrough various biological barriers to reach their target cells, andthe affinity of vectors for cell surface receptors is relegated to beingjust one component of the overall gene delivery vector design.
5. Conclusions
We have demonstrated that AAV2 is capable of reverse trans-duction from a variety of ECM protein surfaces. We observed highlevels of gene expression on HFN and laminin surfaces. Most inter-estingly, when normalized to total virus deposition, we observedramatically higher gene expression on laminin surfaces. Thisexpression cannot be explained by increased cell transcriptionalactivity or vector internalization, suggesting that other more com-plex biological mechanisms may be responsible. Further investiga-tions into the interaction of AAV and ECM proteins could lead toimportant insights that may be useful for applications rangingfrom regenerative medicine to in vivo gene therapy.
Disclosures
The authors acknowledge that they have no conflicts of interestto disclose.
Acknowledgements
This material is based upon work supported by the National Sci-ence Foundation under grant no. 0955536 and a National ScienceFoundation Graduate Research Fellowship to K.I.M. The authorswould like to thank Arum Han and Jaewon Park (Texas A&M Uni-versity) for PDMS stamp fabrication.
Appendix A. Figures with essential colour discrimination
Certain figures in this article, particularly Fig. 2, are difficult tointerpret in black and white. The full colour images can be foundin the on-line version, at http://dx.doi.org/10.1016/j.actbio.2012.06.038.
Appendix B. Supplementary data
Supplementary data associated with this article can be found,in the online version, at http://dx.doi.org/10.1016/j.actbio.2012.06.038.
4078 K.I. McConnell et al. / Acta Biomaterialia 8 (2012) 4073–4079

Author's personal copy
References
[1] Discher DE, Mooney DJ, Zandstra PW. Growth factors, matrices, and forcescombine and control stem cells. Science 2009;324:1673.
[2] Houchin-Ray T, Whittlesey KJ, Shea LD. Spatially patterned gene delivery forlocalized neuron survival and neurite extension. Mol Ther 2007;15:705.
[3] Gersbach CA, Coyer SR, Le Doux JM, Garcia AJ. Biomaterial-mediatedretroviral gene transfer using self-assembled monolayers. Biomaterials2007;28:5121.
[4] Shea LD, Smiley E, Bonadio J, Mooney DJ. DNA delivery from polymer matricesfor tissue engineering. Nat Biotechnol 1999;17:551.
[5] Ziauddin J, Sabatini DM. Microarrays of cells expressing defined cDNAs. Nature2001;411:107.
[6] Pannier AK, Shea LD. Controlled release systems for DNA delivery. Mol Ther2004;10:19.
[7] Wu Z, Asokan A, Samulski RJ. Adeno-associated virus serotypes: vector tool kitfor human gene therapy. Mol Ther 2006;14:316.
[8] Moskalenko M, Chen L, van Roey M, Donahue BA, Snyder RO, McArthur JG,et al. Epitope mapping of human anti-adeno-associated virus type 2neutralizing antibodies: implications for gene therapy and virus structure. JVirol 2000;74:1761.
[9] Samulski RJ, Zhu X, Xiao X, Brook JD, Housman DE, Epstein N, et al. Targetedintegration of adeno-associated virus (AAV) into human chromosome 19.EMBO J 1991;10:3941.
[10] Weitzman MD, Young Jr SM, Cathomen T, Samulski RJ. Targeted integration byadeno-associated virus. Methods Mol Med 2003;76:201.
[11] Nair V. Retrovirus-induced oncogenesis and safety of retroviral vectors. CurrOpin Mol Ther 2008;10:431.
[12] Philpott NJ, Giraud-Wali C, Dupuis C, Gomos J, Hamilton H, Berns KI, et al.Efficient integration of recombinant adeno-associated virus DNA vectorsrequires a p5-rep sequence in cis. J Virol 2002;76:5411.
[13] Xie Q, Bu W, Bhatia S, Hare J, Somasundaram T, Azzi A, et al. The atomicstructure of adeno-associated virus (AAV-2), a vector for human gene therapy.Proc Natl Acad Sci USA 2002;99:10405.
[14] Wallis C, Yang CS, Melnick JL. Effect of cations on thermal inactivation ofvaccinia, herpes simplex, and adenoviruses. J Immunol 1962;89:41.
[15] Kronenberg S, Bottcher B, von der Lieth CW, Bleker S, Kleinschmidt JA. Aconformational change in the adeno-associated virus type 2 capsid leads to theexposure of hidden VP1 N termini. J Virol 2005;79:5296.
[16] Shi W, Bartlett JS. RGD inclusion in VP3 provides adeno-associated virus type 2(AAV2)-based vectors with a heparan sulfate-independent cell entrymechanism. Mol Ther 2003;7:515.
[17] Maheshri N, Koerber JT, Kaspar BK, Schaffer DV. Directed evolution of adeno-associated virus yields enhanced gene delivery vectors. Nat Biotechnol2006;24:198.
[18] Li W, Asokan A, Wu Z, Van Dyke T, Diprimio N, J SJ, et al. Engineering andselection of shuffled AAV genomes: a new strategy for producing targetedbiological nanoparticles. Mol Ther 2008.
[19] Muller OJ, Kaul F, Weitzman MD, Pasqualini R, Arap W, Kleinschmidt JA, et al.Random peptide libraries displayed on adeno-associated virus to select fortargeted gene therapy vectors. Nat Biotechnol 2003;21:1040.
[20] Ito H, Koefoed M, Tiyapatanaputi P, Gromov K, Goater JJ, Carmouche J, et al.Remodeling of cortical bone allografts mediated by adherent rAAV-RANKL andVEGF gene therapy. Nat Med 2005;11:291.
[21] Jang JH, Koerber JT, Gujraty K, Bethi SR, Kane RS, Schaffer DV. Surfaceimmobilization of hexa-histidine-tagged adeno-associated viral vectors forlocalized gene delivery. Gene Ther 2010;17:1384.
[22] Kim JS, Chu HS, Park KI, Won JI, Jang JH. Elastin-like polypeptide matrices forenhancing adeno-associated virus-mediated gene delivery to human neuralstem cells. Gene Ther 2011;19:329.
[23] Dong X, Tian W, Wang G, Dong Z, Shen W, Zheng G, et al. Establishment of anAAV reverse infection-based array. PLoS One 2010;5:e13479.
[24] McConnell KI, Schweller RM, Diehl MR, Suh J. Live-cell microarray surfacecoatings supporting reverse transduction by adeno-associated viruses.Biotechniques 2011;51:255.
[25] McConnell KI, Slater JH, Han A, West JL, Suh J. Microcontact printing for co-patterning cells and viruses for spatially controlled substrate-mediated genedelivery. Soft Matter 2011;7:4993.
[26] Mukhatyar VJ, Salmeron-Sanchez M, Rudra S, Mukhopadaya S, Barker TH,Garcia AJ, et al. Role of fibronectin in topographical guidance of neuriteextension on electrospun fibers. Biomaterials 2011;32:3958.
[27] Dubey G, Mequanint K. Conjugation of fibronectin onto three-dimensionalporous scaffolds for vascular tissue engineering applications. Acta Biomater2011;7:1114.
[28] Opie SR, Warrington Jr KH, Agbandje-McKenna M, Zolotukhin S, Muzyczka N.Identification of amino acid residues in the capsid proteins of adeno-associated virus type 2 that contribute to heparan sulfate proteoglycanbinding. J Virol 2003;77:6995.
[29] Wu Z, Asokan A, Grieger JC, Govindasamy L, Agbandje-McKenna M, SamulskiRJ. Single amino acid changes can influence titer, heparin binding, and tissuetropism in different adeno-associated virus serotypes. J Virol 2006;80:11393.
[30] Keselowsky BG, Collard DM, Garcia AJ. Surface chemistry modulatesfibronectin conformation and directs integrin binding and specificity tocontrol cell adhesion. J Biomed Mater Res 2003;66:247.
[31] Wistuba A, Kern A, Weger S, Grimm D, Kleinschmidt JA. Subcellularcompartmentalization of adeno-associated virus type 2 assembly. J Virol1997;71:1341.
[32] Liu Y, Senger DR. Matrix-specific activation of Src and Rho initiates capillarymorphogenesis of endothelial cells. FASEB J 2004;18:457.
[33] Lan MA, Gersbach CA, Michael KE, Keselowsky BG, Garcia AJ. Myoblastproliferation and differentiation on fibronectin-coated self assembledmonolayers presenting different surface chemistries. Biomaterials 2005;26:4523.
[34] Hanenberg H, Xiao XL, Dilloo D, Hashino K, Kato I, Williams DA. Colocalizationof retrovirus and target cells on specific fibronectin fragments increasesgenetic transduction of mammalian cells. Nat Med 1996;2:876.
[35] Carstanjen D, Dutt P, Moritz T. Heparin inhibits retrovirus binding tofibronectin as well as retrovirus gene transfer on fibronectin fragments. JVirol 2001;75:6218.
[36] Shin S, Salvay DM, Shea LD. Lentivirus delivery by adsorption to tissueengineering scaffolds. J Biomed Mater Res A 2010;93:1252.
[37] Lee HJ, Lee YS, Kim HS, Kim YK, Kim JH, Jeon SH, et al. Retronectin enhanceslentivirus-mediated gene delivery into hematopoietic progenitor cells.Biologicals 2009;37:203.
[38] Bello YM, Falabella AF, Eaglstein WH. Tissue-engineered skin. Current status inwound healing. Am J Clin Dermatol 2001;2:305.
[39] Heydarkhan-Hagvall S, Schenke-Layland K, Dhanasopon AP, Rofail F, Smith H,Wu BM, et al. Three-dimensional electrospun ECM-based hybrid scaffolds forcardiovascular tissue engineering. Biomaterials 2008;29:2907.
[40] Kang Y, Kim S, Khademhosseini A, Yang Y. Creation of bony microenvironmentwith CaP and cell-derived ECM to enhance human bone-marrow MSC behaviorand delivery of BMP-2. Biomaterials 2011;32:6119.
[41] De Laporte L, Yan AL, Shea LD. Local gene delivery from ECM-coatedpoly(lactide-co-glycolide) multiple channel bridges after spinal cord injury.Biomaterials 2009;30:2361.
[42] Nguyen JB, Sanchez-Pernaute R, Cunningham J, Bankiewicz KS. Convection-enhanced delivery of AAV-2 combined with heparin increases TK gene transferin the rat brain. Neuroreport 2001;12:1961.
[43] Dalkara D, Kolstad KD, Caporale N, Visel M, Klimczak RR, Schaffer DV, et al.Inner limiting membrane barriers to AAV-mediated retinal transduction fromthe vitreous. Mol Ther 2009;17:2096.
[44] Candiello J, Balasubramani M, Schreiber EM, Cole GJ, Mayer U, Halfter W, et al.Biomechanical properties of native basement membranes. FEBS J 2007;274:2897.
K.I. McConnell et al. / Acta Biomaterialia 8 (2012) 4073–4079 4079
Related Documents






![[VII]. Regulation of Gene Expression Via Signal Transduction Reading List VII: Signal transduction Signal transduction in biological systems.](https://static.cupdf.com/doc/110x72/56649e385503460f94b28319/vii-regulation-of-gene-expression-via-signal-transduction-reading-list-vii.jpg)




