Photochemistry and Photobiology, 1999, 69(6): 686-693 The Human Melanocyte as a Particular Target for UVA Radiation and an Endpoint for Photoprotection Assessment Laurent Marrot*, Jean-Philippe Belaidi, Jean-Roch Meunier, Philippe Perez and Catherine Agapakis-Causse L‘Oreal Advanced Research, Life Sciences Research, Aulnay-Sous-Bois, France Received 4 January 1999; accepted 5 March 1999 ABSTRACT The induction of DNA breaks by UVA (320-400 nm) in the nucleus of normal human melanocytes in culture was investigated using single cell gel electrophoresis, also called the comet assay. Endogenous pigment and/or mel- anin-related molecules were found to enhance DNA breakage: comets were more intense in melanocytes than in fibroblasts, in cells with high melanin content or after stimulation of melanogenesis by supplying tyrosine in the culture medium. After UVA doses where strong comets were observed, neither cytotoxicity nor stimulation of ty- rosinase activity were detected. However, the accumula- tion of p53 protein suggested that cells reacted to geno- toxic stress under these experimental conditions. The same approach was used to compare two sunscreens with identical sun protection factors but different UVA pro- tection factors. The results presented in this paper sug- gest that human melanocytes may be used as a target cell to evidence broadspectrum photoprotection. Moreover, these data appear to be helpful in getting a better un- derstanding of the role of sunlight in the initiating steps of melanocyte transformation. INTRODUCTION Although the role of UV radiation in the etiology of non- melanoma skin cancer is now well established (1,2), the re- lationship between melanoma incidence and sunlight expo- sure is still a matter of debate. Epidemiological studies show strong evidence for increased risk related to sunburn, but the connection with total sun exposure is unclear (3). Experi- mental studies are limited by the lack of suitable animal models. Commonly used animals, such as mice, are not sus- ceptible to melanoma induction upon exposure to UV alone without combined treatment by chemical carcinogens (4). However, chronic UVB exposure of the South American opossum (Monoddphis durnesfica) results in the formation of cutaneous melanoma (S), and UVA was shown to be ef- fective in stimulating melanocytic hyperplasia in the skin of this animal (6). An action spectrum for the induction of mel- anoma has also been determined in a fish model where a role for UVA and visible radiation was reported (7). Contrary to nonmelanoma skin cancer, the mechanisms involved in the initiating steps of melanoma are not clear. For instance, the role of the p53 tumor suppressor gene is well documented in squamous cell carcinoma (SCC)t where p53 is mutated with a frequency of about 90% or in basal cell carcinoma (BCC) with a frequency of about SO%, and the mechanisms that lead to skin tumors have been described in detail (8,9). In the case of melanoma, only a few studies about mutations have been published and many of them have concluded that pS3 is unlikely to play a major role (10-12). Another possible tumor suppressor gene, the p16CDKN”1NK4 gene, is also suspected to be involved in melanoma suscep- tibility: its deletion andor mutation has been reported, but here again, diverging results were obtained (13-15). Muta- tions in the N-ras oncogene have also been described in pa- tients with cutaneous melanoma, and the involvement of UV-induced dipyrimidine sites was suggested (16). The role of DNA damage in the development of mela- noma is highly probable. Patients with the DNA repair de- ficiency disease xeroderma pigmentosum (XP) are several hundred times more susceptible to melanoma. In this case, the anatomic distribution of tumors is not the same as in carcinomas, suggesting differences in the mechanisms of in- duction (17). Furthermore, when the p53 gene is mutated in melanoma, mutations known as UV signatures are predom- inant and they might result from pyrimidine dimers induced by sunlight in the genome of melanocytes (18). On the other hand, the role of melanin in photoprotection, if any, can also be questioned. It is well established that individuals with dark skin are less susceptible to sun-induced skin cancer than those with fair skin (19). However, recent studies have shown that epidermal tanning is not always ef- fective for preventing skin cancer in the mouse model (20) and that UV-induced melanogenesis in skin types I1 or 111 offers only a moderate protection against erythema (21). ”To whom correspondence hhould be adressed at: L’Oreal Advanced Research. Life Science Research. Investigative Toxicology De- partment. Genotoxicity Group. I. avenue Eugene Schueller, BP22. 93601 Aulnay-Sous-Bois cedex, France. Fax: 33.1.48.68.91.80: e-mail: [email protected] 1999 American Societ! for Photobiology 003 I -8655/99 $5.00+0.00 tAhhreviations: BCC, basal cell carcinoma: DOPA, dihydroxyphe- nylalanine: MSH, melanocyte-stimulating hormone; NHEM, nor- mal human melanocyte: PAGE, polyacrylamide gel electrophoresis: PBS, phosphate-buffered saline: SCC, squamous cell carcinoma: SDS, sodium dodecyl sulfate: SPF, sun protection factor: XP, xe- roderma pigmentosum. 686

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Photochemistry and Photobiology, 1999, 69(6): 686-693
The Human Melanocyte as a Particular Target for UVA Radiation and an Endpoint for Photoprotection Assessment
Laurent Marrot*, Jean-Philippe Belaidi, Jean-Roch Meunier, Philippe Perez and Catherine Agapakis-Causse L‘Oreal Advanced Research, Life Sciences Research, Aulnay-Sous-Bois, France
Received 4 January 1999; accepted 5 March 1999
ABSTRACT The induction of DNA breaks by UVA (320-400 nm) in the nucleus of normal human melanocytes in culture was investigated using single cell gel electrophoresis, also called the comet assay. Endogenous pigment and/or mel- anin-related molecules were found to enhance DNA breakage: comets were more intense in melanocytes than in fibroblasts, in cells with high melanin content or after stimulation of melanogenesis by supplying tyrosine in the culture medium. After UVA doses where strong comets were observed, neither cytotoxicity nor stimulation of ty- rosinase activity were detected. However, the accumula- tion of p53 protein suggested that cells reacted to geno- toxic stress under these experimental conditions. The same approach was used to compare two sunscreens with identical sun protection factors but different UVA pro- tection factors. The results presented in this paper sug- gest that human melanocytes may be used as a target cell to evidence broadspectrum photoprotection. Moreover, these data appear to be helpful in getting a better un- derstanding of the role of sunlight in the initiating steps of melanocyte transformation.
INTRODUCTION
Although the role of UV radiation in the etiology of non- melanoma skin cancer is now well established (1,2), the re- lationship between melanoma incidence and sunlight expo- sure is still a matter of debate. Epidemiological studies show strong evidence for increased risk related to sunburn, but the connection with total sun exposure is unclear ( 3 ) . Experi- mental studies are limited by the lack of suitable animal models. Commonly used animals, such as mice, are not sus- ceptible to melanoma induction upon exposure to UV alone without combined treatment by chemical carcinogens (4). However, chronic UVB exposure of the South American opossum (Monoddphis durnesfica) results in the formation of cutaneous melanoma ( S ) , and UVA was shown to be ef-
fective in stimulating melanocytic hyperplasia in the skin of this animal (6). An action spectrum for the induction of mel- anoma has also been determined in a fish model where a role for UVA and visible radiation was reported (7).
Contrary to nonmelanoma skin cancer, the mechanisms involved in the initiating steps of melanoma are not clear. For instance, the role of the p53 tumor suppressor gene is well documented in squamous cell carcinoma (SCC)t where p53 is mutated with a frequency of about 90% or in basal cell carcinoma (BCC) with a frequency of about SO%, and the mechanisms that lead to skin tumors have been described in detail (8,9). In the case of melanoma, only a few studies about mutations have been published and many of them have concluded that pS3 is unlikely to play a major role (10-12). Another possible tumor suppressor gene, the p16CDKN”1NK4 gene, is also suspected to be involved in melanoma suscep- tibility: its deletion andor mutation has been reported, but here again, diverging results were obtained (13-15). Muta- tions in the N-ras oncogene have also been described in pa- tients with cutaneous melanoma, and the involvement of UV-induced dipyrimidine sites was suggested (16).
The role of DNA damage in the development of mela- noma is highly probable. Patients with the DNA repair de- ficiency disease xeroderma pigmentosum (XP) are several hundred times more susceptible to melanoma. In this case, the anatomic distribution of tumors is not the same as in carcinomas, suggesting differences in the mechanisms of in- duction (17). Furthermore, when the p53 gene is mutated in melanoma, mutations known as UV signatures are predom- inant and they might result from pyrimidine dimers induced by sunlight in the genome of melanocytes (18).
On the other hand, the role of melanin in photoprotection, if any, can also be questioned. It is well established that individuals with dark skin are less susceptible to sun-induced skin cancer than those with fair skin (19). However, recent studies have shown that epidermal tanning is not always ef- fective for preventing skin cancer in the mouse model (20) and that UV-induced melanogenesis in skin types I1 or 111 offers only a moderate protection against erythema (21).
”To whom correspondence hhould be adressed at: L’Oreal Advanced Research. Life Science Research. Investigative Toxicology De- partment. Genotoxicity Group. I . avenue Eugene Schueller, BP22. 93601 Aulnay-Sous-Bois cedex, France. Fax: 33.1.48.68.91.80: e-mail: lmarrot@ loreal-recherche.com 1999 American Societ! f o r Photobiology 003 I -8655/99 $5.00+0.00
tAhhreviations: BCC, basal cell carcinoma: DOPA, dihydroxyphe- nylalanine: MSH, melanocyte-stimulating hormone; NHEM, nor- mal human melanocyte: PAGE, polyacrylamide gel electrophoresis: PBS, phosphate-buffered saline: SCC, squamous cell carcinoma: SDS, sodium dodecyl sulfate: SPF, sun protection factor: XP, xe- roderma pigmentosum.
686

Photochemistry and Photobiology, 1999, 69(6) 687
0 1 0 tw 310 y o ) 6 o s w I w
WAVELENQTH (nm)
Figure 2. Spectral power distribution of solar UV or UVA from the solar simulator used in the experiments.
Figure 1. Morphological aspect of cultured normal human mela- nocytes used in the experiments (magnification: X200).
Similar discrepancies can also be found in studies with cell cultures: some authors have proposed that intracellular mel- anin protects from pyrimidine dimers induction (22) while others did not find any effects of the pigment (23). One must also consider that melanin precursors and melanin polymer itself are both photoreactive (24-27). Upon illumination, a process of photooxidation can occur leading to the formation of H202 (28). This compound can reach the nucleus and induce strand breaks or oxidative damage via the Fenton reaction. The biosynthesis of melanin is confined within a specialized organelle, the melanosome, where it is supposed to be isolated from the other cellular components. However, intermediate products such as dihydroxyindole can escape and can be detected in urine and plasma. Moreover, photo- oxidative stress may damage the melanosome membrane and promote the leakage of reactive chemicals. Indeed, it has been suggested that damaged melanosomes could be asso- ciated with cytotoxic phenomena sometimes observed in melanoma cells (29).
In such a context, more studies about the impact of UV radiation on normal human melanocytes are required in or- der to determine the molecular events triggering the first steps of melanoma development. In vitro systems could be helpful because it is now possible to culture human mela- nocytes using conditions where their behavior is very similar to the in vivo situation, at least with regard to melanin pro- duction in response to UV light and a-melanocyte-stimulat- ing hormone (MSH) stimulation (30,31). Thus, the use of melanocytes in vitro constitutes an interesting approach for characterizing the genotoxic effects of photooxidative stress in pigmented cells.
In order to get a better understanding of the molecular events involved in damaging nuclear DNA of normal human melanocytes exposed to solar UV (290400 nm) or whole UVA (320-400 nm), we have used single cell gel electro- phoresis. This test, also named the comet assay, is a simple and visual technique for measuring DNA breakage in indi- vidual cells (32). It has been extensively used for the anal- ysis of the genotoxic effects of environmental chemicals (33) or of UV components of the solar spectrum (34,35). The aim of this study is to investigate the role of intracellular melanin and melanin-related molecules in the induction of DNA pho- tooxidative damage. In parallel, cellular proliferation, stim- ulation of melanogenesis and the status of p53 protein are
evaluated as endpoints of cellular response to phototoxicity. Finally, considering the melanocyte as a very particular tar- get for sunlight, we use this approach to demonstrate that broadspectrum sunscreens with a high and stable effective- ness in filtering out UVA wavelengths are required to ensure good photoprotection of the skin.
MATERIALS and METHODS Chemicals. Phosphate-buffered saline (PBS) was from Gibco-BRL. Media for human cells were from Clonetics Inc. Agarose for the comet assay was the low melting Incertm agarose from FMC. Excell gels sodium dodecyl sulfate (SDS) from Pharmacia were used for SDS-polyacrylamide gel electrophoresis (PAGE). Nitrocellulose membranes (Hybond-Cplus, Amersham) were used for protein trans- fer. Other chemicals were from Sigma. Both sunscreens A and B were prepared and characterized in the laboratories of L’Oreal Ap- plied Research (Centre Zviak, Clichy, France).
Cells and culturing conditions. Cells were normal human mela- nocytes (NHEM) from neonates (Clonetics Inc.). We used two cau- casian strains with different melanin contents: NHEM 560 (average melanin content in picograms: 20 pg/cell) and NHEM 4528 (average melanin content: 30 pgfcell). Cells were cultured at 37°C in a 5% C 0 2 atmosphere in conditions where they can respond to aMSH and UV irradiation as described by Im et al. (31). The morphological aspect of melanocytes cultured in these conditions is shown in Fig. I . When tyrosine was used in order to stimulate melanogenesis, it was added in the medium for 5 days at the concentration of 1 mM. When necessary, the number of viable cells was assessed by cell numeration using a Coulter counter (Coultronix).
Light source and spectral measurements. The light source was a solar simulator from Oriel equipped with a 1000 W xenon short arc lamp. When solar UV (UVB + UVA) was studied, a WG 32011.5 mm filter was used. A WG 335f3 mni filter was used for UVA. In both cases, a dichroic mirror removed infrared and the main part of visible light. The beam size was 152 X 152 mm. The nature and fluence of UV reaching the samples were analyzed with spectrora- diometer Instaspec 111 (Oriel). Figure 2 shows the spectral power distribution of solar UV or UVA used in the experiments. Integration of the area under the spectrum gave the following results: with WG 320 filter, the average irradiance was 10 W/m2 for UVB (290-320 nm), 20 W/m2 for UVA2 (320-340 nm) and 72 W/m2 for UVAl (34WOO nm). With WG 335 filter, the average irradiance was 9 W/m2 for UVA2 and 68.5 Wfml for UVAl.
Irradiation procedure. When the comet assay was performed im- mediately after exposure, melanocytes were first embedded in an agarose-PBS microgel and irradiated in cold PBS (4°C). When ex- posure was followed by post-treatment incubation in growth medi- um, cells were incubated in their initial medium after irradiation in cold PBS in 60 mm culture dishes. Sunscreens were spread on a quartz slide with a 20 p n edge (designed for us by Hellma). In these conditions, the applied amount was around 2 mg/cm2. The slides were placed over the cells during exposure when photopro- tection was assessed.
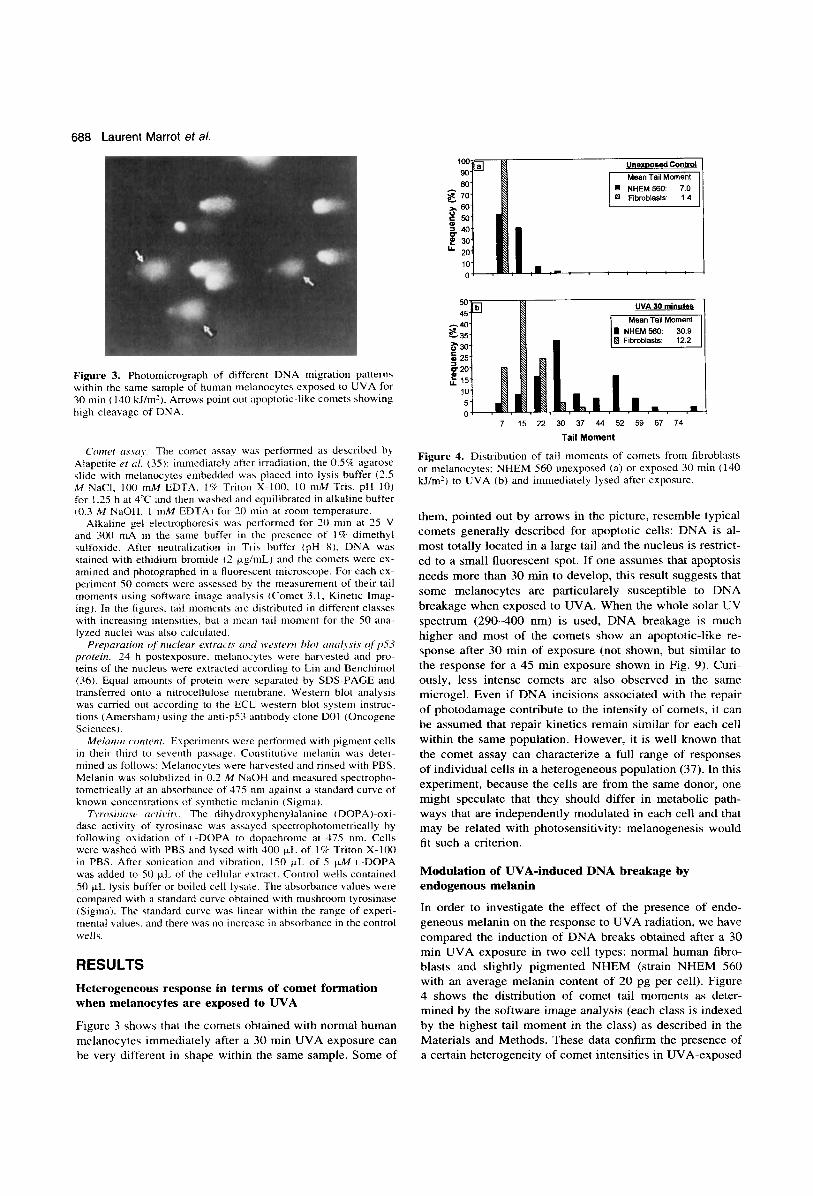
688 Laurent Marrot et a/.
Mean Tail Moment . NHEM560 70 Fibroblasts
%Go
Figure 3. Photomicrograph of different DNA migration patterns within the same sample of human nielanocytes exposed to UVA for 30 min (140 kl/m'). Arrows point out apoptotic-like comets showing high cleavage of DNA.
C017iet m y q y . The comet assay was performed as described by Alapetite er ul. (35): immediately after irradiation. the 0.5% agarose slide with melanocytes embedded wa\ placed into lysis buffer (3.5 M NaCI. 100 mM EDTA. 1Q Triton X-100. 10 mM Tris. pH 10) for 1.35 h at 3'C and then washed and equilibrated in alkaline buffer (0.3 M NaOH. I mM EDTA) for 20 min at rooni temperature.
Alkaline gel electrophoresiq was performed for 10 niin at 25 V and 300 mA in the same buffer in the presencc of 1% dimethyl sulfoxide. After neutralization in Tris buffer ( p H 8). DNA was stained with ethidium bromide ( 2 pg/mL) and the comets were ex- amined and photographed in a fluorescent microscope. For each ex- periment SO comets were assessed by the measurement of their tail moments using software image analysis (Comet 3.1. Kinetic lmag- ing). In the figures. tail moments are distributed in different classes with increasing intensities, but a nican tail moment for the 50 ana- IyLed nuclei was also calculated. f reparution of nlccletrr e.rtrcIc't.7 t i r i d n'esterrl blot frrltr~\sis of'p.53
proteirz. 24 h postexposure. melanocytes were harvested and pro- teins of the nucleus were extracted according to Lin and Benchiniol (36). Equal amounts of protein were separated by SDS-PAGE and transferred onto a nitrocellulose membrane. Western blot analysis was carried out according to the ECL western blot system instruc- tions (Amershani) using the anti-pS3 antibody clone DO1 (Oncogene Sciences).
Mehi7iiI c'ontertt. Experiments were performed with pigment cells in their third to seventh paswge. Constitutive melanin was deter- mined as follows: Melanocytes were hanested and rinsed with PBS. Melanin was solubiliLed in 0.2 M NaOH and measured spectropho- tometrically at an absorbance of 475 nm against a standard curve of known concentrations of synthetic melanin (Sigma).
Tyrosinuse m,tivif.v. The dihydroxyphenylalanine (DOPA)-oxi- dase activity of tyrosinase was yed spectrophotonietrically by following oxidation of i.-DOPA to dopachronie at 475 nm. Cells were washed with PBS and lysed with 400 p L of I % Triton X-100 in PBS. After sonication and vibration. IS0 pL of 5 FM L-DOPA was added to 50 p L of the cellular extract. Control wells contained 50 pL lysis buffer or boilcd cell lysate. The absorbance values were compared with a standard curve obtained with mushroom tyrosinase (Sigma). The standard curve was linear within the range of experi- mental values. and there was no increase in absorbance in the control wells.
RESULTS Heterogeneous response in terms of comet formation when melanocytes are exposed to UVA
Figure 3 shows that the comets obtained with normal human melanocytes immediately after a 30 min UVA exposure can be very different in shape within the same sample. Some of
UVA 30 minutes
Mean Tail Moment
B Fibroblasts:
5 25
15 10 5 0
s 20
7 15 22 30 37 44 52 59 67 74 Tail Moment
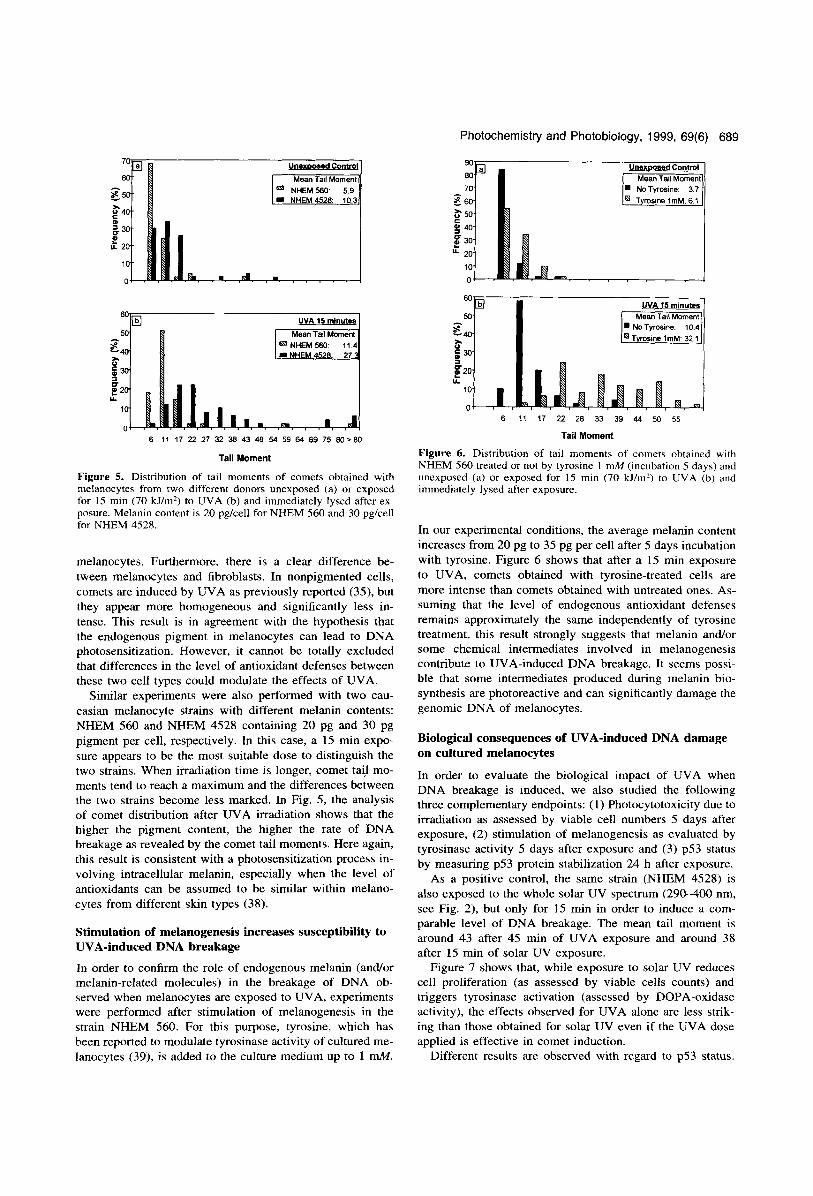
Figure 4. Distribution of tail moments of comets from fibroblasts or melanocytes; NHEM 560 unexposed (a) or exposed 30 min ( 140 lillm') to UVA (b) and immediately lysed after exposure.
them, pointed out by arrows in the picture, resemble typical comets generally described for apoptotic cells: DNA is al- most totally located in a large tail and the nucleus is restrict- ed to a small fluorescent spot. If one assumes that apoptosis needs more than 30 min to develop, this result suggests that some melanocytes are particularely susceptible to DNA breakage when exposed to UVA. When the whole solar UV spectrum (290-400 nm) is used, DNA breakage is much higher and most of the comets show an apoptotic-like re- sponse after 30 min of exposure (not shown, but similar to the response for a 45 min exposure shown in Fig. 9). Curi- ously, less intense comets are also observed in the same microgel. Even if DNA incisions associated with the repair of photodamage contribute to the intensity of comets, it can be assumed that repair kinetics remain similar for each cell within the same population. However, it is well known that the comet assay can characterize a full range of responses of individual cells in a heterogeneous population (37). In this experiment, because the cells are from the same donor, one might speculate that they should differ in metabolic path- ways that are independently modulated in each cell and that may be related with photosensitivity: melanogenesis would fit such a criterion.
Modulation of UVA-induced DNA breakage by endogenous melanin
In order to investigate the effect of the presence of endo- geneous melanin on the response to UVA radiation, we have compared the induction of DNA breaks obtained after a 30 min UVA exposure in two cell types: normal human fibro- blasts and slightly pigmented NHEM (strain NHEM 560 with an average melanin content of 20 pg per cell). Figure 4 shows the distribution of comet tail moments as deter- mined by the software image analysis (each class is indexed by the highest tail moment in the class) as described in the Materials and Methods. These data confirm the presence of a certain heterogeneity of comet intensities in UVA-exposed
Unexwsed Control
Mean Tail Moment
NHEM4528. 103
6 UVA 15 minutes
Mean Tail Moment
I N H 452 ’ 27.
6 11 17 22 21 32 38 43 48 54 59 64 69 75 80 ,80
Tail Moment
Figure 5. Distribution of tail moments of comets obtained with melanocytes from two different donors unexposed (a) or exposed for 15 min (70 kJ/m2) to UVA (b) and immediately lysed after ex- posure. Melanin content is 20 pg/cell for NHEM 560 and 30 pg/cell for NHEM 4528.
melanocytes. Furthermore, there is a clear difference be- tween melanocytes and fibroblasts. In nonpigmented cells, comets are induced by UVA as previously reported (35), but they appear more homogeneous and significantly less in- tense. This result is in agreement with the hypothesis that the endogenous pigment in melanocytes can lead to DNA photosensitization. However, it cannot be totally excluded that differences in the level of antioxidant defenses between these two cell types could modulate the effects of UVA.
Similar experiments were also performed with two cau- casian melanocyte strains with different melanin contents: NHEM 560 and NHEM 4528 containing 20 pg and 30 pg pigment per cell, respectively. In this case, a 15 min expo- sure appears to be the most suitable dose to distinguish the two strains. When irradiation time is longer, comet tail mo- ments tend to reach a maximum and the differences between the two strains become less marked. In Fig. 5, the analysis of comet distribution after UVA irradiation shows that the higher the pigment content, the higher the rate of DNA breakage as revealed by the comet tail moments. Here again, this result is consistent with a photosensitization process in- volving intracellular melanin, especially when the level of antioxidants can be assumed to be similar within melano- cytes from different skin types (38).
Stimulation of melanogenesis increases susceptibility to UVA-induced DNA breakage
In order to confirm the role of endogenous melanin (and/or melanin-related molecules) in the breakage of DNA ob- served when melanocytes are exposed to UVA, experiments were performed after stimulation of melanogenesis in the strain NHEM 560. For this purpose, tyrosine, which has been reported to modulate tyrosinase activity of cultured me- lanocytes (39), is added to the culture medium up to 1 mM.
Photochemistry and Photobiology, 1999, 69(6)
UVA 15 minutes Mean Tail Moment
6 11 17 22 28 33 39 44 50 55
Tail Moment
Figure 6. Distribution of tail moments of comets obtained
689
with NHEM 560 treated or not by tyrosine 1 mM (incubation 5 days) and unexposed (a) or exposed for 15 min (70 kJ/m’) to UVA (b) and immediately lysed after exposure.
In our experimental conditions, the average melanin content increases from 20 pg to 35 pg per cell after 5 days incubation with tyrosine. Figure 6 shows that after a 15 min exposure to UVA, comets obtained with tyrosine-treated cells are more intense than comets obtained with untreated ones. As- suming that the level of endogenous antioxidant defenses remains approximately the same independently of tyrosine treatment, this result strongly suggests that melanin andor some chemical intermediates involved in melanogenesis contribute to UVA-induced DNA breakage. It seems possi- ble that some intermediates produced during melanin bio- synthesis are photoreactive and can significantly damage the genomic DNA of melanocytes.
Biological consequences of UVA-induced DNA damage on cultured melanocytes
In order to evaluate the biological impact of UVA when DNA breakage is induced, we also studied the following three complementary endpoints: (1) Photocytotoxicity due to irradiation as assessed by viable cell numbers 5 days after exposure, ( 2 ) stimulation of melanogenesis as evaluated by tyrosinase activity 5 days after exposure and (3) p53 status by measuring p53 protein stabilization 24 h after exposure.
As a positive control, the same strain (NHEM 4528) is also exposed to the whole solar UV spectrum (290-400 nm, see Fig. 2), but only for 15 min in order to induce a com- parable level of DNA breakage. The mean tail moment is around 43 after 45 min of UVA exposure and around 38 after 15 min of solar UV exposure.
Figure 7 shows that, while exposure to solar UV reduces cell proliferation (as assessed by viable cells counts) and triggers tyrosinase activation (assessed by DOPA-oxidase activity), the effects observed for UVA alone are less strik- ing than those obtained for solar UV even if the UVA dose applied is effective in comet induction.
Different results are observed with regard to p53 status.
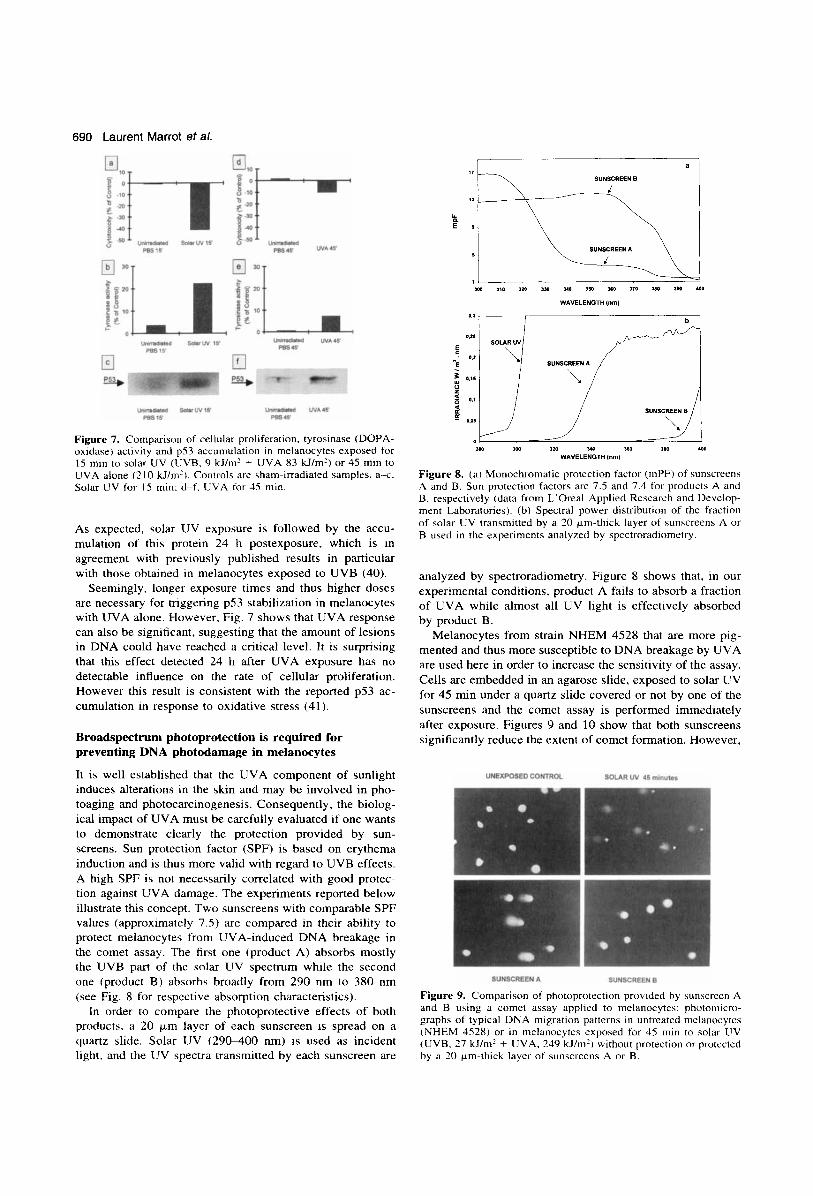
690 Laurent Marrot et al.
Figure 7. Comparison of cellular proliferation. tyrosinase (DOPA- oxidase) activity and p53 accumulation in melanocytes exposed for 15 min to solar UV (LJVB, 9 kJ/m2 + UVA 83 kJ/m') or 45 min to UVA alone (2 10 kJ/m'). Controls are sham-irradiated samples. a x . Solar UV for IS min: d-f. EVA for 45 min.
As expected, solar UV exposure is followed by the accu- mulation of this protein 24 h postexposure, which is in agreement with previously published results in particular with those obtained in melanocytes exposed to UVB (40).
Seemingly, longer exposure times and thus higher doses are necessary for triggering p53 stabilization in melanocytes with UVA alone. However, Fig. 7 shows that UVA response can also be significant, suggesting that the amount of lesions in DNA could have reached a critical level. It is surprising that this effect detected 24 h after UVA exposure has no detectable influence on the rate of cellular proliferation. However this result is consistent with the reported p53 ac- cumulation in response to oxidative stress (41 ).
Broadspectrum photoprotection is required for preventing DNA photodamage in melanocytes
It is well established that the UVA component of sunlight induces alterations in the skin and may be involved in pho- toaging and photocarcinogenesis. Consequently, the biolog- ical impact of UVA must be carefully evaluated if one wants to demonstrate clearly the protection provided by sun- screens. Sun protection factor (SPF) is based on erythema induction and is thus more valid with regard to UVB effects. A high SPF is not necessarily correlated with good protec- tion against UVA damage. The experiments reported below illustrate this concept. Two sunscreens with comparable SPF values (approximately 7.5) are compared in their ability to protect melanocytes from UVA-induced DNA breakage in the comet assay. The first one (product A) absorbs mostly the UVB part of the solar UV spectrum while the second one (product B) absorbs broadly from 290 nm to 380 nm (see Fig. 8 for respective absorption characteristics).
In order to compare the photoprotective effects of both products. a 20 pm layer of each sunscreen is spread on a quartz slide. Solar UV (290-400 nm) is used as incident light. and the UV spectra transmitted by each sunscreen are
SUNSCREENB ;:p, , \ \\..\ +\ SUNSCREEN A
SOLAR W r SUNSCREENA / I
0 i----'--. yo 160 a m 410
/ 2M 1W 120
WAVELENGTH Inm)
Figure 8. ( a ) Monochromatic protection factor (rnPF) of sunscreens A and B. Sun protcction factors are 7.5 and 7.4 for products A and B. respectively (data from L'Oreal Applied Research and Develop- ment Laboratories). (b) Spectral power distribution of the fraction of solar LJV transmitted by a 20 km-thick layer of sunscreens A or B used in the experiments analyzed by spectroradiometry.
analyzed by spectroradiometry. Figure 8 shows that, in our experimental conditions, product A fails to absorb a fraction of UVA while almost all UV light is effectively absorbed by product B.
Melanocytes from strain NHEM 4528 that are more pig- mented and thus more susceptible to DNA breakage by UVA are used here in order to increase the sensitivity of the assay. Cells are embedded in an agarose slide, exposed to solar UV for 45 min under a quartz slide covered or not by one of the sunscreens and the comet assay is performed immediately after exposure. Figures 9 and 10 show that both sunscreens significantly reduce the extent of comet formation. However,
UNEXPOSEDCONTROL SOLAR UV 45 minutes
SUNSCREEN A SUNSCREENB
Figure 9. Comparison of photoprotection provided by sunscreen A and B using it comet assay applied to melanocytes: photoniicro- graphs of typical DNA migration patterns in untreated melanocytes (NHEM 4528) or in melanocytes exposed for 45 min to solar LIV (UVB, 27 kJ/m' + UVA. 249 kJ/m') without protection or protected by a 20 km-thick layer of sunscreens A or B.

Photochemistry and Photobiology, 1999, 69(6) 691
80
70 - 60 - 50 e 40 30
s h
; 20 10
0
80- Solar UV 6 minutes Mean Tail Moment - 60
7 13 19 24 30 36 42 48 54 60 Tail Moment
Figure 10. Distribution of tail moments of comets from melano- cytes (NHEM 4528): (a) unexposed cells or cells exposed for 45 min to solar UV without protection; (b) cells protected by a 20 km- thick layer of sunscreens A or B.
a difference can be observed between product A and product B. Product A does not totally abrogate the induction of DNA breaks and in this case, the mean tail moment remains higher than that of the unirradiated control or of sample protected by product B. We speculate that the remaining comets de- tected, despite the protection provided by sunscreen A, cor- respond to melanocytes that would produce UVA-induced apoptotic-like comets in the absence of protection. This re- sult supports the idea that some melanocytes would be more photosensitive within the same population. The amount of light transmitted by product A is then effective enough for inducing a biologically detectable photooxidative stress in DNA, even if UVB is removed. These results demonstrate that SPF alone is not adequate to evaluate photoprotection and that a precise assessment of UVA protection factor is an additional essential requirement.
DISCUSSION Pigmentation is generally considered as a photoprotective process either for the whole skin or at the level of individual keratinocytes in which melanin is transferred (42). This con- cept is supported by the fact that DNA repair can trigger melanin synthesis, melanogenesis being then regarded as a part of a eukaryotic DNA damage-responsive SOS like sys- tem (43). Moreover, Yohn et al. have shown that melano- cytes from black skin are more resistant to UVA cytotoxicity than melanocytes from white skin (44). However, Noz et al. have proposed a role for endogenous melanin in the induc- tion of UVB-induced photodamage in human melanocytes as detected by comet assay (45). Recently, Wenczl et al. have shown that endogenous melanin can induce breaks in the DNA of pigment cells when exposed to UVA and that stimulation of pigmentation enhances this photosentization process (46). Furthermore, in the study of the action spec- trum for melanoma induction in the fish model, Setlow et
al. have interpreted the involvement of wavelengths greater than 360 nm by a possible interaction with melanin (7).
Our results confirm that the melanocyte must be regarded as a very particular target cell for sunlight and, more spe- cifically, for UVA. Indeed, the pool of reactive intermediates involved in melanogenesis could behave as an endogeneous photosensitizer and generate an oxidative stress when irra- diated. Moreover, in the case of Caucasian human melano- cytes, the photoinstability of pheomelanin must also be con- sidered as a source of free radicals (47). Noteworthy is also the fact that melanocytes could not be particularly well sup- plied with antioxidant enzymes. They are, for instance, less resistant than keratinocytes or fibroblasts to H,O,-induced cytotoxicity (38). If endogenous melanin or related mole- cules could produce H202 during UVA exposure, the high level of strand breaks detected in our comet assay for UVA- irradiated melanocytes would not be surprising. We cannot exclude that part of the DNA breakage detected in our ex- periments occurs as a consequence of the diffusion of re- active molecules released during cell lysis within the agarose microgel. Work is in progress in order to evaluate the con- tribution of such possible indirect DNA-damaging reactions. In any case, our results suggest that the melanocyte acquires a prooxidant status when exposed to UVA, a status in which many biochemical insults, including DNA damage, could oc- cur.
Moreover, in a population of melanocytes derived from the same donor, some cells appear to be more susceptible to DNA breakage induced by UVA. We propose that these cells are involved in a precise step of the melanogenesis pathway, a step during which highly photoreactive inter- mediates are produced. Further studies are required in order to elucidate the importance of such a process in the initiation of melanocyte transformation.
One might also wonder about the biological consequences of UVA-induced DNA strand breaks. In our experiments, at doses where comets are relatively intense, neither cytotox- icity nor melanogenesis stimulation have been detected. However, accumulation of p53 protein is observed, but for a higher dose than when the whole solar UV spectrum is used. This result could suggest that UVA impact is perceived as not negligible by the melanocyte in terms of intracellular signalling, and this response is in agreement with the con- tribution of UVA to genotoxicity of sunlight. It is also note- worthy that the doses delivered here are relatively low if one considers the total amount of UVA received by human be- ings in 1 day, especially in summer, during extensive sun- bathing.
Our results show that hazard increases when pigmentation is starting, suggesting that skin undergoing tanning could be more exposed to photogenotoxic stress. This raises the ques- tion of photoprotection. Sunscreens are generally evaluated by their ability to prevent sunburn and, as a consequence, the SPF is essentially related to UVB effects on the skin. What would be the best protection against solar UVA is often not clearly defined. The experiments reported here us- ing two sunscreens with comparable SPF but different UVA protection factors demonstrate that it is essential to cover broadly the whole UV range of sunlight and this must be done with adequately balanced formulations to avoid a sit- uation where high efficacy in the UVB region (erythema

692 Laurent Marrot et a/.
prevention) would allow longer exposure to unfiltered UVA. Our results also underline the necessity of photostability. Product A can be compared to a nonphotostable sunscreen that would lose its effectiveness after a few hours of expo- sure. We suggest being particularly careful when the skin- tanning process is stimulated, especially for light skin pho- totypes, because development of pigmentation might be as- sociated with increased photooxidative potential in melano- cytes. Some epidemiological studies have suggested that a positive association exists between sunscreen use and mel- anoma incidence (48). Among the possible bias, it could be interesting to consider the fact that some people stop to use sunscreen as soon as their tanning starts.
Finally, this work shows how in vitro studies can be of help to get information at the molecular level, especially for very specific kinds of cells like melanocytes. Of course, many factors related to the interaction with other cell types in the skin are not considered here. It is also obvious that immunological considerations are crucial in photocarcino- genesis. However, it would have been very difficult to isolate quickly the necessary amount of melanocytes from skin ex- plants to perform the experiments described here. The ob- jective is to have a model as informative as possible for a better understanding of phototoxicity mechanisms in pig- mented cells (49). This is why culture conditions must be particularly well defined (and improved) in order to maintain the very specific behavior of this peculiar cell type. On the other hand, such an approach is also an interesting tool for the study of biological effects of UVA in vitro. It should be used complementary to other methods, as for instance three- dimensional slun equivalents (50) , to assess the effectiveness and safety of photoprotection.
Ac~nou.ledKements-Dr. A~erbeck (Institut Curie. Paris. France) is gratefully acknowledged for critical reading of the manuscript and helpful discussions. Drs. Candau. Forestier and Refrkgier (L’Oreal Applied Research. Centre Zviak. Clichy, France) are acknowledged for providing sunscreens with all the related data (spectra, SPF) and helpful discussions. Thanks are due to J . Caradec. N. Lequesne and A. Reinhardt for their help in preparing the manuscript.
REFERENCES 1. Urbach. F. (1997) Ultraviolet radiation and skin cancer of hu-
2. English, D. R.. B. K. Armstrong, A. Kricker and C. Fleming ( 1997) Sunlight and cancer. Cancer Causrs Control 8. 271-283.
3. Elwood, J. M. ( 1996) Melanoma and sun exposure. Semin. On- col. 23. 650466 .
4. Kusewitt. D. F. and R. D. Ley (1996) Animal models of mel- anoma. Cancer Sun,. 26, 35-70,
5 . Ley, R. D., L. A. Applegate. R. S. Padilla and T. D. Stuart ( 1989) Liltraviolet radiation-induced malignant melanoma in Monodelphis domestiw. Phorochem. Photohiol. 50. 1-5.
6. Ley, R. D. (1997) Ultraviolet radiation A induced precursors of cutaneous melanoma in Monodelphi.! domesticu. Cancer Res. 57. 3682-3684.
7 . Setlow. R. €3.. E. Grist, K. Thompson and A. D. Woodhead ( 1 993) Wavelengths effective i n induction of malignant mela- noma. Proc. Nirtl. Auid. St.i USA 90. 6 6 6 M 6 7 0 .
8. Daya-Grosjean. L., N. Dumaz and A. Sarasin (1995) The spec- ificity of p53 mutation spectra in sunlight induced human can- cers. J. Pltntochem. Photobiol B Biol. 28, 1 15- 124.
9. Brash, D. E.. A. Ziepler, A. S Jonason. J. A. Simon. S. Kunala and D. J. Leffell (1996) Sunlight and sunburn in human skin cancer: pS3. apoptosi.; and tumor promotion. J . Im~e.st. Dennu- r o l . Sxnip. Pro(.. 1. 136-141.
mans. J . P/l~Jt<JchCIlI, PkUt~JhifJl. B BIol. 40. 3-7.
10. Lubbe. J . , M. Reichel, G. Burg and P. Kleihues (1994) Absence of p53 gene mutations in cutaneous melanoma. J . Invest. Der- mri!ol. 102, 8 19-82 1.
11. Montano, X. . M. Shamsher, P. Whitehead, K. Dawson and J . Newton (1994) Analysis of p53 in human cutaneous melanoma cell lines. Oncogene 9, 1455-1459.
12. Hartmann, A., H. Blaszyk. J. S. Cunningham, R. M. Mac- Govern, J. S. Schroeder, S. D. Helander, M. R. Pittelkow. S. S. Somnier and J . S. Kovach ( I 996) Overexpression and mutations of p53 in metastatic malignant melanomas. Int. J . Cancer 67,
13. Healy. E., S. Sikkink and J. L. Rees (1996) Infrequent mutation of p161NK4 in sporadic melanoma. J . Invest. Dermcrtol. 107, 3 18- 321.
14. Herbst. R. A,, R. Cutzmer, F. Matiaske, S. Mommert, A. Kapp, J. Weiss, K. C. Arden and K. Cavenee (1997) Further evidence for ultraviolet light induction of CDKN2 ( ~ 1 6 ” ~ ‘ ) mutations in sporadic melanoma in vivo (letter). J. Inl9est. Dermatol. 108, 950.
15. Matsumura, Y., C. Nishigori, T. Yagi, S. lmamura and H. Tak- ebe (1998) Mutations of p16 and p15 tumor suppressor genes and replication errors contribute independently to the pathogen- esis of sporadic malignant melanoma. Arch. Dermatol. Res. 290, 175-180.
16. Jiveskog, S., B. Ragnarsson-Olding, A. Platz and U. Ringborg (1998) N-ras mutations are common in melanomas from sun- exposed skm of humans but rare in mucosal membranes or un- exposed skin. J . Invest. Dermatol. 111, 757-761.
17. Kraemer, K. H., M. M. Lee, A. D. Andrews and W. C. Lambert ( 1993) The role of sunlight and DNA repair in melanoma and non-melanoma skin cancer. Arch. Dermatol. 130. 1018-1021.
18. Weiss, J., M. Heine. K. C. Arden, B. Korner, H. Pilch, R. A. Hebst and E. G. Jung (1995) Mutation and expression of p53 in malignant melanomas. Rec. Results Cancer Rex 139, 137- 154.
19. Kollias, N., R . M. Sayre, L. Zeise and M. R Chedekel (1991) Photoprotection by melanin. J . Photochem. Photohiol. B B i d . 9, 135-160.
20. Kipp, C. , E. J. Lewis and A. R Young (1998) Furocoumarin- induced epidermal melanogenesis does not protect against skin photocarcinogenesis in hairless mice. Phorochem. Photobiol. 67, 126132.
21. Sheehan. J. M., C . S. Potten and A. R Young (1998) Tanning in human skin types I1 and 111 offers modest photoprotection against erythema. Pkotochem. Photobiol. 68, 588-592.
22. Kobayashi. N., T. Muramatsu, Y. Yamashina, T. Shirai. T. Ohn- ishi and T. Mori (1993) Melanin reduces ultraviolet-induced DNA damage formation and killing rate in cultured human mel- anoma cells. J . Invest. Dermatol. 101, 685489 .
13. De Leeuw, S. M., J. W. I. M. Simons, B. J. Vermeer and A. A. Schothorst ( 1995) Comparison of melanocytes and keratinocytes in ultraviolet-induced DNA damage per minimum erythema dose sunlight: applicability of ultraviolet action spectra for risk estimates. J . Invest. Dermutol. 105, 259-263.
24. Koch, W. H. and M. R. Chedekel (1986) Photoinitiated DNA damage by melanogenic intermediates in vitro. Photochem. Photohiol. 44, 703-7 10.
25. Huselton, C. A. and H. Z . Hill (1990) Melanin photosensitised ultraviolet light WVC) DNA damage in pigmented cells. En- &,iron. Mol. Mutagen. 16, 3 7 4 3 .
26. Routaboul. C., C. L. Serpentini, P. Msika, J . P. Cesarini and N. Paillous ( 1995) Photosensitization of supercoiled DNA damage by 5,6-dihydroxyindole-2-carboxylic acid, a precursor of CLI-
melanin. Photodiem. Photobiol. 62, 469475 . 27. Reszka, K. and K. Jimbow (1993) Electron donnor and acceptor
properties of melanin pigments in the skin. In Oxidutiw Stress in Dermatology (Edited by J. Fuchs and L. Paker), pp. 287- 320. M. Dekker, New York.
28. Korytowski. W., B. Pilas, T. Sarna and B. Kalyanaraman (1987) Photoinduced generation of hydrogen peroxide and hydroxyl radicals in melanins. Photochem. Photobiol. 45, 185-1 90.
29. Borovansky, J., P. Mirejovsky and P. A. Riley (1991) Possible relationship between abnormal melanosome structure and cy-
3 13-3 17.

Photochemistry and Photobiology, 1999, 69(6) 693
totoxic phenomena in malignant melanoma. Neoplasma 38, 393400.
30. Hedley, S. J., D. J. Gamkrodger, A. P. Weetman and S. Macneil (1998) a-MSH and melanogenesis in normal human adult me- lanocytes. Pigment Cell Res. 11, 45-56.
31. Im, S., 0. Moro, F. Peng, E. E. Medrano, J. Cornelius, G . Bab- cock, J. J Nordlund and Z . A. Abdel-Malek (1998) Activation of the cyclic AMP pathway by a-melanotropin mediates the response of human melanocytes to ultraviolet B radiation. Can- cer Res. 58, 47-54.
32. Tice. R. R. (1995) The single cell geVcomet assay: a microgel electrophoretic technique for the detection of DNA damage and repair in individual cells. In Environmental Mufagenesis (Edited by D. H. Phillips and S. Venett), pp. 315-339. Bios, Oxford.
33. Belpaeme, K., K. Cooreman and M. Kirsch-Volders (1998) De- velopment and validation of the in vivo alkaline comet assay for detecting genomic damage in marine flatfish. Mutat. Res. 415. 167-184.
34. Arlett. C. F., J. E. Lowe, S. A. Harcourt, A. P. W. Waugh, J. Cole, L. Roza, B. Diffey, T. Mori, 0. Nikaido and M. H. L. Green (1993) Hypersensitivity of human lymphocytes to UV-B and solar irradiation. Cancer Res. 53, 609414.
35. Alapetite, C., E. Moustacchi, T. Wachter and E. Sage (1996) Use of the alkaline comet assay to detect DNA repair deficien- cies in human fibroblasts exposed to UVC, UVB, UVA and y- rays. Int. J. Radiat. Biol. 69, 359-369.
36. Lin, Y. and S. Benchimol (1995) Cytokines inhibit p53-medi- ated apoptosis but not p53-mediated GI arrest. Mol. Cell. B id .
37. Olive, P. L., P. J. Johnston. J. P. Banath and R. E. Durand (1998) The comet assay: a new method to examine heteroge- neity associated with solid tumors. Nature Med. 4, 103-105.
38. Yohn, J. J., D. A. Noms, D. G. Yrastorza, I. J. Buno, J. A. Leff, S. S. Hake and J. E. Repine (1991) Disparate antioxidant en- zyme activities in cultured human cutaneous fibroblasts, kera- tinocytes and melanocytes. J. Invest. Dennatol. 97, 405409.
39. Smit, N. P. M., H. Van Der Meulen, H. K. Koerten, R. M. Kolb, A. M. Mommaas, E. G. W. M. Lentjes and S. Pavel (1997) Melanogenesis in cultured melanocytes can be substantially in- fluenced by L-tyrosine and L-cysteine. J. Invest. Dermatol. 109, 796-800.
40. Barker, D., K. Dixon, E. E. Medrano, D. Smalara, S. Im, D.
15, 6045-6054.
Mitchell. G. Babcock and Z. A. Abdel-Malek (1995) Compar- ison of the response of human melanocytes with different mel- anin contents to ultraviolet B irradiation. Cancer Rex 55,404 1 - 4046.
41. Vile, G . F. (1997) Active oxygen species mediate the solar ul- traviolet-radiation dependent increase in the tumor suppressor protein p53 in human skin fibroblasts. FEBS Lrtr. 412, 70-74.
42. Kobayashi, N., A. Nakagawa, T. Muramatsu. Y. Yamashina. T. Shirai, M. W. Hashimoto, Y. Ishigaki, T. Ohnishi and T. Mori (1998) Supranuclear melanin caps reduce ultraviolet induced DNA photoproducts in human epidermis. J. Invest. Dermatol. 110, 806-810.
43. Eller, M. S.. K. Ostrom and B. A. Gilchrest (1996) DNA dam- age enhances melanogenesis. Proc. Natl. Acad. Sci. USA 93, 1087- 1092.
44. Yohn. J. J., M. B. Lyons and D. A. Norris (1992) Cultured human melanocytes from black and white donors have different sunlight and ultraviolet A radiation sensitivities. J . Invest. Der- matol. 99, 454459.
45. Noz, K. C., M. Bauwens, P. P. W. Van Buul. H. Vrolijk, A. A. Schothorst, S. Pavel, H. J. Tanke and B. Vermeer (1996) Comet assay demonstrates a higher ultraviolet B sensitivity to DNA damage in dysplastic nevus cells than in common melanocytic nevus cells and foreskin melanocytes. J . Invest. Dermatol. 106, 1198-1202.
46. Wenczl, E., G . P. Van der Schans, L. Roza, R. M. Kolb, A. J. Timmerman, N. P. M. Smit, S. Pavel and A. A. Schothorst ( 1998) (Pheo)melanin photosensitizes UVA-induced DNA dam- age in cultured human melanocytes. J . Invest. Dermatol. 111, 678-682.
47. Menon, I. A., S. Persad, N. S. Ranadive and H. F. Haberman (1983) Effects of ultraviolet-visible irradiation in the presence of melanin isolated from human black or red hair upon Ehrlich ascites carcinoma cells. Cancer Res. 43, 3165-3169.
48. Donawho, C. and P. Wolf (1996) Sunburn, sunscreen and mel- anoma. Curr. Opin. Oncol. 8, 159-166.
49. Linge, C. (1996) Relevance of in vitro melanocytic cell studies to the understanding of melanoma. Cancer Sun,. 26, 71-88.
50. Bernerd, F. and D. Asselineau (1998) UVA exposure of human skin reconstructed in vitro induces apoptosis of dermal fibro- blasts: subsequent connective tissue repair and implications i n photoaging. Cell Death Differ. 5 , 792-802.
Related Documents











