The High-Lakes Project Nathalie A. Cabrol, 1,2 Edmond A. Grin, 1,2 Guillermo Chong, 3 Edwin Minkley, 4 Andrew N. Hock, 5 Youngseob Yu, 6 Leslie Bebout, 1 Erich Fleming, 1 Donat P. Ha ¨der, 7 Cecilia Demergasso, 8 John Gibson, 9 Lorena Escudero, 3 Cristina Dorador, 8 Darlene Lim, 1,2 Clayton Woosley, 1,2 Robert L. Morris, 2 Cristian Tambley, 10 Victor Gaete, 11 Matthieu E. Galvez, 12 Eric Smith, 13 Ingrid Uskin-Peate, 14 Carlos Salazar, 15 G. Dawidowicz, 16 and J. Majerowicz 16 Received 30 June 2008; revised 29 June 2009; accepted 29 July 2009; published 5 November 2009. [1] The High Lakes Project is a multidisciplinary astrobiological investigation studying high-altitude lakes between 4200 m and 6000 m elevation in the Central Andes of Bolivia and Chile. Its primary objective is to understand the impact of increased environmental stress on the modification of lake habitability potential during rapid climate change as an analogy to early Mars. Their unique geophysical environment and mostly uncharted ecosystems have added new objectives to the project, including the assessment of the impact of low-ozone/high solar irradiance in nonpolar aquatic environments, the documentation of poorly known ecosystems, and the quantification of the impact of climate change on lake environment and ecosystem. Data from 2003 to 2007 show that UV flux is 165% that of sea level with maximum averaged UVB reaching 4 W/m 2 . Short UV wavelengths (260–270 nm) were recorded and peaked at 14.6 mW/m 2 . High solar irradiance occurs in an atmosphere permanently depleted in ozone falling below ozone hole definition for 33–36 days and between 30 and 35% depletion the rest of the year. The impact of strong UVB and UV erythemally weighted daily dose on life is compounded by broad daily temperature variations with sudden and sharp fluctuations. Lake habitat chemistry is highly dynamical with notable changes in yearly ion concentrations and pH resulting from low and variable yearly precipitation. The year-round combination of environmental variables define these lakes as end-members. In such an environment, they host ecosystems that include a significant fraction of previously undescribed species of zooplankton, cyanobacterial, and bacterial populations. Citation: Cabrol, N. A., et al. (2009), The High-Lakes Project, J. Geophys. Res., 114, G00D06, doi:10.1029/2008JG000818. 1. Introduction [2] The study of terrestrial analog environments supports the design of investigation strategies and life detection methods for planetary exploration. Over the past decades, orbital and ground missions have provided geologic, mor- phologic, mineralogic, and chemical lines of evidence show- ing that water was abundant early in the history of Mars and that the planet was habitable for life as we know it [e.g., Squyres et al., 2004; Knoll et al., 2005; McLennan et al., 2005; Des Marais et al., 2005]. However, phyllosilicates (clays) disappeared from the stratigraphic record around 3.7 Ga ago [Bibring et al., 2005] replaced by sulfate deposits, then by ahydrated ferric oxides (olivine) about 3.2 Ga ago [Christensen et al., 2003; Mustard et al., 2005]. This suc- cession suggests that no further large-scale weathering oc- curred after that period but transient and localized aqueous processes likely associated with high-obliquity cycles were still possible. There is evidence supporting water activity in JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 114, G00D06, doi:10.1029/2008JG000818, 2009 Click Here for Full Article 1 Space Science and Astrobiology Division, NASA Ames Research Center, Moffett Field, California, USA. 2 SETI Carl Sagan Center, Mountain View, California, USA. 3 Centro de Investigacio ´n Cientı ´fica y Tecnolo ´gica para Minerı ´a, Santiago, Chile. 4 Department of Biological Sciences, Carnegie Mellon University, Pittsburgh, Pennsylvania, USA. 5 Department of Earth and Space Sciences, University of California, Los Angeles, California, USA. 6 Department of Civil and Environmental Engineering, Carnegie Mellon University, Pittsburgh, Pennsylvania, USA. Copyright 2009 by the American Geophysical Union. 0148-0227/09/2008JG000818$09.00 7 Department Botanik, University of Erlangen, Erlangen, Germany. 8 Centro de Biotecnologı ´a, Universidad Cato ´lica del Norte, Antofagasta, Chile. 9 Marine Research Laboratories, Tasmanian Aquaculture and Fisheries Institute, University of Tasmania, Hobart, Tasmania, Australia. 10 CHEP, Santiago, Chile. 11 Departamento de Quı ´mica, Universidad Cato ´lica del Norte, Antofagasta, Chile. 12 Ecole Normale Supe ´rieure, Paris, France. 13 Discoverer Ketty Lund Exploration Vessel, Key West, Florida, USA. 14 Geoscience Department, University of Iowa, Iowa City, Iowa, USA. 15 Clı ´nica Mutual de Seguridad, Punta Arenas, Chile. 16 ESRI, Meudon, France. G00D06 1 of 20

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The High-Lakes Project
Nathalie A. Cabrol,1,2 Edmond A. Grin,1,2 Guillermo Chong,3 Edwin Minkley,4
Andrew N. Hock,5 Youngseob Yu,6 Leslie Bebout,1 Erich Fleming,1 Donat P. Hader,7
Cecilia Demergasso,8 John Gibson,9 Lorena Escudero,3 Cristina Dorador,8 Darlene Lim,1,2
Clayton Woosley,1,2 Robert L. Morris,2 Cristian Tambley,10 Victor Gaete,11
Matthieu E. Galvez,12 Eric Smith,13 Ingrid Uskin-Peate,14 Carlos Salazar,15
G. Dawidowicz,16 and J. Majerowicz16
Received 30 June 2008; revised 29 June 2009; accepted 29 July 2009; published 5 November 2009.
[1] The High Lakes Project is a multidisciplinary astrobiological investigation studyinghigh-altitude lakes between 4200 m and 6000 m elevation in the Central Andes of Boliviaand Chile. Its primary objective is to understand the impact of increased environmentalstress on the modification of lake habitability potential during rapid climate change as ananalogy to early Mars. Their unique geophysical environment and mostly unchartedecosystems have added new objectives to the project, including the assessment of the impactof low-ozone/high solar irradiance in nonpolar aquatic environments, the documentationof poorly known ecosystems, and the quantification of the impact of climate change on lakeenvironment and ecosystem. Data from 2003 to 2007 show that UV flux is 165% thatof sea level with maximum averaged UVB reaching 4 W/m2. Short UV wavelengths(260–270 nm) were recorded and peaked at 14.6 mW/m2. High solar irradiance occurs inan atmosphere permanently depleted in ozone falling below ozone hole definition for33–36 days and between 30 and 35% depletion the rest of the year. The impact of strongUVB and UV erythemally weighted daily dose on life is compounded by broad dailytemperature variations with sudden and sharp fluctuations. Lake habitat chemistry is highlydynamical with notable changes in yearly ion concentrations and pH resulting from lowand variable yearly precipitation. The year-round combination of environmental variablesdefine these lakes as end-members. In such an environment, they host ecosystems thatinclude a significant fraction of previously undescribed species of zooplankton, cyanobacterial,and bacterial populations.
Citation: Cabrol, N. A., et al. (2009), The High-Lakes Project, J. Geophys. Res., 114, G00D06, doi:10.1029/2008JG000818.
1. Introduction
[2] The study of terrestrial analog environments supportsthe design of investigation strategies and life detectionmethods for planetary exploration. Over the past decades,orbital and ground missions have provided geologic, mor-phologic, mineralogic, and chemical lines of evidence show-ing that water was abundant early in the history of Marsand that the planet was habitable for life as we know it [e.g.,
Squyres et al., 2004; Knoll et al., 2005; McLennan et al.,2005; Des Marais et al., 2005]. However, phyllosilicates(clays) disappeared from the stratigraphic record around3.7 Ga ago [Bibring et al., 2005] replaced by sulfate deposits,then by ahydrated ferric oxides (olivine) about �3.2 Ga ago[Christensen et al., 2003; Mustard et al., 2005]. This suc-cession suggests that no further large-scale weathering oc-curred after that period but transient and localized aqueousprocesses likely associated with high-obliquity cycles werestill possible. There is evidence supporting water activity in
JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 114, G00D06, doi:10.1029/2008JG000818, 2009ClickHere
for
FullArticle
1Space Science and Astrobiology Division, NASA Ames ResearchCenter, Moffett Field, California, USA.
2SETI Carl Sagan Center, Mountain View, California, USA.3Centro de Investigacion Cientıfica y Tecnologica para Minerıa, Santiago,
Chile.4Department of Biological Sciences, Carnegie Mellon University,
Pittsburgh, Pennsylvania, USA.5Department of Earth and Space Sciences, University of California,
Los Angeles, California, USA.6Department of Civil and Environmental Engineering, Carnegie Mellon
University, Pittsburgh, Pennsylvania, USA.
Copyright 2009 by the American Geophysical Union.0148-0227/09/2008JG000818$09.00
7Department Botanik, University of Erlangen, Erlangen, Germany.8Centro de Biotecnologıa, Universidad Catolica del Norte, Antofagasta,
Chile.9Marine Research Laboratories, Tasmanian Aquaculture and Fisheries
Institute, University of Tasmania, Hobart, Tasmania, Australia.10CHEP, Santiago, Chile.11Departamento de Quımica, Universidad Catolica del Norte, Antofagasta,
Chile.12Ecole Normale Superieure, Paris, France.13Discoverer Ketty Lund Exploration Vessel, Key West, Florida, USA.14Geoscience Department, University of Iowa, Iowa City, Iowa, USA.15Clınica Mutual de Seguridad, Punta Arenas, Chile.16ESRI, Meudon, France.
G00D06 1 of 20

later geological periods [Carr, 1996;Malin and Edgett, 2000;Baker, 2001; Murray et al., 2005] (and many others) butonly episodically and as lower-magnitude events possiblydriven by magmatic pulses [Dohm et al., 2007] and/oroliquity changes [Haberle et al., 2001; Jakosky et al., 2003].Within the past five years, newly formed deposits in CentauriMontes could be consistent with limited flow in present days,although they do not constitute on their own a definitive proof[Malin et al., 2006].[3] Within that context, and using Earth as an analogy,
lakes on early Mars could be envisioned as favorable sites forlife, if any, its development and preservation in the fossilrecord [e.g., Farmer, 1994, 1995, 2000; Des Marais andFarmer, 1995; Cabrol and Grin, 2001, 2004]. While theyultimately disappeared, how long they remained habitableas their water column shrunk may have controlled life’sability to transition to new environments, i.e., by adaptingto more protected habitats that offered radiation refuge andmoisture retention, such as within sediments, rocks (endoliths),or underground.[4] Understanding better the magnitude and impact of
environmental stress on aquatic life during rapid climatechange and the threshold of habitability in lakes is, thus, partof an essential step in the design of future missions that willsearch for traces of life on Mars. This question motivated thedevelopment of the High Lakes Project (HLP), a multidisci-plinary investigation of high-altitude lakes located between4200m and 6000m elevation in the Central Andes of Boliviaand Chile, in an environment presenting some of the bestterrestrial analogy to early Mars.[5] Here, we present the main results of HLP as of the end
of 2007, especially focusing on the environmental factors thatmake the studied bodies of water unique and representative ofan end-member class of lakes. This article also introducesstudies stemming from HLP and other investigations of highaltitude lakes presented in this Special Issue.[6] Section 2 presents the overall goals and objectives of
HLP, and section 3 the study sites in their regional context.Field and laboratory investigation methods are detailed insection 4. Results are discussed in section 5, i.e., climate(section 5.1), geophysical environment (section 5.2), andchemical evolution (section 5.3). Detailed analyses, com-parison to other terrestrial lake environments, and analogyto Mars can be found in the works of Rose et al. [2009],Dorador et al. [2009], and Farias et al. [2009]. Section 5.4summarizes the results of the biological investigations [e.g.,Dorador et al., 2009; Fleming et al., 2009]. Preliminaryresults on present-day phytoplankton populations were pre-sented by Acs et al. [2003], Kiss et al. [2004], and Cabrolet al. [2007a]. We conclude on how this Mars analog projectis also contributing to a better understanding of the impactof current climate change on Earth’s biosphere.
2. Goals and Objectives
[7] After a first field reconnaissance in 2002, HLP hasbeen supported by the NASA Astrobiology Institute (NAI)since 2003 and has a range of scientific objectives encom-passing terrestrial and planetary sciences. They are to:[8] 1. Explore and characterize the habitability potential of
high altitude lakes in an environment presenting analogy toearly Mars [Cabrol et al., 2009b], including: Low air tem-
perature with high daily amplitude and sudden, sharp, fluc-tuations due to a thin atmosphere (480–600 mb); permanentlow ozone concentration and high solar irradiance; strongnegative water balance; aridity; seasonal ice cover; andsulfur-rich, volcanic, and hydrothermal environment. Thegeophysical environment is coupled with a climate rapidlychanging in the Andes leading to the loss of aquatic habitat[Bradley et al., 2006]. Altogether, these environmentalfactors present a unique analogy to conditions on Mars asthe planet was transitioning from a wetter to a drier climate3.7–3.2 Ga ago. These altiplanic lakes also share with earlyMartian lakes the increased impact of UV radiation in arapidly shrinking water column and the fast modification ontheir water chemistry. As a result, they provide a snapshotin time of lakes approaching threshold conditions for habitatsustainability relevant to early Mars [Cabrol et al., 2007b].[9] 2. Assess the impact of low ozone/high solar irradiance
in nonpolar aquatic environments. Seasonal ozone-hole for-mation over polar and high latitude regions are a cause formajor concern and their impact on terrestrial and aquatic lifeis being closely monitored. HLP brings a new perspective onthe issue by providing data from sites where ozone depletionis a permanent factor and UV radiation substantially strongerthan in polar environments year-round. As such, high-altitude lakes hold critical clues about the threshold of lifehabitat and survivability for other exposed terrestrial aquaticecosystems.[10] 3. Document and map poorly known ecosystems
before they disappear. Because of a rapidly changing climate,many lakes in the Central Andes and Altiplano are likelyto disappear within a few decades if current trends persist.HLP has started to inventory ecosystems that had yet to bedocumented for the most part because of altitude and isola-tion. Not surprisingly, reconnaissance samples showed asignificant fraction of previously undescribed species amongothers already known from South America, Antarctica, andTibet [Fleming et al., 2009; Dorador et al., 2009]. The studyof fossil microbialites associated with paleolake terraces isalso leading to the characterization of biosignatures and geo-signatures of ancient life.[11] 4. Quantify the impact of climate change on lake
environment and ecosystem. Earth’s biosphere is subjectedto a climate change the magnitude of which remains to begrasped. HLP provides representative data on a part of theAndes severely affected by Global Warming. For instance, inthe past 50 years, precipitation has decreased 50% in Chile.Most of the South American glaciers from Columbia,Bolivia, Chile, and Argentina up to 25�S are drasticallyreducing their volume and retreating seven times faster com-pared to 13 years ago [Bradley et al., 2006]. Altitude and siteisolation make such data rare, albeit essential, in the contextof assessing the impact of climate change at global scale.[12] Objectives 2–4 were not originally part of HLP which
was primarily a geophysical investigation of a Mars analog.Rather, they became natural developments of the project overthe years, as monitoring revealed a unique geophysical envi-ronment and mostly uncharted ecosystems.
3. Study Sites
[13] Since 2002, HLP has undertaken both detailed andreconnaissance investigations of eight lakes in the Altiplano
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
2 of 20
G00D06

and high-Andes between 4200 m and 6000 m (Table 1),including geophysical, environmental, limnological, and bio-logical studies. The lakes occupy the summit craters ofvolcanoes, topographic lows, and tectonic basins. They arelocated in the barren altiplanic plateau of the Potosi region insouthwest Bolivia, with the exception of Poquentica 150 kmsouth of La Paz, and in the Chilean Altiplano at the contactwith the Atacama desert (Figure 1).[14] Lake Licancabur (Figure 1a) is located in the sum-
mit crater of a 5970 m high composite volcano constructedlargely in late Pleistocene from basaltic andesitic flows andconsidered potentially active [Marinovic and Lahsen, 1984;de Silva and Francis, 1991]. After a reconnaissance in 2002,a UV dosimeter and a meteorological station (detailedin section 4.1) were positioned on the lake shore in 2003.HLP deployed a scuba diving expedition in 2006 [Morriset al., 2007] which returned sediment and biological sam-ples (zooplankton, phytoplankton, bacteria) allowing a pre-liminary assessment of the lake’s ecosystem.[15] Geological, geophysical, limnological, and biological
investigations have been ongoing at Laguna Verde andLaguna Blanca (Figure 1b) since 2002. The two lakes arelocated at the foot of Licancabur. They were one large singlebody of water at their maximum peak volume at the end ofPleistocene [Risacher and Fritz, 1991; Cabrol et al., 2006].This period is marked in the stratigraphic record by thepresence of a 100 km2 terraced field of fossil microbialitessurrounding the present shores. The two lakes are stillconnected today by a small channel but are already twophysically and limnologically very different bodies of water(section 5.3). A UV dosimeter and a meteorological stationidentical to those of Licancabur were deployed in 2003 on theshore of Laguna Blanca and have been logging data since
with the exception of 2005 due to dosimeter maintenance and2007 as the site was not visited that year. The two lakes aresupplied by hydrothermal springs (Table 1).[16] Escalante (Figure 1c) is a 5700 m high edifice and
is an andesitic-dacitic volcano with Holocene lava flows[Deruelle, 1978; Marinovic and Lahsen, 1984]. It is located20 km north of Licancabur. According to local reports,Escalante was still harboring a small crater lake in 2004.The HLP team only found a dry lakebed in the summit craterduring exploration in 2006. A series of small ponds, mostlikely from seasonal snowmelt, occupy the north and westouter rims and were sampled for water and sediment.[17] Aguas Calientes (Figure 1d) is characterized by red
waters, and this lake is nestled in the summit crater of the5930 m high Aguas Calientes volcano. This volcano is alsoknown as Simba, its native name used hereafter to avoidconfusion with Salar de Aguas Calientes also surveyed byHLP. Simba, a composite cone, is part of a 500 km2 fieldof two intersecting volcanic chains, the Cordon de PuntasNegras and the Cordon Chalviri in Chile. The chains containsmall cones, lava domes, lava flows, maars, and 25 vents,many of those of Holocene age [de Silva and Francis, 1991]or historical such as Lascar [Gonzalez-Ferran, 1995], anactive volcano contiguous to Simba which erupts regularly.Following main wind direction, water vapor clouds arebrought frequently to Simba’s crater lake from Lascar alongwith occasional sulfuric acid clouds. Simba has no record ofhistoric activity. The most recent porphyritic andesite lavaflows are Holocene but most of the volcano’s history is pre-Holocene [Ramırez and Gardeweg, 1982]. Its investigationstarted in 2006 with the deployment of a geophysical stationand the sampling of water and lake sediment for microbialpopulation survey. In 2007, the UV dosimeter was retrieved
Table 1. Sites Characteristics
Physical ParameterLag.a
BlancaLag.Verde
Lag.Lejıa
Salar deAguas Calientes
LicancaburLake (v)b Simba (v)
PoquenticaLake (v)
EscalantePonds
Latitude (�S) 22.47.00 22.47.32 23.30.00 23.07.00 22.50.07 23.21.47 18.44.05 22.72.05Longitude (�W) 67.47.00 67.49.16 67.42.00 67.25.00 67.53.07 67.40.55 62.58.28 22.43.10Lake elevation (m) 4340 4340 4325 4200 6000 5870 5750 5700Area (m2) 3,5�106 7,5�106 2�106 134�106 7�103 2.5�103 2,5�103 5�102Depth (m) 0.5 5.4 1.2 0.5 5.2 TBD TBD 0.3UVR (% sea level) 140 140 142 145 165 158 155 150Maximum air temperature (�C) +13 +10 [12]c [13] +9 +9 [�25] [7.7]Minimum air temperature (�C) �25 �30 �28 TBD �40 TBD no data no dataPd (mm/yr) e e f 155/146 98/118 146/146 871/837 e
Water temperatureg (�C) 12–36 13–20 3–10.6 8–18h 4.3–14 [9] [4] [1]pH 7.2–8.42i 9.0–8.19i 6.9j 6.9j 8.4–6.9k 4.9–3.9l 5m 7n
Ice cover variable rare TBD TBD Apr/Sep Apr/Sep Apr/Sep Apr/NovInvestigation type station station reconnaissance reconnaissance station station reconnaissance reconnaissanceInvestigation start date 2002 2002 2007 2007 2002 2006 2005 2006Country Bolivia Bolivia Chile Chile Bolivia/Chile Chile Bolivia/Chile Bolivia
aLaguna.bVolcano summit crater lake.cBrackets indicate that data are from point measurements collected during austral spring. No yearly data integration.dAverage yearly precipitation (median/mean) between 2002 and 2007 (monthly TRMM 3B43 V6, see section 4.1).eValues included in the cell resolution for Licancabur.fValues included in the cell resolution for Simba.gIncludes spring and lake water temperature.hThe range is representative of the springs documented in 2007.iMeasurements made in 2002–2004.jMeasurements made in 2007.kMeasurements made in 2002–2006.lMeasurements made in 2006–2007.mMeasurements made in 2005.nMeasurements made in 2006.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
3 of 20
G00D06

and replaced by a station dedicated to short UV wavelengths(section 4.1). The 2007 investigation was interrupted by the7.8 magnitude earthquake that struck Chile on 15 November,related avalanches, and threatening venting of Lascar. Themicrobial population is currently being inventoried [Escuderoet al., 2007; Dorador et al., 2009].[18] Laguna Lejıa (Figure 1e) is located 10 km south of
Simba, Lejıa and is one of the two most recent investigationsites. Water, sediment, phytoplankton, and zooplankton weresampled for the first time in 2007. Like Laguna Verde andLaguna Blanca, the lake has extensive paleo-terraces show-ing a pre-Holocene lake about five times larger and 15–25 mdeeper than at present [Grosjean, 1994; Risacher et al., 1999,2003]. The terraces are covered by fossil microbialites, coral-like, and rounded stromatolitic structures. In addition toaridity and high solar irradiance common to all studied areas,Laguna Lejıa’s ecosystem is impacted by the regular erup-tions of Lascar only 5 km to the north. The latest majoreruption of April 1993 released large amounts of ash andpyroclast into the lake. Thousands of altiplanic (so-called icy)flamingoes grazed in Lejıa prior to the eruption and did notreturn until a few years ago. Small groups were observedduring the 2007 field campaign suggesting ecosystem recov-ery although plankton netting retrieved very few specimenof zooplankton even on grazing spots.[19] Salar de Aguas Calientes (Figure 1f) is located 30 km
southeast of Simba, and it is the largest lake investigated(Table 1). The first reconnaissance took place in November
2007 [Dorador et al., 2009]. A substantial part of the lake’surface area is now dry. Many small lateral ponds, not all ofthem connected to the main body of water, are supplied byhydrothermal springs (Table 1). Plankton netting resulted inthe capture of an abundant population of zooplankton com-parable in density to that of Laguna Blanca and Licancaburin Bolivia (section 5.4).[20] Lake Poquentica, situated 150 km southwest of La Paz
in Bolivia, was explored only once by HLP in 2005 and isnestled in the summit crater of the Poquentica volcano at5850 m. Other programmatic priorities have prevented HLPfrom returning so far and because of limited data, Poquenticais not discussed here.
4. Field and Laboratory Methods
[21] Data collection includes field samples (rocks, sedi-ment, water, macrobial and microbial life), geophysical andmeteorological data compiled year-round from field dosim-eters, data loggers and stations, point measurements collectedduring field campaigns, andmodeling [Hock, 2008]. The datalibrary is completed by using 0.25� � 0.25� resolution dataproducts and raw data from the OzoneMonitoring Instrument(OMI) onboard the NASA Aura spacecraft. Erythemal UV ismonitored with Eldonet stations on the ground (section 4.1)and OMI data for regional context. Yearly precipitation issurveyed through the analysis of the Monthly Tropical Rain-fall Measuring Mission data (TRMM 3B43 V6) from the
Figure 1. (a) Licancabur summit lake from the summit rim on the Chilean side (2004); (b) (left) LagunaVerde and (right) Laguna Blanca as viewed from the summit of Licancabur (2004); (c) Escalante main pond(2006); (d) Simba summit lake (2006); (e) Laguna Lejıa and, in the background, the volcanoes (left) Lascarventing water vapor, (middle) Simba, and (right) Acamarachi (2007); (f) south margin of the Salar de AguasCalientes (2007). See Table 1 for details about location, dimensions, and other characteristics for each lake.Credit photos: High Lakes Project.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
4 of 20
G00D06
TRMMOnline Visualization andAnalysis System (TOVAS).TOVAS was created and is supported by the NASA GoddardSpace Flight Center (GSFC) Sciences Data and InformationServices Center (GES DISC). Satellite imagery (15–90 m/pxlresolution ASTER Vis/NIR/TIR 15 channels and Shuttleimages) are used to identify paleoshores and terraces whererelevant [Cabrol et al., 2003, 2006, 2007b].
4.1. Geophysics and Meteorology
[22] Solar irradiance is monitored with Eldonet dosimeters[Hader and Lebert, 2006]. These ground stations are three-channel wave band dosimeters with filter functions cover-ing the UVB (280–315 nm (Commission Internationaled’Eclairage, CIE, definition), UVA (315–400 nm), andPhotosynthetic Active Radiation, PAR, (400–700 nm). Theyalso log temperature. The dosimeters measure the irradiancesin the defined wavelength band. The filter function matchesthe selected wavelengths. The UVB filter has a peak wave-length at 296 nm, a half-band of 30 nm, a tenth width of55 nm, maximal transmission of 24%, and a diameter of25mm.TheUVAand PAR filters are broadband filters 20mmin diameter. PAR covers the visible range (93% transmission,50% average 450–675 m, 92% transmission at 722 nm and84% at 531 nm). Additional details including calibration,conversion to erythemal UV, and description of the EldonetNetwork are found in the works of Hader et al. [1999] andHader and Lebert [2006].[23] Two instruments are deployed on the shores of the
Licancabur lake and at Laguna Blanca at the foot of theLicancabur volcano. The location of the two dosimetersseparated by 1,500 m in elevation in the same area wasselected in 2003 to analyze the variation of UV radiation withelevation [Hock, 2008]. In 2006, a similar instrument waspositioned at the summit of Simba. In collaboration with theUniversity of Erlangen, Germany, HLP deployed anotherdosimeter in 2007 on this volcano, this one dedicated to 260–270 nm after detection of short UV wavelengths with SolarLight Company (SLC) sensors in 2006 [Cabrol et al., 2008,2009a]. UV diffuse attenuation coefficient (kd) in the waterfor UVA,UVB, and PAR ismeasured using SLC sensors withPAR (400–700 nm), UVA (320–420 nm), and UVB (280–
370 nm). Temperature, relative humidity, and wind speed aresurveyed with HOBO 12-channel meteorological stationslocated at the summit of Licancabur and at Laguna Blanca.
4.2. Water Chemistry
[24] Water sampling is performed on a regular base duringfield campaigns, always in November, and at identicalsampling spots to prevent data discrepancy from location.In situ and laboratory measurements include pH, tempera-ture, water transparency from Secchi disk with calibratedline, total dissolved solids (TDS), ionic concentration, nutri-ent, metal, specific conductance, silicon, dissolved inorganiccarbon (DIC) and dissolved organic carbon (DOC). Mostanalyses are outsourced to Actlabs (Ontario).
4.3. Bathymetry
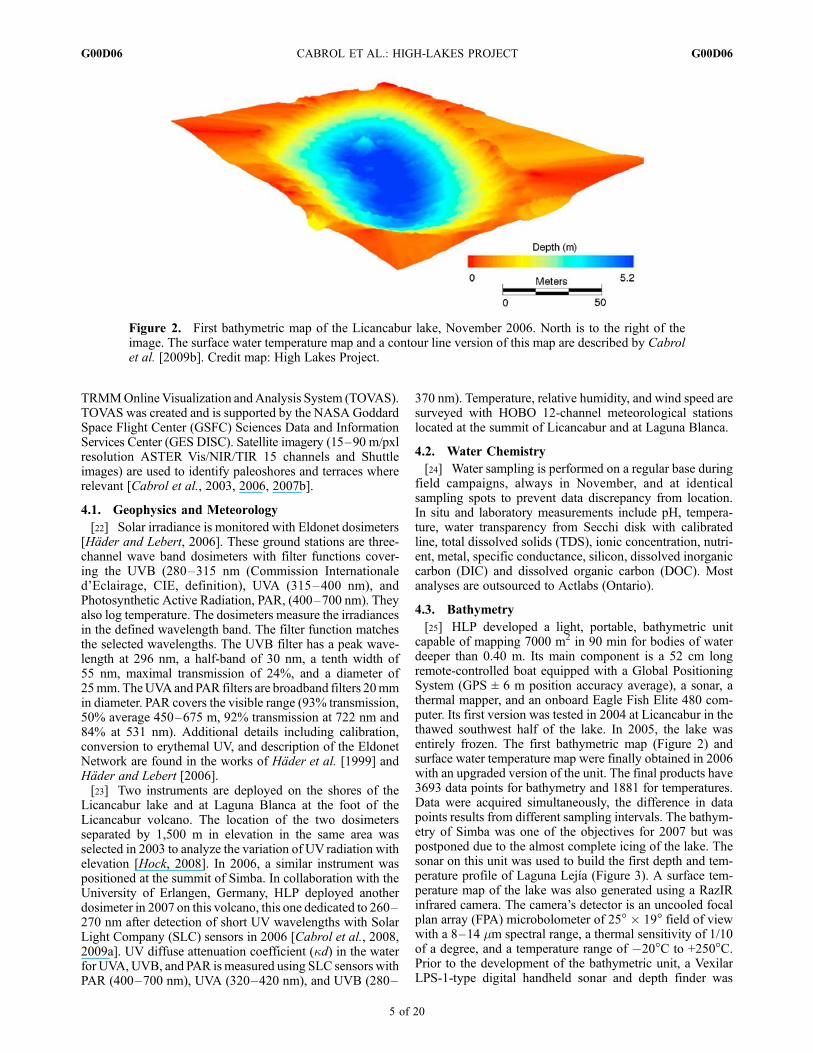
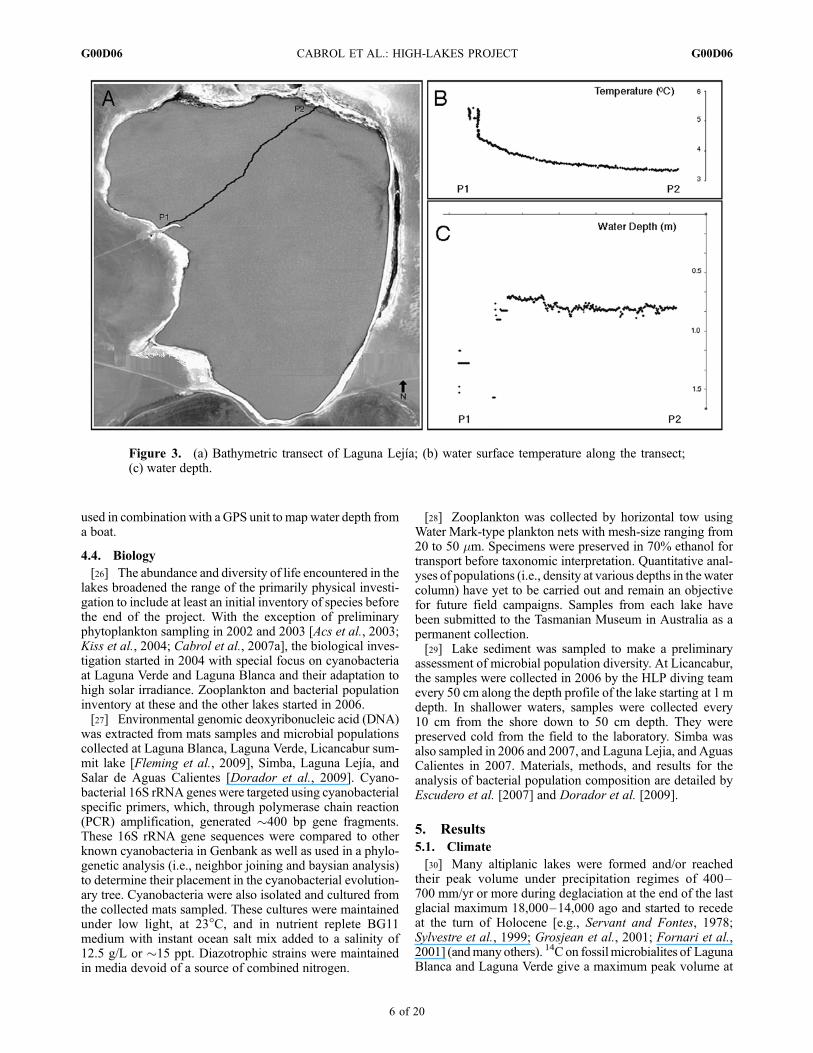
[25] HLP developed a light, portable, bathymetric unitcapable of mapping 7000 m2 in 90 min for bodies of waterdeeper than 0.40 m. Its main component is a 52 cm longremote-controlled boat equipped with a Global PositioningSystem (GPS ± 6 m position accuracy average), a sonar, athermal mapper, and an onboard Eagle Fish Elite 480 com-puter. Its first version was tested in 2004 at Licancabur in thethawed southwest half of the lake. In 2005, the lake wasentirely frozen. The first bathymetric map (Figure 2) andsurface water temperature map were finally obtained in 2006with an upgraded version of the unit. The final products have3693 data points for bathymetry and 1881 for temperatures.Data were acquired simultaneously, the difference in datapoints results from different sampling intervals. The bathym-etry of Simba was one of the objectives for 2007 but waspostponed due to the almost complete icing of the lake. Thesonar on this unit was used to build the first depth and tem-perature profile of Laguna Lejıa (Figure 3). A surface tem-perature map of the lake was also generated using a RazIRinfrared camera. The camera’s detector is an uncooled focalplan array (FPA) microbolometer of 25� � 19� field of viewwith a 8–14 mm spectral range, a thermal sensitivity of 1/10of a degree, and a temperature range of �20�C to +250�C.Prior to the development of the bathymetric unit, a VexilarLPS-1-type digital handheld sonar and depth finder was
Figure 2. First bathymetric map of the Licancabur lake, November 2006. North is to the right of theimage. The surface water temperature map and a contour line version of this map are described by Cabrolet al. [2009b]. Credit map: High Lakes Project.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
5 of 20
G00D06
used in combination with a GPS unit to map water depth froma boat.
4.4. Biology
[26] The abundance and diversity of life encountered in thelakes broadened the range of the primarily physical investi-gation to include at least an initial inventory of species beforethe end of the project. With the exception of preliminaryphytoplankton sampling in 2002 and 2003 [Acs et al., 2003;Kiss et al., 2004; Cabrol et al., 2007a], the biological inves-tigation started in 2004 with special focus on cyanobacteriaat Laguna Verde and Laguna Blanca and their adaptation tohigh solar irradiance. Zooplankton and bacterial populationinventory at these and the other lakes started in 2006.[27] Environmental genomic deoxyribonucleic acid (DNA)
was extracted from mats samples and microbial populationscollected at Laguna Blanca, Laguna Verde, Licancabur sum-mit lake [Fleming et al., 2009], Simba, Laguna Lejıa, andSalar de Aguas Calientes [Dorador et al., 2009]. Cyano-bacterial 16S rRNA genes were targeted using cyanobacterialspecific primers, which, through polymerase chain reaction(PCR) amplification, generated �400 bp gene fragments.These 16S rRNA gene sequences were compared to otherknown cyanobacteria in Genbank as well as used in a phylo-genetic analysis (i.e., neighbor joining and baysian analysis)to determine their placement in the cyanobacterial evolution-ary tree. Cyanobacteria were also isolated and cultured fromthe collected mats sampled. These cultures were maintainedunder low light, at 23�C, and in nutrient replete BG11medium with instant ocean salt mix added to a salinity of12.5 g/L or �15 ppt. Diazotrophic strains were maintainedin media devoid of a source of combined nitrogen.
[28] Zooplankton was collected by horizontal tow usingWater Mark-type plankton nets with mesh-size ranging from20 to 50 mm. Specimens were preserved in 70% ethanol fortransport before taxonomic interpretation. Quantitative anal-yses of populations (i.e., density at various depths in the watercolumn) have yet to be carried out and remain an objectivefor future field campaigns. Samples from each lake havebeen submitted to the Tasmanian Museum in Australia as apermanent collection.[29] Lake sediment was sampled to make a preliminary
assessment of microbial population diversity. At Licancabur,the samples were collected in 2006 by the HLP diving teamevery 50 cm along the depth profile of the lake starting at 1 mdepth. In shallower waters, samples were collected every10 cm from the shore down to 50 cm depth. They werepreserved cold from the field to the laboratory. Simba wasalso sampled in 2006 and 2007, and Laguna Lejia, and AguasCalientes in 2007. Materials, methods, and results for theanalysis of bacterial population composition are detailed byEscudero et al. [2007] and Dorador et al. [2009].
5. Results
5.1. Climate
[30] Many altiplanic lakes were formed and/or reachedtheir peak volume under precipitation regimes of 400–700 mm/yr or more during deglaciation at the end of the lastglacial maximum 18,000–14,000 ago and started to recedeat the turn of Holocene [e.g., Servant and Fontes, 1978;Sylvestre et al., 1999; Grosjean et al., 2001; Fornari et al.,2001] (andmanyothers). 14C on fossilmicrobialites of LagunaBlanca and Laguna Verde give a maximum peak volume at
Figure 3. (a) Bathymetric transect of Laguna Lejıa; (b) water surface temperature along the transect;(c) water depth.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
6 of 20
G00D06

13,240 BP [Cabrol et al., 2006] consistent with previouswork of Risacher et al. [1999, 2003] showing high waterlevels at Laguna Lejıa in Chile during the same period. Theseresults support a more humid climate over the Central Andesduring pre-Holocene. Radiocarbon and stratigraphy alsosuggest an increased water loss 8,290 BP-6,300 BP in theLicancabur area, with an average of 24 m in 2,000 years(1.2 mm/yr) compared to 0.28 mm/yr 13,240 BP-8,290 BP[Cabrol et al., 2006]. This decline is currently accelerated byincreased evaporation [Hock, 2008] associated with GlobalWarming [Bradley et al., 2006]. Conditions at each sites werefound to be dictated mainly by altitude, the yearly reach of thealtiplanic winter, the proximity to the Atacama desert (40 kmon average), and to a lesser extent, winter weather systemsfrom the Pacific ocean.[31] Total yearly precipitation is �100 mm/yr on average
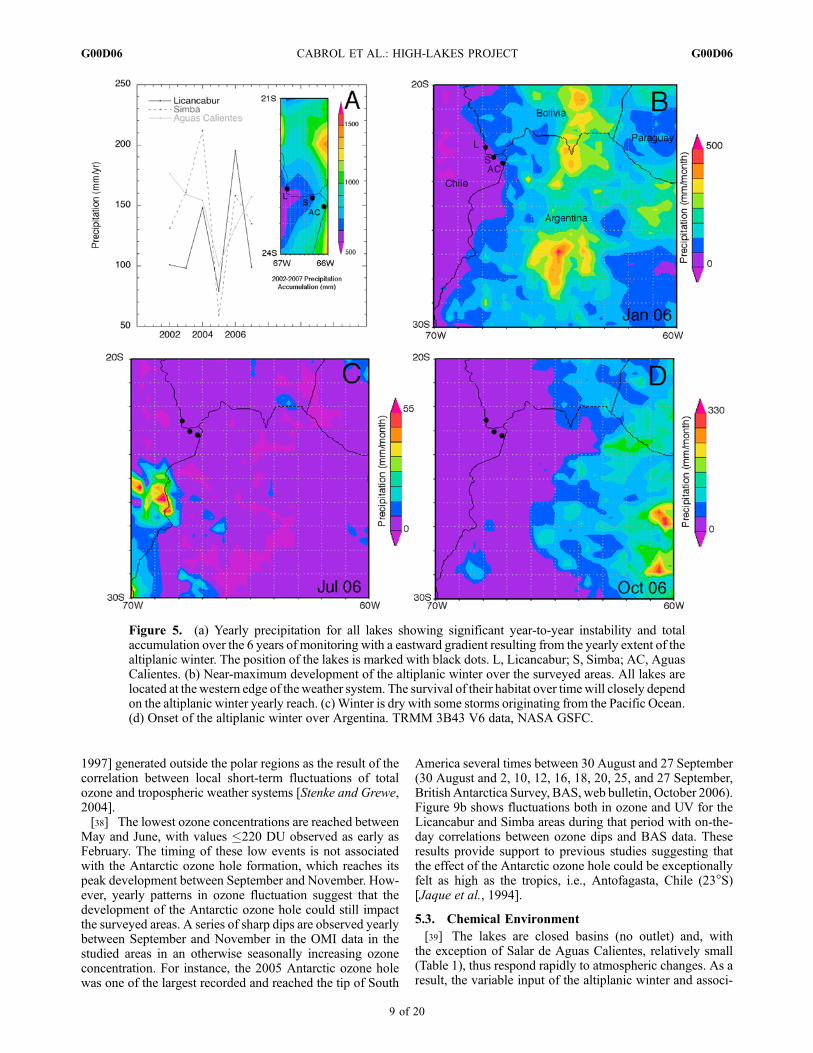
in the Licancabur area with exceptionally dry or wet years(76–195 mm), 146 mm/year, and 155 mm/yr at Simbaand Aguas Calientes, respectively (Figure 4). Months withno measurable precipitation are common at Licancabur andSimba. Most of the precipitation accumulation occurs duringaustral spring and summer as the so-called altiplanic winterbrings humid air originating from Argentina to the area(Figure 5). Altiplanic winter, here, is a local term used onlyto refer to increased precipitation, not to the season. Thelocation of the lakes at the western fringe of its zone ofinfluence and near the Atacama makes its yearly fluctuations(magnitude and wind direction) key to the survival of theseaquatic habitats. Between 2002 and 2007, the sites accumu-lated an average of 80% of their total yearly precipitationfrom this event alone (Figure 4) and, although they areonly dispersed within half a degree in latitude and longitude,they show a noticeable increase in precipitation eastwardly(Figure 5a).[32] Evaporation was modeled following Hamon’s [1961]
method using air temperature, relative humidity, and watertemperature data from the Licancabur and Laguna Blancameteorological stations and underwater data loggers. Mod-eling methods and results are detailed by Hock [2008] andpredict a strong, 500–1000 mm/yr, negative water balance.While substantial evaporation was confirmed through obser-vation (Figure 6), the trend is not monotonical. In spring2006, after several years of consecutive decline, the waterlevel of Licancabur, Laguna Blanca, and Laguna Verderose back higher than their 2002 level following the mostabundant precipitation accumulation in 5 years (195 mm, theequivalent to two regular years).[33] Six year of precipitation survey suggest that drier
years might be the result of a late onset of the altiplanicwinter. Depending on location, the sites are also affecteddifferently by the winter Pacific storms (June–August). Theirinfluence is more pronounced at Licancabur than at Simba orthe more eastern Salar de Aguas Calientes, with a maximumof 20%, 9.4%, and 3.5 of their yearly total accumulation,respectively (Figures 4 and 5). This suggests that the smallamount of moisture crossing the Atacama immediatelyprecipitates as it ascends the Andean peaks and does notreach much farther inland. The proximity of the Atacama alsobrings sustained aridity with low (30%) yearly average rela-tive humidity compounded by desiccation from daily windsregularly reaching 40–60 km/h with higher gusts.
5.2. Physical Environment
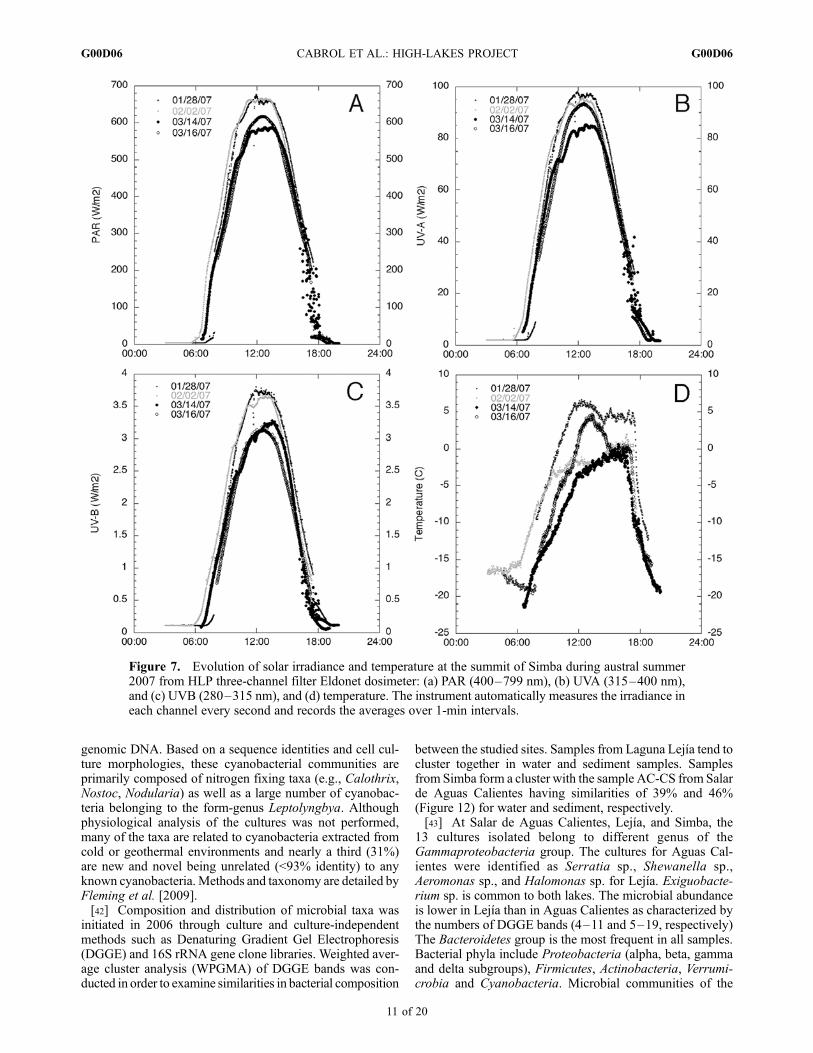
[34] Average cold temperatures, strong solar irradiance,and permanent ozone depletion characterize the environmentat all lakes. The meteorological stations recorded yearlytemperature extremes of �40�C to +5�C at Licancabur and�25�C to +13�C at Laguna Blanca. Out-of-normal rangetemperatures were occasionally monitored during field cam-paigns. Unusually warm or cold temperatures were recordedduring theNovember field campaigns, i.e., +9�C at Licancabur(6000 m) at noon and +18�C at Laguna Blanca (4340 m) at2:00 P.M. in 2006, and night temperatures between �18�Cto�25�C on the Altiplano near Lejıa (4325 m) over a periodof 10 days in 2007. Yearly surveys are discussed by Hock[2008].[35] Maximum (hourly) averaged PAR, UVA, and UVB
reached up to 663 W/m2, 95 W/m2, and 3.7 W/m2, respec-tively, at the summit of Simba in 2007 (Figures 7a–7c).Although 165% that of sea level, these values are still not thehighest expected at this site as the data shown is for January–March 2007. Other data including summer solstice are cur-rently under review. Maximum erythemally weighted dailydose at Laguna Blanca was 17 KJ/m2 and 23 KJ/m2 atLicancabur, three times the values recorded by OMI for thisregion and latitude. Short UV wavelengths (260–270 nm)peaked at 9.6 and 14.6 mW/m2 between noon and 1:00 P.M.at Licancabur and Simba, respectively [Cabrol et al., 2008].[36] Temperature and UV vary along the same gradients
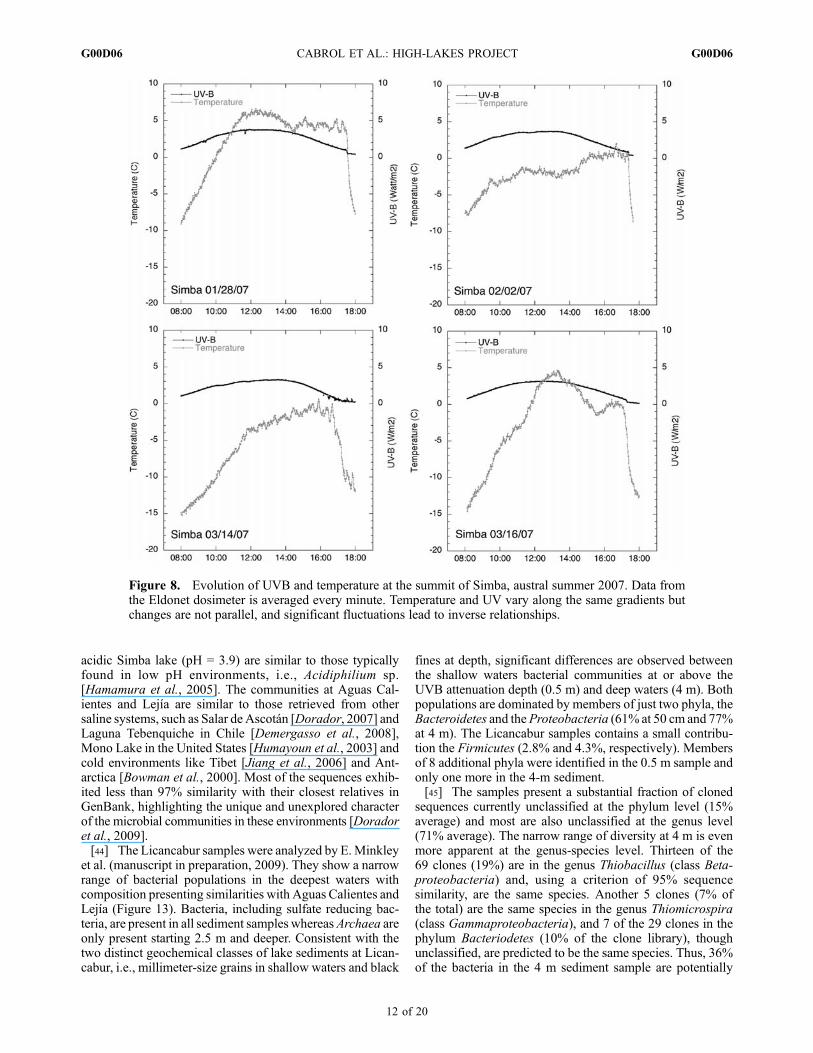
but changes are not parallel over space and time. Suddentemperature variations in scattered cloud skies are common inthe 480–600mb atmosphere (Figure 8). Inverse relationshipsbetween UVand temperature are observed during those fluc-tuations. At Simba, summer 2007 showed daily temperaturegradients between 3.75 and 6.3�C per hour, with significantfluctuation and recovery up to 2�Cwithin oneminute in somecases. Previous studies have shown that such fluctuationssubject aquatic ecosystems to high and unpredictable UV/Tratios, impairing the ability of microbial organisms to repairDNA [e.g.,Garcia-Pichel et al., 1994; Karentz, 1994; Santaset al., 1998; Cockell and Blaustein, 2001; Willamson et al.,2002]. As a result, a working hypothesis was that the impacton ecosystems would be more significant in lakes having theshallowest water column, possibly in terms of abundanceand/or diversity of species. Observation and data suggest amore complex process.While UVand temperature are criticalfactors, other environmental conditions such as lake chem-istry, and the timing and interplay of environmental cycles areequally important for habitat survival (see discussion).[37] Altitude and latitude generate an atmosphere perma-
nently depleted in ozone. The average yearly total ozoneconcentration in the Licancabur area (the OMI cell resolutionincludes Laguna Blanca and Laguna Verde) and the Simbaarea (includes Laguna Lejıa and Aguas Calientes) is compa-rable with 247 and 250 Dobson Units (DU), respectively(Figure 9a). Both regions show between 30 and 40% deple-tion (260–220 DU) for 76.5% of the year. OMI daily min-imum ozone data show that ozone hole definition (�220 DU)is reached over a month per year (33–36 days) on aver-age. The lowest value recorded in the Licancabur area was205 DU on 10 March 2006. Daily fluctuations and recoverypatterns [Cabrol et al., 2009b] are consistent with models ofozoneminihole formation [Newman et al., 1988; James et al.,
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
7 of 20
G00D06
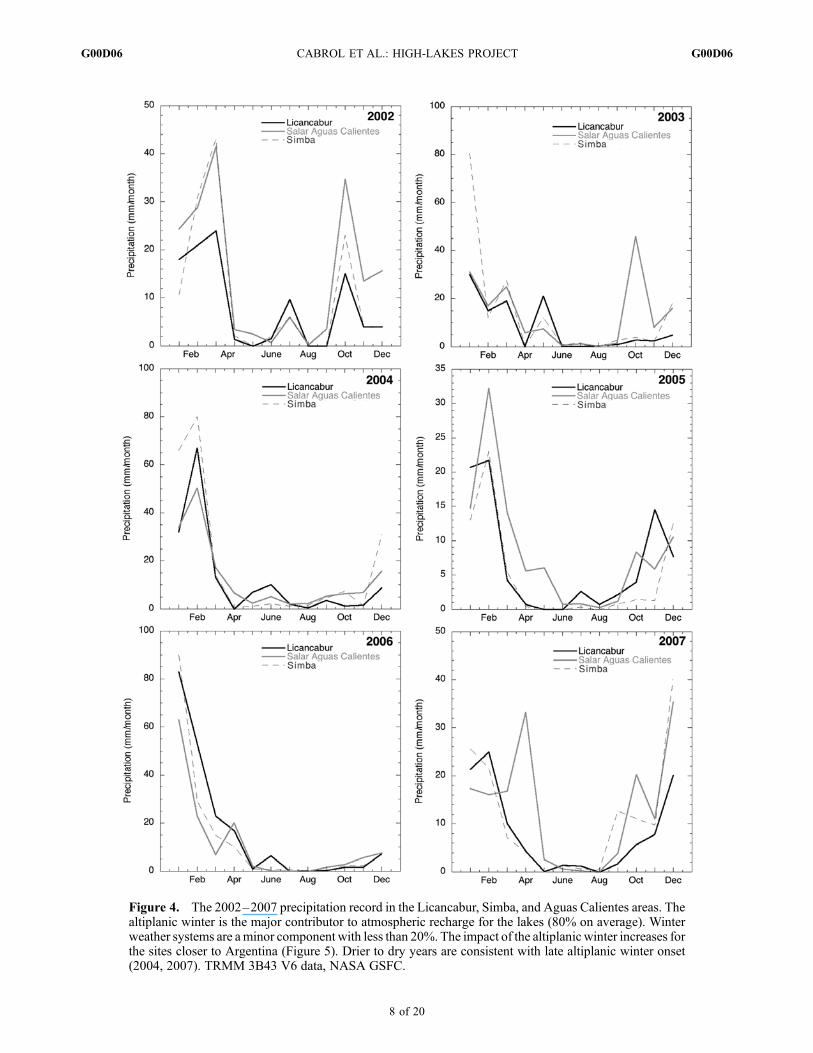
Figure 4. The 2002–2007 precipitation record in the Licancabur, Simba, and Aguas Calientes areas. Thealtiplanic winter is the major contributor to atmospheric recharge for the lakes (80% on average). Winterweather systems are aminor component with less than 20%. The impact of the altiplanic winter increases forthe sites closer to Argentina (Figure 5). Drier to dry years are consistent with late altiplanic winter onset(2004, 2007). TRMM 3B43 V6 data, NASA GSFC.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
8 of 20
G00D06
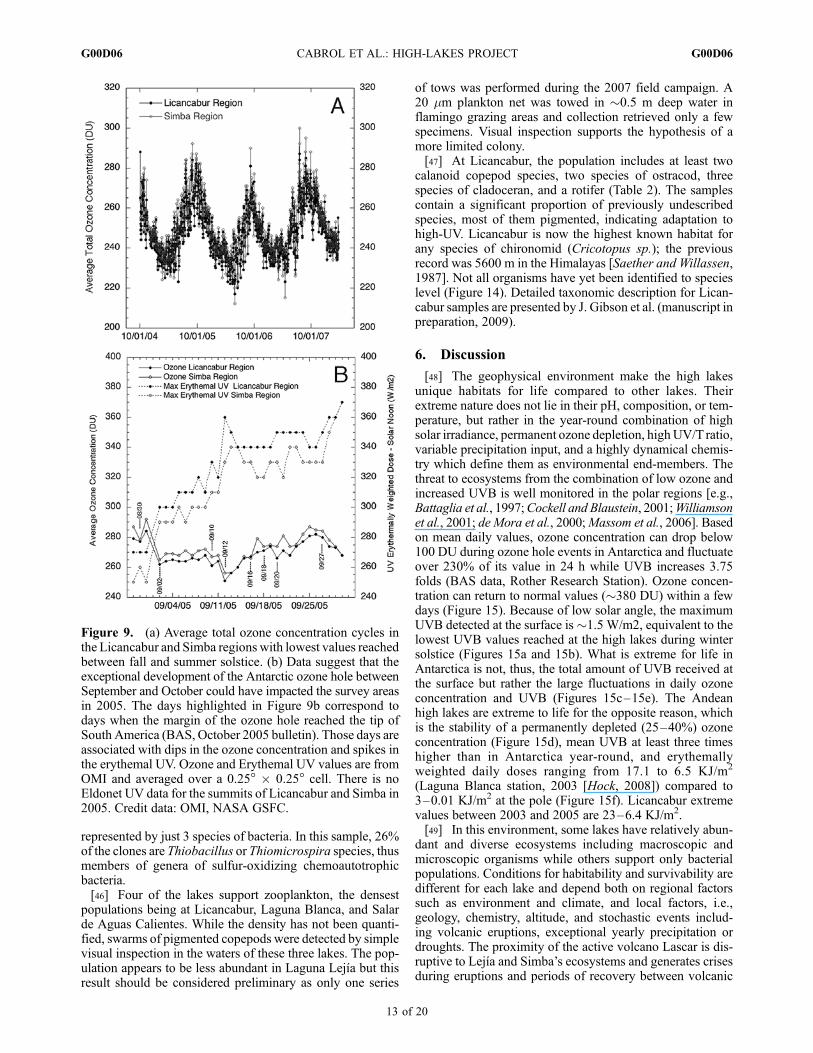
1997] generated outside the polar regions as the result of thecorrelation between local short-term fluctuations of totalozone and tropospheric weather systems [Stenke and Grewe,2004].[38] The lowest ozone concentrations are reached between
May and June, with values �220 DU observed as early asFebruary. The timing of these low events is not associatedwith the Antarctic ozone hole formation, which reaches itspeak development between September and November. How-ever, yearly patterns in ozone fluctuation suggest that thedevelopment of the Antarctic ozone hole could still impactthe surveyed areas. A series of sharp dips are observed yearlybetween September and November in the OMI data in thestudied areas in an otherwise seasonally increasing ozoneconcentration. For instance, the 2005 Antarctic ozone holewas one of the largest recorded and reached the tip of South
America several times between 30 August and 27 September(30 August and 2, 10, 12, 16, 18, 20, 25, and 27 September,British Antarctica Survey, BAS, web bulletin, October 2006).Figure 9b shows fluctuations both in ozone and UV for theLicancabur and Simba areas during that period with on-the-day correlations between ozone dips and BAS data. Theseresults provide support to previous studies suggesting thatthe effect of the Antarctic ozone hole could be exceptionallyfelt as high as the tropics, i.e., Antofagasta, Chile (23�S)[Jaque et al., 1994].
5.3. Chemical Environment
[39] The lakes are closed basins (no outlet) and, withthe exception of Salar de Aguas Calientes, relatively small(Table 1), thus respond rapidly to atmospheric changes. As aresult, the variable input of the altiplanic winter and associ-
Figure 5. (a) Yearly precipitation for all lakes showing significant year-to-year instability and totalaccumulation over the 6 years of monitoring with a eastward gradient resulting from the yearly extent of thealtiplanic winter. The position of the lakes is marked with black dots. L, Licancabur; S, Simba; AC, AguasCalientes. (b) Near-maximum development of the altiplanic winter over the surveyed areas. All lakes arelocated at the western edge of the weather system. The survival of their habitat over timewill closely dependon the altiplanic winter yearly reach. (c) Winter is dry with some storms originating from the Pacific Ocean.(d) Onset of the altiplanic winter over Argentina. TRMM 3B43 V6 data, NASA GSFC.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
9 of 20
G00D06

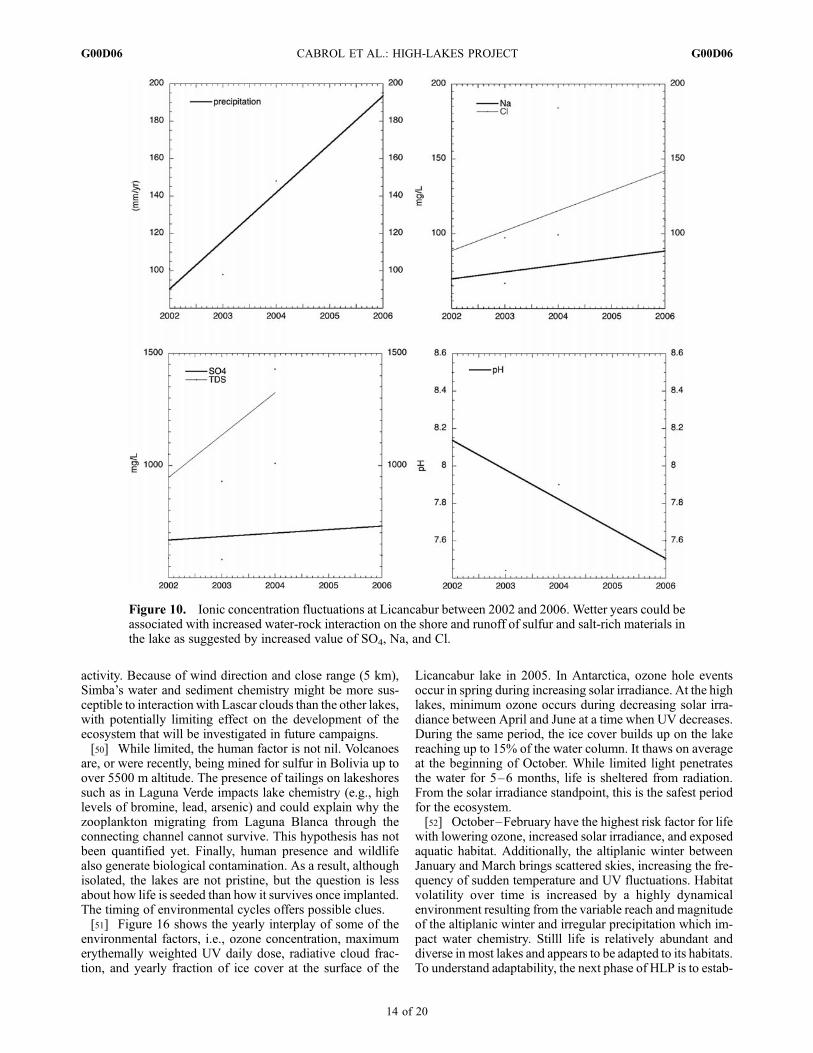
ated hydrological disequilibrium generates a dynamicalchemical habitat. This is confirmed bymeasurements of ionicconcentrations at Licancabur (Figure 10) which follow thevariation in yearly precipitation and suggest increased water-rock interaction during wetter years (increased Na, Cl, TDS,SO4). The pH of Licancabur, Laguna Verde, and LagunaBlanca is also variable. Yearly in situ measurements (2002–2004) indicate 8.4–7.6 at Licancabur; 9.0–8.19 at LagunaVerde. The trend is reversed for Laguna Blanca, with 7.2–8.42. Increase is also notable in the amount of Total Dis-solved Solids (TDS in mg/L): 22,400–23,050 for LagunaBlanca; 117,500–199,000 for Laguna Verde, and 1050–1368 for the Licancabur summit lake. Unlike the other twolakes, Laguna Blanca is becoming more basic while Lican-cabur and Laguna Verde are becoming more acidic. In somecases, changes in pH appear to be positively correlated tochanges in TDS, thus likely linked to climate variability (e.g.,precipitation), while in others this correlation appears to benegative or not well determined. This suggests that otherfactors, i.e., geothermal input through spring discharge, arelikely to also play a role that remains to be quantified.[40] Chemical composition also dictates how UV pene-
trates the water column. Everything being equal, deeperwaters provide a better shelter for life that can migrate inthe water column as a strategy against high solar irradiance[e.g., Hargreaves, 2003; Boeing et al., 2004; Leech et al.,2005a, 2005b; Fischer et al., 2006]. However, water columndepth does not appear to be the primary factor for surviv-ability in all surveyed lakes (see discussion). Laguna Blancaand Licancabur have equally relatively abundant and diverse
ecosystems; the former is the shallowest lake studied (0.5 macross) and the latter is 10 times deeper. Licancabur has17 times less TDS, clear waters, and a dissolved organiccarbon (DOC) concentration of 2.44 mg L�1. As a result, UVattenuation occurs higher in the water column at LagunaBlanca. The diffuse attenuation coefficient (kd) for thevarious UV wavelengths are (Licancabur/Laguna Blanca):kdUVB = 0.53/0.077 m; kdUVA = 1.10/0.28 m; kdPAR = 10.1/0.88 m [Hock, 2008]. For both lakes, kPAR actually occursdeeper than the current maximum depth. As a result, photo-synthesis is still possible without exposure to harmful radia-tion below kdUVB. In other lakes, composition overridesdepth, such as in Laguna Verde (5.4 m) where zooplanktonmigrating fromLaguna Blanca does not survive (see section 6).Water chemistry and solar irradiance are discussed by A. Hocket al. (manuscript in preparation, 2009).
5.4. Life
[41] Despite high solar irradiance and high UV/T ratio,all lakes have developed abundant pigmented periphytonabove 0.5 m depth. Cyanobacterial composition of a varietyof microbial mats present in Laguna Blanca, Laguna Verdeand Licancabur were characterized (Figure 11). Multiplesamples were collected from different mat communities inand around these lakes, which include those from the outflowof a spring north of Laguna Verde, mats along a hydrationtransect in Laguna Blanca, and benthic mats from theLicancabur summit lake. From these samples, 78 cyanobac-terial cultures were isolated in addition to �600 cyanobacte-rial 16s rRNA gene sequences extracted from environmental
Figure 6. Fluctuation of water level at Licancabur from (top) 2002 to 2006 with low precipitation wellmarked in 2005, when the lake reached its lowest level. Credit photo: High Lakes Project.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
10 of 20
G00D06
genomic DNA. Based on a sequence identities and cell cul-ture morphologies, these cyanobacterial communities areprimarily composed of nitrogen fixing taxa (e.g., Calothrix,Nostoc, Nodularia) as well as a large number of cyanobac-teria belonging to the form-genus Leptolyngbya. Althoughphysiological analysis of the cultures was not performed,many of the taxa are related to cyanobacteria extracted fromcold or geothermal environments and nearly a third (31%)are new and novel being unrelated (<93% identity) to anyknown cyanobacteria. Methods and taxonomy are detailed byFleming et al. [2009].[42] Composition and distribution of microbial taxa was
initiated in 2006 through culture and culture-independentmethods such as Denaturing Gradient Gel Electrophoresis(DGGE) and 16S rRNA gene clone libraries. Weighted aver-age cluster analysis (WPGMA) of DGGE bands was con-ducted in order to examine similarities in bacterial composition
between the studied sites. Samples from Laguna Lejıa tend tocluster together in water and sediment samples. Samplesfrom Simba form a cluster with the sample AC-CS from Salarde Aguas Calientes having similarities of 39% and 46%(Figure 12) for water and sediment, respectively.[43] At Salar de Aguas Calientes, Lejıa, and Simba, the
13 cultures isolated belong to different genus of theGammaproteobacteria group. The cultures for Aguas Cal-ientes were identified as Serratia sp., Shewanella sp.,Aeromonas sp., and Halomonas sp. for Lejıa. Exiguobacte-rium sp. is common to both lakes. The microbial abundanceis lower in Lejıa than in Aguas Calientes as characterized bythe numbers of DGGE bands (4–11 and 5–19, respectively)The Bacteroidetes group is the most frequent in all samples.Bacterial phyla include Proteobacteria (alpha, beta, gammaand delta subgroups), Firmicutes, Actinobacteria, Verrumi-crobia and Cyanobacteria. Microbial communities of the
Figure 7. Evolution of solar irradiance and temperature at the summit of Simba during austral summer2007 from HLP three-channel filter Eldonet dosimeter: (a) PAR (400–799 nm), (b) UVA (315–400 nm),and (c) UVB (280–315 nm), and (d) temperature. The instrument automatically measures the irradiance ineach channel every second and records the averages over 1-min intervals.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
11 of 20
G00D06
acidic Simba lake (pH = 3.9) are similar to those typicallyfound in low pH environments, i.e., Acidiphilium sp.[Hamamura et al., 2005]. The communities at Aguas Cal-ientes and Lejıa are similar to those retrieved from othersaline systems, such as Salar de Ascotan [Dorador, 2007] andLaguna Tebenquiche in Chile [Demergasso et al., 2008],Mono Lake in the United States [Humayoun et al., 2003] andcold environments like Tibet [Jiang et al., 2006] and Ant-arctica [Bowman et al., 2000]. Most of the sequences exhib-ited less than 97% similarity with their closest relatives inGenBank, highlighting the unique and unexplored characterof the microbial communities in these environments [Doradoret al., 2009].[44] The Licancabur samples were analyzed by E.Minkley
et al. (manuscript in preparation, 2009). They show a narrowrange of bacterial populations in the deepest waters withcomposition presenting similarities with Aguas Calientes andLejıa (Figure 13). Bacteria, including sulfate reducing bac-teria, are present in all sediment samples whereas Archaea areonly present starting 2.5 m and deeper. Consistent with thetwo distinct geochemical classes of lake sediments at Lican-cabur, i.e., millimeter-size grains in shallow waters and black
fines at depth, significant differences are observed betweenthe shallow waters bacterial communities at or above theUVB attenuation depth (0.5 m) and deep waters (4 m). Bothpopulations are dominated by members of just two phyla, theBacteroidetes and theProteobacteria (61% at 50 cm and 77%at 4 m). The Licancabur samples contains a small contribu-tion the Firmicutes (2.8% and 4.3%, respectively). Membersof 8 additional phyla were identified in the 0.5 m sample andonly one more in the 4-m sediment.[45] The samples present a substantial fraction of cloned
sequences currently unclassified at the phylum level (15%average) and most are also unclassified at the genus level(71% average). The narrow range of diversity at 4 m is evenmore apparent at the genus-species level. Thirteen of the69 clones (19%) are in the genus Thiobacillus (class Beta-proteobacteria) and, using a criterion of 95% sequencesimilarity, are the same species. Another 5 clones (7% ofthe total) are the same species in the genus Thiomicrospira(class Gammaproteobacteria), and 7 of the 29 clones in thephylum Bacteriodetes (10% of the clone library), thoughunclassified, are predicted to be the same species. Thus, 36%of the bacteria in the 4 m sediment sample are potentially
Figure 8. Evolution of UVB and temperature at the summit of Simba, austral summer 2007. Data fromthe Eldonet dosimeter is averaged every minute. Temperature and UV vary along the same gradients butchanges are not parallel, and significant fluctuations lead to inverse relationships.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
12 of 20
G00D06
represented by just 3 species of bacteria. In this sample, 26%of the clones are Thiobacillus or Thiomicrospira species, thusmembers of genera of sulfur-oxidizing chemoautotrophicbacteria.[46] Four of the lakes support zooplankton, the densest
populations being at Licancabur, Laguna Blanca, and Salarde Aguas Calientes. While the density has not been quanti-fied, swarms of pigmented copepods were detected by simplevisual inspection in the waters of these three lakes. The pop-ulation appears to be less abundant in Laguna Lejıa but thisresult should be considered preliminary as only one series
of tows was performed during the 2007 field campaign. A20 mm plankton net was towed in �0.5 m deep water inflamingo grazing areas and collection retrieved only a fewspecimens. Visual inspection supports the hypothesis of amore limited colony.[47] At Licancabur, the population includes at least two
calanoid copepod species, two species of ostracod, threespecies of cladoceran, and a rotifer (Table 2). The samplescontain a significant proportion of previously undescribedspecies, most of them pigmented, indicating adaptation tohigh-UV. Licancabur is now the highest known habitat forany species of chironomid (Cricotopus sp.); the previousrecord was 5600 m in the Himalayas [Saether and Willassen,1987]. Not all organisms have yet been identified to specieslevel (Figure 14). Detailed taxonomic description for Lican-cabur samples are presented by J. Gibson et al. (manuscript inpreparation, 2009).
6. Discussion
[48] The geophysical environment make the high lakesunique habitats for life compared to other lakes. Theirextreme nature does not lie in their pH, composition, or tem-perature, but rather in the year-round combination of highsolar irradiance, permanent ozone depletion, highUV/T ratio,variable precipitation input, and a highly dynamical chemis-try which define them as environmental end-members. Thethreat to ecosystems from the combination of low ozone andincreased UVB is well monitored in the polar regions [e.g.,Battaglia et al., 1997;Cockell and Blaustein, 2001;Williamsonet al., 2001; de Mora et al., 2000;Massom et al., 2006]. Basedon mean daily values, ozone concentration can drop below100 DU during ozone hole events in Antarctica and fluctuateover 230% of its value in 24 h while UVB increases 3.75folds (BAS data, Rother Research Station). Ozone concen-tration can return to normal values (�380 DU) within a fewdays (Figure 15). Because of low solar angle, the maximumUVB detected at the surface is�1.5 W/m2, equivalent to thelowest UVB values reached at the high lakes during wintersolstice (Figures 15a and 15b). What is extreme for life inAntarctica is not, thus, the total amount of UVB received atthe surface but rather the large fluctuations in daily ozoneconcentration and UVB (Figures 15c–15e). The Andeanhigh lakes are extreme to life for the opposite reason, whichis the stability of a permanently depleted (25–40%) ozoneconcentration (Figure 15d), mean UVB at least three timeshigher than in Antarctica year-round, and erythemallyweighted daily doses ranging from 17.1 to 6.5 KJ/m2
(Laguna Blanca station, 2003 [Hock, 2008]) compared to3–0.01 KJ/m2 at the pole (Figure 15f). Licancabur extremevalues between 2003 and 2005 are 23–6.4 KJ/m2.[49] In this environment, some lakes have relatively abun-
dant and diverse ecosystems including macroscopic andmicroscopic organisms while others support only bacterialpopulations. Conditions for habitability and survivability aredifferent for each lake and depend both on regional factorssuch as environment and climate, and local factors, i.e.,geology, chemistry, altitude, and stochastic events includ-ing volcanic eruptions, exceptional yearly precipitation ordroughts. The proximity of the active volcano Lascar is dis-ruptive to Lejıa and Simba’s ecosystems and generates crisesduring eruptions and periods of recovery between volcanic
Figure 9. (a) Average total ozone concentration cycles inthe Licancabur and Simba regions with lowest values reachedbetween fall and summer solstice. (b) Data suggest that theexceptional development of the Antarctic ozone hole betweenSeptember and October could have impacted the survey areasin 2005. The days highlighted in Figure 9b correspond todays when the margin of the ozone hole reached the tip ofSouth America (BAS, October 2005 bulletin). Those days areassociated with dips in the ozone concentration and spikes inthe erythemal UV. Ozone and Erythemal UV values are fromOMI and averaged over a 0.25� � 0.25� cell. There is noEldonet UV data for the summits of Licancabur and Simba in2005. Credit data: OMI, NASA GSFC.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
13 of 20
G00D06
activity. Because of wind direction and close range (5 km),Simba’s water and sediment chemistry might be more sus-ceptible to interaction with Lascar clouds than the other lakes,with potentially limiting effect on the development of theecosystem that will be investigated in future campaigns.[50] While limited, the human factor is not nil. Volcanoes
are, or were recently, being mined for sulfur in Bolivia up toover 5500 m altitude. The presence of tailings on lakeshoressuch as in Laguna Verde impacts lake chemistry (e.g., highlevels of bromine, lead, arsenic) and could explain why thezooplankton migrating from Laguna Blanca through theconnecting channel cannot survive. This hypothesis has notbeen quantified yet. Finally, human presence and wildlifealso generate biological contamination. As a result, althoughisolated, the lakes are not pristine, but the question is lessabout how life is seeded than how it survives once implanted.The timing of environmental cycles offers possible clues.[51] Figure 16 shows the yearly interplay of some of the
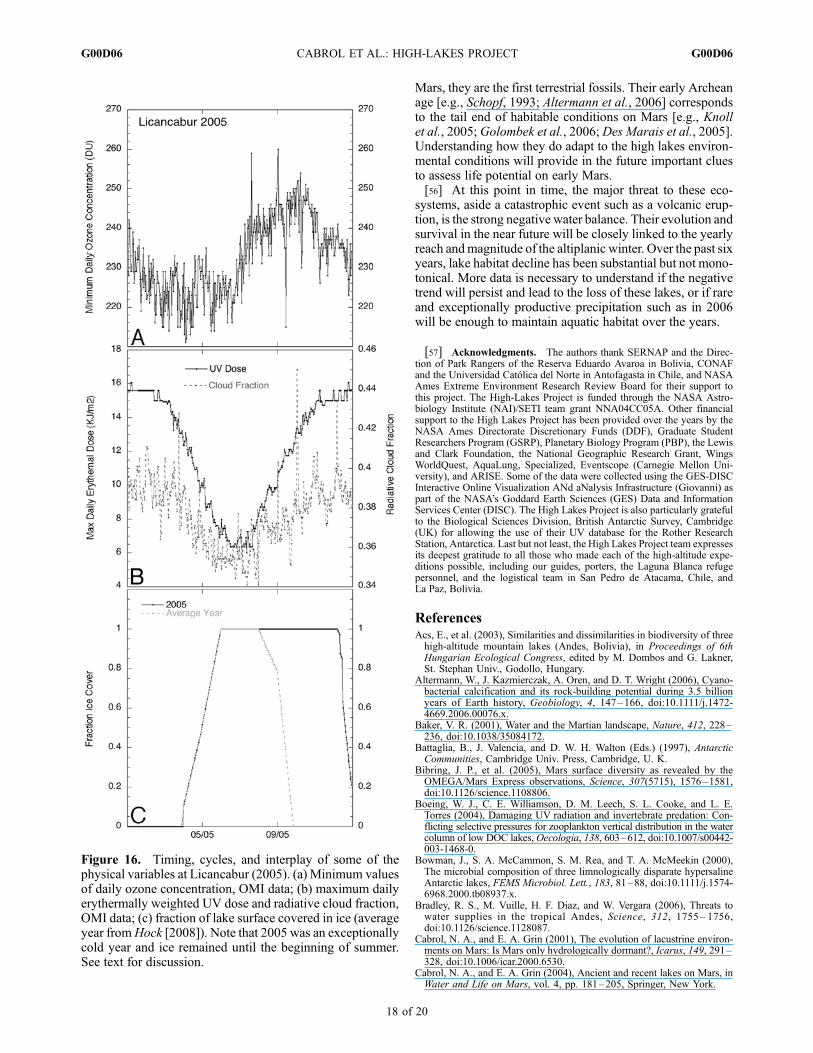
environmental factors, i.e., ozone concentration, maximumerythemally weighted UV daily dose, radiative cloud frac-tion, and yearly fraction of ice cover at the surface of the
Licancabur lake in 2005. In Antarctica, ozone hole eventsoccur in spring during increasing solar irradiance. At the highlakes, minimum ozone occurs during decreasing solar irra-diance between April and June at a time when UV decreases.During the same period, the ice cover builds up on the lakereaching up to 15% of the water column. It thaws on averageat the beginning of October. While limited light penetratesthe water for 5–6 months, life is sheltered from radiation.From the solar irradiance standpoint, this is the safest periodfor the ecosystem.[52] October–February have the highest risk factor for life
with lowering ozone, increased solar irradiance, and exposedaquatic habitat. Additionally, the altiplanic winter betweenJanuary and March brings scattered skies, increasing the fre-quency of sudden temperature and UV fluctuations. Habitatvolatility over time is increased by a highly dynamicalenvironment resulting from the variable reach and magnitudeof the altiplanic winter and irregular precipitation which im-pact water chemistry. Stilll life is relatively abundant anddiverse in most lakes and appears to be adapted to its habitats.To understand adaptability, the next phase of HLP is to estab-
Figure 10. Ionic concentration fluctuations at Licancabur between 2002 and 2006. Wetter years could beassociated with increased water-rock interaction on the shore and runoff of sulfur and salt-rich materials inthe lake as suggested by increased value of SO4, Na, and Cl.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
14 of 20
G00D06
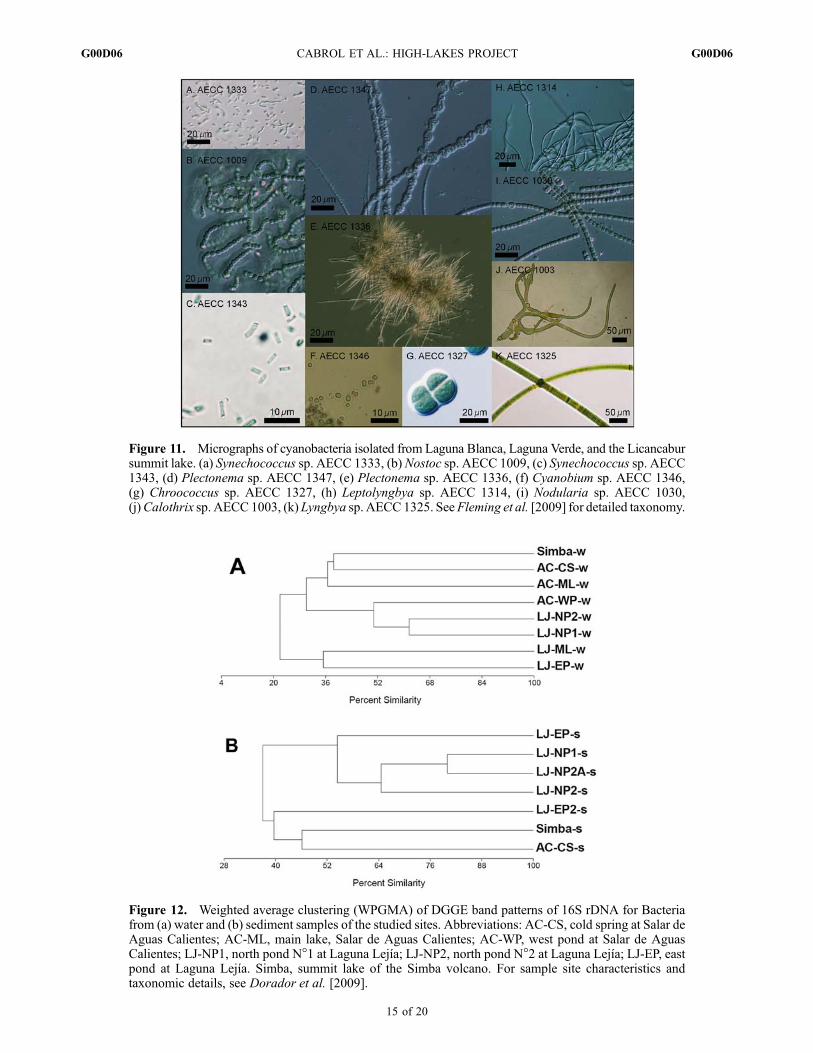
Figure 11. Micrographs of cyanobacteria isolated from Laguna Blanca, Laguna Verde, and the Licancabursummit lake. (a) Synechococcus sp. AECC 1333, (b)Nostoc sp. AECC 1009, (c) Synechococcus sp. AECC1343, (d) Plectonema sp. AECC 1347, (e) Plectonema sp. AECC 1336, (f) Cyanobium sp. AECC 1346,(g) Chroococcus sp. AECC 1327, (h) Leptolyngbya sp. AECC 1314, (i) Nodularia sp. AECC 1030,(j)Calothrix sp. AECC 1003, (k) Lyngbya sp. AECC 1325. SeeFleming et al. [2009] for detailed taxonomy.
Figure 12. Weighted average clustering (WPGMA) of DGGE band patterns of 16S rDNA for Bacteriafrom (a) water and (b) sediment samples of the studied sites. Abbreviations: AC-CS, cold spring at Salar deAguas Calientes; AC-ML, main lake, Salar de Aguas Calientes; AC-WP, west pond at Salar de AguasCalientes; LJ-NP1, north pond N�1 at Laguna Lejıa; LJ-NP2, north pond N�2 at Laguna Lejıa; LJ-EP, eastpond at Laguna Lejıa. Simba, summit lake of the Simba volcano. For sample site characteristics andtaxonomic details, see Dorador et al. [2009].
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
15 of 20
G00D06

lish the connections between the environmental variables andcycles, lake habitat, life’s characteristics, and the biologicalcycles.
7. Conclusion
[53] The primary goal of HLP in investigating the highlakes was to characterize the habitability potential of aquaticenvironments subjected to rapid climate change, and compiledata that could help understand the transformations takingplace inMartian lakes at theNoachian/Hesperian transition inanalogous conditions 3.5 Ga ago. In the process, the projectaccumulated data that show the impact of Earth’s climatechange on lakes in the Central Andes and their ecosystemsin regions where data is scarce due to isolation and difficultaccess.[54] HLP results suggest that in one of the best environ-
mental analogs to earlyMars, including aUV flux double thatof present-day Mars at the equator and UVB only half thatof the red planet [Hock, 2008], solar irradiance, albeit critical,
is only one among many variables (e.g., chemistry, salt con-tents, DOC, water column depth, water supply, temperature)that define habitability potential. The interplay of thesevariables is unique for each body of water. As a result,habitability potential and survivability depend equally onthe lake’s own characteristics and on external factors, such asenvironment and climate, as it must have been the case onearly Mars.[55] Some of the lakes host complex ecosystems, including
zooplankton irrelevant to Mars. Cyanobacteria and bacterialcommunities are, on the other hand, of potential relevance.Although it can be argued that cyanobacteria never existed on
Figure 13. Bacterial composition of Licancabur sediment from (a) a depth of 0.5m; percentages are basedon a total of 72 clones out of 96 sequenced after eliminating chimeras; (b) a depth of 4.0 m based on69 clones out of 96 sequenced. See E. Minkley et al. (manuscript in preparation, 2009) for completedescription, taxonomy, and cladograms.
Table 2. Zooplankton Diversity and Abundancea
Salar AguasCalientes Licancabur
LagunaLejıa
LagunaBlanca
Amphipod 1Chironomid 1 1 1Cladoceran 3 1Copepod 2 2 1Leech 1Nematode 1Ostracod 1+ 2 1Rotifer 2 1Testate amoeba 1Trichopteran 1Preliminary Total 10+ 10 3 1
aPreliminary results. The taxonomic details are provided by J. Gibson et al.(manuscript in preparation, 2009).
Figure 14. Ostracod from the Licancabur lake. Previouslyundescribed species of the genus Amphycitris. Identificationand taxonomy performed by Ricardo Lourenco Pinto, Uni-versidade de Sao Paulo, Brazil. See J. Gibson et al. (manu-script in preparation, 2009) for detailed taxonomy.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
16 of 20
G00D06
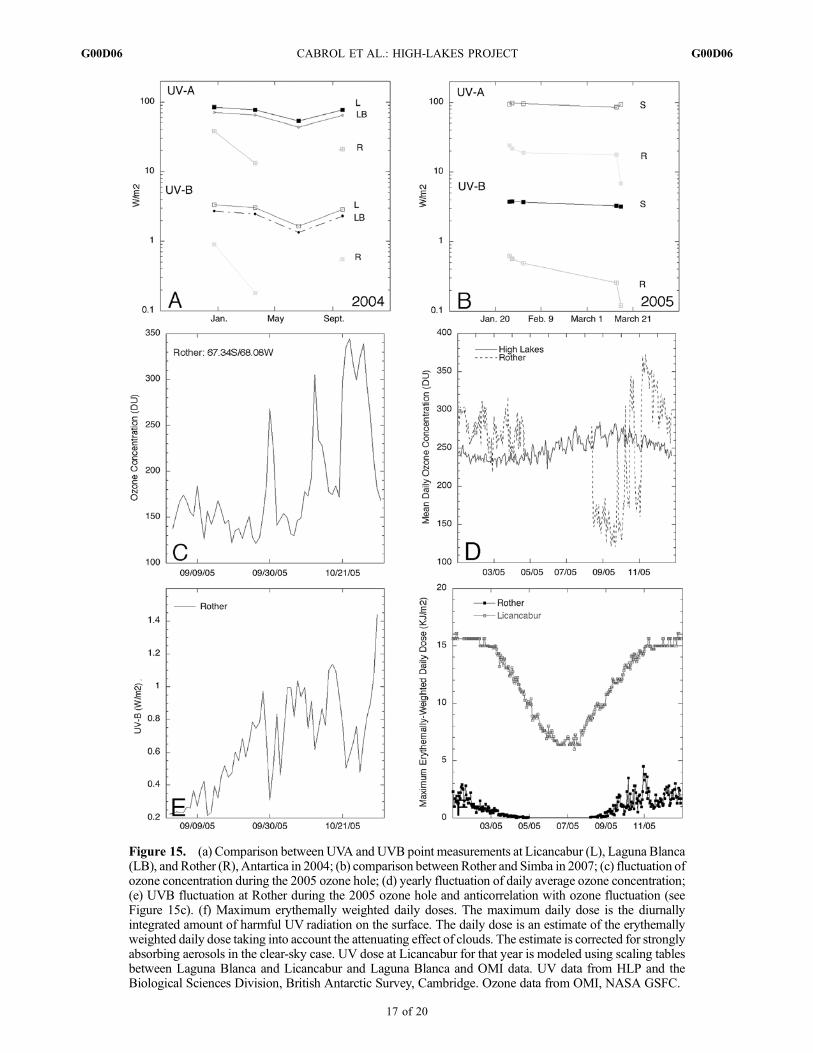
Figure 15. (a) Comparison between UVA and UVB point measurements at Licancabur (L), Laguna Blanca(LB), and Rother (R), Antartica in 2004; (b) comparison between Rother and Simba in 2007; (c) fluctuation ofozone concentration during the 2005 ozone hole; (d) yearly fluctuation of daily average ozone concentration;(e) UVB fluctuation at Rother during the 2005 ozone hole and anticorrelation with ozone fluctuation (seeFigure 15c). (f) Maximum erythemally weighted daily doses. The maximum daily dose is the diurnallyintegrated amount of harmful UV radiation on the surface. The daily dose is an estimate of the erythemallyweighted daily dose taking into account the attenuating effect of clouds. The estimate is corrected for stronglyabsorbing aerosols in the clear-sky case. UV dose at Licancabur for that year is modeled using scaling tablesbetween Laguna Blanca and Licancabur and Laguna Blanca and OMI data. UV data from HLP and theBiological Sciences Division, British Antarctic Survey, Cambridge. Ozone data from OMI, NASA GSFC.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
17 of 20
G00D06
Mars, they are the first terrestrial fossils. Their early Archeanage [e.g., Schopf, 1993; Altermann et al., 2006] correspondsto the tail end of habitable conditions on Mars [e.g., Knollet al., 2005; Golombek et al., 2006; Des Marais et al., 2005].Understanding how they do adapt to the high lakes environ-mental conditions will provide in the future important cluesto assess life potential on early Mars.[56] At this point in time, the major threat to these eco-
systems, aside a catastrophic event such as a volcanic erup-tion, is the strong negative water balance. Their evolution andsurvival in the near future will be closely linked to the yearlyreach andmagnitude of the altiplanic winter. Over the past sixyears, lake habitat decline has been substantial but not mono-tonical. More data is necessary to understand if the negativetrend will persist and lead to the loss of these lakes, or if rareand exceptionally productive precipitation such as in 2006will be enough to maintain aquatic habitat over the years.
[57] Acknowledgments. The authors thank SERNAP and the Direc-tion of Park Rangers of the Reserva Eduardo Avaroa in Bolivia, CONAFand the Universidad Catolica del Norte in Antofagasta in Chile, and NASAAmes Extreme Environment Research Review Board for their support tothis project. The High-Lakes Project is funded through the NASA Astro-biology Institute (NAI)/SETI team grant NNA04CC05A. Other financialsupport to the High Lakes Project has been provided over the years by theNASA Ames Directorate Discretionary Funds (DDF), Graduate StudentResearchers Program (GSRP), Planetary Biology Program (PBP), the Lewisand Clark Foundation, the National Geographic Research Grant, WingsWorldQuest, AquaLung, Specialized, Eventscope (Carnegie Mellon Uni-versity), and ARISE. Some of the data were collected using the GES-DISCInteractive Online Visualization ANd aNalysis Infrastructure (Giovanni) aspart of the NASA’s Goddard Earth Sciences (GES) Data and InformationServices Center (DISC). The High Lakes Project is also particularly gratefulto the Biological Sciences Division, British Antarctic Survey, Cambridge(UK) for allowing the use of their UV database for the Rother ResearchStation, Antarctica. Last but not least, the High Lakes Project team expressesits deepest gratitude to all those who made each of the high-altitude expe-ditions possible, including our guides, porters, the Laguna Blanca refugepersonnel, and the logistical team in San Pedro de Atacama, Chile, andLa Paz, Bolivia.
ReferencesAcs, E., et al. (2003), Similarities and dissimilarities in biodiversity of threehigh-altitude mountain lakes (Andes, Bolivia), in Proceedings of 6thHungarian Ecological Congress, edited by M. Dombos and G. Lakner,St. Stephan Univ., Godollo, Hungary.
Altermann, W., J. Kazmierczak, A. Oren, and D. T. Wright (2006), Cyano-bacterial calcification and its rock-building potential during 3.5 billionyears of Earth history, Geobiology, 4, 147–166, doi:10.1111/j.1472-4669.2006.00076.x.
Baker, V. R. (2001), Water and the Martian landscape, Nature, 412, 228–236, doi:10.1038/35084172.
Battaglia, B., J. Valencia, and D. W. H. Walton (Eds.) (1997), AntarcticCommunities, Cambridge Univ. Press, Cambridge, U. K.
Bibring, J. P., et al. (2005), Mars surface diversity as revealed by theOMEGA/Mars Express observations, Science, 307(5715), 1576–1581,doi:10.1126/science.1108806.
Boeing, W. J., C. E. Williamson, D. M. Leech, S. L. Cooke, and L. E.Torres (2004), Damaging UV radiation and invertebrate predation: Con-flicting selective pressures for zooplankton vertical distribution in the watercolumn of low DOC lakes, Oecologia, 138, 603–612, doi:10.1007/s00442-003-1468-0.
Bowman, J., S. A. McCammon, S. M. Rea, and T. A. McMeekin (2000),The microbial composition of three limnologically disparate hypersalineAntarctic lakes, FEMS Microbiol. Lett., 183, 81–88, doi:10.1111/j.1574-6968.2000.tb08937.x.
Bradley, R. S., M. Vuille, H. F. Diaz, and W. Vergara (2006), Threats towater supplies in the tropical Andes, Science, 312, 1755 – 1756,doi:10.1126/science.1128087.
Cabrol, N. A., and E. A. Grin (2001), The evolution of lacustrine environ-ments on Mars: Is Mars only hydrologically dormant?, Icarus, 149, 291–328, doi:10.1006/icar.2000.6530.
Cabrol, N. A., and E. A. Grin (2004), Ancient and recent lakes on Mars, inWater and Life on Mars, vol. 4, pp. 181–205, Springer, New York.
Figure 16. Timing, cycles, and interplay of some of thephysical variables at Licancabur (2005). (a) Minimum valuesof daily ozone concentration, OMI data; (b) maximum dailyerythermally weighted UV dose and radiative cloud fraction,OMI data; (c) fraction of lake surface covered in ice (averageyear fromHock [2008]). Note that 2005 was an exceptionallycold year and ice remained until the beginning of summer.See text for discussion.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
18 of 20
G00D06

Cabrol, N. A., et al. (2003), First results of the expedition to the highest lakeon earth: Studying a Martian paleolake in Bolivia and the survivalstrategies developed by living organisms, Lunar Planet. Sci. Conf.34th, 1140.
Cabrol, N. A., A. N. Hock, M. Sunagua, and E. A. Grin (2006), Evolutionof aqueous habitat and life in high-altitude lakes during rapid climatechange: Astrobiological methods and geo and biosignatures, Lunar Planet.Sci. Conf. 37th, 1016.
Cabrol, N. A., et al. (2007a), Signatures of habitats and life in Earth’s high-altitude lakes: Clues to Noachian aqueous environments on Mars, in TheGeology of Mars, Evidence From Earth-Based Analogs, edited byM. Chapman, chap. 14, pp. 349–370, Cambridge Univ. Press, Cambridge,U. K.
Cabrol, N. A., E. A. Grin, and A. N. Hock (2007b), Mitigation of environ-mental extremes as a possible indicator of extended habitat sustainabilityfor lakes on early Mars, Proc. SPIE Conf., 1, 15 pp.
Cabrol, N. A., et al. (2008), The High-Lakes Project (HLP): Dynamics ofdeclining lakes, habitat sustainability, and life in early Mars analog envi-ronment, AbSciCon 2008, 3894258.
Cabrol, N. A., D. T. Andersen, C. R. Stoker, P. Lee, C. P. McKay, and D. S.Wettergreen (2009a), Other analogs to Mars: High altitude, subsurface,desert, and polar environments, in Life in Antarctic Deserts and OtherCold Dry Environments: Astrobiological Analogues, edited by P. Doran,Cambridge Univ. Press, Cambridge, U. K., in press.
Cabrol, N. A., et al. (2009b), Lake environment and life under permanentozone depletion and high UV radiation at Licancabur, Central Andes, inLakes on Mars, edited by N. A. Cabrol and E. A. Grin, Elsevier, NewYork, in press.
Carr, M. H. (1996), Water on Mars, 229 pp., Oxford Univ. Press, NewYork.
Christensen, P. R., et al. (2003), Morphology and composition of the sur-face of Mars: Mars odyssey THEMIS results, Science, 300, 2056–2061,doi:10.1126/science.1080885.
Cockell, C. S., and A. R. Blaustein (Eds.) (2001), Ecosystems, Evolution,and Ultraviolet Radiation, 221 pp., Springer, New York.
Demergasso, C., L. Escudero, E. O. Casamayor, G. Chong, V. Balague, andC. Pedros-Alio (2008), Novelty and spatio– temporal heterogeneity in thebacterial diversity of hypersaline Lake Tebenquiche (Salar de Atacama),Extremophiles, doi:10.1007/s00792-008-0153-y.
de Mora, S., , S. Demers, and M. Vernet (Eds.) (2000), The Effects of UVRadiation in the Marine Environment, vol. 10, 324 pp., Cambridge Univ.Press, Cambridge, U. K.
Deruelle, B. (1978), Calc-alkaline and shoshonitic lavas from five Andeanvolcanoes (between latitudes 21�45( and 24�300S) and the distribution ofthe Plio-Quaternary volcanism of the south-central and southern Andes,J. Volcanol. Geotherm. Res., 3, 281–298, doi:10.1016/0377-0273(78)90039-2.
de Silva, S., and P. Francis (1991), Volcanoes of the Central Andes, Springer,Berlin.
Des Marais, D. J., and J. D. Farmer (1995), The search for extinct life, in AnExobiological Strategy for Mars Exploration, edited by M. Meyer andJ. Kerridge, pp. 21–25, NASA, Washington, D. C.
Des Marais, D. J., et al. (2005), Astrobiology and the basaltic plains inGusev crater, Lunar Planet. Sci. Conf. 36th, 2353.
Dohm, J. M., V. R. Baker, S. Maruyama, and R. C. Anderson (2007), Traitsand evolution of the Tharsis superplume, Mars, in Superplumes: BeyondPlate Tectonics, pp. 523–536, Springer, New York.
Dorador, C. (2007), Microbial communities in high altitude altiplanic wet-lands in northern Chile: Phylogeny, diversity and function, Ph.D. thesis,170 pp., Max Planck Inst. fur Evolutionsbiologie, Christian-Albrechts-Univ., Kiel, Germany.
Dorador, C., D. Meneses, V. Urtuvia, C. Demergasso, I. Vila, K.-P. Witzel,and J. F. Imhoff (2009), Diversity of Bacteroidetes in high-altitude salineevaporitic basins in northern Chile, J. Geophys. Res., 114, G00D05,doi:10.1029/2008JG000837.
Escudero, L., G. Chong, C. Demergasso, M. E. Farias, N. A. Cabrol, E. A.Grin, E. Minkley Jr., and Y. Yu (2007), Investigating microbial diversityand UV radiation impact at the high altitude lake Aguas Calientes, Chile,Proc. SPIE Conf., 1, 12 pp.
Farias, M. E., V. Fernandez-Zenoff, R. Flores, O. Ordonez, and C. Estevez(2009), Impact of solar radiation on bacterioplankton in Laguna Vilama,a hypersaline Andean lake (4650 m), J. Geophys. Res., 114, G00D04,doi:10.1029/2008JG000784.
Farmer, J. D. (1994), Landing site catalog, NASA Rep.,, 1238, pp. 363–366.Farmer, J. D. (1995), Mars exopaleontology, Palaios, 10(3), 197–198,doi:10.2307/3515252.
Farmer, J. D. (2000), Strategies for the astrobiological exploration of Mars,in Concepts and Approaches for Mars Exploration, part 1, Lunar Planet.Inst. Contrib., 1062, p. 110–112.
Fischer, J. M., J. Nicolai, C. E. Williamson, A. D. Persaud, and R. S.Lockwood (2006), Effects of ultraviolet radiation on diel vertical migra-tion of crustacean zooplankton: An in situ mesocosm experiment, Hydro-biologia, 563, 217–224, doi:10.1007/s10750-005-0007-x.
Fleming, E. D., et al. (2009), Characterization of cyanobacteria from anatural high ultraviolet radiation environment in Laguna Blanca, Bolivia,J. Geophys. Res., doi:10.1029/2008JG000817, in press.
Fornari, M., F. Risacher, and G. Feraud (2001), Dating of paleolakes in thecentral Altiplano of Bolivia, Palaeogeogr. Palaeoclimatol. Palaeoecol.,172(3/4), 269–282, doi:10.1016/S0031-0182(01)00301-7.
Garcia-Pichel, F., M. Mechling, R. Castenholz, and W. Diel (1994), Migra-tions of microorganisms within a benthic, hypersaline mat community,Appl. Environ. Microbiol., 60(5), 1500–1511.
Golombek, M., et al. (2006), Geology of the Gusev cratered plains from theSpirit rover traverse, J. Geophys. Res., 111, E02S07, doi:10.1029/2005JE002503.
Gonzalez-Ferran, O. (1995), Volcanes de Chile, 635 pp., Inst. Geogr. Militar,Santiago.
Grosjean, M. (1994), Paleohydrology of the Laguna Leijıa (north Chileanaltiplano) and climatic implications for late glacial times, Palaeogeogr.Palaeoclimatol. Palaeoecol., 109(1), 89–100, doi:10.1016/0031-0182(94)90119-8.
Grosjean, M., et al. (2001), Mid-Holocene (ca. 6000–3500 14C yr B.P.)climates in the Atacama Altiplano of the south Central Andes: Humidor dry?—Limnogeological, geomorphological, hydrological and vegeta-tional evidence for widespread mid-Holocene aridity, paper presented atWorkshop on Central Andes Paleoclimatology, Natl. Sci. Found., Tucson,Ariz., 11–16 Jan.
Haberle, R. M., C. P. McKay, J. Schaeffer, N. A. Cabrol, E. A. Grin, A. P.Zent, and R. Quinn (2001), On the possibility of liquid water on present-day Mars, J. Geophys. Res., 106(E10), 23,317–23,326, doi:10.1029/2000JE001360.
Hader, D.-P., and M. Lebert (2006), Eldonet–European-Light DosimeterNetwork, in Environmental UV Radiation: Impact on Ecosystems andHuman Health and Predictive Models, edited by F. Ghetti, G. Checcucci,and J. F. Bornman, NATO Sci. Ser., 57, 95–108.
Hader, D.-P.,M.Lebert, R.Marangoni, andG.Colombetti (1999), ELDONET-European light dosimeter network hardware and software, J. Photochem.Photobiol. B, 52, 51–58, doi:10.1016/S1011-1344(99)00102-5.
Hamamura, N., S. H. Olson, D. Ward, and W. P. Inskeep (2005), Diversityand functional analysis of bacterial communities associated with naturalhydrocarbon seeps in acidic soils at Rainbow Springs, YellowstoneNational Park, Appl. Environ. Microbiol., 71, 5943–5950, doi:10.1128/AEM.71.10.5943-5950.2005.
Hamon,W. R. (1961), Estimating potential evapotranspiration, J. HydraulicsDiv, Proc. Am. Soc. Civ. Eng., 87, 107–120.
Hargreaves, B. R. (2003), Water column optics and penetration of UVR, inUV Effects in Aquatic Organisms and Ecosystems, edited by E. W. Helblingand H. E. Zagarese, pp. 59–105, R. Soc. of Chem., Cambridge, U. K.
Hock, A. N. (2008), Licancabur Volcano and Life in the Atacama: Envir-onmental physics and analogies to Mars, Ph.D. thesis, Univ. of Calif.,Los Angeles.
Humayoun, S. B., N. Bano, and J. T. Hollibaugh (2003), Depth distributionof microbial diversity in Mono Lake, a meromictic soda lake in Califor-nia, Appl. Environ. Microbiol., 69, 1030–1042, doi:10.1128/AEM.69.2.1030-1042.2003.
Jakosky, B. M., B. M. Haberle, and R. E. Arvidson (2003), The changingpicture of volatiles and climate on Mars, Science, 310(5753), 1439–1440, doi:10.1126/science.1118031.
James, P. M., D. Peters, and K. M. Greisinger (1997), A study of ozonemini-hole formation using a tracer advection model driven by barotropicdynamics,Meteorol. Atmos. Phys., 64, 107–121, doi:10.1007/BF01044132.
Jaque, F., et al. (1994), Ground-based ultraviolet radiation measurementsduring springtime in the Southern Hemisphere, Europhys. Lett., 28(4),289–293, doi:10.1209/0295-5075/28/4/012.
Jiang, H., H. Dong, G. Zhang, B. Yu, L. Chapman, and M. Fields (2006),Microbial diversity in water and sediment of Lake Chaka, an athalassoha-line lake in northwestern China, Appl. Environ. Microbiol., 72, 3832–3845, doi:10.1128/AEM.02869-05.
Karentz, D. (1994), Ultraviolet tolerance mechanisms in Antarctic marineorganisms, in Ultraviolet Radiation in Antarctica: Measurements andBiological Effects, Antarctic Res. Ser., vol. 62, edited by C. S. Weilerand P. A. Penhale, pp. 93–100, AGU, Washington, D. C.
Kiss, K. T., et al. (2004), Habitats extremes pour les communautes dediatomees dans les lacs de haute altitude (Laguna Blanca et lac de crateredu volcan Licancabur, Bolivie), paper presented at 23eme Colloque del’ADLaF, Orleans, France.
Knoll, A. H., M. Carr, B. Clark, and D. J. Des Marais (2005), J. D. Farmerand 8 authors. An astrobiological perspective on Meridiani planum, EarthPlanet. Sci. Lett., 240, 179–189, doi:10.1016/j.epsl.2005.09.045.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
19 of 20
G00D06

Leech, D. M., A. T. Padeletti, and C. E. Williamson (2005a), Zooplanktonbehavioral responses to solar UV radiation vary within and among lakes,J. Plankton Res., 27, 461–471, doi:10.1093/plankt/fbi020.
Leech, D. M., C. E. Williamson, R. E. Moeller, and B. R. Hargreaves(2005b), Effects of ultraviolet radiation on the seasonal vertical distributionof zooplankton: A database analysis, Arch. Hydrobiol., 162, 445–464,doi:10.1127/0003-9136/2005/0162-0445.
Malin, M. C., and K. S. Edgett (2000), Sedimentary rocks of early Mars,Science, 290, 1927–1937, doi:10.1126/science.290.5498.1927.
Malin, M. C., K. S. Edgett, L. V. Posiolova, S. M. McColley, and E. Z. NoeDobrea (2006), Present-day impact cratering impact and contemporarygully activity on Mars, Science, 314(5805), 1573–1577, doi:10.1126/science.1135156.
Marinovic, N., and A. Lahsen (1984), Hoja Calama, Region de Antofagasta,Carta Geologica de Chile, 58, 1:250000, Serv. Nac. Geol. Min.
Massom, R., et al. (2006), Extreme anomalous atmospheric circulation inthe West Antarctic Peninsula region in austral spring and summer 2001/2,and its profound impact on sea ice and biota, J. Clim. Res., 19, 3544–3571, doi:10.1175/JCLI3805.1.
McLennan, S. M., et al. (2005), Provenance and diagenesis of the evaporite-bearing Burns formation,Meridiani Planum,Mars, Earth Planet. Sci. Lett.,240, 95–121, doi:10.1016/j.epsl.2005.09.041.
Morris, R. L., R. Berthold, and N. Cabrol (2007), Diving at extreme alti-tude: Dive planning and execution during the 2006 High-Lakes scienceexpedition, paper presented at Diving for Science Symposium, Am.Acad. of Underwater Sci., Miami, Fla.
Murray, J. B., et al. (2005), Evidence from the Mars Express High Resolu-tion Stereo Camera for a frozen sea close to Mars’ equator, Nature, 434,352–355, doi:10.1038/nature03379.
Mustard, J. F., et al. (2005), Olivine and pyroxene diversity in the crust ofMars, Science, 307(5715), 1594–1597, doi:10.1126/science.1109098.
Newman, P. A., L. R. Lait, and M. R. Schoeberl (1988), The morphologyand meteorology of southern hemisphere spring total ozone mini-holes,Geophys. Res. Lett., 15, 923–926, doi:10.1029/GL015i008p00923.
Ramırez, C. F., and M. Gardeweg (1982), Hoja Toconao, Carta Geologicade Chile, SNGM, Santiago.
Risacher, F., and B. Fritz (1991), Geochemistry of Bolivian salars, Lipez,southern Altiplano: Origin of solutes and brine evolution, Geochim.Cosmochim. Acta, 55, 687–705, doi:10.1016/0016-7037(91)90334-2.
Risacher, F., H. Alonso, and C. Salazar (1999), Geoquımica de aguas encuencas cerradas, I, II, III Regiones, Chile, Tech. Open File Rep. S.I.T. 51,Ministerio de Obras Publicas, Direccion General de Aguas, Santiago,Chile.
Risacher, F., H. Alonso, and C. Salazar (2003), The origin of brines andsalts in Chilean salars: A hydrochemical review, Earth Sci. Rev., 63,249–293, doi:10.1016/S0012-8252(03)00037-0.
Rose, K. C., C. E. Williamson, S. G. Schladow, M. Winder, and J. T. Oris(2009), Patterns of spatial and temporal variability of UV transparencyin Lake Tahoe, California-Nevada, J. Geophys. Res., 114, G00D03,doi:10.1029/2008JG000816.
Saether, O. A., and E. Willassen (1987), Four new species of DiamesaMeige, 1835 (Diptera, Chironimidae) from the glaciers of Nepal, Entomol.Scand., 29, 189–203.
Santas, R., P. Santas, C. Lianou, and A. Korda (1998), Community re-sponses to UV radiation. II. Effects of solar UVB on field-grown diatomassemblages of the Carribean, Mar. Biol. Berlin, 131(1), 163 –171,doi:10.1007/s002270050307.
Schopf, J. W. (1993), Microfossils of the Early Archean Apex chert: Newevidence for the antiquity of life, Science, 260, 640–646, doi:10.1126/science.260.5108.640.
Servant, M., and J. C. Fontes (1978), Les lacs quaternaires des hauts plateauxdes Andes boliviennes, Premieres interpretations paleoclimatiques, Cah.O. R. S. T. O. M., Ser. Geol., 10(1), 9–23.
Squyres, S. W., R. E. Arvidson, J. F. Bell III, J. Bruckner, and N. A. Cabrol(2004), The Opportunity rover’s Athena science investigation at MeridianiPlanum, Mars, Science, 306(3), 1698 – 1703, doi:10.1126/science.1106171.
Stenke, A., and V. Grewe (2004), Impact of dynamically induced ozonemini-hole events on PSC formation and chemical ozone destruction, Adv.Space Res., 33(7), 1062–1067, doi:10.1016/j.asr.2003.08.001.
Sylvestre, F., M. Servant, S. Servant-Vildary, C. Causse, and C. Fournier(1999), Lake-level chronology on the southern Bolivian Altiplano(18 degrees-23 degrees S) during late-glacial time and the early Holocene,Quat. Res., 51, 54–66, doi:10.1006/qres.1998.2017.
Williamson, C. E., P. J. Neale, G. Grad, H. J. De Lange, and B. R.Hargreaves (2001), Beneficial and detrimental effects of UV on aquaticorganisms: Implications of spectral variation, Ecol. Appl., 11, 1843–1857,doi:10.1890/1051-0761(2001)011[1843:BADEOU]2.0.CO;2.
Williamson, G., H. De Lange, and S. Gilroy (2002), Temperature-dependentultraviolet responses in zooplankton: Implications of climate change,Limnol. Oceanogr., 47(6), 1844–1848.
�����������������������L. Bebout, N. A. Cabrol, E. Fleming, E. A. Grin, D. Lim, and C. Woosley,
Space Science and Astrobiology Division, NASA Ames Research Center,MS 245-3, Moffett Field, CA 94035-1000, USA. ([email protected])G. Chong and L. Escudero, Centro de Investigacion Cientıfica y
Tecnologica para Minerıa, Santiago, Chile.G. Dawidowicz and J. Majerowicz, ESRI, 21 rue des Capucins, F-92190
Meudon, France.C. Demergasso and C. Dorador, Centro de Biotecnologıa, Universidad
Catolica del Norte, Avenida Angamos 0610, Antofagasta, Chile.V. Gaete, Departamento de Quımica, Universidad Catolica del Norte,
Avenida Angamos 0610, Antofagasta, Chile.M. E. Galvez, Ecole Normale Superieure, 24 rue Lhomond, F-75005,
Paris, France.J. Gibson, Marine Research Laboratories, Tasmanian Aquaculture and
Fisheries Institute, University of Tasmania, Private Bag 49, Hobart, Tas7001, Australia.D. P. Hader, Department Botanik, University of Erlangen, Staudtstrasse 5,
D-91058 Erlangen, Germany.A. N. Hock, Department of Earth and Space Sciences, University of
California, 595 Charles Young Drive East, Los Angeles, CA 90095, USA.E. Minkley, Department of Biological Sciences, Carnegie Mellon
University, Pittsburgh, PA 15213-3890, USA.R. L. Morris, SETI Carl Sagan Center, 515 North Whisman Road,
Mountain View, CA 94043, USA.C. Salazar, Clınica Mutual de Seguridad, Avenida Espana 01890, Punta
Arenas, Chile.E. Smith, Discoverer Ketty Lund Exploration Vessel, 330 Julia Street,
Key West, FL 33040, USA.C. Tambley, CHEP, Avenida Del Valle 961, Office 2707, Huechuraba,
Santiago, Chile.I. Uskin-Peate, Geoscience Department, University of Iowa, 121
Trowbridge Hall, Iowa City, IA 52242, USA.Y. Yu, Department of Civil and Environmental Engineering, Carnegie
Mellon University, Pittsburgh, PA 15213-3890, USA.
G00D06 CABROL ET AL.: HIGH-LAKES PROJECT
20 of 20
G00D06
Related Documents











