JMB—MS 660 Cust. Ref. No. CAM 126/95 [SGML] J. Mol. Biol. (1995) 251, 41–49 The HEM Proteins: A Novel Family of Tissue-specific Transmembrane Proteins Expressed from Invertebrates Through Mammals with an Essential Function in Oogenesis Stefan Baumgartner 1 , Doris Martin 1 , Ruth Chiquet-Ehrismann 1 Jill Sutton 2 , Alpana Desai 2 , Irene Huang 2 , Kikuya Kato 3 and Robert Hromas 2 * We report the identification of a new family of proteins, termed the HEM 1 Friedrich Miescher Institute P.O. Box 2543, CH-4002 family, which show distinct expression patterns in blood cells and the central nervous system. Through the isolation and characterization of the Basel, Switzerland corresponding brain-specific Drosophila (dhem-2 ) and rat orthologues 2 Department of Medicine (Hem-2), and through the detection of the Caenorhabditis elegans Hem-2 IB-442, Indiana University orthologue in the database, we show that this family is conserved throughout Medical Center, Indianapolis evolution. HEM proteins show a conserved length ranging from 1118 to 1126 IN 46202, USA amino acid residues. Moreover, they are at least 35% identical with each other and harbour several conserved membrane-spanning domains, indicative for 3 Okayama Cell Switching their location on the cell surface. One of the members, the Drosophila Project, Research Development orthologue dhem-2 , was analysed in detail for its spatial expression pattern Corporation of Japan, Pasteur during development and for its mutant phenotype. dhem-2 is expressed Building 4F, 103-5 Tanaka maternally in the oocyte and shows uniform expression during the first half Monzen-cho, Sakyo-ku, Kyoto of embryogenesis, but becomes restricted to the brain and the nervous 606, Japan system during late embryogenesis, consistent with the expression of its vertebrate orthologue in the brain. One P-element insertion, located 39 base-pairs downstream from the dhem-2 transcription start site, causes female sterility, due to the fact that developmental processes in the oocyte are disturbed. Of the vertebrate HEM family members, the mammalian Hem-1 gene is expressed only in cells of hematopoietic origin, while Hem-2 is preferentially expressed in brain, heart, liver and testis. 7 1995 Academic Press Limited Keywords: HEM family; transmembrane protein; evolution; Drosophila; *Corresponding author maternal lethal Introduction Many genes appear to have been assembled from a pool of modules that are widely shared (Doolittle, 1992). It appears that there are convenient structural units that are used over and over again, the same modules sometimes being used to perform different tasks. Genes can be grouped into families based on these shared structural modules. Recently, we cloned a human cDNA, termed Hem-1, which showed an expression pattern restricted to blood cells (Hromas et al ., 1991). Kato (1990) reported a murine partial cDNA clone, termed mh19, that showed a specific expression pattern in mouse brain. When comparing the predicted sequences of these two proteins, we noticed a high degree of similarity, suggesting that they belong to a family of proteins with distinct expression patterns. Extending the search for further members, we have been able to identify six members from Caenorhabdi- tis elegans to humans. We present evidence that this small family, termed the HEM family, is conserved to a high degree from invertebrates through mammals. Moreover, members appear to be specifically expressed in distinct tissues, such as in the hematopoietic or central nervous system. In addition, the patterns of expression are developmen- tally regulated. Our analysis suggests that the primary sequence has been retained, and that the Correspondence concerning Drosophila HEM genes should be addressed to S.B. Correspondence concerning the vertebrate HEM genes should be addressed to R.H. 0022–2836/95/310041–09 $08.00/0 7 1995 Academic Press Limited

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JMB—MS 660 Cust. Ref. No. CAM 126/95 [SGML]
J. Mol. Biol. (1995) 251, 41–49
The HEM Proteins: A Novel Family of Tissue-specificTransmembrane Proteins Expressed fromInvertebrates Through Mammals with an EssentialFunction in Oogenesis
Stefan Baumgartner 1, Doris Martin 1, Ruth Chiquet-Ehrismann 1
Jill Sutton 2, Alpana Desai 2, Irene Huang 2, Kikuya Kato 3 andRobert Hromas 2*
We report the identification of a new family of proteins, termed the HEM1Friedrich Miescher InstituteP.O. Box 2543, CH-4002 family, which show distinct expression patterns in blood cells and the
central nervous system. Through the isolation and characterization of theBasel, Switzerlandcorresponding brain-specific Drosophila (dhem-2) and rat orthologues2Department of Medicine (Hem-2), and through the detection of the Caenorhabditis elegans Hem-2
IB-442, Indiana University orthologue in the database, we show that this family is conserved throughoutMedical Center, Indianapolis evolution. HEM proteins show a conserved length ranging from 1118 to 1126IN 46202, USA amino acid residues. Moreover, they are at least 35% identical with each other
and harbour several conserved membrane-spanning domains, indicative for3Okayama Cell Switchingtheir location on the cell surface. One of the members, the DrosophilaProject, Research Developmentorthologue dhem-2, was analysed in detail for its spatial expression patternCorporation of Japan, Pasteurduring development and for its mutant phenotype. dhem-2 is expressedBuilding 4F, 103-5 Tanakamaternally in the oocyte and shows uniform expression during the first halfMonzen-cho, Sakyo-ku, Kyotoof embryogenesis, but becomes restricted to the brain and the nervous606, Japansystem during late embryogenesis, consistent with the expression of itsvertebrate orthologue in the brain. One P-element insertion, located 39base-pairs downstream from the dhem-2 transcription start site, causesfemale sterility, due to the fact that developmental processes in the oocyteare disturbed. Of the vertebrate HEM family members, the mammalianHem-1 gene is expressed only in cells of hematopoietic origin, while Hem-2is preferentially expressed in brain, heart, liver and testis.
7 1995 Academic Press Limited
Keywords: HEM family; transmembrane protein; evolution; Drosophila;*Corresponding author maternal lethal
Introduction
Many genes appear to have been assembled froma pool of modules that are widely shared (Doolittle,1992). It appears that there are convenient structuralunits that are used over and over again, the samemodules sometimes being used to perform differenttasks. Genes can be grouped into families based onthese shared structural modules.
Recently, we cloned a human cDNA, termedHem-1, which showed an expression patternrestricted to blood cells (Hromas et al., 1991). Kato
(1990) reported a murine partial cDNA clone, termedmh19, that showed a specific expression patternin mouse brain. When comparing the predictedsequences of these two proteins, we noticed a highdegree of similarity, suggesting that they belong to afamily of proteins with distinct expression patterns.Extending the search for further members, we havebeen able to identify six members from Caenorhabdi-tis elegans to humans. We present evidence that thissmall family, termed the HEM family, is conserved toa high degree from invertebrates through mammals.Moreover, members appear to be specificallyexpressed in distinct tissues, such as in thehematopoietic or central nervous system. Inaddition, the patterns of expression are developmen-tally regulated. Our analysis suggests that theprimary sequence has been retained, and that the
Correspondence concerning Drosophila HEM genesshould be addressed to S.B.
Correspondence concerning the vertebrate HEMgenes should be addressed to R.H.
0022–2836/95/310041–09 $08.00/0 7 1995 Academic Press Limited
JMB—MS 660
HEM Family of Evolutionary Conserved Proteins42
function may be conserved. One member of thisfamily, dhem-2, has an essential function in oogenesis.P-element mutants of dhem-2 are maternally lethal.
Results
Cloning and sequence analysis of the dhem -2and the Hem-2 gene
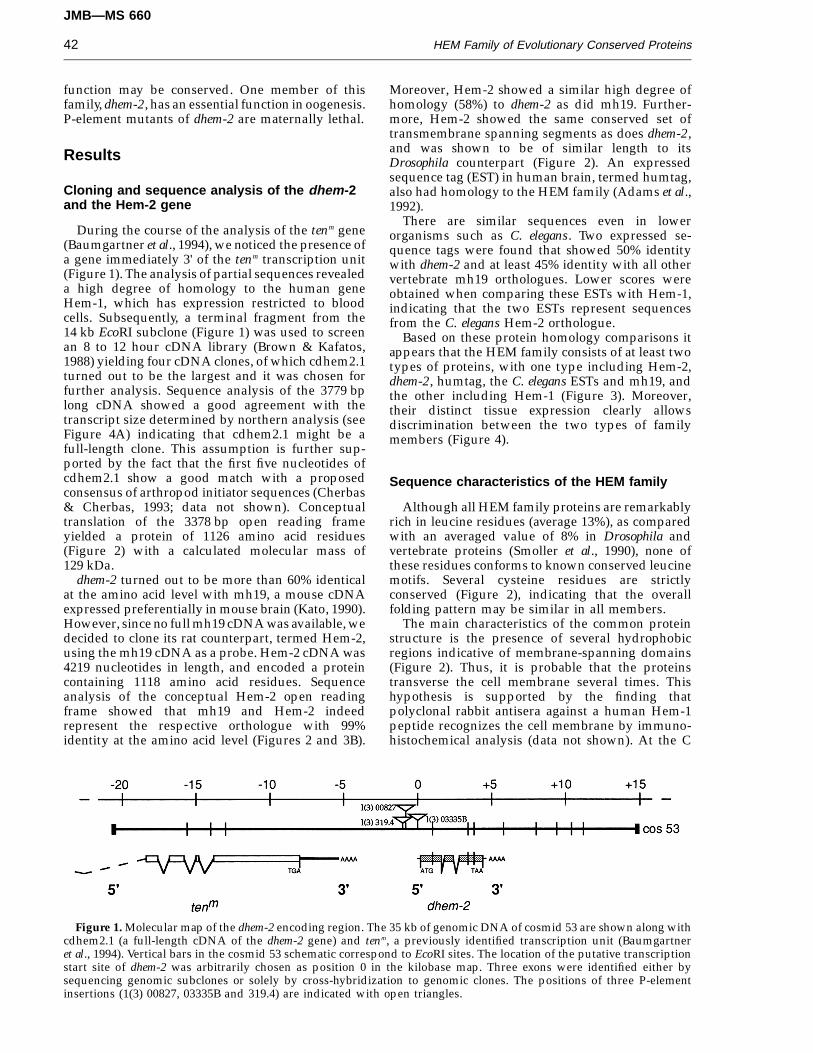
During the course of the analysis of the tenm gene(Baumgartner et al., 1994), we noticed the presence ofa gene immediately 3' of the tenm transcription unit(Figure 1). The analysis of partial sequences revealeda high degree of homology to the human geneHem-1, which has expression restricted to bloodcells. Subsequently, a terminal fragment from the14 kb EcoRI subclone (Figure 1) was used to screenan 8 to 12 hour cDNA library (Brown & Kafatos,1988) yielding four cDNA clones, of which cdhem2.1turned out to be the largest and it was chosen forfurther analysis. Sequence analysis of the 3779 bplong cDNA showed a good agreement with thetranscript size determined by northern analysis (seeFigure 4A) indicating that cdhem2.1 might be afull-length clone. This assumption is further sup-ported by the fact that the first five nucleotides ofcdhem2.1 show a good match with a proposedconsensus of arthropod initiator sequences (Cherbas& Cherbas, 1993; data not shown). Conceptualtranslation of the 3378 bp open reading frameyielded a protein of 1126 amino acid residues(Figure 2) with a calculated molecular mass of129 kDa.
dhem-2 turned out to be more than 60% identicalat the amino acid level with mh19, a mouse cDNAexpressed preferentially in mouse brain (Kato, 1990).However, since no full mh19 cDNA was available, wedecided to clone its rat counterpart, termed Hem-2,using the mh19 cDNA as a probe. Hem-2 cDNA was4219 nucleotides in length, and encoded a proteincontaining 1118 amino acid residues. Sequenceanalysis of the conceptual Hem-2 open readingframe showed that mh19 and Hem-2 indeedrepresent the respective orthologue with 99%identity at the amino acid level (Figures 2 and 3B).
Moreover, Hem-2 showed a similar high degree ofhomology (58%) to dhem-2 as did mh19. Further-more, Hem-2 showed the same conserved set oftransmembrane spanning segments as does dhem-2,and was shown to be of similar length to itsDrosophila counterpart (Figure 2). An expressedsequence tag (EST) in human brain, termed humtag,also had homology to the HEM family (Adams et al.,1992).
There are similar sequences even in lowerorganisms such as C. elegans. Two expressed se-quence tags were found that showed 50% identitywith dhem-2 and at least 45% identity with all othervertebrate mh19 orthologues. Lower scores wereobtained when comparing these ESTs with Hem-1,indicating that the two ESTs represent sequencesfrom the C. elegans Hem-2 orthologue.
Based on these protein homology comparisons itappears that the HEM family consists of at least twotypes of proteins, with one type including Hem-2,dhem-2, humtag, the C. elegans ESTs and mh19, andthe other including Hem-1 (Figure 3). Moreover,their distinct tissue expression clearly allowsdiscrimination between the two types of familymembers (Figure 4).
Sequence characteristics of the HEM family
Although all HEM family proteins are remarkablyrich in leucine residues (average 13%), as comparedwith an averaged value of 8% in Drosophila andvertebrate proteins (Smoller et al., 1990), none ofthese residues conforms to known conserved leucinemotifs. Several cysteine residues are strictlyconserved (Figure 2), indicating that the overallfolding pattern may be similar in all members.
The main characteristics of the common proteinstructure is the presence of several hydrophobicregions indicative of membrane-spanning domains(Figure 2). Thus, it is probable that the proteinstransverse the cell membrane several times. Thishypothesis is supported by the finding thatpolyclonal rabbit antisera against a human Hem-1peptide recognizes the cell membrane by immuno-histochemical analysis (data not shown). At the C
Figure 1. Molecular map of the dhem-2 encoding region. The 35 kb of genomic DNA of cosmid 53 are shown along withcdhem2.1 (a full-length cDNA of the dhem-2 gene) and tenm, a previously identified transcription unit (Baumgartneret al., 1994). Vertical bars in the cosmid 53 schematic correspond to EcoRI sites. The location of the putative transcriptionstart site of dhem-2 was arbitrarily chosen as position 0 in the kilobase map. Three exons were identified either bysequencing genomic subclones or solely by cross-hybridization to genomic clones. The positions of three P-elementinsertions (1(3) 00827, 03335B and 319.4) are indicated with open triangles.

JMB—MS 660
HEM Family of Evolutionary Conserved Proteins 43
Figure 2. Alignment of the amino acid sequences of all known HEM family proteins. Amino acid sequence of dhem-2and Hem-2 and alignment of sequences of all known members of the Hem-family. Each lane contains 100 residues alignedwith each other. Only partial sequences were available from humtag, mh19 and CeEST. Conserved cysteine residues arein bold. One cysteine group, beginning at 675 in Hem-1, has a stringently conserved motif CXHXXXCXHEXC, that isreminescent of zinc-chelating structures. There are runs of basic residues that are also highly conserved, beginning at 340and 634 in Hem-1. Potential membrane-spanning segments are underlined.

JMB—MS 660
HEM Family of Evolutionary Conserved Proteins44
Figure 3. Relationship of the HEM family proteins. A,Schematic alignment of available sequences from allmembers of the HEM family. Note that humtag, CeESTand mh19 do not overlap. B, Comparison of aminoacid sequences of members of the Hem family as a matrixof identical amino acid residues. Due to the lack ofoverlapping sequences of humtag, mh19 and CeEST, novalue is indicated.
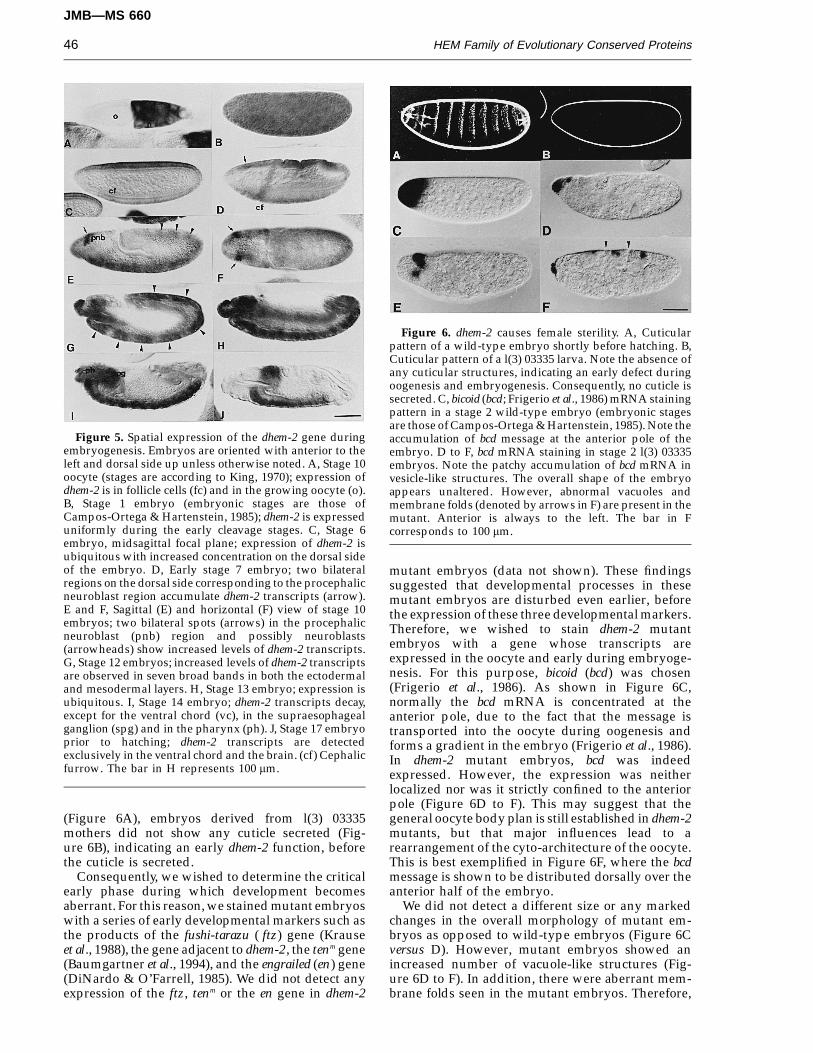
first detected in the ovaries at stage 10 (King, 1970)where the nurse cells are rapidly synthesizingmRNA (Figure 5A). After fertilization and eggdeposition, transcripts are present uniformly at highlevels throughout the embryo (Figure 5B). Maternaldhem-2 transcript levels decrease rapidly during thefirst two hours of embryogenesis, and at stage 6, thezygotic expression shows a uniform pattern withincreased levels on the dorsal side of the embryo(Figure 5C). During gastrulation and germbandextension (stages 6 to 8), transcripts are stillexpressed uniformly. However, an anterior dorsalregion at the presumptive procephalic neuroblastsshows enhanced levels of dhem-2 transcription (Fig-ure 5D, E and F). At the extended germband stage(stage 12, Figure 5G) a uniform pattern is observed.However, seven patches (arrowheads in Figure 5G)show slightly higher levels of transcripts. Aftergermband retraction (stage 13, Figure 5H) expressionis uniform again. Slightly later (stage 14, Figure 5I),dhem-2 transcripts decay, except for the ventral chord,the supraesophageal ganglion and in the pharynx. Atstage 16 shortly before hatching, strong staining isstill observed in the ventral cord, in the brain (Fig-ure 5J). This pattern persists during late stages ofembryogenesis and during first instar larval stage(data not shown).
Expression analysis of vertebrate HEM genes
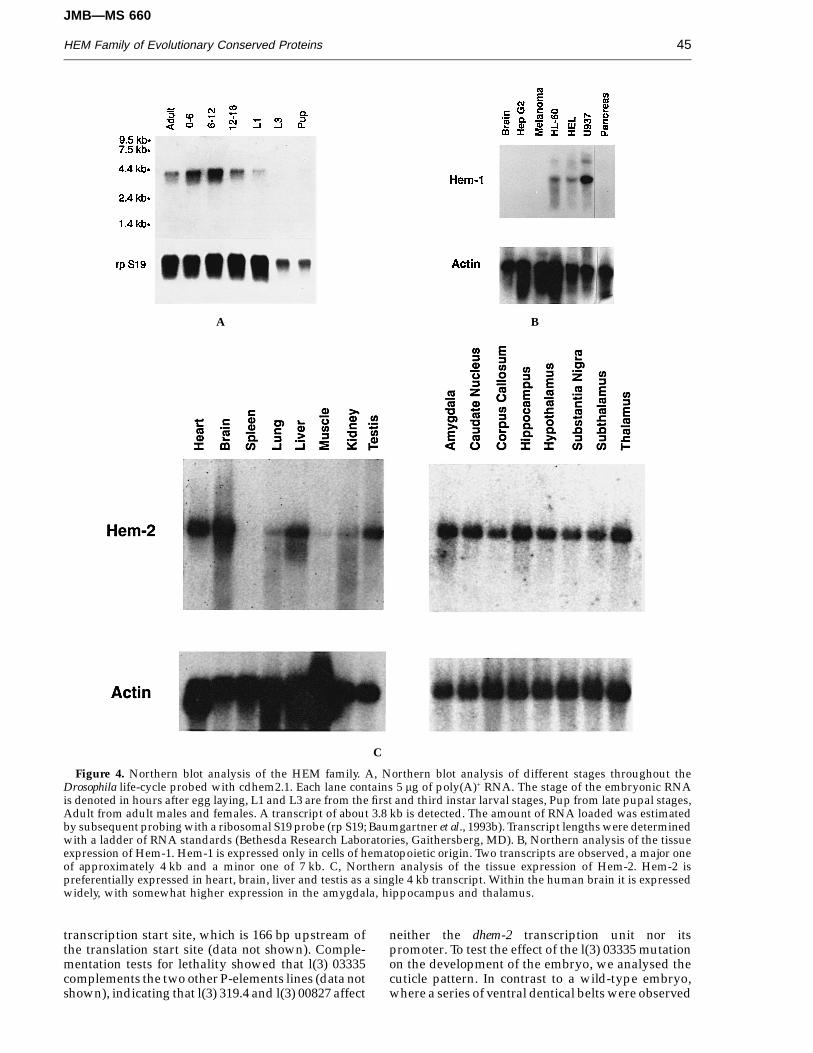
Hem-1 was previously found to be expressedpreferentially in cells of hematopoietic origin(Hromas et al., 1991). This was confirmed byNorthern analysis as seen in Figure 4B. A majortranscript of 4 kb is seen with a 7 kb transcriptpresent in lesser abundance. Hem-1 was expressed inthe myeloid leukemia line HL-60, the erythroidleukemia line HEL and the monocytic leukemia lineU937. It was not expressed in human brain, HepG2hepatoma cells or human pancreatic carcinoma cells,despite prolonged exposure of the Northern blot.Thus, Hem-1 expression was tightly restricted toblood cells.
Hem-2 was expressed in a wider range of tissues(Figure 4C). it was most highly expressed in brain,heart, liver and testis. Within the brain, it was alsowidely expressed. However, there was somewhathigher expression in the amygdala, hippocampusand thalamus. Hem-2 had only one transcript, of 4 kbin size.
Mutations in the dhem-2 gene cause amaternal lethal mutation
In a search for the function of the dhem-2 gene, wewere able to identify three independent lethalP-element insertion lines that all showed a singleP-element insertion around the putative promoterregion of dhem-2. As shown in Figure 1, l(3) 319.4and l(3) 00827 showed an insertion of the P-element 1.8 kb and 1.65 kb upstream of the puta-tive transcription site, respectively. l(3) 03335 showedthe insertion 39 bp downstream of the putative
terminus, the four last transmembrane segmentsappear clustered (Figure 2), indicating short intra-and extracellular loops between the transmembranedomains in this region. Neither member of the familyrevealed a clear signal cleavage site near thepredicted initiator methionine residue, suggestingthat the first hydrophobic stretch (around residue140) may be used as a signal peptide cleavage site(Von Heijne, 1986). We do not know, however,whether the N terminus is on the extracellular sideof the membrane, or whether it is intracellular.
Developmental and spatial expression of thedhem-2 gene: dhem-2 is expressed maternally
Northern blot analysis reveals a transcript of about3.8 kb in length (Figure 4A), which is dynamicallyexpressed during the Drosophila life-cycle. Thedhem-2 transcript is strongly expressed during earlystages of development, in particular during the first12 hours of embryogenesis. dhem-2 is expressed alsoin adults, mainly because dhem-2 is expressedmaternally (Figure 5A).
To analyze the spatial distribution of the dhem-2gene, we hybridized digoxigenin-labelled probes toovaries of females and to embryos (Baumgartner &Chiquet-Ehrismann, 1993a) dhem-2 transcripts are
JMB—MS 660
HEM Family of Evolutionary Conserved Proteins 45
A B
C
Figure 4. Northern blot analysis of the HEM family. A, Northern blot analysis of different stages throughout theDrosophila life-cycle probed with cdhem2.1. Each lane contains 5 mg of poly(A)+ RNA. The stage of the embryonic RNAis denoted in hours after egg laying, L1 and L3 are from the first and third instar larval stages, Pup from late pupal stages,Adult from adult males and females. A transcript of about 3.8 kb is detected. The amount of RNA loaded was estimatedby subsequent probing with a ribosomal S19 probe (rp S19; Baumgartner et al., 1993b). Transcript lengths were determinedwith a ladder of RNA standards (Bethesda Research Laboratories, Gaithersberg, MD). B, Northern analysis of the tissueexpression of Hem-1. Hem-1 is expressed only in cells of hematopoietic origin. Two transcripts are observed, a major oneof approximately 4 kb and a minor one of 7 kb. C, Northern analysis of the tissue expression of Hem-2. Hem-2 ispreferentially expressed in heart, brain, liver and testis as a single 4 kb transcript. Within the human brain it is expressedwidely, with somewhat higher expression in the amygdala, hippocampus and thalamus.
transcription start site, which is 166 bp upstream ofthe translation start site (data not shown). Comple-mentation tests for lethality showed that l(3) 03335complements the two other P-elements lines (data notshown), indicating that l(3) 319.4 and l(3) 00827 affect
neither the dhem-2 transcription unit nor itspromoter. To test the effect of the l(3) 03335 mutationon the development of the embryo, we analysed thecuticle pattern. In contrast to a wild-type embryo,where a series of ventral dentical belts were observed
JMB—MS 660
HEM Family of Evolutionary Conserved Proteins46
Figure 5. Spatial expression of the dhem-2 gene duringembryogenesis. Embryos are oriented with anterior to theleft and dorsal side up unless otherwise noted. A, Stage 10oocyte (stages are according to King, 1970); expression ofdhem-2 is in follicle cells (fc) and in the growing oocyte (o).B, Stage 1 embryo (embryonic stages are those ofCampos-Ortega & Hartenstein, 1985); dhem-2 is expresseduniformly during the early cleavage stages. C, Stage 6embryo, midsagittal focal plane; expression of dhem-2 isubiquitous with increased concentration on the dorsal sideof the embryo. D, Early stage 7 embryo; two bilateralregions on the dorsal side corresponding to the procephalicneuroblast region accumulate dhem-2 transcripts (arrow).E and F, Sagittal (E) and horizontal (F) view of stage 10embryos; two bilateral spots (arrows) in the procephalicneuroblast (pnb) region and possibly neuroblasts(arrowheads) show increased levels of dhem-2 transcripts.G, Stage 12 embryos; increased levels of dhem-2 transcriptsare observed in seven broad bands in both the ectodermaland mesodermal layers. H, Stage 13 embryo; expression isubiquitous. I, Stage 14 embryo; dhem-2 transcripts decay,except for the ventral chord (vc), in the supraesophagealganglion (spg) and in the pharynx (ph). J, Stage 17 embryoprior to hatching; dhem-2 transcripts are detectedexclusively in the ventral chord and the brain. (cf) Cephalicfurrow. The bar in H represents 100 mm.
Figure 6. dhem-2 causes female sterility. A, Cuticularpattern of a wild-type embryo shortly before hatching. B,Cuticular pattern of a l(3) 03335 larva. Note the absence ofany cuticular structures, indicating an early defect duringoogenesis and embryogenesis. Consequently, no cuticle issecreted. C, bicoid (bcd; Frigerio et al., 1986) mRNA stainingpattern in a stage 2 wild-type embryo (embryonic stagesare those of Campos-Ortega & Hartenstein, 1985). Note theaccumulation of bcd message at the anterior pole of theembryo. D to F, bcd mRNA staining in stage 2 l(3) 03335embryos. Note the patchy accumulation of bcd mRNA invesicle-like structures. The overall shape of the embryoappears unaltered. However, abnormal vacuoles andmembrane folds (denoted by arrows in F) are present in themutant. Anterior is always to the left. The bar in Fcorresponds to 100 mm.
mutant embryos (data not shown). These findingssuggested that developmental processes in thesemutant embryos are disturbed even earlier, beforethe expression of these three developmental markers.Therefore, we wished to stain dhem-2 mutantembryos with a gene whose transcripts areexpressed in the oocyte and early during embryoge-nesis. For this purpose, bicoid (bcd) was chosen(Frigerio et al., 1986). As shown in Figure 6C,normally the bcd mRNA is concentrated at theanterior pole, due to the fact that the message istransported into the oocyte during oogenesis andforms a gradient in the embryo (Frigerio et al., 1986).In dhem-2 mutant embryos, bcd was indeedexpressed. However, the expression was neitherlocalized nor was it strictly confined to the anteriorpole (Figure 6D to F). This may suggest that thegeneral oocyte body plan is still established in dhem-2mutants, but that major influences lead to arearrangement of the cyto-architecture of the oocyte.This is best exemplified in Figure 6F, where the bcdmessage is shown to be distributed dorsally over theanterior half of the embryo.
We did not detect a different size or any markedchanges in the overall morphology of mutant em-bryos as opposed to wild-type embryos (Figure 6Cversus D). However, mutant embryos showed anincreased number of vacuole-like structures (Fig-ure 6D to F). In addition, there were aberrant mem-brane folds seen in the mutant embryos. Therefore,
(Figure 6A), embryos derived from l(3) 03335mothers did not show any cuticle secreted (Fig-ure 6B), indicating an early dhem-2 function, beforethe cuticle is secreted.
Consequently, we wished to determine the criticalearly phase during which development becomesaberrant. For this reason, we stained mutant embryoswith a series of early developmental markers such asthe products of the fushi-tarazu ( ftz) gene (Krauseet al., 1988), the gene adjacent to dhem-2, the tenm gene(Baumgartner et al., 1994), and the engrailed (en) gene(DiNardo & O’Farrell, 1985). We did not detect anyexpression of the ftz, tenm or the en gene in dhem-2

JMB—MS 660
HEM Family of Evolutionary Conserved Proteins 47
we conclude that dhem-2 has a critical functionduring growth of the oocyte. Consequently, due toaberrant expression of bcd, no clear body axis isestablished in the embryo and the embryos probablydie during the first hours of development.
Discussion
The HEM family of conserved proteinsduring evolution
We have confirmed the existence of a small familyof proteins that is highly conserved from invert-ebrates through mammals. To date, the familyconsists of two types of proteins, which areexpressed in distinct tissues. One type is preferen-tially expressed in the central nervous system andoocytes, while the other is preferentially expressed inhematopoietic cells.
Related protein sequences may be classified intotwo types: orthologous sequences, which are foundin different species where the differences reflectspecies divergence, and paralogous sequences,which are derived from a gene duplication, typicallybefore separation of the species. Hence, we proposethat dhem-2, mh19, Hem-2, humtag and the two C.elegans ESTs are respective orthologues, while Hem-1is the corresponding paralogue. This implies that acommon ancestor species existed, which may haveharboured a prototype of a HEM gene. Since thenematodes harbour a copy of the hem-2 orthologue,it follows that this ancestral type must have existedvery early. Subsequently, gene duplications events, incombination with the development of tissuespecification, such as the formation of the centralnervous system or blood cells, could have led to theappearance of two protein types.
The function of the Hem-2 genes may beconserved during evolution
We have shown that dhem-2 is expressedmaternally. Furthermore, one lethal P-elementinsertion in the dhem-2 5' untranslated region wasshown to lead to female sterility, indicating that thedhem-2 gene is essential for Drosophila oogenesis andearly embryogenesis.
The involvement of the Hem-2 genes in the earlyprocesses during oogenesis is supported by studieson C. elegans genetics. A closer inspection of the twoC. elegans EST clones revealed that the C. elegans hem-2orthologue resides in a chromosomal region that hadpreviously been subjected to a mutagenesis screen(Clark et al., 1988). A number of lethal complementa-tion groups were established, one of which (let 99)caused a maternal lethal phenotype (Clark et al.,1988; Charest et al., 1990). Thus, it is possible that let99 is encoded by the C. elegans Hem-2 orthologue,indicating that the function of the Hem-2 gene alsomay be conserved in nematodes.
The findings mentioned above indicate that thevertebrate Hem-2 orthologues could have similar
functions early in oogenesis and embryogenesis.Indeed, we were intrigued by the early phenotype inoocytes. There is no clear explanation as to how atransmembrane protein can act in the growingoocyte. The abnormal membrane folds and vacuolesseen in the mutant embryos imply a disruption ofmembrane structure. However, since the dhem-2mRNA is synthesized in nurse cells but not in folliclecells, and is distributed uniformly in the oocyte(Figure 5A and B), we cannot exclude the possibilitythat maternal dhem-2 functions at the RNA level.Indeed, the finding that bcd expression was disruptedin the dhem-2 mutant suggests that dhem-2 may havea role in development prior to bcd, and may directlyor indirectly regulate bcd expression.
Furthermore, the later expression pattern ofdhem-2, as well as its vertebrate orthologue, suggestsdiverse functions during development. In the case ofdhem-2, a later requirement during neurogenesis islikely. However, the analysis of such a function ishampered by the early effect of dhem-2 in oocytes. Incontrast, expression of the Hem-1 gene has beenshown to be restricted to blood cells. Therefore, itprobably exerts a highly specialized function duringthe development of the blood cells.
Evolutionary considerations
The HEM family appears to be conservedthroughout evolution, being found even in lowerorganisms such as nematodes. However, thehomology of the nematode orthologue to thevertebrate orthologue is lower than the Drosophilaorthologue to its vertebrate counterpart. No HEMprotein orthologue has been found so far in plants,fungi or bacteria.
Our analysis indicates that a Drosophila orthologueof Hem-1 may exist as well. This putative gene isexpected to be expressed in the Drosophilahemocytes, which form a population of isolated cellsthat are scattered throughout the entire hemolymphspace (Rizki, 1978). In contrast to vertebrate bloodcells these hemocytes do not have the capacity tocarry oxygen. They do, however, have phagocyticproperties.
Although the mh19 cDNA was picked up in ascreen for brain-specific genes, the analysis of its ratorthologue Hem-2 showed expression in severalother tissues. Besides brain, Hem-2 was expressed inheart, liver and testis. The expression of Hem-2 inbrain and germ cells is consistent with dhem-2expression in the developing brain and oocytes ofDrosophila. However, we were not able to detectdhem-2 expression in the heart analogue of the flies,in the cardiac cells.
The idea that many proteins are constructed fromvarious modules, each identifiable by a consensussequence that is sometimes related to the exonstructure of the gene, is becoming increasinglyfamiliar (Bork, 1992). In our case, however, we do nothave any evidence that domains of the HEM familyof proteins are shared with other proteins. Rather, itappears to constitute a family that has retained its

JMB—MS 660
HEM Family of Evolutionary Conserved Proteins48
autonomy in the protein world. The function of thisfamily remains an intriguing question. Since it doesnot share domain homology to other protein families,there are few clues as to its function in cellularmetabolism.
Accession numbers
The EMBL accession number for dhem-2 cDNA isX80028, for human Hem-1 M58285, for rat Hem-2X80029, for mouse mh19 X61453, for humtagEST00609 M78461 and for the two C. elegans ESTsequences D27398 and D34785.
Materials and Methods
DNA and RNA techniques
Southern and Northern blot analyses were performed bystandard procedures (Maniatis et al., 1989). RNA wasextracted by the guanidium thiocyanate/phenol/chloro-form extraction method described by Chomczynski &Sacchi (1987). Poly(A)+ RNA was isolated using aPharmacia Kit (Pharmacia, Piscataway, NJ).
Isolation of genomic and cDNA clones
A 0.6 kb SacII–EcoRI terminal fragment of DT 24 of tenm
(Baumgartner et al., 1994) was used to isolate genomicclones of dhem-2 from a Drosophila cosmid library(Stratagene, La Jolla, CA) and cDNA clones from an 8 to12 hour cDNA library (Brown & Kafatos, 1988). To isolateHem-2, radiolabelled mh19 cDNA was used to screen asize-selected (greater than 4.0 kb) rat brain cDNA libraryconstructed in lambda Zap according to the manufac-turer’s instructions (Stratagene). Three cDNA clones wereisolated that contained inserts equivalent in size to thetranscripts seen on Northern analysis.
Sequencing
Genomic fragments and cDNAs were subcloned into theEcoRI site of pBluescript (Stratagene). Both strands weredideoxy-sequenced using the Sequenase Kit (U.S.Biochemicals, Cleveland, OH) and a nested deletion kit(Pharmacia).
Whole-mount in situ hybridization
Whole-mount in situ hybridizations were conductedusing digoxigenin-labelled cdhem2.1 (Figure 1) andc53.46.6 (bcd; Frigerio et al., 1986) following the protocol ofTautz & Pfeiffle (1989), as we have previously used(Baumgartner & Chiquet-Ehrismann, 1993a).
Fly stocks
All alleles (1(3) 00827, 03335 and 319.4) were obtainedin two screens for P-element insertion lines (Karpen& Spradling, 1992; N. Walter, unpublished results). Weperformed a complementation analysis for lethalitybetween the insertion lines as a matrix of each line againstthe rest of the alleles. These crosses showed that l(3) 03335complements the two others lines, indicating that l(3) 319.4and l(3) 00827 cause a lethal mutation, but not in thedhem-2 gene. To generate revertants of l(3) 03335, P[PZ]was mobilized with the endogenous D2–3 transposase
(Robertson et al., 1988) and several non-lethal revertantlines were obtained. These were found, by Southernanalysis and PCR, to probably represent perfect P-elementexcisions. The cuticular phenotype of these revertants wasindistinguishable from wild-type.
Immunohistochemistry
Peroxidase immunohistochemistry was performed aswe previously described (Hromas et al., 1993; Baumgart-ner et al., 1994). Polyclonal rabbit antisera were raised atPocono Rabbit Farm (Canadensis, PA) according to theirsynthetic peptide protocol. The antigen used was a peptidecontaining the first 35 amino residues of Hem-1 coupled toKeyhole limpet hemocyanin.
AcknowledgementsWe thank Markus Noll for providing libraries, Patrick
Matthias for human DNA and Joachim Wittbrodt forzebrafish DNA. We thank the Gehring laboratory forproviding anti-Ftz and anti-En antibodies. We thank JohnDaly for critically reading the manuscript. The secretarialhelp of Luigina Beffa is warmly acknowledged. R.H. is aScholar of the Leukemia Society of America, and issupported by NIH HL 48914.
ReferencesAdams, M. D., Brubuick, M., Kerlavage, A. R., Moreno, R.,
Kelley, J. M., Utterbach, T. R., Nagle, J. W., Fields, C.& Venter, J. C. (1992). Sequence identification of 2375human brain genes. Nature, 355, 632–634.
Baumgartner, S. & Chiquet-Ehrismann, R. (1993a). Tena, aDrosophila gene related to tenascin, shows selectivetranscript localization. Mech. Dev. 40, 165–176.
Baumgartner, S., Martin, D. & Chiquet-Ehrismann, R.(1993b). Drosophila ribosomal protein S19 sequence.Nucl. Acids Res. 21, 3897.
Baumgartner, S., Martin, D., Hagios, C. & Chiquet-Ehris-mann, R. (1994). Tenm, a Drosophila gene related totenascin, is a new pair-rule gene. EMBO J. 13,3728–3740.
Bork, O. (1992). Mobile modules and motifs. Curr. Opin.Struct. Biol. 2, 413–421.
Brown, N. H. & Kafatos, F. C. (1988). Functional cDNAlibraries from Drosophila embryos. J. Mol. Biol. 203,425–437.
Campos-Ortega, J. A. & Hartenstein, V. (1985). TheEmbryonic Development of Drosophila melanogaster,Springer, Berlin.
Charest, D. L., Clark, D. V., Green M. E. & Baillie, D. L.(1990). Genetic and fine-structure analysis of unc-26(IV) and adjacent regions in Caenorhabditis elegans. Mol.Gen. Genet. 221, 459–465.
Cherbas, L. & Cherbas, P. (1993). The arthropod initiator:the capsite consensus plays an important role intranscription. Insect Biochem. Mol. Biol. 23, 81–90.
Chomczynski, P. & Sacchi, N. (1987). Single step method ofRNA isolation by acid guanidium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162, 156–159.
Clark, D. V., Rogalski, T. M., Donati, L. M. & Baillie, D. L.(1988). The unc 22 (IV) region of Caenorabditis elegans:Genetic analysis of lethal mutations. Genetics, 119,345–353.
DiNardo, S. & O’Farrell, P. J. (1985). Development of

JMB—MS 660
HEM Family of Evolutionary Conserved Proteins 49
embryonic pattern in D. melanogaster as revealed byaccumulation of the nuclear engrailed protein. Cell, 43,59–69.
Doolittle R. F. (1992). Reconstructing history with aminoacid sequences. Protein Sci. 1, 191–200.
Frigerio, G., Burri, M., Bopp, D., Baumgartner, S. & Noll,M. (1986). Structure of the segmentation gene pairedand the Drosophila PRD gene set as part of a genenetwork. Cell, 47, 735–746.
Hromas, R., Collins, S., Raskind, W., Deaven, L. &Kaushansky, K. (1991). Hem-1, a potential membraneprotein, with expression restricted to blood cells.Biochim. Biophys. Acta, 1090, 241, 244.
Hromas, R., Orazi, A., Neiman, R. S., Maki, R., VanBeveran, C., Moore, J. & Klemsz, M. (1993).Hematopoietic lineage and stage-restricted ex-pression of the ETS oncogene family member PU.1.Blood, 82, 2998–3004.
Karpen, G. H. & Spradling, A. C. (1992). Analysis ofsubtelomeric heterochromatin in a Drosophilaminichromosome Dp 1187 by single P elementinsertion mutagenesis. Genetics, 132, 737–753.
Kato, K. (1990). A collection of cDNA clones with specificexpression patterns in mouse brain. Eur. J. Neurosci. 2,704–711.
King, R. C. (1970). Ovarian Development in Drosophilamelanogaster, Academic Press, New York.
Krause, H. M., Klemenz, R. & Gehring, W. J. (1988).Expression, modification, and localization of thefishi-tarazu protein in Drosophila embryos. Genes Dev.2, 1021–1036.
Maniatis, T., Fritsch, E. F. & Sambrook, J. (1989). MolecularCloning: A Laboratory Manual, Cold Spring HarborLaboratory press, Cold Spring Harbor, NY.
Rizki, T. M. (1978). The circulatory system and associatedcells and tissue. In: Genetics and Biology of Drosophila,vol. 2b, pp. 397–452, Academic Press, New York.
Robertson, H. N., Preston, C. R., Phillis, R. W.,Johnson-Schlitz, D. M., Benz, W. K. & Engels, W.(1988). A stable genomic source of P-elementtransposase in Drosophila. Genetics, 118, 461–470.
Smoller, D., Friedel, C., Schmid, A., Bettler, D., Lam, L. &Yedvobnick, B. (1990). The Drosophila neurogeniclocus mastermind encodes a nuclear protein unusuallyrich in amino acids homopolymers. Genes Dev. 4,1688–1700.
Tautz, D. & Pfeiffle, C. (1989). A non-radioactive in situhybridization method for the localization of specificRNAs in Drosophila reveals translational control ofthe segmentation gene hunchback. Chromosoma, 98,81–85.
Von Heijne, G. (1986). A new method for predictingsignal sequence cleavage sites. Nucl. Acids Res. 14,4683–4690.
Edited by J. Karn
(Received 20 March 1995; accepted 11 May 1995)
Related Documents









![BMC Bioinformatics BioMed CentralMembrane proteins are a large fraction (about 30%) of all the proteins found in living organisms [1]. Knowledge of the location of transmembrane (TM)](https://static.cupdf.com/doc/110x72/5fca0e6942501b0531378c2a/bmc-bioinformatics-biomed-central-membrane-proteins-are-a-large-fraction-about.jpg)

