This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Acta Tropica 115 (2010) 194–199
Contents lists available at ScienceDirect
Acta Tropica
journa l homepage: www.e lsev ier .com/ locate /ac ta t ropica
The giant African snail Achatina fulica as natural intermediate host ofAngiostrongylus cantonensis in Pernambuco, northeast Brazil
S.C. Thiengoa,∗, A. Maldonadob, E.M. Motac, E.J.L. Torresd, R. Caldeirae, O.S. Carvalhoe,A.P.M. Oliveiraa, R.O. Simõesb, M.A. Fernandeza, R.M. Lanfredid,1
a Laboratório de Malacologia, Instituto Oswaldo Cruz/Fiocruz, Av. Brasil 4365 Manguinhos 21.040-900, RJ, Brazilb Laboratório de Biologia e Parasitologia de Mamíferos Silvestres Reservatórios, Instituto Oswaldo Cruz/Fiocruz, Av. Brasil 4365 Manguinhos 21.040-900, RJ, Brazilc Laboratório de Patologia, Instituto Oswaldo Cruz/Fiocruz, Av. Brasil 4365 Manguinhos 21.040-900, RJ, Brazild Laboratório de Biologia de Helmintos Otto Wucherer, Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, RJ, Brazile Laboratório de Helmintologia e Malacologia Médica, Centro de Pesquisas René Rachou/Fiocruz, Av. Augusto Lima, 1715, 30190-002, Belo Horizonte, MG, Brazil
a r t i c l e i n f o
Article history:Received 2 September 2009Received in revised form 7 January 2010Accepted 8 January 2010Available online 18 January 2010
Keywords:Achatina fulicaPernambucoAngiostrongylus cantonensisMetastrongylidaeEpidemiologyInvasive species
a b s t r a c t
The human cases of eosinophilic meningitis recently reported from Brazil have focused the attention ofthe public health agencies on the role the introduced snail Achatina fulica plays as hosts of the metas-trongylid nematodes. Determining the potential of this snail to host and develop infective larval stages ofmetastrongylids in the wild and identify the species harbored by them is crucial for designing effectivecontrol measures. Here we assess if A. fulica may act as intermediate host of A. cantonensis at the peridomi-ciliary areas of a patient’s house from state of Pernambuco (PE), who was diagnosed with eosinophilicmeningitis and a history of ingesting raw molluscs. Larvae obtained from naturally infected A. fulica wereorally administered to Rattus norvegicus. The worms were collected from the pulmonary artery and brain,and were morphologically characterized and compared to the Japan isolate of A. cantonensis. Adult wormsand infective L3 larvae (PE isolate) recovered from A. fulica specimens were also analyzed by polymerasechain reaction and restriction fragment length polymorphism of ITS2 region from rDNA and comparedto A. cantonensis (ES isolate), A. vasorum (MG isolate) and A. costaricensis (RS isolate). The large size of thespicules (greater than those observed in other species of Angiostrongylus) and the pattern of the bursalrays agree with the original species description by Chen (1935). Furthermore, the morphology of the PEisolate was similar to that of Japan isolate. The PCR-RFLP profiles obtained were distinctive among speciesand no variation in patterns was detected among adult individuals from A. cantonensis isolates from PEand ES. The importance of A. fulica as an intermediate host of eosinophilic menigoencepahlitis in Brazilis emphasized.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Achatina (Lissachatina) fulica Bowdich, 1822, the giant Africansnail, has been introduced throughout the tropics and subtropicsand has been considered an important snail pest in these regions.Introduced to Brazil in the 1980s for commercial purposes (“escar-got” farming) A. fulica is now widespread in at least 24 of 26Brazilian states and the Federal District (Brasília), including theAmazonian region and natural reserves, where it is a pest in orna-mental gardens, vegetable gardens, and small-scale agriculture(Thiengo et al., 2007).
∗ Corresponding author. Fax: +55 21 2560 2357.E-mail address: [email protected] (S.C. Thiengo).
1 In memoriam.
Brazil is currently experiencing the explosive phase of the A.fulica invasion. This rapid spread throughout almost the entirecountry followed a pattern similar to the initial introduction in thestate of Paraná, that is, distribution for commercial purposes fol-lowed by the release of the snails into the wild when people gaveup the enterprise (Thiengo et al., 2007).
In addition to the impact on ecosystem health and potentialcompetition with native terrestrial molluscs, A. fulica can alsoact as an intermediate host of nematodes of medical and vet-erinary importance (Thiengo et al., 2008; Carvalho et al., 2003).Among them are, Angiostrongylus cantonensis (Chen, 1935), anematode that can cause eosinophillic meningoencephalitis andAngiostrongylus costaricensis Morera and Céspedes, 1971, whichcauses abdominal angiostrongylosis. The global dispersal of A. can-tonensis, endemic to some Asian countries and Pacific Islands, isclosely associated with the spread of A. fulica (Kliks and Palumbo,1992; Lv et al., 2009b). In 2007, Brazil reported its first two cases of
0001-706X/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.actatropica.2010.01.005

Author's personal copy
S.C. Thiengo et al. / Acta Tropica 115 (2010) 194–199 195
Table 1Measurements (�m) of L1 larvae of Angiostrongylus cantonensis (PE, Brazil isolate) recovered from Rattus norvegicus feces experimentally infected and L3 larvae from Achatinafulica collected in Pernambuco state.
Characteristics n L1 n L3
Mean ± SE Range Mean ± SE Range
Body length 25 254.4 ± 23.2 304.3–206 34 460.4 ± 31.8 544.6–460.3Width 25 10.9 ± 2.4 15.1–5.2 34 24.8 ± 3.8 31.9–18.5Esophagus 24 108.1 ± 14.8 137.6–71.2 34 168.4 ± 11.2 189.0–129.6Nerve ring 8 40.2 ± 12.3 66.4–27.2 30 90.9 ± 27.0 76.3–51.0Excretory pore 25 70.8 ± 17.2 101.4–32.3 33 157.9 ± 28.8 200–79.6Genital primordium 17 9.82 ± 4.1 18.1–5.2 14 40.8 ± 14.0 65.9–22.2Anus–tail 15 26.1 ± 9.4 57.6–12.3 29 41.2 ± 10.4 89.3–27.7
this zoonosis in the municipality of Cariacica, state of Espírito Santo.Both instances were the result of ingestion of raw veronicellid slugs(Caldeira et al., 2007).
Populations of A. fulica, generally with many large individu-als, occur in urban areas and are often a nuisance because of thehigh density of their populations, which often leads people to seekassistance in controlling them from local public health institutions.For the past seven years, the Laboratório de Malacologia do Insti-tuto Oswaldo Cruz/Fiocruz (LRNM), National Reference for MedicalMalacology, has examined samples of A. fulica sent by the Brazil-ian health and environmental agencies for larvae of Angiostrongylusspp.
We report the first finding of A. fulica naturally infected with A.cantonensis larvae in northeast Brazil and present the results of lightmicroscopy morphology of adult and larvae nematodes obtained byexperimental infection as well as the molecular diagnostic of boththird-stage larvae (L3) and adults. We also discuss the epidemio-logical aspects of eosinophilic meningitis in Brazil.
2. Material and methods
In March 2008 a 6-year-old female, with anamnesis of ingestingraw molluscs from her yard, in the municipality of Escada, stateof Pernambuco (PE), was diagnosed with meningitis in the hos-pital Correto Picanco in Recife, PE, Brazil. Three months later, a26-year-old female from the municipality of Olinda, PE, was diag-nosed with the same condition. Similarly to the other patient, theanamnesis emphasized the ingestion of raw molluscs from her yard.Therefore, in June and September 2008 collection of terrestrial andfreshwater molluscs in peridomiciliary areas of the two patients’houses was undertaken by us and health agents from the Secre-taria Estadual de Saúde de Pernambuco. In all 1544 molluscs werecollected randomly. The cephalopodal mass of each of the molluscswas individually minced and digested in a 0.7% HCl solution for6 h. The digested samples were placed in a Baermann apparatusand allowed to sediment overnight. Infective A. cantonensis larvaeobtained from A. fulica collected in Olinda were used to infect 30three-month-old Rattus norvegicus, Wistar strain, at a concentra-tion of 100 L3/animal (Youssif and Ibrahim, 1978).
Nematodes collected from the brain and the pulmonary artery30 and 120 days after infection, respectively, were washed in phys-iologic solution and fixed in hot AFA solution (2% glacial aceticacid, 3% formaldehyde, and 95% ethanol). Specimens (10 males and10 females) from each of those sites were cleared in lactophenol,mounted on slides in lactophenol solution and examined under alight microscope. Drawings for the morphometric analyses weremade using a light microscope with the aid of a camera lucida.Taxonomic identification of nematodes was based on morphologi-cal and morphometric parameters following Anderson (1978) andAsh (1970). Specimens of A. cantonensis isolate from Akita, Japanwere used for comparison. Measurements are given in micrometers(unless otherwise stated).
Ten male and 18 female adult helminthes, and a pool of L3 werefixed in >90% ethanol and washed three times in distilled waterprior to use in molecular analysis. Genomic DNA (gDNA) extrac-tions were undertaken using the Wizard Genomic Purification Kit(PROMEGA, Madison, USA) according to manufacturer’s instruc-tions. Comparative material was obtained from other nematodesamples provided by the Laboratório de Helmintologia e Malacolo-gia Médica do Centro de Pesquisas René Rachou, Fiocruz including:A. cantonensis (from Cariacica, state of Espírito Santo), A. costari-censis (from state of Rio Grande do Sul) and A. vasorum (from the
Fig. 1. Light microscopy of third-stage larvae (L3) of Angiostrongylus cantonensis.(a) Anterior end showing knob-like tips (KT), rod-like structure (RS), esophagus (E),excretory pore (EP); (b) posterior end showing tail pointed tip (TPT) and anus (A).
Author's personal copy
196 S.C. Thiengo et al. / Acta Tropica 115 (2010) 194–199
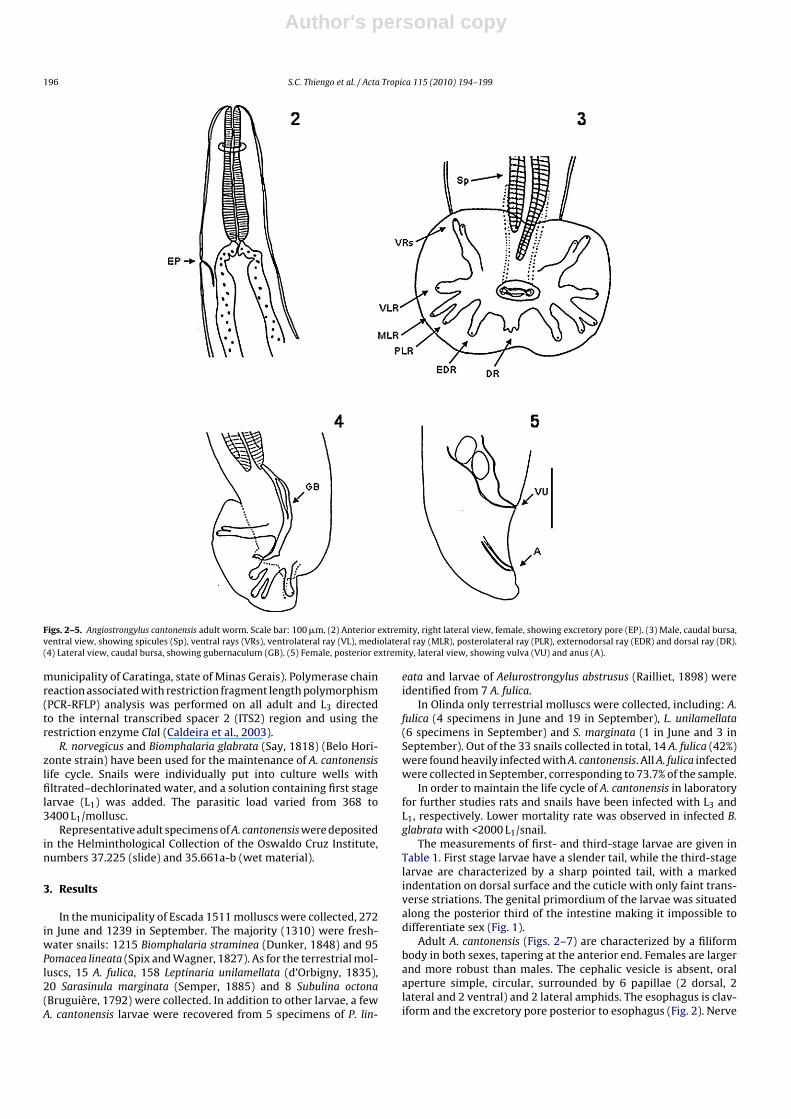
Figs. 2–5. Angiostrongylus cantonensis adult worm. Scale bar: 100 �m. (2) Anterior extremity, right lateral view, female, showing excretory pore (EP). (3) Male, caudal bursa,ventral view, showing spicules (Sp), ventral rays (VRs), ventrolateral ray (VL), mediolateral ray (MLR), posterolateral ray (PLR), externodorsal ray (EDR) and dorsal ray (DR).(4) Lateral view, caudal bursa, showing gubernaculum (GB). (5) Female, posterior extremity, lateral view, showing vulva (VU) and anus (A).
municipality of Caratinga, state of Minas Gerais). Polymerase chainreaction associated with restriction fragment length polymorphism(PCR-RFLP) analysis was performed on all adult and L3 directedto the internal transcribed spacer 2 (ITS2) region and using therestriction enzyme ClaI (Caldeira et al., 2003).
R. norvegicus and Biomphalaria glabrata (Say, 1818) (Belo Hori-zonte strain) have been used for the maintenance of A. cantonensislife cycle. Snails were individually put into culture wells withfiltrated–dechlorinated water, and a solution containing first stagelarvae (L1) was added. The parasitic load varied from 368 to3400 L1/mollusc.
Representative adult specimens of A. cantonensis were depositedin the Helminthological Collection of the Oswaldo Cruz Institute,numbers 37.225 (slide) and 35.661a-b (wet material).
3. Results
In the municipality of Escada 1511 molluscs were collected, 272in June and 1239 in September. The majority (1310) were fresh-water snails: 1215 Biomphalaria straminea (Dunker, 1848) and 95Pomacea lineata (Spix and Wagner, 1827). As for the terrestrial mol-luscs, 15 A. fulica, 158 Leptinaria unilamellata (d’Orbigny, 1835),20 Sarasinula marginata (Semper, 1885) and 8 Subulina octona(Bruguière, 1792) were collected. In addition to other larvae, a fewA. cantonensis larvae were recovered from 5 specimens of P. lin-
eata and larvae of Aelurostrongylus abstrusus (Railliet, 1898) wereidentified from 7 A. fulica.
In Olinda only terrestrial molluscs were collected, including: A.fulica (4 specimens in June and 19 in September), L. unilamellata(6 specimens in September) and S. marginata (1 in June and 3 inSeptember). Out of the 33 snails collected in total, 14 A. fulica (42%)were found heavily infected with A. cantonensis. All A. fulica infectedwere collected in September, corresponding to 73.7% of the sample.
In order to maintain the life cycle of A. cantonensis in laboratoryfor further studies rats and snails have been infected with L3 andL1, respectively. Lower mortality rate was observed in infected B.glabrata with <2000 L1/snail.
The measurements of first- and third-stage larvae are given inTable 1. First stage larvae have a slender tail, while the third-stagelarvae are characterized by a sharp pointed tail, with a markedindentation on dorsal surface and the cuticle with only faint trans-verse striations. The genital primordium of the larvae was situatedalong the posterior third of the intestine making it impossible todifferentiate sex (Fig. 1).
Adult A. cantonensis (Figs. 2–7) are characterized by a filiformbody in both sexes, tapering at the anterior end. Females are largerand more robust than males. The cephalic vesicle is absent, oralaperture simple, circular, surrounded by 6 papillae (2 dorsal, 2lateral and 2 ventral) and 2 lateral amphids. The esophagus is clav-iform and the excretory pore posterior to esophagus (Fig. 2). Nerve
Author's personal copy
S.C. Thiengo et al. / Acta Tropica 115 (2010) 194–199 197
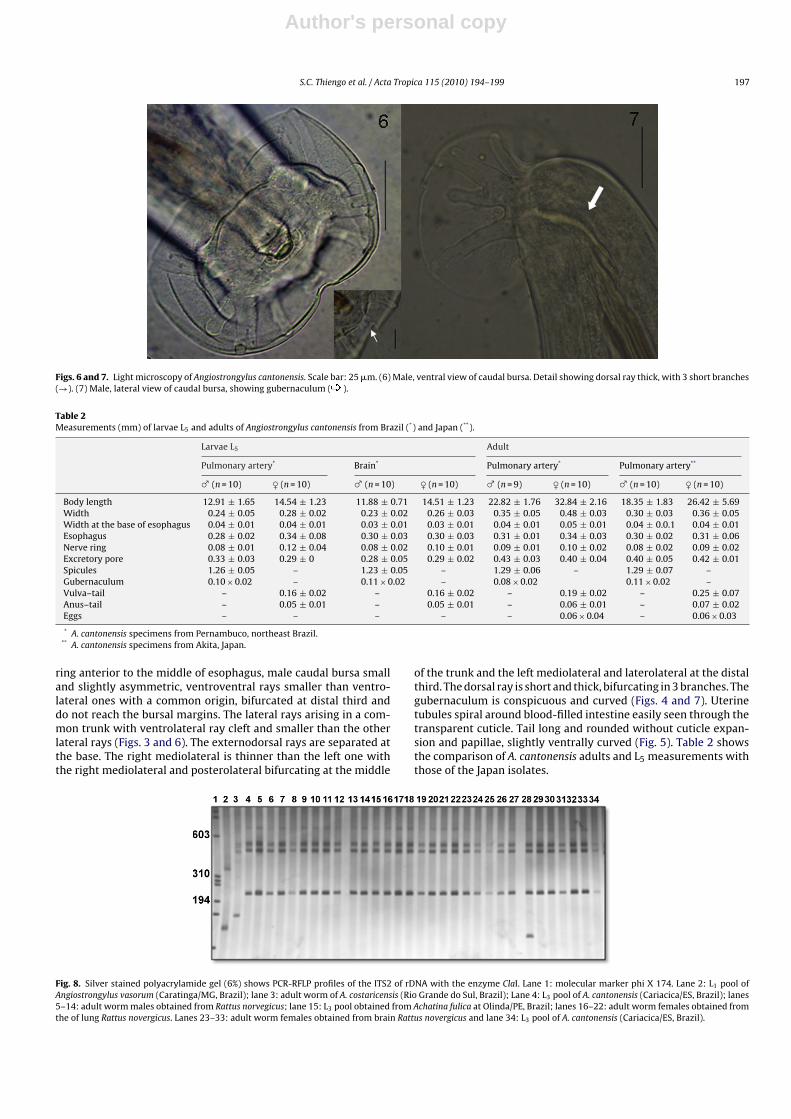
Figs. 6 and 7. Light microscopy of Angiostrongylus cantonensis. Scale bar: 25 �m. (6) Male, ventral view of caudal bursa. Detail showing dorsal ray thick, with 3 short branches(→). (7) Male, lateral view of caudal bursa, showing gubernaculum ( ).
Table 2Measurements (mm) of larvae L5 and adults of Angiostrongylus cantonensis from Brazil (*) and Japan (**).
Larvae L5 Adult
Pulmonary artery* Brain* Pulmonary artery* Pulmonary artery**
♂ (n = 10) ♀ (n = 10) ♂ (n = 10) ♀ (n = 10) ♂ (n = 9) ♀ (n = 10) ♂ (n = 10) ♀ (n = 10)
Body length 12.91 ± 1.65 14.54 ± 1.23 11.88 ± 0.71 14.51 ± 1.23 22.82 ± 1.76 32.84 ± 2.16 18.35 ± 1.83 26.42 ± 5.69Width 0.24 ± 0.05 0.28 ± 0.02 0.23 ± 0.02 0.26 ± 0.03 0.35 ± 0.05 0.48 ± 0.03 0.30 ± 0.03 0.36 ± 0.05Width at the base of esophagus 0.04 ± 0.01 0.04 ± 0.01 0.03 ± 0.01 0.03 ± 0.01 0.04 ± 0.01 0.05 ± 0.01 0.04 ± 0.0.1 0.04 ± 0.01Esophagus 0.28 ± 0.02 0.34 ± 0.08 0.30 ± 0.03 0.30 ± 0.03 0.31 ± 0.01 0.34 ± 0.03 0.30 ± 0.02 0.31 ± 0.06Nerve ring 0.08 ± 0.01 0.12 ± 0.04 0.08 ± 0.02 0.10 ± 0.01 0.09 ± 0.01 0.10 ± 0.02 0.08 ± 0.02 0.09 ± 0.02Excretory pore 0.33 ± 0.03 0.29 ± 0 0.28 ± 0.05 0.29 ± 0.02 0.43 ± 0.03 0.40 ± 0.04 0.40 ± 0.05 0.42 ± 0.01Spicules 1.26 ± 0.05 – 1.23 ± 0.05 – 1.29 ± 0.06 – 1.29 ± 0.07 –Gubernaculum 0.10 × 0.02 – 0.11 × 0.02 – 0.08 × 0.02 0.11 × 0.02 –Vulva–tail – 0.16 ± 0.02 – 0.16 ± 0.02 – 0.19 ± 0.02 – 0.25 ± 0.07Anus–tail – 0.05 ± 0.01 – 0.05 ± 0.01 – 0.06 ± 0.01 – 0.07 ± 0.02Eggs – – – – – 0.06 × 0.04 – 0.06 × 0.03
* A. cantonensis specimens from Pernambuco, northeast Brazil.** A. cantonensis specimens from Akita, Japan.
ring anterior to the middle of esophagus, male caudal bursa smalland slightly asymmetric, ventroventral rays smaller than ventro-lateral ones with a common origin, bifurcated at distal third anddo not reach the bursal margins. The lateral rays arising in a com-mon trunk with ventrolateral ray cleft and smaller than the otherlateral rays (Figs. 3 and 6). The externodorsal rays are separated atthe base. The right mediolateral is thinner than the left one withthe right mediolateral and posterolateral bifurcating at the middle
of the trunk and the left mediolateral and laterolateral at the distalthird. The dorsal ray is short and thick, bifurcating in 3 branches. Thegubernaculum is conspicuous and curved (Figs. 4 and 7). Uterinetubules spiral around blood-filled intestine easily seen through thetransparent cuticle. Tail long and rounded without cuticle expan-sion and papillae, slightly ventrally curved (Fig. 5). Table 2 showsthe comparison of A. cantonensis adults and L5 measurements withthose of the Japan isolates.
Fig. 8. Silver stained polyacrylamide gel (6%) shows PCR-RFLP profiles of the ITS2 of rDNA with the enzyme ClaI. Lane 1: molecular marker phi X 174. Lane 2: L1 pool ofAngiostrongylus vasorum (Caratinga/MG, Brazil); lane 3: adult worm of A. costaricensis (Rio Grande do Sul, Brazil); Lane 4: L3 pool of A. cantonensis (Cariacica/ES, Brazil); lanes5–14: adult worm males obtained from Rattus norvegicus; lane 15: L3 pool obtained from Achatina fulica at Olinda/PE, Brazil; lanes 16–22: adult worm females obtained fromthe of lung Rattus novergicus. Lanes 23–33: adult worm females obtained from brain Rattus novergicus and lane 34: L3 pool of A. cantonensis (Cariacica/ES, Brazil).

Author's personal copy
198 S.C. Thiengo et al. / Acta Tropica 115 (2010) 194–199
PCR-RFLP analysis of the ITS2 rDNA confirmed the identity of alladult worms and L5 obtained from the brain and lungs of R. norvegi-cus and infective larvae recovered from A. fulica as A. cantonensis(Fig. 8, lanes 5–33). The band from the adult worm in lane 28 exhib-ited polymorphism when compared with the other specimens dueto an additional band.
4. Discussion
The comparison of A. cantonensis adults and L5 measurementswith those of the Japan isolates showed a high degree of mor-phometric similarity (Table 2), confirming morphological andmolecular analyses. Similarly to the L3 larvae well compared tothose described by Ash (1970). The only variation observed in adultworms was an additional band (Fig. 8, lane 28). As there are manycopies of the rDNA cistrons in the genome, we believe that thevariation in adult worm profile could be due to heterogeneity, i.e.,the presence of two or more sequence types of the ITS in a singleorganism. This variation has been also described in other nema-tode parasites (Stevenson et al., 1995; Gasser et al., 1996) suchas Haemonchus contortus (Rudolphi, 1803) Cobb, 1898 and Ancy-lostoma caninum (Ercolani, 1859).
A. cantonensis and other congeneric species such as A. costari-censis have low specificity for their intermediate hosts and manydifferent freshwater and terrestrial mollusc species had been foundnaturally infected (Wallace and Rosen, 1969; Graeff-Teixeira et al.,1993; Caldeira et al., 2007; Lv et al., 2009a,b). Among them, A. fulicaplays an important role in the worldwide transmission of A. canto-nensis (Kliks and Palumbo, 1992; Thiengo, 2007; Lv et al., 2009b).The finding of infected specimens of A. fulica in the neighborhoodof the patients in both the southeast (Caldeira et al., 2007) andnortheast regions of Brazil reinforces the emerging public healththreat presented by this species and closely related congenerics inNeotropical regions. In addition, A. fulica was found infected withother nematodes of veterinary importance such as A. abstrusus inseven Brazilian States from the southeast, northeast and midwestregions (Thiengo et al., 2008).
Similar to A. fulica, ampullariid species, including some fromthe Neotropics, mainly Pomacea canaliculata (Lamarck, 1822) intro-duced in Hawaii and Asia for commercial proposes, are implicatedin numerous cases and outbreaks of eosinophilic meningitis inthose regions, especially China (Tsai et al., 2001; Lv et al., 2009a,b).In the present study, although the parasitic index of P. lineata frommunicipality of Escada was low (5.26%) comparing with that foundin the A. fulica from Olinda (42%), the patient probably got infectedfrom the consumption of undercooked P. lineata, according to itsclinical history.
It is noteworthy to mention that although there have been 4cases of eosinophilic meningitis resulting from the ingestion ofundercooked, or raw molluscs in Brazil, it is not common practicefor Brazilians to consume molluscs prepared in this manner. This isin contrast to the common practice of eating raw or undercookedsnails in many parts of Asia (Lv et al., 2009a,b). This points to theneed for a different focus for preventing infections in Brazil, as mostinfections will probably result from inadvertent consumption of thelarvae via unwashed vegetables and other produce. This is also theknown mechanism of transmission in several cases of the zoonosisin Hawaii (Hollingsworth et al., 2007).
In spite of its global spread and the potential public health impli-cations, the morphology of A. cantonensis has never been reviewed.This morphological study of this species isolated from Pernam-buco confirms a new geographical occurrence and adds taxonomiccharacters for the accurate taxonomic identification.
Both nematodes from the brain and pulmonary artery presentsimilar morphometry. However, the adults from pulmonary arteryat 120 days post infection are longer and wider than L5 and the
excretory pore is situated posterior to the junction of esophaguswhereas it is at the level of the esophagus base in L5. The mor-phology of the caudal bursa and the size of the spicules were notvariable among worms of differing ages, which permits the use ofthese parameters for species identification. The large size of thespicules [larger than those observed in any other Angiostrongylusspp (Souza et al., 2009)] and the patterns of the bursal rays agreewith the species described by Chen (1935). Shorter morphologi-cal variability between Japan and Pernambuco population may beattributed to intraspecific variation.
A. cantonensis isolated from Brazil and Japan presented guber-naculums and Alicata (1968) reported the same structure inspecimens from Hawaii. Although the gubernaculum had been rep-resented in the original figure given by Chen (1935) there is nomention of this structure in his paper.
In Brazil there have been few studies of the impact of the intro-duction of A. fulica on the natural environment as well as its role inpublic health. Control measures, health education and surveillancein vulnerable areas for A. cantonensis introduction such as shippingports are needed to prevent the further spread and outbreaks ofeosinophilic meningitis.
Acknowledgements
We are grateful to Dr. Carlos Graeff-Teixeira from Laboratoryof Molecular Parasitology and Parasite Biology, Pontifical CatholicUniversity, Rio Grande do Sul, Brazil, who lent us Angiostrongy-lus cantonensis specimens from Akita, Japan. Also, all the staff ofthe Secretaria Estadual de Saúde de Pernambuco and LACEN/PEfor their support during the field investigations in that region. Wealso want to thank Paulo Sergio Pires, Gisele Resende Espindola andAndreia Natividade da Silva for technical assistance.
References
Alicata, J.E., 1968. The Gubernaculatum of Angiosrongylus cantonensis (Chen). J. Par-asitol. 54, 1193.
Anderson, R.C., 1978. Keys to genera of the superfamily Metastrongyloidea. In:Anderson, R.C., Chabaud, A.G., Willmott, S. (Eds.), CIH Keys to the NematodesParasites of Vertebrates. Common wealth Agricultural Bureaux, Farnham Royal,Bucks, U.K., pp. 1–40.
Ash, R.L., 1970. Diagnostic morphology of the third-stage larvae of Angiostrongyluscantonensis, Angiostrongylus vasorum, Angiostrongylus abstrusus and Anafilaroidesrostratus (Nematoda: Metastrongyloidea). J. Parasitol. 56, 249–253.
Caldeira, R.L., Carvalho, O.S., Mendonca, C.L.F., Graeff-Teixeira, C., Silva, M.C.F.,Bem, R., Maurer, R., Lima, W.S., Lenzi, H.L., 2003. Molecular differentiation ofAngiostrongylus costaricensis, A. cantonensis, and A. vasorum by polymerase chainreaction-restriction fragment length polymorphism. Mem. Inst. Oswaldo Cruz98, 1039–1043.
Caldeira, R.L., Mendonca, C.L.F., Goveia, C.O., Lenzi, H.L., Graeff-Teixeira, C., Lima,W.S., Mota, E.M., Pecora, I.L., Medeiros, A.M.Z., Carvalho, O.S., 2007. First Recordof molluscs naturally infected with Angiostrongylus cantonensis (Chen, 1935)(Nematoda: Metastrongylidae) in Brazil. Mem. Inst. Oswaldo Cruz 102, 887–889.
Carvalho, O.S., Teles, H.M.S., Mota, E.M., Mendonca, C.L.G.F., Lenzi, H.L., 2003. Poten-tiality of Achatina fulica Bowdich, 1822 (Mollusca: Gastropoda) as intermediatehost of the Angiostrongylus costaricensis Morera & Céspedes 1971. Rev. Soc. Bras.Med. Trop. 36, 743–745.
Chen, H.T., 1935. Un nouveau nematode pulmonare. Pulmonema cantonensis, N. G.,N sp. des rats de Canton. Ann. Parasitol. Hum. Comp. 4, 312–317.
Gasser, R.B., Stewart, L.E., Speare, R., 1996. Genetic markers in ribosomal DNA fromhookworm identification. Acta Trop. 62, 15–21.
Graeff-Teixeira, C., Thiengo, S.C., Thomé, J.W., Medeiros, A.B., Camillo-Coura, L., Agos-tini, A.A., 1993. On the diversity of mollusc intermediate hosts of Angiostrongyluscostaricensis Morera & Céspedes, 1971 in southern Brazil. Mem. Inst. OswaldoCruz 88, 487–489.
Hollingsworth, R.G., Kaneta, R., Sullivan, J., Bishop, H.S., Qvarnstrom, Y., Silva, A.F.,Robinson, D.G., 2007. Distribution of Parmarion cf. martensi (Pulmonata: Heli-carionidae), a new semi-slug pest on Hawaii Island, and its potential as a vectorfor humam Angiostrongyliasis. Pacific Sci. 61 (4), 457–467.
Kliks, M.M., Palumbo, N.E., 1992. Eosinophilic meningitis beyond the Pacific Basin:the global dispersal of a peridomestic zoonosis caused by Angiostrongylus can-tonensis, the nematode lungworm of rats. Soc. Sci. Med. 34, 199–212.
Lv, S., Zhang, Y., Liu, H., Zhang, C., Steinmann, P., Zhou, X., Utzinger, J., 2009a.Angiostrongylus cantonensis: morphological and behavioral investigation withinthe freshwater snail Pomacea canaliculata. Parasitol. Res. 3 (2), 368.

Author's personal copy
S.C. Thiengo et al. / Acta Tropica 115 (2010) 194–199 199
Lv, S., Zang, Y., Liu, H., hu, L., Yang, K., Steinmann, P., Chen, Z., Wang, L., Utzinger,J., Zhou, X., 2009b. Invasive snails and an emerging infectious disease: resultsfrom the first national survey on Angiostrongylus cantonensis in China. PLos NTD3 (2), 1–8.
Souza, J., Simões, R., Thiengo, S.C., Lima, W., Mota, E., Rodrigues-Silva, R., Lanfredi,R.M., Maldonado, A., 2009. A new Metastrongilidae species (Nematoda): a lung-worm from Akodon montensis (Rodentia: Sigmodontinae) in Brazil. J. Parasitol.95 (6), 1507–1511.
Stevenson, L.A., Chilton, N.B., Gasser, R.B., 1995. Differentiation of Haemonchus placeifrom H. contortus (Nematoda, Trichostrongylidae) by the ribosomal DNA 2ndinternal transcribed spacer. Int. J. Parasitol. 25, 483–488.
Thiengo, S.C., 2007. Helmintoses de interesse médico-veterinário transmitidas pormoluscos no Brasil. IN: Tópicos em Malacologia - Ecos do XVIII Encontro Brasileirode Malacologia. pp. 287–294.
Thiengo, S.C., Faraco, F.A., Salgado, N.C., Cowie, R., Fernandez, M.A., 2007. Rapidspread of an invasive snail in Soth America: the giant African snail, Achatinafulica, in Brasil. Biol. Invasions 9, 693–702.
Thiengo, S.C., Fernandez, M.A., Torres, E.J.L., Coelho, P.M., Lanfredi, R.M., 2008. FirstRecord of a Metastrongyloidea Aelurostrongylus abstrusus larvae in Achatina (Lis-sachatina) fulica (Mollusca, Achatinidae) in Brazil. J. Invert. Pathol. 98, 34–39.
Tsai, H.C., Liu, Y.C., Kunin, C.M., Lee, S.S., Chen, Y.S., Lin, H.H., Tsai, T.H., Lin, W.R.,Huang, C.K., Yen, M.Y., Yen, C.M., 2001. Eosinophilic meningitis caused byAngiostrongylus cantonensis: report of 17 cases. Am. J. Med. 111, 109–114.
Wallace, G.D., Rosen, L., 1969. Studies on eosinophilic meningitis. V. Molluscan hostsof Angiostrongylus cantonensis on the Pacific Islands. Am. J. Trop. Med. Hyg. 18,206–261.
Youssif, F., Ibrahim, A., 1978. The first record of Angiostrongylus cantonensis fromEgypt. Parasitol. Res. 56, 73–80.
Related Documents











