The genesis of actively growing siliceous stromatolites: Evidence from Lake Specchio di Venere, Pantelleria Island, Italy Marianna Cangemi a , Adriana Bellanca a, ⁎, Sara Borin b , Laurence Hopkinson c , Francesca Mapelli b , Rodolfo Neri a a Dipartimento di Chimica e Fisica della Terra ed Applicazioni alle Georisorse e ai Rischi Naturali (CFTA), Università degli Studi di Palermo, Via Archirafi 36, 90123 Palermo, Italy b Department of Food Science and Microbiology, Consorzio Nazionale Interuniversitario per le Scienze del Mare, Ulr Università degli Studi di Milano, 20133 Milan, Italy c School of Environment and Technology, University of Brighton, Cockcroft Building, Lewes Road, Brighton BN2 4GJ, UK 440 abstract article info Article history: Received 8 March 2010 Received in revised form 28 June 2010 Accepted 29 June 2010 Editor: J. Fein Keywords: Stromatolites Saline lake Hydrothermal activity Silica mineralization Early diagenesis Microbial communities DNA content This study documents the attributes of siliceous stromatolites growing in the Lake Specchio di Venere, on the volcanic island of Pantelleria, Italy, in a setting characterized by very shallow cold waters and pools and by scattered hydrothermal activity, which exhales mainly CO 2 , at emission point temperatures of 34 to 58 °C. The saturation indexes indicate that the lake waters are saturated with respect to tridymite, cristobalite, chalcedony and quartz, and slightly undersaturated with respect to amorphous silica. Common roughly laminated and poorly lithified stromatolites show scanning electron microscope (SEM) evidence for silicified microbial mat structures, including biofilms, filamentous and coccoid cells, and extracellular polymeric substances (EPS). The screening of bacterial 16S rRNA libraries indicates that autotrophic and heterotrophic bacterial communities colonize surface and core levels of the stromatolites. Locally the stromatolites show a granular non-porous fabric, where filaments and silica sheets are not apparent. Inhomogeneity in the stromatolite fabric corresponds with varying DNA content and different structures of the colonizing bacterial communities, non-porous stromatolite levels being microbially colonized to a lesser extent. Based on Fourier Transform (FT)–Raman and FT-infrared investigations, the laminated stromatolite contains early diagenetic tridymite in addition to amorphous silica (opal-A), whereas the non-porous stromatolite shows an essentially amorphous character. The laminated stromatolite is thought to form at relatively low rates of silica precipitation and with a possible microbial mediation in terms of microbial cells and their EPS accelerating the onset of amorphous silica nucleation. It is suggested that high porosity probably favoured a consistent flux of silica-rich fluids that triggered the opal-A to tridymite transformation, thus strengthening the preservation of biosignals. Non-porous stromatolite growth could reflect temporal or localised changes in environmental conditions that caused variations in the degree of silica saturation in the lake waters inducing abiotic silica accumulation. Accelerated opal-A deposition could have obscured primary filamentous fabrics and limited the flux of pore fluid needed for sustaining the process of silica maturation. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Stromatolites (i.e. laminated microbialites) are organosedimen- tary deposits produced by the mediation of growth and activity of microorganisms. Cycling microbial communities facilitate mineral precipitation, trap and cement small grains with extracellular organic material, and promote lithification (Kalkowsky, 1908; Awramik, 1984; Burne and Moore, 1987; Reid et al., 2000; Tucker, 2001; Baumgartner et al., 2009; Budakoglu, 2009). They were common in the Precambrian and represent some of the earliest macroscopic evidence of life in the fossil record (Schopf, 1993), potentially containing important paleoenvironmental and biological information (Andres and Reid, 2006). Living, modern stromatolites are considered their contemporary analogues based on morphology and hence their study has the potential to elucidate the mechanisms of colonization of the early Earth. The most widespread modern and fossil stromatolites display carbonate compositions, and numerous studies have recognized the biogenically mediated origin of specific mineralized fabrics, textures, and mineral assemblages (Reid et al., 2000; Riding, 2000; Webb and Kamber, 2000; Van Lith et al., 2003, Allwood et al., 2006; Olivier and Boyet, 2006; Cavallazzi et al., 2007). Siliceous stromatolites have been reported from many active geothermal systems (Campbell et al., 2002; Rodgers et al., 2004, Handley et al., 2005; Jones et al., 2005; Handley et al., 2008). In these settings microbial activity can result in microbial fossilization, chemical sediment formation, and silica transport (Yee et al., 2003; Benning et al., 2004a, b; Konhauser et al., 2004; Handley et al., 2008), although the precise role of Chemical Geology 276 (2010) 318–330 ⁎ Corresponding author. Tel.: +39 091 23861637; fax: +39 091 6168376. E-mail address: [email protected] (A. Bellanca). 0009-2541/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.chemgeo.2010.06.017 Contents lists available at ScienceDirect Chemical Geology journal homepage: www.elsevier.com/locate/chemgeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chemical Geology 276 (2010) 318–330
Contents lists available at ScienceDirect
Chemical Geology
j ourna l homepage: www.e lsev ie r.com/ locate /chemgeo
The genesis of actively growing siliceous stromatolites: Evidence from Lake Specchiodi Venere, Pantelleria Island, Italy
Marianna Cangemi a, Adriana Bellanca a,⁎, Sara Borin b, Laurence Hopkinson c,Francesca Mapelli b, Rodolfo Neri a
a Dipartimento di Chimica e Fisica della Terra ed Applicazioni alle Georisorse e ai Rischi Naturali (CFTA), Università degli Studi di Palermo, Via Archirafi 36, 90123 Palermo, Italyb Department of Food Science and Microbiology, Consorzio Nazionale Interuniversitario per le Scienze del Mare, Ulr Università degli Studi di Milano, 20133 Milan, Italyc School of Environment and Technology, University of Brighton, Cockcroft Building, Lewes Road, Brighton BN2 4GJ, UK 440
⁎ Corresponding author. Tel.: +39 091 23861637; faxE-mail address: [email protected] (A. Bellanca).
0009-2541/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.chemgeo.2010.06.017
a b s t r a c t
a r t i c l e i n f oArticle history:Received 8 March 2010Received in revised form 28 June 2010Accepted 29 June 2010
Editor: J. Fein
Keywords:StromatolitesSaline lakeHydrothermal activitySilica mineralizationEarly diagenesisMicrobial communitiesDNA content
This study documents the attributes of siliceous stromatolites growing in the Lake Specchio di Venere, on thevolcanic island of Pantelleria, Italy, in a setting characterized by very shallow cold waters and pools and byscattered hydrothermal activity, which exhales mainly CO2, at emission point temperatures of 34 to 58 °C.The saturation indexes indicate that the lake waters are saturated with respect to tridymite, cristobalite,chalcedony and quartz, and slightly undersaturated with respect to amorphous silica. Common roughlylaminated and poorly lithified stromatolites show scanning electron microscope (SEM) evidence for silicifiedmicrobial mat structures, including biofilms, filamentous and coccoid cells, and extracellular polymericsubstances (EPS). The screening of bacterial 16S rRNA libraries indicates that autotrophic and heterotrophicbacterial communities colonize surface and core levels of the stromatolites. Locally the stromatolites show agranular non-porous fabric, where filaments and silica sheets are not apparent. Inhomogeneity in thestromatolite fabric corresponds with varying DNA content and different structures of the colonizing bacterialcommunities, non-porous stromatolite levels being microbially colonized to a lesser extent. Based on FourierTransform (FT)–Raman and FT-infrared investigations, the laminated stromatolite contains early diagenetictridymite in addition to amorphous silica (opal-A), whereas the non-porous stromatolite shows anessentially amorphous character. The laminated stromatolite is thought to form at relatively low rates ofsilica precipitation and with a possible microbial mediation in terms of microbial cells and their EPSaccelerating the onset of amorphous silica nucleation. It is suggested that high porosity probably favoured aconsistent flux of silica-rich fluids that triggered the opal-A to tridymite transformation, thus strengtheningthe preservation of biosignals. Non-porous stromatolite growth could reflect temporal or localised changes inenvironmental conditions that caused variations in the degree of silica saturation in the lake waters inducingabiotic silica accumulation. Accelerated opal-A deposition could have obscured primary filamentous fabricsand limited the flux of pore fluid needed for sustaining the process of silica maturation.
: +39 091 6168376.
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Stromatolites (i.e. laminated microbialites) are organosedimen-tary deposits produced by the mediation of growth and activity ofmicroorganisms. Cycling microbial communities facilitate mineralprecipitation, trap and cement small grains with extracellular organicmaterial, and promote lithification (Kalkowsky, 1908; Awramik,1984; Burne and Moore, 1987; Reid et al., 2000; Tucker, 2001;Baumgartner et al., 2009; Budakoglu, 2009). They were common inthe Precambrian and represent some of the earliest macroscopicevidence of life in the fossil record (Schopf, 1993), potentiallycontaining important paleoenvironmental and biological information
(Andres and Reid, 2006). Living, modern stromatolites are consideredtheir contemporary analogues based on morphology and hence theirstudy has the potential to elucidate the mechanisms of colonization ofthe early Earth.
The most widespread modern and fossil stromatolites displaycarbonate compositions, and numerous studies have recognized thebiogenically mediated origin of specific mineralized fabrics, textures,and mineral assemblages (Reid et al., 2000; Riding, 2000; Webb andKamber, 2000; Van Lith et al., 2003, Allwood et al., 2006; Olivier andBoyet, 2006; Cavallazzi et al., 2007). Siliceous stromatolites have beenreported from many active geothermal systems (Campbell et al.,2002; Rodgers et al., 2004, Handley et al., 2005; Jones et al., 2005;Handley et al., 2008). In these settings microbial activity can result inmicrobial fossilization, chemical sediment formation, and silicatransport (Yee et al., 2003; Benning et al., 2004a, b; Konhauser etal., 2004; Handley et al., 2008), although the precise role of

319M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
microorganisms in silica precipitation and its diagenetic maturation isincompletely known. Mechanisms invoked have included processesacting in modern hot spring settings at moderate to low temperatures(less than 73 °C), where microbial activity is common (Handley et al.,2005). Even if there is no evidence to suggest that prokaryotes activelyprecipitate silica (Walters et al., 1977; Mountain et al., 2003;Konhauser et al., 2004), functional groups on their cell walls andwithin extracellular polymeric substances (EPS) are thought toprovide reactive sites for biologically passive silica depositionfollowed by autocatalytic silica polymerization (Krumbein andWerner, 1983; Urrutia and Beveridge, 1993; Westall et al., 1995;Konhauser and Ferris, 1996; Farmer, 1999; Phoenix et al., 2000;Benning et al., 2004a, b). Silica accumulation has been demonstratedto be proportional to the availability of electrostatically favourablesubstrates (Rimstidt and Barnes, 1980; Fleming, 1986) provided by afilamentous network (Nicholson and Aquino, 1989; Handley et al.,2005). In contrast, possible abiotic mechanisms refer to silicaprecipitation induced by (1) rapid cooling to ambient temperatures,(2) changes in pH, and (3) increased silica oversaturation andpolymerization caused by combined cooling and evaporation or bywind-induced waves providing subaerial transport of silica chargedwaters (White et al., 1956; Iler, 1973; Brown and McDowell, 1983;Mroczek and Reeves, 1994; Jones et al., 1997; Renaut et al., 1998;Mountain et al., 2003; Handley et al., 2005). Whether microbialsurfaces play a crucial role in silica fixation or have a negligible effecton silica nucleation is still controversial. There is however agreementon the need to further investigate mechanisms that control silica–microbe interactions (e.g., Yee et al., 2003).
Actively growing siliceous stromatolites have been recently foundin the Lake Specchio di Venere, on the volcanic island of Pantelleria,Italy (Fig. 1), in a setting characterized by very shallow cold watersand pools and by scattered hydrothermal activity. Owing to theirextensive deposition, variable morphology, association with a salinelake ecosystem the stromatolites have the potential to provide new
Fig. 1. General geological setting of Pantelleria Island, Italy (simplified f
insight into the processes involved in themicrobial fossilization and toenhance recognition of geobiologically relevant imprints in ancientrocks. In this paper, we use an integrated approach combiningpetrographic, geochemical, mineralogical andmolecular microbiologystudies with the aim of contributing to the understanding of biotic andabiotic factors controlling the initial growth and the early diagenetictransformation of the silica stromatolites.
2. Geological setting and lake features
The island of Pantelleria is a quiescent strato-volcano located inthe Strait of Sicily, about 100 km SW of Sicily and 70 km NE of Tunisia,on the axis of the Sicily Channel Rift Zone (Fig. 1). The island is entirelycovered by volcanic products from both effusive and explosiveactivity, with dominant peralkaline rhyolites (“pantellerites”) andtrachytes and minor alkali basalts (Civetta et al., 1984). At present,volcanic activity is limited to low temperature fumarolic emissionsand thermal springs reaching temperature up to 90 °C (Parello et al.,2000; Favara et al., 2001; Aiuppa et al., 2007). Many of thesehydrothermal rises occur in the north-east island sector, particularlyin the area of Lake Specchio di Venere. This is an endorheic lake locatedinside a calderic depression (Caldera Cinque Denti, Fig. 1) which has asub-circular shape, being ca 450 m long and ca 350 m wide, and amaximum depth of 12.5 m (Bocchi et al., 1988) with steeper slopes inits north-eastern zone (Fig. 2A). The surface area is strongly controlledby the rainfall rate, varying from 136.000 to ca 200.000 m2 (Aiuppaet al., 2007). In addition to runoff, the lake receives a contributionfrom the thermal aquifer and loses the water surplus through seepageto groundwater (Aiuppa et al., 2007). Hydrothermal activity, exhalingmainly CO2 (98%) together with low percentages of N2, O2, Ar, CH4, H2
and He (D'Alessandro et al., 1994; Parello et al., 2000; Aiuppa et al.,2007) affects the south-west lake shoreline in proximity to theintersection of the underground water table with the topographicsurface. At the emission points, temperatures are between 34 and
rom Rotolo et al., 2007) showing the location of the sampling area.

320 M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
58 °C. The water depth and the shoreline undergo seasonal adjust-ments as a function of the yearly hydrological balance. The lakewatershave a quite homogeneous pH (ca 9) and temperatures, measured atthe lake surface near the stromatolites, ranging between ca 25 °C(May 2008) and 14.50 °C (December 2008). Based on Eh valuesmeasured on the same area, the lake waters are strongly oxidizing.
The siliceous stromatolites grow in shallow marginal waters nearthe southwestern lake shoreline (Fig. 2A, red arrow) and, althoughshowing diverse fabrics, they generally consist of roughly laminated,poorly lithified layers defined by the presence of abundant fenestralpores and varying in colour from orange-red (top) to green (middlepart) to reddish-brown (bottom) in relation to the presence ofdifferent benthic ecosystems. Locally, laminar structures are notevident and the deposit presents intercalated sinter texturedhorizons.
The stromatolites form a smooth slightly indented bank (Fig. 2B)which ranges in thickness from ca 2 up to ca 30 cm gently slopingdown into the lake waters in a series of terraces to a depth of 2 m,beyond which point the lake bottom slope increases abruptly. Thesubmerged stromatolites appear as large domes or columns rising ca8 cm from the lake bottom (Fig. 2C). Exposed stromatolites constitutea platform-like deposit around the lake margin, in places formingledges that extend from the shoreline. In this case, the stromatolitesare capped by carbonate material mainly constituted by aragonite andhydromagnesite (Cangemi, 2009). In the intertidal zone, where thestromatolite bank is at its thinnest, domal structures are aligned alongthe shoreline (Fig. 2D). In summer, when the lake level is shallower,these structures are subjected to intense drying forming mud crack-like structures due to desiccation of the domes.
3. Materials and methods
The sampling sites extend along a south-west transect parallel tothe lake margin, expanding close to the rising of CO2 fromhydrothermal vents (Fig. 2A). In May and December 2008, in situwater electrolytic conductivity, temperature, Eh and pH measure-ments were obtained, water samples were collected for analysis, andthe stromatolites were sampled. Stromatolite samples were picked upfrom the bank and immediately stored in airtight plastic bags uponcollection. A representative sample of the submerged stromatoliteshas been subsampled at centimetre scale and labelled A to G from topto bottom (Fig. 2E). Levels A, B, E, F and G are roughly laminatedwhereas levels C and D appear as sinter textured horizons. Samples tobe analysed for microbial community were isolated from the surfaceof the stromatolites as described below.
Petrographic features were investigated by a polarizing micro-scope, using epoxy-impregnated samples. In addition, small fracturedsamples were mounted on stubs, coated with a layer of graphite andobserved using a Scanning Electron Microscope (SEM) LEO 440 withEDS system OXFORD ISIS Link and Si (Li) PENTAFET detector. Bulksample mineralogy was determined by powder X-ray diffraction(XRD) using a Philips PW14 1373 with Cu-Kα radiation filtered by amonochromator crystal and a scanning speed of 2°2θ/min. To furtherelucidate the nature of poorly crystalline and semi-amorphous silicaphases Fourier Transform Infrared Spectrometry (FTIR) and FourierTransform–Raman (FT–Raman) were applied. Raman measurementswere performed at room temperature with a Bruker RAMII FourierTransform Raman spectrometer, employing a 500 mW Nd:YAG laser(1064 nm) as light source and having 5 cm−1 spectral resolution. Thespectral accumulations is 500, no averaging, spectral handling andcorrection. The ATR-FTIR measurements were performed on a Tensor27 spectrometer (Bruker Optics). The spectrometer was completelysoft-controlled by the OPUS software version 6.0 provided by BrukerOptics. The spectrum of each sample was obtained by taking theaverage of 32 scans at a resolution of 4 cm−1. It was acquired between
4000 and 400 cm−1, with scanner velocity of 10 kHz and abackground of 32 scans.
Water samples used for determination of dissolved major andtrace elements were collected in either LD-PE (low-density polyeth-ylene, for major elements) or HD-PE (high-density polyethylene, fortrace elements) bottles, which were pre-washed with ultrapureHNO3. Waters were stored after filtration, using 0.45 μm Milliporefilter, and acidification to pH ca 2 with ultrapure concentrated HNO3.Unacidified samples were stored for analysis of anions. Aliquots, notfiltered and not acidified, were used for alkalinity titration with HCl(0.1 N). The pH was measured with an 8102BN Ross combination pHelectrode. Na, K, Mg and Ca cations, Cl and SO4 anions weredetermined by ionic chromatography (Dionex model DX-120) withNaHCO3/Na2CO3 eluant for the anionic column CS12A. Ionic balancewas computed for each sample taking into account major species; allsamples exhibited imbalance lower than 5%. Silica was determined byICP-OES, Horiba Jobin Yvon equipment, model ULTIMA 2. Theanalytical precision measured as relative standard deviation was 2%.Field and laboratory physico-chemical data were elaborated by usingthe geochemical code PHREEQC (version 2.15.0; Parkhurst andAppelo, 1999) and the Lawrence Livermore National Laboratory(LLNL) database in order to calculate saturation indexes (SI) of severalmineralogical phases for lake water solutions, according to theequation: SI=log IAP− log K, where IAP is the ionic activity productof the specific reaction and K is the equilibrium constant of dissolutionreaction.
To perform microbial community analysis, submerged stromato-lites were dissected with sterile scalpels on the basis of the visiblelamination, collecting levels of different colour and texture. Sampleswere frozen within a maximum of 4h from collection and stored at−20 °C in sterile containers until analysis. Surficial water wascollected from the lake and immediately filtered to collect the cellson sterile mixed cellulose esters Sterivex filters having 0.22 μm poresize (Millipore, Milan, Italy). The filters were stored at −20 °C untilanalysis. In the laboratory, total DNA was extracted using acommercial kit and following the instructions of the manufacturer(FastDNA™ SPIN Kit for Soil, BIO 101 Systems Q-BIO gene; CA, USA).Total DNA was extracted from water using an established protocol(Murray et al. 1998). Extracted DNA was quantified using QubitFluorometer and Quant-iT dsDNA HS assay kit following themanufacturer instructions (Stratagene, TX, USA). 16S rRNA genewas amplified from the extracted DNA and 16S rRNA gene librarieswere built using universal bacterial PCR primers as previouslydescribed (Borin et al., 2009). Forty-five clones were partiallysequenced from each library and the obtained 800 bp long sequenceshave been deposited in GeneBank under accession numbersFN687011–FN687143. Sequences were checked using the CHECK_-CHIMERA program to determine the presence of hybrid sequences(Maidak et al., 2001). Phylogenetic affiliations were preliminarilyobtained using BLASTN (Altschul et al., 1997) and confirmed with thenaïve Bayesian rRNA Classifier provided by the Ribosomal DatabaseProject website (Maidak et al. 2001). Bacterial diversity was analysedby Amplified Ribosomal Intergenic Spacer Analysis (ARISA) finger-printing performed in triplicate on the total DNA of each fraction withthe protocol set up by Cardinale et al. (2004). Downstreammultivariate statistical analysis of the obtained fingerprints wasperformed by the application of Detrended Correspondence Analysis(DCA) using the software Multi Variate Statistic Package, version3.13n (Kovach Computing Services, UK).
4. Results
4.1. Stromatolite imaging by light microscopy and SEM
In thin section, siliceous stromatolites predominantly consist of adense semiopaque microcrystalline silica matrix (micrite-like) with

Fig. 2. Field photographs. (A) South-west panoramic view of Lake Specchio di Venere showing the siliceous stromatolite zone (red arrow); on the background, the Mediterranean Sea.(B) Submerged and emerged stromatolites (whitish bank) located close to CO2 vents along the southwestern lake shore (grey bar on the right=30 cm). (C) Underwaterphotography showing a group of domal and columnar stromatolites covered by microbial mat (yellow bar, centre to top on the right,=5.5 cm). To note gas bubbles emerging fromthe light green microbial mat. (D) Alignment of domal forms in the intertidal zone. (E) Siliceous stromatolite sample from the southwestern lake shore. Subsampled levels, ca 1 cmthick, are identified by black dashed lines and letters (A to G, top to bottom).
321M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
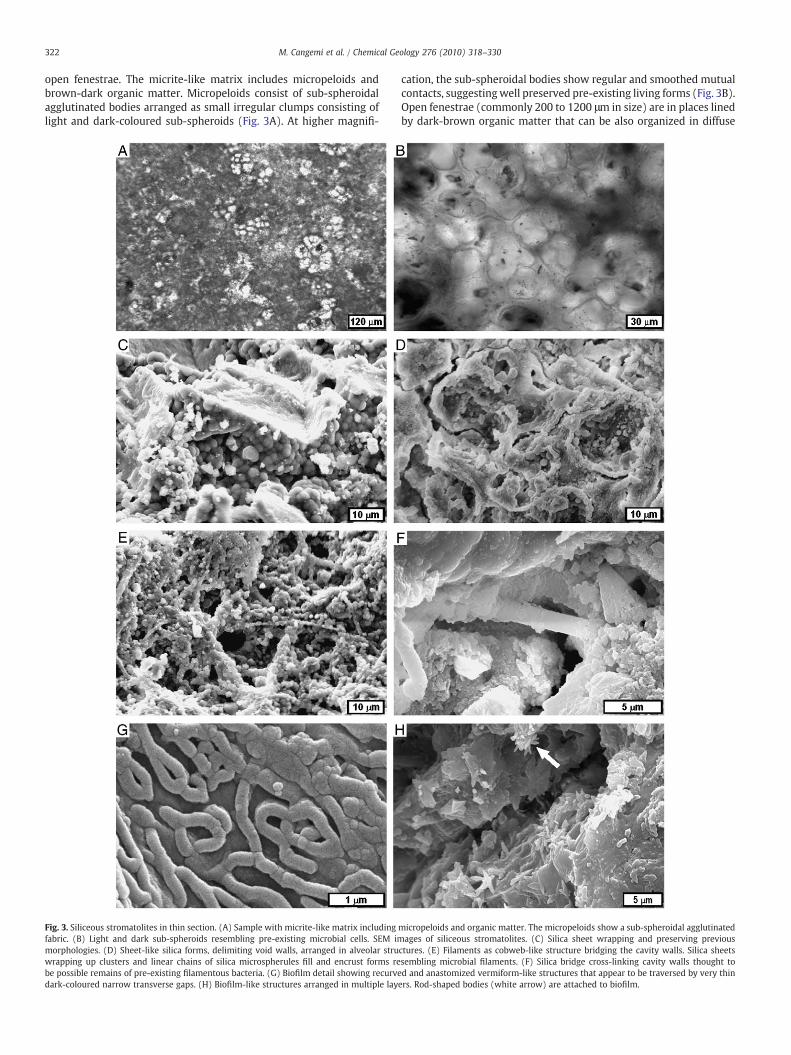
322 M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
open fenestrae. The micrite-like matrix includes micropeloids andbrown-dark organic matter. Micropeloids consist of sub-spheroidalagglutinated bodies arranged as small irregular clumps consisting oflight and dark-coloured sub-spheroids (Fig. 3A). At higher magnifi-
Fig. 3. Siliceous stromatolites in thin section. (A) Sample with micrite-like matrix includingfabric. (B) Light and dark sub-spheroids resembling pre-existing microbial cells. SEM immorphologies. (D) Sheet-like silica forms, delimiting void walls, arranged in alveolar struwrapping up clusters and linear chains of silica microspherules fill and encrust forms rebe possible remains of pre-existing filamentous bacteria. (G) Biofilm detail showing recurvedark-coloured narrow transverse gaps. (H) Biofilm-like structures arranged in multiple lay
cation, the sub-spheroidal bodies show regular and smoothed mutualcontacts, suggestingwell preserved pre-existing living forms (Fig. 3B).Open fenestrae (commonly 200 to 1200 μm in size) are in places linedby dark-brown organic matter that can be also organized in diffuse
micropeloids and organic matter. The micropeloids show a sub-spheroidal agglutinatedages of siliceous stromatolites. (C) Silica sheet wrapping and preserving previous
ctures. (E) Filaments as cobweb-like structure bridging the cavity walls. Silica sheetssembling microbial filaments. (F) Silica bridge cross-linking cavity walls thought tod and anastomized vermiform-like structures that appear to be traversed by very thiners. Rod-shaped bodies (white arrow) are attached to biofilm.
323M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
brown irregular clumps. Bioaccumulation of undeterminated speciesof diatoms is locally observed.
Under the SEM, samples from the roughly laminated stromatolitelevels (A, B, E, F and G in Fig. 2E) and other similar stromatolitesamples are characterized by the presence of silica sheets, wrappingand preserving previous morphologies, that act as an initial templatefor the stromatolite growth (Fig. 3C). These sheet-like forms locallycover void walls and are arranged in alveolar structures (Fig. 3D). Thestromatolite contains filaments arranged in cobweb-like structuresbridging the cavity walls (Fig. 3E) and locally showing tubular shapes.Moreover, tubular organic structures, putatively ascribed to filamen-tous microorganisms, occur inside the voids (Fig. 3F). Gently foldedbiofilm-like structures line the cavities. An enlarged view of thebiofilm (Fig. 3G) shows anastomized vermicular structures (about0.5 μm in diameter) dispersed and encapsulated in a homogeneousblanket covering free surfaces. Silica microspherules (up to 5 μm indiameter) are deposited within filaments or are accumulated uponthe filamentous mat (Fig. 3E). Microspherule coalescence generatesclusters or chains encrusting the filaments, according to their three-dimensional arrangement (Fig. 3E).
The sinter textured horizons (C and D in Fig. 2E) are characterizedby the presence of biofilm-like structures arranged in multiple layers(Fig. 3H). Locally, micron-sized rod-like bodies are attached to orextend from the biofilm (Fig. 3H). Unlike the other stromatolite levelsexamined, filaments and silica sheets are not apparent in samples Cand D.
4.2. XRD and spectroscopic analysis
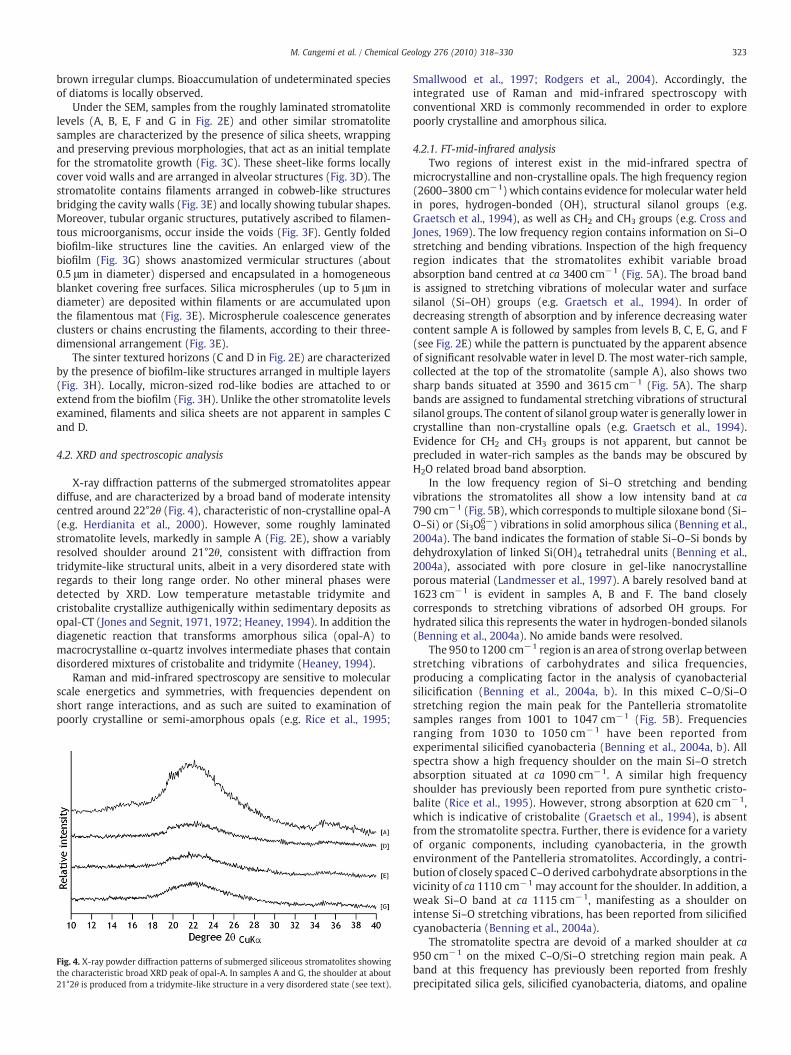
X-ray diffraction patterns of the submerged stromatolites appeardiffuse, and are characterized by a broad band of moderate intensitycentred around 22°2θ (Fig. 4), characteristic of non-crystalline opal-A(e.g. Herdianita et al., 2000). However, some roughly laminatedstromatolite levels, markedly in sample A (Fig. 2E), show a variablyresolved shoulder around 21°2θ, consistent with diffraction fromtridymite-like structural units, albeit in a very disordered state withregards to their long range order. No other mineral phases weredetected by XRD. Low temperature metastable tridymite andcristobalite crystallize authigenically within sedimentary deposits asopal-CT (Jones and Segnit, 1971, 1972; Heaney, 1994). In addition thediagenetic reaction that transforms amorphous silica (opal-A) tomacrocrystalline α-quartz involves intermediate phases that containdisordered mixtures of cristobalite and tridymite (Heaney, 1994).
Raman and mid-infrared spectroscopy are sensitive to molecularscale energetics and symmetries, with frequencies dependent onshort range interactions, and as such are suited to examination ofpoorly crystalline or semi-amorphous opals (e.g. Rice et al., 1995;
Fig. 4. X-ray powder diffraction patterns of submerged siliceous stromatolites showingthe characteristic broad XRD peak of opal-A. In samples A and G, the shoulder at about21°2θ is produced from a tridymite-like structure in a very disordered state (see text).
Smallwood et al., 1997; Rodgers et al., 2004). Accordingly, theintegrated use of Raman and mid-infrared spectroscopy withconventional XRD is commonly recommended in order to explorepoorly crystalline and amorphous silica.
4.2.1. FT-mid-infrared analysisTwo regions of interest exist in the mid-infrared spectra of
microcrystalline and non-crystalline opals. The high frequency region(2600–3800 cm−1) which contains evidence formolecular water heldin pores, hydrogen-bonded (OH), structural silanol groups (e.g.Graetsch et al., 1994), as well as CH2 and CH3 groups (e.g. Cross andJones, 1969). The low frequency region contains information on Si–Ostretching and bending vibrations. Inspection of the high frequencyregion indicates that the stromatolites exhibit variable broadabsorption band centred at ca 3400 cm−1 (Fig. 5A). The broad bandis assigned to stretching vibrations of molecular water and surfacesilanol (Si–OH) groups (e.g. Graetsch et al., 1994). In order ofdecreasing strength of absorption and by inference decreasing watercontent sample A is followed by samples from levels B, C, E, G, and F(see Fig. 2E) while the pattern is punctuated by the apparent absenceof significant resolvable water in level D. The most water-rich sample,collected at the top of the stromatolite (sample A), also shows twosharp bands situated at 3590 and 3615 cm−1 (Fig. 5A). The sharpbands are assigned to fundamental stretching vibrations of structuralsilanol groups. The content of silanol groupwater is generally lower incrystalline than non-crystalline opals (e.g. Graetsch et al., 1994).Evidence for CH2 and CH3 groups is not apparent, but cannot beprecluded in water-rich samples as the bands may be obscured byH2O related broad band absorption.
In the low frequency region of Si–O stretching and bendingvibrations the stromatolites all show a low intensity band at ca790 cm−1 (Fig. 5B), which corresponds to multiple siloxane bond (Si–O–Si) or (Si3O9
6−) vibrations in solid amorphous silica (Benning et al.,2004a). The band indicates the formation of stable Si–O–Si bonds bydehydroxylation of linked Si(OH)4 tetrahedral units (Benning et al.,2004a), associated with pore closure in gel-like nanocrystallineporous material (Landmesser et al., 1997). A barely resolved band at1623 cm−1 is evident in samples A, B and F. The band closelycorresponds to stretching vibrations of adsorbed OH groups. Forhydrated silica this represents the water in hydrogen-bonded silanols(Benning et al., 2004a). No amide bands were resolved.
The 950 to 1200 cm−1 region is an area of strong overlap betweenstretching vibrations of carbohydrates and silica frequencies,producing a complicating factor in the analysis of cyanobacterialsilicification (Benning et al., 2004a, b). In this mixed C–O/Si–Ostretching region the main peak for the Pantelleria stromatolitesamples ranges from 1001 to 1047 cm−1 (Fig. 5B). Frequenciesranging from 1030 to 1050 cm− 1 have been reported fromexperimental silicified cyanobacteria (Benning et al., 2004a, b). Allspectra show a high frequency shoulder on the main Si–O stretchabsorption situated at ca 1090 cm−1. A similar high frequencyshoulder has previously been reported from pure synthetic cristo-balite (Rice et al., 1995). However, strong absorption at 620 cm−1,which is indicative of cristobalite (Graetsch et al., 1994), is absentfrom the stromatolite spectra. Further, there is evidence for a varietyof organic components, including cyanobacteria, in the growthenvironment of the Pantelleria stromatolites. Accordingly, a contri-bution of closely spaced C–O derived carbohydrate absorptions in thevicinity of ca 1110 cm−1 may account for the shoulder. In addition, aweak Si–O band at ca 1115 cm−1, manifesting as a shoulder onintense Si–O stretching vibrations, has been reported from silicifiedcyanobacteria (Benning et al., 2004a).
The stromatolite spectra are devoid of a marked shoulder at ca950 cm−1 on the mixed C–O/Si–O stretching region main peak. Aband at this frequency has previously been reported from freshlyprecipitated silica gels, silicified cyanobacteria, diatoms, and opaline
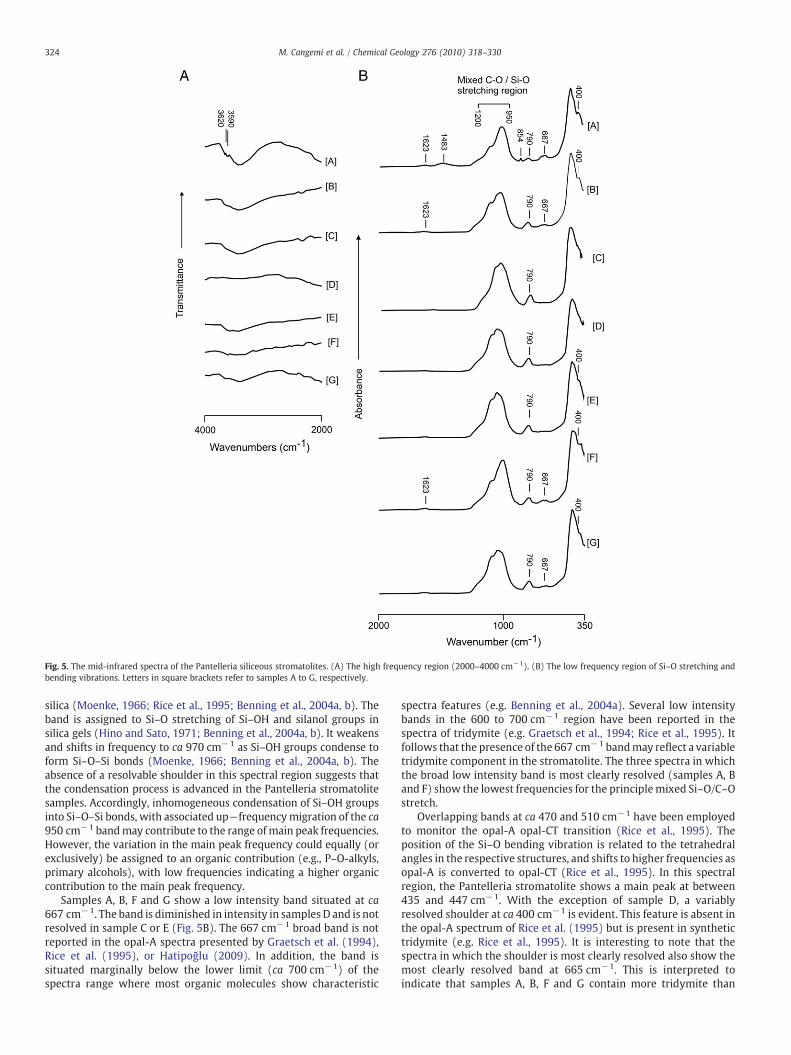
Fig. 5. The mid-infrared spectra of the Pantelleria siliceous stromatolites. (A) The high frequency region (2000–4000 cm−1). (B) The low frequency region of Si–O stretching andbending vibrations. Letters in square brackets refer to samples A to G, respectively.
324 M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
silica (Moenke, 1966; Rice et al., 1995; Benning et al., 2004a, b). Theband is assigned to Si–O stretching of Si–OH and silanol groups insilica gels (Hino and Sato, 1971; Benning et al., 2004a, b). It weakensand shifts in frequency to ca 970 cm−1 as Si–OH groups condense toform Si–O–Si bonds (Moenke, 1966; Benning et al., 2004a, b). Theabsence of a resolvable shoulder in this spectral region suggests thatthe condensation process is advanced in the Pantelleria stromatolitesamples. Accordingly, inhomogeneous condensation of Si–OH groupsinto Si–O–Si bonds, with associated up−frequencymigration of the ca950 cm−1 bandmay contribute to the range of main peak frequencies.However, the variation in the main peak frequency could equally (orexclusively) be assigned to an organic contribution (e.g., P–O-alkyls,primary alcohols), with low frequencies indicating a higher organiccontribution to the main peak frequency.
Samples A, B, F and G show a low intensity band situated at ca667 cm−1. The band is diminished in intensity in samples D and is notresolved in sample C or E (Fig. 5B). The 667 cm−1 broad band is notreported in the opal-A spectra presented by Graetsch et al. (1994),Rice et al. (1995), or Hatipoğlu (2009). In addition, the band issituated marginally below the lower limit (ca 700 cm−1) of thespectra range where most organic molecules show characteristic
spectra features (e.g. Benning et al., 2004a). Several low intensitybands in the 600 to 700 cm−1 region have been reported in thespectra of tridymite (e.g. Graetsch et al., 1994; Rice et al., 1995). Itfollows that the presence of the 667 cm−1 bandmay reflect a variabletridymite component in the stromatolite. The three spectra in whichthe broad low intensity band is most clearly resolved (samples A, Band F) show the lowest frequencies for the principlemixed Si–O/C–Ostretch.
Overlapping bands at ca 470 and 510 cm−1 have been employedto monitor the opal-A opal-CT transition (Rice et al., 1995). Theposition of the Si–O bending vibration is related to the tetrahedralangles in the respective structures, and shifts to higher frequencies asopal-A is converted to opal-CT (Rice et al., 1995). In this spectralregion, the Pantelleria stromatolite shows a main peak at between435 and 447 cm−1. With the exception of sample D, a variablyresolved shoulder at ca 400 cm−1 is evident. This feature is absent inthe opal-A spectrum of Rice et al. (1995) but is present in synthetictridymite (e.g. Rice et al., 1995). It is interesting to note that thespectra in which the shoulder is most clearly resolved also show themost clearly resolved band at 665 cm−1. This is interpreted toindicate that samples A, B, F and G contain more tridymite than

325M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
samples C, D and E. Sample A contains two low intensity bands (854and 1483 cm−1) which cannot be assigned to crystalline or non-crystalline opal. The bands are however broadly coincident with thev2 and v3 internal modes of aragonite (White, 1974). XRD patterns donot provide evidence for the occurrence of aragonite; however, theconcentration of Sr, that is a major impurity component in aragonite(Sunagawa et al., 2007), is low (40 mg kg−1) in this sample butslightly higher than in other stromatolite samples (ca 15 mg kg−1;Cangemi, 2009). On this basis, the presence of small amounts ofaragonite cannot be ruled out.
4.2.2. FT–Raman spectroscopic analysisIn commonwith sedimentary opals, the stromatolite shows a variety
of broad low intensity bands (Fig. 6), consistent with a generallywidespread amorphous character (Smallwood et al., 1997), with strongsignals at ca 460 to 490 cm−1, typical of amorphous silica (Ilieva et al.,2007). Samples G and F, and to a lesser extent sample A show lowintensity doublets or multiplets in the region 200 to 500 cm−1,including a clearly resolved broad band situated at 335 cm−1.Raman scattering in the spectra at ca 350 cm−1 is indicative oftridymite-like structural units (Ilieva et al., 2007). Samples G and F,
Fig. 6. The FT–Raman spectra of the Pantelleria siliceous stromatolites. Band assign-ments are compiled from various sources (Kingma and Hemley, 1994; Smallwood et al.,1997; Ilieva et al., 2007). Letters in square brackets refer to samples A to G, respectively.
and to a lesser extent sample A also show ill-defined low intensity bandsat ca 460 and 400 cm−1. Several peaks in the spectra of α-tridymite(assigned to bending modes of Si–O–Si units) occur in this region, themost prominent of which are at 403, 422, 449 and 457 cm−1 (e.g. Ilievaet al., 2007). Accordingly, spectroscopic data is consistent with shortrange tridymite ordering in these samples. FT–Raman bands indicativeof tridymite ordering approach background or else are absent forsamples E to B.
Samples A, B, F and G (and possibly E and D) show a broad lowintensity band situated between 1070 and 1080 cm−1. This band isassigned to Si–O–Si asymmetric stretch and is observed in the spectra ofpure α-tridymite and α-cristobalite (Kingma and Hemley, 1994;Smallwood et al., 1997). The band also broadly coincides with the v1internalmodeof aragonite (e.g.,White, 1974). The stromatolite samplesshowabroadasymmetric peak, in the750 to850 cm−1 region (Fig. 6). Aband at 786 cm−1 has previously been described from a range of opals,and is assigned to Si–O–Si symmetric stretch for three and fourmemberrings (Smallwood et al., 1997). The phase α-tridymite shows a doubletat 785 cm−1 and a weaker band at 820 cm−1, whereas α-cristobaliteshows a single peak at 785 cm−1. Accordingly, the asymmetry of the786 cm−1 band suggests that tridymite short range ordering isabundant relative to, or to the exclusion of a cristobalite component.Furthermore cristobalite-typearrangements are characterizedbystrongsignals at 418 and 231 cm−1 (Kingma and Hemley, 1994; Ilieva et al.,2007), neither of which are evident in the stromatolite spectra.
The 786 cm−1 band is poorly resolved in samples C and D; the twosamples are also devoid ofmultiple broad bands in the 200 to 400 cm−1
region (Fig. 6), suggesting that the samples are essentially non-crystalline. The spectra show to varying extents weak broad bandscentred at ca 970 cm−1 and ca 1380 cm−1. Bands situated at similarfrequencies have been reported from natural opals, but not from purecrystalline or amorphous silica (Smallwood et al., 1997) suggesting thatthey reflect the presence of impurities. For example bands centred at ca960 cm−1 in natural opals have been assigned to metal ion silicates(McMillan and Wolf, 1995; Smallwood et al., 1997).
The stromatolites spectra show a band at ca 680 cm−1. Theintensity of the band is greatest in samples A and G. The band is notpresent in the opaline spectra reported by Ilieva et al. (2007).However, the band closely coincides with several low intensityinfrared active bands reported for opal-CT (e.g. Rice et al., 1995), andthe broad low intensity band measured at ca 667 cm−1 in somestromatolite FTIR spectra. Accordingly, it is possible that themeasuredband is Raman and infrared active, and is associated with theemergence of tridymite-type units from the amorphous precursor.
4.3. Microbiology
Total DNA has been extracted from four different levels of thesubmerged stromatolite representative sample and from lake super-ficial waters collected at the sampling site (Table 1). For a comparison,additional DNA data have been obtained from six levels in anotherstromatolite sample (SS-2 in Table 1). The quantity of total nucleicacids extractable from an environmental sample can be considered aproxy of biological colonization. The highest concentrations, up to1.6 μg DNA g−1 of stromatolite, have been retrieved in the uppersuperficial levels, consistent with the presence of a thin bright greenband putatively attributable to endolytic algal/cyanobacterial biofilm,positioned just underneath the superficial crust (Oren et al., 1995;Paerl et al., 2001). DNA concentrations in the superficial levels arehigher than in the lake water (see Table 1), suggesting that thestromatolite surface is an enrichment environment for microorgan-isms. In the intermediate sinter textured horizons, DNA is at least oneorder of magnitude less than in the surface levels, indicating thatthese parts of the stromatolites are microbially colonized, but to alesser extent. In some of the gelatinous material present in theintermediate levels, the extracted DNA is under the detection level,

Table 1Description, DNA extraction yield and results of amplification of the universal bacterialgene 16S rRNA of samples analysed for microbial colonization.
Sample Level Description Code DNAconcentration
16S rRNAamplification
ARISA
SS-1 A Superficialbrown level andunderneath thinbrilliant greenlevel
Top 26.8 ng/g + +
D Browngelatinous level
BGL 7.8 ng/g + +
D/E Green gelatinouslevel
GGL 12.4 ng/g + +
G Bottom level Bottom b0.1 ng/g + +W Superficial water SW 7.86 ng/ml + +SS-2 Superficial
brown level andunderneath thinbrilliant greenlevel
Top 1626 ng/g + +
White granularlevel
WGL 170.6 ng/g + +
Greenish level GL 38.2 ng/g + +Green-brownishgelatinous level
GBGL b0.1 ng/g + +
White level WL b0.1 ng/g + +Bottom level Bottom b0.1 ng/g + +
SS-1 = siliceous stromatolite (representative); W = water; SS-2 = siliceousstromatolite (additional).
Fig. 7. Bacterial structure and diversity in stromatolites from Lake Specchio di Venere,Pantelleria. Detrended Corrispondance Analysis (DCA) of the ARISA profiles obtainedfrom different levels in the representative stromatolite sample shown in Fig. 3 (A) andin an additional sample (B). This ordination method calculates the similarity betweenthe samples, represented by the spacing between them in the 2 axis plot. See Table 1 forthe code key; solid curves encircle samples from intermediate levels.
Table 2Phylogenetic identification of the clones from 16S rRNA libraries and relativeabundance of each taxon for the analysed samples.
Phylogenetic group Bottom Top GGL
Unclassified Bacteria 27 11 23Alphaproteobacteria Unidentified 2 2 2
Rhodobacterales 2 14Sphingomonadales 2Rhizobiales 16 7
Betaproteobacteria Unidentified 4Deltaproteobacteria Unidentified 7 2
Desulfobacterales 7 9Syntrophobacterales 2 2
Gamma-proteobacteria Unidentified 9 7 16Cyanobacteria 32Chromatiales 2 7 7Acidobacteriales 11 2Actinobacteria 5 11Bacteroidetes 5 7Chloroflexi 9 2Verrucomicrobiales 4 7Spirochaetales 2 9Nitrospirales 2Clostridiales 2
GGL = gelatinous greenish level.
326 M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
but in enough quantity for the amplification of the bacterial universalgene 16S rRNA, thus indicating that the bacterial colonization expandsin all the levels, even if at different extents (Table 1).
ARISA DNA fingerprinting has the potential to discriminate atspecies/subspecies level the different bacterial assemblages present ina sample (Cardinale et al., 2004). This technique has been applied onthe extracted DNA to describe the structure of the bacterialcommunities inhabiting the different stromatolite levels. The ordina-tion method Detrended Correspondence Analysis (DCA), previouslyused for the statistical analysis of environmental microbial diversity(e.g. Culman et al., 2008; Borin et al., 2010), was applied on theobtained fingerprints to evaluate the degree of similarity between thebacterial communities. The stromatolites appear to be colonized bybacterial communities taxonomically stratified, given that levels withdifferent morphologies contain peculiar communities. Nevertheless,not all the levels presenting the same morphology in differentstromatolites have harboured similar bacterial communities (data notshown). However, when analysed separately, samples from thedifferent stromatolites show similar trends (Fig. 7) with superficiallevels harbouring bacterial communities different from those of theintermediate levels, which, in contrast, have more similar communi-ties clustering together in the DCA plot. The upper and lower levels ofthe stromatolites are differently colonized, consistent with differentenvironmental characteristics in the upper part (top), exposeddirectly to solar irradiation and to the lake water column, and in thebottom part, partially embedded in the sediments.
Bacterial 16S rRNA gene libraries have been constructed fordetailed analyses of taxonomic diversity of bacteria colonizing upperand lower levels of the representative stromatolite sample and thegelatinous greenish material from an intermediate level D/E (Top,Bottom and CGL in Table 2). A total of 135 partial sequences, 800 bplong on average, have been aligned against the public sequencedatabase Ribosomal Database Project (RDP). The phylogeneticaffiliation of the sequences is shown in Table 2. Between 11 and 27%of the sequences did not find significant homology with any of thesequences contained in RDP, thus belonged to bacteria not previouslydescribed. The investigated samples exhibit high taxonomic diversity,with 19 major bacterial Phyla identified, confirming the ARISA
fingerprinting results. Statistical. ∫-Libshuff analysis based on ashuffling approach (Singleton et al., 2001) indicate that the 3 librariesare significantly different (Pb0.05). The photosynthetic taxa, Cyano-bacteria and Chromatiales, constitute the most abundant groups of thetop level (39% of the total clones, Table 2), consistent with the higher
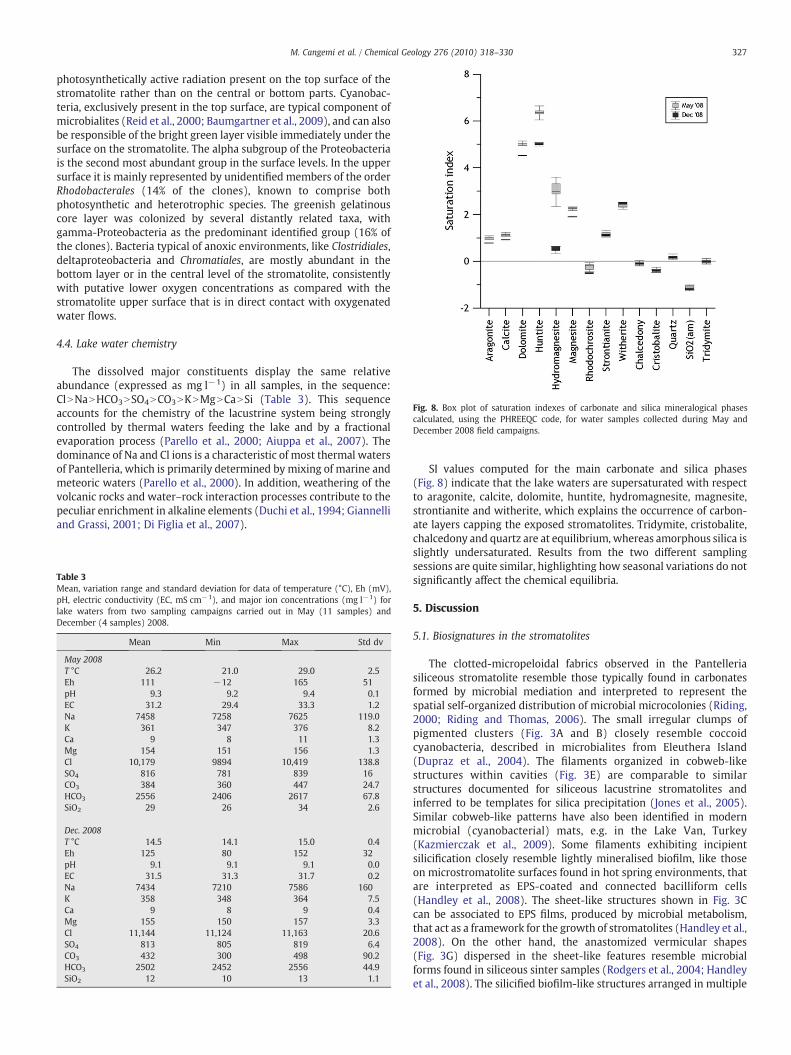
Fig. 8. Box plot of saturation indexes of carbonate and silica mineralogical phasescalculated, using the PHREEQC code, for water samples collected during May andDecember 2008 field campaigns.
327M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
photosynthetically active radiation present on the top surface of thestromatolite rather than on the central or bottom parts. Cyanobac-teria, exclusively present in the top surface, are typical component ofmicrobialites (Reid et al., 2000; Baumgartner et al., 2009), and can alsobe responsible of the bright green layer visible immediately under thesurface on the stromatolite. The alpha subgroup of the Proteobacteriais the second most abundant group in the surface levels. In the uppersurface it is mainly represented by unidentified members of the orderRhodobacterales (14% of the clones), known to comprise bothphotosynthetic and heterotrophic species. The greenish gelatinouscore layer was colonized by several distantly related taxa, withgamma-Proteobacteria as the predominant identified group (16% ofthe clones). Bacteria typical of anoxic environments, like Clostridiales,deltaproteobacteria and Chromatiales, are mostly abundant in thebottom layer or in the central level of the stromatolite, consistentlywith putative lower oxygen concentrations as compared with thestromatolite upper surface that is in direct contact with oxygenatedwater flows.
4.4. Lake water chemistry
The dissolved major constituents display the same relativeabundance (expressed as mg l−1) in all samples, in the sequence:ClNNaNHCO3NSO4NCO3NKNMgNCaNSi (Table 3). This sequenceaccounts for the chemistry of the lacustrine system being stronglycontrolled by thermal waters feeding the lake and by a fractionalevaporation process (Parello et al., 2000; Aiuppa et al., 2007). Thedominance of Na and Cl ions is a characteristic of most thermal watersof Pantelleria, which is primarily determined by mixing of marine andmeteoric waters (Parello et al., 2000). In addition, weathering of thevolcanic rocks and water–rock interaction processes contribute to thepeculiar enrichment in alkaline elements (Duchi et al., 1994; Giannelliand Grassi, 2001; Di Figlia et al., 2007).
Table 3Mean, variation range and standard deviation for data of temperature (°C), Eh (mV),pH, electric conductivity (EC, mS cm−1), and major ion concentrations (mg l−1) forlake waters from two sampling campaigns carried out in May (11 samples) andDecember (4 samples) 2008.
Mean Min Max Std dv
May 2008T °C 26.2 21.0 29.0 2.5Eh 111 −12 165 51pH 9.3 9.2 9.4 0.1EC 31.2 29.4 33.3 1.2Na 7458 7258 7625 119.0K 361 347 376 8.2Ca 9 8 11 1.3Mg 154 151 156 1.3Cl 10,179 9894 10,419 138.8SO4 816 781 839 16CO3 384 360 447 24.7HCO3 2556 2406 2617 67.8SiO2 29 26 34 2.6
Dec. 2008T °C 14.5 14.1 15.0 0.4Eh 125 80 152 32pH 9.1 9.1 9.1 0.0EC 31.5 31.3 31.7 0.2Na 7434 7210 7586 160K 358 348 364 7.5Ca 9 8 9 0.4Mg 155 150 157 3.3Cl 11,144 11,124 11,163 20.6SO4 813 805 819 6.4CO3 432 300 498 90.2HCO3 2502 2452 2556 44.9SiO2 12 10 13 1.1
SI values computed for the main carbonate and silica phases(Fig. 8) indicate that the lake waters are supersaturated with respectto aragonite, calcite, dolomite, huntite, hydromagnesite, magnesite,strontianite and witherite, which explains the occurrence of carbon-ate layers capping the exposed stromatolites. Tridymite, cristobalite,chalcedony and quartz are at equilibrium, whereas amorphous silica isslightly undersaturated. Results from the two different samplingsessions are quite similar, highlighting how seasonal variations do notsignificantly affect the chemical equilibria.
5. Discussion
5.1. Biosignatures in the stromatolites
The clotted-micropeloidal fabrics observed in the Pantelleriasiliceous stromatolite resemble those typically found in carbonatesformed by microbial mediation and interpreted to represent thespatial self-organized distribution of microbial microcolonies (Riding,2000; Riding and Thomas, 2006). The small irregular clumps ofpigmented clusters (Fig. 3A and B) closely resemble coccoidcyanobacteria, described in microbialites from Eleuthera Island(Dupraz et al., 2004). The filaments organized in cobweb-likestructures within cavities (Fig. 3E) are comparable to similarstructures documented for siliceous lacustrine stromatolites andinferred to be templates for silica precipitation (Jones et al., 2005).Similar cobweb-like patterns have also been identified in modernmicrobial (cyanobacterial) mats, e.g. in the Lake Van, Turkey(Kazmierczak et al., 2009). Some filaments exhibiting incipientsilicification closely resemble lightly mineralised biofilm, like thoseon microstromatolite surfaces found in hot spring environments, thatare interpreted as EPS-coated and connected bacilliform cells(Handley et al., 2008). The sheet-like structures shown in Fig. 3Ccan be associated to EPS films, produced by microbial metabolism,that act as a framework for the growth of stromatolites (Handley et al.,2008). On the other hand, the anastomized vermicular shapes(Fig. 3G) dispersed in the sheet-like features resemble microbialforms found in siliceous sinter samples (Rodgers et al., 2004; Handleyet al., 2008). The silicified biofilm-like structures arranged in multiple
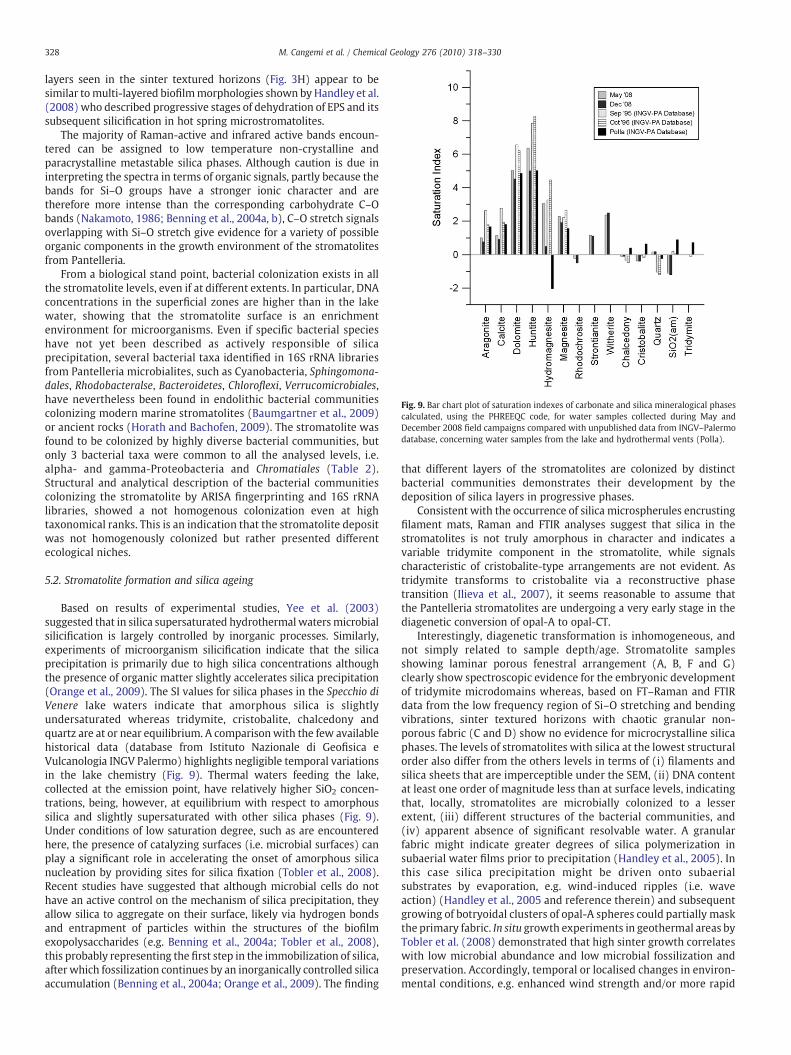
Fig. 9. Bar chart plot of saturation indexes of carbonate and silica mineralogical phasescalculated, using the PHREEQC code, for water samples collected during May andDecember 2008 field campaigns compared with unpublished data from INGV–Palermodatabase, concerning water samples from the lake and hydrothermal vents (Polla).
328 M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
layers seen in the sinter textured horizons (Fig. 3H) appear to besimilar tomulti-layered biofilmmorphologies shown by Handley et al.(2008) who described progressive stages of dehydration of EPS and itssubsequent silicification in hot spring microstromatolites.
The majority of Raman-active and infrared active bands encoun-tered can be assigned to low temperature non-crystalline andparacrystalline metastable silica phases. Although caution is due ininterpreting the spectra in terms of organic signals, partly because thebands for Si–O groups have a stronger ionic character and aretherefore more intense than the corresponding carbohydrate C–Obands (Nakamoto, 1986; Benning et al., 2004a, b), C–O stretch signalsoverlapping with Si–O stretch give evidence for a variety of possibleorganic components in the growth environment of the stromatolitesfrom Pantelleria.
From a biological stand point, bacterial colonization exists in allthe stromatolite levels, even if at different extents. In particular, DNAconcentrations in the superficial zones are higher than in the lakewater, showing that the stromatolite surface is an enrichmentenvironment for microorganisms. Even if specific bacterial specieshave not yet been described as actively responsible of silicaprecipitation, several bacterial taxa identified in 16S rRNA librariesfrom Pantelleria microbialites, such as Cyanobacteria, Sphingomona-dales, Rhodobacteralse, Bacteroidetes, Chloroflexi, Verrucomicrobiales,have nevertheless been found in endolithic bacterial communitiescolonizing modern marine stromatolites (Baumgartner et al., 2009)or ancient rocks (Horath and Bachofen, 2009). The stromatolite wasfound to be colonized by highly diverse bacterial communities, butonly 3 bacterial taxa were common to all the analysed levels, i.e.alpha- and gamma-Proteobacteria and Chromatiales (Table 2).Structural and analytical description of the bacterial communitiescolonizing the stromatolite by ARISA fingerprinting and 16S rRNAlibraries, showed a not homogenous colonization even at hightaxonomical ranks. This is an indication that the stromatolite depositwas not homogenously colonized but rather presented differentecological niches.
5.2. Stromatolite formation and silica ageing
Based on results of experimental studies, Yee et al. (2003)suggested that in silica supersaturated hydrothermal watersmicrobialsilicification is largely controlled by inorganic processes. Similarly,experiments of microorganism silicification indicate that the silicaprecipitation is primarily due to high silica concentrations althoughthe presence of organic matter slightly accelerates silica precipitation(Orange et al., 2009). The SI values for silica phases in the Specchio diVenere lake waters indicate that amorphous silica is slightlyundersaturated whereas tridymite, cristobalite, chalcedony andquartz are at or near equilibrium. A comparisonwith the few availablehistorical data (database from Istituto Nazionale di Geofisica eVulcanologia INGV Palermo) highlights negligible temporal variationsin the lake chemistry (Fig. 9). Thermal waters feeding the lake,collected at the emission point, have relatively higher SiO2 concen-trations, being, however, at equilibrium with respect to amorphoussilica and slightly supersaturated with other silica phases (Fig. 9).Under conditions of low saturation degree, such as are encounteredhere, the presence of catalyzing surfaces (i.e. microbial surfaces) canplay a significant role in accelerating the onset of amorphous silicanucleation by providing sites for silica fixation (Tobler et al., 2008).Recent studies have suggested that although microbial cells do nothave an active control on the mechanism of silica precipitation, theyallow silica to aggregate on their surface, likely via hydrogen bondsand entrapment of particles within the structures of the biofilmexopolysaccharides (e.g. Benning et al., 2004a; Tobler et al., 2008),this probably representing the first step in the immobilization of silica,after which fossilization continues by an inorganically controlled silicaaccumulation (Benning et al., 2004a; Orange et al., 2009). The finding
that different layers of the stromatolites are colonized by distinctbacterial communities demonstrates their development by thedeposition of silica layers in progressive phases.
Consistent with the occurrence of silica microspherules encrustingfilament mats, Raman and FTIR analyses suggest that silica in thestromatolites is not truly amorphous in character and indicates avariable tridymite component in the stromatolite, while signalscharacteristic of cristobalite-type arrangements are not evident. Astridymite transforms to cristobalite via a reconstructive phasetransition (Ilieva et al., 2007), it seems reasonable to assume thatthe Pantelleria stromatolites are undergoing a very early stage in thediagenetic conversion of opal-A to opal-CT.
Interestingly, diagenetic transformation is inhomogeneous, andnot simply related to sample depth/age. Stromatolite samplesshowing laminar porous fenestral arrangement (A, B, F and G)clearly show spectroscopic evidence for the embryonic developmentof tridymite microdomains whereas, based on FT–Raman and FTIRdata from the low frequency region of Si–O stretching and bendingvibrations, sinter textured horizons with chaotic granular non-porous fabric (C and D) show no evidence for microcrystalline silicaphases. The levels of stromatolites with silica at the lowest structuralorder also differ from the others levels in terms of (i) filaments andsilica sheets that are imperceptible under the SEM, (ii) DNA contentat least one order of magnitude less than at surface levels, indicatingthat, locally, stromatolites are microbially colonized to a lesserextent, (iii) different structures of the bacterial communities, and(iv) apparent absence of significant resolvable water. A granularfabric might indicate greater degrees of silica polymerization insubaerial water films prior to precipitation (Handley et al., 2005). Inthis case silica precipitation might be driven onto subaerialsubstrates by evaporation, e.g. wind-induced ripples (i.e. waveaction) (Handley et al., 2005 and reference therein) and subsequentgrowing of botryoidal clusters of opal-A spheres could partially maskthe primary fabric. In situ growth experiments in geothermal areas byTobler et al. (2008) demonstrated that high sinter growth correlateswith low microbial abundance and low microbial fossilization andpreservation. Accordingly, temporal or localised changes in environ-mental conditions, e.g. enhanced wind strength and/or more rapid

329M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
evaporation, might have caused variations in the degree of silicaoversaturation in the Specchio di Venere lake waters. To explain theapparent absence of a tridymite component in the sinter texturedhorizons, it is suggested that a non-porous fabric could limit the fluxof pore fluids which should sustain the process of silica maturation. Itis commonly thought that loss of water is an integral part of the opal-A to opal-CT transformation and, therefore, opal-CT always containsless water than opal-A. However, it has been demonstrated that theamount of water initially bound into the opal-A can vary dependingon different environmental conditions at the time of its formation(Jones and Renaut, 2004, 2007). Consistent with the model of Jonesand Renaut (2004), “dry” opal-A in the sinter textured horizons ofPantelleria stromatolite could result by transformation of “wet” opal-A following desiccation on a period of (or in places where) lakewaters evaporated to dryness.
6. Conclusion
The study shows that the laminar, porous stromatolites of upperand lower levels, showing filamentous network, silica sheets and/orfenestrae, formed at relatively low rates of silica deposition andwith apossible microbial mediation in terms of reactive cell surfaces and EPSproviding sites for the silica fixation. The resulting high porosityprobably favoured a consistent flux of silica-rich fluids that triggeredopal-A to trydimite diagenetic transformation, thus strengthening thepreservation of biosignals. Granular non-porous sinter texturedhorizons, showing minor evidence of microbial colonization and noapparent filament networks in the siliceous matrix, formed at higherdeposition rates. Probably, in this case, the growing process is mainlygoverned by chemical factors. It is also hypothesized that thestructure of stromatolites varies with changes of environmentalconditions, which may control the nature of precipitated silica(degree of silica polymerization). Owing to the concurrence ofinhomogenous fabric and microbial abundance and diversity, furtherresearch on the colonizing microbial communities could address thepossible role of microbiota on the different structural arrangements ofstromatolites.
Acknowledgements
We are grateful to LAMP Research Team (Department DSFA,Palermo University) for facilitating part of FT–Raman analyses, and tothe management of INGV Palermo for access to their chemicaldatabase. We would like to thank Agostino Rizzi (CNR IDPA, MilanUniversity) for help in preliminary SEM investigations. Thanks arealso due to Paolo Madonia (INGV Palermo) for underwater images,and to Salvatore Francofonte and Lorenzo Brusca (INGV Palermo) forassistance duringmajor and trace element determinations. Commentsby two anonymous reviewers on a previous version of this manuscriptwere extremely helpful. This work is part of the Ph.D. thesis of MCsupported by MIUR Funds; additional financial support was providedby MIUR (ex-quota 60%) Grants to AB and RN.
References
Aiuppa, A., D'Alessandro, W., Gurrieri, S., Madonia, P., Parello, F., 2007. Hydrologic andgeochemical survey of the lake “Specchio di Venere” (Pantelleria island, SouthernItaly). Environmental Geology 53, 903–913.
Allwood, A.C., Walter, M.R., Kamber, B.S., Marshall, C.P., Burch, I.W., 2006. Stromatolitereef from the Early Archaean era of Australia. Nature 441, 714–718.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman, D.J.,1997. Gapped BLAST and PSI-BLAST: a new generation of protein database searchprograms. Nucleic Acids Research 25, 3389–3402.
Andres, M.S., Reid, R.P., 2006. Growth morphologies of modern marine stromatolites: acase of study from Highborne Cay, Bahamas. Sedimentary Geology 185, 319–328.
Awramik, S.M., 1984. Ancient stromatolites and microbial mats. In: Cohen, Y.,Castenholz, R.W., Halvorson, H.O. (Eds.), Microbial Mats: Stromatolites. A.R. LissInc, New York, pp. 1–22.
Baumgartner, L.K., Spear, J.R., Buckley, D.H., Pace, N.R., Reid, P., Dupraz, C., Visscher, P.T.,2009. Microbial diversity in modern marine stromatolites, Highborne Cay,Bahamas. Environmental Microbiology 11, 2710–2719.
Benning, L.G., Phoenix, V.R., Yee, N., Konhauser, K.O., 2004a. The dynamics ofcyanobacterial silification: an infraredmicro-spectroscopic investigation. Geochimicaet Cosmochimica Acta 68, 743–757.
Benning, L.G., Phoenix, V.R., Yee, N., Tobin, M.J., 2004b. Molecular characterisation ofcyanobacterial silicificationusing synchrotron infraredmicro-spectroscopy.Geochimicaet Cosmochimica Acta 68, 729–741.
Bocchi, G., Gabbianelli, G., Lanzafame, G., Lucchini, F., Rabbi, E., Rossi, P.L., 1988.Relazione sui rilievi eseguiti sul “lago Specchio di Venere”, Pantelleria. Bulletin ofthe Italian National Group for Volcanology 4, 63–73.
Borin, S., Brusetti, L., Daffonchio, D., Delaney, E., Baldi, F., 2009. Biodiversity ofprokaryotic communities in sediments of different sub-basins of the Venice lagoon.Research in Microbiology 160, 307–314.
Borin, S., Ventura, S., Tambone, F., Mapelli, F., Schubotz, F., Brusetti, L., Scaglia, B.,D'Acqui, L.P., Solheim, B., Turicchia, S., Marasco, R., Hinrichs, K.U., Baldi, F., Adani, F.,Daffonchio, D., 2010. Rock weathering creates oases of life in a high arctic desert.Environmental Microbiology 12, 293–303.
Brown, K.L., McDowell, G.D., 1983. pH control of silica scaling. Proceedings of the5th NZ Geothermal Workshop. University of Auckland, Auckland, New Zealand,pp. 1157–1162.
Budakoglu, M., 2009. Comparison of recent siliceous and carbonate mat developmenton the shore of hyper-alkaline Lake Van and Mt. Nemrut Soğuk Lake, NE Anatolia,Turkey. Geomicrobiology Journal 26, 146–160.
Burne, R.V., Moore, L.S., 1987. Microbialites; organosedimentary deposits of benthicmicrobial communities. Palaios 2, 241–254.
Campbell, K.A., Rodgers, K.A., Brotheridge, J.M.A., Browne, P.R.L., 2002. An unusualmodern silica-carbonate sinter from Pavlova spring, Ngatamariki, New Zealand.Sedimentology 49, 835–854.
Cangemi M., 2009. Petrographic and geochemical evidence of biomineralization inseep-related carbonates from the Sicily Strait and in siliceous stromatolites fromSpecchio di Venere lake, Pantelleria Island. Ph.D. Thesis, Palermo University, Italy,86 pp.
Cardinale, M., Brusetti, L., Quatrini, P., Borin, S., Puglia, A.M., Rizzi, A., Zanardini, E.,Sorlini, C., Corselli, C., Daffonchio, D., 2004. Comparison of different primer sets forthe automated ribosomal intergenic spacer analysis (ARISA) of complex bacterialcommunities. Applied and Environmental Microbiology 70, 6147–6156.
Cavallazzi, J.R.P., Filho, O.K., Stuermer, S.L., Rygiewicz, P.T., de Mendonca, M.M.,2007. Screening and selecting arbuscular mycorrhizal fungi for inoculatingmicropropagated apple rootstocks in acid soils. Plant Cell, Tissue and OrganCulture 90, 117–129.
Civetta, L., Cornette, Y., Crisci, G., Gillot, P.Y., Orsi, G., Requeiros, C.S., 1984. Geology,geochronology and chemical evolution of the island of Pantelleria. GeologicalMagazine 121, 541–562.
Cross, A.D., Jones, R.A., 1969. An Introduction to Practical Infra-red Spectroscopy, Thirdedition. Butterworths, London, U.K.
Culman, S.W., Gauch, H.G., Blackwood, C.B., Thies, J.E., 2008. Analysis of T-RFLP datausing analysis of variance and ordination methods: a comparative study. Journal ofMicrobiological Methods 75, 55–63.
D'Alessandro, W., Dongarrà, G., Gurrieri, S., Parello, F., Valenza, M., 1994. Geochemicalcharacterization of naturally occurring fluids on the island of Pantelleria.Mineralogica et Petrographica Acta 27, 91–102.
Di Figlia, M.G., Bellanca, A., Neri, R., Stefansson, A., 2007. Chemical weathering ofvolcanic rocks at the island of Pantelleria, Italy: information from soil profile andsoil solution investigations. Chemical Geology 246, 1–18.
Duchi, V., Campana, M.E., Minissale, A., Thompson, M., 1994. Geochemistry of thermalfluids on the volcanic isle of Pantelleria, southern Italy. Applied Geochemistry 9,147–160.
Dupraz, C., Visscher, P.T., Baumgartner, L.K., Reid, R.P., 2004. Microbe–mineralinteractions: early carbonate precipitation in a hypersaline lake (Eleuthera Island,Bahamas). Sedimentology 51, 745–765.
Farmer, J., 1999. Taphonomic modes in microbial fossilization. Size Limits of Very SmallMicroorganisms: Proceedings of aWorkshop. National Academy Press, WashingtonDC, pp. 94–102.
Favara, R., Giammanco, S., Inguaggiato, S., Pecoraino, G., 2001. Preliminary estimate ofCO2 output from Pantelleria Island volcano (Sicily, Italy): evidence of active mantledegassing. Applied Geochemistry 16, 883–894.
Fleming, B.A., 1986. Kinetics of reaction between silicic acid and amorphous silicasurfaces in NaCl solutions. Journal of Colloid and Interface Science 110, 40–64.
Giannelli, G., Grassi, S., 2001. Water–rock interaction in the active geothermal system ofPantelleria, Italy. Chemical Geology 181, 113–130.
Graetsch, H., Gies, H., Topalović, I., 1994. NMR, XRD and IR study on microcrystallineopal. Physics and Chemistry of Minerals 21, 166–175.
Handley, K.M., Campbell, K.A., Mountain, B.W., Browne, P.R.L., 2005. Abiotic–bioticcontrols on the origin and development of spicular sinter: in situ growthexperiments, Champagne Pool, Waiotapu, New Zealand. Geobiology 3, 93–114.
Handley, K.M., Turner, S.J., Campbell, K.A., Mountain, B.W., 2008. Silicifying biofilmexopolymers on a hot-spring microstromatolite: templating nanometer-thicklaminae. Astrobiology 8, 747–770.
Hatipoğlu, M., 2009. Moganite and quartz inclusions in the nano-structured Anatolianfire opals from Turkey. Journal of African Earth Science 54, 1–21.
Heaney, P.J., 1994. Structure and chemistry of the low-pressure silica polymorphs. In:Heaney, P.J., Prewitt, C.T., Gibbs, G.V. (Eds.), Silica, Physical Behavior, Geochemistryand Material Applications. Reviews in Mineralogy, vol. 29. Mineralogical Society ofAmerica, Washington, DC, pp. 1–40.

330 M. Cangemi et al. / Chemical Geology 276 (2010) 318–330
Herdianita, N.R., Browne, P.R.L., Rodgers, K.A., Campbell, K.A., 2000. Mineralogical andtextural changes accompanying ageing of silica sinter. Mineralium Deposita 35,48–62.
Hino, M., Sato, T., 1971. Infrared absorption spectra of silica gel–water, water–d2, andwater–18O systems. Bulletin. Chemical Society of Japan 44, 33–37.
Horath, T., Bachofen, R., 2009. Molecular characterization of an endolithic microbialcommunity in dolomite rock in the Central Alps (Switzerland). Microbial Ecology58, 290–306.
Iler, R.K., 1973. Colloidal silica. Surface and Colloid Science 6, 1–100.Ilieva, A., Mihailova, B., Tsintsov, Z., Petrov, O., 2007. Structural state of microcrystalline
opals. American Mineralogist 92, 1325–1333.Jones, B., Renaut, R.W., 2004. Water content of opal-A: implications for the origin of
laminae in geyserite and sinter. Journal of Sedimentary Research 74, 117–128.Jones, B., Renaut, R.W., 2007. Microstructural changes accompanying the opal-A to
opal-CT transition: new evidence from the siliceous sinters of Geysir, Haukadalur,Iceland. Sedimentology 54, 921–948.
Jones, J.B., Segnit, E.R., 1971. The nature of opal I. Nomenclature and constituent phases.Australian Journal of Earth Sciences 18, 57–68.
Jones, J.B., Segnit, E.R., 1972. Genesis of cristobalite and tridymite at low temperatures.Australian Journal of Earth Sciences 18, 419–422.
Jones, B., Renaut, R.W., Rosen, M.R., 1997. Vertical zonation of biota in microstromatolitesassociated with hot springs, North Island, New Zealand. Palaios 12, 220–236.
Jones, B., Renaut, R.W., Konhauser, K.O., 2005. Genesis of large siliceous stromatolites atFlying Pan Lake, Waimangu geothermal field, North Island, New Zealand.Sedimentology 52, 1229–1252.
Kalkowsky, E., 1908. Oolith und Stromatolith im nord-deutschen Buntsandstein:Deutsch Geologische Gesellschaft. Zeitschrift 60, 68–125.
Kazmierczak, J., Altermann, W., Kremera, B., Kemped, S., Erikssonb, P.G., 2009.Mass occurrence of benthic coccoid cyanobacteria and their role in the productionof Neoarchean carbonates of South Africa. Precambrian Research. doi:10.1016/j.precamres.2009.02.002.
Kingma, K.J., Hemley, R.J., 1994. Raman spectroscopic study of microcrystalline silica.American Mineralogist 79, 269–273.
Konhauser, K.O., Ferris, F.G., 1996. Diversity of iron and silica precipitation by microbialmats in hydrothermal waters, Iceland; implications for Precambrian iron formations.Geology 24, 323–326.
Konhauser, K.O., Jones, B., Phoenix, V.R., Ferris, G., Renaut, R.W., 2004. The microbialrole in hot spring silicification. Ambio 33 (8), 552–558.
Krumbein, W.E., Werner, D., 1983. The microbial silica cycle. In: Krumbein, W.E. (Ed.),Microbial Geochemistry. Blackwell Scientific Publications, New York, pp. 125–157.
Landmesser, H., Kosslick, H., Storek, W., Fricke, R., 1997. Interior surface hydroxylgroups in ordered mesoporous silicates. Solid State Ionics 101–103, 271–277.
Maidak, B.L., Cole, J.R., Lilburn, T.G., Parker Jr, C.T., Saxman, P.R., Farris, R.J., 2001. TheRDP-II (Ribosomal Database Project). Nucleic Acids Research 29, 173–174.
McMillan, P.F., Wolf, G.H., 1995. Vibrational spectroscopy of silicate liquids. Reviews inMineralogy and Geochemistry 32, 247–315.
Moenke, H., 1966. Mineralspektren 2. Die Ultrarotabsorption häufiger und paragenetischoder wirtschaftlich wichtiger Carbonat-, Borat-, Sulfat-, Chromat-, Phosphat-,... 400–4000 cm hochminus 1 (25–2,5 m) (Unbekannter Einband). Akademie-Verlag, Berlin,Germany.
Mountain, B.W., Benning, L.G., Boerema, J.A., 2003. Experimental studies on NewZealand hot spring sinters: rates of growth and textural development. CanadianJournal of Earth Sciences 40, 1643–1667.
Mroczek, E.K., Reeves, R.R., 1994. The effect of colloidal silica on silica scaling fromgeothermal fluid. Proceedings of the 16th NZ Geothermal Workshop. GeothermalInstitute, University of Auckland, Auckland, New Zealand, pp. 97–101.
Murray, A.E., Preston, C.M., Massana, R., Taylor, L.T., Blakis, A., Wu, K., DeLong, E.F., 1998.Seasonal and spatial variability of bacterial and Archaeal assemblages in the coastalwaters near Anvers Island, Antarctica. Applied and Environmental Microbiology64 (7), 2585–2595.
Nakamoto, K., 1986. Infrared and Raman Spectra of Inorganic and CoordinationCompounds. Wiley, New York. pp. 272–280.
Nicholson, K., Aquino, C.G., 1989. Life in geothermal systems: a key to sinter formationand recognition? Proceedings of the 11th NZ Geothermal Workshop. GeothermalInstitute, University of Auckland, Auckland, New Zealand, pp. 143–148.
Olivier, N., Boyet, M., 2006. Rare earth and trace elements of microbialites in UpperJurassic coral- and sponge-microbialite reefs. Chemical Geology 230, 105–123.
Orange, F., Westall, F., Disnar, J.-R., Prieur, D., Bienvenu, N., Le Romancer, M., Défarge, C.H.,2009. Experimental silicification of the extremophilic Archaea Pyrococcus abyssi andMethanocaldococcus jannaschii: applications in the search for evidence of life in earlyEarth and extraterrestrial rocks. Geobiology 7, 403–418.
Oren, A., Khul, M., Karsten, U., 1995. An endoevaporitic microbial mat within a gypsumcrust: zonation of phototrophs, photopigments, and light penetration. MarineEcology Progress Series 128, 151–159.
Paerl, H.W., Steppe, T.F., Reid, P., 2001. Bacterially mediated precipitation in marinestromatolites. Environmental Microbiology 3 (2), 123–130.
Parello, F., Allard, P., D'Alessandro, W., Federico, C., Jean-Baptiste, P., Catani, O., 2000.Isotope geochemistry of Pantelleria volcanic fluids, Sicily Channel rift: a mantlevolatile end-member for volcanism in southern Europe. Earth and PlanetaryScience Letters 180, 325–339.
Parkhurst, D.L., Appelo, C.A.J., 1999. User's guide to PHREEQC (version 2) — a computerprogram for speciation, batch-reaction, one-dimensional transport, and inversegeochemical modelling calculations. U.S. Geological Survey Water — ResourcesInvestigations Report 99–4259. . 312 pp.
Phoenix, V.R., Adams, D.G., Konhauser, K.O., 2000. Cyanobacterial viability duringhydrothermal biomineralisation. Chemical Geology 169, 329–338.
Reid, R.P., Visscher, P.T., Decho, A.W., Stolz, J.F., Beboutk, B.M., Dupraz, C., Macintyre, I.G.,Paerl, H.W., Pinckney, J.L., Prufert-Beboutk, L., Steppe, T.F., DesMaraisk, D.J., 2000.The role of microbes in accretion, lamination and early lithification of modernmarine stromatolites. Nature 406, 989–992.
Renaut, R.W., Jones, B., Tiercelin, J.-J., 1998. Rapid in situ silicification of microbes atLoburu hot springs, Lake Bogoria, Kenya Rift Valley. Sedimentology 45, 1083–1103.
Rice, S.B., Freund, H., Huang, W.L., Clouse, J.A., Isaacs, C.M., 1995. Application of Fouriertransform infrared spectroscopy to silica diagenesis: the opal-A to opal-CTtransformation. Journal of Sedimentary Research 65, 639–647.
Riding, R., 2000. Microbial carbonates: the geological record of calcified bacterial-algalmats and biofilms. Sedimentology 47, 179–214.
Riding, R., Thomas, S., 2006. Stromatolite reef crusts, Early Cretaceous, Spain: bacterialorigin of in situ-precipitation peloid microspar? Sedimentology 53, 23–34.
Rimstidt, J.D., Barnes, H.L., 1980. The kinetics of silica–water reactions. Geochimica etCosmochimica Acta 44, 1683–1699.
Rodgers, K.A., Browne, P.R.L., Buddle, T.F., Cook, K.L., Greatrex, R.A., Hampton, W.A.,Herdianita, N.R., Holland, G.R., Lynne, B.Y., Martin, R., Newton, Z., Pastars, D.,Sannazarro, Z.L., Teece, C.I.A., 2004. Silica phases in sinters and residues fromgeothermal fields of New Zealand. Earth Science Reviews 66, 1–61.
Rotolo, S.G., La Felice, S., Mangalaviti, A., Landi, P., 2007. Geology and petrochemistry ofthe recent (b25 ka) silicic volcanism at Pantelleria Island. Bollettino. SocietaGeologica Italiana 126, 191–208.
Schopf, J.W., 1993. Microfossils of the early Archean apex chert: new evidence of theantiquity of life. Science 260, 640–646.
Singleton, D.R., Furlong, M.A., Rathbun, S.L., Whitman, W.B., 2001. Quantitativecomparisons of 16S rRNA gene sequence libraries from environmental samples.Applied and Environmental Microbiology 67, 4373–4376.
Smallwood, A.G., Thomas, P.S., Ray, A.S., 1997. Characterisation of sedimentary opals byFourier transform Raman spectroscopy. Spectrochimica Acta 53, 2341–2345.
Sunagawa, I., Takahashi, Y., Imai, H., 2007. Strontium and aragonite–calcite precipitation.Journal of Mineralogical and Petrological Sciences 102, 174–181.
Tobler, D.J., Stefánsson, A., Benning, L.G., 2008. In-situ grown silica sinters in Icelandicgeothermal areas. Geobiology 6, 481–502.
Tucker, M.E., 2001. Sedimentary Petrology. Blackwell Scientific Pubblication, Oxford.262 pp.
Urrutia, M.M., Beveridge, T.J., 1993. Mechanism of silicate binding to the bacterial cellwall in Bacillus subtilis. Journal of Bacteriology 175, 1936–1945.
Van Lith, Y., Warthmann, R., Vasconcelos, C., McKenzie, J.A., 2003. Microbial fossilizationin carbonate sediments: a result of the bacterial surface involvement in dolomiteprecipitation. Sedimentology 50, 237–245.
Walters, C.C., Margulis, L., Barghoorn, E.S., 1977. On the experimental silicification ofmicroorganisms. I. Microbial growth on organosilicon compounds. PrecambrianResearch 5, 241–248.
Webb, G.E., Kamber, B.S., 2000. Rare earth elements in Holocene reefal microbialites: anew shallow seawater proxy. Geochimica et Cosmochimica Acta 64, 1557–1565.
Westall, F., Boni, L., Guerzoni, E., 1995. The experimental silicification ofmicrooganisms.Palaeontology 38, 495–528.
White, W.B., 1974. The carbonate minerals. In: Farmer, V.C. (Ed.), MineralogicalMonograph 4. The infrared spectra of minerals. Mineralogical Society, pp. 227–282.
White, D.E., Brannock, W.W., Murata, K.J., 1956. Silica in hot-spring waters. Geochimicaet Cosmochimica Acta 10, 27–59.
Yee, N., Phoenix, V.R., Konhauser, K.O., Benning, L.G., Ferris, F.G., 2003. The effect ofcyanobacteria on silica precipitation at neutral pH: implications for bacterialsilicification in geothermal hot springs. Chemical Geology 199, 83–90.
Related Documents











