Journal of Neuroscience Methods 140 (2004) 39–46 The GABA uptake inhibitor -alanine reduces pilocarpine-induced tremor and increases extracellular GABA in substantia nigra pars reticulata as measured by microdialysis Keita Ishiwari, Susana Mingote, Merce Correa 1 , Jennifer T. Trevitt 2 , Brian B. Carlson, John D. Salamone ∗ Department of Psychology, University of Connecticut, Storrs, CT 06269-1020, USA Received 30 October 2003; accepted 29 March 2004 Abstract Substantia nigra pars reticulata (SNr) is a major output nucleus of the basal ganglia that receives GABAergic projections from neostriatum and globus pallidus. Previous research has shown that local pharmacological manipulations of GABA in SNr can influence tremulous jaw movements in rats. Tremulous jaw movements are defined as rapid vertical deflections of the lower jaw that resemble chewing but are not directed at a particular stimulus, and evidence indicates that these movements share many characteristics with parkinsonian tremor in humans. In order to investigate the role of GABA in motor functions related to tremor, the present study tested the GABA uptake blocker -alanine for its ability to reduce pilocarpine-induced tremulous jaw movements. In a parallel experiment, the effect of an active dose of -alanine on dialysate levels of GABA in SNr was assessed using microdialysis methods. GABA levels in dialysis samples were measured using high performance liquid chromatography with electrochemical detection. -Alanine (250–500 mg/kg) significantly reduced tremulous jaw movements induced by pilocarpine (4.0 mg/kg). Moreover, systemic administration of -alanine at a dose that reduced tremulous jaw movements (500 mg/kg) resulted in a substantial increase in extracellular levels of GABA in SNr compared to the pre-injection baseline. Thus, the present results are consistent with the hypothesis that GABAergic tone in SNr plays a role in the regulation of tremulous jaw movements. This research may lead to a better understanding of how parkinsonian symptoms are modulated by SNr GABA mechanisms. © 2004 Elsevier B.V. All rights reserved. Keywords: Basal ganglia; Striatonigral; Acetylcholine; Parkinson’s disease; Muscarinic; Dialysis 1. Introduction Substantia nigra pars reticulata (SNr) is a mesencephalic nucleus that serves several important functions in the overall circuitry of the basal ganglia (Bevan et al., 1996; Bolam et al., 2000; Trevitt et al., 2001). Although substantia nigra pars compacta contains dopamine (DA) cell bodies that project to ∗ Corresponding author. Tel.: +1 860 486 4302; fax: +1 860 486 2760. E-mail address: [email protected] (J.D. Salamone). 1 Present address: Area de Psicobiologia, Campus Riu Sec, Universitat Jaume I, 12079 Castell ´ o, Spain. 2 Present address: Department of Psychology, California State University, Fullerton, CA, USA. neostriatum, the SNr is ventral to the compacta, and receives GABAergic projections from both neostriatum and lateral globus pallidus (Parent et al., 1984; Loopjuit and Van der Kooy, 1985; Gerfen, 1992; Parent and Hazrati, 1993; Fallon and Laughlin, 1995). GABAergic neurons that originate in SNr project to thalamic motor nuclei and various brainstem motor areas (Faull and Mehler, 1978; Fallon and Laughlin, 1995; Kha et al., 2001), and therefore SNr is thought to be one of the major relay nuclei through which basal ganglia out- put is conveyed to other motor regions of the brain. Several studies have focused upon the behavioral functions of fore- brain DA systems (e.g., McCullough et al., 1993; Salamone, 1996; Salamone et al., 1994, 1997; Sokolowski et al., 1998), yet much less is known about the behavioral functions of 0165-0270/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.jneumeth.2004.03.030

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Neuroscience Methods 140 (2004) 39–46
The GABA uptake inhibitor�-alanine reduces pilocarpine-inducedtremor and increases extracellular GABA in substantia nigra pars
reticulata as measured by microdialysis
Keita Ishiwari, Susana Mingote, Merce Correa1, Jennifer T. Trevitt2,Brian B. Carlson, John D. Salamone∗
Department of Psychology, University of Connecticut, Storrs, CT 06269-1020, USA
Received 30 October 2003; accepted 29 March 2004
Abstract
Substantia nigra pars reticulata (SNr) is a major output nucleus of the basal ganglia that receives GABAergic projections from neostriatuma ulous jawm but are notd r in humans.Ia el rformancel ucedb g/kg)r results arec arch mayl©
K
1
ncac
J
F
eivesralerllon
intemlin,beout-eral
fore-e,98s of
0d
nd globus pallidus. Previous research has shown that local pharmacological manipulations of GABA in SNr can influence tremovements in rats. Tremulous jaw movements are defined as rapid vertical deflections of the lower jaw that resemble chewingirected at a particular stimulus, and evidence indicates that these movements share many characteristics with parkinsonian tremo
n order to investigate the role of GABA in motor functions related to tremor, the present study tested the GABA uptake blocker�-alanine for itsbility to reduce pilocarpine-induced tremulous jaw movements. In a parallel experiment, the effect of an active dose of�-alanine on dialysat
evels of GABA in SNr was assessed using microdialysis methods. GABA levels in dialysis samples were measured using high peiquid chromatography with electrochemical detection.�-Alanine (250–500 mg/kg) significantly reduced tremulous jaw movements indy pilocarpine (4.0 mg/kg). Moreover, systemic administration of�-alanine at a dose that reduced tremulous jaw movements (500 mesulted in a substantial increase in extracellular levels of GABA in SNr compared to the pre-injection baseline. Thus, the presentonsistent with the hypothesis that GABAergic tone in SNr plays a role in the regulation of tremulous jaw movements. This reseead to a better understanding of how parkinsonian symptoms are modulated by SNr GABA mechanisms.
2004 Elsevier B.V. All rights reserved.
eywords:Basal ganglia; Striatonigral; Acetylcholine; Parkinson’s disease; Muscarinic; Dialysis
. Introduction
Substantia nigra pars reticulata (SNr) is a mesencephalicucleus that serves several important functions in the overallircuitry of the basal ganglia (Bevan et al., 1996; Bolam etl., 2000; Trevitt et al., 2001). Although substantia nigra parsompacta contains dopamine (DA) cell bodies that project to
∗ Corresponding author. Tel.: +1 860 486 4302; fax: +1 860 486 2760.E-mail address:[email protected] (J.D. Salamone).
1 Present address: Area de Psicobiologia, Campus Riu Sec, Universitataume I, 12079 Castello, Spain.2 Present address: Department of Psychology, California State University,ullerton, CA, USA.
neostriatum, the SNr is ventral to the compacta, and recGABAergic projections from both neostriatum and lateglobus pallidus (Parent et al., 1984; Loopjuit and Van dKooy, 1985; Gerfen, 1992; Parent and Hazrati, 1993; Faand Laughlin, 1995). GABAergic neurons that originateSNr project to thalamic motor nuclei and various brainsmotor areas (Faull and Mehler, 1978; Fallon and Laugh1995; Kha et al., 2001), and therefore SNr is thought toone of the major relay nuclei through which basal gangliaput is conveyed to other motor regions of the brain. Sevstudies have focused upon the behavioral functions ofbrain DA systems (e.g.,McCullough et al., 1993; Salamon1996; Salamone et al., 1994, 1997; Sokolowski et al., 19),yet much less is known about the behavioral function
165-0270/$ – see front matter © 2004 Elsevier B.V. All rights reserved.oi:10.1016/j.jneumeth.2004.03.030

40 K. Ishiwari et al. / Journal of Neuroscience Methods 140 (2004) 39–46
SNr GABA. The SNr is very rich in GABA, and the predom-inant postsynaptic GABA receptor in SNr is the GABAAsubtype (Bowery et al., 1987). Pharmacological modulationof GABAA receptor function in SNr can affect various as-pects of behavioral function, including locomotor activity,startle, catalepsy and lever pressing (Di Chiara et al., 1998;Scheel-Kruger et al., 1981a, 1981b; Scheel-Kruger, 1983;Cools et al., 1983; Baumeister et al., 1988; Timmerman andAbercrombie, 1996; Koch et al., 2000; Correa et al., 2003).GABAA receptor blockade has been shown to reduce mo-tor activity, while stimulation of GABAA function in SNrinduced motor activity and produced effects consistent withantiparkinsonian actions in animal models (Trevitt et al.,2002; Wichmann et al., 2001). Moreover, it has been sug-gested that some of the behavioral effects of DA D1 agonistsare mediated by GABA release in SNr (Starr and Starr, 1989;Matuszewich and Yamamoto, 1999; Trevitt et al., 2002). Lo-cal infusion of the D1 agonist SKF 82958 into SNr substan-tially elevated extracellular GABA as measured by micro-dialysis (Trevitt et al., 2002). The stimulation of locomotionand suppression of tremor induced by SKF 82958 were re-versed by infusions of the GABAA receptor antagonist bicu-culline directly into SNr (Mayorga et al., 1999; Trevitt et al.,2002).
One of the animal procedures that is used to study motorf nts.T ionso s ofP -c ove-m tiont wss anb e,1 ofD skin,1 an-t one,1 a eta isma anti-c onea n-d or inhi .,1 edj rkin-sb angliac so-n A int ssiono thatw etelyr e
into SNr (Mayorga et al., 1999). Injections of bicuculline di-rectly into the SNr were shown to induce tremulous jaw move-ments (Salamone et al., 1998). Local infusions of the GABAAagonist muscimol into SNr were able to completely blockcholinomimetic-induced jaw movements, while infusions ofmuscimol into entopeduncular nucleus failed to affect jawmovement activity (Finn et al., 1997b). Implantations of en-gineered GABA-synthesizing cells into SNr also significantlyreduced cholinomimetic-induced tremulous jaw movements,whereas implantations of these cells into a dorsal brainstemcontrol site did not (Carlson et al., 2003). Thus, several linesof evidence indicate that SNr GABA is involved in the modu-lation of tremulous jaw movement activity. The present stud-ies were undertaken to investigate the behavioral functionsof GABA using a combination of behavioral pharmacologyand microdialysis methods. As described above, previous re-search has shown that local pharmacological manipulationsof GABA in SNr can influence tremulous jaw movements,suggesting that increasing GABAergic tone in SNr may al-leviate parkinsonian symptoms. To examine this possibility,the present study tested the GABA uptake blocker�-alaninefor its ability to reduce pilocarpine-induced tremulous jawmovements. In addition, parallel neurochemical studies in-vestigated the effect of systemic injections of a behaviourallyactive dose of�-alanine on extracellular levels of GABAi beens theG dO hass ari-eA ys-t e-i rallya BAi
2
2
, In-dTl erea andw e ofL
2m
laceia the
unctions of the basal ganglia is tremulous jaw movemeremulous jaw movements are rapid vertical deflectf the lower jaw that have many of the characteristicarkinsonian tremor (Salamone et al., 1998). The pharmaological induction and suppression of tremulous jaw ments is characterized by an acetylcholine/DA interac
hat is similar to that seen in parkinsonism (for revieeeSalamone et al., 1998, 2001). These movements ce induced by striatal DA depletions (Jicha and Salamon991; Finn et al., 1997a), pharmacological depletionA (Steinpreis and Salamone, 1993; Salamone and Ba996), and by acute or sub-chronic administration of DA
agonists (Steinpreis et al., 1993; Steinpreis and Salam993; Trevitt et al., 1999; Wisniecki et al., 2003; Correl., 2004). The jaw movements induced by DA antagonnd DA depletion can be reduced by antiparkinsonianholinergic drugs (Steinpreis and Salamone, 1993; Salamnd Baskin, 1996). Cholinomimetics, which are known to iuce or exacerbate parkinsonian symptoms such as tremumans (Ott and Lannon, 1992; Salamone et al., 2001), also
nduce tremulous jaw movements in rats (Salamone et al986, 1990, 1998, 2001). Because cholinomimetic-induc
aw movements in rats can be reduced by various antipaonian drugs (Cousins et al., 1997; Salamone et al., 1998), thisehavioral test has been used to study the role of basal gircuitry in motor functions that may be related to parkinian tremor. Several studies have implicated SNr GAB
he regulation of tremulous jaw movements. The suppref cholinomimetic-induced tremulous jaw movementsas produced by the D1 agonist SKF 82958 was compl
eversed by infusion of the GABAA antagonist bicucullin
n SNr samples using microdialysis methods. SNr hashown to be rich in GABA transporters, particularly ofAT-1 and GAT-3 subtypes (Ikegaki et al., 1994; Wang anng, 1999). Moreover, research with SNr synaptosomes
hown that GABA transport in SNr can be inhibited by a vty of compounds, including�-alanine (Bahena-Trujillo andrias-Montano, 1999). Thus, it was hypothesized that s
emic administration of�-alanine should reduce pilocarpinnduced tremulous jaw movements, and that a behaviouctive dose of�-alanine should also elevate levels of GA
n SNr dialysate samples.
. Materials and methods
.1. Animals
Male Sprague–Dawley rats (Harlan Sprague Dawleyianapolis, IN) weighing 280–330 g were used (totaln= 20).he rat colony was maintained at 23◦C with a 12-h/12-h
ight/dark cycle (lights on at 07:00 h). Animal protocols wpproved by the Institutional Animal Care Committee,ere in accordance with the Guide for the Care and Usaboratory Animals (National Research Council).
.2. Observations of pilocarpine-induced tremulous jawovements
Observations of animals in the first experiment took pn a clear Plexiglas chamber (30 cm× 30 cm× 30 cm) withwire mesh floor. The chamber was elevated 42 cm from

K. Ishiwari et al. / Journal of Neuroscience Methods 140 (2004) 39–46 41
bottom of the table top, which allowed for the viewing ofthe animal from several angles. Tremulous jaw movementswere defined as rapid vertical deflections of the lower jaw thatresemble chewing but are not directed at any particular stim-ulus (Salamone et al., 1998). Each individual deflection ofthe jaw was recorded using a mechanical hand counter, by anobserver who was blind to the experimental condition of theanimal being observed. Separate studies with two observersdemonstrated an interrater reliability ofr = 0.96 (p < 0.05)using these methods. For assessment of tremulous jaw move-ments, rats received injections of 4.0 mg/kg pilocarpine, andwere observed during a 5-min period beginning 10 min afterpilocarpine injection.
2.3. Implantations of dialysis cannulae and probes
For cannulae implantations in the dialysis experiment, ratswere anesthetized with a solution (1.0 ml/kg, i.p.) that con-tained ketamine (100 mg/ml) and xylazine (20 mg/ml). Theincisor bar on the stereotax was set to 5.0 mm above the in-teraural line. Rats received unilateral implantation (with sidebalanced across rats) of a 10-mm guide cannula purchasedfrom Bioanalytical Systems, Inc. (BAS; West Lafayette,IN). All guide cannula were placed 2.0 mm above the ros-tral SNr at the following coordinates: AP−3.0 mm (fromb -t uredt ic ce-m can-n ior toi n of1 tion.E ctives r in-s nula,w im-p lenet F)w te of2
2
hro-m SA,N sentsiC ual-p lumn( e-t itions etec-t V.P us-i le-
ophile. GABA, OPA, and sulfite react to form a stable, elec-trochemically active GABA-isoindole sulfonate derivative.Fifty microliters of dialysate or a standard solution (10−7 MGABA), and 20�l derivatizing reagent (22.0 mg OPA dis-solved in 0.5 ml ethanol, with 0.5 ml 0.0313 M sodium sul-fite and 9.0 ml 0.1 M sodium tetraborate), were mixed andallowed to react for 15 min. The sample was then assayed us-ing HPLC [mobile phase = 100 mM sodium phosphate buffer,8% methanol, 10.0�l 4.0 M EDTA, set to pH 4.9; flow rate= 1.0 ml/min]. After the elution of GABA, an injection of40�l ethanol was made to remove late-eluting peaks. Stan-dards of GABA were assayed before and after the dialysissamples.
2.5. Experiment 1: effects of�-alanine onpilocarpine-induced jaw movements
A group of rats (n = 13) was used to assess the effects ofacute�-alanine injection on pilocarpine-induced jaw move-ments. Animals were tested once a week for 3 weeks. Ontest days each animal received an injection of 4.0 mg/kg pi-locarpine (10 min before testing) to induce tremulous jawmovements, as well as a dose of�-alanine or vehicle 20 minbefore testing. The following doses of�-alanine were used:vehicle, 250.0 and 500.0 mg/kg;�-alanine was dissolved in0 seda reat-m perw atsw pinei for5 ec-t ve-m in-j be-c menta
2G
wass hiss BAS;W ysiss ingp MN ht esw ddi-t g/kg� ABAc icald s col-l anes-t ts in
regma), LM±1.8 mm (from midline), DV−7.2 mm (venral from skull surface). The guide cannulae were seco the skull using stainless-steel screws and cranioplastent. A stainless-steel stylet was inserted through eachula to ensure its patency throughout the experiment. Pr
nsertion of the dialysis probe, rats were given an injectio.5 mg/kg acepromazine maleate to produce mild sedaach rat then had a concentric dialysis probe (2.0 mm aurface; BAS) inserted through the guide cannula. Afteertion, the probe extended 2.0 mm beyond the guide canith the active surface located in the SNr. At the time oflantation, the dialysis probe was attached via polyethy
ubing to a fluid swivel. Artificial cerebrospinal fluid (aCSas continuously perfused through the tubing at a ra.0�l/min.
.4. Neurochemical analysis of GABA
GABA was assayed using high-performance liquid catography (HPLC) and electrochemical detection (Eew Bedford, MA). The HPLC methods used for the pretudy were originally developed byTrevitt et al. (1999), us-ng a modification of the method published byPhillips andox (1997). The HPLC system comprised a Waters diston pump, a precolumn filter, a reverse-phase coVelosep, C-18, 3 mm× 100 mm), an electrochemical dector (Coulochem II model 5014A), and a data acquistation (ESA). Detector parameters were as follows: dor 1 = +0.35 V, detector 2 = +0.60 V, guard cell = +0.65re-injection derivatization of GABA was accomplished
ng o-phthalidialdehyde (OPA) with sulfite as the nuc
.3% tartaric acid, and this vehicle solution also was us the vehicle control treatment. Each rat received all tents in a randomly varied order, with one injectioneek. All drugs were administered via i.p. injection. Rere placed in the observation chamber after pilocar
njection for a 10-min habituation, and were observedmin during the period 10–15 min after pilocarpine inj
ion. Pilocarpine injections generally produce jaw moents during a period ranging from 5 to 45 min after
ection, and the 10–15 min time period was selectedause this represents the period of maximal jaw movectivity.
.6. Experiment 2: effects of�-alanine on extracellularABA
As described above, a single unilateral guide cannulaurgically implanted above SNr. After recovery from turgery (7–10 days), rats had dialysis probes inserted (est Lafayette, IN) through the guide cannulae. Dial
amples were collected every 30 min on the day followrobe implantation. Artificial cerebrospinal fluid (147.2 maCl, 1.2 mM CaCl2, 4.0 mM KCl) was pumped throug
he probe at a rate of 2.0�l/min. Three baseline samplere conducted prior to drug injection, and then four a
ional samples were collected after injection of 500.0 m-alanine. Samples were frozen and later analyzed for Gontent using reverse-phase HPLC with electrochemetection as described above. After the last sample wa
ected, the probes were removed and the rats werehetized and perfused. Only rats with verified placemen
42 K. Ishiwari et al. / Journal of Neuroscience Methods 140 (2004) 39–46
SNr were used for statistical analysis of the neurochemistrydata (n = 7).
2.7. Histology
Several days after each dialysis session, each animal wasanesthetized with CO2 and perfused with physiological salinefollowed by a 10% formalin solution. The brains were re-moved and stored in formalin, and then were cut with a cryo-stat in 50-�m slices, which were mounted on glass micro-scope slides. Following mounting, slides were stained withCresyl Violet, and cannulae placements were determined us-ing a microscope.
2.8. Data analysis
For the first experiment, tremulous jaw movement datawere analyzed using repeated measures analysis of variance(ANOVA). Planned comparisons using the overall error termwere used to assess the differences between each dose andthe control condition, which kept the total number of com-parisons to the number of conditions minus one (Keppel,1982; pp. 106–124). For the dialysis experiment, repeatedmeasures ANOVA was used to analyze the neurochemicaldata, and planned comparisons were conducted to comparee e.
3
3p
itor� ucedt dt alleP cantd0
3G
,a ent 1t asedd re-i nif-i BAl in-d reds dc e-m
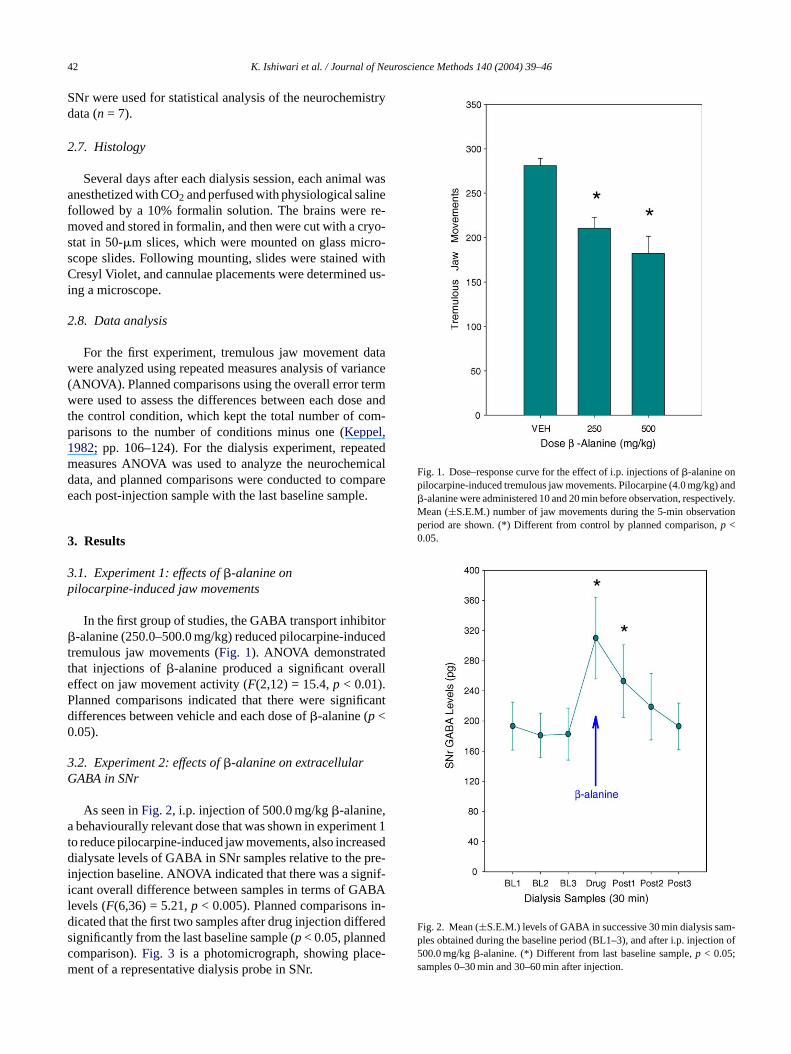
Fig. 1. Dose–response curve for the effect of i.p. injections of�-alanine onpilocarpine-induced tremulous jaw movements. Pilocarpine (4.0 mg/kg) and�-alanine were administered 10 and 20 min before observation, respectively.Mean (±S.E.M.) number of jaw movements during the 5-min observationperiod are shown. (*) Different from control by planned comparison,p <0.05.
Fig. 2. Mean (±S.E.M.) levels of GABA in successive 30 min dialysis sam-ples obtained during the baseline period (BL1–3), and after i.p. injection of500.0 mg/kg�-alanine. (*) Different from last baseline sample,p < 0.05;samples 0–30 min and 30–60 min after injection.
ach post-injection sample with the last baseline sampl
. Results
.1. Experiment 1: effects of�-alanine onilocarpine-induced jaw movements
In the first group of studies, the GABA transport inhib-alanine (250.0–500.0 mg/kg) reduced pilocarpine-ind
remulous jaw movements (Fig. 1). ANOVA demonstratehat injections of�-alanine produced a significant overffect on jaw movement activity (F(2,12) = 15.4,p < 0.01).lanned comparisons indicated that there were signifiifferences between vehicle and each dose of�-alanine (p <.05).
.2. Experiment 2: effects of�-alanine on extracellularABA in SNr
As seen inFig. 2, i.p. injection of 500.0 mg/kg�-alaninebehaviourally relevant dose that was shown in experim
o reduce pilocarpine-induced jaw movements, also increialysate levels of GABA in SNr samples relative to the p
njection baseline. ANOVA indicated that there was a sigcant overall difference between samples in terms of GAevels (F(6,36) = 5.21,p < 0.005). Planned comparisonsicated that the first two samples after drug injection diffeignificantly from the last baseline sample (p< 0.05, planneomparison).Fig. 3 is a photomicrograph, showing placent of a representative dialysis probe in SNr.

K. Ishiwari et al. / Journal of Neuroscience Methods 140 (2004) 39–46 43
Fig. 3. Photomicrograph of Nissl-stained coronal section through midbrain showing placements of dialysis probe in SNr of a representative rat. SNc (substantianigra pars compacta), SNr (substantia nigra pars reticulata), cp (cerebral peduncle). Calibration bar = 0.5 mm.
4. Discussion
The results of the first experiment demonstrated that i.p.injection of the GAT inhibitor�-alanine reduced pilocarpine-induced tremulous jaw movements. Both doses of�-alanine(i.e., 250.0 and 500.0 mg/kg) produced a significant reduc-tion of jaw movement activity relative to injection of pilo-carpine plus vehicle. Although�-alanine has several phar-macological effects, including actions on other transmitterssuch as glycine (Mori et al., 2002), additional lines of ev-idence suggest that�-alanine is acting on GABA systemsto suppress jaw movement activity. Another GAT inhibitor,NO-711, also was shown recently to suppress pilocarpine-induced tremulous jaw movements (Ishiwari et al., 2003).NO-711 was much more potent than�-alanine (i.e., NO-711 suppressed jaw movements at doses of 2.5–5.0 mg/kg),which is consistent with an action of these drugs on GAT-1 transporters. Also,�-alanine was shown in experiment2 to elevate extracellular GABA at one of the same doses(i.e., 500 mg/kg) that also suppressed jaw movement activ-ity (see discussion below). These observations are consis-tent with previous studies showing that elevation of GABAtransmission within the SNr can reduce cholinomimetic-induced tremulous jaw movements. Intranigral injections ofthe GABAA agonist muscimol, in doses as low as 6 ng,w ents( -t leus,o duc-i ,w ess ells
into SNr also could reduce pilocarpine-induced jaw move-ments (Carlson et al., 2003). Of course, in the present study�-alanine was injected systemically, so it is not clear thatthe SNr is the locus of action for the behavioral effects ofthis drug. For that reason, additional research should inves-tigate the effects of intracranial injections of GAT inhibitorsdirectly into SNr, and should characterize a wider range ofcompounds.
The microdialysis experiment demonstrated that injectionof 500.0 mg/kg�-alanine, a dose which reduced jaw move-ment activity in the first experiment, produced a significant el-evation of GABA in SNr dialysis samples. The drug-inducedincrease in dialysate GABA peaked within the first sampleafter injection, resulting in a mean percentage increase ofmore than 66% relative to the last baseline sample. GABAlevels remained significantly elevated during the second post-injection sample, and thereafter declined back to baseline.These results demonstrate that a behaviorally relevant doseof �-alanine is capable of elevating dialysate levels of GABAin SNr in vivo. A previous microdialysis experiment showedthat GAT inhibitors, including tiagabine (30�M) and NNC05-2045 (100�M), increased dialysate levels of GABA inhippocampus when perfused through the microdialysis probein halothane anesthetized rats (Dalby, 2000). That experi-ment focused upon the hippocampal actions of these drugsb . Thep n el-e forG ichG mi-c archh GAT-
ere shown to block pilocarpine-induced jaw movemFinn et al., 1997b). Moreover, GABAA receptor stimulaion in other brain areas, such as entopeduncular nucr brainstem areas dorsal to SNr, was ineffective at re
ng jaw movements (Finn et al., 1997b). More recentlyork with engineered conditionally immortalized cell linhowed that transplantations of GABA-synthesizing c
ecause of their possible utility as anticonvulsant agentsresent study demonstrates that GAT inhibition also cavate GABA levels in SNr, and suggests a potential roleAT inhibitors as tremorolytic agents. It is not clear whAT subtype is involved in the behavioral and neuroche
al effects reported in the present study. Anatomical reseas indicated that SNr has a dense expression of the

44 K. Ishiwari et al. / Journal of Neuroscience Methods 140 (2004) 39–46
1 and GAT-3 GABA transporter subtypes (Ikegaki et al.,1994; Wang and Ong, 1999). Although �-alanine is moreselective for the GAT-3 subtype (Clark and Amara, 1994), itdoes act on GAT-1 as well, and in vitro work indicates that�-alanine can inhibit GAT-1 transport on SNr synaptosomes(Bahena-Trujillo and Arias-Montano, 1999). Although thehigher potency of NO-711 for suppressing tremulous jawmovements relative to�-alanine (Ishiwari et al., 2003) sug-gests that GAT-1 inhibition may be involved in this effect,future studies with a wider range of compounds would benecessary to reach a definitive conclusion about the partic-ular GAT subtype that mediates the behavioral and neuro-chemical effects of�-alanine observed in the present experi-ments. Moreover, additional studies will be necessary to de-termine whether neurons or glia are the source of the increasein dialysate GABA levels that is induced by administrationof various GAT inhibitors.
The present results are consistent with the hypothesisthat GABAergic tone in SNr plays a role in the regu-lation of tremulous jaw movements. The use of micro-dialysis methods for studying extracellular GABA maylead to a better understanding of how diverse motor pro-cesses such as tremor, locomotion and skilled voluntarymovements are modulated by SNr GABA mechanisms(Timmerman and Westerink, 1995; Biggs et al., 1995; Morarie a-t rreae BAi ofb rgict pal-l 6;W ber,1r tionst nists,D ularG hea s fort neo mp-t
A
Na-t theN
R
B idarma-
Neu-
Baumeister AA, Hawkins MF, Anderson-Moore LL, Anticich TG, Hig-gins TD, Griffin P. Effects of bilateral injection of GABA into thesubstantia nigra on spontaneous behavior and measures of analgesia.Neuropharmacology 1988;27:817–21.
Bevan MD, Smith AD, Bolam JP. The substantia nigra as a site of synap-tic integration of functionally diverse information arising from theventral pallidum and the globus pallidus in the rat. Neurosci Lett1996;75:5–12.
Bianchi L, Colivicchi MA, Bolam JP, Della Corte L. The release ofamino acids from rat neostriatum and substantia nigra in vivo:a dual microdialysis probe analysis. Neuroscience 1998;87:171–80.
Biggs CS, Fowler LJ, Whitton PS, Starr MS. Impulse-dependent andtetrodotoxin-sensitive release of GABA in the rat’s substantia nigraas measured by microdialysis. Brain Res 1995;684:172–8.
Bolam JP, Hanley JJ, Booth PA, Bevan MD. Synaptic organization of thebasal ganglia. J Anat 2000;196:527–42.
Bowery NG, Hudson AL, Price GW. GABAA and GABAB receptor sitedistribution in the rat central nervous system. Neuroscience 1987;20:365–83.
Carlson BB, Behrstock SP, Tobin A, Salamone JD. Brain implantations ofengineered GABA-releasing cells suppress tremor in an animal modelof Parkinsonism. Neuroscience 2003;119:927–32.
Clark JA, Amara SG. Stable expression of a neuronal gamma-aminobutyric acid transporter, GAT-3, in mammalian cells demon-strates unique pharmacological properties and ion dependence. MolPharmacol 1994;46:550–7.
Cools AR, Jaspers R, Kolasiewicz W, Sontag KH, Wolfarth S. Substantianigra as a station that not only transmits, but also transforms, in-coming signals for its behavioural expression: striatal dopamine and
Brain
C igraverience
C J,omo-
operi-Res
C entsf an-
D ofogy
D nglia
D gralmine
F The215–
F la-989–
F nnerrgicnd itsEur J
F tic ofl stri-:243–
t al., 1996; O’Connor, 1998; Bianchi et al., 1998; Muszewich and Yamamoto, 1999; Trevitt et al., 2002; Cot al., 2003). Research on the motor functions of SNr GA
s particularly important in view of the various modelsasal ganglia circuitry that emphasize the role of GABAe
ransmission in output structures such as medial globusidus and SNr (Delong, 1990; Gerfen, 1992; Vila et al., 199
ichmann and DeLong, 1996; Ferre et al., 1996; Hau998; Wichmann et al., 1999; Obeso et al., 2000). Additionalesearch will be necessary to study the effects of condihat induce parkinsonian symptoms, such as DA antagoA depletions and cholinomimetic drugs, on extracellABA in SNr. Moreover, it will be important to evaluate tctions of antiparkinsonian agents from various classe
heir potential actions on nigral GABA. Ultimately, this lif research may help to elucidate how parkinsonian sy
oms are modulated by SNr GABA transmission.
cknowledgements
This work was supported by grants to J.S. from theional Institute of Neurological Disorders and Stroke, andational Institute of Mental Health.
eferences
ahena-Trujillo R, Arias-Montano JA. [3H] gamma-aminobutyric actransport in rat substantia nigra pars reticulata synaptosomes: phcological characterization and phorbol ester-induced inhibition.rosci Lett 1999;274:119–22.
GABA-mediated responses of pars reticulata neurons. BehavRes 1983;7:39–49.
orrea M, Mingote SM, Wisniecki A, Salamone JD. Substantia npars reticulata GABA is involved in the regulation of operant lepressing: pharmacological and microdialysis studies. Neurosc2003;119:759–66.
orrea M, Wisniecki A, Betz A, Dobson DR, O’Neill MF, O’Neill Met al. The adenosine A2A antagonist KF 17837 reverses the loctor suppression and tremulous jaw movements induced by haldol in rats: possible relevance to parkinsonism. Behav Brain2004;148:47–54.
ousins MS, Carriero DL, Salamone JD. Tremulous jaw moveminduced by the acetylcholinesterase inhibitor tacrine: effects otiparkinsonian drugs. Eur J Pharmacol 1997;322:137–45.
alby NO. GABA-level increasing and anticonvulsant effectsthree different GABA uptake inhibitors. Neuropharmacol2000;39:2399–407.
elong MR. Primate models of movement disorders of basal gaorigin. Trends Neurosci 1990;13:281–5.
i Chiara G, Morelli M, Porceddu ML, Gessa GL. Evidence that niGABA mediates behavioral responses elicited by striatal dopareceptor stimulation. Life Sci 1998;23:2045–52.
allon JH, Laughlin SE. Substantia nigra. In: Paxinos G, editor.rat nervous system. San Diego: Academic Press; 1995. p.37.
aull RL, Mehler WR. The cells of origin of nigrotectal, nigrothamic and nigrostriatal projections in the rat. Neuroscience 1978;3:1002.
erre S, O’Connor WT, Svenningsson P, Bjorklund L, Lindberg J, TiB, et al. Dopamine D1 receptor mediated facilitation of GABAeneurotransmission in the rat striatoentopeduncular pathway amodulation by adenosine A1 receptor-mediated mechanisms.Neurosci 1996;8:1545–53.
inn M, Jassen A, Baskin P, Salamone JD. Tremulous characterisvacuous jaw movements induced by pilocarpine and ventrolateraatal dopamine depletions. Pharmacol Biochem Behav 1997a;579.

K. Ishiwari et al. / Journal of Neuroscience Methods 140 (2004) 39–46 45
Finn M, Mayorga AJ, Salamone JD. Involvement of pallidal and nigralGABA mechanisms in the generation of tremulous jaw movements inrats. Neuroscience 1997b;80:535–44.
Gerfen CR. The neostriatal mosaic: multiple levels of organization in thebasal ganglia. Ann Rev Neurosci 1992;15:285–320.
Hauber W. Involvement of basal ganglia transmitter systems in movementinitiation. Prog Neurobiol 1998;56:507–40.
Ikegaki N, Saito N, Hashima M, Tanaka C. Production of spe-cific antibodies against GABA transporter subtypes (GAT1, GAT2,GAT3) and their application to immunocytochemistry. Mol Brain Res1994;26:47–54.
Ishiwari K, Carlson BB, Correa M, Mingote SM, Arizzi MN, SalamoneJD. Behavioral and microdialysis studies of GABA in substantia nigrapars reticulata: effects of GABA uptake inhibition in an animal modelof parkinsonian tremor. Soc Neurosci Abstr 2003.
Jicha GA, Salamone JD. Vacuous jaw movements and feeding deficitsin rats with ventrolateral striatal dopamine depletion: possiblerelation to parkinsonian symptoms. J Neurosci 1991;11:3822–9.
Keppel G. Design and analysis: a researcher’s handbook. EnglewoodCliffs, NJ: Prentice-Hall; 1982.
Kha HT, Finkelstein DI, Tomas D, Drago J, Pow DV, Horne MK. Projec-tions from the substantia nigra pars reticulata to the motor thalamus ofthe rat: single axon reconstructions and immunohistochemical study.J Comp Neurol 2001;440:20–30.
Koch M, Fendt M, Kretschmer BD. Role of substantia nigra pars reticulatain sensorimotor gating, measured by prepulse inhibition of startle inrats. Behav Brain Res 2000;117:153–62.
Loopjuit LD, Van der Kooy D. Organization of the striatum: collateral-ization of its efferent axons. Brain Res 1985;348:86–99.
M by
M Stri-ffectsrats.
M cum-erantchem
M naludiedstri-er-:89–
M ousysiol
O a J,son’s
O udied
O eu-
P tput.
P oni-Res
P mma-im-
lec-
S thod-tivity
of nucleus accumbens dopamine. J Neurosci Methods 1996;64:137–49.
Salamone JD, Lalies MD, Channell SL, Iversen SD. Behavioural andpharmacological characterization of the mouth movements induced bymuscarinic agonists in the rat. Psychopharmacology 1986;88:467–71.
Salamone JD, Johnson CJ, McCullough LD, Steinpreis RE. Lateral striatalcholinergic mechanisms involved in oral motor activities in the rat.Psychopharmacology 1990;102:529–34.
Salamone JD, Baskin P. Vacuous jaw movements induced by acute re-serpine and low-dose apomorphine: possible model of parkinsoniantremor. Pharmacol Biochem Behav 1996;53:179–83.
Salamone JD, Cousins MS, McCullough LD, Carriero DL, Berkowitz RL.Nucleus accumbens dopamine release increases during instrumentallever pressing for food but not food consumption. Pharmacol BiochemBehav 1994;49:25–31.
Salamone JD, Cousins MS, Snyder BJ. Behavioral functions of nucleusaccumbens dopamine: Empirical and conceptual problems with theanhedonia hypothesis. Neurosci Biobehav Rev 1997;21:341–59.
Salamone JD, Mayorga AJ, Trevitt JT, Cousins MS, Conlan A, Nawab A.Tremulous jaw movements in rats: a model of Parkinsonian tremor.Prog Neurobiol 1998;56:591–611.
Salamone JD, Correa M, Carlson BB, Wisniecki A, Mayorga AJ, Nisen-baum E, et al. Neostriatal muscarinic receptor subtypes involved in thegeneration of tremulous jaw movements in rodents: implications forcholinergic involvement in parkinsonism. Life Sci 2001;68:2584–97.
Scheel-Kruger J, Magelund G, Olianas MC. Role of GABA in the stri-atal output system: globus pallidus, nucleus entopeduncularis, substan-tia nigra and nucleus subthalamicus. Adv Biochem Psychopharmacol1981a;30:165–86.
Scheel-Kruger J, Magelund G, Olianas M. The role of GABA in the basalndeless;
S enceasaltors.
S leusin the
S BA-9–51.
S ducedopo-
S reser-t age
T id inrats:
T den-d be-91.
T r as-. In:itor-3.
T parsects
T JD.nismsl stud-
V al.utyric
atuszewich L, Yamamoto BK. Modulation of GABA releasedopamine in the substantia nigra. Synapse 1999;32:29–36.
ayorga AJ, Trevitt JT, Conlan A, Gianutsos G, Salamone JD.atal and nigral mechanisms involved in the antiparkinsonian eof SKF 82958 (APB): studies of tremulous jaw movements inPsychopharmacology 1999;143:72–81.
cCullough LD, Cousins MS, Salamone JD. The role of nucleus acbens dopamine in responding on a continuous reinforcement opschedule: a neurochemical and behavioral study. Pharmacol BioBehav 1993;46:581–6.
orari M, O’Connor WT, Ungerstedt U, Bianchi C, Fuxe K. Functioneuroanatomy of the nigrostriatal and striatonigral pathways as stwith dual probe microdialysis in the awake rat-II. Evidence foratalN-methyl-d-aspartate receptor regulation of striatonigral GABAgic transmission and motor function. Neuroscience 1996;7297.
ori M, Gahwiler BH, Gerber U.�-alanine and taurine as endogenagonists at glycine receptors in rat hippocampus in vitro. J Ph2002;539:191–200.
beso JA, Rodriguez-Oroz MC, Rodriguez M, Lanciego JL, ArtiedGonzolo N, et al. Pathophysiology of the basal ganglia in Parkindisease. Trends Neurosci 2000;23:S8–19.
’Connor WT. Functional neuroanatomy of the basal ganglia as stby dual-probe microdialysis. Nucl Med Biol 1998;25:743–6.
tt BR, Lannon MC. Exacerbation of parkinsonism by tacrine. Clin Nropharm 1992;15:322–5.
arent A, Hazrati LN. Common structural organization of two ounuclei of primate basal ganglia. Trends Neurosci 1993;16:308–9
arent A, Bouchard C, Smith Y. The striatopallidal and striatgral projections: two distinct fiber systems in primate. Brain1984;303:385–90.
hillips JB, Cox BM. Release of endogenous glutamate and gaaminobutyric acid from rat striatal tissue slices measured by anproved method of high-performance liquid chromatography with etrochemical detection. J Neurosci Methods 1997;75:207–14.
alamone JD. The behavioral neurochemistry of motivation: meological and conceptual issues in studies of the dynamic ac
ganglia and limbic system for behaviour. In: DeFeudis FV, MaP, editors. Amino Acid Neurotransmitters. New York: Raven Pr1981b.
cheel-Kruger J. The GABA receptors and animal behaviour: evidthat GABA transmits and mediates dopaminergic function in the bganglia and limbic system. In: Enna SJ, editor. The GABA recepClifton, NJ: The Humana Press; 1983. p. 215–56.
okolowski JD, Conlan A, Salamone JD. A microdialysis study of nucaccumbens core and shell dopamine during operant respondingrat. Neuroscience 1998;86:1001–9.
tarr MS, Starr BS. Circling evoked by intranigral SKF 38393: a GAmediated D-1 response? Pharmacol Biochem Behav 1989;32:84
teinpreis RE, Baskin P, Salamone JD. Vacuous jaw movements inby sub-chronic administration of haloperidol: interactions with sclamine. Psychopharmacology 1993;111:99–105.
teinpreis RE, Salamone JD. Effects of acute haloperidol andpine administration on vacuous jaw movements in three differengroups of rats. Pharmacol Biochem Behav 1993;46:405–9.
immerman W, Westerink BH. Extracellular gamma-aminobutyric acthe awake substantia nigra measured by microdialysis in awakeeffects of various stimulants. Neurosci Lett 1995;197:21–4.
immerman W, Abercrombie ED. Amphetamine-induced release ofdritic dopamine in substantia nigra pars reticulata: D1-mediatehavioral and electrophysiological effects. Synapse 1996;23:280–
revitt JT, Carlson BB, Salamone JD. Electrochemical methods fosessing extracellular GABA in substantia nigra pars reticulataRollema H, Abercrombie E, Dulzer D, Zackheim J, editors. Moning molecules in neuroscience. Newark, NJ: Rutgers; 1999. p. 9
revitt JT, Carlson BB, Nowend K, Salamone JD. Substantia nigrareticulata is a highly potent site of action for the behavioral effof SCH 23390. Psychopharmacology 2001;156:32–41.
revitt JT, Carlson BB, Correa M, Keene A, Morales M, SalamoneInteractions between dopamine D1 receptors and GABA mechain substantia nigra pars reticulata: neurochemical and behavioraies. Psychopharmacology 2002;159:229–37.
ila M, Herrero MT, Levy R, Faucheux B, Ruberg M, Guillen J, etConsequences of nigrostriatal denervation on the gamma-aminob

46 K. Ishiwari et al. / Journal of Neuroscience Methods 140 (2004) 39–46
acidic neurons of substantia nigra pars reticulata and superior collicu-lus in parkinsonian syndromes. Neurology 1996;46:802–9.
Wang XS, Ong WY. A light and electron microscopic study of GAT-1 inthe monkey basal ganglia. J Neurocytol 1999;28:1053–61.
Wichmann T, DeLong MR. Functional and pathophysiological models ofbasal ganglia. Curr Opin Neurobiol 1996;6:751–8.
Wichmann T, Bergmann H, Starr PA, Subramanian T, Watts RL, DelongMR. Comparison of MPTP-induced changes in spontaneous neuronal
discharge in the internal pallidal segment and in the substantia nigrapars reticulata in primates. Exp Brain Res 1999;125:397–409.
Wichmann T, Kliem MA, DeLong MR. Antiparkinsonian and behavioraleffects of inactivation of the substantia nigra pars reticulata in hemi-parkinsonian primates. Exp Neurol 2001;167:410–24.
Wisniecki A, Correa M, Arizzi MN, Ishiwari K, Salamone JD. Motor ef-fects of GABAa antagonism in globus pallidus: Studies of locomotionand tremulous jaw movements. Psychopharmacology 2003;170:140–9.
Related Documents











