The first terrestrial isopod (Crustacea: Isopoda: Oniscidea) from Cretaceous Burmese amber of Myanmar Pierre Broly a, * ,S ebastien Maillet b , Andrew J. Ross c, * a Unit ed’Ecologie Sociale, Universit e Libre de Bruxelles, Campus de la Plaine, Bruxelles, Belgium b Universit e Lille1 des Sciences et Technologies, UMR 8217 CNRS G eosyst emes, B^ at. SN5, Avenue Paul Langevin, 59 655, Villeneuve-d’Ascq, France c Department of Natural Sciences, National Museum of Scotland, Chambers St. Edinburgh EH1 1JF, UK article info Article history: Received 23 December 2014 Accepted in revised form 28 February 2015 Available online Keywords: Myanmar Burmese amber Synocheta Woodlice Fossil New species Palaeobiogeography abstract This paper represents the first formal description of a Cretaceous terrestrial isopod (Oniscidea). This unique specimen, a well preserved female in Burmese amber from Myanmar, is described as Myan- mariscus deboiseae gen. nov. sp. nov. It belongs to the clade Synocheta Legrand, 1946 based on the poorly differentiated flagellum of antenna, its small body size and underdeveloped eyes. Its familial position remains uncertain because this group is primarily classified using male reproductive struc- tures. However, comparison with the biogeographical distribution of Recent species of Oniscidea suggests it most probably belongs to the Styloniscidae Vandel, 1952. This specimen and other records of Oniscidea in Cretaceous amber from East Europe indicate that this remarkable group of crustaceans was already highly diversified by the Cretaceous, thus supporting an earlier origin for the group. Finally, this strictly terrestrial Mesozoic isopod represents the earliest formal record of land coloni- zation within Crustacea. © 2015 Elsevier Ltd. All rights reserved. 1. Introduction Within Crustacea, the Oniscidea (Isopoda) are remarkable or- ganisms due to their ability to live on land (Hornung, 2011). Currently, terrestrial isopods, commonly named woodlice, are one of the major groups of soil macrofauna (e.g. Gongalsky et al., 2005; Paoletti & Hassall, 1999; Hornung et al., 2007; Podgaiski et al., 2007) and have an important role in soil process and soil ecosys- tems, particularly as litter decomposers (e.g. Wieser, 1978; Hassall et al., 1987; Kautz & Topp, 2000; Zimmer, 2002; Quadros & Araujo, 2008). Today their distribution is cosmopolitan, extending from the supralittoral level to hot deserts and high altitudes (Vandel, 1960; Warburg, 1993; Linsenmair, 2007). They are partic- ularly common in forest leaf litters, grasslands and sites managed by humans, e.g. gardens. For a long time, terrestrial isopods (Oniscidea) were believed to be a relatively recent group probably because, despite the relatively large size of the suborder (over 3600 species; Schmalfuss, 2003), the fossil record of Oniscidea is quite scarce and, until recently, was restricted to the Cenozoic (Broly et al., 2013). Woodlice are crustaceans, so their exoskeleton is mainly composed of calcium carbonate (CaCO 3 )(Becker et al., 2005; Neues et al., 2007; Hild et al., 2008) with minor amounts of materials resistant to chemi- cal and physical degradation, in particular chitin or sclerotized structures (Girling, 1979; and see Briggs, 1999; Gupta & Briggs, 2011). Therefore, the probability of burial and fossilization in the continental realm is low for these terrestrial crustaceans. However, one particular material can lead to a good preserva- tion of ancient woodlice, and that is amber. Currently, the major part of the oniscidean fossil record comes from Cenozoic amber (Broly et al., 2013). Sometimes, the biomineralized cuticle may be superficially degraded by acidic compounds of fresh resins (Schmalfuss, 1984a), however many ambers, such as Rovno, Baltic, Bitterfeld and Dominican amber have well preserved woodlice through time and several species have been described (Spahr, 1993; Schmalfuss, 1980, 1984a). The regular capture of macroscopic invertebrates in resin began in the Early Cretaceous, with only one record of this known before then (Schmidt et al., 2012). This probably explains why the start of the oniscidean fossil record is only known from Early Cretaceous amber (Aptian-Cenomanian) while a Carboniferous origin of this group is assumed (Broly et al., 2013). To our knowledge, Cretaceous amber with woodlice inclusions is only known from three countries * Corresponding authors. E-mail addresses: [email protected] (P. Broly), [email protected] (A.J. Ross). Contents lists available at ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/CretRes http://dx.doi.org/10.1016/j.cretres.2015.02.012 0195-6671/© 2015 Elsevier Ltd. All rights reserved. Cretaceous Research 55 (2015) 220e228

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Cretaceous Research 55 (2015) 220e228
Contents lists avai
Cretaceous Research
journal homepage: www.elsevier .com/locate/CretRes
The first terrestrial isopod (Crustacea: Isopoda: Oniscidea) fromCretaceous Burmese amber of Myanmar
Pierre Broly a, *, S�ebastien Maillet b, Andrew J. Ross c, *
a Unit�e d’Ecologie Sociale, Universit�e Libre de Bruxelles, Campus de la Plaine, Bruxelles, Belgiumb Universit�e Lille1 des Sciences et Technologies, UMR 8217 CNRS G�eosyst�emes, Bat. SN5, Avenue Paul Langevin, 59 655, Villeneuve-d’Ascq, Francec Department of Natural Sciences, National Museum of Scotland, Chambers St. Edinburgh EH1 1JF, UK
a r t i c l e i n f o
Article history:Received 23 December 2014Accepted in revised form 28 February 2015Available online
Keywords:MyanmarBurmese amberSynochetaWoodliceFossilNew speciesPalaeobiogeography
* Corresponding authors.E-mail addresses: [email protected] (P. Broly)
http://dx.doi.org/10.1016/j.cretres.2015.02.0120195-6671/© 2015 Elsevier Ltd. All rights reserved.
a b s t r a c t
This paper represents the first formal description of a Cretaceous terrestrial isopod (Oniscidea). Thisunique specimen, a well preserved female in Burmese amber from Myanmar, is described as Myan-mariscus deboiseae gen. nov. sp. nov. It belongs to the clade Synocheta Legrand, 1946 based on thepoorly differentiated flagellum of antenna, its small body size and underdeveloped eyes. Its familialposition remains uncertain because this group is primarily classified using male reproductive struc-tures. However, comparison with the biogeographical distribution of Recent species of Oniscideasuggests it most probably belongs to the Styloniscidae Vandel, 1952. This specimen and other records ofOniscidea in Cretaceous amber from East Europe indicate that this remarkable group of crustaceanswas already highly diversified by the Cretaceous, thus supporting an earlier origin for the group.Finally, this strictly terrestrial Mesozoic isopod represents the earliest formal record of land coloni-zation within Crustacea.
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
Within Crustacea, the Oniscidea (Isopoda) are remarkable or-ganisms due to their ability to live on land (Hornung, 2011).Currently, terrestrial isopods, commonly named woodlice, are oneof the major groups of soil macrofauna (e.g. Gongalsky et al., 2005;Paoletti & Hassall, 1999; Hornung et al., 2007; Podgaiski et al.,2007) and have an important role in soil process and soil ecosys-tems, particularly as litter decomposers (e.g. Wieser, 1978; Hassallet al., 1987; Kautz & Topp, 2000; Zimmer, 2002; Quadros &Araujo, 2008). Today their distribution is cosmopolitan, extendingfrom the supralittoral level to hot deserts and high altitudes(Vandel, 1960; Warburg, 1993; Linsenmair, 2007). They are partic-ularly common in forest leaf litters, grasslands and sites managedby humans, e.g. gardens.
For a long time, terrestrial isopods (Oniscidea) were believed tobe a relatively recent group probably because, despite the relativelylarge size of the suborder (over 3600 species; Schmalfuss, 2003),the fossil record of Oniscidea is quite scarce and, until recently, wasrestricted to the Cenozoic (Broly et al., 2013). Woodlice are
, [email protected] (A.J. Ross).
crustaceans, so their exoskeleton is mainly composed of calciumcarbonate (CaCO3) (Becker et al., 2005; Neues et al., 2007; Hildet al., 2008) with minor amounts of materials resistant to chemi-cal and physical degradation, in particular chitin or sclerotizedstructures (Girling, 1979; and see Briggs, 1999; Gupta & Briggs,2011). Therefore, the probability of burial and fossilization in thecontinental realm is low for these terrestrial crustaceans.
However, one particular material can lead to a good preserva-tion of ancient woodlice, and that is amber. Currently, the majorpart of the oniscidean fossil record comes from Cenozoic amber(Broly et al., 2013). Sometimes, the biomineralized cuticle may besuperficially degraded by acidic compounds of fresh resins(Schmalfuss, 1984a), however many ambers, such as Rovno, Baltic,Bitterfeld and Dominican amber have well preserved woodlicethrough time and several species have been described (Spahr, 1993;Schmalfuss, 1980, 1984a).
The regular capture of macroscopic invertebrates in resin beganin the Early Cretaceous, with only one record of this known beforethen (Schmidt et al., 2012). This probably explains why the start ofthe oniscidean fossil record is only known from Early Cretaceousamber (Aptian-Cenomanian) while a Carboniferous origin of thisgroup is assumed (Broly et al., 2013). To our knowledge, Cretaceousamberwithwoodlice inclusions is only known from three countries

P. Broly et al. / Cretaceous Research 55 (2015) 220e228 221
in the world: Myanmar (Hukawng Valley; Ross et al., 2010 and thispaper), Spain (�Alava; Delcl�os et al., 2007) and France (Charente-Maritime; Perrichot, 2004).
There is a large amount of published literature about the di-versity of insects and other successful terrestrial arthropods, suchas spiders, in amber (e.g. Grimaldi & Engel, 2005; Selden & Penney,2010), but very little has been written about crustaceans in amberand especially about woodlice. The evolutionary history of thisgroup is nevertheless of particular importance to better understandthe terrestrialization processes in Crustacea. Woodlice may also beuseful for interpreting palaeoenvironments because of the strongdegree of specialization of the species in terrestrial environments(Edney, 1954; Schmalfuss, 1984b; Warburg, 1993).
Herein, we provide the first formal description of a Cretaceousoniscid, based on a single specimen found in Burmese amber(Cenomanian) from Myanmar. This specimen, a well preserved fe-male, provides important clues for increased knowledge of thediversification and evolutionary history of this remarkable group ofstrictly terrestrial crustaceans. Some notes about the possibleecology of the described species are also provided.
2. Material and methods
Specimen is preserved in a small piece (10 � 9 � 4 mm) ofyellow-coloured Burmese amber, or Burmite. A small fragmentedarthropod co-occurs with the specimen. The piece of amber comesfrom Noije Bum in the Hukawng Valley, northern Myanmar(Burma; Fig. 1). For more details on this locality and its stratigraphy,see Cruickshank & Ko (2003). The amber of this area comes from abed dated as early Cenomanian in age (about 99 Ma; Shi et al.,2012), however the amber itself could be older as it may havetaken a while to be deposited.
The piece containing the oniscid fossil is from the Scott Ander-son Collection, deposited in the National Museums Scotland (NMS),registered as NMS G.2010.20.42.
The specimen was examined under Zeiss Stemi 2000-C andMeiji Techno RZ stereomicroscopes. Figures and drawings weredrawn with the CorelDRAW 12.0 software. It was photographedusing a Wild Makroscope M420 (fitted with Apozoom objective)with Nikon D700 digital SLR camera.
3. Systematic paleontology
Class: Malacostraca Latreille, 1802Order: Isopoda Latreille, 1817Suborder: Oniscidea Latreille, 1802Section: Synocheta Legrand, 1946Family: ?Styloniscidae Vandel, 1952Subfamily: ?Styloniscinae Vandel, 1952Genus Myanmariscus gen. nov.Derivation of name. “Myanmariscus”: fromMyanmar, the countryof origin of the type species.Type species. Myanmariscus deboiseae sp. nov.Diagnosis. A Cretaceous oniscid with the following features: fe-male with elongated body. Pleon narrower than pereon. Ceph-alon and pereionites distinctly tuberculated. Eyes absent.Cephalon with small lateral lobes and vertex delimited by agroove. Flagellum of the antenna with 6 indistinct articles.Sensorial and locomotor appendages with sparse setae. Promi-nent uropods with endopodite slightly shorter than exopodite.Male unknown.Myanmariscus deboiseae sp. nov.(Figs. 2AeD, 3AeG)Derivation of name. “deboiseae”: after Emma De Boise, wife ofAndrew J. Ross.
Holotype.NMS G.2010.20.42. Female specimenwell preserved inthree dimensions, housed in the National Museums CollectionCentre, National Museums Scotland, Edinburgh (NMS). ScottAnderson Collection.Type locality and age.Noije Bum, Hukawng Valley, Myanmar (seeCruickshank& Ko, 2003); early Cenomanian (see Shi et al. 2012).Diagnosis. As for genus (monotypic new genus).Description. Female without marsupium. Maximal dimensions:2.9 mm (L), 1.0 mm (W). Colourless. Elongated body shape withpleon narrower than pereon (see Figs. 2AeB, 3A). Body dorso-laterally compressed, unable to roll into a ball.Trapezoidal cephalon (0.4 � 0.5 mm) with small quadrangularlateral lobes. Eyes absent. Cephalon distinctly granulated; tu-bercles with a quite irregular disposition. Vertex separated fromthe front part of cephalon by a depression (Figs. 2C, 3B).Pereon composed of 7 pereonites with fairly well developedepimera. Cephalon well set into pereonite I. Two (or three)rows of bumps on posterior part of each pereonite. Lateralmargins of pereonites almost horizontal. Dense and stockytriangular scale-setae particularly visible on this part. Per-eonites I to IV with convex hind margins and rounded postero-lateral corners. Pereonites V to VII with concave hind marginswith postero-lateral corners increasingly acute and pointingbackwards. Outline of edges of pereon and pleon with a step-ped line.Pleon composed of 5 smooth pleonites. Pleonites I and II framedby pereonite VII. Pleonites III to V slightly incurved with smallpostero-lateral corners pointing backwards.Telson with broadly rounded apex and slightly concave sides.Antennula not visible. Antenna (Fig. 3C) of 0.7 mm lengthcomposed of a peduncle of 5 articles and a flagellum. The threebasal peduncular articles are relatively short. Peduncular seg-ments more or less spiny. Inner edge of the fifth pedunculararticle serrated. Articles 3 to 5 covered by some setae. Poorlydivided flagellumwith at least 6 indistinct proximal articles andterminated by a fine seta. Flagellum longer than the fifthpeduncular article. Mouthparts not sufficiently visible (andundissectible) to finely discern form and structural organizationof buccal pieces.Pereopods (Fig. 3DeF) simple with sparse setae.Pleopodes poorly distinct (Figs. 2D and 3G). Only exopodites offirst and second pleopods substantially fully observable. Noobservable pleopod endopodites.Uropods prominent. Exopodite of uropod conical, robust in itsbasis and fine in its apical part, terminated by a seta. Endopoditeof uropod almost as long as exopodite (respectively 0.3 mm and0.4 mm) and inserting at the same level.Remarks. Based on the single female available, we established anew genus and a new species within Synocheta despite a lack ofunique morphological characters and especially of the usualdiagnostic male features (see discussion). Therefore, the diag-nosis of this new species does not allow a clear morphologicaldistinction from current species of Synocheta (see discussion),an approach which may be debatable due to the taxonomicconfusion it causes. Comparisonwith female specimens of othermodern Synocheta is difficult because only males are usuallydescribed in the literature. However, as it stands, the specimendescribed herein is of an outstanding importance by its age (65million years older than the next oldest described species) andby the knowledge brings to the understanding of the terrestri-alization process and chronology of the evolution of Isopoda.Conversely, keeping it in open nomenclature is less helpful,especially for discussions of terrestrial isopod phylogeny andorigination. Therefore, because of this, we argue that the spec-imen should be named.

Fig. 1. Location of the studied locality providing amber (star) and distribution of continental blocks constitutive of Myanmar (after Metcalfe, 2013).
P.Brolyet
al./Cretaceous
Research55
(2015)220
e228
222

Fig. 2. Photomicrographs of Myanmariscus deboiseae gen. nov. sp. nov. (NMS G.2010.20.42) in Burmese amber. (A) habitus, dorsal view; (B) habitus, ventral view; (C) anterior part,dorsal view; (D) posterior part, ventral view. Scale bar: 0.5 mm.
P. Broly et al. / Cretaceous Research 55 (2015) 220e228 223
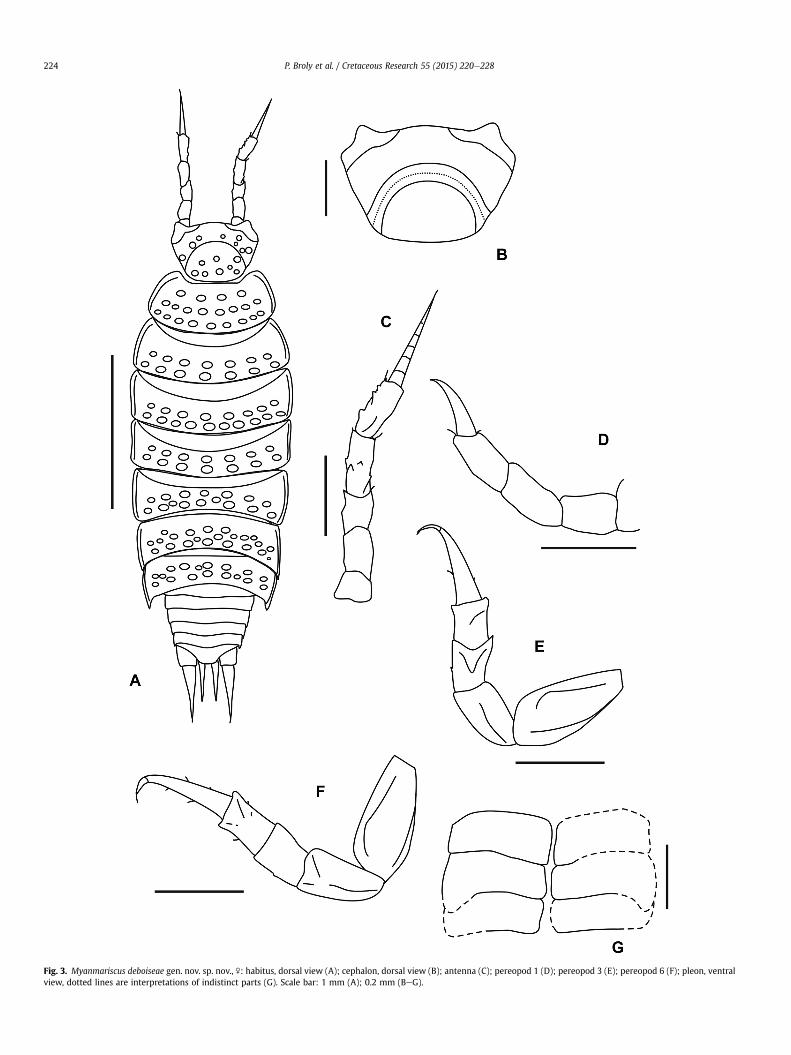
Fig. 3. Myanmariscus deboiseae gen. nov. sp. nov., \: habitus, dorsal view (A); cephalon, dorsal view (B); antenna (C); pereopod 1 (D); pereopod 3 (E); pereopod 6 (F); pleon, ventralview, dotted lines are interpretations of indistinct parts (G). Scale bar: 1 mm (A); 0.2 mm (BeG).
P. Broly et al. / Cretaceous Research 55 (2015) 220e228224

P. Broly et al. / Cretaceous Research 55 (2015) 220e228 225
Habitat. Currently, all the Synocheta are restricted to wet ormoist habitats. The absence of eyes, the small size and the dorsalornamentation of the specimen (tuberculate) suggest a cave oran endogean mode of life as usual for many current Synocheta(Schmalfuss, 1977, 1984b). We exclude the hypothesis of a truecave species which seems highly improbable for getting trappedin amber. More probably, Myanmariscus deboiseae gen. nov. sp.nov was a true soil dwelling species and presented an endogeanway of life in the deeper layers of leaf-litters.
4. Discussion
Myanmariscus deboiseae gen. nov. sp. nov. belongs to theterrestrial isopods (Oniscidea) especially due to the segmentationof the body and its seven pairs of legs.
Themonophyletic group of Oniscidea is divided into five groups:Ligiidae (Diplocheta), Tylidae (Tylida), Mesoniscidae (Microcheta),Synocheta and Crinocheta (Schmalfuss, 2003; Schmidt, 2008).Synocheta and Crinocheta constitute the higher phylogenetic levelswithin Oniscidea (Schmalfuss, 1989; Erhard, 1998; Schmidt, 2008).
Within Oniscidea, Myanmariscus deboiseae can be readilyassigned to the Synocheta Legrand, 1946 according to its weaklyindividualized flagellum on antenna, its small body size, and itsunderdeveloped eyes. Until now, fossils of Synocheta were onlyrecorded up to Eocene (Perkovsky et al., 2010; Weitschat &Wichard, 2010). Currently, the Synocheta group includes about630 species, i.e. approximately 16% of the Oniscidea species(Schmidt, 2008). Members of the group Synocheta present severalapomorphies, including fused male genital ducts in distal part, eyeswith at most 3 ommatidia or articles of the antennal flagellumweakly individualized (Tabacaru & Danielopol, 1996; reviewed inSchmidt, 2008).
The Synocheta group contains currently two large and commonfamilies (Styloniscidae and Trichoniscidae), and three highlyspecialized families (Schoebliidae, Titanidae and Turanoniscidae)(Tabacaru, 1993; Schmalfuss, 2003). Nothing indicates that Syn-ocheta is not a monophyletic group (Schmalfuss, 1989; Tabacaru,1993; Erhard, 1998; Michel-Salzat & Bouchon, 2000; Mattern2003). The phylogenetic relations within the Synocheta are pri-marily based on the structure of the reproductive system, andmoreparticularly on male first pleopods (Vandel, 1952; Tabacaru, 1993;Schmalfuss, 2005). Notably, the families Trichoniscidae and Stylo-niscidae are anatomically very similar; the main, and only surecharacter, allowing the separation of these families is themorphology and development of the muscles of the endopodite ofthe male first pleopods (Vandel, 1952, 1960). Although Vandelrecognised later (1975) that the dichotomous nature of this featureis probably too excessive and restrictive, it is still widely used today(Tabacaru, 1993; Schmalfuss, 2005).
Among Synocheta, the familial assignation of Myanmariscusdeboiseae is problematic. The extant termitophilous familiesSchoebliidae, Titanidae and Turanoniscidae can be readily excludedbecause they are highly specialized taxa presenting non-conformistforms (see Budde-Lund, 1909; Vandel, 1952 and Kensley, 1971 forfigures). Thus,Myanmariscus deboiseaemay be assigned to only oneof two extant families: Trichoniscidae or Styloniscidae. Unfortu-nately, the separation of these two large families is tenuous andessentially based on the male sexual features (see above; Vandel,1952; Tabacaru, 1993; Schmalfuss, 2005). Within Trichoniscidae,two of the three subfamilies can however be readily excluded: theHaplophthalminae due to their highly developed ornamentationand well developed epimera of pleonites 3e5 or 4e5 (Vandel,1960); and the Buddelundiellinae due to their ability to roll into aball (Vandel, 1960). Within Styloniscidae, two of the three
subfamilies can be readily excluded: the Notoniscinae, due to theirtergal ornamentation and well developed epimera of pleonites 3e5or 4e5 (similar to Haplophthalminae; Vandel, 1952); and theKuscheloniscinae (only 1 species), due to their high continuitybetween pereon and pleon and perpendicularity of the sides oftergites (Strouhal, 1961). So this leaves only two potential sub-families for Myanmariscus deboiseae: Trichoniscinae (Trichonisci-dae) and Stlyloniscinae (Styloniscidae). Both subfamilies have asmooth or tuberculated body (without longitudinal ridges) andpleon distinctly narrower than pereon because of the reduction ofthe pleonites 1 to 5, as defined by Vandel (1952). Only the malesexual characters allow the discrimination of these two familieswith certainty (Vandel, 1952, 1960; Tabacaru, 1993; Schmalfuss,2005). So, the female features of the fossil specimen describedherein make any conclusion impossible about its precise taxonomicposition in one of these two families and subfamilies. The poorvisibility of significant morphological details as mouthparts orantennula does not help to solve this problem.
Myanmariscus deboiseae morphologically differs from moderngenera found in the same geographical area, as Thailandoniscus(southern Thailand; Dalens, 1989) and Trogloniscus (southernChina; Taiti & Xue, 2012), respectively due to the smooth dorsumplus number of articles of flagellum (Dalens, 1989) and due tocephalon lobes development plus number of articles of flagellum(Taiti & Xue, 2012). Myanmariscus deboiseae could be more closelyrelated to recent Indoniscus and some species of Styloniscus due todorsum ornamentation and antennal features but species of theselast genera have three ommatidia whereas the fossil specimenappears to lack eyes.
Regarding their current distribution, the synochetan familiespresent a high geographical speciation. Today, the Trichoniscidaedominate all the holarctic zone (Fig. 4; Vandel, 1960; Jass &Klausmeier, 2000; Schmalfuss, 2003). In contrast, the Stylonisci-dae are restricted to a Gondwanan distribution (Fig. 4; Vandel,1952; Ferrara & Taiti, 1979; Leistikow and W€agele, 1999;Schmalfuss, 2003).
Regarding the present day geographical distribution of syn-ochetan families (Fig. 4), we argue in favour of the tentativeplacement of Myanmariscus deboiseae in the Styloniscidae. Inaddition, we can speculate on a Laurasian origin of Trichoniscidaeand a Gondwanan origin of Styloniscidae, after vicariance of acommon pangean population. Palaeogeographical reconstructionsindicate that present-day Southeast Asia is composed of a set ofterranes originated from northwestern Gondwana (Metcalfe, 1998,2013). Myanmar is mainly composed of two Gondwanan terranes:Sibumasu and West Burma. Myanmariscus deboiseae is recordedfrom Noije Bum, located on the West Burma terrane (Fig. 1). Ac-cording to the commonly accepted model, rifting of the WestBurma terrane led to its separation from northwest Australia and toits northwards drifting during Late Triassic to Late Jurassic; theWest Burma terrane finally collided with the Eurasian margin(Sibumasu terrane) during the Late Cretaceous (Metcalfe, 1998;Heine & Müller, 2005; Seton et al., 2012). Another recent modelsuggested a Devonian rifting of the West Burma terrane and aTriassic collision with Eurasia (Metcalfe, 2013). These palaeogeo-graphical reconstructions show that Burma is of gondwanan originand support the assignation of the Cretaceous Burmese amberwoodlouse described herein to the Styloniscidae. However, thisshould be taken with caution: due to the paucity of both Oniscideafossils and phylogenetic data on group divergence, it is impossibleto indisputably assign Myanmariscus deboiseae to Styloniscidae onthis single palaeobiogeographic argument (see Murienne et al.,2014).
For a long time, terrestrial isopods (Oniscidea) were undeserv-edly believed to be a relatively recent group due to their poor fossil
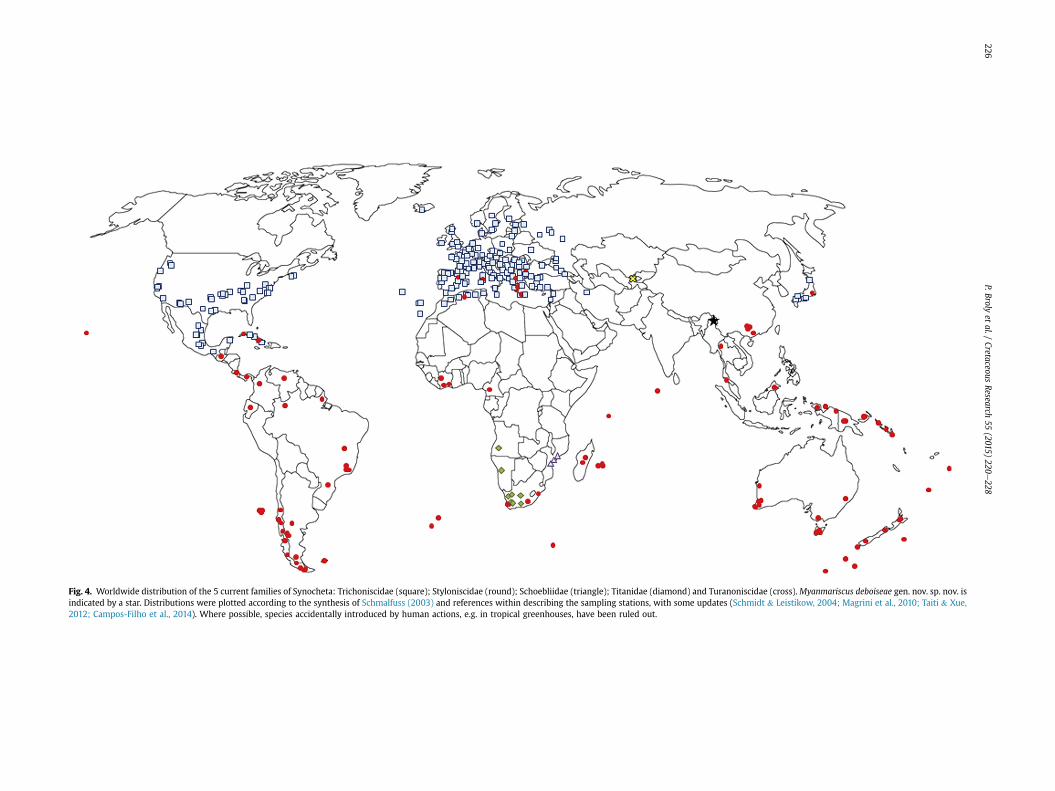
Fig. 4. Worldwide distribution of the 5 current families of Synocheta: Trichoniscidae (square); Styloniscidae (round); Schoebliidae (triangle); Titanidae (diamond) and Turanoniscidae (cross).Myanmariscus deboiseae gen. nov. sp. nov. isindicated by a star. Distributions were plotted according to the synthesis of Schmalfuss (2003) and references within describing the sampling stations, with some updates (Schmidt & Leistikow, 2004; Magrini et al., 2010; Taiti & Xue,2012; Campos-Filho et al., 2014). Where possible, species accidentally introduced by human actions, e.g. in tropical greenhouses, have been ruled out.
P.Brolyet
al./Cretaceous
Research55
(2015)220
e228
226

P. Broly et al. / Cretaceous Research 55 (2015) 220e228 227
record in regard to the whole size of the group (Broly et al., 2013).Because of this, each new discovery provides valuable informationabout the evolutionary history of this group. First observations onCretaceous specimens from Charente (France) suggest the identi-fication of the family Ligiidae (figure inN�eraudeau 2008). Currently,Ligiidae (Diplocheta) constitutes the most basal group in Oniscidea(Schmidt, 2008), with species mostly restricted to supralittoralareas (Carefoot & Taylor, 1995; Taiti et al., 2003). By contrast, theBurmese specimen described herein belongs to Synocheta, thesecond highest phylogenic division within Oniscidea (Schmidt,2008). These results suggest that terrestrial isopods were alreadyhighly diversified and widely distributed on Earth at least from theEarly Cretaceous. These elements argue for an old origin of Onis-cidea, in pre-Pangean times (see Broly et al., 2013).
Except Oniscidea, the greater part of the fossil record of Isopodahas been recorded from aquatic environments. Many current ma-rine species are intertidal (especially in Sphaeromatidae; e.g.Holdich & Jones, 1973), but the only isopods presenting a strictlyterrestrial mode of life belong to the Oniscidea. Some authorsspeculated about the terrestrial origin of few fossil specimens ortraces earlier than the Cretaceous; however, these are preserved insequences from inter/supratidal or shallow-water environments(Upper Jurassic: Gaillard et al., 2005; Uchman et al., 2011; LowerCretaceous: Wieder and Feldmann, 1992) and were probably notcompletely free from the aquatic world. In this respect, the spec-imen described herein constitutes the earliest evidence of fullyterrestrial life in Isopoda. Likewise, it constitutes also the earliestformal evidence of fully terrestrial colonization in Crustacea. Untilnow, the fossil record of strictly terrestrial crustaceans, includingAmphipoda, Isopoda and Decapoda, was recent and only knownfrom the Tertiary (reviewed in Dunlop et al., 2013; Richardson &Araujo, 2015). Therefore, the Cretaceous specimen describedherein considerably improves the calibration of the evolutionaryhistory of this large and ecologically important group ofarthropods.
5. Conclusion
Among the few inclusions reported in Cretaceous deposits(Myanmar: Ross et al., 2010; and present study; Northern Spain:Alonso et al., 2000 and Delcl�os et al., 2007; South Western France:Perrichot, 2004), this study constitutes the first detailed descriptionof an oniscid fossil from the Mesozoic.
The specimen can be readily assigned to the Synocheta based onits small body size, its underdeveloped eyes and its weakly sub-divided flagellum of antenna. Despite a doubtful familial placementon single morphological data, the specimen, formally described asMyanmariscus deboiseae gen. nov. sp. nov., is tentatively assigned toStyloniscidae based upon its geographical origin, the current dis-tribution of Synocheta and palaeogeographic reconstructions.Some notes about the possible endogean mode of life of thedescribed species are also provided.
Myanmariscus deboiseae gen. nov. sp. nov. is of outstandingimportance in understanding the evolutionary history of Oniscidea.The relative high phylogenetic position of Synocheta within Onis-cidea indicates that these crustaceans were already highly diver-sified as early as the Cenomanian. This first Burmese amber record,added to the European amber specimens (from France and Spain),indicates that the Oniscidea group was widely distributed duringthe Early Cretaceous. These results support a pre-Pangean origin ofOniscidea.
Finally, this Cretaceous specimen, being a water-free organism,represents the earliest formal trace of land colonization in Crus-tacea, therefore considerably improving our knowledge of theevolutionary history of this large group of arthropods.
Acknowledgements
We are grateful to J.-P. Parent (ICL), B. Milhau (ICL), A. Giurgincaand I. Tabacaru for their valuable comments about familial place-ment of the specimen. We gratefully acknowledge the two anon-ymous reviewers for their constructive recommendations. Also,many thanks are addressed to B. Crighton (NMS volunteer) fortaking the photographs, and to R. Broly for translation of theGerman literature.We alsowant to acknowledge D. Ouvrard (NHM)and M. Judson (MNHN) for couriering and temporarily storing thespecimen. Pierre Broly is supported by a FRIA grant (Fonds pour laRecherche dans l’Industrie et dans l’Agriculture, FRS-FNRS).
References
Alonso, J., Arillo, A., Barr�on, E., Corral, J.C., Grimalt, J., L�opez, J.F., Martínez-Delcl�os, X.,Ortu~no, V., Pe~nalver, E., Trinc~ao, P.R., 2000. A new fossil resin with biologicalinclusions in Lower Cretaceous deposits from �Alava (Northern Spain, Basque-Cantabrian Basin). Journal of Paleontology 74, 158e178.
Becker, A., Ziegler, A., Epple, M., 2005. The mineral phase in the cuticles of twospecies of Crustacea consists of magnesium calcite, amorphous calcium car-bonate, and amorphous calcium phospate. Dalton Transactions 10, 1814e1820.
Budde-Lund, G., 1909. Zoologische und anthropologische Ergebnisse im westlichenund zentralen Südafrika, vol. 2, IX. Isopoden (I), Land-Isopoden. Denkschriftender medizinisch-naturwissenschaftlichen Gesellschaft zu Jena 14, 53e70.
Briggs, D.E.G., 1999. Molecular taphonomy of animal and plant cuticles: Selectivepreservation and diagenesis. Philosophical Transactions of the Royal Society B354, 7e17.
Broly, P., Deville, P., Maillet, S., 2013. The origin of terrestrial isopods (Crustacea:Isopoda: Oniscidea). Evolutionary Ecology 27, 461e476.
Campos-Filho, I.S., Araujo, P.B., Bichuette, M.E., Trajano, E., Taiti, S., 2014. Terrestrialisopods (Crustacea: Isopoda: Oniscidea) from Brazilian caves. Zoological Journalof the Linnean Society 172, 360e425.
Carefoot, T.H., Taylor, B.E., 1995. Ligia: A prototypal terrestrial isopod. In:Alikhan, A.M. (Ed.), Terrestrial Isopod Biology. Crustacean Issues, Rotterdam,pp. 47e60.
Cruickshank, R.D., Ko, K., 2003. Geology of an amber locality in the Hukawng Valley,Northern Myanmar. Journal of Asian Earth Sciences 21, 441e455.
Dalens, H., 1989. Sur un nouveau genre d’oniscoide «aquatique» provenant du sud-est Asiatique: Thailandoniscus annae (Isopoda, Oniscidea, Styloniscidae). Spixi-ana 12, 1e6.
Delcl�os, X., Arillo, A., Pe~nalver, E., Barr�on, E., Soriano, C., L�opez Del Valle, R.,Bern�ardez, E., Corral, C., Ortu~no, V., 2007. Fossiliferous amber deposits from theCretaceous (Albian) of Spain. Comptes Rendus Palevol 6, 135e149.
Dunlop, J.A., Scholtz, G., Selden, P.A., 2013. Water-to-land transitions. In: Minelli, A.,et al. (Eds.), Arthropod Biology and Evolution. Springer Berlin Heidelberg,Berlin, pp. 417e439.
Edney, E., 1954. Woodlice and the land habitat. Biological Reviews 29, 185e219.Erhard, F., 1998. Phylogenetic relationships within the Oniscidea (Crustacea, Iso-
poda). Israel Journal of Zoology 44, 303e309.Ferrara, F., Taiti, S., 1979. A check-list of terrestrial isopods from Africa (south of the
Sahara). Monitore zoologico italiano, Nuova Serie, Supplemento 12, 89e215.Gaillard, C., Hantzpergue, P., Vannier, J., Margerard, A.-L., Mazin, J.-M., 2005. Isopod
trackways from the Crayssac Lagerst€atte, Upper Jurassic, France. Palaeontology48, 947e962.
Girling, M.A., 1979. Calcium carbonate-replaced arthropods from archaeologicaldeposits. Journal of Archaeological Science 6, 309e320.
Gongalsky, K.B., Savin, F.A., Pokarzhevskii, A.D., Filimonova, Z.V., 2005. Spatial dis-tribution of isopods in an oakebeech forest. European Journal of Soil Biology 41,117e122.
Grimaldi, D.A., Engel, M.S., 2005. Evolution of the Insects. Cambridge UniversityPress, New York.
Gupta, N.S., Briggs, D.E.G., 2011. Taphonomy of animal organic skeletons throughtime. In: Allison, P.A., Bottjer, D.J. (Eds.), Taphonomy: Process and Bias ThroughTime, Topics in Geobiology, 32, pp. 199e221.
Hassall, M., Turner, J.G., Rands, M.R.W., 1987. Effects of terrestrial isopods on thedecomposition of woodland leaf litter. Oecologia 72, 597e604.
Heine, C., Müller, R.D., 2005. Late Jurassic rifting along the Australian NorthwestShelf: margin geometry and spreading ridge configuration. Australian Journal ofEarth Science 52, 27e39.
Hild, S., Othmar, M., Ziegler, A., 2008. Spatial distribution of calcite and amorphouscalcium carbonate in the cuticle of the terrestrial crustaceans Porcellio scaberand Armadillidium vulgare. Journal of Structural Biology 163, 100e108.
Holdich, D.M., Jones, D.A., 1973. The systematics and ecology of a new genus of sandbeach isopod (Sphaeromatidae) from Kenya. Journal of Zoology 171, 385e395.
Hornung, E., 2011. Evolutionary adaptation of oniscidean isopods to terrestrial life:Structure, physiology and behavior. Terrestrial Arthropod Reviews 4, 95e130.
Hornung, E., T�othm�er�esz, B., Magura, T., 2007. Changes of isopod assemblages alongan urbanerural gradient in Hungary. European Journal of Soil Biology 43,158e161.

P. Broly et al. / Cretaceous Research 55 (2015) 220e228228
Jass, J., Klausmeier, B., 2000. Endemics and immigrants: North American terrestrialisopods (Isopoda, Oniscidea) north of Mexico. Crustaceana 73, 771e799.
Kautz, G., Topp, W., 2000. Acquisition of microbial communities and enhancedavailability of soil nutrients by the isopod Porcellio scaber (Latr.) (Isopoda:Oniscidea). Biology and Fertility of Soils 31, 102e107.
Kensley, B., 1971. Termitophilous isopods from southern Africa. Annals of the SouthAfrican Museum 57, 131e147.
Leistikow, A., W€agele, J.W., 1999. Checklist of the terrestrial isopods of the NewWorld (Crustacea, Isopoda, Oniscidea). Revista Brasileira de Zoologia 16, 1e72.
Linsenmair, K.E., 2007. Sociobiology of terrestrial isopods. In: Duffy, J.E., Thiel, M.(Eds.), Evolutionary ecology of social and sexual systems: crustaceans as modelorganisms. Oxford University Press, New York, pp. 339e364.
Magrini, M.J., Araujo, P.B., Uehara-Prado, M., 2010. Crustacea, Isopoda, OniscideaLatreille, 1802: New continent record and distribution extension in Brazil. CheckList 6, 217e219.
Mattern, D., 2003. New aspects in the phylogeny of the Oniscidea inferred frommolecular data. In: Sfenthourakis, S., Araujo de, P.B., Hornung, E., et al. (Eds.),The biology of terrestrial Isopods V. Crustaceana Monographs, pp. 23e37.
Metcalfe, I., 1998. Palaeozoic and Mesozoic geological evolution of the SE Asia re-gion: multidisciplinary constraints and implications for biogeography. In:Hall, R., Holloway, J.D. (Eds.), Biogeography and Geology of SE Asia. Backhuys,Leiden, pp. 25e41.
Metcalfe, I., 2013. Gondwana dispersion and Asian accretion: tectonic and palae-ogeographic evolution of easternTethys. Journal of Asian Earth Sciences 66,1e33.
Michel-Salzat, A., Bouchon, D., 2000. Phylogenetic analysis of mitochondrial LSUrRNA in oniscids. Comptes Rendus de l'Acad�emie des Sciences 323, 827e837.
Murienne, J., Daniels, S.R., Buckley, T.R., Mayer, G., Giribet, G., 2014. A living fossiltale of Pangaean biogeography. Proceedings of the Royal Society B 281,20132648.
N�eraudeau, D., 2008. Nouveaux regards sur l’�evolution et la biodiversit�e pass�ee. In:Grappin, C., Cardin, P., Goff�e, B., et al. (Eds.), Terre, Plan�ete myst�erieuse. LeCherche Midi �editeur, Paris, pp. 128e137.
Neues, F., Ziegler, A., Epple, M., 2007. The composition of the mineralized cuticle inmarine and terrestrial isopods: a comparative study. CrystEngComm 9,1245e1251.
Paoletti, M.G., Hassall, M., 1999. Woodlice (Isopoda: Oniscidae): their potential forassessing sustainability and use as bioindicators. Agriculture, Ecosystems andEnvironment 74, 157e165.
Perkovsky, E.E., Zosimovich, V.Y., Vlaskin, A.P., 2010. Rovno amber. In: Penney, D.(Ed.), Biodiversity of fossils in amber from the major world deposits. Siri Sci-entific Press, Manchester, pp. 116e136.
Perrichot, V., 2004. Early Cretaceous amber from south-western France: insight intothe Mesozoic litter fauna. Geologica Acta 2, 9e22.
Podgaiski, R.L., Ott, R., Ganade, G., 2007. Ocupaç~ao de microh�abitats artificiais porinvertebrados de solo em um fragmento florestal no sul do Brasil. NeotropicalBiology and Conservation 2, 71e79.
Quadros, A.F., Araujo, P.B., 2008. An assemblage of terrestrial isopods (Crustacea) insouthern Brazil and its contribution to leaf litter processing. Revista Brasileirade Zoologia 25, 58e66.
Richardson, A., Araujo, P.B., 2015. Lifestyles of Terrestrial Crustaceans. In: Thiel, M.,Walting, L. (Eds.), The Natural History of the Crustacea, Lifestyles and FeedingBiology. Oxford University Press, New York, pp. 299e336.
Ross, A., Mellish, C., York, P., Crighton, B., 2010. Burmese Amber. In: Penney, D. (Ed.),Biodiversity of fossils in amber from the major world deposits. Siri ScientificPress, Manchester, pp. 208e235.
Schmalfuss, H., 1977. Morphologie und Funktion der tergalen L€angsrippen beiLandisopoden (Oniscoidea, Isopoda, Crustacea). Zoomorphologie 86, 155e167.
Schmalfuss, H., 1980. Die ersten Landasseln aus Dominikanischem Bernstein miteiner systematisch phylogenetischen Revision der Familie Sphaeroniscidae(Stuttgarter Bernsteinsammlung: Crustacea, Isopoda, Oniscoidea). StuttgarterBeitr€age zur Naturkunde, Serie B 61, 1e12.
Schmalfuss, H., 1984a. Two new species of the terrestrial isopod genus Pseudarma-dillo from Dominican amber (Amber-Collection Stuttgart: Crustacea, Isopoda,Pseudarmadillidae). Stuttgarter Beitr€age zur Naturkunde, Serie B 102, 1e14.
Schmalfuss, H., 1984b. Eco-morphological strategies in terrestrial isopods. In:Sutton, S.L., Holdich, D.M. (Eds.), The Biology of Terrestrial Isopods. TheZoological Society of London. Clarendon Press, Oxford, pp. 49e63.
Schmalfuss, H., 1989. Phylogenetics in Oniscidea. Monitore zoologico italiano,Nuova Serie, Monografia 4, 3e27.
Schmalfuss, H., 2003. World catalog of terrestrial isopods (Isopoda: Oniscidea).Stuttgarter Beitr€age zur Naturkunde, Serie A 654, 1e341.
Schmalfuss, H., 2005. Utopioniscus kuehni n. gen., n. sp. (Isopoda: Oniscidea: Syn-ocheta) from Submarine Caves in Sardinia. Stuttgarter Beitr€age zur Naturkunde,Serie A 677, 1e21.
Schmidt, C., 2008. Phylogeny of the terrestrial Isopoda (Oniscidea): a review.Arthropod Systematics & Phylogeny 66, 191e226.
Schmidt, C., Leistikow, A., 2004. Catalogue of genera of the terrestrial Isopoda(Crustacea: Isopoda: Oniscidea). Steenstrupia 28, 1e118.
Schmidt, A.R., Jancke, S., Lindquist, E.E., Ragazzi, E., Roghi, G., Nascimbene, P.C.,Schmidt, K., Wappler, T., Grimaldi, D.A., 2012. Arthropods in amber from theTriassic Period. Proceedings of the National Academy of Sciences of the UnitedStates of America 109, 14796e14801.
Selden, P.A., Penney, D., 2010. Fossil spiders. Biological Reviews 85, 171e206.Seton, M., Mueller, R.D., Zahirovic, S., Gaina, C., Torsvik, T., Shepard, G., Talsma, A.,
Gurnis, M., Turner, M., Chandler, M., 2012. Global continental and ocean basinreconstructions since 200 Ma. Earth-Science Reviews 113, 212e270.
Shi, G., Grimaldi, D.A., Harlow, G.E., Wang, J., Wang, J., Yang, M., Lei, W., Li, X., 2012.Age constraint on Burmese amber based on U-Pb dating of zircons. CretaceousResearch 37, 155e163.
Spahr, U., 1993. Erg€anzungen und Berichtigungen zu R. Keilbachs Bibliographie undListe der Bernsteinfossilien - Verschiedene Tiergruppen, ausgenommen Insectaund Araneae. Stuttgarter Beitr€age zur Naturkunde, Serie B 194, 1e77.
Strouhal, H., 1961. Die Oniscoideen-Fauna der Juan Fern�andez - Inseln (Crustacea,Isopoda terrestria). Annalen des Naturhistorischen Museums in Wien 64,185e244.
Tabacaru, I., 1993. Sur la classification des Trichoniscidae et la position syst�ematiquede Thaumatoniscellus orghidani Tabacaru, 1973 (Crustacea, Isopoda, Oniscidea).Travaux de l'Institut de Sp�eologie “Emile Racovitza” 32, 43e85.
Tabacaru, I., Danielopol, D.L., 1996. Phylog�enie des Isopodes terrestres. ComptesRendus de l’Acad�emie des Sciences, Sciences de la vie 319, 71e80.
Taiti, S., Xue, Z., 2012. The cavernicolous genus Trogloniscus nomen novum, withdescriptions of four new species from southern China (Crustacea, Oniscidea,Styloniscidae). Tropical Zoology 25, 183e209.
Taiti, S., Arnedo, M.A., Lew, S.E., Roderick, G.K., 2003. Evolution of terrestriality inHawaiian species of the genus Ligia (Isopoda, Oniscidea). In: Sfenthourakis, S.,Araujo de, P.B., Hornung, E., et al. (Eds.), The biology of terrestrial Isopods V.Crustaceana Monographs, pp. 85e102.
Uchman, A., Hu, B., Wang, Y., Song, H., 2011. The trace fossil Diplopodichnus from theLower Jurassic lacustrine sediments of central China and the isopod Arma-dillidium vulgare (Pillbug) lebensspuren as its recent analogue. Ichnos 18,147e155.
Vandel, A., 1952. Les Trichoniscides (Crustac�es; Isopodes) de l'h�emisph�ere austral.Leur place syst�ematique. Leur int�eret biog�eographique. M�emoires du Mus�eumNational d'Histoire Naturelle, S�erie A : Zoologie 6, 1e116.
Vandel, A., 1960. Faune de France 64 : les Isopodes Terrestres, premi�ere partie.Lechevallier, Paris.
Vandel, A., 1975. Les Isopodes terrestres et le Gondwana. M�emoires du Mus�eumNational d'Histoire Naturelle, S�erie A : Zoologie 88, 107e112.
Warburg, M.R., 1993. Evolutionary Biology of Land Isopods. Springer-Verlag, Berlin.Weitschat, W., Wichard, W., 2010. Baltic Amber. In: Penney, D. (Ed.), Biodiversity of
fossils in amber from the major world deposits. Siri Scientific Press, Manchester,pp. 80e115.
Wieder, R.W., Feldmann, R.M., 1992. Mesozoic and Cenozoic fossil isopods of NorthAmerica. Journal of Paleontology 66, 958e972.
Wieser, W., 1978. Consumer strategies of terrestrial gastropods and isopods.Oecologia 36, 191e201.
Zimmer, M., 2002. Nutrition in terrestrial isopods (Isopoda: Oniscidea): anevolutionary-ecological approach. Biological Reviews 77, 455e493.
Related Documents











