The enzyme-binding region of human GM2-activator protein Michaela Wendeler 1 , Norbert Werth 1 , Timm Maier 2 , Guenter Schwarzmann 1 , Thomas Kolter 1 , Maike Schoeniger 1 , Daniel Hoffmann 3 , Thorsten Lemm 1 , Wolfram Saenger 2 and Konrad Sandhoff 1 1 Kekule ´ -Institut fu ¨ r Organische Chemie und Biochemie der Universita ¨t Bonn, Germany 2 Institut fu ¨ r Chemie, Abt. Kristallographie, Freie Universita ¨t Berlin, Germany 3 Forschungszentrum caesar, Bonn, Germany Keywords GM2-activator; ganglioside degradation; b-hexosaminidase A; lipid transfer; lysosome Correspondence K. Sandhoff, Kekule ´ -Institut fu ¨ r Organische Chemie und Biochemie der Universita ¨t Bonn, Gerhard-Domagk-Str. 1, D-53121 Bonn, Germany Fax: +49 228 73 7778 Tel: +49 228 73 5346 E-mail: [email protected] (Received 8 November 2005, revised 21 December 2005, accepted 28 December 2005) doi:10.1111/j.1742-4658.2006.05126.x The GM2-activator protein (GM2AP) is an essential cofactor for the lyso- somal degradation of ganglioside GM2 by b-hexosaminidase A (HexA). It mediates the interaction between the water-soluble exohydrolase and its membrane-embedded glycolipid substrate at the lipid–water interface. Functional deficiencies in this protein result in a fatal neurological storage disorder, the AB variant of GM2 gangliosidosis. In order to elucidate this cofactor’s mode of action and identify the surface region of GM2AP responsible for binding to HexA, we designed several variant forms of this protein and evaluated the consequences of these mutations for lipid- and enzyme-binding properties using a variety of biophysical and functional studies. The point mutants D113K, M117V and E123K showed a drastic- ally decreased capacity to stimulate HexA-catalysed GM2 degradation. However, surface plasmon resonance (SPR) spectroscopy showed that the binding of these variants to immobilized lipid bilayers and their ability to solubilize lipids from anionic vesicles were the same as for the wild-type protein. In addition, a fluorescence resonance energy transfer (FRET)- based assay system showed that these variants had the same capacity as wild-type GM2AP for intervesicular lipid transfer from donor to acceptor liposomes. The concentration-dependent effect of these variants on hydro- lysis of the synthetic substrate 4-methylumbelliferyl-2-acetamido-2-deoxy-6- sulfo-b-d-glucopyranoside (MUGS) indicated a weakened association with the enzyme’s a subunit. This identifies the protein region affected by these mutations, the single short a helix of GM2AP, as the major determinant for the interaction with the enzyme. These results further confirm that the function of GM2AP is not restricted to a biological detergent that simply disrupts the membrane structure or lifts the substrate out of the lipid plane. In contrast, our data argue in favour of the critical importance of distinct activator–hexosaminidase interactions for GM2 degradation, and corrobor- ate the view that the activator ⁄ lipid complex represents the true substrate for the degrading enzyme. Abbreviations BMP, bis(monoacylglycero)phosphate; FRET, fluorescence resonance energy transfer; GM1, ganglioside GM1; GM2, ganglioside GM2; GM2AP, GM2-activator protein; GSLs, glycosphingolipids; HexA, b-hexosaminidase A; HexB: b-hexosaminidase B; LUV, large unilamellar vesicle; MUG, 4-methylumbelliferyl-2-acetamido-2-deoxy-b-D-glucopyranoside; MUGS, 4-methylumbelliferyl-2-acetamido-2-deoxy-6-sulfo-b- D-glucopyranoside; NBD, 7-nitrobenz-2-oxa-1,3-diazol-4-yl; PtdCho, phosphatidylcholine; PE, dioleoyl glycero phosphoryl ethanolamine; SAP, sphingolipid activator protein; SPR, surface plasmon resonance. 982 FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The enzyme-binding region of human GM2-activatorproteinMichaela Wendeler1, Norbert Werth1, Timm Maier2, Guenter Schwarzmann1, Thomas Kolter1,Maike Schoeniger1, Daniel Hoffmann3, Thorsten Lemm1, Wolfram Saenger2 and Konrad Sandhoff1
1 Kekule-Institut fur Organische Chemie und Biochemie der Universitat Bonn, Germany
2 Institut fur Chemie, Abt. Kristallographie, Freie Universitat Berlin, Germany
3 Forschungszentrum caesar, Bonn, Germany
Keywords
GM2-activator; ganglioside degradation;
b-hexosaminidase A; lipid transfer; lysosome
Correspondence
K. Sandhoff, Kekule-Institut fur Organische
Chemie und Biochemie der Universitat
Bonn, Gerhard-Domagk-Str. 1, D-53121
Bonn, Germany
Fax: +49 228 73 7778
Tel: +49 228 73 5346
E-mail: [email protected]
(Received 8 November 2005, revised 21
December 2005, accepted 28 December
2005)
doi:10.1111/j.1742-4658.2006.05126.x
The GM2-activator protein (GM2AP) is an essential cofactor for the lyso-
somal degradation of ganglioside GM2 by b-hexosaminidase A (HexA). It
mediates the interaction between the water-soluble exohydrolase and its
membrane-embedded glycolipid substrate at the lipid–water interface.
Functional deficiencies in this protein result in a fatal neurological storage
disorder, the AB variant of GM2 gangliosidosis. In order to elucidate this
cofactor’s mode of action and identify the surface region of GM2AP
responsible for binding to HexA, we designed several variant forms of this
protein and evaluated the consequences of these mutations for lipid- and
enzyme-binding properties using a variety of biophysical and functional
studies. The point mutants D113K, M117V and E123K showed a drastic-
ally decreased capacity to stimulate HexA-catalysed GM2 degradation.
However, surface plasmon resonance (SPR) spectroscopy showed that the
binding of these variants to immobilized lipid bilayers and their ability to
solubilize lipids from anionic vesicles were the same as for the wild-type
protein. In addition, a fluorescence resonance energy transfer (FRET)-
based assay system showed that these variants had the same capacity as
wild-type GM2AP for intervesicular lipid transfer from donor to acceptor
liposomes. The concentration-dependent effect of these variants on hydro-
lysis of the synthetic substrate 4-methylumbelliferyl-2-acetamido-2-deoxy-6-
sulfo-b-d-glucopyranoside (MUGS) indicated a weakened association with
the enzyme’s a subunit. This identifies the protein region affected by these
mutations, the single short a helix of GM2AP, as the major determinant
for the interaction with the enzyme. These results further confirm that the
function of GM2AP is not restricted to a biological detergent that simply
disrupts the membrane structure or lifts the substrate out of the lipid plane.
In contrast, our data argue in favour of the critical importance of distinct
activator–hexosaminidase interactions for GM2 degradation, and corrobor-
ate the view that the activator ⁄ lipid complex represents the true substrate
for the degrading enzyme.
Abbreviations
BMP, bis(monoacylglycero)phosphate; FRET, fluorescence resonance energy transfer; GM1, ganglioside GM1; GM2, ganglioside GM2;
GM2AP, GM2-activator protein; GSLs, glycosphingolipids; HexA, b-hexosaminidase A; HexB: b-hexosaminidase B; LUV, large unilamellar
vesicle; MUG, 4-methylumbelliferyl-2-acetamido-2-deoxy-b-D-glucopyranoside; MUGS, 4-methylumbelliferyl-2-acetamido-2-deoxy-6-sulfo-b-
D-glucopyranoside; NBD, 7-nitrobenz-2-oxa-1,3-diazol-4-yl; PtdCho, phosphatidylcholine; PE, dioleoyl glycero phosphoryl ethanolamine;
SAP, sphingolipid activator protein; SPR, surface plasmon resonance.
982 FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS

Glycosphingolipids (GSLs) are characteristic compo-
nents of eukaryotic plasma membranes. Over the last
20 years, their highly complex degradation pathway in
lysosomes and the metabolic diseases associated with
inherited defects in this pathway have been intensively
investigated [1]. In the case of GSLs with rather short
oligosaccharide head-groups of four or fewer sugar resi-
dues, these membrane-embedded substrates are not suf-
ficiently accessible for the degradation by water-soluble
enzymes, and the exohydrolases need the assistance of
small glycoprotein cofactors, the sphingolipid activator
proteins (SAPs) [2]. For hydrolytic conversion of gan-
glioside GM2 catalysed by b-hexosaminidase A (HexA,
EC 3.2.1.52), the GM2-activator protein is required
[3,4].
Three different isoforms of human lysosomal b-hexo-saminidases are known: b-hexosaminidase A (HexA),
the heterodimer of the noncovalently linked a chainand b chain, and the homodimeric isoenzymes b-hex-osaminidase B (HexB, bb) and b-hexosaminidase S
(aa). Despite the a and b subunits being 60% identical
in their primary structure and functionally very similar,
they show distinct specificity [5]. Only HexA is able to
degrade the physiologically most important substrate,
ganglioside GM2, at significant rates in the presence of
the GM2 activator. Defects in any of the three genes
encoding the polypeptides involved in GM2 degrada-
tion, the a and b subunit of HexA or the GM2AP,
result in the accumulation of nondegraded glycolipids
within the lysosomal compartment and the develop-
ment of severe neurodegenerative storage diseases
known as GM2-gangliosidoses [4,6,7].
In its mature form present in the lysosomes, the
GM2 activator is a small glycoprotein of 162 amino
acids [8]. It is known to bind avidly to a variety of ani-
onic lipids in vitro [9], to be able to extract several gly-
cosphingolipid monomers from micelles or liposomes
and to transport them as soluble 1 : 1 complexes
between donor and acceptor membranes [10]. The
membrane activity of GM2AP was found to depend
critically on several physiological parameters, most
notably a lateral pressure of the lipid bilayer below
15–25 mNÆm)1 [11] and the presence of anionic lipids,
in particular bis(monoacylglycero)phosphate (BMP)
[12]. Recently, a novel function of GM2AP in the con-
text of glycolipid antigen presentation via CD1d was
identified [13].
For the function of GM2AP in the lysosomal GM2
hydrolysis, the term ‘liftase’ was coined: it recognizes
the lipid substrate within the membrane plane and lifts
it out of the lipid bilayer, thereby presenting it to the
water-soluble enzyme for degradation [14]. In addition,
it has been suggested that GM2AP modifies the
conformation of the trisaccharide unit of ganglioside
GM2, thus facilitating the enzymatic hydrolysis of the
terminal N-acetyl-d-galactosamine (GalNAc) residue
[15]. Additional protein–protein interactions between
GM2AP and HexA were implied in the catalysis of
GM2 degradation [16,17]. Several lines of evidence
indicate that GM2AP interacts with the a subunit of
HexA, but the presence of the b subunit enhances
binding [5,16,18,19]. The crystal structure of mature
GM2AP expressed in Escherichia coli [20], as well as
that of lipid complexes of GM2AP [21,22], revealed a
novel protein fold, denoted b cup, which consists of an
eight-stranded antiparallel b-pleated sheet forming a
spacious hydrophobic cavity, as well as several surface
loops and a single short a helix. The dimensions of the
central pocket are such that it can accommodate the
ceramide tail of GM2. Using photoaffinity labelling,
we were able to establish that the most flexible of the
surface loops, the chain segment V153–L163 constitutes
the part of the activator protein that directly interacts
with the ganglioside substrate [23]. (The amino acid
numbering used here refers to the complete, prepro,
form of GM2AP, including residues 1–31 which are
removed by proteolytic processing. Please note that
Protein Data Bank entry 1G13 describing the crystal
structure of mature GM2AP expressed in E. coli, and
also Wright et al. [21], differ from this nomenclature;
they assign the number 1 to S32, which represents the
N-terminus of mature GM2AP.) This interaction might
be crucial for stabilization of the position of the glycoli-
pid within the spacious cavity, thereby ensuring the
correct orientation of the tetrasaccharide head-group
with respect to the degrading enzyme’s active site.
The goal of this study was to elucidate the interac-
tion between cofactor and enzyme and to develop a
comprehensive model for the GM2 degradation pro-
ceeding at a phase frontier on intralysosomal mem-
branes. Previously, attempts to identify the lipid- and
enzyme-binding region of GM2AP were hampered
because assay systems for GM2AP, which measure its
stimulatory potential for the degradation of GM2 by
HexA, reflect simultaneously both the interaction of
GM2AP with GM2 and the interaction of the
GM2AP ⁄GM2 complex with the enzyme. To delineate
in detail the separate effect of the introduced muta-
tions on the lipid- and the enzyme-binding of this
cofactor, we combined a variety of functional and bio-
physical analyses.
Crystallographic analysis of the homodimeric iso-
enzyme HexB [24,25] and subsequent comparative
modelling studies of heterodimeric HexA [25] suggested
that the dimer interface of the enzyme forms the dock-
ing site for the GM2AP ⁄GM2 complex. Furthermore,
M. Wendeler et al. Enzyme-binding region of GM2-activator protein
FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS 983

based on the crystal structures of the enzyme [24,25]
and of GM2AP [20–22], computational modelling and
theoretical docking studies identified the single short
ahelix of GM2AP as the region most likely to be in
direct contact with the enzyme. We therefore expressed
a series of site-directed mutants of GM2AP with amino
acid exchanges in this region of the protein. Only vari-
ants with conformations identical to the wild-type
(WT) GM2AP, as judged by CD spectroscopy, were
considered. To study the stimulatory potential of
GM2AP variants for ganglioside GM2 degradation by
HexA, conventional micellar assays, as well as a deter-
gent-free, liposomal assay system [12], were employed.
The influence on the hydrolysis of the synthetic sub-
strate 4-methylumbelliferyl-2-acetamido-2-deoxy-6-sulfo-
b-d-glucopyranoside (MUGS), which is degraded by
the a subunit of HexA and which hydrolysis is inhibited
by GM2AP [5], allowed probing the variant activator’s
binding to the enzyme. To study the membrane activity
and lipid-binding properties, SPR spectroscopy with
immobilized lipid bilayers was performed. Finally, a
recently developed assay based on fluorescence reson-
ance energy transfer (FRET) [26] enabled us to observe
in real-time the ability for intervesicular lipid transfer
from donor to acceptor vesicles.
This allowed identification of the a-helical region of
GM2AP as the major determinant in interacting with
the enzyme. Our data further confirm conclusively the
critical importance of distinct protein–protein interac-
tions in GM2 degradation and corroborate the view
that the activator ⁄ glycolipid complex represents the
true substrate for the degrading enzyme.
Results
To identify the protein region of GM2AP that inter-
acts with HexA in the lysosomal degradation of gan-
glioside GM2, we performed site-directed mutagenesis
of GM2AP and evaluated the biophysical and func-
tional properties of the variant proteins.
Initial homology modelling of HexA based on the
recently published crystal structure of human HexB
[25], and subsequent theoretical docking studies of the
GM2AP structure [20,21] suggested that the a helix of
GM2AP plays a major role in binding to the enzyme.
In this helix, comprising amino acids F111 to P120,
every fourth residue points in the same direction and
the side chains of D113 and M117 were found to be
oriented away from the GM2AP core towards the
putative interface with the enzyme. The first residue
following the GM2AP helix, which likewise points
towards this suggested interaction region, was E123.
The long and charged side chain of glutamate is
known to be particularly well suited for protein–pro-
tein interactions. In contrast, the residues between the
end of the helix and E123 (P120, T121, G122) most
likely serve a structural role only, allowing the loop to
adopt a bent conformation.
We therefore introduced the following mutations in
this region of the protein: D113K, D113A, D113Y,
M117V, E123K, E123A and E123Y. All variant pro-
tein forms were readily produced in the insect cell ⁄baculovirus expression vector system (BEVS) and puri-
fied to homogeneity by cation exchange and sub-
sequent Ni-NTA chromatography. Protein yields of
GM2AP mutants were in the range 4–7 mg purified
protein per litre of expression supernatant, compared
with 7–9 mg of the WT protein.
To monitor any possible disturbances in the protein
fold upon amino acid exchange, we subjected all
protein variants to UV circular dichroism (CD) spectro-
scopy (Fig. 1). Although secondary structure pre-
dictions suggested identical conformations for the
above-described GM2AP variants, it was found that
the point mutants D113A, D113Y, E123A and E123Y
showed altered CD spectra. In contrast, the mutations
M117V, D113K and E123K resulted in CD spectra
identical to that of the WT protein. Only these vari-
ants of GM2AP were examined in all further studies.
To assess the potential of the variant GM2AP forms
for the stimulation of HexA catalysed hydrolysis of
GM2, two established assays, involving either micellar
or liposomal glycolipid substrate, were performed. As
shown in Fig. 2, the mutant forms of GM2AP exam-
ined here showed a drastically decreased ability to
stimulate the hydrolysis of GM2 by HexA. In the mi-
cellar system (Fig. 2A), the point mutant M117V dis-
played 25–26%, the variant D113K 4–5% and E123K
3–4% of the WT capacity to stimulate the enzymatic
degradation of GM2. In the liposomal system (Fig. 2B),
which we introduced to mimic more closely the in vivo
reaction conditions on the surface of intralysosomal
vesicles and membrane structures [12], these results were
confirmed and the differences between WT protein and
mutants were even more pronounced. In this system,
the variant M117V showed 7–8% of the WT activity,
whereas the mutants D113K and E123K displayed
almost a complete loss of activity.
To assess the membrane activity and lipid-binding
behaviour of the GM2AP variants, their interaction
with immobilized lipid bilayers was measured using
SPR spectroscopy in a Biacore instrument (Fig. 3). All
mutant protein forms showed binding to the same
extent as the WT protein. In the presence of the ani-
onic lipid BMP, which is known to occur in inner
membranes of the acidic compartments and which
Enzyme-binding region of GM2-activator protein M. Wendeler et al.
984 FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS

stimulates membrane activity of GM2AP [12], all
mutants showed a comparable pronounced decrease in
the SPR signal during the subsequent injection of pro-
tein-free buffer. This response has been interpreted as
the solubilization of lipids from the surface of immobi-
Fig. 2. Stimulation of HexA-catalysed ganglioside GM2 degradation
by WT and variant GM2AP. (A) Substrate hydrolysis measured in a
micellar assay system. (B) GM2 degradation measured in a lipo-
somal assay system with LUVs containing 50 mol% PtdCho,
20 mol% Chol, 10 mol% GM2 and 20 mol% BMP. Values repre-
sent means of duplicate measurements; deviations observed were
< 5%.
Fig. 3. Interaction of WT and variant GM2AP with immobilized vesi-
cles measured by SPR spectroscopy. Negatively charged LUVs
containing 20 mol% Chol, 50 mol% PtdCho, 10 mol% GM2 and
20 mol% BMP, were immobilized on a Pioneer L1 sensorchip.
GM2AP (2 lM) was injected at a flow rate of 20 lLÆmin)1 in 50 mM
sodium citrate buffer, pH 4.2, for 180 s, followed by the injection
of protein-free buffer. The measurement started with the protein
injection.
A
B
C
Fig. 1. UV circular dichroism (CD) spectra of wild-type (WT) GM2AP
and variant proteins examined in this study. (A) WT GM2AP and
mutant M117V. (B) WT GM2AP and D113 mutants. (C) WT GM2AP
and E123 mutants. Spectra were obtained at 10 �C, at a protein
concentration of 0.5 mgÆmL)1 in 15 mM sodium phosphate, pH 7.0,
100 mM NaCl.
M. Wendeler et al. Enzyme-binding region of GM2-activator protein
FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS 985
lized membrane structures via the action of bound
GM2AP [12], either by directly extracting lipids or by
destabilizing the bilayers with a concomitant loss of
lipids. The point mutants examined here were found to
interact with immobilized lipid bilayers in the same
way as WT GM2AP.
To study directly the lipid-transfer properties of
GM2AP, we recently introduced a novel FRET-based
assay system [26]. A new class of fluorescent glyco-
sphingolipid analogue, either 2-(N-(7-nitrobenz-2-oxa-
1,3-diazol-4-yl)) (NBD)–GM1 or 2-NBD–GM2, serves
as FRET-donor for rhodamine–dioleoyl glycero phos-
phoryl ethanolamine (PE) as acceptor which is initially
localized within the same model membrane and thereby
quenches NBD fluorescence. It has been shown previ-
ously that GM1 and GM2 are transferred almost
equally well between vesicles by GM2AP [10]. These
novel fluorescent gangliosides are easily incorporated
into model membranes, where they exhibit an extremely
low ‘off rate’ and are recognized and degraded by HexA
in the presence of GM2AP, similar to their natural ana-
logue. Upon addition of catalytic amounts of GM2AP
to donor vesicles containing NBD–gangliosides, the
activator-mediated glycolipid transfer from donor to
acceptor liposomes increases the NBD fluorescence,
permitting continuous real-time monitoring of GM2AP
transfer activity. As shown in Fig. 4, the point mutants
examined here showed the same ability as WT GM2AP
to transfer NBD–GM1 from donor to acceptor vesicles.
The initial rate of NBD transfer and the capacity
observed under equilibrium conditions are identical.
It has previously been shown that, in the absence of
ganglioside GM2, the GM2 activator protein inhibits
hydrolysis of the synthetic fluorogenic substrate
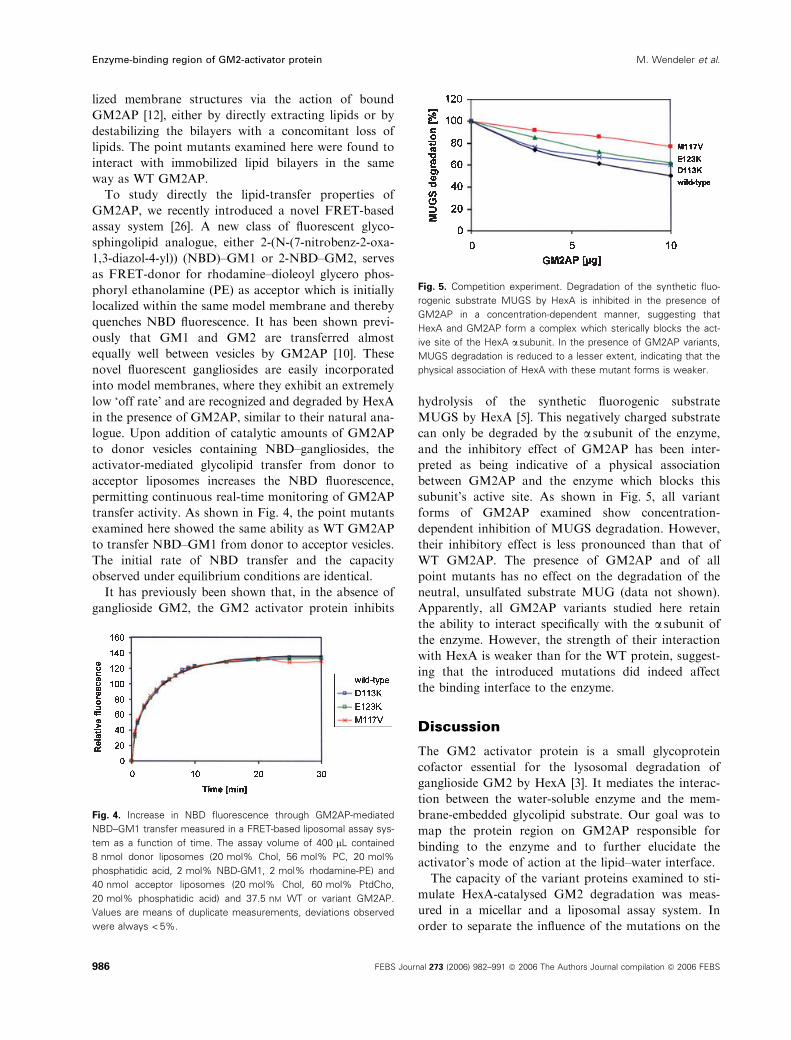
MUGS by HexA [5]. This negatively charged substrate
can only be degraded by the a subunit of the enzyme,
and the inhibitory effect of GM2AP has been inter-
preted as being indicative of a physical association
between GM2AP and the enzyme which blocks this
subunit’s active site. As shown in Fig. 5, all variant
forms of GM2AP examined show concentration-
dependent inhibition of MUGS degradation. However,
their inhibitory effect is less pronounced than that of
WT GM2AP. The presence of GM2AP and of all
point mutants has no effect on the degradation of the
neutral, unsulfated substrate MUG (data not shown).
Apparently, all GM2AP variants studied here retain
the ability to interact specifically with the a subunit ofthe enzyme. However, the strength of their interaction
with HexA is weaker than for the WT protein, suggest-
ing that the introduced mutations did indeed affect
the binding interface to the enzyme.
Discussion
The GM2 activator protein is a small glycoprotein
cofactor essential for the lysosomal degradation of
ganglioside GM2 by HexA [3]. It mediates the interac-
tion between the water-soluble enzyme and the mem-
brane-embedded glycolipid substrate. Our goal was to
map the protein region on GM2AP responsible for
binding to the enzyme and to further elucidate the
activator’s mode of action at the lipid–water interface.
The capacity of the variant proteins examined to sti-
mulate HexA-catalysed GM2 degradation was meas-
ured in a micellar and a liposomal assay system. In
order to separate the influence of the mutations on the
Fig. 4. Increase in NBD fluorescence through GM2AP-mediated
NBD–GM1 transfer measured in a FRET-based liposomal assay sys-
tem as a function of time. The assay volume of 400 lL contained
8 nmol donor liposomes (20 mol% Chol, 56 mol% PC, 20 mol%
phosphatidic acid, 2 mol% NBD-GM1, 2 mol% rhodamine-PE) and
40 nmol acceptor liposomes (20 mol% Chol, 60 mol% PtdCho,
20 mol% phosphatidic acid) and 37.5 nM WT or variant GM2AP.
Values are means of duplicate measurements, deviations observed
were always < 5%.
Fig. 5. Competition experiment. Degradation of the synthetic fluo-
rogenic substrate MUGS by HexA is inhibited in the presence of
GM2AP in a concentration-dependent manner, suggesting that
HexA and GM2AP form a complex which sterically blocks the act-
ive site of the HexA a subunit. In the presence of GM2AP variants,
MUGS degradation is reduced to a lesser extent, indicating that the
physical association of HexA with these mutant forms is weaker.
Enzyme-binding region of GM2-activator protein M. Wendeler et al.
986 FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS

lipid- and enzyme-binding properties, we tested the
membrane activity of GM2AP using SPR spectroscopy
and probed the lipid-transfer properties in a FRET-
based assay system. The ability to bind specifically the
a subunit of HexA could be assessed by studying
the competition of GM2AP and its variants with the
hydrolysis of the synthetic fluorogenic hexosaminidase
substrate MUGS.
Our data clearly indicate that mutations introduced
into the a-helical region of GM2AP drastically
decreased the ability of the activator to stimulate
HexA-catalysed degradation of GM2. However, the
membrane activity of these variants, as well as their
lipid-binding and transfer ability, was identical to that
of the WT protein. All variants could still bind speci-
fically to the a subunit of HexA, but their association
with the enzyme was weaker than for WT GM2AP.
This clearly identifies the region affected by these
mutations, the single a helix of GM2AP, as the major
surface epitope interacting with the enzyme.
That the mutants D113K and E123K exhibit the
most drastically reduced stimulatory activity for GM2
degradation and show the weakest binding to HexA
might be attributed to the reversal of local charge
resulting from the exchange of acidic amino acids to
lysine. In addition to possible electrostatic repulsion,
all salt bridges and hydrogen bonds which are possible
between positively charged residues on HexA and
amino acids D113 and E123 in WT GM2AP can no
longer be formed.
Figure 6 shows the position of the examined point
mutations at the interface between activator and HexA,
as a result of theoretical docking studies based on the
known crystal structures of GM2AP [20–22] and HexB
[24,25]. Our results are consistent with the location of
the ceramide tail of GM2 as identified by crystallo-
graphic analysis of lipid complexes of GM2AP [21],
which positions the tetrasaccharide head-group of the
substrate in direct vicinity of the enzyme’s active site.
Among patients with variant AB of GM2 gangliosi-
dosis, whose severe neurological storage of ganglioside
GM2 results from the functional deficiency of
GM2AP, no case has been found in which the muta-
tion occurred directly within the a-helical region of
GM2AP. However, the biochemical consequence of
one fatal mutation, the amino acid exchange C138R,
was thought to be mainly a disrupted interaction with
the enzyme [17], and this might well be affected by
altered orientation of the helix. The crystal structure of
GM2AP [20,21] and the known disulfide bond connec-
tivity [27] indicate that the disulfide bond formed by
C138 with C112 forms a ‘clamp’ that structurally
restrains the activator’s helical domain. It is therefore
most likely that the mutation C138R distorts the orien-
tation of the helix and compromises the activator’s
ability to interact with the enzyme.
In addition to mapping the activator’s binding inter-
face with the enzyme, our data confirm the critical
importance of distinct protein–protein interactions in
lysosomal GM2 degradation. The activator’s role is
not limited to that of a biological detergent that simply
disrupts the membrane structure. In contrast, the acti-
vator ⁄ glycolipid complex represents the true Michaelis
substrate for the degrading enzyme.
Experimental procedures
Materials
For insect cell culture, serum-free IPL-41 medium was pur-
chased from Invitrogen Life Technologies (Carlsbad, CA).
The baculovirus transfer vector pAcMP3, linearized baculo-
virus DNA (BaculoGold), the transfection kit and the insect
cell lines were from Pharmingen, obtained via BD Bio-
sciences (Erembodegem, Belgium). Phosphatidylcholine (egg
yolk, PtdCho) and cholesterol (Chol) were purchased from
Sigma (Taufkirchen, Germany). BMP was obtained from
Avanti Polar Lipids (Alabaster, AL), Lichroprep RP18 was
Fig. 6. Model for the interface in the ternary complex of GM2AP,
GM2 and HexA resulting from theoretical docking studies based on
the known crystal structures of GM2AP [20–22] and HexB [25]. In
the GM2AP structure b sheets are blue, the single a helix is green,
and regions exhibiting conformational differences in free and ligand-
bound form are red. The position of the bound ligand GM2 follows
the model presented in Fig. 4 of Wright et al. [21], as far as the
lipid tail is concerned. Regarding the tetrasaccharide head-group,
however, we consider a more ‘strained’ orientation as presented
here, more likely. The picture was generated using MOLSCRIPT [30].
M. Wendeler et al. Enzyme-binding region of GM2-activator protein
FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS 987

from Merck (Darmstadt, Germany). The Pioneer L1 Chip
was purchased from Biacore (Uppsala, Sweden). All other
chemicals and solvents were of analytical grade or the high-
est purity available.
Protein expression and mutagenesis
Recombinant GM2AP was expressed in BEVS as described
previously [28]. Site-directed mutagenesis using the vector
pAcMP3-GM2APHis6 [28] as a template was performed
using the QuikChange site-directed mutagenesis kit from
Stratagene (La Jolla, CA). Synthetic oligonucleotides were
obtained from Sigma and Invitrogen Life Technologies.
Mutations encoding the following amino acid exchanges
were introduced into the cDNA of GM2AP: D113K,
D113A, D113Y, M117V, E123K, E123A, E123Y. All
expression vectors encoded a hexahistidine tag at the C-ter-
minus of GM2AP. The resulting constructs encoding point
mutants of GM2AP were fully sequenced using an ABI 310
sequencer and the BigDye cycle sequencing kit, both from
Applied Biosystems (Foster City, CA). Recombinant bacu-
loviruses were then generated by cotransfection of Sf9 cells
with the respective transfer vector and linearized BaculoGold
viral DNA according to the manufacturer’s protocol. Pure
viral stocks were obtained using the end-point dilution
method.
For protein expression, Sf9 cells grown in serum-free
IPL-41 to a cell density of 1.5 · 106 cellsÆmL)1 were infec-
ted with a viral stock at a multiplicity of infection of 5. The
medium was harvested 96 h after infection.
Protein purification
Proteins were purified, with modification, following the
method described by Wendeler et al. [28]. Briefly, the super-
natant was first subjected to perfusion chromatography on
a cation exchange resin (Poros HS) using a BIOCADSprint
System HPLC workstation (Applied Biosystems). Fractions
containing recombinant GM2AP were combined and then
further purified by immobilized metal ion affinity chroma-
tography on Ni-NTA-agarose (Qiagen, Hilden, Germany).
The eluted proteins were analysed by electrophoresis on a
12.5% tricine-SDS ⁄ polyacrylamide gel and visualized by
silver staining. The polyclonal antibody raised against WT
GM2AP [29] recognized all GM2AP mutants. Amino acid
exchanges in the recombinant proteins were confirmed by
ESI-TOF-MS.
Micellar GM2AP assay
In a micellar in vitro system, the activity of recombinant
GM2AP was tested by measuring the stimulation of hex-
osaminidase A-catalysed hydrolysis of [3H]GM2 tritium-
labelled in its terminal GalNAc moiety [3]. In 40 lL of
100 mm sodium citrate buffer containing 2.5 lg BSA,
10 nmol [3H]GM2 were incubated with 80 mU HexA (puri-
fied from post mortem human liver) in the presence of 3 lgrecombinant GM2AP for 1 h. Reactions were stopped by
adding 40 lL methanol. Liberated [3H]GalNAc was isola-
ted using self-packed RP18 cartridges: 0.5 mL LiChroprep
RP18 (Merck) was applied to a glass Pasteur pipette, pre-
viously stuffed with a small amount of glass wool. The
material was equilibrated by washing subsequently with
2 · 1 mL chloroform ⁄methanol 1 : 1 (v ⁄ v), 2 · 1 mL
methanol and 2 · 1 mL chlorofom ⁄methanol ⁄ 0.1 m KCl
3 : 47 : 48 (v ⁄ v ⁄ v). The assay solution was then applied
and the flow-through collected. Soluble [3H]GalNAc was
then eluted with 2 · 1 mL water, and the eluate combined
with the flow-through and 10 mL scintillation liquid.
Radioactivity in the effluents was measured in a scintilla-
tion counter (Packard). One activator unit is defined as the
amount of GM2AP that stimulates the degradation of
1 nmol GM2 per minute and enzyme unit.
Liposomal assay systems
To mimic more closely the reaction conditions encountered
on intralysosomal vesicles and membrane structures, the
degradation of membrane-bound ganglioside GM2 by hex-
osaminidase and GM2AP was studied in a detergent-free,
liposomal assay system as described previously [12].
Vesicle prepapration
Large unilamellar vesicles (LUVs) of 100 nm were prepared
by the following procedure. Appropriate aliquots of the
lipid solutions PtdCho (50 mm, toluol ⁄ ethanol 2 : 1 v ⁄ v),BMP (10 mm, chloroform ⁄methanol 1 : 1 v ⁄ v), Chol
(25.6 mm, chloroform ⁄methanol 2 : 1 v ⁄ v) and GM2 trit-
ium-labelled in its terminal GalNAc moiety (0.5 mm, tolu-
ol ⁄ ethanol 1 : 1 v ⁄ v) were mixed and dried in a stream of
nitrogen. The lipid mixture was dissolved to a total lipid
concentration of 2 mm in sodium citrate buffer (50 mm,
pH 4.2) and freeze–thawed 10 times in liquid nitrogen to
ensure solute equilibration between trapped and bulk
solutions. Unilamellar vesicles were prepared by passage
through two polycarbonate filters (pore size, 100 nm; Aves-
tin) mounted in tandem in a mini-extruder (Liposo-Fast;
Avestin, Ottawa, Canada) a total of 19 times.
Liposomal assay
The standard incubation mixture using GM2 as substrate
contained the following components in a final volume of
50 lL: BSA (50 lgÆmL)1), sodium citrate buffer (pH 4.2,
50 mm), unilamellar liposomes (total lipid concentration:
1 mm), GM2AP (2 lm) and HexA (25 mU). Liposomes
had the following composition: [3H]GM2 (10 mol%,
Enzyme-binding region of GM2-activator protein M. Wendeler et al.
988 FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS

1.8 CiÆmol)1), Chol (20 mol%), PtdCho (50 mol%) and
BMP (20 mol%). The standard incubation conditions were
37 �C for 30 min, and the enzyme assays were stopped by
the addition of 50 lL methanol. Terminated enzyme assays
were loaded onto a reverse-phase column (RP18, 1 mL)
equilibrated with a solution of chloroform ⁄methanol ⁄ 0.1 m
KCl (3 : 48 : 47, v ⁄ v ⁄ v). The column was eluted with 2 mL
of the same solvent, and the radioactivity in the effluents
was measured in a scintillation counter.
Determination of HexA activity with the
fluorogenic substrates 4-methylumbelliferyl-
2-acetamido-2-deoxy-b-D-glucopyranoside and its
sulfated derivative MUGS
The activity of HexA towards the synthetic substrates
4-methylumbelliferyl-2-acetamido-2-deoxy-b-d-glucopyrano-side (MUG) and its sulfated derivative MUGS was meas-
ured essentially as described previously [5]. For routine
activity measurements, MUG was used as substrate under
the following standard conditions. In a volume of 40 lL,10 mm citrate buffer, pH 4.2, 2 mm substrate, 6 lg BSA
and an appropriate amount of HexA were incubated at
37 �C for 30 min. One enzyme unit is defined as the
amount of hexosaminidase that generates 1 lmolÆmin)1 of
4-methylumbelliferone.
MUGS was used in competition experiments designed to
study the association of GM2AP with the a subunit of
HexA. In a volume of 40 lL, 10 mm citrate buffer, pH 4.2,
2 · 10)5 m substrate, 6 lg BSA and an appropriate amount
of HexA were incubated at 37 �C for 30 min in the absence
or presence of various amounts of GM2AP (up to 10 lg).Reactions were stopped by the addition of 5 vol. of a 0.2 m
glycine, 0.2 m Na2CO3 solution and the generated 4-methyl-
umbelliferone determined by measuring the fluorescence at
440 nm after excitation at 365 nm.
UV CD spectroscopy
CD spectroscopy was used to monitor potential distur-
bances of the protein fold upon introduction of the individ-
ual mutations. CD spectra in the range of 250 to 200 nm
were acquired on a JASCO J600 spectropolarimeter with
four accumulations in a 0.1 cm cuvette at 10 �C for solu-
tions of 0.5 mgÆmL)1 of the mutant proteins in 15 mm
sodium phosphate, 100 mm NaCl, pH 7. Data processing
and evaluation was carried out with jasco software as
recommended by the manufacturer.
SPR spectroscopy
Biomolecular interaction analyses (BIA, SPR spectroscopy)
were carried out at 25 �C with a Bialite instrument (Bia-
core) on a Pioneer L1 Chip. For SPR spectroscopy, lipo-
somes were prepared as described above, but diluted to a
lipid concentration of 0.5 mm in NaCl ⁄Pi (10 mm phos-
phate, 140 mm NaCl, 10 mm KCl pH 7.4). These liposomes
contained unlabelled GM2.
First, LUVs (with a total lipid concentration of 0.5 mm),
diluted in NaCl ⁄Pi, were immobilized on the sensorchip by
two injections at a flow rate of 5 lLÆmin)1 (first injection
60 lL, second injection 20 lL) as described previously [12].
This resulted in a RU shift of 5000–7000 RU. Ten micro-
litres of NaOH (25 mm) was then injected at 100 lLÆmin)1
to remove multilamellar structures and to stabilize the base-
line, resulting in a RU shift of ~ 20–50 RU. Sixty micro-
litres of GM2AP (1 or 2 lm) in running buffer (50 mm
sodium citrate buffer, pH 4.2) were injected into the flow
cells at a rate of 20 lLÆmin)1 with a dissociation time of
180 s. Measurements started with the injection of GM2AP.
FRET-based assay for GM2AP
The fluorescent analogue 2-NBD–GM1 was synthesized as
described previously [26]. Large unilamellar donor vesicles
were prepared with the following composition: PtdCho
(56 mol%), Chol (20 mol%), phosphatidic acid (20 mol%),
2-NBD–GM1 (2 mol%) and rhodamine–PE (2 mol%).
Acceptor vesicles comprised PtdCho (60 mol%), Chol
(20 mol%) and phosphatidic acid (20 mol%). The final
donor vesicle concentration in the assay mixture was
20 nmolÆmL)1, the concentration of acceptor vesicles was
100 nmolÆmL)1 in a total volume of 400 lL in 50 mm citrate
buffer, pH 4.2. The transfer of NBD–GM1 was started by
the addition of GM2AP to a final concentration of 37.5 nm
(WT or point mutants). Fluorescence measurements were
performed at 28 �C using a quartz cuvette in a Shimadzu
RF5000 instrument (Kyoto, Japan) with an excitation wave-
length of 480 nm and an emission wavelength of 522 nm.
Without the addition of GM2AP, no increase in fluorescence
was observed over a period of 2 h. For each experimental
time point over the interval ranging from 0.5 to 30 min, the
instrument’s shutter was opened for only 3–4 s, resulting in
a total of maximal 52 s illumination of NBD–GM2. Under
these conditions, photobleaching was negligible.
Acknowledgements
We thank Dr Christina Schuette, Luebeck, for provi-
ding purified HexA, and Dr Joerg Hoernschemeyer,
Loerrach, for performing ESI-TOF-MS. This study
was supported by the Deutsche Forschungsgemeinsc-
haft, SFB 284 and SA-257 ⁄21-1.
References
1 Kolter T & Sandhoff K (1999) Sphingolipids – their
metabolic pathways and the pathobiochemistry of
M. Wendeler et al. Enzyme-binding region of GM2-activator protein
FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS 989

neurodegenerative diseases. Angew Chem Int Ed 38,
1532–1568.
2 Sandhoff K, Kolter T & Harzer K (2001) Sphingolipid
activator proteins. In The Metabolic and Molecular
Bases of Inherited Disease, 8th edn (Scriver C, Beaudet
AL, Sly W & Valle D, eds), pp. 3371–3389. McGraw
Hill, New York.
3 Conzelmann E & Sandhoff K (1979) Purification and
characterization of an activator protein for the degrada-
tion of glycolipids GM2 and GA2 by hexosaminidase
A. Hoppe-Seyler Z Physiol Chem 360, 1837–1849.
4 Gravel RA, Kaback MM, Proia RL, Sandhoff K,
Suzuki K & Suzuki K (2001) The GM2 gangliosi-
doses. In The Metabolic and Molecular Bases of Inher-
ited Disease, 8th edn (Scriver C, Beaudet AL, Sly WS
& Valle D, eds), pp. 3827–3876. McGraw Hill, New
York.
5 Kytzia HJ & Sandhoff K (1985) Evidence for two dif-
ferent active sites on human b-hexosaminidase A. J Biol
Chem 260, 7568–7572.
6 Conzelmann E & Sandhoff K (1978) AB variant of
infantile GM2 gangliosidosis: deficiency of a factor
necessary for stimulation of hexosaminidase A-catalyzed
degradation of ganglioside GM2 and glycolipid GA2.
Proc Natl Acad Sci USA 75, 3979–3983.
7 Platt FM & Walkley SU (2004) Lysosomal defects and
storage. In Lysosomal Disorders of the Brain (Platt FM
& Walkley SU, eds), pp. 32–49. Oxford University
Press, New York.
8 Glombitza GJ, Becker E, Kaiser HW & Sandhoff K
(1997) Biosynthesis, processing, and intracellular trans-
port of GM2 activator protein in human epidermal ker-
atinocytes. The lysosomal targeting of the GM2
activator is independent of a mannose-6-phosphate sig-
nal. J Biol Chem 272, 5199–5207.
9 Hama Y, Li YT & Li SC (1997) Interaction of the
GM2 activator protein with glycosphingolipids. J Biol
Chem 272, 2828–2833.
10 Conzelmann E, Burg J, Stephan G & Sandhoff K
(1982) Complexing of glycolipids and their transfer
between membranes by the activator protein for degra-
dation of lysosomal ganglioside GM2. Eur J Biochem
123, 455–464.
11 Giehl A, Lemm T, Bartelsen O, Sandhoff K & Blume A
(1999) Interaction of the GM2-activator protein with
phospholipid-ganglioside bilayer membranes and with
monolayers at the air–water interface. Eur J Biochem
261, 650–658.
12 Werth N, Schuette CG, Wilkening G, Lemm T & Sand-
hof K (2001) Degradation of membrane-bound ganglio-
side GM2 by beta-hexosaminidase A. Stimulation by
GM2 activator protein and lysosomal lipids. J Biol
Chem 276, 12685–12690.
13 Zhou D, Cantu C 3rd, Sagiv Y, Schrantz N, Kulkarni
AB, Qi X, Mahuran DJ, Morales CR, Grabowski GA,
Benlagha K et al. (2004) Editing of CD1d-bound lipid
antigens by endosomal lipid transfer proteins. Science
303, 523–527.
14 Furst W & Sandhoff K (1992) Activator proteins and
topology of lysosomal sphingolipid catabolism. Biochim
Biophys Acta 1126, 1–16.
15 Li YT, Li SC, Hasegawa A, Ishida H, Kiso M, Bernardi
A, Brocca P, Raimondi L & Sonnino S (1999) Struc-
tural basis for the resistance of Tay-Sachs ganglioside
GM2 to enzymatic degradation. J Biol Chem 274,
10014–10018.
16 Yadao F, Hechtman P & Kaplan F (1997) Formation
of a ternary complex between GM2 activator protein,
GM2 ganglioside and hexosaminidase A. Biochim
Biophys Acta 1340, 45–52.
17 Xie B, Rigat B, Smiljanic-Georgijev N, Deng H &
Mahuran DJ (1998) Biochemical characterization of the
Cys138Arg substitution associated with the AB variant
form of GM2 gangliosidosis: evidence that Cys138 is
required for the recognition of the GM2 ganglioside
complex by hexosaminidase A. Biochemistry 37, 814–
821.
18 Meier EM, Schwarzmann G, Furst W & Sandhoff K
(1991) The human GM2 activator protein – a substrate
specific cofactor of b-hexosaminidase A. J Biol Chem
266, 1879–1887.
19 Zarghooni M, Bukovac S, Tropak M, Calhan J &
Mahuran D (2004) An alpha-subunit loop structure is
required for GM2-activator protein binding by beta-
hexosaminidase A. Biochem Biophys Res Commun 324,
1048–1052.
20 Wright CS, Li SC & Rastinejad F (2000) Crystal struc-
ture of human GM2-activator protein with a novel
b-cup topology. J Mol Biol 304, 411–422.
21 Wright CS, Zhao Q & Rastinejad F (2003) Structural
analysis of lipid complexes of GM2-activator protein.
J Mol Biol 331, 951–964.
22 Wright CS, Mi LZ & Rastinejad F (2004) Evidence for
lipid packaging in the crystal structure of the GM2-acti-
vator complex with platelet activating factor. J Mol Biol
342, 585–592.
23 Wendeler M, Hoernschemeyer J, Hoffmann D, Kolter
T, Schwarzmann G & Sandhoff K (2003) Photoaffinity
labelling of human GM2-activator protein – mechanistic
insight into ganglioside GM2 degradation. Eur J Bio-
chem 271, 614–627.
24 Mark BL, Mahuran DJ, Cherney MM, Zhao D, Knapp
S & James MNG (2003) Crystal structure of human
beta-hexosaminidase B: understanding the molecular
basis of Sandhoff and Tay-Sachs disease. J Mol Biol
327, 1093–1109.
25 Maier T, Strater N, Schuette CG, Klingenstein R,
Sandhoff K & Saenger W (2003) The X-ray crystal
structure of human beta-hexosaminidase B provides new
insights into Sandhoff disease. J Mol Biol 328, 669–681.
Enzyme-binding region of GM2-activator protein M. Wendeler et al.
990 FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS

26 Schwarzmann G, Wendeler M & Sandhoff K (2005)
Synthesis of novel NBD-GM1 and NBD-GM2 for the
transfer-activity of GM2-activator protein by a FRET-
based assay system. Glycobiology 15, 1302–1311.
27 Schuette CG, Lemm T, Glombitza GJ & Sandhoff K
(1998) Complete localization of disulfide bonds in GM2
activator protein. Protein Sci 7, 1039–1045.
28 Wendeler M, Lemm T, Weisgerber J, Hoernschemeyer
J, Bartelsen O, Schepers U & Sandhoff K (2003)
Expression of recombinant human GM2-activator
protein in insect cells: purification and characterization
by mass spectrometry. Prot Expr Purif 27, 259–266.
29 Klima H, Klein A, van Echten G, Schwarzmann G,
Suzuki K & Sandhoff K (1993) Over-expression of a
functionally active human GM2-activator protein in
Escherichia coli. Biochem J 292, 571–576.
30 Kraulis PJ (1991) MOLSCRIPT: a program to produce
both detailed and schematic plots of protein structures.
J Appl Crystallogr 24, 946–950.
M. Wendeler et al. Enzyme-binding region of GM2-activator protein
FEBS Journal 273 (2006) 982–991 ª 2006 The Authors Journal compilation ª 2006 FEBS 991
Related Documents











