The endocannabinoid system in the brain of Carassius auratus and its possible role in the control of food intake M. Valenti,* E. Cottone, R. Martinez,à N. De Pedro,à M. Rubio,à M. P. Viveros,à M. F. Franzoni, M.J. Delgadoà and V. Di Marzo* *Endocannabinoid Research Group, Istituto di Chimica Biomolecolare, C.N.R., Pozzuoli (NA), Italy Dipartimento di Biologia Animale e dell’Uomo, Universita ` degli Studi di Torino, Torino, Italy àDepartamento de Fisiologı ´a (Fisiologı ´a Animal II), Facultad de Biologı ´a, Universidad Complutense, Madrid, Spain Abstract Cannabinoid receptors and the endocannabinoids ananda- mide and 2-arachidonoylglycerol have been suggested to regulate food intake in several animal phyla. Orthologs of the mammalian cannabinoid CB 1 and CB 2 receptors have been identified in fish. We investigated the presence of this endo- cannabinoid system in the brain of the goldfish Carassius auratus and its role in food consumption. CB 1 -like immuno- reactivity was distributed throughout the goldfish brain. The prosencephalon showed strong CB 1 -like immunoreactivity in the telencephalon and the inferior lobes of the posterior hypothalamus. Endocannabinoids were detected in all brain regions of C. auratus and an anandamide-hydrolysing enzy- matic activity with features similar to those of mammalian fatty acid amide hydrolase was found. Food deprivation for 24 h was accompanied by a significant increase of anandamide, but not 2-arachidonoylglycerol, levels only in the telencepha- lon. Anandamide caused a dose-dependent effect on food intake within 2 h of intraperitoneal administration to satiated fish and significantly enhanced or reduced food intake at low (1 pg/g body weight) or intermediate (10 pg/g) doses, respectively, the highest dose tested (100 pg/g) being inac- tive. We suggest that endocannabinoids might variously contribute to adaptive responses to food shortage in fish. Keywords: anandamide, cannabinoid, CB 1 receptor, fish, food intake, receptors. J. Neurochem. (2005) 95, 662–672. The cannabinoid CB 1 receptor (Matsuda et al. 1990) is one of the most abundant G-protein-coupled receptors in the CNS and is believed to be responsible for the majority of the ‘central’ actions of D 9 -tetrahydrocannabinol, the major psychoactive component of Cannabis sativa and marijuana (Gaoni and Mechoulam 1964). D 9 -tetrahydrocannabinol also activates cannabinoid CB 2 receptors, which are also coupled to G-proteins of the G i/o family but are absent in the brain under physiological conditions, when they are almost uniquely found in immune tissues and cells (Galiegue et al. 1995). CB 1 and CB 2 receptors share very little homology, thus pointing to a very early separation of the two encoding genes during animal phylogeny. In fact, the only cannabinoid receptor ortholog fully sequenced in an invertebrate species, the sea squirt Ciona intestinalis (chordata), exhibits low homology with both mammalian subtypes (Elphick et al. 2003). Nevertheless, the orthologs of cannabinoid receptors that have been found so far in non-mammalian vertebrates, including fish, amphibians and birds (McPartland and Glass 2003), are mostly CB 1 orthologs , except for the CB 2 ortholog identified in the fish Fugu rubripes (Elphick 2002) which, contrary to a previous report (Yamaguchi et al. 1996), does not express only CB 1 orthologs. Endogenous ligands of cannabinoid receptors, or endocannabinoids, have been identified in all phyla studied and they are derived from arachidonic acid. The best studied endocannabinoids are N-arachidonoylethanolamide (anandamide) and 2-arachido- noylglycerol (2-AG) (Devane et al. 1992; Mechoulam et al. 1995; Sugiura et al. 1995), which are biosynthesized from Received March 11, 2005; revised manuscript received May 28, 2005 accepted May 29, 2005. Address correspondence and reprint requests to V. Di Marzo, Endo- cannabinoid Research Group, Istituto di Chimica Biomolecolare, C.N.R. Via Campi Flegrei 34, 80078, Pozzuoli (NA), Italy. E-mail: [email protected] Abbreviations used: 2-AG, 2-arachidonoylglycerol; bw, body weight; CB 1 -LI-IR, CB 1 -like immunoreactivity; FAAH, fatty acid amide hydrolase; NArPE, N-arachidonoyl-phosphatidylethanolamine. Journal of Neurochemistry , 2005, 95, 662–672 doi:10.1111/j.1471-4159.2005.03406.x 662 ȑ 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The endocannabinoid system in the brain of Carassius auratusand its possible role in the control of food intake
M. Valenti,* E. Cottone,� R. Martinez,� N. De Pedro,� M. Rubio,�M. P. Viveros,�M. F. Franzoni,�M.J. Delgado� and V. Di Marzo*
*Endocannabinoid Research Group, Istituto di Chimica Biomolecolare, C.N.R., Pozzuoli (NA), Italy
�Dipartimento di Biologia Animale e dell’Uomo, Universita degli Studi di Torino, Torino, Italy
�Departamento de Fisiologıa (Fisiologıa Animal II), Facultad de Biologıa, Universidad Complutense, Madrid, Spain
Abstract
Cannabinoid receptors and the endocannabinoids ananda-
mide and 2-arachidonoylglycerol have been suggested to
regulate food intake in several animal phyla. Orthologs of the
mammalian cannabinoid CB1 and CB2 receptors have been
identified in fish. We investigated the presence of this endo-
cannabinoid system in the brain of the goldfish Carassius
auratus and its role in food consumption. CB1-like immuno-
reactivity was distributed throughout the goldfish brain. The
prosencephalon showed strong CB1-like immunoreactivity in
the telencephalon and the inferior lobes of the posterior
hypothalamus. Endocannabinoids were detected in all brain
regions of C. auratus and an anandamide-hydrolysing enzy-
matic activity with features similar to those of mammalian fatty
acid amide hydrolase was found. Food deprivation for 24 h
was accompanied by a significant increase of anandamide,
but not 2-arachidonoylglycerol, levels only in the telencepha-
lon. Anandamide caused a dose-dependent effect on food
intake within 2 h of intraperitoneal administration to satiated
fish and significantly enhanced or reduced food intake at low
(1 pg/g body weight) or intermediate (10 pg/g) doses,
respectively, the highest dose tested (100 pg/g) being inac-
tive. We suggest that endocannabinoids might variously
contribute to adaptive responses to food shortage in fish.
Keywords: anandamide, cannabinoid, CB1 receptor, fish,
food intake, receptors.
J. Neurochem. (2005) 95, 662–672.
The cannabinoid CB1 receptor (Matsuda et al. 1990) is oneof the most abundant G-protein-coupled receptors in the CNSand is believed to be responsible for the majority of the‘central’ actions of D9-tetrahydrocannabinol, the majorpsychoactive component of Cannabis sativa and marijuana(Gaoni and Mechoulam 1964). D9-tetrahydrocannabinol alsoactivates cannabinoid CB2 receptors, which are also coupledto G-proteins of the Gi/o family but are absent in the brainunder physiological conditions, when they are almostuniquely found in immune tissues and cells (Galiegue et al.1995). CB1 and CB2 receptors share very little homology,thus pointing to a very early separation of the two encodinggenes during animal phylogeny. In fact, the only cannabinoidreceptor ortholog fully sequenced in an invertebrate species,the sea squirt Ciona intestinalis (chordata), exhibits lowhomology with both mammalian subtypes (Elphick et al.2003). Nevertheless, the orthologs of cannabinoid receptorsthat have been found so far in non-mammalian vertebrates,including fish, amphibians and birds (McPartland and Glass
2003), are mostly CB1 orthologs, except for the CB2 orthologidentified in the fish Fugu rubripes (Elphick 2002) which,contrary to a previous report (Yamaguchi et al. 1996), doesnot express only CB1 orthologs. Endogenous ligands ofcannabinoid receptors, or endocannabinoids, have beenidentified in all phyla studied and they are derived fromarachidonic acid. The best studied endocannabinoids areN-arachidonoylethanolamide (anandamide) and 2-arachido-noylglycerol (2-AG) (Devane et al. 1992; Mechoulam et al.1995; Sugiura et al. 1995), which are biosynthesized from
Received March 11, 2005; revised manuscript received May 28, 2005accepted May 29, 2005.Address correspondence and reprint requests to V. Di Marzo, Endo-
cannabinoid Research Group, Istituto di Chimica Biomolecolare, C.N.R.Via Campi Flegrei 34, 80078, Pozzuoli (NA), Italy.E-mail: [email protected] used: 2-AG, 2-arachidonoylglycerol; bw, body weight;
CB1-LI-IR, CB1-like immunoreactivity; FAAH, fatty acid amidehydrolase; NArPE, N-arachidonoyl-phosphatidylethanolamine.
Journal of Neurochemistry, 2005, 95, 662–672 doi:10.1111/j.1471-4159.2005.03406.x
662 � 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

phospholipid precursors to be immediately released from thecell in order to activate their targets and then rapidly removedfrom the extracellular space by selective reuptake into thecell followed by enzymatic hydrolysis (Di Marzo et al.2004). The cannabinoid receptors, the endocannabinoids andthe proteins for their biosynthesis and degradation constitutethe endocannabinoid system (Di Marzo et al. 2004).
Cannabinoid receptor orthologs have been identified inmost vertebrate phyla and in a urochordate species, thesea squirt C. intestinalis (Elphick et al. 2003), whereas ininvertebrates the action of endocannabinoids seems to bemostly mediated by receptors with little or no homology withthe mammalian receptors. In the coelenterate Hydra vulgaris,anandamide inhibits a typical feeding response, i.e. the timeduring which the mouth stays open following exposure toglutathione, a chemical stimulus that mimics exposure to prey.This effect of anandamide is blocked by the CB1 antagonistrimonabant (SR141716A). Importantly, specific binding sitesfor rimonabant, with high affinity also for anandamide,are found in H. vulgaris (De Petrocellis et al. 1999). InC. intestinalis, endocannabinoids are found in cerebralganglia, where specific binding sites for cannabinoid agonistsalso exist. Accordingly, a CB1/CB2 receptor ortholog wasfound in this organism, containing all the amino acidsnecessary for binding to these agonists (Elphick et al. 2003).One such agonist retards the reopening time of theC. intestinalis buccal siphon after mechanical stimulation-induced mouth closure (Matias et al. 2005). Therefore, it ispossible that the endocannabinoid system also controls theneuronal mechanisms that regulate the feeding behavior ofsea squirts.
In non-mammalian vertebrates there is one example of theinvolvement of the endocannabinoid system in the control offeeding behavior. In the songbird zebra finch, brain 2-AGlevels are increased following food deprivation and concurin inhibiting neural processes involved in new songlearning via a CB1 ortholog previously cloned in this bird(Soderstrom et al. 2004). These data suggest that endo-cannabinoids in birds behave as orexigenic mediators, like inmammals. In fact, in rodents, endocannabinoid levels alsoincrease, after short periods of food deprivation, in brainareas involved in the incentive or appetitive aspects of foodintake, i.e. the nucleus accumbens and hypothalamus but notcerebellum (Kirkham et al. 2002), and CB1 receptors arenecessary for optimal food intake after a brief starvation(Di Marzo et al. 2001). The present study was aimed atinvestigating the presence and possible role in food intake ofendocannabinoids in the teleost fish Carassius auratus. First,we looked at the distribution of CB1-like immunoreactivity(CB1-LI-IR) in the forebrain of this fish. Next, we assessedwhether C. auratus brain contains endocannabinoids and iftheir levels vary as observed in birds and rats following fooddeprivation and refeeding. Finally, we studied the effect ofanandamide on food intake.
Materials and methods
Immunohistochemical studies
The experiments were performed under the guidelines established
by the Italian law for animal welfare. Adult specimens of
C. auratus (n ¼ 8) of both sexes were deeply anesthetized with
tricaine methanesulfonate (1 : 1000, MS222, Sandoz Ltd, Basel,
Switzerland). The CNS was removed and fixed for 14 h in
paraformaldehyde (4%) in phosphate buffer (0.1 M, pH 7.4),
cryoprotected, embedded in Killik medium (Bio-Optica, Milan,
Italy) and frozen in liquid isopentane. Coronal sections (12 lmthick) of the CNS, obtained with a cryostat, were mounted on
3-aminopropyl-triethoxysilan-coated slides and stored at 4�C until
use. For molecular biology experiments, the C. auratus CNS was
rapidly dissected out and immediately frozen at )80�C.CNS sections were incubated with an affinity-purified primary
polyclonal antibody raised against the N-terminus of the rat CB1
receptor (a kind gift of Prof. K. Mackie, University of Washington,
Seattle, WA, USA; 1 : 800 dilution in 0.01 M phosphate-buffered
saline, 0.1% Triton-X100). The immunoreactivity was revealed by
the biotin–avidin complex system and H2O2/3,3¢-diaminobenzidine-
tetrahydrochloride as substrate/chromogen. The sections were
observed under a light microscope (Axioskop, Zeiss, Gottingen,
Germany) and then photographed. Images were processed with the
software PhotoShop 7.0 (Adobe Systems Incorporated, Mountain
View, CA, USA). The specificity of the anti-CB1 antibody was
previously assessed in rat (Tsou et al. 1998) and evaluated in
C. auratus by incubating the sections with the anti-CB1 antibody
(1 : 800 dilution) pre-adsorbed for 14 h at 25�C with the immun-
izing fusion protein (1 lg/mL); moreover, the specificity of the
technique was confirmed by omitting the primary antibody.
Nomenclature was mainly referred, with modifications, to Meek
and Nieuwenhuys (1998).
For western immunoblotting, carried out to further assess the
specificity of the antibody used, proteins were extracted from
C. auratus brain following the procedure described by Tsou et al.(1998). Briefly, brains were homogenized in a fivefold (v/w) excess
of a buffer containing 25 mM HEPES, 1 mM EDTA, 6 mM MgCl2and protease inhibitor cocktail (Sigma, St Louis, MO, USA),
pH 7.4. Homogenates were spun for 5 min at 1000 g and the
supernatants were then collected. Pellets were rehomogenized in the
same buffer and centrifuged as above and the supernatants were then
pooled. Alternatively, total proteins were extracted from C. auratusand rat CNS by using a boiling buffer containing 2.5% sodium
dodecyl sulfate and 125 mM Tris-HCl, pH 6.8. The protein
concentration was determined by means of the bicinchoninic acid
technique (Pierce, Rockford, IL, USA); 100 lg of total proteins
were loaded on a 10% polyacrylamide gel, separated by sodium
dodecyl sulfate–polyacrylamide gel electrophoresis and then blot-
ted onto a polyvinylidene difluoride membrane (Amersham
Biosciences, Little Chalfont, UK). Western blotting was performed
by using as a primary antibody an affinity-purified polyclonal
antiserum raised against the N-terminus of the rat CB1 diluted
1 : 500 in Tris-buffered saline, 5% bovine serum albumin. As a
control, the anti-CB1 N-terminus antibody (1 : 500 dilution) was
pre-adsorbed for 14 h at 25�C with the immunizing fusion protein
(10 lg/mL). After the incubation with an anti-rabbit IgG horseradish
peroxidase-linked antiserum, the reaction was revealed with the
Endocannabinoid system in brain of Carassius auratus 663
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

ECL Plus Western blotting detection reagent and Hyperfilm-ECL
autoradiography film (Amersham Biosciences). The GelDoc gel
documentation system and the software Quantity One (Bio-Rad
Laboratories, Hercules, CA, USA) were used to determine the
molecular weight of the specific CB1 band on the basis of the
migration of the Precision plus protein dual color molecular weight
standards (Bio-Rad Laboratories).
Sequencing of a PCR cDNA fragment from Carassius auratus
mRNA encoding a CB1-like receptor
Total RNA was extracted from Ca. auratus CNS using the TRIZOLreagent (Life Technologies, Rockville, MD, USA) and following the
manufacturer’s instructions. DNA contaminants were eliminated by
digestion with RQ1 RNase-free DNase (Promega, Madison, WI,
USA). cDNA was synthesized from total RNA using M-MLV
reverse transcriptase (USBiological, Swampscott, MA, USA) and
random examers. A degenerate primer PCR was performed using
RedTaq DNA polymerase (Sigma) in the presence of 1.5 mM MgCl2at an annealing temperature of 50�C for 30 cycles. Carassiusauratus CB1 was amplified using a 5¢ sense degenerate primer [5¢-TACCACTTCAT(T/C/A)GGCAGC(T/C)T-3¢] and a 3¢ antisense
degenerate primer [5¢-(A/G)CA(G/A)AA(G/C/T)GCAAA(G/C/A)A(T/C)(C/A)GTCT-3¢] complementary to the sequences codifying
for regions of the second and seventh transmembrane domains of rat
CB1, respectively (see also Cottone et al. 2005). The 690-bp
amplification product was cloned into pGEM-T-easy vector using
the pGEM-T-easy Vector System (Promega). JM109 high efficiency
competent cells were transformed and recombinant colonies
were identified by blue/white color screening and restriction
digestion; selected recombinant clones (pGEM-T-easy-CB1Car)were sequenced. The deduced nucleotide sequence of the cloned
C. auratus CB1 650-bp fragment is available at GenBank
(Accession no. AY674057). In order to establish the degree of
identity of the cannabinoid receptors among the different species,
both the nucleotide and the amino acid sequence of C. auratus CB1
were aligned with the CB1A sequence of F. rubripes available in the
GenBank database using ClustalW multiple alignment computer
programs. The BOX-SHADE alignment graphics program was used
to highlight amino acid sequence similarities.
Determination of endocannabinoid levels
Goldfish [34.68 ± 1.63 g average body weight (bw)], fed with a
daily ration of 1% bw food, were divided into three experimental
groups (n ¼ 12 goldfish/group): (1) control group, receiving food in
excess 2 h before killing; (2) fasting group, deprived of food for 24 h
and (3) fasting and refed group, deprived for 24 h and receiving food
in excess for 2 h before killing. Fish were killed by decapitation and
the cerebellum, telencephalon and hypothalamus were dissected on
ice and immediately stored at )80�C until extracted as described
previously (Matias et al. 2003). Briefly, tissues were homogenized in
and extracted with chloroform : methanol : Tris-HCl (50 mM,
pH 7.5, 2 : 1 : 1, v/v) containing internal standards (10 pmol of
[2H]8 anandamide, [2H]4 N-palmitoylethanolamine and 100 pmol of
[2H]5-2-arachidonyl glycerol, obtained from Cayman Chemicals,
Ann Arbor, MI, USA). The lipid-containing organic phase was
collected and dried. The mass of dried extract was weighed
accurately with an analytical balance in 1.5-mL Eppendorf tubes
and pre-purified by open-bed chromatography on silica gel. Fractions
eluted with 9 : 1 chloroform : methanol were analysed by liquid
chromatography-atmospheric pressure chemical ionization-mass
spectrometry using an HPLC apparatus coupled to an LCMS-2010
quadruple mass spectrometry via an atmospheric pressure chemical
ionization interface (all Shimadzu, Tokyo, Japan). Mass spectrom-
etry analyses were carried out in the selected ion-monitoring as
described previously (Di Marzo et al. 2001). The temperature of the
atmospheric pressure chemical ionization source was 400�C and the
HPLC column was a Phenomenex (5 mm, 150 · 4.5 mm) reverse-
phase column, eluted as previously described (Di Marzo et al. 2001).Anandamide (retention time 13.5 min) and 2-AG (retention time
15.0 min) quasi-molecular ions (m/z ¼ 348.0 and 379.3) were
quantified by isotope dilution with the above-mentioned deuterated
standards (same retention times and m/z ¼ 356.0 and 384.3) and
their amounts in picomoles or nanomoles normalized per milligram
of lipid extract. Two liquid chromatography-mass spectrometry
peaks for both deuterated and undeuterated mono-arachidonoylglyc-
erol were found at retention times of 15.0 and 16.9 min, respectively,
corresponding to 2-AG and 1(3)-orechidenoyl-glycerol, in agree-
ment with the previous observation that 2-AG undergoes isomeri-
zation during the purification procedure. Therefore, the amounts of
2-AG were calculated by adding the amounts of the two isomers.
Silica column fractions eluted with 1 : 1 chloroform : methanol,
which contain the N-acyl-phosphatidylethanolamines, were proc-
essed with Streptomyces chromofuscus phospholipase D to yield the
corresponding N-acylethanolamines in a quantitative manner, as
described previously (Di Marzo et al. 2001). Therefore, the
anandamide biosynthetic precursor N-arachidonoyl-phosphatidyleth-anolamine (NArPE) was quantified by measuring the amounts,
by means of liquid chromatography-atmospheric pressure chemical
ionization-mass spectrometry, of the anandamide produced from its
hydrolysis by the phospholipase D.
Assays for anandamide hydrolysing activity
These assays were carried out as described previously (De
Petrocellis et al. 1999) using [14C]ethanolamine-labeled ananda-
mide as substrate and membranes from C. auratus whole brain
prepared as described previously (De Petrocellis et al. 1999).
Membranes were incubated at 37�C either with increasing concen-
trations of substrate (1, 4, 10, 50 and 100 lM) at pH 9.0, with 4 lMsubstrate in buffers at different pH (De Petrocellis et al. 1999) or inthe presence of two distinct inhibitors of mammalian fatty acid
amide hydrolase (FAAH), OL-135 and arachidonoylserotonin (see
Di Marzo et al. 2004 for a review). Following the incubation for
30 min, the assay mixture was extracted with chloroform : meth-
anol (2 : 1 by volume) and the amount of [14C]ethanolamine
released into the aqueous phase was used to quantify the
anandamide amidase activity.
Feeding experiments
Four groups of satiated goldfish (13.9 ± 0.5 g bw, n ¼ 8 goldfish/
group) were intraperitoneally injected with 10 lL vehicle/g bw at the
following doses of anandamide: 0, 1, 10 or 100 pg/g bw. Fish were
anesthetized before the i.p. injection in water containing tricaine
methanesulfonate (MS-222, 1/10 000) and the injections were
performedwhen loss of equilibriumwas observed. Individual goldfish
were transferred to 5-L aquaria. Fish recovered equilibrium and
normal swimming activity in anesthetic-free water within 1–2 min
664 M. Valenti et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

of the injections. Once recovered, the animals received pre-weighed
food in excess (5% bw). Food intake (FI) was measured for 2 h
post-injection and calculated as follows: FI ¼ Wi ) (Wf · F), whereWi ¼ initial dry food weight and Wf ¼ remaining dry food weight.
F was previously calculated in the absence of fish to determine the
reduction in the weight of food pellets due to water dissolution after 2
and 6 h in the aquaria (Lopez-Patino et al. 1999).
Fish cortisol level and activity measurements
Two groups (n ¼ 8/group) of satiated goldfish (30.1 ± 0.9 g bw)
were injected i.p. with either vehicle (10 lL/g bw) or anandamide
(10 pg/g bw). The vehicle composition was (ethanol : cremo-
phor : saline) (1 : 1 : 18), with fish saline being made up of
20 mg Na2CO3/100 mL of 0.6% NaCl. The cremophor was from
Fluka (Buchs, Switzerland). Blood samples from caudal veins
were collected at 2 h post-injection and plasma cortisol levels
were measured in duplicate by radioimmunoassay using a
commercial kit (DSL 2000, Diagnostic Systems Laboratories,
Sinsheim, Germany). The validity of this radioimmunoassay has
been previously determined for cortisol titres in goldfish plasma
(De Pedro et al. 1997). Locomotor activity was recorded
automatically by two infrared sensors located on one side of
the aquaria. Every time the fish interrupted light beams it
generated an output signal that was transferred via an interface to
a computer that recorded and stored the data at 10-min intervals.
Results
Carassius auratus brain expresses CB1-like receptors
We have analysed the distribution of the CB1-LI-IR inC. auratus CNS by means of a primary antibody raisedagainst the CB1 receptor amino-terminus. The specificity ofsuch an antibody has already been demonstrated in rats (Tsouet al. 1998). Specificity controls performed on goldfish brainsections, employing the CB1 antiserum pre-adsorbed withthe immunizing protein [Fig. 1(c) to be compared with theCB1-LI-IR shown in Fig. 1(b)] or omitting the primaryantibody, resulted in the complete absence of immuno-staining. Moreover, western-blotting analysis performedusing the same antibody revealed a CB1-like receptor as aprotein with an apparent molecular weight of �53 kDa,identical to one of the two bands found in the brain of the rat(Fig. 1j); the specific band was not detected when incubatingthe blot with the antibody pre-adsorbed with the immunogenand it corresponds to the molecular weight of non-modifiedmammalian CB1 receptors. The following immunohisto-chemical observations concern the prosencephalic areasreported as involved in the fish feeding response.
TelencephalonThe CB1-LI-IR was distributed through all of the goldfishprosencephalon, including the olfactory bulbs. In particular,in the telencephalon the immunostaining is observed in cellbodies (15–20 lm diameter) and nerve terminals which arealmost dispersed in the central portion of the dorsal area
(Figs 1d and e) and through the ventral area. Proceedingcaudally, plentiful immunopositive varicosities are observedin the ventral area surrounding negative perikarya, as shownin Fig. 1(f).
HypothalamusA strong CB1-LI-IR (more intense than that described in anyother brain area) is seen in the inferior lobes of the posteriorhypothalamus (Fig. 1g). Several labeled cells are neuronsseemingly in contact with cerebrospinal fluid and sendingtheir intraventricular processes into the lateral recess of thethird ventricle (Fig. 1h), while the cell bodies and nerveterminals more deeply located within the hypothalamic wallpossibly belong to the so-called nucleus diffusus of thehypothalamic inferior lobes (Fig. 1g). In the dorsal part ofsuch a nucleus a dense CB1-LI-IR innervation characterizedby thin punctuate terminations is shown in Fig. 1(i).
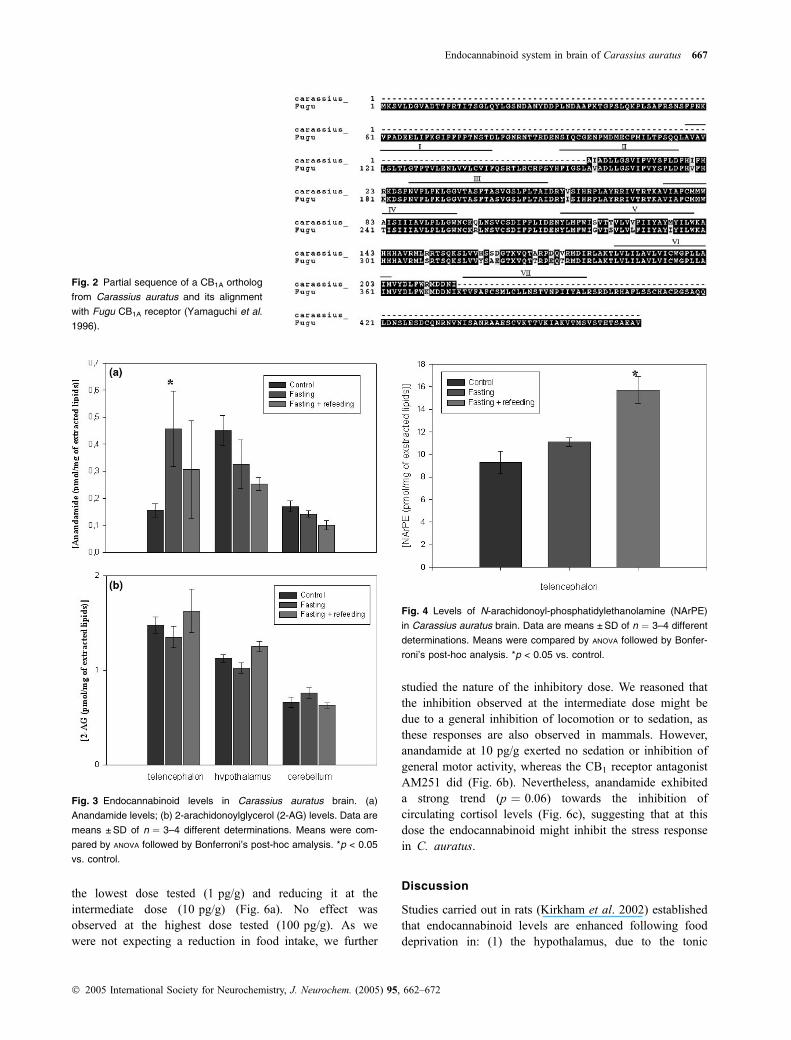
Partial sequence of a Carassius auratus CB1 ortholog
Carassius auratus mRNAwas found here to amplify by PCRa transcript whose partial sequence yielded an amino acidsequence (GenBank no. AY674057) that was almost iden-tical to that of the previously reported Fugu CB1A receptor(Yamaguchi et al. 1996) between amino acids 189 and 374(Fig. 2). In particular, residues necessary for the interactionwith cannabinoid receptor ligands (McPartland and Glass2003), such as the 263–268 CSXXFP sequence in FuguCB1A important for binding CP55,940 or other residuesinvolved in the binding with anandamide, THC, HU-210 andCP55,940 (e.g. residues F188 and F190) or for CB1
functional activity (e.g. residues F199, Y275 and W299),were all conserved.
Endocannabinoid levels in Carassius auratus
Both anandamide and 2-AG were detected in all brainregions of C. auratus. Anandamide was highest in thehypothalamus (p < 0.05), whereas 2-AG was highest in thetelencephalon (p < 0.05) (Figs 3a and b). Food deprivationwas accompanied by a significant, more than twofold,increase of anandamide but not 2-AG levels, but only inthe telencephalon (Figs 3a and b). However, refeeding didnot significantly reduce anandamide levels in this region,although after this treatment the concentration of anandamidein the telencephalon was not significantly different fromcontrol fish. As anandamide was the only endocannabinoidwhose levels changed during food deprivation, we alsoanalysed the levels of its biosynthetic precursor NArPE. Thiscompound was again detected in all brain regions of controlC. auratus (data not shown), thus suggesting that in thisfish anandamide is produced through the same pathway asin mammals. As previously observed in mammals, thelevels of NArPE were at least one order of magnitude higherthan anandamide levels (Fig. 4), as expected from theprecursor–product relationship between the two compounds.
Endocannabinoid system in brain of Carassius auratus 665
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

Food deprivation was accompanied by a non-statisticallysignificant increase of NArPE in the telencephalon andrefeeding resulted in a further increase of this compound(Fig. 4), thus possibly explaining why anandamide levelswere not reduced to normal after refeeding.
Carassius auratus brain contains a fatty acid amide
hydrolase-like enzyme
Membranes prepared from C. auratus whole brainwere found to actively hydrolyse [14C]-anandamide to[14C]-ethanolamine in a pH-dependent manner. Maximalactivity was found between pH 9 and 10 and a relative peakof activity at pH 6 (Fig. 5a), exactly as in the case of
mammalian FAAH. The enzyme followed Michaelis-Mentenkinetics, as indicated by the Linewaver-Burk curve shown inFig. 5(b) and exhibited an apparent Km of 16.6 ± 1.9 lM andapparent Bmax of 0.3 ± 0.05 nmol/mg/min (means ± SEM,n ¼ 3). Finally, two selective FAAH inhibitors, OL-135(10 nM) and arachidonoylserotonin (10 lM), inhibited thehydrolysis of [14C]-anandamide at pH 9 by 35.4 ± 5.1 and55.0 ± 6.2%, respectively (means ± SEM, n ¼ 3).
Effect of anandamide on food intake, locomotor activity
and plasma cortisol levels
Anandamide, injected intraperitoneally in C. auratus,exerted a dual effect on food intake by stimulating it at
Fig. 1 Immunochemical detection of a CB1-like receptor in Carassius
auratus brain. (a) Line drawing of the goldfish brain, lateral view.
(b) Coronal section showing thin CB1-like-immunoreactive thin nerve
terminals in the mesencephalic tegmentum. (c) The same area, in a
section consecutive to (b), after staining with the primary antibody
pre-adsorbed with the immunogen; no immunolabelings are observed.
(d) Schematic representation of a coronal section of the telencepha-
lon, at the level indicated in (a) (vertical bar), showing CB1-like-
immunolabeled neurons (d) and terminals (small dots) in the
telencephalon. (e) CB1-immunolabeled neurons in the central portion
of the dorsal area at the border with the ventral area of the telen-
cephalon [coronal section at the level schematized in (d)]. (f) Besides
CB1-immunolabeled neurons, a number of positive nerve terminals
and varicosities surrounding negative perikarya (arrow) are shown in
the ventral area of the telencephalon [coronal section at a more
caudal level than (d)]. (g) Schematic representation of a coronal
section of the hypothalamus, at the level indicated in (a), showing
CB1-like-immunolabeled neurons (d) and terminals (small dots)
through the hypothalamus. (h) CB1-like-immunopositive ependymal
cells and liquor contacting neurons sending their periventricular pro-
cesses (arrows) into the lateral recess of the third ventricle [coronal
section at the level schematized in (g)]. Immunostained cell bodies are
also seen in the subependymal layers of the periventricular gray. (i)
Abundant CB1-immunostained varicosities through the nucleus diffu-
sus (dorsal part) of the hypothalamus inferior lobe. (j) Western
immunoblot of the proteins from the brain of the rat and C. auratus
(using two protein extraction protocols, see text). The image from gels
exposed to the antibody pre-adsorbed with immunogen is also shown.
Calibration bars: b and c, 100 lm; e, f, h and i, 50 lm. DC, area
dorsalis telencephali pars centralis; DD, area dorsalis telencephali
pars dorsalis; M, Mauthner’s fiber; mv, mesencephalic ventricle; NDLI,
nucleus diffusus lobi inferioris; NG, nucleus glomerulosus; OT, optic
tectum; PC, posterior commissure; RL, recessus lateralis; VC, valvula
cerebelli; VD, area ventralis telencephali pars dorsalis; VL, area
ventralis telencephali pars lateralis; VV, area ventralis telencephali
pars ventralis.
666 M. Valenti et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672
the lowest dose tested (1 pg/g) and reducing it at theintermediate dose (10 pg/g) (Fig. 6a). No effect wasobserved at the highest dose tested (100 pg/g). As wewere not expecting a reduction in food intake, we further
studied the nature of the inhibitory dose. We reasoned thatthe inhibition observed at the intermediate dose might bedue to a general inhibition of locomotion or to sedation, asthese responses are also observed in mammals. However,anandamide at 10 pg/g exerted no sedation or inhibition ofgeneral motor activity, whereas the CB1 receptor antagonistAM251 did (Fig. 6b). Nevertheless, anandamide exhibiteda strong trend (p ¼ 0.06) towards the inhibition ofcirculating cortisol levels (Fig. 6c), suggesting that at thisdose the endocannabinoid might inhibit the stress responsein C. auratus.
Discussion
Studies carried out in rats (Kirkham et al. 2002) establishedthat endocannabinoid levels are enhanced following fooddeprivation in: (1) the hypothalamus, due to the tonic
Fig. 2 Partial sequence of a CB1A ortholog
from Carassius auratus and its alignment
with Fugu CB1A receptor (Yamaguchi et al.
1996).
Fig. 3 Endocannabinoid levels in Carassius auratus brain. (a)
Anandamide levels; (b) 2-arachidonoylglycerol (2-AG) levels. Data are
means ±SD of n ¼ 3–4 different determinations. Means were com-
pared by ANOVA followed by Bonferroni’s post-hoc amalysis. *p < 0.05
vs. control.
Fig. 4 Levels of N-arachidonoyl-phosphatidylethanolamine (NArPE)
in Carassius auratus brain. Data are means ±SD of n ¼ 3–4 different
determinations. Means were compared by ANOVA followed by Bonfer-
roni’s post-hoc analysis. *p < 0.05 vs. control.
Endocannabinoid system in brain of Carassius auratus 667
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

inhibitory effect exerted by leptin on endocannabinoid levelsin this region (Di Marzo et al. 2001), which is mostlyinvolved in the appetite-inducing behavior that follows fooddeprivation and (2) the limbic forebrain, which contains thenucleus accumbens and where endocannabinoids appear toplay a reinforcing function on the motivational aspects offood intake. Accordingly, injection of endocannabinoids inthe nucleus accumbens and hypothalamus enhances foodintake (Jamshidi and Taylor 2001; Kirkham et al. 2002).Also in a bird species a brief period of food deprivationcauses a significant enhancement of endocannabinoid levels,although only in brain regions involved in the motivationalaspects of food consumption and in the learning of songs(Soderstrom et al. 2004). In this study we investigated thepresence of the endocannabinoid system in the goldfishC. auratus and addressed the possibility that this system
plays a role in the physiology of feeding behaviors in fishafter a brief period of food deprivation.
We studied cannabinoid CB1 receptor distribution inC. auratus brain. Although both CB1 and CB2 orthologshave been found in fish, we focused on CB1 receptors asthese are the only cannabinoid receptor subtype involved infood intake in mammals. Indeed, we amplified and
Fig. 5 Preliminary characterization of an anandamide hydrolase
activity in Carassius auratus brain. (a) Lineweaver-Burk curve for
substrate dependence of enzymatic activity, calculated using 100 lg
of proteins. From this curve it was possible to calculate an apparent
Km of 16.6 lM and an apparent Bmax of 0.3 nmol/mg/min. (b) pH
dependence of the amidase activity, calculated using a 4 lM
concentration of substrate and 100 lg of proteins.
Fig. 6 (a) Food intake (mg) after intraperitoneal administration of
anandamide [1, 10 and 100 pg/g body weight (bw)] at 2 h post-injec-
tion (n ¼ 8/group) in goldfish. (b) Effect of intraperitoneal administra-
tion of anandamide (10 pg/g bw) and AM251 (1 lg/g bw) on total
motor activity in goldfish (n ¼ 6/group). (c) Plasma cortisol levels (lg/
dL) at 2 h post-injection in goldfish (n ¼ 8/group) intraperitoneally
injected with vehicle (control) or anandamide (10 pg/g bw). Data
are expressed as means ±SEM. Means were compared by
ANOVA followed by Student-Newman-Keuls post-hoc comparison.
**p < 0.001, *p < 0.05 vs. control.
668 M. Valenti et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

sequenced a large part of the encoding region of a FuguCB1A gene ortholog that contained all of the amino acidresidues that are most conserved between mammalian CB1
and Fugu CB1A receptors. Our observations largely confirmthe description of CB1 immunostaining recently reported inthe brain of another bony fish, the cichlid Pelvicachromispulcher (Cottone et al. 2005), although in the present studymost attention was focused on the CB1-LI-IR distributedthroughout the goldfish prosencephalic areas known to beinvolved in the fish feeding control. In particular, we foundan abundant CB1-LI-IR in the central and ventral areas of thetelencephalon as well as in the hypothalamic inferior lobessurrounding the third ventricle lateral recesses. For arelatively long time the interest of comparative neuroscien-tists has been focused on fish brain areas involved in thefeeding response, although neural mechanisms underlyingfood intake are still under investigation. Electrophysiologicalstudies (Demski and Knigge 1971; Demski 1973) pointedto the bony fish hypothalamus, near the lateral recess ofthe third ventricle, as an important area responsible for thecontrol of feeding behavior. Furthermore, lesions of thelateral areas of the hypothalamus were found to causeaphagia in C. auratus (Roberts and Savage, 1978). However,teleost fish possess an elaborate telencephalon with multiplemigrated nuclear groups and a telencephalic involvement incomplex behavioral patterns has been demonstrated byelectrophysiology. Feeding responses have, in fact, beenevoked by electrical stimulation of the medioventral part ofthe telencephalic hemisphere (Demski and Northcutt 1983).There is evidence indicating that several neuropeptidesinvolved in feeding regulation in fish show a wide distribu-tion in prosencephalic areas (De Pedro and Bjornsson 2001).Recently, it has been suggested that neural circuits neuro-chemically identified within the telencephalon and pre-opticarea, such as for example the prosencephalic neuropeptide Yinnervation, are deeply implicated in goldfish appetite control(Narnaware et al. 2000) which, in fish as in mammals, isunder multifactorial regulation (Lin et al. 2000). On the otherhand, the present data show that CB1-LI-IR is found inalmost the same prosencephalic areas where a well-devel-oped neuropeptide Y neuronal system was described inC. auratus by immunohistochemistry (Pontet et al. 1989). Ingoldfish, using a pharmacological approach, neuropeptide Ywas demonstrated to be a physiological brain orexigenicsignal as in mammals (Lopez-Patino et al. 1999) and thisstimulatory effect on feeding seems to be dependent oninteractions with other neuropeptides (De Pedro et al. 2000;Bernier and Peter 2001; Volkoff and Peter 2001). Moreover,neuropeptide Y gene expression in goldfish prosencephalonis influenced by the composition of the diet (Narnaware andPeter 2002). In summary, we have shown here that CB1-likereceptors are expressed in all those brain areas of C. auratuspreviously suggested to intervene in the control of foodintake.
We have also reported here the capability of a fish speciesto synthesize brain endocannabinoids. During the prepar-ation of this work, the presence of anandamide and 2-AGwas recently reported also in the brain of another fishspecies, Pimephales promelas (Rademacher et al. 2005).Although these findings were predictable, due to thereported presence of the two major endocannabinoids inall animal phyla investigated so far, we also observed herethat the levels of anandamide, but not 2-AG, are augmentedby 2.5-fold after food deprivation in the telencephalon butnot in the hypothalamus nor in a brain area, the cerebellum,which is not directly involved in the control of food intake.In rats following brief food deprivation, anandamide levelswere found to change in the brain region, the limbicforebrain, which, like the telencephalon in fish, is involvedmore in the motivational than appetitive aspects of foodintake (Kirkham et al. 2002), whereas the other endocann-abinoid, 2-AG, is also increased in the hypothalamus.However, as outlined above, in fish (and, to some extent,also in mammals), there is no clear distinction in the roleplayed by these two brain areas in the control of energybalance. While in the rat endocannabinoid levels return tobasal levels during normal food consumption (Kirkhamet al. 2002), anandamide levels in the goldfish telencephalonafter refeeding were found here to decrease non-signifi-cantly, although they were not significantly different fromlevels in goldfish fed ad libitum. Apart from obviousdifferences between the two phyla, it is possible that such adiscrepancy is due to the different food deprivation/refeed-ing protocols used in the two studies. Also, the levels ofanandamide in the telencephalon of refed goldfish werecharacterized by marked variability, which may haveprevented us from observing a significant difference fromthe levels in either food-deprived or ad libitum-fed fish. Wealso found that the levels of the anandamide biosyntheticprecursor NArPE were increased after refeeding and not, asexpected, decreased, thus possibly buffering anandamidelevels against a stronger decrease following energy replen-ishment in C. auratus. In turn, the levels of fatty acidprecursors necessary for arachidonic acid and N-arachido-noylphosphatidylethanolamine biosynthesis are likely to beaugmented after refeeding, thus explaining why this condi-tion enhanced, rather than diminished, the tissue concentra-tion of the anandamide precursor.
Having shown that CB1-like receptors, and increasedanandamide concentrations to activate them, are present in aC. auratus brain region involved in the control of foodintake, we needed to demonstrate that anandamide doesaffect energy intake in this fish. Indeed, we found that, at thelowest dose tested, the endocannabinoid causes more than atwofold stimulation of food consumption over 2 h fromadministration. The dose of 1 pg/g which exerted this effectis certainly attainable in vivo following a food deprivationeven shorter than that used in this study (see above). At a
Endocannabinoid system in brain of Carassius auratus 669
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

higher dose (10 pg/g), anandamide suppressed food intake.This rather surprising effect is not likely to be due to asedative effect or to a non-selective impairment of fish motoractivity as it could be assessed by simply monitoring fishmotor functions. However, it must be emphasized that we didnot measure any motor parameter of specific relevance tofeeding (e.g. oromotor control and coordination). In mice,very low doses of systemic anandamide are sufficient toinduce appetite (Hao et al. 2000), whereas another putativeendocannabinoid, noladin ether, also exhibits a biphasicdose–response curve on food intake and bw (Avraham et al.2005). It is tempting to speculate that brain endocannabinoidlevels are regulated, following food shortage, in a way thatthey first enhance food intake in order to re-establish energyhomeostasis; subsequently, in the presence of persistent fooddeprivation (which possibly leads to even higher endocann-abinoid levels), their role may be to help the organism toadapt to this condition by recovering from the stress causedby the lack of energy and, at the same time, by inhibiting thedrive to eat. Some support for this hypothesis comes from theobservation that the anorexic dose of anandamide exhibited astrong trend towards inhibiting plasma cortisol levels, whichwould be in agreement with a similar effect recentlydescribed for 2-AG on corticosterone serum levels in rats(Patel et al. 2004). Another endocannabinoid-based adaptivemechanism reported in rodents and their offspring might berepresented by the reduction, rather than enhancement, ofhypothalamic endocannabinoid levels that follows prolonged(i.e. several days) semistarvation (Hanus et al. 2003; Matiaset al. 2003), which again, by reducing the need to consumefood, might help the animal to adapt to food shortage. Wehave no evidence other than that reported here to support apossible dual role of endocannabinoids following fooddeprivation, as we used only one type of protocol of limitedaccess to food and could not test the effect of anandamide infood-deprived fish. However, it is clear that the endocanna-binoid system plays important roles in both the control ofenergy intake and the recovery from stress, two conditionsthat may be strictly related or even compete with each otherduring food shortage (Di et al. 2003). It is likely that one ofthese two roles of the endocannabinoids may over-ride theother depending on the type and duration of food deprivation.
It should be underlined that, at the highest dose tested,anandamide exerted no overt effect on food intake inC. auratus. The only explanation that we can provide forthis phenomenon is that, at high doses, anandamide becomesa substrate for a hydrolysing enzyme that we have identifiedin a fish species for the first time in this study. This enzymeexhibits features similar to those of mammalian FAAH, i.e.an optimal alkaline pH and sensitivity to arachidonoylsero-tonin and OL-135, two selective FAAH inhibitors, thussuggesting that the C. auratus anandamide amidase might bestructurally similar to FAAH. However, we provided nomolecular data to support this suggestion. Importantly, we
found that the apparent Km for anandamide of this enzyme isvery similar to values found for FAAH enzymes in mammalsand is significantly higher than the concentrations probablyattained in C. auratus following the injection of anandamide.However, it is possible that, at the highest dose tested,anandamide does become a substrate for this enzyme and issubsequently inactivated.
Although we have shown here the presence of CB1-likereceptors and of endogenous cannabinoid ligands in areas ofC. auratus deputed to the control of food intake and havedescribed the effect on food consumption of the onlyendocannabinoid whose levels change during food depriva-tion, we have not provided any evidence that this latter effect isdue to the interaction of anandamide with CB1-like receptors.The part of the sequence of the Ca. auratus CB1A orthologidentified here does contain all the highly conserved aminoacids known to be necessary for a functional interaction ofligands (including anandamide) with the mammalian CB1
receptor (McPartland and Glass 2003) and is therefore verylikely to be activated by anandamide.We also performed someexperiments with the CB1 receptor antagonists SR141716Aand, particularly, AM251 but could not find an inactive dose ofthese compounds that could also reverse the effects ofanandamide on food intake. In fact, AM251 strongly reducedfood intake in C. auratus (data not shown), which is to beexpected if endocannabinoids tonically stimulate appetite infish and is in agreement with thewidely reported effects of CB1
antagonists in mammals (see Black 2004 for review). How-ever, we found that AM251 also inhibits goldfish generalmotor activity and we could not separate this effect of theantagonist from its anorexic action (although, again,we did notmeasure more specific motor parameters that are of directrelevance to feeding behaviors). In summary, also in view ofthe several molecular targets that have been proposed foranandamide inmammals, we cannot assert conclusively that itsdual effects onC. auratus food intake, described above, are dueto activation of theCB1-like receptors detected here in the brainof this fish nor that endocannabinoids tonically stimulateenergy intake in this teleost.
In conclusion, we have reported here that the brain of thegoldfish C. auratus contains CB1-like receptors and putativeendogenous ligands for these receptors. The distribution ofCB1 immunoreactivity in the brain and the type of changesobserved following food deprivation for one of the two majorendocannabinoids, anandamide, point to a role of the endo-cannabinoid system in the control of energy intake in fish, aspreviously also suggested in birds and as widely demonstratedin mammals. The dual effect of anandamide on food intakesuggests that this endogenous compound may not only act tore-establish energy balance but also help goldfish copewith thelack of food, depending on whether or not this becomesavailable again. Further studies are necessary in order toinvestigate whether anandamide effects in C. auratus areindeed mediated by CB1-like receptors.
670 M. Valenti et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

Acknowledgements
The authors are grateful to Prof. Ken Mackie (University of
Washington, Seattle, WA, USA) for the gift of CB1 antibodies.
Financial support from the Italian Ministry of University, Research
and Technology [COFIN no. 2003059955-002(2003) and ex-60%
2005 to MFF] and from the Spanish Ministry of Science and
Technology (project AGL2004-08137-C04-01 to MJD) is acknow-
ledged.
References
Avraham Y., Ben Menachem A., Okun A., Zlotarav O., Abel N.,Mechoulam R. and Berry E. M. (2005) Effects of the endo-cannabinoid noladin ether on body weight, food consumption,locomotor activity, and cognitive index in mice. Brain Res. Bull.65, 117–123.
Bernier N. J. and Peter R. E. (2001) Appetite-suppressing effects ofurotensin I and corticotropin-releasing hormone in goldfish(Carassius auratus). Neuroendocrinology 73, 248–260.
Black S. C. (2004) Cannabinoid receptor antagonists and obesity. Curr.Opin. Invest. Drugs 5, 389–394.
Cottone E., Forno S., Campantico E., Guastalla A., Viltono L., MackieK. and Franzoni M. F. (2005) Expression and distribution of CB1cannabinoid receptors in the central nervous system of the Africancichlid fish Pelvicachromis pulcher. J. Comp. Neurol. 485, 293–303.
De Pedro N., Alonso-Gomez A. L., Gancedo B., Valenciano A. I.,Delgado M. J. and Alonso-Bedate M. (1997) Effect of a-helical-CRF[9)41] on feeding in goldfish: involvement of cortisol andcatecholamines. Behav. Neurosci. 111, 398–403.
De Pedro N. and Bjornsson B. T. (2001) Regulation of food intake byneuropeptides and hormones, in Food Intake in Fish (Houlihan D.,Boujard T. and Jobling M., eds), Chap. 12, pp. 269–296. BlackwellScience, Oxford.
De Pedro N., Lopez-Patino M. A., Guijarro A. I., Pinillos M. L., DelgadoM. J. and Alonso-Bedate M. (2000) NPY receptors and opioidergicsystem are involved in NPY-induced feeding in goldfish. Peptides21, 1495–1502.
De Petrocellis L., Melck D., Bisogno T., Milone A. and Di Marzo V.(1999) Finding of the endocannabinoid signalling system in Hydra,a very primitive organism: possible role in the feeding response.Neuroscience 92, 377–387.
Demski L. S. (1973) Feeding and aggressive behavior evoked byhypothalamic stimulation in a cichlid fish. Comp. Biochem.Physiol. 44, 685–692.
Demski L. S. and Knigge K. M. (1971) The telencephalon and hypo-thalamus of the bluegill (Lepomis macrochirus): evoked feeding,aggressive and reproductive behavior with representative frontalsections. J. Comp. Neurol. 143, 1–16.
Demski L. S. and Northcutt R. G. (1983) The terminal nerve: a newchemosensory system in vertebrates?. Science 220, 435–437.
Devane W. A., Hanus L., Breuer A., Pertwee R. G., Stevenson L. A.,Griffin G., Gibson D., Mandelbaum A., Etinger A. and MechoulamR. (1992) Isolation and structure of a brain constituent that binds tothe cannabinoid receptor. Science 258, 1946–1949.
Di S., Malcher-Lopes R., Halmos K. C. and Tasker J. G. (2003) Non-genomic glucocorticoid inhibition via endocannabinoid release inthe hypothalamus: a fast feedback mechanism. J. Neurosci. 23,4850–4857.
Di Marzo V., Bifulco M. and De Petrocellis L. (2004) The endocann-abinoid system and its therapeutic exploitation. Nat. Rev. DrugDiscov. 3, 771–784.
Di Marzo V., Goparaju S. K., Wang L. et al. (2001) Leptin-regulatedendocannabinoids are involved in maintaining food intake. Nature410, 822–825.
Elphick M. R. (2002) Evolution of cannabinoid receptors in vertebrates:identification of a CB(2) gene in the puffer fish Fugu rubripes.Biol. Bull. 202, 104–107.
Elphick M. R., Satou Y. and Satoh N. (2003) The invertebrate ancestryof endocannabinoid signalling: an orthologue of vertebrate cann-abinoid receptors in the urochordate Ciona intestinalis. Gene 302,95–101.
Galiegue S., Mary S., Marchand J., Dussossoy D., Carriere D., CarayonP., Bouaboula M., Shire D., Le Fur G. and Casellas P. (1995)Expression of central and peripheral cannabinoid receptors inhuman immune tissues and leukocyte subpopulations. Eur. J.Biochem. 232, 54–61.
Gaoni Y. and Mechoulam R. (1964) Isolation, structure and partialsynthesis of an active constituent of hashish. J. Am. Chem. Soc. 86,1646.
Hanus L., Avraham Y., Ben-Shushan D., Zolotarev O., Berry E. M. andMechoulam R. (2003) Short-term fasting and prolonged semistar-vation have opposite effects on 2-AG levels in mouse brain. BrainRes. 983, 144–151.
Hao S., Avraham Y., Mechoulam R. and Berry E. M. (2000) Low doseanandamide affects food intake, cognitive function, neurotrans-mitter and corticosterone levels in diet-restricted mice. Eur. J.Pharmacol. 392, 147–156.
Jamshidi N. and Taylor D. A. (2001) Anandamide administration into theventromedial hypothalamus stimulates appetite in rats. Br. J.Pharmacol. 134, 1151–1154.
Kirkham T. C., Williams C. M., Fezza F. and Di Marzo V. (2002)Endocannabinoid levels in rat limbic forebrain and hypothalamusin relation to fasting, feeding and satiation: stimulation of eating by2-arachidonoyl glycerol. Br. J. Pharmacol. 136, 550–557.
Lin X., Volkoff H., Narnaware Y., Bernier N. J., Peyon P. and Peter R. E.(2000) Brain regulation of feeding behavior and food intake infish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 126, 415–434.
Lopez-Patino M. A., Guijarro A. I., Delgado M. J., Alonso-Bedate M.and De Pedro N. (1999) Neuropeptide Y has a stimulatory actionon feeding behaviour in goldfish (Carassius auratus). Eur. J.Pharmacol. 377, 147–153.
Matias I., Leonhardt M., Lesage J., De Petrocellis L., Dupouy J. P.,Vieau D. and Di Marzo V. (2003) Effect of maternal under-nutri-tion on pup body weight and hypothalamic endocannabinoid lev-els. Cell Mol. Life Sci. 60, 382–389.
Matias I., McPartland J. M. and Di Marzo V. (2005) Occurrence andpossible biological role of the endocannabinoid system in the seasquirt Ciona intestinalis. J. Neurochem. 93, 1141–1156.
Matsuda L. A., Lolait S. J., Brownstein M. J., Young A. C. and BonnerT. I. (1990) Structure of a cannabinoid receptor and functionalexpression of the cloned cDNA. Nature 346, 561–564.
McPartland J. M. and Glass M. (2003) Functional mapping of cann-abinoid receptor homologs in mammals, other vertebrates, andinvertebrates. Gene 312, 297–303.
Mechoulam R., Ben-Shabat S., Hanus L. et al. (1995) Identification ofan endogenous 2-monoglyceride, present in canine gut, that bindsto cannabinoid receptors. Biochem. Pharmacol. 50, 83–90.
Meek J. and Nieuwenhuys R. (1998) Holosteans and Teleosteans, in TheCentral Nervous System of Vertebrates (Nieuwenhuys R., TenDonkelaar H. J. and Nicholson C., eds), Vol. 2, pp. 759–938.Springer, Berlin, Germany.
Narnaware Y. K. and Peter R. E. (2002) Influence of diet composition onfood intake and neuropeptide Y (NPY) gene expression in goldfishbrain. Regul. Pept. 103, 75–83.
Endocannabinoid system in brain of Carassius auratus 671
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672

Narnaware Y. K., Peyon P. P., Lin X. and Peter R. E. (2000) Regulationof food intake by neuropeptide Y in goldfish. Am. J. Physiol.Regul. Integr. Comp. Physiol. 279, R1025–R1034.
Patel S., Roelke C. T., Rademacher D. J., Cullinan W. E. and HillardC. J. (2004) Endocannabinoid signaling negatively modulatesstress-induced activation of the hypothalamic-pituitary-adrenalaxis. Endocrinology 145, 5431–5438.
Pontet A., Danger J. M., Dubourg P., Pelletier G., Vaudry H., Calas A.and Kah O. (1989) Distribution and characterization of neuro-peptide Y-like immunoreactivity in the brain and pituitary of thegoldfish. Cell Tissue Res. 255, 529–538.
Rademacher D. J., Weber D. N. and Hillard C. J. (2005) Waterborne leadexposure affects brain endocannabinoid content in male but notfemale fathead minnows (Pimephales promelas). Neurotoxicology26, 9–15.
Roberts M. G. and Savage G. E. (1978) Effects of hypothalamic lesionson the food intake of the goldfish (Carassius auratus). BrainBehav. Evol. 15, 150–164.
Soderstrom K., Tian Q., Valenti M. and Di Marzo V. (2004) Endo-cannabinoids link feeding state and auditory perception-relatedgene expression. J. Neurosci. 24, 10 013–10 021.
Sugiura T., Kondo S., Sukagawa A., Nakane S., Shinoda A., Itoh K.,Yamashita A. and Waku K. (1995) 2-Arachidonoylglycerol: apossible endogenous cannabinoid receptor ligand in brain. Bio-chem. Biophys. Res. Commun. 215, 89–97.
Tsou K., Brown S., Sanudo-Pena M. C., Mackie K. and Walker J. M.(1998) Immunohistochemical distribution of cannabinoid CB1receptors in the rat central nervous system. Neuroscience 83, 393–411.
Volkoff H. and Peter R. E. (2001) Interactions between orexin A, NPYand galanin in the control of food intake of the goldfish, Carassiusauratus. Regul. Pept. 101, 59–72.
Yamaguchi F., Macrae A. D. and Brenner S. (1996) Molecular cloning oftwo cannabinoid type 1-like receptor genes from the puffer fishFugu rubripes. Genomics 35, 603–605.
672 M. Valenti et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 95, 662–672
Related Documents











