ADVANCES IN COGNITIVE PSYCHOLOGY RESEARCH ARTICLE http://www.ac-psych.org 2011 (special issue) • volume 7 • 132-141 132 e effects of spatial and temporal cueing on metacontrast masking Maximilian Bruchmann, Philipp Hintze, and Simon Mota Institute for Biomagnetism and Biosignalanalysis, University of Muenster, Germany visual masking, metacontrast, spatial cueing, temporal cueing We studied the effects of selective attention on metacontrast masking with 3 different cueing ex- periments. Experiments 1 and 2 compared central symbolic and peripheral spatial cues. For sym- bolic cues, we observed small attentional costs, that is, reduced visibility when the target appeared at an unexpected location, and attentional costs as well as benefits for peripheral cues. All these effects occurred exclusively at the late, ascending branch of the U-shaped metacontrast masking function, although the possibility exists that cueing effects at the early branch were obscured by a ceiling effect due to almost perfect visibility at short stimulus onset asynchronies (SOAs). In Ex- periment 3, we presented temporal cues that indicated when the target was likely to appear, not where. Here, we also observed cueing effects in the form of higher visibility when the target ap- peared at the expected point in time compared to when it appeared too early. However, these effects were not restricted to the late branch of the masking function, but enhanced visibility over the complete range of the masking function. Given these results we discuss a common effect for different types of spatial selective attention on metacontrast masking involving neural subsystems that are different from those involved in temporal attention. Corresponding author: Maximilian Bruchmann, Institute for Biomagnetism and Biosignalanalysis, University of Münster, Malme- dyweg 15, 48149 Münster, Germany. Phone: +49 251 83-52547. Fax: +49 251 83-56874. E-mail: [email protected] ABSTRACT KEYWORDS DOI • 10.2478/v10053-008-0093-1 INTRODUCTION Attending to a stimulus and becoming aware of it go hand in hand in everyday life. Yet, awareness and attention are not identical (Lamme, 2003). For example, it is known from patients suffering from lesions of their primary visual cortex that attention can have an effect on detec- ting stimuli in the patients’ blind visual field of which these patients remain unaware (blindsight; Kentridge, Heywood, & Weiskrantz, 1999; Kentridge, Nijboer, & Heywood, 2008). In healthy subjects, awareness can be manipulated by employing metacontrast masking, which is one classical type of visual backward masking. Awareness of a briefly flashed target stimulus can be de- creased or even completely prevented by the following presentation of a surrounding masking stimulus (for an overview, see e.g., Breitmeyer & Ögmen, 2000, 2006; Enns & Di Lollo, 2000). When visibility of the target stimulus is plotted against the stimulus onset asynchrony (SOA) of target and mask, one typically obtains a U-shaped masking function. It has been argued that the reason for a U-shape is the superposition of at least two processes (Michaels & Turvey, 1979; Reeves, 1982; Turvey, 1973): At the descending branch (SOA = 0 ms up to about 60 ms) sub- jects perceive target and mask as one stimulus whose visibility in the area of the target decreases while the visibility of the mask does not change substantially. Michaels and Turvey (1979, p. 1) called this inte- gration by common synthesis, that is, due to their temporal proximity, target and mask “yield one iconic representation” comprising features of both stimuli. At the ascending branch of the masking function, sub- jects are progressively better able to detect a temporal separation of the two stimulus events. e visibility of the target increases monotonically with the likelihood with which targets and masks are perceived as sepa- rate events (Michaels & Turvey, 1979; Reeves, 1982). e dissociation of the descending and ascending part is mirrored in the effects of selective attention on metacontrast. Boyer and Ro (2007) used a version of Posner’s classical symbolic cueing paradigm (Posner, 1980) in which arrows are presented before the target-mask

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141132
The effects of spatial and temporal cueing on metacontrast maskingMaximilian Bruchmann, Philipp Hintze, and Simon Mota
institute for Biomagnetism and Biosignalanalysis, University of Muenster, germany
visual masking,
metacontrast, spatial
cueing, temporal cueing
We studied the effects of selective attention on metacontrast masking with 3 different cueing ex-periments. experiments 1 and 2 compared central symbolic and peripheral spatial cues. For sym-bolic cues, we observed small attentional costs, that is, reduced visibility when the target appeared at an unexpected location, and attentional costs as well as benefits for peripheral cues. All these effects occurred exclusively at the late, ascending branch of the U-shaped metacontrast masking function, although the possibility exists that cueing effects at the early branch were obscured by a ceiling effect due to almost perfect visibility at short stimulus onset asynchronies (soAs). in ex-periment 3, we presented temporal cues that indicated when the target was likely to appear, not where. here, we also observed cueing effects in the form of higher visibility when the target ap-peared at the expected point in time compared to when it appeared too early. however, these effects were not restricted to the late branch of the masking function, but enhanced visibility over the complete range of the masking function. given these results we discuss a common effect for different types of spatial selective attention on metacontrast masking involving neural subsystems that are different from those involved in temporal attention.
corresponding author: Maximilian Bruchmann, institute for
Biomagnetism and Biosignalanalysis, University of Münster, Malme-
dyweg 15, 48149 Münster, germany. Phone: +49 251 83-52547.
Fax: +49 251 83-56874. e-mail: [email protected]
AbstrAct
Keywords
doi • 10.2478/v10053-008-0093-1
IntroductIon
Attending to a stimulus and becoming aware of it go hand in hand in
everyday life. Yet, awareness and attention are not identical (Lamme,
2003). For example, it is known from patients suffering from lesions of
their primary visual cortex that attention can have an effect on detec-
ting stimuli in the patients’ blind visual field of which these patients
remain unaware (blindsight; Kentridge, Heywood, & Weiskrantz, 1999;
Kentridge, Nijboer, & Heywood, 2008).
In healthy subjects, awareness can be manipulated by employing
metacontrast masking, which is one classical type of visual backward
masking. Awareness of a briefly flashed target stimulus can be de-
creased or even completely prevented by the following presentation of
a surrounding masking stimulus (for an overview, see e.g., Breitmeyer
& Ögmen, 2000, 2006; Enns & Di Lollo, 2000). When visibility of the
target stimulus is plotted against the stimulus onset asynchrony (SOA)
of target and mask, one typically obtains a U-shaped masking function.
It has been argued that the reason for a U-shape is the superposition of
at least two processes (Michaels & Turvey, 1979; Reeves, 1982; Turvey,
1973): At the descending branch (SOA = 0 ms up to about 60 ms) sub-
jects perceive target and mask as one stimulus whose visibility in the
area of the target decreases while the visibility of the mask does not
change substantially. Michaels and Turvey (1979, p. 1) called this inte-
gration by common synthesis, that is, due to their temporal proximity,
target and mask “yield one iconic representation” comprising features
of both stimuli. At the ascending branch of the masking function, sub-
jects are progressively better able to detect a temporal separation of the
two stimulus events. The visibility of the target increases monotonically
with the likelihood with which targets and masks are perceived as sepa-
rate events (Michaels & Turvey, 1979; Reeves, 1982).
The dissociation of the descending and ascending part is mirrored
in the effects of selective attention on metacontrast. Boyer and Ro
(2007) used a version of Posner’s classical symbolic cueing paradigm
(Posner, 1980) in which arrows are presented before the target-mask

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141133
sequence. When arrows pointed to the correct position of the target,
detection performance was increased (compared to when they pointed
to the wrong position). This effect appeared exclusively at the ascend-
ing branch of the masking function. Similarly, Neumann and Scharlau
(2007) found that presenting a distracting task-irrelevant stimulus con-
tralateral to the target decreased target detectability at the ascending
branch. Tata (2002) used peripheral cues in a metacontrast paradigm,
that is, target position was indicated by a stimulus appearing at the
exact location of the consecutive target and found increased detection
rates if the target location was validly cued. Tata kept the SOA constant
at 80 ms, which is typically in the range of the ascending branch of the
masking function. In general, the finding that effects of attention do
not simply counteract the awareness-reducing effects of metacontrast
is further evidence that attention and awareness are qualitatively dif-
ferent concepts.
The studies discussed above used either central symbolic or peri-
pheral flanking cues to direct attention to the target location (or away
from it). In none of the cited studies symbolic and flanking cues were
compared in a single experiment, or in two otherwise comparable ex-
periments. With the first two of the reported experiments we addressed
this question. Symbolic and flanking cues were compared within the
same paradigm. Note that we refrain from calling these cue types endog-
enous and exogenous cues, respectively, because exogenous cueing re-
quires the cues to be uninformative (i.e., a ratio of 1:1 of valid and invalid
cues; Carrasco, 2011) and in the present experiments we used informa-
tive cues, both in the symbolic and flanker cueing tasks. Compared to
previous studies we further improved the design in two ways: We mo-
nitored eye movements by means of an eye-tracker and we included a
neutral cueing condition to be able to dissociate attentional benefits and
costs. Differentiating between costs and benefits may allow conclusions
about the way attentional resources are assigned to the visual input.
It further remains to be clarified whether enhanced visibility at
the late branch of the masking function is only found when subjects
attend to the correct location of the target. The third experiment ex-
tends the study of effects of attention on metacontrast by providing the
subject with (valid or invalid) information not about where the target
appears but when it will appear. Temporal cueing has been shown to
enhance behavioral performance (Coull & Nobre, 1998; Nobre, 2001).
Studies measuring event-related EEG potentials suggest that temporal
cues facilitate performance by enhancing early visual processing steps
(Correa, Lupiáñez, Madrid, & Tudela, 2006), especially if the task is
perceptually demanding. With this comparison of the effects of three
different types of selective attention on metacontrast masking we seek
to clarify how specifically selective attention interacts with the modula-
tion of conscious stimulus perception by visual masking.
ExpErImEnts 1 and 2: EffEcts of cEntral symbolIc and pErIphEral flankIng cuEs on mEtacontrast maskIng
All reported experiments were adapted versions of the experimental
design used by Bruchmann, Breitmeyer, and Pantev (2010). Targets and
masks consisted of sinusoidal gratings with a Gaussian envelope. The
participants’ task was to rate the visibility of the target subjectively on a
5-point scale. As reported recently by Albrecht, Klapötke, and Mattler
(2010), subjects may show Type-A (i.e., monotonous) or Type-B
(U-shaped) masking functions depending on their individual strategy.
In our previous studies (Bruchmann et al., 2010), we had observed that
with Gaussian blurred stimuli in combination with a subjective ratings
task in which targets were presented on every trial (except for a few
control trials), subjects did not appear to engage in different strategies.
+
+
+
+
+
+
30 ms
200 ms 30 ms
30 ms
flanker cueingsymbolic cueing
CT-SOA = 250 ms
CT-SOA = 80 ms
TM-SOA = 0 - 170 ms
TM-SOA = 0 - 170 ms
Figure 1.
trial sequences used in the symbolic and flanker cueing experiments. symbolic cues were presented for 200 ms, flanker cues for 30 ms. the cues were followed by a blank interval of 50 ms, resulting in a cue-to-target-stimulus onset asynchrony (ct-soA) of 250 ms and 80 ms, respectively. cues could be valid, invalid, or neutral (double headed arrow / flankers on both sides) with a ratio of 3:1:1. targets and masks were presented for 30 ms. on each trial the target-mask- stimulus onset asynchrony (tM-soA) was chosen randomly be-tween 0 and 170 ms.

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141134
Because of the similarity of both tasks we will first describe the
design and methods of both and then report the results. Figure 1
shows exemplary trial sequences for the symbolic and flanker cueing
experiments. For each of the two cue types we chose cue-to-target-
SOAs (CT-SOAs) that are in the typical range reported in the literature
(Carrasco, 2011; Cheal, Lyon, & Hubbard, 1991; Hein, Rolke, & Ulrich,
2006; Ling & Carrasco, 2006; Müller & Rabbitt, 1989). For symbolic
cues we used a CT-SOA = 250 ms and for flanking cues we used a
CT-SOA = 80 ms.
SubjectsSix subjects (five female, one male) participated in both experiments.
One half of the subjects started with the symbolic cueing experiment,
and the other half with the flanker cueing experiment. All had normal
or corrected to normal vision and no history of neurological or psy-
chiatric diseases. Their age was 22 to 24 years (M = 23, SD = 0.7). Four
subjects were right-handed, two left-handed. The subjects gave their
informed consent and volunteered for participation and were paid 9 €
per hour. All procedures were carried out according to the declaration
of Helsinki and were approved by the ethical committee of the medical
faculty of the University of Münster.
Apparatus and stimuliThe experiment was run using SR Research Experiment Builder (SR
Research Ltd., version 1.6.1). Stimuli were presented on a Samsung
SyncMaster 1100P screen at a resolution of 1,024 × 768 pixel and 100
Hz, at a viewing distance of 65 cm. The subjects responded by pressing
one of five buttons on an external response box. The participants were
instructed to focus on a central fixation mark. To monitor the subject’s
eye-movements, a head-based SR Research Ltd. EyeLink II eye-track-
ing device (version 2.22), was used. Once the focus deviated more than
2 degrees of visual angle (°) from the central fixation mark, a warning
message was displayed and the respective trial was reintegrated into
the condition list at a random point for later presentation. As target
stimuli, Gabor patches with a diameter of 2° (measured from -2.5 to
2.5 SD of the Gaussian envelope) were used. As mask, a grating annulus
with a Gaussian envelope was used. The diameter of the Gaussian enve-
lope was 2°. Targets and masks were centered randomly 5° to the left or
right of the fixation mark. Both had a spatial frequency of f = 4 cycles
per degree of visual angle (cpd) and were presented at six orientations:
φ = 0°, 30°, 60°, 90°, 120°, and 150°. The phase of the sinusoidal lumi-
nance modulation was φ = 0° in the target and φ = 180° in the mask,
meaning that each white “stripe” in the target was aligned with a black
“stripe” of the mask and vice versa. The mask was presented at 100%
black-and-white-contrast, the target at 90% black-and-white-contrast.
The background color was middle gray. The symbolic cue stimulus
was the white outline of an arrow, 3° in length. It could point either to
the left or to the right or (in the neutral condition) could be double-
headed, pointing in both directions. The flanker cue stimulus was a
white line 2.5° above and below the possible location of the target stim-
ulus. It could be presented at one or (in the neutral condition) at both
locations.
Symbolic cueing: Procedure
The subjects were instructed to focus on the fixation mark. Trials star-
ted with the symbolic cue stimulus, presented for 200 ms. The interval
between cue offset and target onset was 50 ms. The cue-to-target-SOA
therefore was 250 ms. The cue could be valid (pointing to the correct
side), invalid (pointing to the opposite side), or neutral (pointing to
both sides) at a ratio of 3:1:1. The target was presented either to the left
or to the right of the fixation mark for 30 ms, followed by the mask, also
presented for 30 ms. The SOA between target and mask was either 0,
10, 20, 30, 40, 50, 60, 80, 120, or 170 ms.
Beside the target and mask conditions, we occasionally presented
the target or the mask only. The target only conditions were supposed
to “remind” subjects from time to time what they were supposed to
detect. The mask only conditions were needed to obtain a false alarm
rate (for details, see Results section). The orientation φ of the target and
mask was varied randomly and averaged for the analysis. The result-
ing 72 experimental conditions (3 cueing conditions [valid, invalid,
neutral] × 12 SOAs [10 SOAs + target-/mask-only trials]× 2 screen
sides) were repeated 30 times in case of invalid and neutral trials and
90 times in case of valid trials, adding up to 3,600 trials. The trials were
distributed over five sessions of 1 hr each. In order to get accustomed
to the task, each subject performed 100 practice trials before response
recording started. Subjects were asked to ignore the mask and to rate
the visibility of the target stimulus after each trial, using one of five
buttons, ranging from “0” = not visible to “4” = clearly visible. They were
instructed to maintain a constant rating scheme over the experimental
sessions and to use the full rating scale. The next trial started 200 ms
after the response.
Flanker cueing: ProcedureThe procedure was equivalent to the previous experiment apart from
the cue setup (see Figure 1). The flanker cue stimulus could again be
valid (presented on the correct side), invalid (presented on the opposite
side), or neutral (presented on both sides simultaneously). The cue was
presented for 30 ms and the onset asynchrony between cue and target
stimulus was 80 ms. As in the symbolic cueing conditions the propor-
tions of valid to invalid to neutral trials was 3:1:1.
ResultsIn order to exclude a possible response bias (e.g., deliberately giving
higher ratings in valid trials), we chose not to analyze the raw rating
data. Instead, we chose a signal detection theory approach (Green &
Swets, 1966) where the visibility ratings are treated as detection data
combined with a confidence rating (i.e., the lowest rating was treated
as “target absent, high confidence”, the second lowest rating as “target
absent, low confidence”, up to “target present, high confidence”). To
obtain hit-rates (H), the relative frequency of each rating level in trials
where a target was presented at a given SOA and cueing condition is
first calculated and then summed over rating levels, yielding cumu-
lative conditional probabilities. For k rating levels one obtains k – 1
cumulative hit rates, because the kth level necessarily has a cumulative
probability of p = 1. Similarly, the false alarm rates are calculated for
AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141135
each rating level in trials where only the mask was presented at a given
cueing condition. Since there is no SOA in mask-only trials, the same
false-alarm data is used for all SOAs in a given cueing condition. We
then fitted a receiver operation characteristic (ROC) curve to the cu-
mulative probabilities using the algorithm described by Dorfman and
Berbaum (1986). For the ROCs we assumed a normal distribution of
noise (i.e., internal activation in trials without a target) with μ0= 0 and
σ0 = 1, and a normal distribution of signal + noise (i.e., internal activa-
tion in trials with targets) with μ1 and σ1 as free parameters. The analy-
sis is based on the measure Az, that is, the area under the ROC curve
(see e.g., Wickens, 2001) which ranges from Az = .5 for performance at
chance level to Az = 1 for perfect detection.
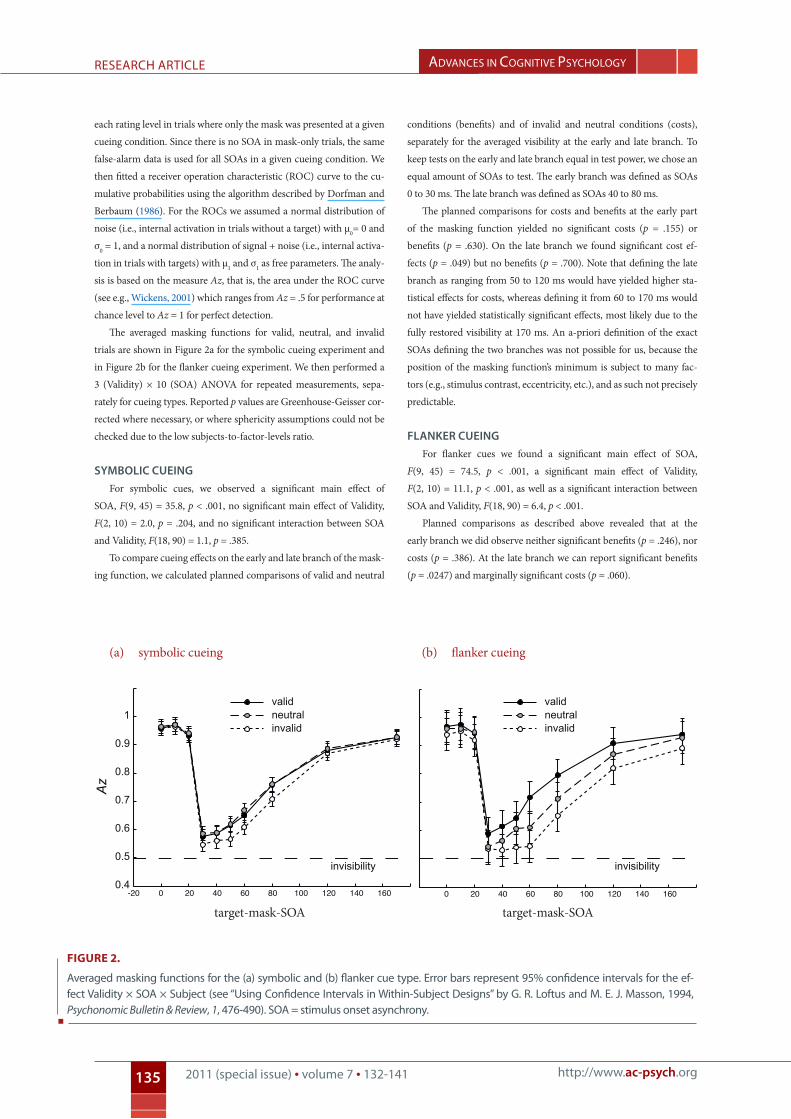
The averaged masking functions for valid, neutral, and invalid
trials are shown in Figure 2a for the symbolic cueing experiment and
in Figure 2b for the flanker cueing experiment. We then performed a
3 (Validity) × 10 (SOA) ANOVA for repeated measurements, sepa-
rately for cueing types. Reported p values are Greenhouse-Geisser cor-
rected where necessary, or where sphericity assumptions could not be
checked due to the low subjects-to-factor-levels ratio.
Symbolic cueingFor symbolic cues, we observed a significant main effect of
SOA, F(9, 45) = 35.8, p < .001, no significant main effect of Validity,
F(2, 10) = 2.0, p = .204, and no significant interaction between SOA
and Validity, F(18, 90) = 1.1, p = .385.
To compare cueing effects on the early and late branch of the mask-
ing function, we calculated planned comparisons of valid and neutral
conditions (benefits) and of invalid and neutral conditions (costs),
separately for the averaged visibility at the early and late branch. To
keep tests on the early and late branch equal in test power, we chose an
equal amount of SOAs to test. The early branch was defined as SOAs
0 to 30 ms. The late branch was defined as SOAs 40 to 80 ms.
The planned comparisons for costs and benefits at the early part
of the masking function yielded no significant costs (p = .155) or
benefits (p = .630). On the late branch we found significant cost ef-
fects (p = .049) but no benefits (p = .700). Note that defining the late
branch as ranging from 50 to 120 ms would have yielded higher sta-
tistical effects for costs, whereas defining it from 60 to 170 ms would
not have yielded statistically significant effects, most likely due to the
fully restored visibility at 170 ms. An a-priori definition of the exact
SOAs defining the two branches was not possible for us, because the
position of the masking function’s minimum is subject to many fac-
tors (e.g., stimulus contrast, eccentricity, etc.), and as such not precisely
predictable.
Flanker cueingFor flanker cues we found a significant main effect of SOA,
F(9, 45) = 74.5, p < .001, a significant main effect of Validity,
F(2, 10) = 11.1, p < .001, as well as a significant interaction between
SOA and Validity, F(18, 90) = 6.4, p < .001.
Planned comparisons as described above revealed that at the
early branch we did observe neither significant benefits (p = .246), nor
costs (p = .386). At the late branch we can report significant benefits
(p = .0247) and marginally significant costs (p = .060).
target-mask-SOA target-mask-SOA
symbolic cueing flanker cueing valid
invalidneutral
valid
invalidneutral
Az
a) b)0 20 40 60 80 100 120 140 160-20 0 20 40 60 80 100 120 140 160
0.4
0.5
0.6
0.7
0.8
0.9
1
invisibilityinvisibility
Figure 2.
Averaged masking functions for the (a) symbolic and (b) flanker cue type. error bars represent 95% confidence intervals for the ef-fect validity × soA × subject (see “Using confidence intervals in Within-subject designs” by g. r. loftus and M. e. J. Masson, 1994, Psychonomic Bulletin & Review, 1, 476-490). soA = stimulus onset asynchrony.
target-mask-SOA target-mask-SOA
(a) (b) symbolic cueing flanker cueing

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141136
Discussion
The results on symbolic cueing effects partly replicate those of Boyer
and Ro (2007), as we also found significant differences after the SOA
of optimal masking. In addition, we see that the cueing effect is com-
pletely due to attentional costs rather than benefits, that is, compared to
the neutral condition, visibility is not enhanced when arrows indicate
the correct location of the target, but visibility is reduced when arrows
point to the wrong location. The effect is small, similar to Boyer and Ro’s
result. Flanker cues exhibit a larger effect on the masking function. We
observed attentional costs as well as benefits. In all cases, effects were
restricted to the late branch of the masking function. To compare the
effects of both experiments we calculated an additional 2 (Experiment)
× 3 (Validity) × 10 (SOA) ANOVA. As expected, the three-way interac-
tion was significant, F(18, 90) = 2.3, p = .005, confirming that flanker
cueing effects were substantially larger than symbolic cueing effects.
It may be argued that effects at the early part of the masking func-
tion could have been obscured by a ceiling effect at SOAs = 0 to 20 ms
and that intermediate levels of visibility would have obtained between
SOAs of 20 and 30 ms. Due to the monitor refresh rate we were bound
to a spacing of SOAs by at least 10 ms. Thus, we were not able to cover
the early branch in more detail. To check for a possible ceiling effect we
analyzed the inter-individual variation in visibility at the early part (i.e.,
the individual Az averaged over Validity and SOAs of 0 to 20 ms) and
correlated this measure with the attentional costs and benefits (also ave-
raged over SOAs of 0 to 20 ms). Given a substantial variation in average
visibility, a ceiling effect would imply a negative correlation of visibility
and the negative or positive effects of cueing. The observed range of
averaged visibilities was Az = 0.935 to Az = 0.980. For symbolic cue-
ing, the Pearson-correlation coefficient for cueing effects and averaged
visibility was positive but statistically nonsignificant for costs (r = .554,
p = .254), and negative but also nonsignificant for benefits (r = −.391,
p = .444). For flanker cueing, the correlation was negative but statisti-
cally nonsignificant for costs (r = −.281, p = .590) as well as for benefits
(r = −.355, p = .490). Since the inter-individual variation of averaged Az
at the early part of the masking function as well as the sample size were
small, we have to acknowledge that cueing effects at the early part of
the masking function cannot be excluded based on our present data.
ExpErImEnts 3: EffEcts of tEmporal cuEIng on mEtacontrast maskIng
In this experiment, the offset of the fixation mark was used as a tem-
poral cue (see Figure 3). Over the course of the experiment, subjects
were supposed to learn that in most cases the offset of the fixation mark
preceded the target onset by a fixed temporal interval (t1). In the re-
maining trials the subjects’ expectation was violated and the target was
presented after a different temporal interval (t2). This procedure was
first described by Coull and Nobre (1998). It is well known that sub-
jects intuitively establish an accurate representation of the frequency of
events even if not instructed to do so (for a review, see Hasher & Zacks,
1984). The experiment consisted of two sessions between which the
values for t1 and t2 were exchanged.
SubjectsNine subjects (five female, four male) participated in the experiment.
All had normal or corrected to normal vision and no history of neu-
rological or psychiatric diseases. Their age was between 22 and 29
years (M = 25, SD = 2.52). Seven subjects were right-handed, two left-
handed. The subjects gave their informed consent and volunteered for
participation and were paid 9 € per hour. All procedures were carried
out according to the declaration of Helsinki and were approved by the
ethical committee of the medical faculty of the University of Münster.
Apparatus and stimuliThe experiment was run using MATLAB and the PsychophysicsToolbox
(Brainard, 1997). Stimuli were presented on a ViewSonic G90fB CRT
monitor at 100 Hz and a resolution of 1,024 × 768 pixels at a view-
ing distance of 80 cm. The mean brightness of the monitor was set
to approximately 50 cd/m2 (Imin = 0.413 cd/m2, Imax = 100.201 cd/m2).
Participants gave their responses by pressing one of four buttons on
an external response box. The stimuli were generated as described by
Bruchmann et al. (2010). All stimuli were always presented at the ma-
ximum Michelson contrast of (Imax - Imin) / (Imax + Imin) = 0.992. Stimulus
dimensions were identical to those in the two previous experiments.
Targets and masks had a spatial frequency of f = 2 cpd and were pre-
sented at random orientation, with the target and mask always sharing
the same orientation.
+
+
30 ms
100 or 1000 ms
1000 ms
30 msTM-SOA = 0 - 140 ms
Figure 3.
trial sequences used in the temporal cueing experiment. cueing the time point of target occurrence was achieved by introducing an interval between fixation mark offset and tar-get onset with two fixed durations, t1 and t2, where t1 was eight times more frequent than t2. After a short learning period sub-jects began to expect target occurrence after t1. the interval lengths used for t1 and t2 were 100 ms and 1 s. subjects com-pleted two sessions of the experiment, with 100 ms as t1 in one session and as t2 in the other. tM-soA = target-mask- stimulus onset asynchrony.
AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141137
Procedure
The general procedure (i.e., stimulus durations and dimensions, SOA
randomization, control trials with target or mask only) was identical to
the previous experiments. The fixation mark was shown for 1 s before
it disappeared. The target-mask sequence appeared at random to the
left or the right of the fixation mark after one of two possible CT-SOAs.
The CT-SOAs were 100 ms and 1 s. In each of two experimental ses-
sions per subject one was used eight times more often than the other
(validity 8:1). The order of sessions was balanced across subjects. SOA
varied randomly between 0, 30, 50, 60, 80, 110, and 140 ms. In each
session, the invalid condition comprised 180 trials (2 sides × [7 SOAs
+ 1 target only reference trial + 1 mask only reference trial] × 10 re-
petitions). The valid condition was eight times more frequent than the
invalid, yielding 1,440 trials. In each of two experimental sessions of
90 min, participants completed 1,620 trials. After each trial, the par-
ticipants were asked to rate the visibility of the target with four buttons.
They were instructed to press button “1” if the target was not visible
at all, and button “4” if it was well visible, and to use buttons “2” and
“3” for intermediate visibility. The participants were asked to try using
the full rating scale and to establish a constant rating scheme. Before
starting the main experiment, the participants had 5 min of training to
get familiar with the task.
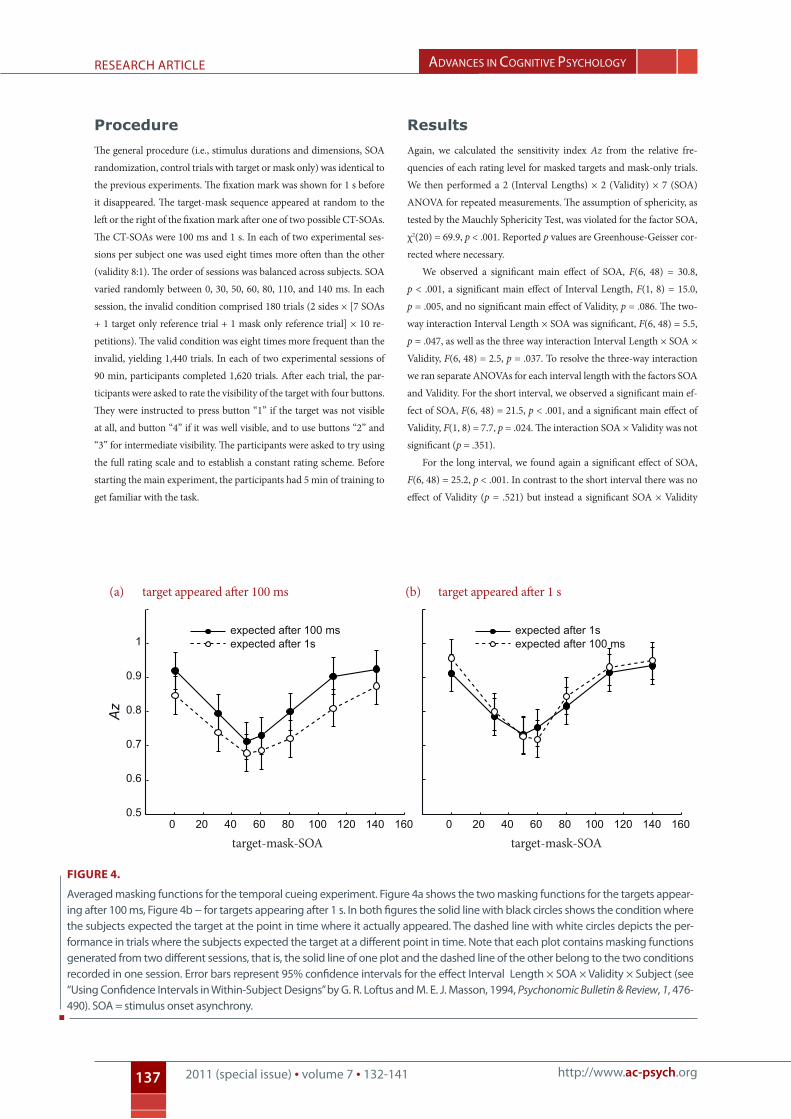
Results
Again, we calculated the sensitivity index Az from the relative fre-
quencies of each rating level for masked targets and mask-only trials.
We then performed a 2 (Interval Lengths) × 2 (Validity) × 7 (SOA)
ANOVA for repeated measurements. The assumption of sphericity, as
tested by the Mauchly Sphericity Test, was violated for the factor SOA,
χ2(20) = 69.9, p < .001. Reported p values are Greenhouse-Geisser cor-
rected where necessary.
We observed a significant main effect of SOA, F(6, 48) = 30.8,
p < .001, a significant main effect of Interval Length, F(1, 8) = 15.0,
p = .005, and no significant main effect of Validity, p = .086. The two-
way interaction Interval Length × SOA was significant, F(6, 48) = 5.5,
p = .047, as well as the three way interaction Interval Length × SOA ×
Validity, F(6, 48) = 2.5, p = .037. To resolve the three-way interaction
we ran separate ANOVAs for each interval length with the factors SOA
and Validity. For the short interval, we observed a significant main ef-
fect of SOA, F(6, 48) = 21.5, p < .001, and a significant main effect of
Validity, F(1, 8) = 7.7, p = .024. The interaction SOA × Validity was not
significant (p = .351).
For the long interval, we found again a significant effect of SOA,
F(6, 48) = 25.2, p < .001. In contrast to the short interval there was no
effect of Validity (p = .521) but instead a significant SOA × Validity
target appeared after 1starget appeared after 100 msexpected after 100 msexpected after 1s
target-mask-SOAtarget-mask-SOA
expected after 1sexpected after 100 ms
0 20 40 60 80 100 120 140 160 0 20 40 60 80 100 120 140 160a) b)
0.5
0.6
0.7
0.8
0.9
1
Az
Figure 4.
Averaged masking functions for the temporal cueing experiment. Figure 4a shows the two masking functions for the targets appear-ing after 100 ms, Figure 4b − for targets appearing after 1 s. in both figures the solid line with black circles shows the condition where the subjects expected the target at the point in time where it actually appeared. the dashed line with white circles depicts the per-formance in trials where the subjects expected the target at a different point in time. note that each plot contains masking functions generated from two different sessions, that is, the solid line of one plot and the dashed line of the other belong to the two conditions recorded in one session. error bars represent 95% confidence intervals for the effect interval length × soA × validity × subject (see “Using confidence intervals in Within-subject designs” by g. r. loftus and M. e. J. Masson, 1994, Psychonomic Bulletin & Review, 1, 476-490). soA = stimulus onset asynchrony.
target-mask-SOA target-mask-SOA
(a) target appeared after 100 ms (b) target appeared after 1 s

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141138
interaction, F(6, 48) = 2.9, p = .047. Post-hoc comparisons at each
SOA for “valid vs. invalid” with Tukey-tests corrected for multiple
application were all not significant (all ps ≥ .155, with the smallest
p value observed at SOA = 0 ms), and could thus not provide certainty
about the reason for the interaction. Descriptively, we observed slightly
higher visibility in invalid trials at short SOAs (0 and 30 ms) and long
SOAs (80, 110, and 140 ms). At intermediate SOAs, this difference was
either not observable (SOA = 50 ms), or reversed (SOA = 60 ms, see
Figure 4b).
To compare cueing effects on the early and late branch of the mask-
ing function, we calculated planned comparisons of valid and invalid
conditions, separately for the averaged visibility of SOAs between
0 and 50 ms (early branch) and SOAs between 60 and 120 ms (late
branch). Again, defining the ascending branch as 80 to 140 ms would
have been less conservative. Since the interaction of SOA and Validity
is only present for the long interval, the statistics actually do not justify
a separate look at the two branches in the short interval. Nevertheless,
we provide the results for the sake of completeness.
For targets presented after 100 ms, there was no significant cueing
effect at the early branch (p = .085). At the late branch target visibility
was significantly higher if the temporal cue was valid (p = .016). For
targets presented after 1 s, there were no significant effects of cueing at
the early (p = .388) or late (p = .692) branch.
DiscussionIn contrast to the effects of spatial cueing (peripheral or symbolic),
temporal cueing can affect the complete masking function. We ob-
served higher visibility ratings for targets appearing after 100 ms when
the target was expected to appear at this time point compared to when
it was expected to appear after 1 s. This effect was not found for targets
appearing after 1 s. A similar finding was reported by Coull and Nobre
(1998): In their temporal cueing study they observed validity effects
in all conditions, except for those in which temporal cues incorrectly
predicted the target’s appearance at the long time interval. As we did,
the authors found no deleterious effect when the subject expected the
target to occur at the short time interval but it actually occurred at the
longer one. The lack of a cueing effect was explained by a “reorientation
of attention” toward the long CT-SOA (Coull & Nobre, 1998). Since
subjects learned that only two intervals were used, omission of the
target at the short interval guaranteed it would occur at the long inter-
val. In line with this interpretation is the observation that the masking
functions for targets appearing after 1 s have the same shape as the
function for targets presented and expected after 100 ms and not as the
function presented but not expected after 100 ms. We conclude that
under all conditions, except when targets appeared unexpectedly early,
attention was present at the moment the target appeared.
Our results further indicate that, depending on SOA, reorienting
attention from the short to the long interval may even increase visibility
as compared to a direct shift towards the long interval. This is indicated
by the SOA × Validity interaction for stimuli presented after 1 s in com-
bination with the descriptively higher visibility ratings at short and long
SOAs in invalid trails compared to valid trials. However, the present
data are insufficient to clarify whether benefits of a temporal reorien-
tation of attention exist and in how far they are modulated by SOA.
gEnEral dIscussIon
Three different types of attentional cues were used to study the ef-
fects of selective attention on metacontrast masking. Both spatial cue
types revealed that expecting targets at the wrong location reduced
target visibility exclusively at the late branch of the masking function.
Additionally, flanker cues, but not symbolic cues, provided attentional
benefits when the correct location was attended, again only at the late
branch. Temporal cues provided a different picture: Expecting targets
later than they actually appear yielded decreased visibility ratings, ir-
respective of SOA. Expecting targets earlier than they actually appear,
did not lower or lift the masking function as a whole. There appeared
to be subtle variations of a cueing effect with SOA, indicating that at
short and long SOAs there was also a benefit from reorienting temporal
attention after the expectancy of an early target had been violated.
The symbolic cues in this study match the classic definition of en-
dogenous cues, which means that they are assumed to trigger a slow,
voluntary shift of attention to the cued location. The flanker cues do
not match the classic definition of exogenous cues since they were
informative. Thus, we cannot exclude that the flankers triggered fast
involuntary as well as slow voluntary attentional shifts. However, en-
dogenous attention takes on average about 300 ms to develop its full
effect (Carrasco, 2011). Since the CT-SOA used for flankers was 80 ms,
we conclude that the major attentional resources contributing to the
observed effects stem from a fast involuntary attentional system.
Interestingly, we observed qualitatively comparable effects of sym-
bolic and flanker cues on metacontrast masking although they can be
assumed to trigger fundamentally different mechanisms of attention
allocation. The difference is merely that flanker cueing effects are larger
and reflect attentional costs as well as benefits, whereas symbolic cue-
ing effects are smaller and appear to reflect only attentional costs.
The conditions under which selective attention is proposed to have
an effect are discussed below. A model dealing with the question how
selective attention may affect visual masking was proposed by Smith
and colleagues (Smith, 2000; Smith, Lee, Wolfgang, & Ratcliff, 2009;
Smith & Wolfgang, 2004). The authors propose that the mask limits
the time target information is represented at a sensorial processing
level. The allocation of attention to the target area causes an increase
of the speed with which sensory information is read out to short-term
memory. With the present results we can add to this model that sym-
bolic and flanker cues appear to have comparable effects, except that
only with flankers we found that a valid cue is better than a neutral cue.
In Smith et al.’s model, attention is described as a spatiotemporal filter,
which corresponds to the classic spotlight metaphor of attention, with
the exception that it is defined by three dimensions: a spatial dimen-
sion, an intensity dimension, and a temporal dimension. To explain
the differences between symbolic and flanker cueing effects we draw
on the finding that size and shape of the attentional spotlight can be in-
fluenced by the type of the cue (Castiello & Umiltà, 1990; Eriksen & St

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141139
James, 1986; Eriksen & Yeh, 1985; Galera & Grünau, 2003). We assume
that the attentional spotlight triggered by symbolic cues is comparably
broad in the spatial dimensions, so that with neutral cues both loca-
tions share some attentional gain. Invalid cues shift the broad focus to
the wrong side, leaving the target side unattended. Valid cues, however,
do not provide significantly more gain than the neutral cue, due to the
broad spatial distribution of attentional resources. In contrast, flanker
cues trigger a spatially sharply focused attentional spotlight. A neutral
cue may provide the same mild attentional gain as a neutral symbolic
cue, but the other cues appear to allocate sharply focused attentional
resources at the cued side, withdrawing resources from the uncued
side. Castiello and Umiltà (1990) presented evidence for a trade-off
between size and efficiency of the attentional spotlight. Hence, we
observe not only larger effects with flanker cues, but attentional costs
as well as benefits. All spatial cues provide sharp temporal information
since the CT-SOA was fixed.
An explanation for the observation that only the late branch of the
masking function is affected by spatial selective attention cannot be
easily deduced from current theories or models of metacontrast mask-
ing. In the classic sustained-transient model (ST-model) by Breitmeyer
and Ganz (1976), metacontrast is a consequence of the interaction
between the delayed sustained signal of the target and the quick tran-
sient signal of the mask. Due to the timing difference of sustained and
transient channels it takes a positive non-zero SOA for a maximum
overlap, and hence maximum inhibitory effects, of target and mask sig-
nals. However, if we look at two points in the masking function that are
equal in visibility, but one left of the masking maximum and one right
of it (i.e., one point on the early branch, the other on the late branch),
the ST-model proposes that the reason for incomplete visibility reduc-
tion is the same in both cases: Partial temporal overlap of sustained
target signals with transient mask signals. Yet, we see that only to the
right of the masking maximum visibility is affected by spatial selective
attention. Other models, such as the lateral inhibition theory (Macknik
& Livingstone, 1998; Macknik & Martinez-Conde, 2007) also do not
propose different mechanisms to explain partial masking at the two
branches. A distinction between the two branches is made by Michaels
and Turvey (1979), by Turvey (1973), and by Reeves (1982). Although
the authors do not fully agree concerning the exact number and types
of processes responsible for the U-shaped masking function, they do
agree that the late branch is characterized by temporal separability of
target and mask appearance. Specifically, it has been shown that for
SOAs beyond the SOA of maximum masking, the likelihood of per-
ceiving two events is higher than perceiving a single event (Michaels
& Turvey, 1979). Temporal separability may thus be the precondition
for any effect of spatial selective attention to take place. Because the
experimental task is to rate target visibility and to ignore the mask, we
assume that the attentional focus adheres to the target only, not to the
mask, and not to an integrated target-mask-object.
The ST-model’s successor, the RECOD-model (Breitmeyer &
Öğmen, 2006), incorporates non-linear feedback loops to explain me-
tacontrast masking. In this model, the late branch is characterized by
an increasing number of uninterrupted re-entrant activity from higher
to lower visual processing stages. This top-down directed information
is likely to be the carrier of attentional information.
All these explanations may be also valid for the effect of temporal
cueing on the late branch of the masking function. However, the effect
of temporal cueing (with targets appearing after 100 ms) was not limi-
ted to the late branch. In our view, this indicates a qualitative difference
between spatial and temporal attention. In general, this assumption is
in line with Nobre’s (2001) review comparing spatial and temporal at-
tention, where the author concludes that the mechanisms behind the
two types are “not simply the same and redundant” (p. 1319). Our in-
terpretation of the present result is that spatial attention interacts with,
or modulates the target-mask interactions that are causing the meta-
contrast phenomenon. Temporal attention, on the other hand, appears
to have an additive effect on target visibility and may involve neural
mechanisms or subsystems that are independent of those engaged in
metacontrast. This hypothesis is supported by neuroimaging results on
metacontrast on the one hand and spatial or temporal attention on the
other: An fMRI study by Haynes, Driver, and Rees (2005) suggests that
visibility reductions by metacontrast coincide with reduced effective
connectivity between primary visual cortex and the fusiform gyrus
(FG). FG has been repeatedly shown to be involved in spatial attention
(Heinze et al., 1994; Hopfinger, Buonocore, & Mangun, 2000). Neural
correlates of temporal attention, on the other hand, as observed by
fMRI (Coull, Frith, Büchel, & Nobre, 2000; Coull & Nobre, 1998) or
PET (Coull & Nobre, 1998), do not involve FG. Of course, we have to
assume that numerous brain areas are engaged in metacontrast and
spatial attention, and that even more brain areas are not involved in
temporal attention. Consequently, showing that neural correlates of the
former two share one brain area that the third one does not share can-
not be treated as proof for FG being the neural locus at which spatial
attention modulates the effectiveness of metacontrast. However, FG
qualifies as a candidate for such a locus.
We conclude that spatial and temporal attention exhibit qualita-
tively different effects on metacontrast masking. Spatial cues leave the
early branch of the metacontrast masking function unchanged, whereas
temporal cues do not. Given the subtle and not yet clarified interaction
of temporal cueing and metacontrast with targets appearing after 1 s,
future experiments have to clarify the role of the exact choice of tem-
poral intervals. Nobre (2001) discusses how not only the absolute dura-
tion of cue-target intervals but also the difference between the chosen
intervals influences the size of the observed attentional effect on choice
reaction time. In combination with metacontrast these interactions
may be even more complicated, as indicated by our present results.
We believe it to be a promising approach to study these interactions in
more detail in order to learn more about the temporal relationships of
stimulus processing and temporal attention.
acknowledgementSThis work was funded by the German Research Foundation (DFG),
BR3832/1-1. We thank Dirk Vorberg for helpful discussions during
planning and analyzing of the experiments and two anonymous review-
ers for valuable suggestions regarding data analysis and interpretation.

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141140
reFerenceSAlbrecht, t., Klapötke, s., & Mattler, U. (2010). individual dif-
ferences in metacontrast masking are enhanced by per-
ceptual learning. Consciousness and Cognition, 19(2), 656-
666.
Boyer, J., & ro, t. (2007). Attention attenuates metacontrast mask-
ing. Cognition, 104(1), 135- 149.
Brainard, d. h. (1997). the Psychophysics toolbox. Spatial Vision,
10(4), 433-436.
Breitmeyer, B. g., & ganz, l. (1976). implications of sustained and
transient channels for theories of visual pattern masking, sac-
cadic suppression, and information processing. Psychological
Review, 83(1), 1-36.
Breitmeyer, B. g., & Ögmen, h. (2000). recent models and find-
ings in visual backward masking: A comparison, review, and
update. Perception & Psychophysics, 62(8), 1572-1595.
Breitmeyer, B. g., & Öğmen, h. (2006). Visual masking: Time slices
through conscious and unconscious vision. new york: oxford
University Press.
Bruchmann, M., Breitmeyer, B. g., & Pantev, c. (2010). Metacontrast
masking within and between visual channels: effects of orien-
tation and spatial frequency contrasts. Journal of Vision, 10(6),
1-14.
carrasco, M. (2011). visual attention: the past 25 years. Vision
Research, 51(13), 1484-1525.
castiello, U., & Umiltà, c. (1990). size of the attentional focus and
efficiency of processing. Acta Psychologica, 73(3), 195-209.
cheal, M., lyon, d. r., & hubbard, d. c. (1991). does attention have
different effects on line orientation and line arrangement dis-
crimination? The Quarterly Journal of Experimental Psychology
Section A, 43(4), 825-857.
correa, Á., lupiáñez, J., Madrid, e., & tudela, P. (2006). temporal
attention enhances early visual processing: A review and
new evidence from event-related potentials. Brain Research,
1076(1), 116-128.
coull, J. t., Frith, c. d., Büchel, c., & nobre, A. c. (2000). orienting
attention in time: Behavioural and neuroanatomical distinction
between exogenous and endogenous shifts. Neuropsychologia,
38(6), 808-819.
coull, J. t., & nobre, A. c. (1998). Where and when to pay atten-
tion: the neural systems for directing attention to spatial loca-
tions and to time intervals as revealed by both Pet and fMri.
The Journal of Neuroscience: The Official Journal of the Society
for Neuroscience, 18(18), 7426-7435.
dorfman, d. d., & Berbaum, K. s. (1986). rscore-J: Pooled rating-
method data: A computer program for analyzing pooled roc
curves. Behavior Research Methods, 18(5), 452-462.
enns, J. t., & di lollo, v. (2000). What’s new in visual masking?
Trends in Cognitive Sciences, 4(9), 345-352.
eriksen, c. W., & st James, J. d. (1986). visual attention within
and around the field of focal attention: A zoom lens model.
Perception & Psychophysics, 40(4), 225-240.
eriksen, c. W., & yeh, y. y. (1985). Allocation of attention in the vi-
sual field. Journal of Experimental Psychology: Human Perception
and Performance, 11(5), 583-597.
galera, c., & grünau, M. W. (2003). size and shape of the at-
tentional spotlight affect efficiency of processing. Journal of
Vision, 3(9), 575.
green, d. M., & swets, J. A. (1966). Signal detection theory and
psychophysics. new york: Wiley.
hasher, l., & Zacks, r. t. (1984). Automatic processing of funda-
mental information: the case of frequency of occurrence. The
American Psychologist, 39(12), 1372-1388.
haynes, J.-d., driver, J., & rees, g. (2005). visibility reflects dyna-
mic changes of effective connectivity between v1 and fusiform
cortex. Neuron, 46(5), 811-821.
hein, e., rolke, B., & Ulrich, r. (2006). visual attention and tempo-
ral discrimination: differential effects of automatic and volun-
tary cueing. Visual Cognition, 13(1), 29-50.
heinze, h. J., Mangun, g. r., Burchert, W., hinrichs, h., scholz,
M., Münte, t. F., et al. (1994). combined spatial and temporal
imaging of brain activity during visual selective attention in
humans. Nature, 372(6506), 543-546.
hopfinger, J., Buonocore, M., & Mangun, g. (2000). the neu-
ral mechanisms of top-down attentional control. Nature
Neuroscience, 3, 284-291.
Kentridge, r. W., heywood, c. A., & Weiskrantz, l. (1999). effects
of temporal cueing on residual visual discrimination in blind-
sight. Neuropsychologia, 37(4), 479-483.
Kentridge, r. W., nijboer, t. c. W., & heywood, c. A. (2008).
Attended but unseen: visual attention is not sufficient for
visual awareness. Neuropsychologia, 46(3), 864-869.
lamme, v. A. F. (2003). Why visual attention and awareness are
different. Trends in Cognitive Sciences, 7(1), 12-18.
ling, s., & carrasco, M. (2006). sustained and transient covert
attention enhance the signal via different contrast response
functions. Vision Research, 46(8-9), 1210-1220.
loftus, g. r., & Masson, M. e. J. (1994). Using confidence intervals
in within-subject designs. Psychonomic Bulletin & Review, 1,
476-490.
Macknik, s. l., & livingstone, M. s. (1998). neuronal correlates of
visibility and invisibility in the primate visual system. Nature
Neuroscience, 1(2), 144-149.
Macknik, s. l., & Martinez-conde, s. (2007). the role of feedback
in visual masking and visual processing. Advances in Cognitive
Psychology, 3, 125-152.
Michaels, c. F., & turvey, M. t. (1979). central sources of visual
masking: indexing structures supporting seeing at a single,
brief glance. Psychological Research, 41(1), 2-61.
Müller, h. J., & rabbitt, P. M. (1989). reflexive and voluntary
orienting of visual attention: time course of activation
and resistance to interruption. Journal of Experimental
Psychology: Human Perception and Performance, 15(2), 315-
330.

AdvAnces in cognitive PsychologyreseArch Article
http://www.ac-psych.org2011 (special issue) • volume 7 • 132-141141
neumann, o., & scharlau, i. (2007). visual attention and the
mechanism of metacontrast. Psychological Research, 71(6),
626-633.
nobre, A. c. (2001). orienting attention to instants in time.
Neuropsychologia, 39(12), 1317- 1328.
Posner, M. i. (1980). orienting of attention. Quarterly Journal of
Experimental Psychology, 32(1), 3.
reeves, A. (1982). Metacontrast U-shaped functions derive from
two monotonic processes. Perception, 11(4), 415-426.
smith, P. l. (2000). Attention and luminance detection: effects of
cues, masks, and pedestals. Journal of Experimental Psychology:
Human Perception and Performance, 26(4), 1401-1420.
smith, P. l., lee, y.-e., Wolfgang, B. J., & ratcliff, r. (2009). Attention
and the detection of masked radial frequency patterns: data
and model. Vision Research, 49(10), 1363-1377.
smith, P. l., & Wolfgang, B. J. (2004). the attentional dynamics of
masked detection. Journal of Experimental Psychology: Human
Perception and Performance, 30(1), 119-136.
tata, M. s. (2002). Attend to it now or lose it forever: selective
attention, metacontrast masking, and object substitution.
Perception & Psychophysics, 64(7), 1028-1038.
turvey, M. t. (1973). on peripheral and central processes in vision:
inferences from an information-processing analysis of masking
with patterned stimuli. Psychological Review, 80(1), 1-52.
Wickens, t. d. (2001). Elementary signal detection theory. oxford:
oxford University Press.
received 16.02.2011 | AccePted 27.09.2011
Related Documents











