Copyright # 2002 John Wiley & Sons, Ltd. Received 26 June 2001 Accepted 16 April 2002 AQUATIC CONSERVATION: MARINE AND FRESHWATER ECOSYSTEMS Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003) Published online 26 September 2002 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/aqc.542 The effects of shore-based diamond-diving on intertidal and subtidal biological communities and rock lobsters in southern Namibia ANDREA PULFRICH a,b , COLLEEN A. PARKINS b and GEORGE M. BRANCH b * a Pisces Environmental Services (Pty) Ltd, 22 Forest Glade, Tokai Rd., Tokai 7945, South Africa b Zoology Department, University of Cape Town, Rondebosch 7701, South Africa ABSTRACT 1. Divers mine shallow-water diamonds on the west coasts of South Africa and Namibia. In the process they cut kelp, suck up gravel that is sorted on the shore and then deposited intertidally, and uncover and overturn subtidal boulders. 2. The impacts of these divers on intertidal and subtidal reefs, and on the population structure of the commercially important rock lobster Jasus lalandii, were monitored for 5 years near L . uderitz in southern Namibia. Sampling was undertaken at unmined reference (control) sites and at sites mined at various times in the past. Percentage cover and densities of benthic organisms were recorded, and the abundance, length-frequencies and sex ratios of rock lobsters determined. 3. High natural variability in benthic community structure made it difficult to distinguish mining impacts even a short time after mining had ceased. Nevertheless, mining reduced the species diversity and abundance of both intertidal and subtidal communities. Recovery did, however, occur within 2 years. During, or immediately after mining, the intertidal community became characterized by the near disappearance of grazers, proliferation of fast-growing, opportunistic foliose algae and decreased cover of filter-feeders. Subsequently, grazers increased, curtailing the algae. In contrast, mining reduced subtidal algal cover. Kelp was cut to facilitate mining, foliose algae were removed and smothered as rocks were overturned, and reef-building filter-feeders diminished. 4. The abundance and population structure of rock lobsters appeared to be largely determined by environmental conditions and habitat suitability. Their abundance and catch per unit effort were more often higher at reference sites and recently mined sites than at sites mined >5 years previously. Sex ratios and mean sizes of rock lobsters were not statistically affected by mining. Mining does alter the biological and physical characteristics of the habitat, and in the process, it may temporarily increase the availability of suitable rock lobster habitats. No negative impacts of shore-based diamond-diving on rock lobsters were detected. Copyright # 2002 John Wiley & Sons, Ltd. KEY WORDS: Diamond mining; Shore-based contractor divers; Disturbance; Rocky shore communities; Rock lobsters; Spiny lobsters *Correspondence to: G.M. Branch, Zoology Department, University of Cape Town, Private Bag, Rondebosch 7701, South Africa. E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Copyright # 2002 John Wiley & Sons, Ltd. Received 26 June 2001Accepted 16 April 2002
AQUATIC CONSERVATION: MARINE AND FRESHWATER ECOSYSTEMS
Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)
Published online 26 September 2002 in Wiley InterScience(www.interscience.wiley.com). DOI: 10.1002/aqc.542
The effects of shore-based diamond-diving on intertidal andsubtidal biological communities and rock lobsters in southern
Namibia
ANDREA PULFRICHa,b, COLLEEN A. PARKINSb and GEORGE M. BRANCHb*aPisces Environmental Services (Pty) Ltd, 22 Forest Glade, Tokai Rd., Tokai 7945, South Africa
bZoology Department, University of Cape Town, Rondebosch 7701, South Africa
ABSTRACT
1. Divers mine shallow-water diamonds on the west coasts of South Africa and Namibia. In theprocess they cut kelp, suck up gravel that is sorted on the shore and then deposited intertidally, anduncover and overturn subtidal boulders.2. The impacts of these divers on intertidal and subtidal reefs, and on the population structure of
the commercially important rock lobster Jasus lalandii, were monitored for 5 years near L .uuderitz insouthern Namibia. Sampling was undertaken at unmined reference (control) sites and at sites minedat various times in the past. Percentage cover and densities of benthic organisms were recorded, andthe abundance, length-frequencies and sex ratios of rock lobsters determined.3. High natural variability in benthic community structure made it difficult to distinguish mining
impacts even a short time after mining had ceased. Nevertheless, mining reduced the species diversityand abundance of both intertidal and subtidal communities. Recovery did, however, occur within 2years. During, or immediately after mining, the intertidal community became characterized by thenear disappearance of grazers, proliferation of fast-growing, opportunistic foliose algae anddecreased cover of filter-feeders. Subsequently, grazers increased, curtailing the algae. In contrast,mining reduced subtidal algal cover. Kelp was cut to facilitate mining, foliose algae were removedand smothered as rocks were overturned, and reef-building filter-feeders diminished.4. The abundance and population structure of rock lobsters appeared to be largely determined by
environmental conditions and habitat suitability. Their abundance and catch per unit effort weremore often higher at reference sites and recently mined sites than at sites mined >5 years previously.Sex ratios and mean sizes of rock lobsters were not statistically affected by mining. Mining does alterthe biological and physical characteristics of the habitat, and in the process, it may temporarilyincrease the availability of suitable rock lobster habitats. No negative impacts of shore-baseddiamond-diving on rock lobsters were detected.Copyright # 2002 John Wiley & Sons, Ltd.
KEY WORDS: Diamond mining; Shore-based contractor divers; Disturbance; Rocky shore communities; Rock
lobsters; Spiny lobsters
*Correspondence to: G.M. Branch, Zoology Department, University of Cape Town, Private Bag, Rondebosch 7701, South Africa.E-mail: [email protected]

INTRODUCTION
Shortly after the discovery of the first diamonds in L .uuderitz in 1908, public access to much of the Namibiancoastline southwards towards the Orange River mouth became restricted (Schneider, 1998), andconsequently it has been maintained in an almost pristine condition. However, at particular sites on thecoast there have been historical perturbations of intertidal and shallow subtidal areas due to diamondmining.
Marine diamonds have been extracted from coastal deposits in the area since early in the 1960s. Innearshore waters, diamonds are distributed in gullies and potholes, having accumulated in gravel beds nearthe bedrock through wave sorting. Shore concession areas under the jurisdiction of the large diamondcompanies are leased to smaller contractors who mine the ore bodies by diver-operated suction pumps,operating directly from the shore down to depths of 10m in small bays. For convenience, we term theseactivities ‘shore-based diver-operated‘ mining, to distinguish them from offshore mining that occurs inmuch deeper water and relies on boats and remotely controlled suction pumps. A shore-based operationtypically consists of 2–3 divers, their land-based assistants, and a tractor modified to drive a rotary classifierand centripetal pump to which a 20-cm diameter suction hose is attached. As the contractors locate thisheavy equipment as close to the sea as possible, access to the water is sometimes achieved by blasting ormechanical damage in the supratidal and intertidal regions. In order to reach the diamondiferous depositsin the surf zone and beyond, kelp (mostly Laminaria pallida) is cut to allow easy movement of the suctionhoses and airlines. The divers, operating on surface-supplied diving equipment, guide the terminal end ofthe hose into the gravel deposits, which are sucked up and delivered to the classifier on the shore. To reachthe deeper deposits, boulders of all sizes usually need to be broken up and overturned, and these may berelocated by divers into rock-piles, or dragged from gullies at low tide by tractor and chains, and depositedat higher tidal levels. Rocks are also uncovered through removal of surrounding gravel. Relocation andremoval of rocks during mining operations have a destructive impact on the associated biota and alters thephysical characteristics of the gullies. The diamond concentrate is separated from the gravel by theclassifier, bagged and transported to central sorting houses. The wastes that remain comprise both oversizegravel and pebbles (‘‘tailings’’), which are allowed to accumulate around the screening units, and smallerparticles (‘‘fines’’), which are returned to the sea as a slurry across the intertidal zone. This waste materialhas smothering and scouring effects. In addition, mechanical disturbance is caused by trampling andabrasion due to the dragging of suction hoses and the movement of machinery.
As part of a coastal biological monitoring programme launched by Namdeb Diamond Corporation (Pty)Ltd at Elizabeth Bay, a study was initiated south of L .uuderitz in early 1994 specifically to examine the effectsof shore-based, diver-operated mining activities on the intertidal and subtidal community structure of rockyreefs. Of particular interest was the potential negative effects of these mining activities on the abundanceand distribution of the commercially important West Coast rock lobster (Jasus lalandii). This papersummarizes the results of 5 years of intertidal and 4 years of subtidal monitoring in the vicinity of ElizabethBay, at sites mined by contractors at various times in the past, and at comparable reference sites.
METHODS
Study area
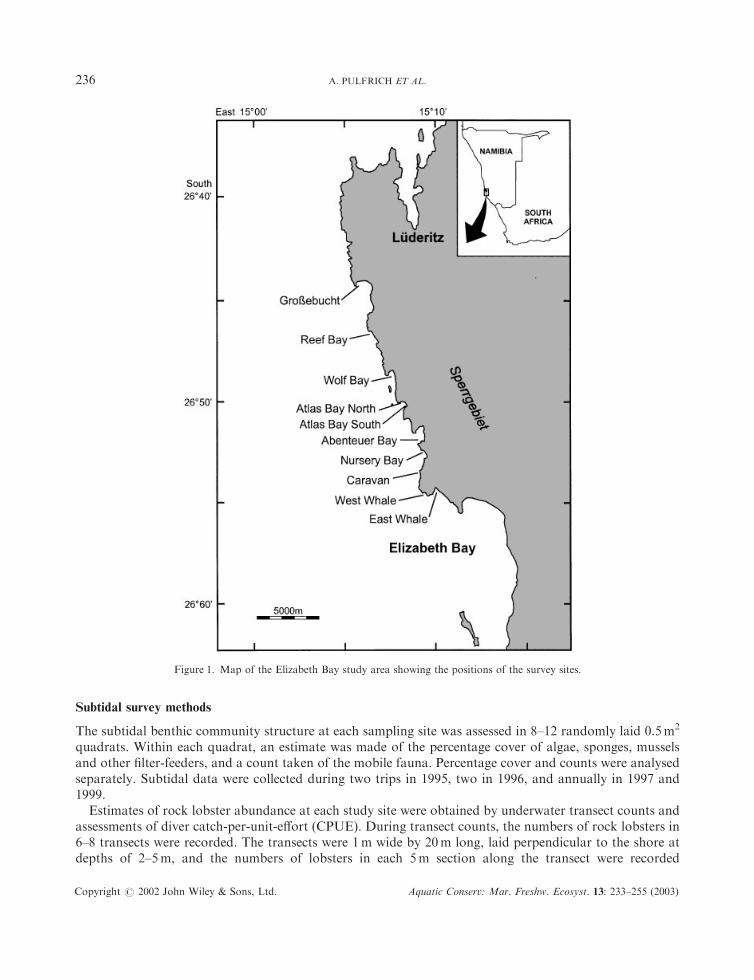
The southern Namibian coastline is a transitional zone between two large cool-temperate marinebiogeographic provinces } the southern Namaqua and northern Namib provinces (Emanuel et al., 1992).The area between L .uuderitz and Elizabeth Bay is characterized mostly by rocky shores, interrupted by small,protected bays and sandy beaches. The open coast is exposed to strong wave action, and most
A. PULFRICH ET AL.234
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

diver-operated mining is limited to sheltered bays to maximize diving opportunities. The communitystructure of the subtidal benthos in the sheltered bays south of L .uuderitz is typical of the West Coast kelpbed environment (see Branch and Griffiths, 1988 for a review). There is a high biomass of kelp (mainlyLaminaria pallida) and an associated understorey community that includes crustose coralline algae andsponges, with upright coralline algae occurring in more silty areas. The Cape reef worm, Gunnarea capensis,covers a large percentage (ca 40%) of the rock under the kelp canopy at some sites. In the intertidal zone ofsheltered bays, limpets (particularly Cymbula granatina) often reach extraordinary population densities andrestrict algal biomass (Bustamante et al., 1995). Filter-feeder biomass is low there, but reaches high levelson the open coast (Bustamante and Branch, 1996), where it is dominated by the mussel Mytilusgalloprovincialis and G. capensis.
Six sheltered sites at which shore-based mining had been conducted were chosen (Figure 1). At three ofthese (Caravan, Nursery Bay and Atlas Bay North), contractors had completed their mining operationsmore than 5 years prior to the commencement of the study, and these sites are referred to as being mined>5 years previous to sampling. At the remaining three sites (East Whale, Wolf Bay and Reef Bay), mininghad ceased less than 6 months before the start of the intertidal study. Subtidal sampling at these sitescommenced within 2 years of mining and continued for a further 4 years. These latter sites are collectivelyreferred to as being mined 52 years previously, although the post-mining period rose to 5 years by the endof the sampling. Two unmined sites (West Whale and Abenteuer Bay), served as reference (control) sites forsubtidal sampling, and Atlas Bay South and Gro�ebucht were used as undisturbed reference sites for theintertidal sampling (Table 1). It should be noted that for the April 1999 sampling period, Wolf Bay (firstmined in 1993) was re-classified as being mined >5 years previously. Re-commencement of mining atNursery Bay and Reef Bay in early 1999 provided the opportunity to study the immediate effects of miningand the short-term path of recovery. For the purposes of analysis, samples taken after the renewed miningwere re-classified as being mined 52 years previously.
Pre-mining baseline data were available for the intertidal rocky-shore communities at Wolf Bay only.This site was extensively mined by contractor divers between December 1993 and early 1994. Prior to this,however, it had served as a reference site in 1993 for a separate study by Pulfrich et al. (2003) on theintertidal impacts of fine sediments discharged from the terrestrial mining plant at Elizabeth Bay. Datacollected in November 1993 thus provided information on the pre-mining structure of the intertidalcommunities in Wolf Bay. At the remaining sampling localities, however, it was not possible to implementthe ‘Before-After/Control-Impact’ (BACI) sampling strategy proposed by Green (1979, 1993) andUnderwood (1992, 1993, 1994). Sites mined at some time in the past could, however, be compared with theundisturbed reference sites. Two of the subtidal sites (Reef Bay and Nursery Bay) were originally mined,but then left untouched for at least 6 years, only to be mined again early in 1999. This provided theopportunity to do a before/after comparison of rock lobster CPUE at these sites (comparing the ‘before’data for September 1995 and May 1996 with the ‘after’ data of April 1999). These sets of data could also bematched against changes at two reference sites (West Whale and Abenteuer Bay) that remained unminedover this period, to provide impact/control comparisons.
Intertidal survey methods
At each intertidal site, three transects were surveyed perpendicular to the shore. On each transect, a total of6–7 quadrats of 0.5m2 were positioned in a stratified manner at regular intervals from mean low-waterspring to mean high-water spring-tide levels to ensure that comparisons between different sites wereequivalent. The quadrats were divided into a regular 50� 50 mm mesh, giving 171 intersecting points in a1� 0.5 m frame. Individual species were identified and recorded under each intersection point to measuretheir percentage cover. Mobile invertebrates in the quadrats were counted. Intertidal sites were sampled onthree occasions during 1994, a further three during 1995 and annually thereafter.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 235
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)
Subtidal survey methods
The subtidal benthic community structure at each sampling site was assessed in 8–12 randomly laid 0.5m2
quadrats. Within each quadrat, an estimate was made of the percentage cover of algae, sponges, musselsand other filter-feeders, and a count taken of the mobile fauna. Percentage cover and counts were analysedseparately. Subtidal data were collected during two trips in 1995, two in 1996, and annually in 1997 and1999.
Estimates of rock lobster abundance at each study site were obtained by underwater transect counts andassessments of diver catch-per-unit-effort (CPUE). During transect counts, the numbers of rock lobsters in6–8 transects were recorded. The transects were 1m wide by 20m long, laid perpendicular to the shore atdepths of 2–5m, and the numbers of lobsters in each 5m section along the transect were recorded
Figure 1. Map of the Elizabeth Bay study area showing the positions of the survey sites.
A. PULFRICH ET AL.236
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)
separately. Results are expressed as the mean number of rock lobsters per square metre. CPUE wasquantified as the number of rock lobsters caught by each of the two divers in two 15 min intervals. Theresults are given as the mean number of animals caught per minute by the two divers. The rock lobsterswere subsequently measured, sexed and returned to the sea.
As a precaution against biases arising from differing capabilities among divers, varying combinationsof the same five divers were used to collect the CPUE data. Furthermore, Students’ t-tests showedno significant differences (p > 0:05) between the catch successes of divers during each sampling period.Pulfrich et al. (2003) have shown that the results from transects (y, countsm�2) and from CPUE (x, no.caughtmin�1) are significantly correlated according to the following equation:
y ¼ �1:08þ exp ð0:29þ 0:18xÞ; r ¼ 0:52 n ¼ 38; p ¼ 0:001
Data analysis
Non-parametric multivariate analyses, particularly hierarchical clustering (Bray and Curtis, 1957) andmulti-dimensional scaling, were applied to assess patterns of association in the intertidal and subtidalbenthic communities, using the software package PRIMER (Plymouth Routines In Multivariate EcologicalResearch) (Clarke, 1993; Clarke and Warwick, 1994). Following the recommendations of Clarke andWarwick (1994), a 4th-root transformation was applied to the data.
The non-parametric analysis of similarity (ANOSIM) was used to test for differences between groups ofsamples. The groups chosen a priori in this study were samples from impacted and reference sites. The teststatistic (R) separated the effects of natural spatial variability in community structure from changesassociated with mining disturbance itself, with significance levels being set at p50:05.
The PRIMER programme SIMPER was used to re-examine the data set in the light of the multivariateresults, to determine which taxa were responsible for any patterns observed in the community structure.Multifactoral ANOVA and non-parametric statistical tests such as Kruskal–Wallis were used to test fordifferences between pairs or groups of samples, employing STATISTICA (Version 6, 2002 edition).
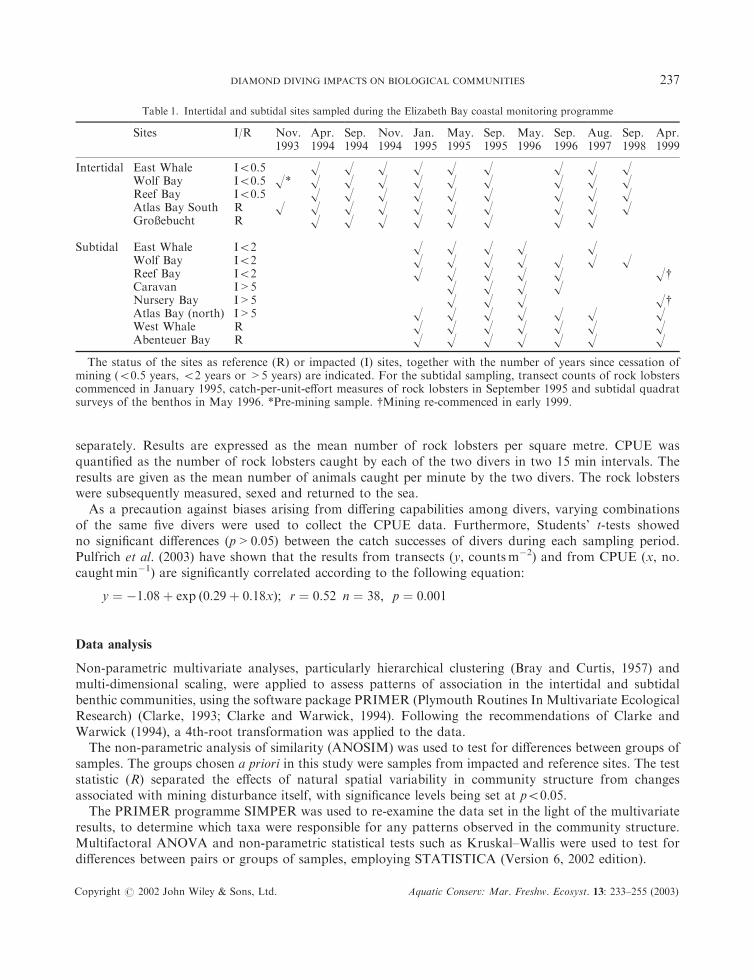
Table 1. Intertidal and subtidal sites sampled during the Elizabeth Bay coastal monitoring programme
Sites I/R Nov.1993
Apr.1994
Sep.1994
Nov.1994
Jan.1995
May.1995
Sep.1995
May.1996
Sep.1996
Aug.1997
Sep.1998
Apr.1999
Intertidal East Whale I50.5 H H H H H H H H HWolf Bay I50.5 H* H H H H H H H H HReef Bay I50.5 H H H H H H H H HAtlas Bay South R H H H H H H H H H HGro�ebucht R H H H H H H H H
Subtidal East Whale I52 H H H H HWolf Bay I52 H H H H H H HReef Bay I52 H H H H H HyCaravan I>5 H H H HNursery Bay I>5 H H H HyAtlas Bay (north) I>5 H H H H H H HWest Whale R H H H H H H HAbenteuer Bay R H H H H H H H
The status of the sites as reference (R) or impacted (I) sites, together with the number of years since cessation ofmining (50.5 years, 52 years or >5 years) are indicated. For the subtidal sampling, transect counts of rock lobsterscommenced in January 1995, catch-per-unit-effort measures of rock lobsters in September 1995 and subtidal quadratsurveys of the benthos in May 1996. *Pre-mining sample. yMining re-commenced in early 1999.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 237
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

RESULTS
Intertidal benthos
A total of 85 species was encountered at the survey sites in the study area. Of these, 54 were algae, and theremaining 31 invertebrates. The major space-occupiers in the reference sites were typical of those onsheltered west-coast shores (Penrith and Kensley, 1970a; Bustamante and Branch, 1995), with the limpetCymbula granatina being dominant in the low-shore and replaced by Scutellastra granularis and Siphonariacapensis in the mid- to high-shore. Filter-feeders were scarce. Several algae were present but not nearly asabundant as they became at impacted sites. A species list is provided in Appendix A.
The ordination plots resulting from the cluster analyses of the intertidal benthic communities at themined and reference sites from 1994 to 1998 are shown in Figure 2. Clustering of individual sites occurredat levels of 60–70% similarity. Due to high variability between transects within sites, clear separation intomined and reference sites was not discernible. Despite this, ANOSIM analyses revealed significantdifferences (Global R-statistic, p50:05) between reference and mined sites in 1994 and 1995 (Table 2(A)).Thereafter, the differences were not significant, implying that recovery is complete in 2–3 years. In 1993,prior to the mining of Wolf Bay, ANOSIM analysis could not detect any significant differences betweenWolf Bay and the other reference site at Atlas Bay South. However, after Wolf Bay was mined in 1994,significant differences existed for a period of 2 years (Table 2(B)). Again, a recovery period of 2–3 years isimplied.
During 1994 and 1995, when the sites were sampled on three occasions during the year, there were nosignificant temporal differences among the intertidal communities at the reference sites (ANOSIM, 1994:R ¼ 0:070, p ¼ 0:193; 1995: R ¼ 0:120, p ¼ 0:096). At mined sites, temporal differences were significantduring 1994, but not in 1995 (1994: R ¼ 0:199, p ¼ 0:004; 1995: R ¼ 0:106, p ¼ 0:064).
Having identified community differences between mined and reference sites using ANOSIM, the data setwas re-arranged into functional groups, and univariate analyses based on the original data set weresubsequently used to establish which functional groups were responsible for the differences. Temporalvariations in the mean percentage cover of filter-feeders, encrusting coralline and foliose algae, and densitiesof grazers and predators are illustrated in Figure 3. A noticeable difference between the mined and referencesites was the lower cover of filter-feeders at the former, especially in the first 2 years post-mining (1994 and1995), after which a gradual increase occurred (Figure 3(C)). This difference was significant for all samplingperiods during 1994 and 1995 (Kruskal–Wallis test, p50.05). Subsequent ANOVAs on the individual filter-feeding species revealed that this was attributable primarily to the significantly higher cover of the tube-building gastropod Dendropoma corallinaceus and the reef-building polychaete Gunnarea capensis(ANOVA, p50:05) at reference sites.
The percentage cover of foliose and crustose coralline algae at the reference sites was initially highlyvariable (Figure 3(C)). This is likely to reflect seasonal changes in abundance of algae, with higher values offoliose algae in autumn (April–May) and early summer (November). From 1995 onwards, sampling wasstandardised and only undertaken in spring (August–September). Values during this season varied littlebetween years until 1998, when a bloom was recorded following mining trials at Atlas Bay late in 1997. Atthe mined sites, particularly at Wolf Bay, this seasonality was overpowered by a significant increase infoliose algal cover for as long as 2 years after mining activities ceased. Following ANOVA (see Table 3),Tukey post-hoc tests showed that at all impacted sites combined, algae increased significantly betweenNovember 1993 and the period spanning from April to November 1994, and then decreased again whencomparing November 1994 with the period September 1995–September 1998 (p50:05 in all cases). WolfBay was analysed separately because it was the only site at which data were gathered before mining. Miningbegan there in April 1994, and significant increases in algae were recorded from November 1994 to May1995 (p50:04 in all cases).
A. PULFRICH ET AL.238
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

Subsequent to mining, decreases in the density of grazers at impacted sites (Figure 3(B)) coincided withincreases in foliose algae (Figure 3(C)). At Wolf Bay, the densities of grazers declined by 40% from the pre-mining levels of November 1993 and those at all mined sites remained at a comparatively low level untilmid-1995. There was a gradual increase in grazer densities at the mined sites between 1995 and 1997, but atReef Bay specifically, a significant decline in grazer population densities was recorded during 1998,concomitant with an increase in foliose algae, suggesting some unrecorded disturbance at this site. Overtime, the mean densities of grazers and cover of foliose algae were negatively correlated at the mined sites(R ¼ �0:805 p50:01).
Stress = 0.08
September 1998
Stress = 0.18
August 1997
Stress = 0.12
September 1996
Stress = 0.16
September 1995
Stress = 0.18
May 1995
Stress = 0.16
January 1995
Stress = 0.16
November 1994
Stress = 0.16
September 1994
Stress = 0.14
April 1994
Reference Sites Contractor-mined SitesGroßebucht
Atlas Bay South
East WhaleWolf BayReef Bay
Figure 2. Ordination plots of the intertidal benthic community associations at contractor-mined and reference sites surveyed fromApril 1994 to September 1998. Clusters forming at 60% Bray–Curtis similarity are circled. Each data point represents a single transect.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 239
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)
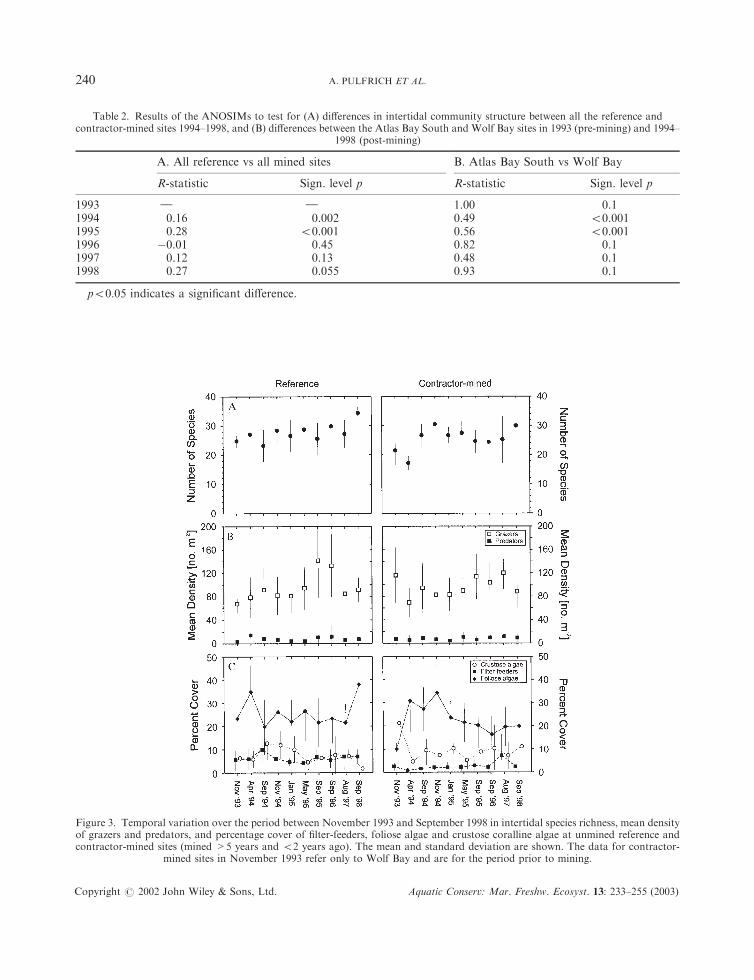
Table 2. Results of the ANOSIMs to test for (A) differences in intertidal community structure between all the reference andcontractor-mined sites 1994–1998, and (B) differences between the Atlas Bay South and Wolf Bay sites in 1993 (pre-mining) and 1994–
1998 (post-mining)
A. All reference vs all mined sites B. Atlas Bay South vs Wolf Bay
R-statistic Sign. level p R-statistic Sign. level p
1993 } } 1.00 0.11994 0.16 0.002 0.49 50.0011995 0.28 50.001 0.56 50.0011996 �0.01 0.45 0.82 0.11997 0.12 0.13 0.48 0.11998 0.27 0.055 0.93 0.1
p50:05 indicates a significant difference.
Figure 3. Temporal variation over the period between November 1993 and September 1998 in intertidal species richness, mean densityof grazers and predators, and percentage cover of filter-feeders, foliose algae and crustose coralline algae at unmined reference andcontractor-mined sites (mined >5 years and 52 years ago). The mean and standard deviation are shown. The data for contractor-
mined sites in November 1993 refer only to Wolf Bay and are for the period prior to mining.
A. PULFRICH ET AL.240
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)
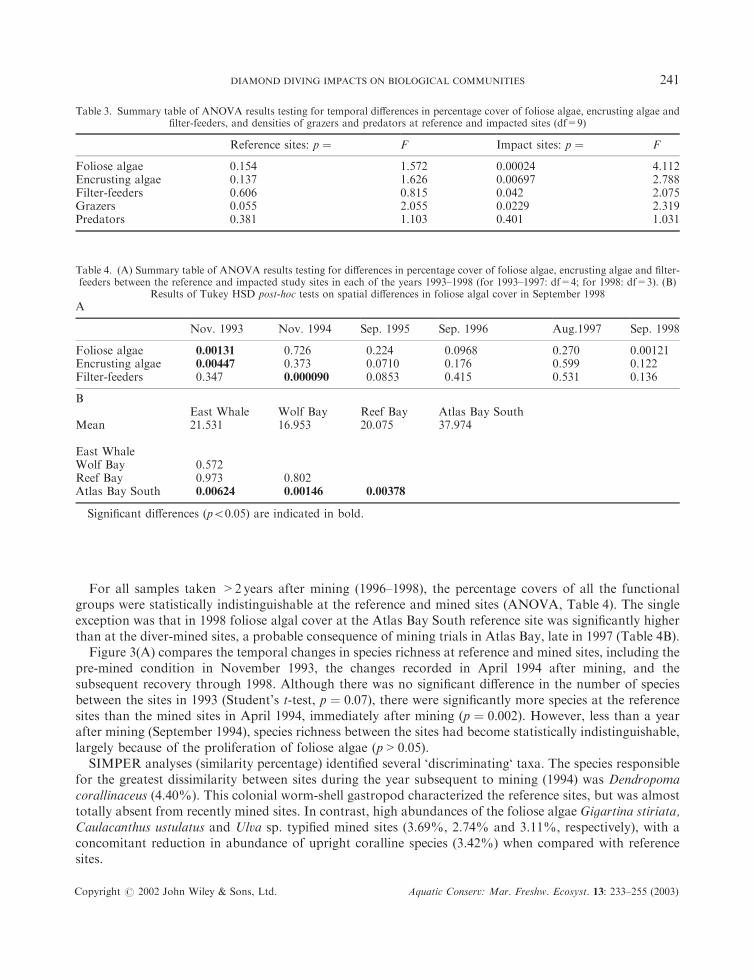
For all samples taken >2years after mining (1996–1998), the percentage covers of all the functionalgroups were statistically indistinguishable at the reference and mined sites (ANOVA, Table 4). The singleexception was that in 1998 foliose algal cover at the Atlas Bay South reference site was significantly higherthan at the diver-mined sites, a probable consequence of mining trials in Atlas Bay, late in 1997 (Table 4B).
Figure 3(A) compares the temporal changes in species richness at reference and mined sites, including thepre-mined condition in November 1993, the changes recorded in April 1994 after mining, and thesubsequent recovery through 1998. Although there was no significant difference in the number of speciesbetween the sites in 1993 (Student’s t-test, p ¼ 0:07), there were significantly more species at the referencesites than the mined sites in April 1994, immediately after mining (p ¼ 0:002). However, less than a yearafter mining (September 1994), species richness between the sites had become statistically indistinguishable,largely because of the proliferation of foliose algae (p > 0:05).
SIMPER analyses (similarity percentage) identified several ‘discriminating‘ taxa. The species responsiblefor the greatest dissimilarity between sites during the year subsequent to mining (1994) was Dendropomacorallinaceus (4.40%). This colonial worm-shell gastropod characterized the reference sites, but was almosttotally absent from recently mined sites. In contrast, high abundances of the foliose algae Gigartina stiriata,Caulacanthus ustulatus and Ulva sp. typified mined sites (3.69%, 2.74% and 3.11%, respectively), with aconcomitant reduction in abundance of upright coralline species (3.42%) when compared with referencesites.
Table 3. Summary table of ANOVA results testing for temporal differences in percentage cover of foliose algae, encrusting algae andfilter-feeders, and densities of grazers and predators at reference and impacted sites (df=9)
Reference sites: p ¼ F Impact sites: p ¼ F
Foliose algae 0.154 1.572 0.00024 4.112Encrusting algae 0.137 1.626 0.00697 2.788Filter-feeders 0.606 0.815 0.042 2.075Grazers 0.055 2.055 0.0229 2.319Predators 0.381 1.103 0.401 1.031
Table 4. (A) Summary table of ANOVA results testing for differences in percentage cover of foliose algae, encrusting algae and filter-feeders between the reference and impacted study sites in each of the years 1993–1998 (for 1993–1997: df=4; for 1998: df=3). (B)
Results of Tukey HSD post-hoc tests on spatial differences in foliose algal cover in September 1998
A
Nov. 1993 Nov. 1994 Sep. 1995 Sep. 1996 Aug.1997 Sep. 1998
Foliose algae 0.00131 0.726 0.224 0.0968 0.270 0.00121Encrusting algae 0.00447 0.373 0.0710 0.176 0.599 0.122Filter-feeders 0.347 0.000090 0.0853 0.415 0.531 0.136
BEast Whale Wolf Bay Reef Bay Atlas Bay South
Mean 21.531 16.953 20.075 37.974
East WhaleWolf Bay 0.572Reef Bay 0.973 0.802Atlas Bay South 0.00624 0.00146 0.00378
Significant differences (p50:05) are indicated in bold.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 241
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

Subtidal benthos
Multivariate cluster analysis performed on the integrated data for all sampling periods (Figure 4)emphasized the high degree of variability in community structure both within and between sites. Wehypothesized that mining would have a significant impact on subtidal benthic communities, and thusexpected to identify statistical differences in the communities at mined compared to unmined reference sites.ANOSIM, however, failed to detect significant differences between the benthic communities at referencesites and those at sites mined in the past, whether they were originally mined >5 or 52 years ago(R ¼ �0:110, p ¼ 0:85). Further ANOSIM comparisons investigating annual inter-site differences inbenthic community structure identified significant differences between the contractor-mined and referencesites for two of the four sampling dates, i.e. September 1996 and April 1999 (Table 5). The difference for thelatter date can be explained by the re-commencement of shore-based mining activities at Nursery and ReefBays early in 1999.
Cluster analyses done on individual quadrats showed that most samples clustered together, whether theywere taken from reference sites, sites mined >5 years ago, or sites mined 52 years previous to the start ofsampling (Figure 5). Some quadrats did separate out, and these outlying samples were all taken on sandy orrocky substrata characterized by low benthic cover (525%). In Nursery Bay and Reef Bay, recently re-mined areas were recognizable by clearly distinguishable rock-piles and boulder aggregations created bydivers to gain access to the deeper gravel deposits. In the process, the benthic cover on the sides and lower
Contractor-mined Sites <2 yrs>5 yrs
Reef Bay
Reference Sites Abenteuer BayWest Whale
East Whale
Atlas Bay NorthNursery BayCaravan
Wolf Bay
Stress=0.15
Bray-Curtis Similarity80 90 100
Figure 4. Dendrogram and ordination plot showing the subtidal benthic community associations in reference and contractor-minedsites for 1996–1999. Each data point represents pooled quadrat samples for 1 year.
A. PULFRICH ET AL.242
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

surfaces of over-turned rocks was disturbed and often buried, and rocks that were uncovered throughremoval of surrounding gravel were initially totally devoid of benthos. These newly exposed barren surfaceswere subsequently recolonized by successional communities including barnacle recruits and algal turf
Table 5. Results of ANOSIMs to test for annual differences in benthic composition between subtidal reference sites and those minedby contractors 55 years and >5 years ago
Sites R-statistic Significance level p (%)
May 1996 0.037 0.093Sep. 1996 0.067 0.025Aug. 1997 0.084 0.057Apr. 1999 0.171 50.001
p50:05 indicates a significant difference in community structure between sites.
Stress=0.19
Bra
y-C
urtis
Sim
ilarit
y
80
70
60
50
40
90
100
Contractor-mined Sites <2 yrs>5 yrs
Reef Bay
Reference Sites Abenteuer BayWest Whale
East Whale
Atlas Bay NorthNursery BayCaravan
Wolf Bay
Figure 5. Dendrogram and ordination plot showing the subtidal benthic community associations in reference and contractor-minedsites in April 1999. Each data point represents a quadrat. Arrows indicate those samples that were taken from subtidal rubble piles left
by miners. All other outlying samples were taken on sandy patches.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 243
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

(A. Pulfrich, pers. obs.). The outlying samples evident in Figure 5 predominantly comprised quadrats takenin such disturbed habitats, indicating a short-term effect.
The taxa identified by the SIMPER analysis as being consistently abundant throughout the reference andmined sites were red algae and filter-feeders, which respectively accounted for 29% and 22–23% of thesimilarity. Conversely, green algae were consistently good discriminators. They accounted for 28.5% of thedifferences in community structure between treatments, being more abundant at sites mined 2–5 yearspreviously than in reference sites or older mined sites.
The percentage cover of kelp, red algae, green algae, coralline algae and sessile filter-feeding organisms isshown in Figure 6. Univariate Kruskal–Wallis tests of mean percentage cover detected no significantdifferences in any of the algal groups, but significantly higher values for filter-feeders (primarily Cape reefworm Gunnarea capensis) during 1996, at the sites mined 55 years ago (May 1996: p ¼ 0:03, September1996, p ¼ 0:03). This effect was particularly obvious at Reef Bay. This pattern persisted during thefollowing year although the difference was not significant.
The re-commencement of contractor-mining at Nursery and Reef Bays in early 1999 resulted in asubstantial decrease in overall cover, and the lowest values recorded throughout the survey (Figure 6). Inparticular, there was a significant decrease in the percentage cover of kelp at Nursery Bay (Mann–WhitneyU-test, p ¼ 0:03) but not at Reef Bay (p ¼ 0:21) and a significant decrease in filter-feeders at both sites(Nursery Bay, p ¼ 0:04; Reef Bay, p ¼ 0:004), compared with levels in previous years at these sites. Thecover of filter-feeders at Wolf Bay similarly decreased significantly in 1999 (Kruskal–Wallis test, p ¼ 0:03)compared to the previous sample taken in August 1997. There were, however, no significant changes in thecover of filter-feeders or kelp during the course of the study at the reference sites, or at Atlas Bay North(mined >5years ago). The cover of red and green foliose algae at recently mined locations also tended to belower relative to that before mining was reinitiated, although the differences were not significant (Kruskal–Wallis test, p > 0:05). Collectively, these results support the significant differences detected betweentreatments by the multivariate ANOSIM.
Throughout the study, no significant differences in population densities of subtidal mobile predators,scavengers or grazers were detected between mined and reference sites (Kruskal–Wallis test, p > 0:05).During April 1999 when sampling was conducted shortly after mining had been reinitiated at Nursery andReef Bays, the densities of grazers and predators were higher at the mined sites, although not significantlyso (grazers: p ¼ 0:08, predators, p ¼ 0:07).
rock lobster surveys
The average abundance of rock lobsters appeared to be highest at either reference sites or those mined 55years ago, and lowest at sites mined >5 years previously (Figure 7). Differences were, however, onlysignificant in September 1996 and April 1999, when the reference site(s) had significantly more lobsters thanany of the mined sites (ANOVA, p50:05, see Table 6). An ANOVA of the integrated data for all yearsconfirmed that abundances were higher at reference than mined sites (p50:001). Two of the sites (Reef Bayand Nursery Bay) were mined again early in 1999, but counts and CPUE immediately after miningremained as high as at any stage of the sampling (Figure 7). A BACI analysis (repeated measures ANOVA)was used to compare the Reef and Nursery Bay pre-mining data (September 1995 and May 1996) and post-mining data (April 1999) with those for equivalent dates for the West Whale and Abenteuer Bay referencesites. This confirmed that there was no significant effect of mining on the CPUE of rock lobsters (F ¼ 0:090,df=2, p ¼ 0:91).
CPUE showed similar patterns. Significant differences in CPUE occurred between the treatments in Mayand September 1996 and in August 1997, with catches at sites mined >5 years previously being lower thanat reference and/or recently mined sites (ANOVA, p50:05). Differences between reference sites and thosemined 55 years ago were insignificant (Student’s t-test, p ¼ 0:77).
A. PULFRICH ET AL.244
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

High temporal variability in rock lobster abundance and CPUE was evident at all sites, with differencesbetween sampling periods being significant (ANOVA, abundance: p50:001; CPUE: p50:001). On averagethe counts and catches of rock lobsters at the West Whale reference site were significantly higher than atAbenteuer Bay, the second reference site (Mann–Whitney U-test, counts: p ¼ 0:00006, CPUE: p ¼ 0:04),indicating high natural variability between sampling locations. The influence of sea conditions at the timeof sampling on diver-recorded densities and catch success must also be kept in mind. Exceptionally highcounts and catches were made at West Whale in 1999 probably due to unusually favourable divingconditions. If these data were omitted from the analysis, counts and catches of rock lobsters at West Whale
Figure 6. Percentage cover of kelp, red algae, green algae, coralline algae and filter-feeding organisms recorded in subtidal quadrats atthe contractor-mined and reference sites between May 1996 and April 1999. Asterisks indicate where no samples were taken. Note:
mining re-commenced in Nursery and Reef Bay in early 1999.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 245
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

were still higher than at Abenteuer Bay, but this difference was then significant for the transect counts only(Mann–Whitney U-test, counts: p ¼ 0:03, CPUE: p ¼ 0:20).
0 0
2 2
4 4
6 6
8 8
0 0
2 2
4 4
6 6
8 8
0 0
2 2
4 4
6 6
8 8
0 0
2 2
4 4
6 6
8 8
0 0
2 2
4 4
6 6
8 8
0 0
2 2
4 4
6 6
8 8
Roc
k-lo
bste
r A
bund
ance
[num
ber
per
m]
2C
PU
E [num
ber caught per minute]
Reference SitesMined <5 yrs ago Mined >5 yrs ago
September 1995
January 1995
May 1996
September 1996
August 1997
April 1999
Eas
t Wha
le
Wol
f Bay
Ree
f Bay
Car
avan
Nur
sery
Bay
Atla
s B
ay N
orth
Wes
t Wha
le
Abe
nteu
er B
ay
*
*
*
*
*
*
*
*
Figure 7. Mean rock lobster abundance (bars) and catch-per-unit-effort (CPUE) (dots) at the contractor-mined and reference sitesbetween January 1995 and April 1999. Asterisks indicate where no samples were taken. The standard deviations are also shown. Note:
Mining re-commenced at Nursery and Reef Bay in early 1999.
A. PULFRICH ET AL.246
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)
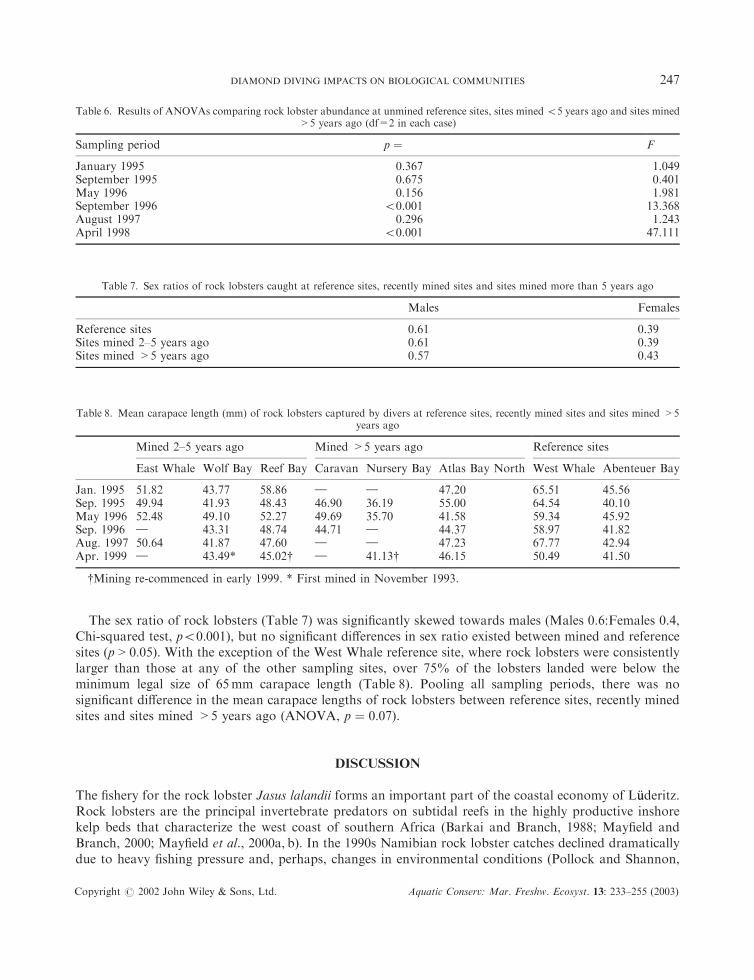
The sex ratio of rock lobsters (Table 7) was significantly skewed towards males (Males 0.6:Females 0.4,Chi-squared test, p50:001), but no significant differences in sex ratio existed between mined and referencesites (p > 0:05). With the exception of the West Whale reference site, where rock lobsters were consistentlylarger than those at any of the other sampling sites, over 75% of the lobsters landed were below theminimum legal size of 65mm carapace length (Table 8). Pooling all sampling periods, there was nosignificant difference in the mean carapace lengths of rock lobsters between reference sites, recently minedsites and sites mined >5 years ago (ANOVA, p ¼ 0:07).
DISCUSSION
The fishery for the rock lobster Jasus lalandii forms an important part of the coastal economy of L .uuderitz.Rock lobsters are the principal invertebrate predators on subtidal reefs in the highly productive inshorekelp beds that characterize the west coast of southern Africa (Barkai and Branch, 1988; Mayfield andBranch, 2000; Mayfield et al., 2000a, b). In the 1990s Namibian rock lobster catches declined dramaticallydue to heavy fishing pressure and, perhaps, changes in environmental conditions (Pollock and Shannon,
Table 7. Sex ratios of rock lobsters caught at reference sites, recently mined sites and sites mined more than 5 years ago
Males Females
Reference sites 0.61 0.39Sites mined 2–5 years ago 0.61 0.39Sites mined >5 years ago 0.57 0.43
Table 8. Mean carapace length (mm) of rock lobsters captured by divers at reference sites, recently mined sites and sites mined >5years ago
Mined 2–5 years ago Mined >5 years ago Reference sites
East Whale Wolf Bay Reef Bay Caravan Nursery Bay Atlas Bay North West Whale Abenteuer Bay
Jan. 1995 51.82 43.77 58.86 } } 47.20 65.51 45.56Sep. 1995 49.94 41.93 48.43 46.90 36.19 55.00 64.54 40.10May 1996 52.48 49.10 52.27 49.69 35.70 41.58 59.34 45.92Sep. 1996 } 43.31 48.74 44.71 } 44.37 58.97 41.82Aug. 1997 50.64 41.87 47.60 } } 47.23 67.77 42.94Apr. 1999 } 43.49* 45.02y } 41.13y 46.15 50.49 41.50
yMining re-commenced in early 1999. * First mined in November 1993.
Table 6. Results of ANOVAs comparing rock lobster abundance at unmined reference sites, sites mined55 years ago and sites mined>5 years ago (df=2 in each case)
Sampling period p ¼ F
January 1995 0.367 1.049September 1995 0.675 0.401May 1996 0.156 1.981September 1996 50.001 13.368August 1997 0.296 1.243April 1998 50.001 47.111
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 247
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

1987; Grobler, 1994). In parallel, marine diamond mining activities have increased along the southernNamibian coastline, leading to perceptions that mining operations had a negative impact on the marineenvironment in general, and on the lobster resource in particular.
Rocky reefs in the region have been poorly studied, with only two descriptive studies (Penrith andKensley, 1970a, b), a biogeographic analysis of algae (Engledow and Bolton, 1992), and a singlequantitative analysis of intertidal biomass (Bustamante and Branch, 1996). The shore-based, diver-operated diamond mining that occurs there is internationally unique (Garnett, 1994), but scientificliterature on the response of marine communities to this mining is sparse. As a consequence, NamdebDiamond Corporation (Pty) Ltd launched the coastal biological monitoring programme described in thispaper, both to help resolve conflicts between the rock lobster and diamond-mining industries, and tobroaden knowledge about Namibian intertidal and subtidal reefs.
Intertidal benthos
The impacts of shore-based mining units described caused an immediate decline in species diversity andabundance in the vicinity of mining activity (Figure 3). However, with the subsequent proliferation offoliose algae at the impacted site after the initial disturbance, the differences in species richness betweenmined and reference sites became statistically indistinguishable in less than a year (Figure 3(A)). Similarshort-term declines in diversity and subsequent proliferations of algae have been reported for intertidalcommunities subjected to natural disturbances such as sand burial, extended aerial exposure and fresh-water floods (Littler and Littler, 1981; Seapy and Littler, 1982; Littler et al., 1983; Branch et al., 1990; VanTamelen, 1996; Airoldi and Virgilio, 1998). Dethier (1984) found that localized disturbances andsubsequent recovery maintains mosaic patterns of diversity, and McQuaid and Dower (1990) documentedsimilar effects of sanding on rocky shores. Sousa (1979a, b) and Van Tamelen (1996) showed that rockmovement may, in fact, increase diversity by the establishment of mixed patches of communities at differentstages of succession.
During mining, and shortly after it ceases, fast-growing, opportunistic, intertidal foliose algae proliferate,associated with a decline in grazers and decreased cover of filter-feeders (Figures 3(B) and (C)). Aftermining ceases, increases in grazer abundance are the next successional stage, and algae begin to diminish. Ina parallel study investigating the impact of discharged fine terrestrial sediments from a mining plant intoElizabeth Bay (Pulfrich et al., 2003), similar declines in grazer densities and concurrent increases in algalcover in affected intertidal and subtidal rocky habitats were reported. Reduced grazer densities withconcomitant dominance of foliose algae typify disturbed rocky shore communities (Littler and Murray,1975; Littler et al., 1983; Branch et al., 1990). In a study addressing the impacts of shore-based, diver-operated mining on the South African Namaqualand coast, Barkai and Bergh (1992) reported that thedensities of several functional groups, particularly coralline algae and filter-feeders, were generally lower atmined sites. However, none of these differences was statistically significant.
Our study clearly indicates several negative effects of contractor-mining on intertidal rocky shores, butalso shows that the intertidal biota appears to recover relatively quickly once mining ceases, becomingstatistically indistinguishable from that of unmined reference sites within 24 months. In general, differenceswere detected between mined and reference sites when univariate analyses were employed (Figure 3), butmultivariate ordination failed to separate mined and reference samples distinctively because of substantialbetween-site variability (Figure 2).
Subtidal benthos
The natural heterogeneity of rocky-reef communities tended to mask any effects of mining. A conclusiveassessment of the actual impacts of mining was thus difficult. The temporal and spatial differences observed
A. PULFRICH ET AL.248
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

amongst the impacted sites, and between the impacted and reference sites are, however, likely to reflect thedevelopment of successional communities in differing stages of recovery at the mined sites.
At the sites mined 2–5 years before sampling, the benthos appeared to have recovered from miningimpacts. Only the community composition of the rubble piles of boulders left by divers remained differentfrom that of reference sites and, indeed, different from the rest of the samples in the mined areas. However,where mining was renewed during the study, there were immediate decreases in algal cover and reductionsin filter-feeders (particularly the polychaete Gunnarea capensis).
The lower cover of kelp recorded at sites that were currently or recently mined supports the findings ofBarkai and Bergh (1992), who established that sites recovering from mining tended to be dominated byyoung kelp plants, whereas the densities of mature kelp were much higher at unmined sites. Kelp beds areutilized extensively by juveniles of many benthic species (Velimirov et al., 1977; Velimirov and Griffiths,1979), and their holdfasts act as nurseries that protect juveniles and sporelings against grazers (Andersonet al., 1997). Along the southern Namibian coastline these shallow-water habitats are thought to serve asimportant rock lobster nursery areas for offshore fishing reefs (Tomalin, 1993). The kelp holdfasts providepreferred refuges for juvenile lobsters (CSIR, 1997) and those of cut plants rot and are broken free by waveaction. Furthermore, the puerulus larvae of J. lalandii settle preferentially in inshore kelp-bed habitats(Tomalin, 1993).
Kelp cutting modifies the habitat because it results in the penetration of greater wave energy in theshallow sublittoral, and modifies light levels (Borchers and Field, 1981). The extent and duration of theimpact of the cutting of Laminaria pallida in Namaqualand and Namibia have not been quantified. Reportsby Simons and Jarman (1981) and Levitt et al. (2002) on harvesting of Ecklonia maxima in the WesternCape (South Africa) suggest that the biomass, plant density and benthos of kelp beds can recover to theiroriginal condition within 2 years. Although cutting kelp causes a localized impact, its severity and durationdepend on the extent and frequency of cutting and the age of the kelp plants cut. Kelp sporelings settle mostsuccessfully at or near the base of adult holdfasts (Velimirov and Griffiths, 1979; Anderson et al., 1997;Levitt et al., 2002), and recovery occurs from the fringe of the cut area. A clear-cut area, or a repeatedly cutarea, will recover more slowly. The loss of shelter caused by divers cutting kelp along the pumping hose andat the pumping site could thus have significant negative effects on the associated fauna and understoreyflora, and important implications for the success of rock lobster recruitment.
In sum, no long-term differences in subtidal benthic composition could be detected between referencesites and those mined >5 years ago, but kelp, other algae and filter-feeders were reduced immediately aftermining and took about 2–3 years to recover. Samples taken in the rubble piles left by divers wereparticularly distinctive, having a very low cover (525%).
Few human activities have effects comparable with those described here, but harvesting of rock-drillingbivalves (Lithophaga lithophaga) requires destruction of rock to extract them. The resulting destabilizationand physical damage to the substratum disrupts the entire system, leading to what is described as‘desertification’ (Fanelli et al., 1994).
Rock lobsters
Counts of rock lobsters were generally higher at reference sites than at any of the mined sites, and catchrates lower at sites mined >5 years previously than in reference or recently mined sites (Figure 7). Thedifferences in abundance were not, however, striking, and were significant on only two of the six samplingoccasions. The population structure of rock lobsters was not affected by mining activities, sex ratios andmean sizes at reference and mined sites being indistinguishable (Tables 6 and 7). These results should beinterpreted with caution, considering the high natural variability in rock lobster population structure,abundance and CPUE, particularly between the West Whale and Abenteuer Bay reference sites. Thevariability between sites most probably reflects spatial differences in suitability of rock lobster habitat, food
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 249
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

availability and other environmental influences, rather than the long-term effects of mining activities. Rocklobsters are highly mobile generalist feeders (Barkai and Branch, 1988; Mayfield et al., 2000a, b) capable ofundertaking onshore–offshore migrations in response to changing environmental conditions (Pollock andShannon, 1987; Jury et al., 1995), and are thus unlikely to be negatively affected in the long-term bylocalized mining activities.
Seasonal onshore–offshore migration of rock lobsters in response to fluctuations in dissolved oxygencontent on the Namibian coast is a well known phenomenon (Newman and Pollock, 1971; Beyers, 1979,Pollock and Beyers, 1981; Bailey et al., 1985; Pollock and Shannon, 1987; Tomalin, 1993). Rock lobstersare found in shallow water during summer when oxygen concentrations in deeper water are at or below2ml/litre�1, the critical concentration for lobsters. The lobsters remain in shallow water until May/June,when they begin to move gradually into deeper water. Although the high temporal variability in lobsterabundances recorded by CPUE and transect counts more than likely reflects variability in divingconditions, the possible influence of seasonal migration patterns of rock lobsters cannot be disregarded.
Other studies addressing the impact of contractor-mining on rock lobster resources on the Namaqualandcoast (Barkai and Bergh, 1992) and at Namibian islands (Tomalin, 1995), concluded that due to naturalheterogeneity, differences in rock lobster abundance between impacted and control sites were insignificantand could not be directly attributed to mining activities. In April 1999, we recorded higher populationdensities of rock lobsters at sites then currently being mined (Reef and Nursery Bays) than at sites minedmore than 5 years previously, and the CPUE at those two sites were at that stage higher than at any othertime (Figure 7). If anything, this suggests that the habitat disturbance created by shore-based mining may insome way be beneficial to rock lobsters.
Whereas the recovery of the subtidal benthic communities seems to be rapid, the mechanical disturbancecaused by mining practices remains noticeable for an extended period. A proportion of the displaced rocksand boulders remain unstable, and the effects of kelp cutting remain visible. It has been postulated that theboulder aggregations resulting from the mining process provide additional shelter for rock lobsters (Barkaiand Bergh, 1992, 1996; CSIR, 1997; Pulfrich and Penney, 1999a). However, Barkai and Bergh (1992) andZoutendyk (1994) have surmised that the consequences of removing and destabilizing rocks in intertidaland shallow subtidal habitats may be severe. They argued that during rough seas such unstable habitatsmay crush rock lobsters that have been attracted to the disturbed benthos and newly created shelters.Although Barkai and Bergh (1992) provided quantitative evidence that there are larger numbers of looserocks at mined than at unmined sites, no correlation was found with the number or size of lobstersassociated with these rocks.
With time, however, wave action redistributes the gravel and sand removed during mining, thereby re-stabilizing the boulders and filling in potential rock lobster shelters (CSIR, 1997; Pulfrich and Penney,1999a). This may have the effect of reducing the rock lobster abundance in an area over the longer-term(>5 years), as suggested by our results.
Shore-based, diver-assisted mining undertaken by contractors is limited to relatively protected small baysalong the southern Namibian coastline, and is opportunistic and highly dependent on weather and seaconditions, with diving being limited to approximately 30 days per year. The time required to mine out anarea depends on the type of substrate and the production of the sediment. While some sites aresystematically mined over a longer period of time, others are periodically revisited as wave action removesexcessive sand overburden and exposes the gravel. Barkai and Bergh (1992) estimated that approximately0.7 km2 of the rocky-reef coastline of Namaqualand is affected annually by the �70 shore-units operatingthere. Whilst �30 units were located in the vicinity of L .uuderitz in the early 1990s, the number of shore-units has now declined to less than 15 (Carter et al., 1998; Pulfrich and Penney, 1999b). The area ofcoastline in Namibia impacted by contractor-mining is thus estimated as 50.2 km2 per year.
Although offshore reefs have never been quantified in southern Namibia, the region affected by shore-based contractor-mining is 322 km in length, and 44% of this comprises rocky shores. Assuming this is
A. PULFRICH ET AL.250
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

representative of the amount of shallow subtidal reef available for shore-based diamond mining, andfurther making the conservative assumption that accessible reefs extend 150m offshore, this allows theestimation that there are approximately 21.3 km2 of reef in the depth range within which diamond diversoperate. This calculation is rough, but it is also conservative because it ignores large areas of offshore reefthat are suitable for rock lobsters but cannot be mined by shore-based divers. Even so, it is clear that bycovering50.2 km2 per year, contractor-mining only takes place on a very small proportion of the reefs thatcan support rock lobsters: approximately 1%.
CONCLUSIONS
The impact of shore-based, diver-operated mining activities on the intertidal and subtidal communities insouthern Namibia appears to be of short duration, and no long-term significant differences in communitystructure between mined and reference sites were evident. Barkai and Bergh (1992) likewise concluded thatthe impacts of contractor-mining on the Namaqualand coast were insignificant compared to mechanicaldisturbance caused by wave action, currents and storms in this variable and dynamic environment. Thenatural variability inherent in benthic communities may outweigh mining impacts within a short periodafter cessation of mining, especially if these impacts occur on small spatial and/or temporal scales. This isparticularly applicable to the potential effects of mining on the shallow-water rock lobster resource. Thereis, however, considerable natural heterogeneity, so that it is difficult to make a conclusive assessment of theactual impacts of contractor-mining on rock lobster abundance. All the evidence suggests, however, that ifmining does have effects on rock lobsters, they are limited to a relatively small proportion of the area of thelobster-grounds and are short-lived. The most obvious and long-lived impacts are associated with theaccumulation of rubble piles left when divers concentrate boulders to be able to work adjacent areas.Although small in extent, they are persistent and support biotic communities that are depauperate anddistinctly different from those in reference areas or in other parts of mined areas. The view taken on theeffects of mining depends on one’s perspective. From a biodiversity conservation perspective, mining clearlydoes have effects, and the longer-lasting influences of rubble piles in particular mean that rehabilitation ofthese is desirable. Alternatively, mining procedures can be altered to ensure that rubble piles are notdeveloped. In terms of the effects of mining on rock lobster populations, however, there is no justificationfor the expense involved in rehabilitation or modification of mining procedures.
ACKNOWLEDGEMENTS
This study was funded through the Contractor Fund of NAMDEB Diamond Corporation (Pty) Ltd. The cheerful anddiligent help of the many field assistants and divers from the University of Cape Town is gratefully acknowledged. Themanager and staff of Elizabeth Bay Mine are thanked for their hospitality and assistance, and for accommodating us inthe old Mine Managers House. Thanks also to the Environmental Officers of Mineral Resources and their assistants,for their help with the administrative logistics of trips, and Kathi Noli and Colette Grobler (Namibian Ministry ofFisheries and Marine Resources) for their interest and assistance. We appreciate the improvements that two anonymousreviewers made to the manuscript.
REFERENCES
Airoldi L, Virgilio M. 1998. Responses of turf-forming algae to spatial variations in the deposition of sediments.MarineEcology Progress Series 165: 271–282.
Anderson RJ, Carrick P, Levitt GJ, Share A. 1997. Holdfasts of adult kelp Ecklonia maxima provide refuges fromgrazing for recruitment of juvenile kelps. Marine Ecology Progress Series 159: 265–273.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 251
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

Bailey GW, Beyers CJ De B, Lipschitz SR. 1985. Seasonal variation of oxygen deficiency in waters off southern SouthWest Africa in 1975 and 1976 and its relation to catchability and distribution of the Cape rock lobster Jasus lalandii.South African Journal of Marine Science 3: 197–214.
Barkai A, Bergh MO. 1992. The effect of marine diamond pumping operations on the littoral and shallow sublittoralbenthos along the South African west coast, Namaqualand region, with special attention to possible effects on therock lobster resource: a pilot study. Report to Marine Diamond Mining Association (MDMA), Cape Town, SouthAfrica, 43 pp.
Barkai A, Bergh MO. 1996. Marine diamond mining along the southern Namibian coastline: possible impacts on, andconflicts with, the local rock lobster fishery. Report for the Namibian Ministry of Fisheries and Marine Resources,Windhoek, Namibia, 30 pp.
Barkai A, Branch GM. 1988. Energy requirements for a dense population of rock lobsters Jasus lalandii: novelimportance of unorthodox food sources. Marine Ecology Progress Series 50: 83–96.
Beyers CJ De B. 1979. Stock assessment and some morphometric and biological characteristics of the rock lobster Jasuslalandii on Marshall Rocks, its main commercial fishing area off South West Africa, 1971–1974. InvestigationalReport of the Sea Fisheries Branch, South Africa 117: 1–26.
Borchers P, Field JG. 1981. The effects of kelp shading on phytoplankton production. Botanica Marina 24: 89–91.Branch GM, Griffiths CL. 1988. The Benguela ecosystem. Part V. The coastal zone. Annual Review of Oceanographyand Marine Biology 26: 395–489.
Branch GM, Eekhout S, Bosman AL. 1990. Short-term effects of the 1988 Orange River floods on the intertidal rockyshore communities of the open coast. Transactions of the Royal Society of South Africa 47: 331–354.
Bray JR, Curtis JT. 1957. An ordination of the upland forest communities of Southern Wisconsin. EcologicalMonographs 27: 325–349.
Bustamante RH, Branch GM. 1995. Maintenance of an exceptional intertidal grazer biomass in South Africa: subsidyby subtidal kelps. Ecology 76: 2314–2329.
Bustamante RH, Branch GM. 1996. Large scale patterns and trophic structure of southern African rocky shores: theroles of geographic variation and wave action. Journal of Biogeography 23: 339–351.
Bustamante RH, Branch GM, Eekhout S. 1995. Maintenance of an exceptional intertidal grazer biomass in SouthAfrica: subsidy by subtidal kelps. Ecology 76: 2314–2329.
Carter RA, Lane S, Wickens P. 1998. Towards responsible environmental management for marine diamond mining inSouth Africa. First Regional Workshop on the Benguela Current Large Marine Ecosystem (BCLME), Cape Town,South Africa, 22–24 July 1998, 9 pp.
Clarke KR. 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal ofEcology 18: 117–143.
Clarke KR, Warwick RM. 1994. Change in marine communities } an approach to statistical analysis and interpretation.Natural Environment Research Council, UK 53 pp.
CSIR. 1997. Environmental management programme report for marine diamond mining in the Namibian Islandconcession. Council for Scientific and Industrial Research Report ENV/S-C 97103. 142 pp.
Dethier M. 1984. Disturbance and recovery in intertidal pools: maintenance of mosaic patterns. Ecological Monographs54: 99–118.
Emanuel BP, Bustamante RH, Branch GM, Eekhout S, Odendaal FJ. 1992. A zoogeographic and functional approachto the selection of marine reserves on the west coast of Africa. South African Journal of Marine Science 12: 341–368.
Engledow H, Bolton JJ. 1992. The biogeography of the seaweed flora of Namibia. In Proceedings of the FirstInternational Workshop on Sustainable Seaweed Resource Development in Sub-Saharan Africa, Windhoek, Namibia,22–29 March 1992, Mshingeni KE, Bolton JJ, Critchley A, Kiangi G. (eds), pp 117–130.
Fanelli G, Piraino F, Belmonte G, Geraci S, Boero F. 1994. Human predation along Apulian rocky coasts (S. E. Italy):desertification caused by Lithophaga lithophaga (Mollusca) fisheries. Marine Ecology Progress Series 110: 1–8.
Garnett RHT. 1994. Marine mining } this is reality. Atlantic Geology 30: 155.Green RH. 1979. Sampling Design and Statistical Methods for Environmental Scientists. Wiley: Chichester, 257 pp.Green RH. 1993. Applications of repeated measures designs in environmental impact and monitoring studies.Australian Journal of Ecology 18: 81–98.
Grobler CAF. 1994. Strict control after the collapse of rock lobster resource. Namibia Brief 18: 93.Jury SH, Howell WH, Watson III WH. 1995. Lobster movement in response to a hurricane. Marine Ecology ProgressSeries 119: 305–310.
Levitt GJ, Anderson RJ, Boothroyd CJT, Kemp FA. 2002. The effects of kelp harvesting on kelp biomass, density andrecruitment and understory community structure at Danger Point (Gansbaai), South Africa. South African Journal ofMarine Science 24: (71–85).
A. PULFRICH ET AL.252
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

Littler MM, Littler DS. 1981. Intertidal macrophyte communities from Pacific Baja California and the upper Gulf ofCalifornia: relatively constant vs. environmentally fluctuating systems. Marine Ecology Progress Series 4: 145–158.
Littler MM, Murray SN. 1975. Impact of sewage on the distribution, abundance and community structure of rockyintertidal macro-organisms. Marine Biology 30: 277–291.
Littler MM, Martz DR, Littler DS. 1983. Effects of recurrent sand deposition on rocky intertidal organisms:importance of substrate heterogeneity in a fluctuating environment. Marine Ecology Progress Series 11: 129–139.
Mayfield S, Branch GM. 2000. Inter-relations among rock lobsters, sea urchins and abalone: implications forcommunity management. Canadian Journal of Fisheries and Aquatic Science 57: 2175–2185,
Mayfield S, Branch GM, Cockcroft AC. 2000a. Relationships among diet, growth rate and food availability for theSouth African rock lobster, Jasus lalandii. Crustaceana 73: 815–834.
Mayfield S, Atkinson LA, Branch GM, Cockcroft AC. 2000b. Variation in the diet of the West Coast Rock Lobster,Jasus lalandii: influence of rock lobster size, sex, capture depth, latitude and moult stage. South African Journal ofMarine Science 22: 57–69.
McQuaid CD, Dower KM. 1990. Enhancement of habitat heterogeneity and species richness on rocky shores inundatedby sand. Oecologia (Berlin) 84: 142–144.
Newman GG, Pollock DE. 1971. Biology and migration of rock lobster Jasus lalandii and their effect on availability atElands Bay, South Africa. Investigational Report of the Sea Fisheries Branch, South Africa 94: 1–24.
Penrith M-L, Kensley BF. 1970a. The constitution of the intertidal fauna of rocky shores of South West Africa. Part I.L .uuderitzbucht. Cimbebasia Series A 1: 191–239.
Penrith M-L, Kensley BF. 1970b. The constitution of the intertidal fauna of rocky shores of South West Africa. Part II.Rocky Point. Cimbebasia Series A 1: 243–267.
Pollock DE, Beyers CJ De B. 1981. Environment, distribution and growth rates of West Coast rock lobster Jasuslalandii (H. Milne Edwards). Transactions of the Royal Society of South Africa 44: 379–400.
Pollock DE, Shannon LV. 1987. Response of rock lobster populations in the Benguela ecosystem to environmentalchange } a hypothesis. South African Journal of Marine Science 5: 887–899.
Pulfrich A, Penney AJ. 1999a. Assessment of the impact of diver-operated nearshore diamond mining on marinebenthic communities near L .uuderitz, Namibia. Final Report to NAMDEB Diamond Corporation (Pty) Ltd.,Oranjemund, Namibia, 40 pp.
Pulfrich A, Penney AJ. 1999b. Interactions between the rock lobster fishery and marine diamond mining along thesouthern African West Coast: a decade in review. GOPA-Cofad Short Term Consultancy Report, AdvisoryAssistance to the Ministry of Fisheries and Marine Resources, Republic of Namibia, 98 pp.
Pulfrich A, Parkins CA, Branch GM, Bustamante RH, Vel!aasquez CR. 2003. The effects of sediment deposits fromNamibian diamond mines on intertidal and subtidal reefs and rock lobster populations. Aquatic Conservation Marineand Freshwater Ecosystems 13: 257–278.
Schneider GIC. 1998. Diamond mining off the coast of Namibia and the marine environment. First RegionalWorkshop on the Benguela Current Large Marine Ecosystem (BCLME), Cape Town, South Africa, 22–24 July 1998,9 pp.
Seapy RR, Littler MM. 1982. Population and species diversity fluctuations in a rocky intertidal community relative tosevere aerial exposure and sediment burial. Marine Biology 71: 87–96.
Simons RH, Jarman NG. 1981. Subcommercial harvesting of a kelp on a South African shore. In Proceedings of the10th International Seaweed Symposium, G .ooteborg, Sweden, August 1980, Levring T (ed.). Walter de Gruyter, Berlin,pp. 731–736.
Sousa WP. 1979a. Disturbance in marine intertidal boulder fields: the non-equilibrium maintenance of species diversity.Ecology 60: 1225–1239.
Sousa WP. 1979b. Experimental investigations of disturbance and ecological succession in a rocky intertidal algalcommunity. Ecological Monographs 49: 227–254.
Tomalin BJ. 1993. Migrations of spiny rock lobsters, Jasus lalandii, at L .uuderitz: Environmental causes, and effects onthe fishery and benthic ecology. MSc thesis, University of Cape Town, 99 pp.
Tomalin BJ. 1995. Specialist study on impacts of diamond mining on rock lobster populations and the fishery. In CSIR1996. Environmental Impact Assessment for marine diamond mining in the Namibian Islands concession, CSIRReport EMAS-C96023, 33 pp.
Underwood AJ. 1992. Beyond BACI: the detection of environmental impact on populations in the real but variableworld. Journal of Experimental Marine Biology and Ecology 161: 145–178.
Underwood AJ. 1993. The mechanics of spatially replicated sampling programmes to detect environmental impacts in avariable world. Australian Journal of Ecology 18: 99–116.
Underwood AJ. 1994. On beyond BACI: sampling designs that might reliably detect environmental disturbances.Ecological Applications 4: 3–15.
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 253
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

Van Tamelen PG. 1996. Algal zonation in tidepools: experimental evaluation of the roles of physical disturbance,herbivory and competition. Journal of Experimental Marine Biology and Ecology 201: 197–231.
Velimirov B, Field JG, Griffiths CL, Zoutendyk P. 1977. The ecology of kelp bed communities in the Benguelaupwelling system. Helgol .aander Meeresuntersuchungen 30: 495–518.
Velimirov B, Griffiths CL. 1979. Wave induced kelp movement and its importance for community structure. BotanicaMarina 22: 169–172.
Zoutendyk P. 1994. Issues and impacts relating to mining operations affecting inter-tidal, rock and sand shores and theneritic environments. In Alexkor environmental management programme, vol. 2. Specialist Studies Report. Council forScientific and Industrial Research Report EMAS-C94037(2), 18 pp.
APPENDIX
List of species recorded at the intertidal study sites near Elizabeth Bay, Namibia
GRAZERS FOLIOSE ALGAEPolyplacophora PhaeophytaChiton nigrovirescens Chordariopsis capensisGastropoda Laminaria pallidaFissurella mutabilis Leathesia difformisHelcion spp. Splachnidium rugosumLittorina africana knysnaensisOxystele spp. ChlorophytaScutellastra argenvillei Bryopsis flanaganiiScutellastra barbara Cladophora contextaCymbula granatina Cladophora spp.Scutellastra granularis Codium fragileCymbula miniata Enteromorpha intestinalisSiphonaria capensis Ulva spp.OpisthobranchiaOnchidella capensis RhodophytaEchinoidea Aeodes orbitosaParechinus angulosus Aristothamnion collabensAsteroidea Carpoblepharus minimaPatiriella exigua Caulacanthus ustulatus
Ceramium spp.FILTER FEEDERS Centroceras clavulatumBivalvia Champia lumbricalisAulacomya ater Coralline (upright)Mytilus galloprovincialis Delisseria papenfussiGastropoda Ectocarpus sp.Crepidula porcellana Gelidium micropterumDendropoma corallinaceus Gigartina bracteataPolychaeta Gigartina scutellataGunnarea capensis Gigartina polycarpa (=radula)Holothuroidea Gigartina stiriataCucumaria rynhardi Grateloupia doryphoraCirripedia Gymnogongrus sp.Chthamalus dentatus Gymnogongrus glomeratus
A. PULFRICH ET AL.254
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)

Cirripedia (Cont.) Rhodophyta (Cont.)Notomegabalanus algicola Gymnogongrus dilatatusOctomeris angulosa Herposiphonia sp.Porifera Iridaea capensisHymeniacedon sp. Nothogenia erinacea
Nothogenia ovalisPREDATORS Pachymena carnosaActinaria Phyllymenia belangeriBunodactis reynaudi Platysiphonia sp.Actinia sp. Pleonosporium sp.Anthothoe sp. Plocamium cornutumAnthopleura sp. Plocamium rigidumPseudactinia sp. Polysiphonia virgata
Porphyra capensisGastropoda Pterosiphonia cloiophyllaBurnupena cincta Scytosiphon sp.Nucella spp. Schizymenia belangeri
Streplocladia captocadaENCRUSTING ALGAE Suhria vittataDiatoms Thamnophyllis discigeraCoralline (crustose) Tayloriella tenebrosaHildenbrandia lecanellieriiLyngbya sp.Ralfsia verrucosa
DIAMOND DIVING IMPACTS ON BIOLOGICAL COMMUNITIES 255
Copyright # 2002 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 13: 233–255 (2003)
Related Documents











