Regular paper The effect of sub-lethal ALA-PDT on the cytoskeleton and adhesion of cultured human cancer cells Anatoly Uzdensky a,b, * , Elona Kolpakova a , Asta Juzeniene a , Petras Juzenas a , Johan Moan a a Institute for Cancer Research, 0310 Montebello, Oslo, Norway b Rostov State University, 344090 Rostov-on-Don, Russia Received 16 July 2004; received in revised form 9 November 2004; accepted 16 November 2004 Available online 15 December 2004 Abstract 5-Aminolevulinic acid (ALA), a precursor of the endogenous photosensitizer protoporphyrin IX, is used in the photodynamic therapy (PDT) of cancer. Sub-lethal ALA-PDT (1-min irradiation with 370–450 nm blue light, 0.6 mW/cm 2 after 2-h incubation with 1 mM ALA) has been earlier shown to change cell morphology and to inhibit both trypsin-induced detachment of cultured cancer cells from the plastic substrata and cell attachment to the bottom of the plastic well plates. In the present study, we found that such treatment of human adenocarcinoma WiDr cells grown in dense colonies stimulated the formation of actin cortex between cells in the colonies and increased the number of actin stress fibres in some, but not in all, cells. However, ALA-PDT did not change the microtubular cytoskeleton in these cells. A similar treatment of glioblastoma D54Mg cells, which grow separately and communicate by protrusions, caused loss of fibrillar actin structures in growth cones, retraction of protrusions, and surface blebbing in some cells. The application of the cytoskeleton inhibitors cytochalasin D, colchicine or taxol showed that the inhibition of trypsin-induced detachment of photosensitized WiDr cells was related to ALA-PDT-induced changes in actin and microtubular cytoskeleton. Some signal transduction processes are suggested to be involved in ALA-PDT-induced changes in cytoskeleton, cell shape, and adhesion. D 2004 Elsevier B.V. All rights reserved. Keywords: 5-Aminolevulinic acid-photodynamic therapy; Cell adhesion; Actin; Tubulin 1. Introduction The photodynamic effect is based on the destruction of stained cells by highly toxic singlet oxygen that is generated by photoexcited dye upon light exposure in the presence of oxygen. This effect is used in cytology as a tool for selective damage of intracellular organelles [1]. Photodynamic therapy (PDT) is a promising anticancer treatment based on light-induced destruction of photosensitized malignant cells and tissues [2,3]. In the last years 5, aminolevulinic acid (ALA), a biochemical precursor of the potent photo- sensitizer protoporphyrin IX (PpIX), has been successfully used for fluorescent diagnosis and photodynamic treatment of cancer. ALA-derived PpIX accumulates in rapidly proliferating cancer cells, thus providing selective destruc- tion of tumours [2,4,5]. PDT has been found to influence cell adhesion [6,7] and to reduce cancer metastases [8–10]. Cell adhesion is related to cancer metastases [11–13]. Therefore, some interest has been focused on the PDT effect on the adhesive properties of cancer cells. Two aspects of cell adhesion are influenced by PDT: (a) cell attachment to a plastic substratum [6,14,15], or to a substratum coated with extracellular matrix proteins [16,17], or to other cells [6,14]; and (b) trypsin-induced detachment of cultured cells from a substratum [6,7,18,19]. Such enzymatic cell detachment, being a routine cell culture procedure, also characterizes cell 0304-4165/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.bbagen.2004.11.011 Abbreviations: ALA, 5-aminolevulinic acid; CD, cytochalasin D; EDTA, ethylenediamine tetraacetate; FCS, foetal calf serum; PBS, phosphate buffered saline; PDT, photodynamic therapy; PpIX, protopor- phyrin IX; TBS, phosphate buffered saline with milk powder and Triton X-100 * Corresponding author. Institute for Neurocybernetics, Rostov State University, 194/1 Stachky ave., NIINK, Rostov-on-Don, 344090, Russia. Tel.: +7 8632 433577; fax: +7 8632 433588. E-mail address: [email protected] (A. Uzdensky). Biochimica et Biophysica Acta 1722 (2005) 43 – 50 http://www.elsevier.com/locate/bba

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

http://www.elsevier.com/locate/bba
Biochimica et Biophysica A
Regular paper
The effect of sub-lethal ALA-PDT on the cytoskeleton and adhesion of
cultured human cancer cells
Anatoly Uzdenskya,b,*, Elona Kolpakovaa, Asta Juzenienea, Petras Juzenasa, Johan Moana
aInstitute for Cancer Research, 0310 Montebello, Oslo, NorwaybRostov State University, 344090 Rostov-on-Don, Russia
Received 16 July 2004; received in revised form 9 November 2004; accepted 16 November 2004
Available online 15 December 2004
Abstract
5-Aminolevulinic acid (ALA), a precursor of the endogenous photosensitizer protoporphyrin IX, is used in the photodynamic therapy
(PDT) of cancer. Sub-lethal ALA-PDT (1-min irradiation with 370–450 nm blue light, 0.6 mW/cm2 after 2-h incubation with 1 mM ALA)
has been earlier shown to change cell morphology and to inhibit both trypsin-induced detachment of cultured cancer cells from the plastic
substrata and cell attachment to the bottom of the plastic well plates. In the present study, we found that such treatment of human
adenocarcinoma WiDr cells grown in dense colonies stimulated the formation of actin cortex between cells in the colonies and increased the
number of actin stress fibres in some, but not in all, cells. However, ALA-PDT did not change the microtubular cytoskeleton in these cells. A
similar treatment of glioblastoma D54Mg cells, which grow separately and communicate by protrusions, caused loss of fibrillar actin
structures in growth cones, retraction of protrusions, and surface blebbing in some cells. The application of the cytoskeleton inhibitors
cytochalasin D, colchicine or taxol showed that the inhibition of trypsin-induced detachment of photosensitized WiDr cells was related to
ALA-PDT-induced changes in actin and microtubular cytoskeleton. Some signal transduction processes are suggested to be involved in
ALA-PDT-induced changes in cytoskeleton, cell shape, and adhesion.
D 2004 Elsevier B.V. All rights reserved.
Keywords: 5-Aminolevulinic acid-photodynamic therapy; Cell adhesion; Actin; Tubulin
1. Introduction
The photodynamic effect is based on the destruction of
stained cells by highly toxic singlet oxygen that is generated
by photoexcited dye upon light exposure in the presence of
oxygen. This effect is used in cytology as a tool for selective
damage of intracellular organelles [1]. Photodynamic
therapy (PDT) is a promising anticancer treatment based
on light-induced destruction of photosensitized malignant
0304-4165/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.bbagen.2004.11.011
Abbreviations: ALA, 5-aminolevulinic acid; CD, cytochalasin D;
EDTA, ethylenediamine tetraacetate; FCS, foetal calf serum; PBS,
phosphate buffered saline; PDT, photodynamic therapy; PpIX, protopor-
phyrin IX; TBS, phosphate buffered saline with milk powder and Triton
X-100
* Corresponding author. Institute for Neurocybernetics, Rostov State
University, 194/1 Stachky ave., NIINK, Rostov-on-Don, 344090, Russia.
Tel.: +7 8632 433577; fax: +7 8632 433588.
E-mail address: [email protected] (A. Uzdensky).
cells and tissues [2,3]. In the last years 5, aminolevulinic
acid (ALA), a biochemical precursor of the potent photo-
sensitizer protoporphyrin IX (PpIX), has been successfully
used for fluorescent diagnosis and photodynamic treatment
of cancer. ALA-derived PpIX accumulates in rapidly
proliferating cancer cells, thus providing selective destruc-
tion of tumours [2,4,5].
PDT has been found to influence cell adhesion [6,7] and
to reduce cancer metastases [8–10]. Cell adhesion is related
to cancer metastases [11–13]. Therefore, some interest has
been focused on the PDT effect on the adhesive properties
of cancer cells. Two aspects of cell adhesion are influenced
by PDT: (a) cell attachment to a plastic substratum
[6,14,15], or to a substratum coated with extracellular
matrix proteins [16,17], or to other cells [6,14]; and (b)
trypsin-induced detachment of cultured cells from a
substratum [6,7,18,19]. Such enzymatic cell detachment,
being a routine cell culture procedure, also characterizes cell
cta 1722 (2005) 43–50
A. Uzdensky et al. / Biochimica et Biophysica Acta 1722 (2005) 43–5044
adhesion. This process may serve as a model of cancer
metastasis because of the involvement of diverse proteolytic
enzymes in the remodelling of extracellular matrix, tumour
invasion, and the formation of metastases.
Cell adhesion to the substrata is mediated by integrins
and other cell adhesion molecules, which link the
extracellular matrix to the intracellular actin cytoskeleton
[20–22]. As recently shown [15,19], non-lethal ALA-PDT,
which inhibits the attachment of cultured cancer cells to
the plastic substratum, as well as trypsin-induced cell
detachment from the plastic surfaces, redistributes aVh3integrin in cultured cancer cells and impairs cell
morphology.
In order to elucidate further the mechanisms underlying
the effect of sub-lethal ALA-PDT treatment on the adhesive
properties of cultured cells [19], we studied the influence of
ALA-PDT on actin and tubulin cytoskeleton in two human
tumour-derived cell lines: adenocarcinoma WiDr and
glioblastoma D54Mg differing in shape and behaviour.
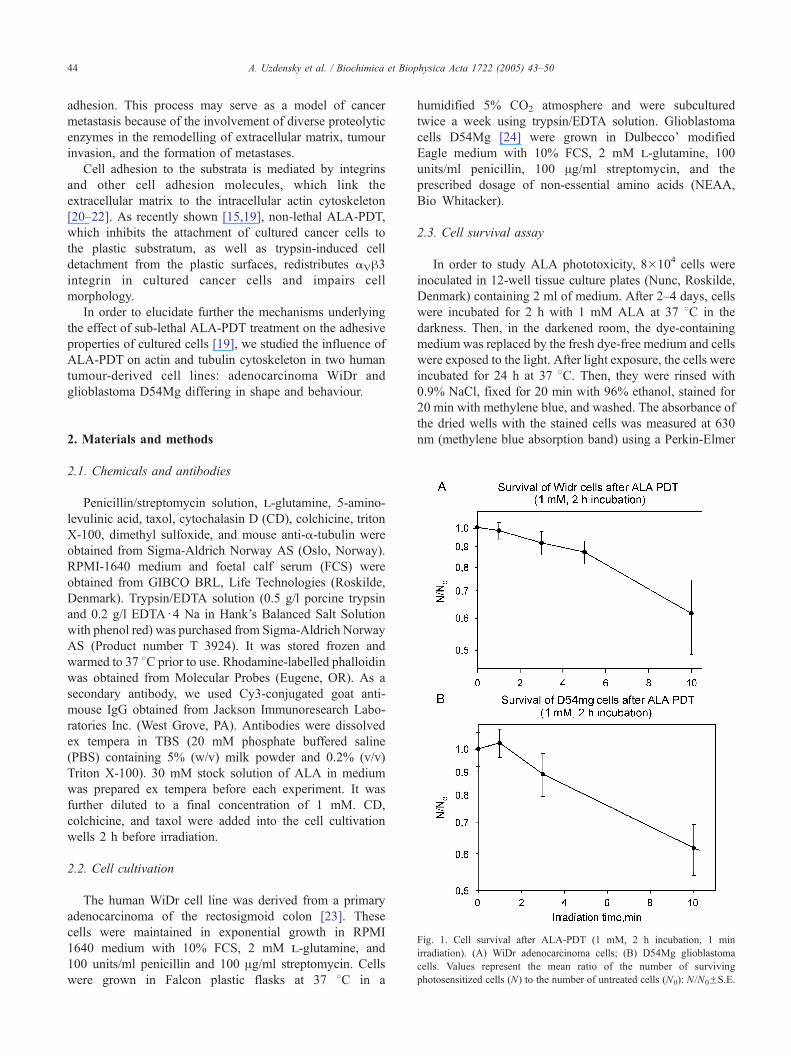
Fig. 1. Cell survival after ALA-PDT (1 mM, 2 h incubation, 1 min
irradiation). (A) WiDr adenocarcinoma cells; (B) D54Mg glioblastoma
cells. Values represent the mean ratio of the number of surviving
photosensitized cells (N) to the number of untreated cells (N0): N/N0FS.E.
2. Materials and methods
2.1. Chemicals and antibodies
Penicillin/streptomycin solution, l-glutamine, 5-amino-
levulinic acid, taxol, cytochalasin D (CD), colchicine, triton
X-100, dimethyl sulfoxide, and mouse anti-a-tubulin were
obtained from Sigma-Aldrich Norway AS (Oslo, Norway).
RPMI-1640 medium and foetal calf serum (FCS) were
obtained from GIBCO BRL, Life Technologies (Roskilde,
Denmark). Trypsin/EDTA solution (0.5 g/l porcine trypsin
and 0.2 g/l EDTAd 4 Na in Hank’s Balanced Salt Solution
with phenol red) was purchased from Sigma-Aldrich Norway
AS (Product number T 3924). It was stored frozen and
warmed to 37 8C prior to use. Rhodamine-labelled phalloidin
was obtained from Molecular Probes (Eugene, OR). As a
secondary antibody, we used Cy3-conjugated goat anti-
mouse IgG obtained from Jackson Immunoresearch Labo-
ratories Inc. (West Grove, PA). Antibodies were dissolved
ex tempera in TBS (20 mM phosphate buffered saline
(PBS) containing 5% (w/v) milk powder and 0.2% (v/v)
Triton X-100). 30 mM stock solution of ALA in medium
was prepared ex tempera before each experiment. It was
further diluted to a final concentration of 1 mM. CD,
colchicine, and taxol were added into the cell cultivation
wells 2 h before irradiation.
2.2. Cell cultivation
The human WiDr cell line was derived from a primary
adenocarcinoma of the rectosigmoid colon [23]. These
cells were maintained in exponential growth in RPMI
1640 medium with 10% FCS, 2 mM l-glutamine, and
100 units/ml penicillin and 100 Ag/ml streptomycin. Cells
were grown in Falcon plastic flasks at 37 8C in a
humidified 5% CO2 atmosphere and were subcultured
twice a week using trypsin/EDTA solution. Glioblastoma
cells D54Mg [24] were grown in Dulbecco’ modified
Eagle medium with 10% FCS, 2 mM l-glutamine, 100
units/ml penicillin, 100 Ag/ml streptomycin, and the
prescribed dosage of non-essential amino acids (NEAA,
Bio Whitacker).
2.3. Cell survival assay
In order to study ALA phototoxicity, 8�104 cells were
inoculated in 12-well tissue culture plates (Nunc, Roskilde,
Denmark) containing 2 ml of medium. After 2–4 days, cells
were incubated for 2 h with 1 mM ALA at 37 8C in the
darkness. Then, in the darkened room, the dye-containing
medium was replaced by the fresh dye-free medium and cells
were exposed to the light. After light exposure, the cells were
incubated for 24 h at 37 8C. Then, they were rinsed with
0.9% NaCl, fixed for 20 min with 96% ethanol, stained for
20 min with methylene blue, and washed. The absorbance of
the dried wells with the stained cells was measured at 630
nm (methylene blue absorption band) using a Perkin-Elmer
A. Uzdensky et al. / Biochimica et Biophysica Acta 1722 (2005) 43–50 45
LS-50 B spectrofluorimeter equipped with a well plate
reader accessory. 3–4 wells were studied for each light
exposure. Relative survival fractions were calculated from:
N=N0 ¼ logIb � logIð Þ= logIb � logIoð Þ;
where I, Io, and Ib are the light intensities passed through
experimental, non-irradiated control, and blank wells,
respectively.
2.4. Cell detachment assay
In the cell detachment experiment, WiDr cells were
subcultured in plastic 12- or 24-well plates (Nunc, Roskilde,
Denmark, or Costar, Corning Inc., Corning, NY) for 3–4
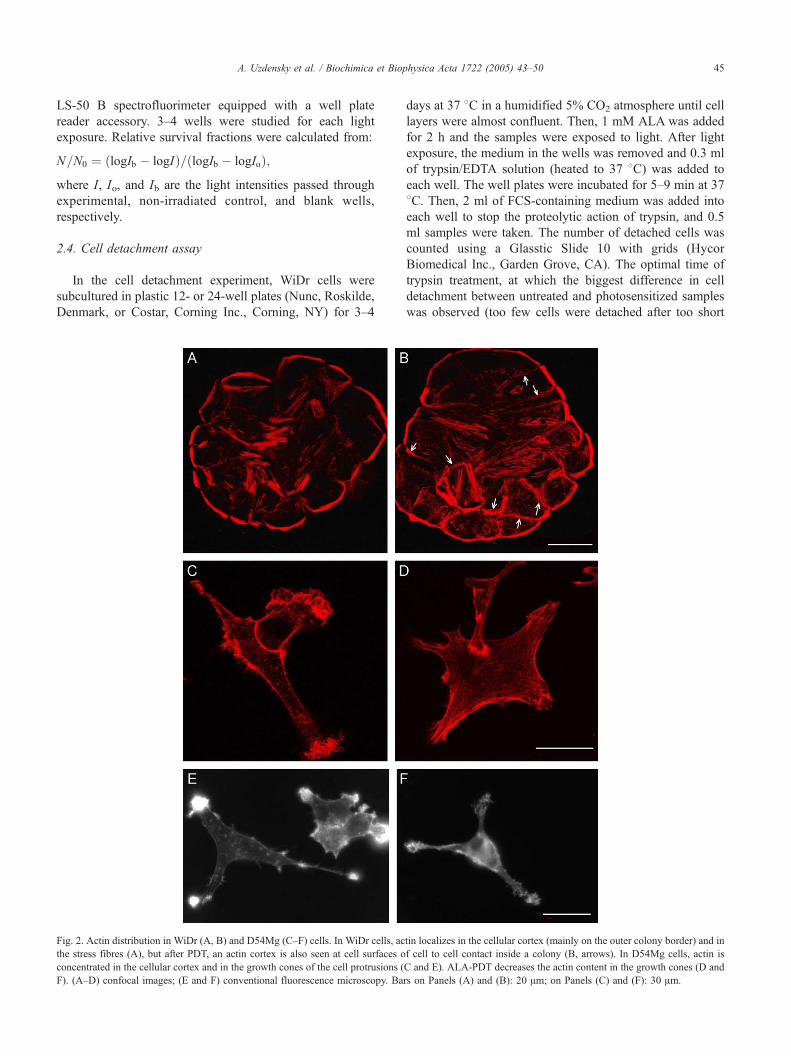
Fig. 2. Actin distribution in WiDr (A, B) and D54Mg (C–F) cells. In WiDr cells, ac
the stress fibres (A), but after PDT, an actin cortex is also seen at cell surfaces o
concentrated in the cellular cortex and in the growth cones of the cell protrusions (
F). (A–D) confocal images; (E and F) conventional fluorescence microscopy. Ba
days at 37 8C in a humidified 5% CO2 atmosphere until cell
layers were almost confluent. Then, 1 mM ALA was added
for 2 h and the samples were exposed to light. After light
exposure, the medium in the wells was removed and 0.3 ml
of trypsin/EDTA solution (heated to 37 8C) was added to
each well. The well plates were incubated for 5–9 min at 37
8C. Then, 2 ml of FCS-containing medium was added into
each well to stop the proteolytic action of trypsin, and 0.5
ml samples were taken. The number of detached cells was
counted using a Glasstic Slide 10 with grids (Hycor
Biomedical Inc., Garden Grove, CA). The optimal time of
trypsin treatment, at which the biggest difference in cell
detachment between untreated and photosensitized samples
was observed (too few cells were detached after too short
tin localizes in the cellular cortex (mainly on the outer colony border) and in
f cell to cell contact inside a colony (B, arrows). In D54Mg cells, actin is
C and E). ALA-PDT decreases the actin content in the growth cones (D and
rs on Panels (A) and (B): 20 Am; on Panels (C) and (F): 30 Am.

Fig. 3. Modification of trypsin detachment of ALA photosensitized WiDr
cells with cytochalasin D (A), colchicine (B), or taxol (C). The
trypsinization efficiency N/N0 (see the legend for Fig. 1) is plotted. ALA
concentration: 1 mM; time of cells incubation with ALA: 2 h. Values
represent the mean N/N0FS.E.
A. Uzdensky et al. / Biochimica et Biophysica Acta 1722 (2005) 43–5046
trypsin exposure but too long exposure caused detachment
of all cells and no difference was observed), varied between
the experiments. It depended on the experimental condi-
tions: temperature, duration of experimental manipulations
leading to cooling of samples, etc. In order to reduce such
variations, the effect of photosensitization was quantified as
the ratio N/N0, where N and N0 are the numbers of the
detached cells in the photosensitized and control samples,
respectively. The duration of trypsinization was determined
visually in each experiment until the detachment of the
majority of control cells.
2.5. Light exposure
Cells incubated with ALA in well plates were exposed to
light from a bank of fluorescent tubes (Model 3026, Applied
Photophysics, London, UK) with an irradiance of 0.7 mW/
cm2 at the position of the cells. The emission of this lamp
was mainly in the wavelength region 370–450 nm, with a
peak at 405 nm, which is close to the maximum of the Soret
band of PpIX in cells.
2.6. Immunofluorescence
For the microscopic study, approximately 104 cells were
seeded into plastic 6-well plates (Nunc, Roskilde, Denmark,
or Costar, Corning Inc., Corning, NY), in which 2–3 round
glass cover slips (10 mm diameter) were placed and allowed
to grow for 2–3 days before photosensitization and staining.
Then, the samples were incubated with 1 mM ALA
dissolved in FCS-containing medium for 2 h. They were
exposed to light for 1 min as described above and fixed for
20 min in 3% paraformaldehyde at room temperature. The
treated cells were washed with PBS, incubated with PBS
containing 30 mM NH4Cl for 10 min and again washed
twice for 10 min in PBS. For actin visualization, the cells
were stained for 20 min with rhodamine-labelled phalloidin
at room temperature. For the visualization of microtubules,
the cells were labelled for 20 min with anti-a-tubulin
dissolved in TBS as described above, washed twice with
PBS for 10 min, and then labelled for 20 min at a room
temperature with secondary goat anti-mouse antibodies
diluted in TBS.
After staining, the cover slips were mounted in Mowiol
(Calbiochem, San Diego, CA). Fluorescence microscopy
was performed using the Zeiss Axioplan microscope (Karl
Zeiss, FRG) equipped with epifluorescence and an oil
immersion objective of 63�. A HBO/100 W mercury lamp
was used for fluorescence excitation. Fluorescence was
observed with an excitation filter 450–490 nm, a dichroic
beam splitter FT 510, and a long pass emission filter N630
nm. Fluorescence images were acquired by a CCD-camera
(Princeton Instruments, Princeton, USA) driven by the
software AnalySis (Soft imaging system GmbH, Munster,
Germany). A Leica TCSNT (Wetzlar, Germany) microscope
was used for confocal microscopy. Images were acquired
with 63� (for glioblastoma cells) or 100� (for adenocarci-
noma cells) objectives and captured with the resolution of
1024�1024 pixels. Montages of images were prepared by
A. Uzdensky et al. / Biochimica et Biophysica Acta 1722 (2005) 43–50 47
using the Adobe PhotoShop 4.0 software (Adobe, Mountain
View, CA).
2.7. Statistics
Standard statistical methods based on Student’s criterion
were used. Results are expressed as meanFS.E.
3. Results
3.1. Phototoxicity of ALA-PDT
The survival curves for the photosensitization of
adenocarcinoma WiDr cells and glioblastoma D54Mg cells
incubated for 2 h with 1 mM ALA are shown in Fig. 1. The
fluences giving 90% survival were about 150 and 110 mJ/
cm2 (light exposures 3.7 and 2.7 min, respectively). 1-min
light exposure killed less than 3% of cells.
3.2. ALA-PDT effect on the intracellular actin cytoskeleton
In tightly packed colonies of untreated WiDr cells,
intracellular actin is manifested as stress fibres and cortex
filaments localized at the cellular periphery (Fig. 2A). In a
colony, the actin cortex was localized mainly at the colony
borders. Almost no cortex-specific staining was observed
inside colonies in the proximity of intercellular contacts
(Fig. 2A). 5–10 min after ALA-PDT (1 min light exposure),
the actin cortex appeared in intercellular regions (Fig. 2B).
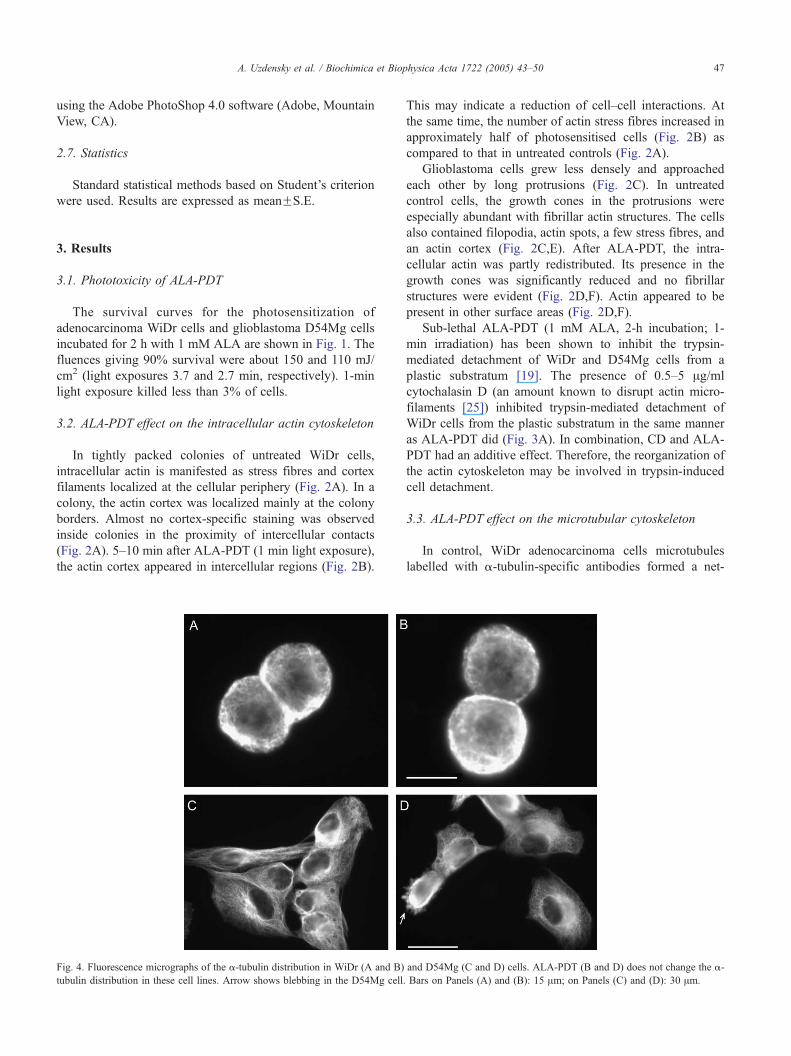
Fig. 4. Fluorescence micrographs of the a-tubulin distribution in WiDr (A and B)
tubulin distribution in these cell lines. Arrow shows blebbing in the D54Mg cell.
This may indicate a reduction of cell–cell interactions. At
the same time, the number of actin stress fibres increased in
approximately half of photosensitised cells (Fig. 2B) as
compared to that in untreated controls (Fig. 2A).
Glioblastoma cells grew less densely and approached
each other by long protrusions (Fig. 2C). In untreated
control cells, the growth cones in the protrusions were
especially abundant with fibrillar actin structures. The cells
also contained filopodia, actin spots, a few stress fibres, and
an actin cortex (Fig. 2C,E). After ALA-PDT, the intra-
cellular actin was partly redistributed. Its presence in the
growth cones was significantly reduced and no fibrillar
structures were evident (Fig. 2D,F). Actin appeared to be
present in other surface areas (Fig. 2D,F).
Sub-lethal ALA-PDT (1 mM ALA, 2-h incubation; 1-
min irradiation) has been shown to inhibit the trypsin-
mediated detachment of WiDr and D54Mg cells from a
plastic substratum [19]. The presence of 0.5–5 Ag/ml
cytochalasin D (an amount known to disrupt actin micro-
filaments [25]) inhibited trypsin-mediated detachment of
WiDr cells from the plastic substratum in the same manner
as ALA-PDT did (Fig. 3A). In combination, CD and ALA-
PDT had an additive effect. Therefore, the reorganization of
the actin cytoskeleton may be involved in trypsin-induced
cell detachment.
3.3. ALA-PDT effect on the microtubular cytoskeleton
In control, WiDr adenocarcinoma cells microtubules
labelled with a-tubulin-specific antibodies formed a net-
and D54Mg (C and D) cells. ALA-PDT (B and D) does not change the a-
Bars on Panels (A) and (B): 15 Am; on Panels (C) and (D): 30 Am.

A. Uzdensky et al. / Biochimica et Biophysica Acta 1722 (2005) 43–5048
work around the nucleus (Fig. 4A). ALA-PDT did not
induce any changes in this network (Fig. 4B).
In the flattened D54Mg glioblastoma cells, the micro-
tubule network was well developed (Fig. 4C). After ALA-
PDT, some cells retracted their lamellipodia and formed
extracellular blebs containing tubulin (Fig. 4D). In these
cells, the microtubular cytoskeleton was, to some extent,
disaggregated so that the tubulin was concentrated in the
perinuclear region (Fig. 4D). Such cells seemed to be
loosely attached to the plastic substratum.
Colchicine is known to depolymerise microtubules [26].
In our experiments, 1 AM colchicine did not significantly
influence the trypsinization efficiency by itself. However, 1–
5 AM colchicine increased the effect of ALA-PDT on
trypsin detachment of WiDr cells (Fig. 3B) independently of
concentration in this range. In contrast to colchicine, taxol
stabilizes microtubules [26]. Together with ALA-PDT, 1
AM taxol decreased the trypsinization efficiency in an
additive manner (Fig. 3C). Therefore, despite the absence of
visible morphological changes in tubulin organization in
WiDr cells, the microtubules may be involved in trypsin-
induced cell detachment.
4. Discussion
Our present and previous data [15,19] show that sub-
lethal ALA-PDT inhibits both trypsin-mediated detachment
of WiDr or D54Mg cells from plastic substrata and
attachment of suspended WiDr cells to plastic. It is known
that cell adhesion and growth depend on the chemical nature
of the substratum, which may vary from one manufacturer
to another [27]. Nevertheless, we did not observe any
noticeable difference between ALA-PDT effect on trypsin-
induced detachment of WiDr cells from Nunc or Costar
well-plates. Therefore, the difference in surface chemistry
was not of significant importance.
The presence of serum in medium is necessary for
adhesion and growth of diverse mammalian cells in culture.
Vitronectin and fibronectin are the major serum proteins
involved in cell adhesion. These proteins adsorbed on the
plastic surface are recognized by the surface receptor
proteins such as integrins [20–22]. The major integrin in
focal contacts is aVh3 [21,28]. It binds specifically to
vitronectin [29]. Redistribution of aVh3 integrin in WiDr
cells after sub-lethal ALA-PDT treatment has been recently
demonstrated [15]. Trypsin-mediated cell detachment is
promoted by the cleavage of extracellular integrin fragments
interacting with the adsorbed serum proteins.
Photodynamic inhibition of trypsin-mediated cell detach-
ment has been suggested to be due to protein cross-linking
caused either by direct photosensitization of the cell
membrane [6], or by the photosensitization of tissue
transglutaminase [18]. We suggest that PDT-induced inhib-
ition of trypsin detachment may be due to mutual integrin
cross-linking within the pre-existing adhesion clusters and
cross-linking of integrins with other proteins in the focal
contacts.
ALA-PDT-induced changes in cell adhesion were
accompanied with the remodelling of the actin cytoskeleton.
Actin fibres are formed after integrin binding to extracellular
ligands. The following integrin clustering leads to the
formation of large actin bundles (stress fibres) at the focal
contacts [21,22]. PDT-induced increase of the number of
stress fibres in WiDr cells may indicate a strengthening of
the cell-substratum contacts. This may be related to photo-
inhibition of cell detachment. We also observed the
formation of the actin cortex at the inner borders of WiDr
cells growing in contact with each other in colonies. This
may indicate the weakening of intercellular contacts and
isolation of cells. Such a pronounced reorganization of the
actin cytoskeleton, including the formation of stress fibres
and remodelling of the actin cortex, may be governed by the
cellular signal transduction system rather than by local
processes triggered by direct PDT targeting of the cell
membrane where integrin-mediated adhesion contacts are
formed. GTPases as Rho and Rac, responsible for the
formation of stress fibres and lamellipodia, respectively, as
well as protein kinase C and phosphatydylinositol 3-kinase,
playing key roles in adhesion signalling [30–33], may
participate in these processes. Their role in photoinduced
cytoskeleton remodelling should be studied in the future.
The microtubules are other cytoskeleton components
responsible for the cell shape. They do not directly link
integrins to actin fibres. Their role in cell adhesion is
assumed to be auxiliary and regulatory [32,33]. For
instance, microtubules growing towards focal adhesions
may deliver the brelaxing factorQ that stimulates the
dissolving of the focal adhesions at the cell periphery
[34]. The disruption of microtubules stimulates the for-
mation of stress fibres and the assembly of adhesion
plaques and triggers the integrin-dependent signalling
cascade [33,35,36]. Microtubules have shown sensitivity
to PDT [37,38]. However, in our experiments, sub-lethal
ALA-PDT did not visibly change the microtubular cyto-
skeleton in WiDr cells. Nevertheless, the experiments with
microtubule modulators indicated their involvement in
ALA-PDT-induced inhibition of trypsin-mediated cell
detachment. It was of interest that both colchicine
(preventing microtubule assembly [26]) and taxol (stabiliz-
ing microtubules [26]) enhanced the ALA-PDT-induced
increase of cell resistance to trypsinization.
Different morphological changes and reorganization of
actin cytoskeleton were observed in ALA-PDT-treated
glioblastoma cells D54Mg. These cells do not form dense
colonies as WiDr cells do and establish intercellular contacts
by cytoplasmic protrusions. After sub-lethal ALA-PDT,
glioblastoma cells lost the fibrillar actin structures in the
growth cones of the protrusions, indicating adhesion
weakening at these sites. Simultaneously, the actin cortex
enlarged in the central cellular regions (Fig. 2D,F). A certain
fraction of the photosensitized D54Mg cells demonstrated

A. Uzdensky et al. / Biochimica et Biophysica Acta 1722 (2005) 43–50 49
surface blebbing and segregation of tubulin-containing
blebs. Some blebs were formed from the retracted protru-
sions in the course of cytoskeleton reorganization and cell
rounding. Blebbing of the cell surface is often accompanied
by the disruption of microtubules [37,38] and the reorgan-
ization of actin microfilaments [39]. Sub-lethal ALA-PDT,
which induced cell blebbing in our experiments, inhibited
trypsin detachment of the D54Mg cells. The hallmarks of
adhesion reduction (loss of fibrillar actin structures in distal
ends of cell protrusions and surface blebbing) and adhesion
strengthening (inhibition of detachment) are not in agree-
ment, and a more detailed study is necessary to resolve this
contradiction.
The observed photodynamic inhibition of cell attachment
and enzymatic detachment are in agreement with the data on
the correlation between attachment rate and organ-coloniz-
ing capacity of cancer cells [40], with the published
observations that PDT decreases the metastatic potential of
surviving cancer cells [10,41], and with reported reduction
of tumour metastases after PDT compared to that after
surgery [8,9]. This gives PDT an advantage compared to
other treatment modalities.
In conclusion, sub-lethal ALA-PDT (1-min blue light
exposure after 2 h cells incubation with 1 mM ALA),
which inhibits the attachment of WiDr cells to plastic and
trypsin-induced cell detachment from plastic substrata,
caused the reorganization of the actin cytoskeleton. The
effects differed in the two cell lines. In WiDr adenocarci-
noma cells, which form densely packed colonies, ALA-
PDT-induced formation of an actin cortex inside the
colonies and increased the number of actin stress fibres in
some cells. The microtubule cytoskeleton appeared to be
unchanged. Similar treatment of glioblastoma D54Mg
cells, which do not form colonies and communicate via
protrusions, caused the loss of fibrillar actin structures in
the growth cones, retraction of protrusions, and surface
blebbing in some, but not in all cells. Cytochalasin D,
colchicine, and taxol influenced ALA-PDT effect on
trypsin-induced detachment of WiDr cells showing the
involvement of actin and microtubule cytoskeleton in this
process. Some signal transduction processes are suggested
to be involved in ALA-PDT-induced changes in cytoske-
leton, cell shape, and adhesion.
Acknowledgements
The present work was supported by the Research
Foundation of the Norwegian Radium Hospital (RF) and
by the Norwegian Cancer Society (DNK).
References
[1] T. Ito, Photodynamic agents as tools for cell biology, Photochem.
Photobiol. Rev. 7 (1983) 141–186.
[2] T. Dougherty, C.J. Gomer, B.W. Henderson, G. Jori, D. Kessel, M.
Korbelik, J. Moan, Q. Peng, Photodynamic therapy, J. Natl. Cancer
Inst. 90 (1998) 889–903.
[3] M. Ochsner, Photophysical and photobiological processes in the
photodynamic therapy of tumours, J. Photochem. Photobiol., B Biol.
39 (1997) 1–18.
[4] Q. Peng, K. Berg, J. Moan, M. Kongshaug, J.M. Nesland, 5-
Aminolevulinic acid-based photodynamic therapy: principles and
experimental research, Photochem. Photobiol. 65 (1997) 235–251.
[5] Q. Peng, T. Warloe, K. Berg, J. Moan, M. Kongshaug, K.-E.
Giercksky, J.M. Nesland, 5-Aminolevulinic acid-based photodynamic
therapy: clinical research and future challenges, Cancer 79 (1998)
2282–2308.
[6] T. Christensen, J. Moan, L. Smedshammer, A. Western, C. Rimington,
Influence of hematoporphyrin derivative (Hpd) and light on the
attachment of cells to the substratum, Photobiochem. Photobiophys.
10 (1985) 53–59.
[7] S.C. Denstman, L.E. Dillehay, J.R. Williams, Enhanced susceptibility
to Hpd-sensitized phototoxicity and correlated resistance to trypsin
detachment in SV40 and IMR-90 cells, Photochem. Photobiol. 43
(1986) 145–147.
[8] C.J. Gomer, A. Ferrario, A.L. Murphee, The effect of localized
photodynamic therapy on the induction of tumour metastasis, Br.
J. Cancer 56 (1987) 27–32.
[9] G. Canti, D. Lattuada, A. Nicolin, P. Taroni, G. Valentini, R. Cubeddu,
Antitumour immunity induced by photodynamic therapy with
aluminium disulfonated phthalocyanines and laser light, Anti-cancer
Drugs 5 (1994) 443–447.
[10] N. Rousset, V. Vonarx, S. Eleouet, J. Carre, E. Kerninon, Y. Lajat, T.
Patrice, Effects of photodynamic therapy on adhesion molecules and
metastasis, J. Photochem. Photobiol., B Biol. 52 (1999) 65–73.
[11] C.W. Evans, Cell adhesion and metastasis, Cell Biol.Int. Rep. 16
(1992) 1–10.
[12] B.R. Zetter, Adhesion molecules in tumour metastasis, Semin. Cancer
Biol. 4 (1993) 219–229.
[13] T. Bogenrieder, M. Herlyn, Axis of evil: molecular mechanisms of
cancer metastasis, Oncogene 22 (2003) 6524–6536.
[14] M.T. Foultier, V. Vonarx-Coinsmann, S. Cordel, A. Combre, T.
Patrice, Modulation of colonic cancer cell adhesiveness by hema-
toporphyrin derivative photodynamic therapy, J. Photochem. Photo-
biol., B. Biol. 23 (1994) 9–17.
[15] A.B. Uzdensky, A. Juzeniene, E. Kolpakova, G.-O. Hjortland, P.
Juzenas, J. Moan, Photosensitization with protoporphyrin IX inhibits
attachment of cancer cells to a substratum, Biochem. Biophys. Res.
Commun. 322 (2004) 452–457.
[16] P. Margaron, R. Sorrenti, J.G. Levy, Photodynamic therapy inhibits
cell adhesion without altering integrin expression, Biochim. Biophys.
Acta 1359 (1997) 200–210.
[17] J.M. Runnels, N. Chen, B. Ortel, D. Kato, T. Hasan, BPD-MA-
mediated photosensitization in vitro and in vivo: cellular adhesion and
h1 integrin expression in ovarian cancer cells, Br. J. Cancer 80 (1999)
946–953.
[18] D.J. Ball, S. Mayhew, D.L. Vernon, M. Griffin, S.B. Brown,
Decreased efficiency of trypsinization of cells following photo-
dynamic therapy: evaluation of a role for tissue transglutaminase,
Photochem. Photobiol. 73 (2001) 47–53.
[19] A.B. Uzdensky, A. Juzeniene, L.-W. Ma, J. Moan, Photodynamic
inhibition of enzymatic detachment of human cancer cells from a
substratum, Biochim. Biophys. Acta 1670 (2004) 1–11.
[20] B.M. Gumbiner, Cell adhesion: the molecular basis of tissue
architecture and morphogenesis, Cell 84 (1996) 345–357.
[21] E. Zamir, B. Geiger, Molecular complexity dynamics of cell–matrix
adhesions, J. Cell. Sci. 114 (2001) 3583–3590.
[22] F.M. Watt, Role of integrins in regulating epidermis adhesion, growth
and differentiation, EMBO J. 15 (2002) 3919–3926.
[23] P. Noguchi, R. Wallace, J. Johnson, E.M. Earley, S. O’Brien, S.
Ferrone, M.A. Pellegrino, J. Milstein, C. Needy, W. Browne, J.

A. Uzdensky et al. / Biochimica et Biophysica Acta 1722 (2005) 43–5050
Petricciani, Characterization of WiDr: a human colon adenocarcinoma
cell line, In Vitro 15 (1979) 401–408.
[24] D.D. Bigner, S.H. Bigner, J. Ponten, B. Westermark, M.S. Mahaley, E.
Ruoslahti, H. Herschman, L.F. Eng, C.J. Wikstrand, Heterogeneity of
genotypic and phenotypic characteristics of fifteen permanent cell
lines derived from human glioblastomas, J. Neuropathol. Exp. Neurol.
40 (1981) 201–229.
[25] L.V. Domnina, V.I. Gelfand, O.Y. Ivanova, E.V. Leonova, O.Y.
Pletyushkina, I.M. Vasiliev, I.M. Gelfand, Effect of small doses of
cytochalasins on fibroblasts: preferential changes of active edges and
focal contacts, Proc. Natl. Acad. Sci. U. S. A. 79 (1982) 7754–7757.
[26] L. Wilson, M.A. Jordan, Pharmacological probes of microtubule
function, in: J. Hyams, C. Lloyd (Eds.), Microtubules, John Wiley and
Sons, New York, 1994, pp. 113–149.
[27] W.K. Scholz, Cell adhesion and growth on coated or modified glass or
plastic surfaces. Nunc. Brand Products, Bulletin 13 (2000) 1–12.
[28] D.I. Leavesley, G.D. Ferguson, E.A. Wayner, D.A. Cheresh, Require-
ment of the integrin aVh3 for carcinoma cell spreading or migration
on vitronectin and fibrinogen, J. Cell Biol. 117 (1992) 1101–1107.
[29] E.G. Hayman, M.D. Pierschbacher, S. Suzuki, E. Ruoslahti, Vitro-
nectin—a major cell attachment-promoting protein in fetal bovine
serum, Exp. Cell Res. 160 (1985) 245–258.
[30] F.G. Giancotti, E. Ruoslahti, Integrin signalling, Science 285 (1999)
1028–1032.
[31] C.K. Miranti, J.S. Brugge, Sensing the environment: a historical
perspective on integrin signal transduction, Nat. Cell Biol. 4 (2002)
83–89.
[32] R.L. Juliano, S. Haskill, Signal transduction from the extracellular
matrix, J. Cell Biol. 120 (1993) 577–585.
[33] A. Bershadsky, A. Chausovsky, E. Becker, A. Lyubimova, B. Geiger,
Involvement of microtubules in the control of adhesion-dependent
signal transduction, Curr. Biol. 6 (1996) 1279–1289.
[34] A.F. Palazzo, G.G. Gundersen, Microtubule-actin cross-talk at focal
adhesions. Science’s STKE, (2002) http://www.stke.org/cgi/content/
full/sigtrans;2002/139/pe31pe31.
[35] A. Kadi, V. Pichard, M. Lehmann, C. Briand, D. Braguer, J. Marvaldi,
J.B. Rognoni, J. Luis, Effect of microtubule disruption on cell
adhesion and spreading, Biochem. Biophys. Res. Commun. 246
(1998) 690–695.
[36] G.G. Gundersen, T.A. Cook, Microtubules and signal transduction,
Curr. Opin. Cell Biol. 11 (1999) 81–94.
[37] K. Berg, J. Moan, J.C. Bommer, J.W. Winkelman, Cellular inhibition
of microtubule assembly by photoactivated sulphonated meso-
tetraphenylporphynes, Int. J. Radiat. Biol. 58 (1990) 3101–3105.
[38] A. Villanueva, M. Canete, S. Nonell, J.L. Borrell, J. Teixido, A.
Juarranz, Photodamaging effect of tetraphenylporphycene in a
human adenocarcinoma cell line, Anti-cancer Drug Des. 11 (1996)
89–99.
[39] J. Huot, F. Houle, S. Rousseau, R.G. Deschesnes, G.M. Shah, J.
Laundry, SAPK2/p38-dependent F-actin reorganization regulates
early membrane blebbing during stress induced apoptosis, J. Cell
Biol. 143 (1998) 1361–1373.
[40] G.A. Turner, Surface properties of metastatic cells, Invasion Meta-
stasis 2 (1982) 197–216.
[41] V. Vonarx, M.T. Foultier, L. Xavier de Brito, L. Anasagasti, L. Morlet,
T. Patrice, Photodynamic therapy decreases cancer colonic cell
adhesiveness and metastatic potential, Res. Exp. Med. 195 (1995)
101–116.
Related Documents