The effect of high air and water temperature on juvenile Mytilus edulis in Prince Edward Island, Canada Neil LeBlanc a, * , Thomas Landry b , Henrik Stryhn a , Re ´jean Tremblay c , Mary McNiven a , Jeff Davidson a a Atlantic Veterinary College, Department of Health Management, University of Prince Edward Island, 550 University Avenue, Charlottetown, Prince Edward Island, Canada C1A 4P3 b Department of Fisheries and Oceans, Science Branch, Mollusc Section, Gulf Fisheries Centre, P.O. Box 5030, Moncton, New Brunswick, Canada E1C 9B6 c Institut des sciences de la mer, Universite ´ du Que ´bec a ` Rimouski, 310 alle ´e des Ursulines, Rimouski, QC, Canada G5L 3A1 Received 24 February 2004; received in revised form 8 July 2004; accepted 28 September 2004 Abstract Mussel aquaculture on Prince Edward Island (PEI), Canada, is an important but relatively new industry. Although seed manipulation using hatcheries for mussel culture occurs on the west coast of North America, seed supply on the east coast of Canada, including Prince Edward Island (PEI), is based solely on wild collection. Two techniques for culling seed (b10 mm) were studied in this experiment to look at the effect on productivity, measured as size, growth and survival. The separate effects of air exposure or high water temperature treatments on a sample of Mytilus edulis spat were examined in the lab and in the field. The in vitro treatments resulted in a ~50% mortality from an air exposure of 11 h at 27 8C and ~75% mortality from a 6-h exposure to 33 8C water. Survivors of each treatment (n =1152) were measured, along with controls (n =2304) and randomly placed in compartmentalized (condo) cages. Cages were deployed on a mussel farm in each of three bays on Prince Edward Island, Canada. Size, growth and survival were monitored over a 10-month period. After the initial treatment, survivors of the air exposure treatment were significantly ( p b0.01) larger than the control. Survivors of the high water temperature treatment were smaller than the control ( p b0.01). Results from the field study showed that the treatments had a significant effect on size, growth and survival. These results suggest that relatively simple husbandry practices of weeding out weaker mussel seed can affect productivity. Given the limited availability for lease expansion in PEI bays, new husbandry practices are an important avenue to investigate, in order for the mussel industry to maximize production capability. D 2004 Elsevier B.V. All rights reserved. Keywords: Mytilus edulis ; Cull; Size; Mortality; Temperature; Air exposure 0044-8486/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.aquaculture.2004.09.035 * Corresponding author. Tel.: +1 902 566 0995; fax: +1 902 566 0823. E-mail address: [email protected] (N. LeBlanc). Aquaculture 243 (2005) 185 – 194 www.elsevier.com/locate/aqua-online

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/aqua-online
Aquaculture 243 (
The effect of high air and water temperature on juvenile
Mytilus edulis in Prince Edward Island, Canada
Neil LeBlanca,*, Thomas Landryb, Henrik Stryhna, Rejean Tremblayc,
Mary McNivena, Jeff Davidsona
aAtlantic Veterinary College, Department of Health Management, University of Prince Edward Island, 550 University Avenue,
Charlottetown, Prince Edward Island, Canada C1A 4P3bDepartment of Fisheries and Oceans, Science Branch, Mollusc Section, Gulf Fisheries Centre, P.O. Box 5030, Moncton,
New Brunswick, Canada E1C 9B6cInstitut des sciences de la mer, Universite du Quebec a Rimouski, 310 allee des Ursulines, Rimouski, QC, Canada G5L 3A1
Received 24 February 2004; received in revised form 8 July 2004; accepted 28 September 2004
Abstract
Mussel aquaculture on Prince Edward Island (PEI), Canada, is an important but relatively new industry. Although seed
manipulation using hatcheries for mussel culture occurs on the west coast of North America, seed supply on the east coast of
Canada, including Prince Edward Island (PEI), is based solely on wild collection. Two techniques for culling seed (b10 mm)
were studied in this experiment to look at the effect on productivity, measured as size, growth and survival. The separate effects
of air exposure or high water temperature treatments on a sample of Mytilus edulis spat were examined in the lab and in the
field. The in vitro treatments resulted in a ~50% mortality from an air exposure of 11 h at 27 8C and ~75% mortality from a 6-h
exposure to 33 8C water. Survivors of each treatment (n=1152) were measured, along with controls (n=2304) and randomly
placed in compartmentalized (condo) cages. Cages were deployed on a mussel farm in each of three bays on Prince Edward
Island, Canada. Size, growth and survival were monitored over a 10-month period. After the initial treatment, survivors of the
air exposure treatment were significantly ( pb0.01) larger than the control. Survivors of the high water temperature treatment
were smaller than the control ( pb0.01). Results from the field study showed that the treatments had a significant effect on size,
growth and survival. These results suggest that relatively simple husbandry practices of weeding out weaker mussel seed can
affect productivity. Given the limited availability for lease expansion in PEI bays, new husbandry practices are an important
avenue to investigate, in order for the mussel industry to maximize production capability.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Mytilus edulis; Cull; Size; Mortality; Temperature; Air exposure
0044-8486/$ - s
doi:10.1016/j.aq
* Correspon
E-mail addr
2005) 185–194
ee front matter D 2004 Elsevier B.V. All rights reserved.
uaculture.2004.09.035
ding author. Tel.: +1 902 566 0995; fax: +1 902 566 0823.
ess: [email protected] (N. LeBlanc).

N. LeBlanc et al. / Aquaculture 243 (2005) 185–194186
1. Introduction
Over the past decade, the mussel aquaculture
industry has become an important part of the economy
in Prince Edward Island (PEI), Canada. Seed supply is
a crucial part of this industry, which depends solely on
natural spat collection. Traditionally, it was believed
that the main broodstock contributing to the seed
production in a bay was the wild population. How-
ever, a recent study in Tracadie Bay, PEI, suggested
that the earliest and largest pulse of larvae comes from
the cultured stock (T. Landry, DFO, personal commu-
nication). This study, therefore, suggests that from a
quantitative standpoint, cultured stocks are self-
sufficient. The experience in Japan with the cultured
scallop, Patinopecten yessoensis indicates that cul-
tured stocks of bivalves can indeed be self-sustaining
(Gosling, 2003).
Historically, in some areas, large-scale mass
mortality has been a problem for growers in the
mussel aquaculture industry (Incze et al., 1980; Mallet
et al., 1990; Myrand and Gaudreault, 1995). Studies
suggest that the seed quality or genetics may be a
cause of these mortalities (Dickie et al., 1984; Mallet
et al., 1990; Myrand and Gaudreault, 1995). Seed
source along with other factors such as stocking
density have been studied in areas of large-scale
mussel culture. These studies have found that the seed
used has a significant effect on production (Fuentes et
al., 1992, 1994; Molares and Fuentes, 1995). Culling
practices are commonly used in hatcheries with the
objective of achieving a more productive crop by
selecting stronger individuals to use as stock (Lestor,
1983). Air exposure and high water temperature are
stresses that cultured mussels are often exposed to
during the farming process, particularly in PEI.
During the summer, waters in the shallow bays of
PEI often reach temperatures of 25 8C (Bernard,
1999). These temperatures come close to reaching a
lethal temperature for Mytilus edulis, which has been
reported in Australia to be 28.2 8C (Wallis, 1975). It
should be noted that since publication of the article by
Wallis (1975), Australia has been reported to have
Mytilus galloprovincialis, rather than M. edulis
(Gosling, 1992). Field experiments in Rhode Island
showed that continuous temperatures above 27 8Chave been found to be lethal to M. edulis (Gonzalez
and Yevich, 1976). Read and Cumming (1967) found
that at 27 8C, 50% mortality occurred in about 3 days.
Since temperature stress is a reality on mussel farms in
PEI, a process that removes mussels susceptible to
high water temperature was investigated. The farming
of mussels requires that mussels be exposed to air for
periods of time throughout the harvest cycle.
Although mussels are an intertidal species, they will
eventually die from air exposure, especially at high
temperatures (Tsuchiya, 1983). For this reason,
selecting for mussels that can better cope during
emersion was the other culling technique investigated
in this study.
The objective of this study was to examine whether
the culling practices employed can result in a higher
quality mussel seed for the grow-out phase. The study
examined the productivity measures, size and sur-
vival, to assess the effects the stress treatments had on
mussel seed immediately and over time in the field.
2. Materials and methods
2.1. Specimen manipulation
2.1.1. Sampling
Wild seed was manually collected from aquacul-
ture gear in St. Peter’s Bay, PEI. Approximately
30,000 individual spat were retained. Seed was
transported to a hatchery in Prince Edward Island.
The sample was divided in half and placed into two
upwellers (40 cm diameter) in a 1.2 m3 tank. Mortality
in these upwellers remained low (b3%) throughout the
course of the experiments. Flow rate of unfiltered tidal
river water varied from 0.1 to 0.5 L/s, with an average
temperature of 24.3 8C and average salinity of 28.2
ppt. In keeping with other literature, a minimum level
of mortality of 50% was targeted (Wallis, 1975; Cotter
et al., 1982; Tremblay et al., 1998), for the purpose of
detecting any effect of treatments.
2.1.2. Air exposure
An air exposure trial was conducted with approx-
imately 5000 individuals taken from the upwellers and
placed on a layer of brown paper towel and spread
over a counter top in the lab. The individuals were left
exposed for 11 h at an average temperature of
27.2F0.5 8C and average relative humidity of
55.6F10%. They were then placed in a recovery tank
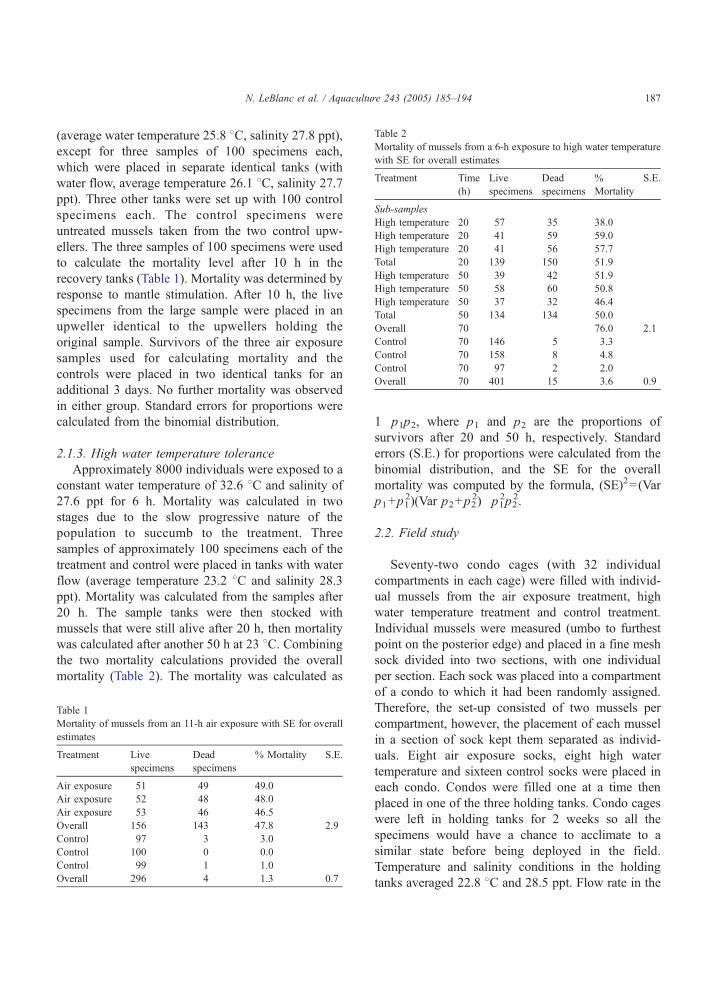
Table 2
Mortality of mussels from a 6-h exposure to high water temperature
with SE for overall estimates
Treatment Time
(h)
Live
specimens
Dead
specimens
%
Mortality
S.E.
Sub-samples
High temperature 20 57 35 38.0
High temperature 20 41 59 59.0
High temperature 20 41 56 57.7
Total 20 139 150 51.9
High temperature 50 39 42 51.9
High temperature 50 58 60 50.8
High temperature 50 37 32 46.4
Total 50 134 134 50.0
Overall 70 76.0 2.1
Control 70 146 5 3.3
Control 70 158 8 4.8
Control 70 97 2 2.0
Overall 70 401 15 3.6 0.9
N. LeBlanc et al. / Aquaculture 243 (2005) 185–194 187
(average water temperature 25.8 8C, salinity 27.8 ppt),except for three samples of 100 specimens each,
which were placed in separate identical tanks (with
water flow, average temperature 26.1 8C, salinity 27.7
ppt). Three other tanks were set up with 100 control
specimens each. The control specimens were
untreated mussels taken from the two control upw-
ellers. The three samples of 100 specimens were used
to calculate the mortality level after 10 h in the
recovery tanks (Table 1). Mortality was determined by
response to mantle stimulation. After 10 h, the live
specimens from the large sample were placed in an
upweller identical to the upwellers holding the
original sample. Survivors of the three air exposure
samples used for calculating mortality and the
controls were placed in two identical tanks for an
additional 3 days. No further mortality was observed
in either group. Standard errors for proportions were
calculated from the binomial distribution.
2.1.3. High water temperature tolerance
Approximately 8000 individuals were exposed to a
constant water temperature of 32.6 8C and salinity of
27.6 ppt for 6 h. Mortality was calculated in two
stages due to the slow progressive nature of the
population to succumb to the treatment. Three
samples of approximately 100 specimens each of the
treatment and control were placed in tanks with water
flow (average temperature 23.2 8C and salinity 28.3
ppt). Mortality was calculated from the samples after
20 h. The sample tanks were then stocked with
mussels that were still alive after 20 h, then mortality
was calculated after another 50 h at 23 8C. Combining
the two mortality calculations provided the overall
mortality (Table 2). The mortality was calculated as
Table 1
Mortality of mussels from an 11-h air exposure with SE for overall
estimates
Treatment Live
specimens
Dead
specimens
% Mortality S.E.
Air exposure 51 49 49.0
Air exposure 52 48 48.0
Air exposure 53 46 46.5
Overall 156 143 47.8 2.9
Control 97 3 3.0
Control 100 0 0.0
Control 99 1 1.0
Overall 296 4 1.3 0.7
1�p1p2, where p1 and p2 are the proportions of
survivors after 20 and 50 h, respectively. Standard
errors (S.E.) for proportions were calculated from the
binomial distribution, and the SE for the overall
mortality was computed by the formula, (SE)2=(Var
p1+p12)(Var p2+p2
2)�p12p2
2.
2.2. Field study
Seventy-two condo cages (with 32 individual
compartments in each cage) were filled with individ-
ual mussels from the air exposure treatment, high
water temperature treatment and control treatment.
Individual mussels were measured (umbo to furthest
point on the posterior edge) and placed in a fine mesh
sock divided into two sections, with one individual
per section. Each sock was placed into a compartment
of a condo to which it had been randomly assigned.
Therefore, the set-up consisted of two mussels per
compartment, however, the placement of each mussel
in a section of sock kept them separated as individ-
uals. Eight air exposure socks, eight high water
temperature and sixteen control socks were placed in
each condo. Condos were filled one at a time then
placed in one of the three holding tanks. Condo cages
were left in holding tanks for 2 weeks so all the
specimens would have a chance to acclimate to a
similar state before being deployed in the field.
Temperature and salinity conditions in the holding
tanks averaged 22.8 8C and 28.5 ppt. Flow rate in the

N. LeBlanc et al. / Aquaculture 243 (2005) 185–194188
holding tanks was approximately 0.2 L/s. Dimensions
of the holding tanks were 2.13 m long�1.22 m
wide�0.91 m deep.
Six cages (containing four condos) were placed
into each of St. Peter’s, New London and Tracadie
Bay (Fig. 1); all three bays are used extensively for
mussel aquaculture. Although the bays are similar in
depth, temperature and salinity (Fisheries and Aqua-
culture Division, 2001; Crane, 2003), three different
field sites were used to detect any interaction between
the treatments and the environment. All were attached
to mussel long lines in the bays at a point where the
depth was 4.6–4.96 m (15–16 ft.). The cages were
sunk to a depth of 1.8–2.1 m (6–7 ft.). Water tem-
perature in all three bays were slightly lower (20.5–
21.5 8C) but similar to the temperature in the
experimental tanks, so no temperature acclimation
was necessary (Fisheries and Aquaculture Division,
2001).
After 4 months, mussels in each of the bays were
measured for length and mortality was counted.
Final measurements and mortality counts were done
at the 10-month period. The experiment was
initiated approximately 6 weeks prior to the normal
seed transfer time on PEI. This was necessary for
practical purposes, such as the availability of
facilities and technical support, including industry
partners.
Fig. 1. Map of Prince Edward Island, Canada, indicating the three bays u
follows: New London Bay (46829.756 N, 63828.185 W), Tracadie Ba
62841.599 W).
2.3. Species determination
A random sample of one hundred and ten mussels
was taken for species characterization as M. edulis or
Mytilus trossulus according to the PCR technique
developed by Rawson et al. (1996).
2.4. Statistical analyses
The effect of the treatments on initial mussel size
was assessed by one-way ANOVA using the length
measurements from the specimens placed in condos
for the field study.
The models used for analyzing the field data used
treatment and bay as fixed factors with condo as a
random factor to account for potential clustering
within condo cages. Mixed ANOVA models were
used for length data (software SAS 8.2; Littell et al.,
1996). Random effects binary logistic regression was
used to analyze mortality (software MLwiN 1.2;
Rasbash et al., 2000). In the analyses, missing
specimens were treated as dead. The mesh socks kept
the mussels separated within in the condo compart-
ments and prevented them from falling out when they
were small enough to fit through spaces in the condo
cage. A small percentage of specimens (b3%) which
showed negative growth were not used, although their
inclusion in the analyses did not affect the signifi-
sed as field sites. Locations of the study sites in the bays were as
y (46823.355 N, 62859.440 W), St. Peter’s Bay (46826.231 N,

N. LeBlanc et al. / Aquaculture 243 (2005) 185–194 189
cance of the results. Shell deposition is permanent,
thus negative growth indicates an error in measuring,
recording, sampling, etc.; therefore, removal of such
data is appropriate. All statistical tests were inter-
preted using a 5% error level, and multiple pairwise
comparisons were carried out by the Bonferroni
method.
Table 3
Adjusted treatment and bay mean mussel sizes (mm; with S.E.),
initially and after 4 and 10 months
Size (mm)
Initial
(n=4608)
Four months
(n=843)
Ten months*
(n=394)
Treatment
Air exposure 8.07(0.05)c 26.55(0.41)b 35.06(0.60)b
High water
temperature
6.49(0.05)a 24.54(0.41)a 32.96(0.63)a
Control 6.81(0.03)b 25.93(0.33)b 31.69(0.56)a
Bay
Tracadie 7.14(0.04)b 24.69(0.37)b 30.91(0.57)a
New London 6.90(0.05)a 25.86(0.52)ab 35.60(0.76)b
St. Peter’s 7.10(0.04)b 26.47(0.42)a 33.20(0.85)ab
Statistical differences within each time period for treatment and bay
are indicated using superscripted letters.a,b,cCategories indicated by the same symbol are not statistically
significant at the 5% level.
* Comparisons for treatment and bay refer to average effects,
due to significant interaction (Fig. 2).
3. Results
3.1. Laboratory culling experiment
3.1.1. Eleven-hour air exposure
The mortality level was 47.8% from a sample size
of 299 specimens (Table 1). The vast majority of the
dead specimens were floating after 10 h in the
recovery tanks. Thirty floating specimens were physi-
cally examined first by exposing to the air to observe
gaping, then gently prying to test valve function and
finally opening the animal to inspect the soft tissue.
None of the mussels that were floating after 10 h in
the recovery tank were found to be alive. Only a few
dead specimens (b10%) were found on the bottom of
the tanks with the live individuals. Mortality in the
control groups was low (Table 1).
3.1.2. Six-hour high water temperature immersion
Mortality determination for the high water temper-
ature was more complicated because death occurred
over a longer period of time compared with the air
exposure (50+ h compared with 10 h) (Table 2). Also,
the vast majority of the dead specimens in the air
exposure treatment floated, facilitating separation, this
was not the case for this treatment. Due to the time
lag, mortality determination for this treatment was
done in two steps, so that dead specimens could be
removed from the upwellers and decomposing mus-
sels did not contribute to mortality.
Mortality from the treatment was 76% while the
mortality in the control groups remained low
(Table 2).
3.1.3. Size of mussels after treatments
Significant differences in mean size were present
between all three treatments (Pb0.01). The stress
treatments had opposite effects with the smaller
mussels surviving the temperature treatment and
larger ones surviving the air exposure (Table 3).
Although selection for mussels placed in each bay
was random, the mussels that went into New London
Bay were significantly smaller than mussels in the
other two bays ( pb0.05). This effect may have arisen
from New London Bay being prepared last. With the
initial sizes being quite small, there may have been an
unconscious tendency to select the larger mussels for
measurement.
3.2. Field study
3.2.1. Size and growth
Analysis was performed to determine the effect of
the treatments on the size of the mussels at each time
period and the growth over that time period. Size was
preferred over growth in the analysis because there
was no significant correlation between the initial size
of the mussels and the size at each time period. In
other words, differential growth of the mussels during
the experiment eliminated any size differences at the
outset.
Data showed a substantial variation among the
condo cages, indicating that the set-up procedure of
randomly placing treatment and control (s) within
each condo was prudent.

Table 4
Mortality of mussels after 10 months in the PEI bays with SE
N % Mortality
Treatment
Air exposure 234 56.8(3.2)
33 8C H2O 228 58.3(3.3)
Control 451 71.8(2.1)
Bay
Tracadie 529 69.4(2.0)
New London 148 55.4(4.1)
St. Peter’s 236 59.7(3.2)
N. LeBlanc et al. / Aquaculture 243 (2005) 185–194190
3.2.2. Size after 4 months
The ordering of the treatments with respect to size
did not change from the initial measurements: The air-
exposed mussels were the largest and the high
temperature exposed mussels the smallest, however,
air-exposed mussels were no longer significantly
larger than control mussels (Table 3).
Mussels in St. Peter’s Bay were significantly larger
than mussels in Tracadie Bay, whereas, mussels in
New London Bay were not different from either of the
other two bays in size (Table 3).
3.2.3. Size after 10 months
Size differences were seen between treatments and
between bays, as well as in a significant interaction
( p=0.03, Fig. 2). The air exposure treated mussels
were significantly larger than the mussels in the high
water temperature treatment and control groups,
averaged across bays (Table 3). Among the bays,
mean mussel size in New London was significantly
larger than mussels in Tracadie Bay, averaged across
treatments ( pb0.05).
Pairwise comparisons for size between the treat-
ments in specific bays showed that air-exposed
mussels in New London Bay were larger than each
of the treatments in Tracadie Bay including controls,
as well as control mussels in New London and St.
Peter’s Bay. Also, the high water temperature treated
mussels in New London Bay were larger than the high
water temperature treated mussels in Tracadie Bay.
Fig. 2. Mean size of mussels after ten months in PEI bays with SE.
Table 5
Random effects logistic regression analysis of mussel mortality after
10 months with respect to bay and treatments
Variable Comparison Estimate S.E. Odds
ratio
p Value
Intercept Reference
(Tracadie, Control)
1.369 0.304
Bay 0.25
St. Peter’s vs.
Tracadie
�0.557 0.535 0.573 0.30
New London
vs. Tracadie
�0.844 0.545 0.430 0.12
Treatment b0.001
Air exposure
vs. Control
�0.762 0.179 0.467 b0.001
High water
temperature
vs. Control
�0.690 0.181 0.502 b0.001
Variance
(condo)
Reference
(Tracadie, Control)
0.724 0.281
3.2.4. Mortality
Mortality was analyzed over the 10-month period
of the experiment (Table 4). Significant differences
among the treatments were found (Table 5). The air
and temperature exposed mussels had significantly
lower mortality than the control mussels at the 5%
level but were not significantly different from each
other.
Although there are larger differences in mortality
among the bays than in the treatments (Table 4), the
random variability of the condo cages prevents any
statistically significant results. Treatments were nested
within the condo cages so the variability within the
condos did not affect the findings for the treatments as
it did for the bays.

N. LeBlanc et al. / Aquaculture 243 (2005) 185–194 191
3.3. Species determination
The Gulf of St. Lawrence is an area where the
distributions of M. edulis and M. trossulus are known
to overlap (Gosling, 1992), however, the level of M.
trossulus in Prince Edward Island has been found to
be quite low (Mallet and Carver, 1995). The results
from the specimens tested in this study found that this
population was 100% M. edulis.
4. Discussion
The laboratory experiment was designed to cull
mussels through selective mortality resulting from
exposure to the stressful conditions of high water
temperature or air exposure. The selective effect of the
treatments was determined by measuring the size and
mortality of treated and control mussels over a 10-
month period in the field. The important factors for
the culling treatments are: The ability to selectively
cull out weaker mussels, the surviving mussels must
be able to fully recover from the treatment, and the
treatments must be feasible on a large scale. By
choosing relatively simple natural factors that affect
mussel populations, all the above criteria were met.
Mytilus is an intertidal species. The most important
factors in determining the upper limits for Mytilus on
rocky intertidal shores are physiological intolerance to
temperature extremes and desiccation (Suchanek,
1985). Given that Mytilus has evolved to be a
successful intertidal species (Lewis, 1997), often to
avoid predation (Ebling et al., 1964; Paine, 1974), it is
more likely that desiccation would not permanently
harm the animals. As for high water temperature, in
this region, water temperatures can reach and even
exceed 25 8C (Bernard, 1999; Myrand and Gau-
dreault, 1995). The hope was that exposure to
temperatures less than 6 8C above lethal temperatures
for a short time period (6 h) would not permanently
harm the mussels that survived the cull. An effective
cull needs to be practical in terms of time to complete
the procedure and it must have a minimal long-term
effect on the productivity of the survivors.
In both cases, the stress treatment did not appear to
permanently harm the surviving mussels, although
growth may have been stunted for some period of
time. With less mortality and comparable, if not
greater, size than the controls, the treated mussels at
the end of the 10-month field experiment appeared to
suffer no permanent damage. However, it still makes
sense to perform a culling treatment when the effect
on mussel growth would be minimal, perhaps in late
fall when growth is already retarded by natural
conditions of declining food and temperature.
The major observation on the immediate effect of
the treatments besides the level of mortality was the
selection for size. Clearly, smaller mussels better
survived the high water temperature treatment and
larger mussels were more resilient to air exposure. A
previous study by Bayne (1984) found that although
small mussels (M. edulis) have a higher maintenance
energy requirement and lose more weight during
starvation, small individuals were at an energetic
advantage in responding to high temperature. The
energetic advantage was calculated using scope for
growth or the energy available for growth and
reproduction. As temperature was increased, smaller
mussels (b0.6 g dry weight) maintained positive scope
for growth while larger mussels (up to 2.0 g dry
weight) did not. The results here indicate that smaller
mussels also had an advantage during acute exposure
to lethal high water temperature.
The survival from the air exposure treatment of the
larger mussels immediately seems to make it a more
favourable cull, as they immediately seemed to have
an advantage in getting to market size. The survival in
air of certain shellfish species, including Mytilus(De
Zwaan, 1977; Livingstone and Bayne, 1977; Widd-
ows and Shick, 1985), has been examined for differ-
ent reasons (Matthews and Mcmahon, 1995; Paukstis
et al., 1999; Bartsch et al., 2000) including long line
culture (Guderley et al., 1994). Survival times in air
for bivalves, including M. edulis, vary greatly depend-
ing on acclimation and various environmental factors
such as temperature and humidity (Tsuchiya, 1983;
Paukstis et al., 1999). These studies on Mytilus
dealing with air exposure and mortality (Tsuchiya,
1983; Eertman et al., 1993; Guderley et al., 1994) did
not mention a physiological advantage larger mussels
had in surviving air exposure. However, this advant-
age was seen in the freshwater bivalve, Dreissena
polymorpha, commonly known as the zebra mussel
(Matthews and Mcmahon, 1995; Paukstis et al.,
1999). Paukstis et al. (1999) stated that larger zebra
mussels (shell length N16 mm) were significantly

N. LeBlanc et al. / Aquaculture 243 (2005) 185–194192
more likely to survive exposure for 36 or 60 h at 20
8C than were smaller individuals.
The field study was set-up to monitor the long-term
effects the treatments had on the mussels’ size and
mortality. Statistically, the treatments had an effect on
size, but there was an interaction between treatments
and the bays. In New London Bay, the air-exposed
mussels had significantly larger size than all other
bay/treatment combinations while in Tracadie Bay, the
high temperature exposed mussels were smaller than
elsewhere. The results suggest that the environment
will in part determine the effect of treatments. The air
exposure treatment had an advantage in size in some
circumstances and never had a negative interaction
with location, suggesting it may be the better
candidate as a culling technique.
There was no correlation between the initial size of
the mussels and the subsequent size measurements.
However measurements from the 4- and 10-month
periods were highly correlated. This raises the
question: Why does the size of the mussel at the start
of the experiment have no bearing on its size later on?
The genotypic factors related to fitness in mussels are
thought to be minor compared with environmental
factors (Seed and Suchanek, 1992). Under heavy
settlement, growth in individuals has been found to
vary by as much as 10-fold (Trevelyan, 1991). It is
hypothesized that the initial sizes were based mainly
on environmental factors (position, crowding, food
availability) and that once placed in similar con-
ditions, genotypic factors played more of a role in
growth.
The treated mussels showed lower mortality than
the controls. This indicates that the mussels that
survived the cull were more fit than those that were
not exposed to any stress treatment. There is an
important issue regarding the nature of the fitness
advantage that needs consideration. Did the differ-
ences seen in the treated mussels result from weaker
specimens being eliminated from the population or
did the treatments condition the survivors to better
withstand environmental stress? Currently, the answer
to this question is unknown. The long-term modifi-
cation of mussel populations to different thermal
regimes has been suggested (Newell and Branch,
1980), and fits the basic premise of evolution. What
about individual modification? It appears there is
some evidence that Mytilus can adjust itself after
experiencing a thermal shock. Buckley et al. (2001)
found that the expression of heat shock proteins
changes in Mytilus based on the previous thermal
history. This study by Buckley puts forward the
possibility that husbandry practices being developed
should investigate conditioning as well as culling as a
potential tool for improving the growth and survival
of farmed mussels.
5. Conclusions
This study found that in mussel seed from St.
Peter’s Bay, PEI:
(1) larger seed survived air exposure and smaller seed
survived exposure to high water temperature.
(2) exposing M. edulis to the air or high water
temperatures affected growth but there was an
interaction between the treatments and the
environment.
(3) mussel samples exposed to the air or high water
temperature had lower mortality than mussels
that had not undergone such treatments.
(4) the experimental husbandry practices examined
in this study affect the productivity of mussels
and this type of work has the potential to benefit
mussel aquaculture; this is especially true in a
place like Prince Edward Island where additional
leasing areas are not available to increase
production.
(5) this study showed an interaction between treat-
ments and the environment therefore further
research on mussel seed husbandry should take
into account as many environmental parameters
as possible.
Acknowledgements
We would like to thank Garth Arsenault for his
technical assistance and the three anonymous
reviewers for their helpful comments. We are also
grateful to Stephen Stewart (Stewart Mussels farms),
Bob Fortune (United Mussel Farms) and Russell
Dockendorff Jr. (PEI Mussel King) for their partic-
ipation in the field study. This work was supported by
the Prince Edward Island Aquaculture Alliance with

N. LeBlanc et al. / Aquaculture 243 (2005) 185–194 193
funds from NRC-IRAP (National Research Council-
Industrial Research Assistance Program), AFRI
(Aquaculture and Fisheries Research Initiative),
PEIAFAF (Prince Edward Island Department of
Agriculture, Fisheries, Aquaculture and Forestry)
and DFO (Department of Fisheries and Oceans).
References
Bartsch, M.R., Waller, D.L., Cope, W.G., Gutreuter, S., 2000.
Emersion and thermal tolerances of three species of unionid
mussels: survival and behavioral effects. J. Shellfish Res. 19,
233–240.
Bayne, B.L., 1984. Responses to environmental stress: tolerance,
resistance and adaptation. Proc. 18th Euro. Mar. Biol. Sym.,
Aug. 1983, pp. 331–349.
T. Bernard, 1999. P.E.I. Mussel Monitoring Program, P.E.I.
D.F.E.F.A. Tech. Rep. Ser. 226.
Buckley, B.A., Owen, M.E., Hofmann, G.E., 2001. Adjusting the
thermostat: the threshold induction temperatures for the heat-
shock response in intertidal mussels (genus Mytilus) changes as
a function of thermal history. J. Exp. Biol. 204, 3571–3579.
Cotter, A.J.R., Phillips, D.J.H., Ahsanullah, M., 1982. The
significance of temperature, salinity and zinc as lethal factors
for the mussel Mytilus edulis in a polluted estuary. Mar. Biol.
68, 135–141.
Crane, C., 2003. Unpublished salinity data (1998–2003) from the
August survey, Prince Edward Island Department of Environ-
ment and Energy, Water Management Division.
De Zwaan, A., 1977. Anaerobic energy metabolism in bivalve
molluscs. Oceanogr. Mar. Biol. Ann. Rev. 15, 103–187.
Dickie, L.M., Boudreau, P.R., Freeman, K.R., 1984. Influences of
stock and site on growth and mortality in the blue mussel
(Mytilus edulis). Can. J. Fish. Aquat. Sci. 41, 134–141.
Ebling, F.J., Kitching, J.A., Muntz, L., Taylor, C.M., 1964. The
ecology of Lough Ine: XIII. Experimental observations of the
destruction of Mytilus edulis and Nucella lapillus by crabs.
J. Anim. Ecol. 33, 73–82.
Eertman, R.H.M., Wagenvoort, A.J., Hummel, H., Smaal, A.C.,
1993. bSurvival in airQ of the blue mussel Mytilus edulis L. as a
sensitive response to pollution-induced environmental stress.
J. Exp. Mar. Biol. 170, 179–195.
Fisheries and Aquaculture Division, Dept. of Fisheries, Aquaculture
and Environment, Prince Edward Island, 2001. P.E.I. mussel
monitoring program 2001 report. Fish. and Aqua. Tech. Rep. Ser.
Fuentes, J., Reyero, I., Zapata, C., Alvarez, G., 1992. Influence of
stock and culture site on growth rate and mortality of mussels
(Mytilus galloprovincialis Lmk.) in Galicia, Spain. Aquaculture
105, 131–142.
Fuentes, J., Reyero, I., Zapata, C., Alvarez, G., 1994. Production
traits of the mussel Mytilus galloprovincialis cultured in Galicia
(NW of Spain): relative effects of source of seed and growing
environment. Aquaculture 122, 19–31.
Gonzalez, J.G., Yevich, P., 1976. Responses of an estuarine
population of the blue mussel Mytilus edulis to heated water
from a steam generating plant. Mar. Biol. 34, 177–189.
Gosling, E.M., 1992. Systematics and distribution. In: Gosling, E.
(Ed.), The Mussel Mytilus: Ecology, Physiology, Genetics and
Culture. Elsevier Science Publishers, Amsterdam.
Gosling, E., 2003. Bivalve Molluscs Biology, Ecology and Culture.
Fishing News Books, Great Britain.
Guderley, H., Demers, A., Couture, P., 1994. Acclimatization of
blue mussel, (Mytilus edulis Linnaeus, 1758) to intertidal
conditions: effects on mortality and gaping during air exposure.
J. Shellfish Res. 13 (2), 379–385.
Incze, L.S., Lutze, R.A., Watling, L., 1980. Relationships between
effects of environmental temperature and seston on growth and
mortality onMytilus edulis in a temperate northern estuary. Mar.
Biol. 57, 147–156.
Lestor, L.J., 1983. Developing a selective breeding program for
penaeid shrimp mariculture. Aquaculture 33, 41–50.
Lewis, J.R., 1977. The role of physical and biological factors in the
distribution and stability of rocky shore communities. In:
Keegan, B.F, Ceidigh, P.O., Boaden, P.J.S. (Eds.), Biology of
Benthic Organisms. Proc.11th Eur. Mar. Biol. Symp., Galway,
Ireland. Pergamon Press, London.
Littell, R.C., Milliken, G.A., Stroup, W.W., Wolfinger, R.D., 1996.
SASR System for Mixed Models. SAS Institute, Cary, NC,
USA.
Livingstone, D.R., Bayne, B.L., 1977. Responses of Mytilus edulis
L. to low oxygen tension: anaerobic metabolism of the posterior
adductor muscle and mantle tissues. J. Comp. Physiol. 114B,
145–153.
Mallet, A.L., Carver, C.E.A., 1995. Comparative growth and
survival patterns of Mytilus trossulus and Mytilus edulis in
Atlantic Canada. Can. J. Fish. Aquat. Sci. 52, 1873–1880.
Mallet, A.L., Carver, C.E.A., Freeman, K.R., 1990. Summer
mortality of the blue mussel in eastern Canada: spatial,
temporal, stock, and age variation. Mar. Ecol., Prog. Ser. 67,
35–41.
Matthews, M.A., Mcmahon, R.F., 1995. Survival of zebra mussels
(Dreissena polymorpha) and Asian clams (Corbicula fluminea)
under extreme hypoxia. U.S. Army Corps of Engineers.
Waterways Experiment Station. Tech. Rep. EL-95-3.
Molares, J., Fuentes, J., 1995. Recruitment of the mussel
Mytilus galloprovincialis on collectors situated on the inter-
tidal zone in the Ria de Arousa (NW Spain). Aquaculture
138, 131–137.
Myrand, B., Gaudreault, J., 1995. Summer mortality of blue
mussels (Mytilus edulis Linneaus, 1758) in the Magdalen
Islands (southern Gulf of St. Lawrence Canada). J. Shellfish
Res. 14, 395–404.
Newell, R.C., Branch, G.M., 1980. The influence of temperature on
the maintenance of metabolic energy balance in marine
invertebrates. Adv. Mar. Biol. 17, 329–396.
Paine, R.T., 1974. Intertidal community structure: experimen-
tal studies on the relationship between a dominant
competitor and its principal predator. Oecologia (Berl.) 15,
93–120.

N. LeBlanc et al. / Aquaculture 243 (2005) 185–194194
Paukstis, G.L., Tucker, J.K., Bronikowski, A.M., Janzen, F.J., 1999.
Survivorship of aerially-exposed zebra mussels (Dressena
polymorpha) under laboratory conditions. J. Freshw. Ecol. 14
(4), 511–517.
Rasbash, J., Browne, W., Goldstein, H., Yang, M., Plewis, I., Healy,
M., Woodhouse, G., Draper, D., Langford, I., Lewis, T., 2000. A
User’s Guide to MLwiN, (Second edition). Institute of Educa-
tion, London.
Rawson, P.D., Joyner, K.L., Meetze, K., Hilbish, T.J., 1996.
Evidence for intragenic recombination within a novel marker
that distinguishes mussels in theMytilus edulis species complex.
Heredity 77, 599–607.
Read, K.R.H, Cumming, K.B., 1967. Thermal tolerance of the
bivalve molluscs Modiolus modiolus (L), Mytilus edulis L., and
Brachidontes demissus Dillwyn. Comp. Biochem. Physiol. 22,
149–155.
Seed, R., Suchanek, T.H., 1992. Population and community ecology
of Mytilus. In: Gosling, E. (Ed.), The Mussel Mytilus: Ecology,
Physiology, Genetics and Culture. Elsevier Science Publishers,
Amsterdam.
Suchanek, T.H., 1985. Mussels and their role in structuring rocky
shore communities. In: Moore, P.G., Seed, R. (Eds.), The
Ecology of Rocky Coasts. Holder and Stoughton, Sevenoaks,
UK.
Tremblay, R., Myrand, B., Guderley, H., 1998. Thermal
sensitivity of organismal and mitochondrial oxygen con-
sumption in relation to susceptibility of blue mussels,
Mytilus edulis (L.), to summer mortality. J. Shellfish Res. 17,
141–152.
G.A. Trevelyan, 1991. Aquacultural ecology of hatchery produced
juvenile bay mussels, Mytilus edulis L. PhD thesis, University
of California, USA.
Tsuchiya, M., 1983. Mass mortality in a population of the mussel
Mytilus edulis L. caused by high temperature on rocky shores.
J. Exp. Mar. Biol. 66, 101–111.
Wallis, R.L., 1975. Thermal tolerance of Mytilus edulis of Eastern
Australia. Mar. Biol. 30, 183–191.
Widdows, J., Shick, J.M., 1985. Physiological responses of Mytilus
edulis and Cardium edule to aerial exposure. Mar. Biol. 85,
217–232.
Related Documents