DEVELOPMENTAL NEUROPSYCHOLOGY, 38(5), 281–300 Copyright © 2013 Taylor & Francis Group, LLC ISSN: 8756-5641 print / 1532-6942 online DOI: 10.1080/87565641.2013.799672 The Development of Mismatch Responses to Mandarin Lexical Tones in Early Infancy Ying-Ying Cheng Instituteof Neuroscience, National Yang-Ming University, Taipei City, Taiwan Hsin-Chi Wu Taipei Tzuchi Hospital, The Buddhist Tzuchi Medical Foundation, New Taipei City, Taiwan, and School of Medicine, Tzu Chi University, Hualien, Taiwan Yu-Lin Tzeng Instituteof Neuroscience, National Yang-Ming University, Taipei City, Taiwan Ming-Tao Yang Far EasternMemorial Hospital, New Taipei City, Taiwan Lu-Lu Zhao Taipei Tzuchi Hospital, The Buddhist Tzuchi Medical Foundation, New Taipei City, Taiwan Chia-Ying Lee Institute of Neuroscience, National Yang-Ming University, Taipei City, Taiwan, The Instituteof Linguistics, Academia Sinica, Taipei City, Taiwan, and Instituteof Cognitive Neuroscience, National Central University, Taoyuan, Taiwan This study examined how maturation and the size of deviance affect the development of mismatch responses to Mandarin lexical tones by a multi-deviant oddball paradigm with both large deviant T1/T3 and small deviant T2/T3 pairs in newborns and 6-month-olds. The T1/T3 pair elicited a pos- itive mismatch response (P-MMR) at birth but an adult-like mismatch negativity (MMN) at 6 months of age. For the T2/T3 pair, no significant MMR was seen in newborns, whereas a P-MMR was found when infants are 6 months old. Results suggest that the developmental trajectories of MMRs are dependent on the neural maturation and the discriminability of tonal changes. Correspondence should be addressed to Chia-Ying Lee, The Institute of Linguistics, Academia Sinica, 128, Section 2, Academia Road 115, Taipei, Taiwan, R.O.C. E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DEVELOPMENTAL NEUROPSYCHOLOGY, 38(5), 281–300Copyright © 2013 Taylor & Francis Group, LLCISSN: 8756-5641 print / 1532-6942 onlineDOI: 10.1080/87565641.2013.799672
The Development of Mismatch Responses to MandarinLexical Tones in Early Infancy
Ying-Ying ChengInstitute of Neuroscience, National Yang-Ming University, Taipei City, Taiwan
Hsin-Chi WuTaipei Tzuchi Hospital, The Buddhist Tzuchi Medical Foundation, New Taipei City, Taiwan,
and School of Medicine, Tzu Chi University, Hualien, Taiwan
Yu-Lin TzengInstitute of Neuroscience, National Yang-Ming University, Taipei City, Taiwan
Ming-Tao YangFar Eastern Memorial Hospital, New Taipei City, Taiwan
Lu-Lu ZhaoTaipei Tzuchi Hospital, The Buddhist Tzuchi Medical Foundation, New Taipei City, Taiwan
Chia-Ying LeeInstitute of Neuroscience, National Yang-Ming University, Taipei City, Taiwan,
The Institute of Linguistics, Academia Sinica, Taipei City, Taiwan, andInstitute of Cognitive Neuroscience, National Central University, Taoyuan, Taiwan
This study examined how maturation and the size of deviance affect the development of mismatchresponses to Mandarin lexical tones by a multi-deviant oddball paradigm with both large deviantT1/T3 and small deviant T2/T3 pairs in newborns and 6-month-olds. The T1/T3 pair elicited a pos-itive mismatch response (P-MMR) at birth but an adult-like mismatch negativity (MMN) at 6 monthsof age. For the T2/T3 pair, no significant MMR was seen in newborns, whereas a P-MMR was foundwhen infants are 6 months old. Results suggest that the developmental trajectories of MMRs aredependent on the neural maturation and the discriminability of tonal changes.
Correspondence should be addressed to Chia-Ying Lee, The Institute of Linguistics, Academia Sinica, 128, Section 2,Academia Road 115, Taipei, Taiwan, R.O.C. E-mail: [email protected]

282 CHENG ET AL.
A growing body of studies suggests that the auditory cortical response to acoustic changesrecorded by event-related potentials (ERPs) in early infancy may serve as early markers for pre-dicting later language and cognitive development. For example, longitudinal studies reportedthat newborns’ ERPs to the processing of stop-consonant vowel (CV) syllables predict theirpre-reading language skills between 2.5 to 6.5 years of age (Guttorm, Leppänen, Hamalainen,Eklund, & Lyytinen, 2010; Guttorm et al., 2005; Molfese & Molfese, 1985, 1997) and theirreading performance at 8 years of age (Molfese, 2000). ERPs to the auditory changes of puretones in newborns (Leppänen et al., 2010) and of stress patterns in 5-month-olds (Weber, Hahne,Friedrich, & Friederici, 2005) also predict their language outcomes. However, the maturationtimetable of auditory change-related cortical responses varies in different sound features, whichplay different weightings across languages. This study investigated how maturation and acousticsimilarity affect the development of mismatch responses (MMRs) to Mandarin lexical tones inadults and infants from birth to 6 months old.
The auditory change-related cortical response can be indexed by an ERP component calledmismatch negativity (MMN). MMN is typically obtained using an auditory oddball paradigm, inwhich a deviant in certain aspects of a sound feature occurs infrequently in a sequence of repeti-tive homogeneous (standard) stimuli (Näätänen, Kujala, & Winkler, 2011; Näätänen, Paavilainen,Rinne, & Alho, 2007). In adults, MMN is a frontal-central distributed negativity peaking between100 msec and 250 msec by subtracting the ERP to the standard from that to the deviant. TheMMN amplitude increases, whereas the peak latency decreases as the discriminability of thestandard and the deviant sounds rise. MMN is hypothesized to index change detection based on asound representation constructed to a repeated auditory input and to reveal whether listeners haveformed sufficient and robust representations of automatic pre-attentive discrimination (Näätänenet al., 2007; Winkler, 2007). Cross-linguistic studies have shown that MMN is sensitive to lan-guage experiences, even for infants one year old and younger. Cheour and colleagues (1998a)reported that Finnish 12-month-olds showed an enhanced MMN response to their native vowelcontrast, as compared with the non-native Estonian vowel contrast, even when the non-native con-trast had a more distinct acoustic difference (Cheour et al., 1998a). Infants as young as 4 monthsof age showed processing advantages to the typical stress pattern in their native language indexedby the presence of mismatch responses (Friederici, Friedrich, & Christophe, 2007). It suggeststhat, in addition to automatic change detection in short-term memory, MMN is sensitive to thelong-term memory trace built on language experience. Most important, the MMN can be elicited,even when a participant does not attend to the stimuli, such as reading a book or watching asilent movie. Thus, the MMN serves as an excellent tool for assessing auditory discrimination,especially for infants and children with limited attention or motivation. Tracing MMN to variousspeech units might demonstrate how infants allocate attention to processing speech at differentages, and can provide information on whether native language speech perception has becomeautomatized.
Although MMN is well established in adults, the polarity and latency of mismatch response ininfants are highly inconsistent across studies. Alho, Sainio, Sajaniemi, Reinikainen, and Naatanen(1990) first reported an adult-like MMN, which has mean peak latencies 296 msec at Fz and270 msec at Cz, for pure tone deviance in quiet sleeping newborns. Cheour et al. found anadult-like MMN peaking at approximately 380 msec to 400 msec in newborns and infants whenresponding to changes in pure tones (Cheour, Kushnerenko, Ceponiene, Fellman, & Näätänen,2002; Cheour et al., 2002). Other studies also demonstrated adult-like MMNs in infants for

MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 283
the duration changes (Brannon, Libertus, Meck, & Woldorff, 2008; Brannon, Roussel, Meck,& Woldorff, 2004) and the changes in speech sounds (Cheour et al., 1998b; Cheour-Luhtanenet al., 1995; Kushnerenko et al., 2001; Martynova, Kirjavainen, & Cheour, 2003). However, suchmismatch negativities in newborns and infants usually persist for a longer interval and in a rela-tively late time window than those typically seen in adult data. Other studies instead reportedthat mismatch responses in infants are often a positivity between 200 msec and 450 msec,rather than an adult-like negativity (Dehaene-Lambertz & Baillet, 1998; Dehaene-Lambertz &Dehaene, 1994; Friederici, Friedrich, & Weber, 2002; Jing & Benasich, 2006; Leppänen, Eklund,& Lyytinen, 1997; Morr, Shafer, Kreuzer, & Kurtzberg, 2002; Novitski, Huotilainen, Tervaniemi,Näätänen, & Fellman, 2007). For example, Leppänen et al. (1997) observed a positive mismatchresponse (P-MMR) peaking between 250 msec and 350 msec to the pure tone change in new-borns. Dehaene-Lambertz and Dehaene (1994) reported that three-month-old infants showeda P-MMR peaking at approximately 390 msec to initial consonant change (/ba/ vs. /ga/).Friederici et al. (2002) examined the mismatch response to syllables varying in vowel duration(short /ba/ vs. long /ba:/) in 2-month-old infants and found a P-MMR peaking at approximately400 msec, especially when considering long syllables as the deviance. Generally, the P-MMRwas found mainly at a younger age and it can be elicited by various speech and non-speechchanges.
The characteristics of the P-MMR remain unclear. Some studies have suggested that the polar-ity of mismatch response depend on maturation factors. For example, Leppänen et al. (2004)reported that the amplitude of mismatch response was positively correlated with physiologicalmaturation in infants. A more mature newborn tends to have a more positive mismatch response.He, Hotson, and Trainor (2007) examined the brain responses of 2- to 4-month-old infants toinfrequent pitch changes of piano tones. Their data showed an increase in the left lateralized pos-itive slow wave at 2 and 3 months of age, whereas a faster adult-like MMN was present in 3- to4-month-old infants. Trainor et al. (2003) reported that the mismatch response turned from pos-itive to negative between 2 and 6 months of age. Kushnerenko, Ceponiene, Balan, Fellman, andNäätänen (2002) longitudinally traced the development of pitch change detection in a group ofinfants from birth until 12 months of age. Their data showed that the adult-like MMN stabilizedbetween 3 to 6 months of age, although a substantial MMN variability from age to age withinthe same infant was observed. These findings imply that the adult-like MMN becomes promi-nent, whereas the P-MMR diminishes as infants age. These observations suggest that the neuralmaturation may play a role in the developmental changes of mismatch responses.
Other studies have suggested that stimulus-related factors, such as short inter-stimulus inter-vals and smaller magnitude of deviance, decrease the discriminability between standard anddeviant, which might be responsible for the polarity transition of mismatch responses. Morret al. (2002) examined how maturation and the size of deviance affect the polarity of mismatchresponse in infants and preschoolers. Their data showed that the majority of infants failed toexhibit an adult-like MMN to the small deviance (1,000 Hz vs. 1,200 Hz). The adult-like MMNcannot be elicited reliably by the small deviance until 4 years of age. Conversely, the adult-likeMMN presents in most infants and preschoolers to the large deviance (1,000 Hz vs. 2,000 Hz).Maurer, Bucher, Brem, and Brandeis (2003) further examined mismatch responses in adultsand 6- to 7-year-old children with substantially smaller frequency (1,000 Hz vs. 1,060 Hz) andphoneme deviance (“ba” vs. “ta” or “da”) with shorter intervals relative to those in most previousstudies. Although a typical frontal-central MMN was evident in adults, the positive mismatch

284 CHENG ET AL.
response remained pertinent in 6- to 7-year-olds (Maurer et al., 2003). These findings suggestedthat both stimulus-related and biological criteria are required for P-MMR to switch to MMN.
Spoken languages vary in phonological structures and speech features, which can be segmen-tal units, such as phonemes, or supra-segmental units, such as word stress and tone. Some featuresare relatively common across different languages. For example, nearly all languages have at leastthree phonemic vowels, /i/, /a/, and /u/. However, some features are unique to certain lan-guages, such as vowel duration for Finnish, word stress for German and English, and lexical tonefor Thai and Chinese. Accurate speech perception is based on language-specific memory tracesof the essential speech units for each language that developed during language acquisition and arerepresented in long-term memory (Näätänen et al., 2007). Previous studies have established thatlanguage experience influences the automatic involuntary processing of speech sounds (Cheouret al., 1998a; Friederici et al., 2007; Naatanen et al., 1997), thus suggesting that MMN can serveas an index of a language-specific memory trace for early identification of children at risk oflanguage deficit (Leppänen et al., 2010; Weber et al., 2005). Lexical tone is an essential featureof Mandarin Chinese. This study aims to uncover the developmental trajectories of mismatchresponses to Mandarin lexical tones in early infancy.
As mentioned, Mandarin Chinese is a tonal language that exploits variations in pitch at thesyllable level to determine different meanings. The four lexical tones can be categorized phono-logically into a high-level tone (T1), a high-rising tone (T2), a low-dipping tone (T3), and ahigh-falling tone (T4). The major sign for discriminating different lexical tones is the contour offundamental frequency (F0). Studies on speech production showed that, although children mas-ter most of the tones by 3 years of age (Hua & Dodd, 2000; Wong, Schwartz, & Jenkins, 2005),each of the lexical tones has a different developmental trajectory. For production, T1 and T4 aremastered earlier than T2 and T3 (Clumeck, 1980; Hua, 2002; Li & Tompson, 1977). Studies ontonal perception also showed a similar developmental trajectory. In a picture-pointing task, 3-year-old children showed higher accuracy in perceiving T1, T2, and T4 (90%, 87%, and 89%,respectively) than that for T3 (70%), in which the most frequent error was misidentifying T3 asT2 (Wong et al., 2005). Multidimensional scaling (MDS) analysis is a statistical technique foranalyzing proximity data in a set of stimuli to reveal the hidden structure of the data. MDS analy-sis of tone perception involves constructing dissimilarity matrices based on direct comparisons oftonal pairs. Using MSD analysis, Gandour and his colleagues have reported that pitch contour andpitch height are crucial for characterizing Mandarin tone perception (Gandour, 1983; Gandour &Harshman, 1978). For pitch contour and direction, T2 is acoustically more similar to T3 thanT1. Tonal discrimination and identification data confirm that T2 and T3, which are acousticallythe most similar, are more often confused with one another, compared to other tonal pairs. Tsao(2008) examined whether acoustic similarity between lexical tones would affect the perceptualdiscrimination performance of 12-month-old infants by using the head-turn technique, in whichinfants are taught to turn their heads to a sound or to a change in sound. This technique is idealfor understanding the speech perception capability of infants between 6 and 18 months of age andhow such capability changes as a function of language experience and development. Tsao (2008)showed that the distinction accuracy between T1 and T3, which is the most acoustically distinctcontrast, is higher than that of the other contrasts (T2 vs. T4 and T2 vs. T3). This suggested thatthe acoustic similarity affects Mandarin lexical tone discrimination as early as 1 year of age.
A few studies concerned mainly with the hemispheric dominance of tonal processing inves-tigated the MMNs for Mandarin lexical tone in adults. A number of studies reported that MMNfor lexical tone was larger in the right fronto-central sites than in the left fronto-central sites (Luo

MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 285
et al., 2006; Ren, Yang, & Li, 2009; Xi, Zhang, Shu, Zhang, & Li, 2010). Luo et al. (2006) con-ducted a dipole analysis to localize the neural generator of MMN for lexical tone. They reportedthat the dipole was significantly stronger in the right hemisphere than in the left hemisphere,and that the hemispheric lateralization of MMN for consonant deviance has a reverse pattern.However, other studies failed to find a significant right lateralization of MMNs for lexical tone(Chandrasekaran, Krishnan, & Gandour, 2007a,b, 2009). Furthermore, Xi et al. (2010) examinedthe categorical perception of Mandarin lexical tone by MMN for within- and across-categorydeviance selected from a 10-interval continuum between T2 and T4. Their data showed thatMMN for cross-category deviance was larger than that for within-category deviance only in theleft-distributed sites. The contradiction on scalp distribution suggested that MMN reflected bothacoustical and phonological processing of tonal changes. Regarding how the size of devianceaffects MMNs for Mandarin lexical tone, Chandrasekaran et al. (2007a) compared MMNs fordifferent tonal pairs. Their data showed that the acoustically distinct T1/T3 pair elicited largerMMN with earlier peak latency than the acoustically similar T2/T3 pair. However, the acous-tical similarity effect on MMN was observed only in the native Chinese group, but not in thenative English group (Chandrasekaran et al., 2007a). Their following studies supported that MMNcould index the experience-dependent neural plasticity to the acoustic features of lexical tones(Chandrasekaran, Gandour, & Krishnan, 2007b; Chandrasekaran et al., 2009).
Although the adult data showed that deviance size affects MMNs for Mandarin lexical tones,the developmental trajectory of this effect remains unclear. Meng et al. (2005) addressed whetherdyslexic children have neural deficits in auditory processing. In a part of their experiments, theyexamined the MMNs to lexical tone by subtracting ERPs to the standard “ba1” (T1) from ERPsto the deviant “ba2” (T2) in 8- to 13-year-old children with and without dyslexia. Their datasuggested that no difference exists in mismatch responses to lexical tone between dyslexic andnormal children. However, they did not examine if a significant mismatch response (the differencebetween the ERPs elicited by standard and that elicited by the deviant) exists in each group.In other words, whether the dyslexic group and their matched control showed significant MMNto the T1/T2 contrast is unclear. A more discreet data analysis is required to investigate themismatch response to lexical tone in early childhood.
Lexical tone awareness has been shown to be strongly associated with Chinese reading(McBride-Chang et al., 2008), suggesting that supra-segmental perception is particularly essen-tial in exploring Chinese reading development and impairment. The current study used MMN,which has been used to index an automatic pre-attentive discrimination of speech sounds, forinvestigating the development of lexical tone perception in early infancy. In addition, whetherthe size of deviance affects the mismatch response to lexical tones remains unclear, especially indevelopmental populations. Tsao (2008) showed that acoustic similarity affects perceptual dis-crimination performance in infants as young as one year of age. Thus, this study examined howboth biological maturation and deviance size affect the mismatch responses to Mandarin lex-ical tone in infants at birth and at 6 months of age. The size-of-deviance effect on mismatchresponses, both the acoustically distinct T1/T3 and the acoustically similar T2/T3 contrastswere used with a multi-deviant oddball paradigm (Näätänen, Pakarinen, Rinne, & Takegata,2004). Two experiments were conducted: Experiment 1 involved determining whether typicalMMN and the size-of-deviance effect can be demonstrated in the mature brain. The results ofthis experiment shall be able to replicate those of Chandrasekaran et al. (2007a) by showingthat a larger deviance (T1/T3 pair) elicits larger and earlier MMN than does a smaller deviance(T2/T3 pair) in adult native Mandarin Chinese speakers. Then, the same paradigm was used

286 CHENG ET AL.
in Experiment 2 to explore the developmental trajectories of mismatch responses from birth to6 months of age and to determine how the difficulty of lexical tone discriminability affects thematuration on mismatch responses. Previous studies have shown that mismatch responses couldswitch from positive to negative with growth in younger age (He et al., 2007; Morr et al., 2002).The transition from a P-MMR to adult-like MMN is expected in mismatch responses to lexicaltones in early infancy, especially for the acoustically distinct T1/T3 contrast.
EXPERIMENT 1: MMNS FOR MANDARIN LEXICAL TONES IN ADULTS
Methods
Participants. Eighteen native speakers of Mandarin Chinese (2 women, age range =18–29) with normal hearing were paid to participate in this study. All the participants are col-lege or graduate students without a history of neurological or psychological issues. The currentstudy was approved by the Human Subject Research Ethics Committee/IRB. Written consentforms were obtained from all participants.
Stimuli. The stimuli consisted of three Mandarin syllables with different lexical tones: yi1,“clothing” (T1), yi2 “aunt” (T2), and yi3 “chair” (T3), which share the same vowel /i/, butcarry different tonal contours. The T3 was assigned as the standard, with T1 and T2 as deviants.The T1/T3 pair represents the larger deviant contrast, and the T2/T3 pair represents the smallerdeviant contrast. All the stimuli are meaningful syllables in Mandarin Chinese. These stimuliwere pronounced by a native female speaker of Mandarin and were recorded at 16 bits, with asampling rate of 44 kHz. Intensity and duration of the stimuli were normalized to 70 dB and250 msec, respectively, with Sony Sound Forge 9.0 software.
Procedure. Participants were seated in a soundproof and electrically shielded room. Theywere instructed to play a puzzle computer game called “super-box” silently while listen passivelyto auditory stimuli. The stimuli were presented at a sound pressure level (SPL) of 70 dB througha set of loudspeakers located approximately 75 cm in front of the participants. The experimen-tal session started with 20 trials of standard, followed by 1,000 trials with 80% standard and20% deviants (10% for each deviant). To establish a memory trace for the standard trial beforeeach deviant, the pseudo-randomized sequence with at least two successive standards betweendeviants was adopted. Such a design has been widely used in MMN studies for both adults andinfants (Cheour et al., 2002; He et al., 2007; Kushnerenko et al., 2007; Shestakova, Huotilainen,Ceponiene, & Cheour, 2003). In each trial, the stimuli lasted 250 msec, with 500 msec ofinter-stimulus interval.
EEG recording and data analysis. The electroencephalogram (EEG) signals were ampli-fied by SYNAMPS2
®(Neuroscan, Inc.) in DC mode, low-pass 100 Hz, and digitized at a
sampling rate of 500 Hz. EEG data were recorded from 64 Ag/AgCl electrodes (QuickCap,Neuromedical Supplies, Sterling, USA), arranged according to the international 10–20 system,including a reference electrode located between Cz and CPz and a ground electrode locatedbetween Fpz and Fz. Six additional electrodes were attached over the left and right mastoids,supra- and infra-orbital ridges of left eye (VEOG), and outer canthi of both eyes (HEOG).

MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 287
For offline analysis, the EEG data were re-referenced to the average of the left and rightmastoids. Continuous EEG was segmented into epochs from 100 msec prior to the onset of thestimulus to 700 msec after onset, and the pre-stimulus interval was used for baseline correction.A 1 Hz to 30 Hz (zero phase shifting, 12 dB/oct) band-pass filter was applied before artifact rejec-tion. The first 20 trials and epochs with artifacts exceeding ±100 µV were rejected. To ensurethat the ERP to standard does not detect any change-detection response, the post deviant stan-dards are usually excluded from the standard averages (see Ahmmed, Clarke, & Adams, 2008;Cheour et al., 2002; He et al., 2007). To ensure further that the numbers of trials between thestandard and deviant were comparable, only the standard trials that were preceded by at leastthree successive standards were included for analyses. For each participant, at least 45 accepteddeviants were required to be included in further analyses. The mean number of accepted deviantswas 76.56, and that of accepted standards was 173.5. The grand averaged ERPs for standard,small deviant, and large deviant were computed for each participant and each electrode.
To evaluate the spatial-temporal ERP differences between standard and each deviant, repeatedmeasures ANOVAs with conditions (standard, large deviant, and small deviant) and electrodesites (F3, FZ, F4, FC3, FCZ, FC4, C3, CZ, and C4) as within-subject factors were conducted in amean amplitude of six successive epochs of 50 msec each, from 50 msec to 350 msec. This pro-cedure allowed for identification of the temporal courses at which the mismatch effect appeared.The planned comparisons were conducted between the large deviant and standard (T1/T3), andbetween the small deviant and the standard (T2/T3), to determine whether both types of deviantselicited significant MMRs. In all ANOVAs, the Greenhouse-Geisser adjustment to the degreesof freedom was applied to correct violations of sphericity associated with repeated measures.Accordingly, the corrected p-value for all the F tests with more than one degree of freedom inthe numerator is reported. Only the effects or interactions showing significant differences (p <
.05) in at least two consecutive time windows were considered meaningful and reported. To eval-uate further the precise moments of MMRs for both T1/T3 and T2/T3 contrasts, MMN peaklatencies obtained from each difference wave for each participant between 100 to 300 msec weresubmitted to a repeated measures ANOVA under particular conditions (T1/T3 and T2/T3) andelectrode sites (F3, FZ, F4, FC3, FCZ, FC4, C3, CZ, and C4) as within-subject factors.
Results
MMN data were derived by subtracting the standard from each deviant, resulting in individualMMNs for T1/T3 and T2/T3. The grand average of MMNs was fronto-to-central-distributedbetween 100 msec and 300 msec after the onset of stimuli (Figure 1). The main effect of thecondition was significant in four consecutive time windows: 100–150 msec (F(2,34) = 30.56, p <
.0001), 150–200 msec (F(2,34) = 54.52, p < .0001), 200–250 msec (F(2,34) = 16.75, p < .0001),and 250–300 msec (F(2,34) = 26.79, p < .0001). Interactions between conditions and sites weresignificant in two consecutive time windows: 100–150 msec (F(16,272) = 3.52, p < .005) and150–200 msec (F(16,272) = 4.05, p < .005). Post hoc analysis revealed that the condition effectwas significant in all selected sites for both time windows (F(2,272) = 100.38–527.07, ps <
.0001). It revealed the difference among standard and two deviants was significant in intervalsfrom 100 to 300 msec. Planned comparison revealed that MMN for T1/T3 was significant in theintervals of 100–150 msec (F(1,34) = 48.67, p < .0001) and 150–200 msec ((F(1,34) = 96.26,
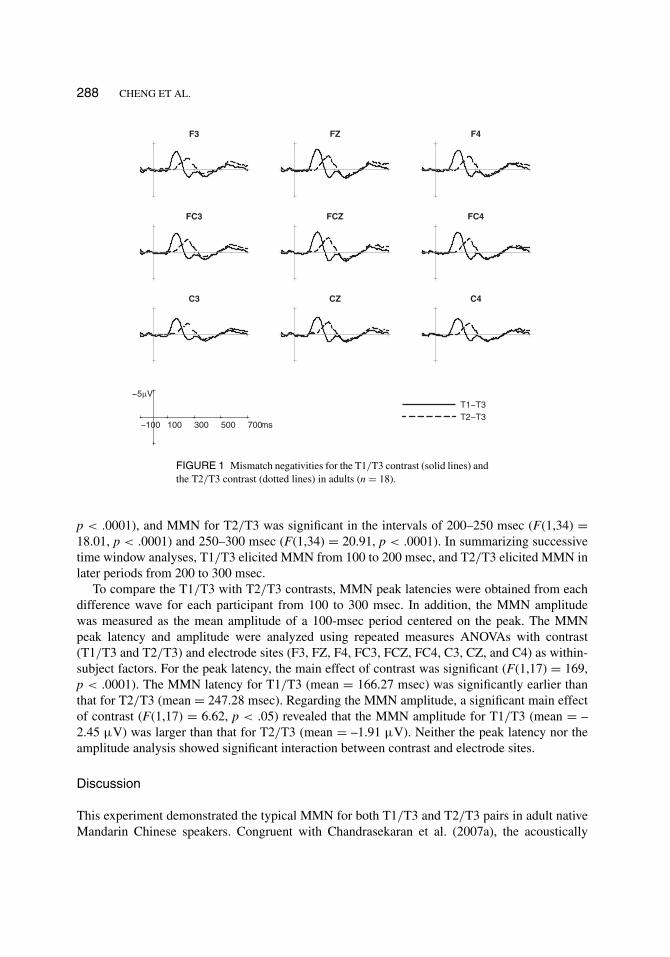
288 CHENG ET AL.
F3 FZ F4
FC3 FCZ FC4
C3 CZ C4
T1−T3T2−T3
−5µV
−100 100 300 500 700ms
FIGURE 1 Mismatch negativities for the T1/T3 contrast (solid lines) andthe T2/T3 contrast (dotted lines) in adults (n = 18).
p < .0001), and MMN for T2/T3 was significant in the intervals of 200–250 msec (F(1,34) =18.01, p < .0001) and 250–300 msec (F(1,34) = 20.91, p < .0001). In summarizing successivetime window analyses, T1/T3 elicited MMN from 100 to 200 msec, and T2/T3 elicited MMN inlater periods from 200 to 300 msec.
To compare the T1/T3 with T2/T3 contrasts, MMN peak latencies were obtained from eachdifference wave for each participant from 100 to 300 msec. In addition, the MMN amplitudewas measured as the mean amplitude of a 100-msec period centered on the peak. The MMNpeak latency and amplitude were analyzed using repeated measures ANOVAs with contrast(T1/T3 and T2/T3) and electrode sites (F3, FZ, F4, FC3, FCZ, FC4, C3, CZ, and C4) as within-subject factors. For the peak latency, the main effect of contrast was significant (F(1,17) = 169,p < .0001). The MMN latency for T1/T3 (mean = 166.27 msec) was significantly earlier thanthat for T2/T3 (mean = 247.28 msec). Regarding the MMN amplitude, a significant main effectof contrast (F(1,17) = 6.62, p < .05) revealed that the MMN amplitude for T1/T3 (mean = –2.45 µV) was larger than that for T2/T3 (mean = –1.91 µV). Neither the peak latency nor theamplitude analysis showed significant interaction between contrast and electrode sites.
Discussion
This experiment demonstrated the typical MMN for both T1/T3 and T2/T3 pairs in adult nativeMandarin Chinese speakers. Congruent with Chandrasekaran et al. (2007a), the acoustically

MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 289
distinct T1/T3 pair elicited an earlier and larger MMN than the acoustically similar T2/T3 did.This corresponds with the findings of behavioral studies on lexical tone perception (Huang &Johnson, 2010; Tsao, 2008; Wong et al., 2005) and suggests that MMN is sensitive to the difficultyof lexical tone discrimination.
MMN has been shown to be influenced by long-term experience (Näätänen et al., 2007).Recent studies have suggested that the lexical properties may also contribute to the MMN effect(Pulvermuller, Shtyrov, Kujala, & Naatanen, 2004). When listeners were presented with word andpseudo-word deviants among pseudo-word standards, the MMN elicited by word deviants weremore pronounced than those elicited by acoustically matched pseudo-word deviants. This MMNenhancement could be caused by the activation of cortical memory traces for words. Consideringthat every Mandarin syllable also corresponds to at least one Chinese character, it is crucial toclarify whether the greater difficulty for T2/T3 than that for T1/T3 is solely because of acousticsimilarity, or may confound with other lexical factors, such as lexical frequency.
For lexical frequency, all three stimuli yi1 (T1), yi2 (T2), and yi3 (T3) used in this study aremeaningful Mandarin syllables. They are matched according to their type frequency; meaningthat each can be written according to 9 to 12 frequently used Chinese homophones, based onthe top 50% of the frequency ranking of Chinese characters. However, regarding the token fre-quency (the sum of character frequency of homophones), yi2 (T2) has a lower token frequency(34,886 per 10 millions) than do yi1 (T1) and yi3 (T3) (155,866 and 111,184 per 10 million,respectively). However, previous studies have suggested that lexical frequency is actually a rel-atively insignificant factor causing MMN enhancement, as compared to the lexical status ofthe stimuli (Pulvermuller, et al., 2001, 2004). Most important, Chandrasekaran and colleagues(2007a) showed that the peak latency of the T2/T3 condition is shifted later when compared tothe T1/T3 condition in both English and Chinese groups. Because the lexical frequency is mean-ingful only for Chinese group, the possible confounding factor from the lexical frequency seemsuntenable. However, in future studies, controlling the various acoustic and lexical parameters inthe speech stimuli that potentially contribute to the mismatch response might be more favorable.
In summary, our adult data demonstrated the typical MMN for both T1/T3 and T2/T3 pairsand suggests that the current experimental design is feasible to elicit MMN. Lee et al. (2012) usedthe same paradigm in 4- to 6-year-old preschoolers and showed that all of the groups exhibitedadult-like MMN to the T1/T3 contrast, but elicited P-MMR to the T2/T3 pair (Lee et al., 2012).Whether adult-like MMN can be found in early infancy remains to be seen.
EXPERIMENT 2: MISMATCH RESPONSE TO MANDARIN LEXICALTONES IN INFANTS
This experiment investigates the mismatch responses to Mandarin lexical tones in infants frombirth to 6 months of age, and examines how the deviance size effect shapes the maturation ofmismatch responses. Previous studies have reported that infants showed either a P-MMR or anadult-like MMN for speech sounds, but the effects of maturation and deviance size on the tran-sition of polarity have never been examined simultaneously. Therefore, we collected mismatchresponses by using the multi-deviant oddball paradigm several days after birth of the infants andat 6 months of age. The maturation effect could be examined by comparing the property of mis-match response in two age groups. Furthermore, the deviance size effect on the emergence and

290 CHENG ET AL.
polarity of mismatch responses could be examined by inspecting the independent response toT1/T3 and T2/T3.
Methods
Participants. Thirty-one full-term infants whose parents were native speakers of MandarinChinese participated in this study. The study protocols were approved by the Human SubjectResearch Ethics Committee/IRB and written consent forms were obtained from the parentsfor their infants’ participation. All the participants had a gestational age (GA) in the range of37 to 41 weeks and a birth weight in the range of 2,200 to 3,760 grams, and their 1- and 5-min Apgar scores were higher than 8. All the participants passed the acoustic emission testfor hearing screening. For newborns, ERP data were collected within 13 days after birth whilethey were asleep. The data of 6 newborns among all the participants (n = 31) were excludedfrom analysis because of setting errors or their failing to fall asleep. Thereafter, data from 25(10 female) newborns were included for final analysis. It was required that all the 31 partic-ipants return for their second stage of ERP recording when they were 6 months old (within15 days). At this stage, eight infants dropped out because they are incapable of staying quietduring data collection. Accordingly, the final analysis included data from 23 6-month-olds, inwhich 13 (3 female) infants were sleeping, and the remaining 10 (7 female) infants were awakeduring the ERP recording. Their cognitive outcomes were assessed by using the Bayley Scalesof Infant Development-Mental Developmental Index (MDI) before the 6-month-old ERP record-ing. All of the participants had a MDI within normal limits (ranging from 85 to 114), except twoinfants in the waking group. The follow-up assessments showed that the two sub-average infantshad a MDI within normal limits when at 12 or 18 months of age.
Stimuli and procedures. The stimuli and the multi-deviant oddball task were the same asthose in Experiment 1. During data recording, sleeping infants were either lying on a bed orheld by their caregivers. The waking infants were seated comfortably on their caregivers’ laps.In following similar procedures that have been used in other infant MMN studies (Morr et al.,2002; Shafer, Yu, & Datta, 2011), silent movies or cartoons were played on a monitor in front ofthe waking infants to engage them and minimize their movement. An experimenter entertainedthe infants with quiet toys whenever they lost interest in the videos.
EEG recording and data analysis. Infants’ EEG signals were amplified by NuAmps(Neuroscan, Inc.) in DC mode, with 100 Hz low-pass and 60 Hz notch filters, and digitized onlineat a rate of 500 Hz. For newborns, Ag/AgCl electrodes were attached to eight scalp sites: FPz,F3, Fz, F4, C3, C4, O1, O2, and left and right mastoids according to the international 10–20 sys-tem. The FPz was considered the ground electrode, and the EEG was referenced online to Fz.Eye movement was monitored with two electrodes: one at the supra-outer canthus of the left eye,and the other at the infra-outer canthus of the right eye. The setting for 6-month-old infants wasthe same as that for neonates, except that all the electrodes were held with an elastic cap.
The procedure for EEG data processing and artifact reject was the same as that used inExperiment 1. The repeated measures ANOVAs with conditions (standard, large deviant, andsmall deviant) and electrode sites (F3, F4, C3, and C4) as within-subject factors were conductedin a mean amplitude of nine consecutive 50 msec time intervals, from 50 msec to 500 msec

MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 291
after stimulus onset, for examining the time course of the condition effects across electrodes.The planned comparisons between the large deviant and standard (T1/T3) and between the smalldeviant and the standard (T2/T3) were also conducted in each electrode. Only effects or inter-actions showing significant differences (p < .05) in at least two consecutive time windows wereconsidered meaningful and reported.
Results
Newborn infants. ERPs did not show N1-P2 deflection for auditory stimuli (Figure 2A).Two-way ANOVA revealed significant main effects of the condition in two separate time win-dows: the 200–250 msec (F(2,48) = 7.41, p = .0018) and the 350–400 msec (F(2,48) = 3.64, p =.0394). The interaction between conditions and electrodes was marginally significant in two con-secutive time windows: 250–300 msec (F(6,144) = 2.17, p = .0858) and 300–350 msec (F(6,144)= 2.19, p = .084). MMRs could be inspected from the difference waves derived by subtractingthe standard from each deviant (Figure 2B). Planned comparisons showed that T1 was signifi-cantly more positive than T3 in four consecutive time windows on F3: 300–350 msec (F(1,24) =7.62, p = .0082), 350–400 msec (F(1,24) = 5.98, p = .0182), 400–450 msec (F(1,24) = 5.57,p = .0224), and 450–500 msec (F(1,24) = 4.61, p = .0369). The difference between T2 andT3 was significant in the 200 msec to 250 msec time window on all electrodes (F(1,24) = 5.91(F3), 5.40 (C3), 5.55 (F4) and 8.08 (C4), ps < .05), although it did not fulfill the criterion of twoconsecutive time windows. In summary, the newborns showed a P-MMR to T1/T3 in 300 msecto 500 msec on the left frontal electrode (F3), but did not show a significant mismatch responseto T2/T3.
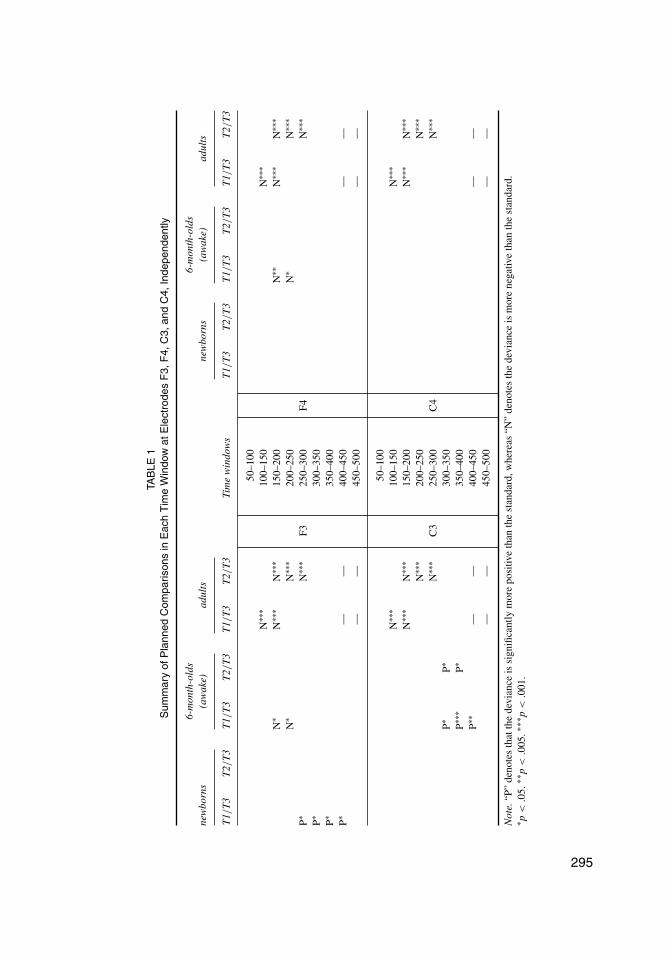
6-month-old infants. ERPs in 6-month-old infants were subdivided into sleeping (n =13) and waking groups (n = 10). ERPs showed typical N1-P2 deflection for auditory stimuliin both subgroups. For the waking group, the two-way ANOVA showed significant main effectsof condition in two consecutive time windows: 150–200 msec (F(2,18) = 6.17, p = .0144) and200–250 msec (F(2,18) = 4.23, p = .0331). The interaction between conditions and electrodeswas significant in three consecutive time windows: 250–300 msec (F(6,54) = 3.62, p = .0177),300–350 msec (F(6,54) = 3.79, p = .0257) and 350–400 msec (F(6,54) = 3.00, p = .0395).As shown in Figure 3B, the T1/T3 difference showed negativity followed by positivity, but theT2/T3 difference showed only positivity. For the T1/T3 pair, planned comparisons showed sig-nificant mismatch negativity in two consecutive time windows on F3 and F4: 150–200 msec(F(1,18) = 7.83 (F3) and 11.37 (F4), ps < .05) and 200–250 msec (F(1,18) = 8.08 (F3) and 4.94(F4), ps < .05), and a significant mismatch positivity in three consecutive time windows on C3:300-350 msec (F(1,18) = 7.02, p = .0163), 350–400 msec (F(1,18) = 15.56, p = .0009) and400–450 msec (F(1,18) = 11.92, p = .0028). For the T2/T3 pair, planned comparisons showed asignificant mismatch positivity in two consecutive time windows on C3: 300–350 msec (F(1,18)= 5.61, p = .0293) and 350–400 msec (F(1,18) = 5.04, p = .0375). A summary of the sig-nificance of planned comparisons is shown in Table 1. For the sleeping group, no significantcondition effect was found. In summary, waking 6-month-old infants showed an adult-like MMNfor T1/T3 in 150 msec to 250 msec on the frontal electrodes (F3 and F4), and a P-MMR to bothT1/T3 and T2/T3 in 300 msec to 450 msec on the left central electrodes (C3).
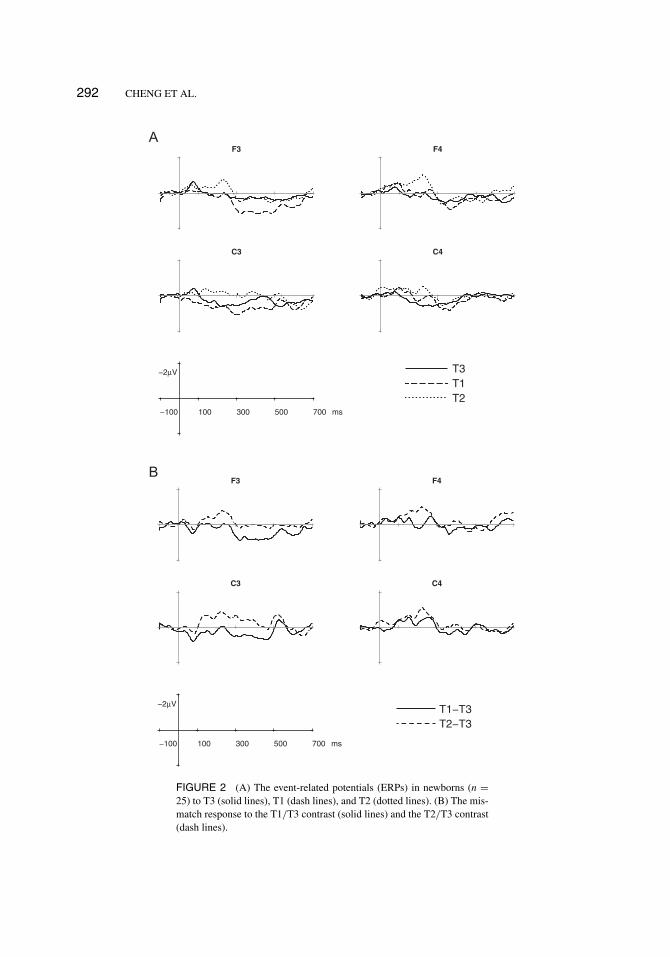
292 CHENG ET AL.
F3 F4
C3 C4
T3T1T2
−2µV
−100 100 300 500 700 ms
A
F3 F4
C3 C4
T1−T3T2−T3
−2µV
−100 100 300 500 700 ms
B
FIGURE 2 (A) The event-related potentials (ERPs) in newborns (n =25) to T3 (solid lines), T1 (dash lines), and T2 (dotted lines). (B) The mis-match response to the T1/T3 contrast (solid lines) and the T2/T3 contrast(dash lines).
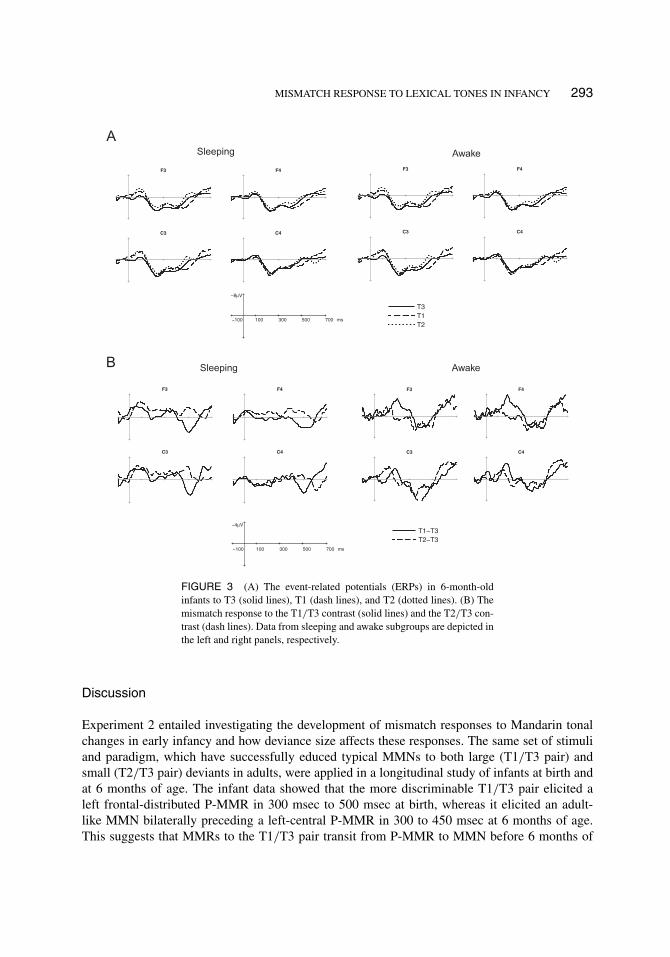
MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 293
F3 F4
C3 C4
Sleeping
F3 F4
C3 C4
T3T1T2
−8µV
−100 100 300 500 700 ms
AAwake
F3 F4
C3 C4
Sleeping
F3 F4
C3 C4
T1−T3T2−T3
−4µV
−100 100 300 500 700 ms
B Awake
FIGURE 3 (A) The event-related potentials (ERPs) in 6-month-oldinfants to T3 (solid lines), T1 (dash lines), and T2 (dotted lines). (B) Themismatch response to the T1/T3 contrast (solid lines) and the T2/T3 con-trast (dash lines). Data from sleeping and awake subgroups are depicted inthe left and right panels, respectively.
Discussion
Experiment 2 entailed investigating the development of mismatch responses to Mandarin tonalchanges in early infancy and how deviance size affects these responses. The same set of stimuliand paradigm, which have successfully educed typical MMNs to both large (T1/T3 pair) andsmall (T2/T3 pair) deviants in adults, were applied in a longitudinal study of infants at birth andat 6 months of age. The infant data showed that the more discriminable T1/T3 pair elicited aleft frontal-distributed P-MMR in 300 msec to 500 msec at birth, whereas it elicited an adult-like MMN bilaterally preceding a left-central P-MMR in 300 to 450 msec at 6 months of age.This suggests that MMRs to the T1/T3 pair transit from P-MMR to MMN before 6 months of

294 CHENG ET AL.
age, which is consistent with the developmental trajectory of MMRs to pure tone (Kushnerenkoet al., 2002). The transition of polarity between newborns and 6-month-olds demonstrated thematuration effect on the polarity of the infants’ mismatch response to pitch contour change inlinguistic context. The MMRs to T2/T3 pair, however, had a developmental trajectory that wasdifferent from the MMRs for T1/T3. The T2/T3 pair elicited no significant MMRs in newbornsand a left central-distributed P-MMR in 6-month-old infants. Congruent with Lee et al. (2012),the data of infants at 6 months of age demonstrated the co-existence of MMN and P-MMR andsuggested that the developmental trajectory of MMR polarity transition can be explained not onlyby citing the maturational factor, but also by referring to the stimuli-related factors (such as thediscriminability of contrast and the size of deviance).
Studies using non-speech pitch change have suggested that the polarity of mismatch responseis dependent on both maturation and stimulus-related factors (He, Hotson, & Trainor, 2009a,2009b). The younger infants would show P-MMR to the changes of lexical tones (especiallyfor the small deviant T2/T3) because of the insufficiently detailed representations to supportautomatic, pre-attentive discrimination. With increasing age, the negative MMR (presumably theemerging MMN) are expected because more children will have developed robust representations.This is especially true for the larger deviant (T1/T3) with relatively high discriminability. Thelarger deviant elicited P-MMR in newborns, and the adult-like MMN became apparent in awakeinfants at 6 months of age. However, the P-MMR remains stable at a time window of approxi-mately 300 msec to 450 msec, from birth to 6 months of age. For the small deviant (T2/T3), theMMR was not presented until 6-months of age. A P-MMR was found in the left central site in 6-month-old infants while they were awake. The transition from P-MMR to MMR seems not only tobe affected by maturation, but also by the deviance size. Furthermore, a stable mismatch responseresembling adult’s MMN is already present early in the development for the larger change in lex-ical tone. The presence of MMN implies that the developed phonological representations in thebrain support an automatic, pre-attentive discrimination. Earlier maturation of discrimination forT1/T3 than that for T2/T3 mirrors the observations of speech acquisition (Tsao, 2008; Wonget al., 2005).
The absence of MMN in newborns was expected because previous studies showed more posi-tive responses to the deviant stimuli, especially in infants under one year of age (He et al., 2007;Kushnerenko et al., 2002; Leppänen et al., 1997; Trainor, Samuel, Desjardins, & Sonnadara,2001; Trainor et al., 2003). For example, Leppänen et al. (1997) observed a P-MMR peakingbetween 250 msec to 350 msec to the pure tonal change in newborns. The studies series byHe et al. demonstrated that MMN became robust in 4-month-old infants, whereas only a broadpositive mismatch response was present in 2-month-olds (He et al., 2007, 2009a, 2009b).
The present findings show that the large deviant T1/T3 elicited P-MMR in sleeping newborns,and the transition from P-MMR to adult-like MMN was evident in waking infants at 6 monthsof age. These results are in agreement with the findings by Moore and colleagues (Moore, 2002;Moore & Linthicum, 2007), which suggest that the neural regions (cortical layers) that are thelikely sources of the adult MMN are insufficiently mature before 3 months of age. The immatureauditory cortices are considered the source of infant MMRs for perinatal infants.
Not only age but also the stimuli-related factors (such as the discriminability of contrast) affectwhen and how the transition from P-MMR to MMN occurs. For example, our data showed thatthe smaller deviant did not elicit any MMR in newborns but a P-MMR in infants at 6 monthsof age. This suggested that MMN could not be found consistently to fine-grained differences in
TAB
LE1
Sum
mar
yof
Pla
nned
Com
paris
ons
inE
ach
Tim
eW
indo
wat
Ele
ctro
des
F3,
F4,
C3,
and
C4,
Inde
pend
ently
new
born
s6-
mon
th-o
lds
(aw
ake)
adul
tsne
wbo
rns
6-m
onth
-old
s(a
wak
e)ad
ults
T1/
T3
T2/
T3
T1/
T3
T2/
T3
T1/
T3
T2/
T3
Tim
ew
indo
ws
T1/
T3
T2/
T3
T1/
T3
T2/
T3
T1/
T3
T2/
T3
50–1
00N
∗∗∗
100–
150
N∗∗
∗N
∗N
∗∗∗
N∗∗
∗15
0–20
0N
∗∗N
∗∗∗
N∗∗
∗N
∗N
∗∗∗
200–
250
N∗
N∗∗
∗P∗
N∗∗
∗F3
250–
300
F4N
∗∗∗
P∗30
0–35
0P∗
350–
400
P∗––
––40
0–45
0––
––––
––45
0–50
0––
––
50–1
00N
∗∗∗
100–
150
N∗∗
∗N
∗∗∗
N∗∗
∗15
0–20
0N
∗∗∗
N∗∗
∗N
∗∗∗
200–
250
N∗∗
∗N
∗∗∗
C3
250–
300
C4
N∗∗
∗P∗
P∗30
0–35
0P∗
∗∗P∗
350–
400
P∗∗
––––
400–
450
––––
––––
450–
500
––––
Not
e.“P
”de
note
sth
atth
ede
vian
ceis
sign
ifica
ntly
mor
epo
sitiv
eth
anth
est
anda
rd,w
here
as“N
”de
note
sth
ede
vian
ceis
mor
ene
gativ
eth
anth
est
anda
rd.
∗ p<
.05.
∗∗p
<.0
05.∗
∗∗p
<.0
01.
295

296 CHENG ET AL.
awake infants and are absent in sleeping newborns. This is supported by Morr et al. (2002), whoreported a P-MMR to the pitch change between 1,000 Hz and 1,200 Hz but an adult-like MMN forthe change between 1,000 Hz and 2,000 Hz in infants within 1 year of age. The P-MMR remainedin older children when fine-grained deviances were used. Maurer et al. (2003) reported P-MMRto a substantially smaller frequency deviance (100 Hz vs. 1,060 Hz or 1,030 Hz) and phonemedeviance (/ba/ vs. /ta/ or /da/) in 6- to 7-year-old children. The present findings of adult-likeMMN for T1/T3 and P-MMR to T2/T3 in 6-month-old infants supported that the degree ofdeviance on lexical tone could modulate the polarity of MMRs. The T2/T3 contrast does notsuffice in salience for 6-month-old infants to elicit an MMN for automatic change detection. Thisis also congruent with the behavioral observation that showed the difficulty in discriminatingbetween T2 and T3 remaining, even at three years of age (Wong et al., 2005). Further studies arerequired to follow the developmental trajectory of the polarity transition in MMRs to T2/T3.
Data from awake 6-month-old infants revealed the coexistence of MMN and P-MMR.However, the adult-like MMN and P-MMR have distinct scalp distributions. The adult-like MMNin 6-month-old infants were bilaterally significant in the frontal electrodes, whereas the P-MMRsin newborns and 6-month-old infants were significant only in left electrodes. These findings sug-gest that the adult-like MMN and P-MMR may reflect different characteristics of processing.Because the MMN usually indexes the automaticity of auditory change detection, the presence ofP-MMR seemingly reflects an immature brain response to auditory changes if it is more likely tobe observed in children or infants at a younger age and in response to small deviance, whichis difficult to discriminate. However, the functional significance of P-MMR remains unclear.A number of studies suggest that the P-MMR might be an analogue of adults’ P3a response,which reflects an involuntary attention switching to a task-irrelevant novel sound (He, Hotson, &Trainor, 2008; Kushnerenko et al., 2002; Shestakova et al., 2003). However, Kushnerenko et al.(2007) examined whether the P3a-like positivity in newborns could be attributed to novelty assuch, or to acoustic characteristics of novel sounds by using various types of novel sounds, suchas novel environmental sounds, white-noise segments, or harmonic tones with a higher pitch orwith higher intensity. They found that newborns are most sensitive to the spectral width of soundsbecause the largest and most reliable P3a was found in response to infrequent white noise seg-ments than to other novel stimuli. This phenomenon was not found in adults, and suggested thatthe P3a-like positivity in infants might be attributed to the spectral richness rather than in responseto novelty as such. Thus, the P-MMR of infants may not be identified with the P3a response seenin adults.
A number of studies have suggested that P3a elicitation is dependent on adequate changedetection, which was indexed by the preceding MMN (Escera, Alho, Winkler, & Näätänen,1998; Friedman, Cycowicz, & Gaeta, 2001). However, no preceding MMN was found in theP-MMRs elicited by T1/T3 in newborns or by T2/T3 in 6-month-old infants. This is in line withthe study conducted by Shafer et al. (2011), which reported a P-MMR to English vowel contrastswithout preceding MMN from 6 to 18 months of age. The P-MMR may not merely reflectthe involuntary attention shifting as the P3a, but also indexes a preliminary change detectionmechanism in infancy.
Our newborn data showed a P-MMR to large deviant (T1/T3), but no significant MMR tosmall deviant (T2/T3). When infants turned into 6-month-old, the MMRs could be detected onlywhen they were awake, but no MMRs could be detected when they were sleeping. The dataimply that alertness affects the presence of MMRs in early infancy. However, it is unclear why

MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 297
MMRs could be detected in sleeping newborns but not in 6-month-old infants when they werealso sleeping.
One possible reason is that the MMN-generating system may be affected in differently innewborns and in infants of 6-month-olds during sleep. Many studies have successfully demon-strated MMN in sleeping newborns (Cheour et al., 2002; Leppänen et al., 2004; Martynovaet al., 2003). However, for adults and older infants, MMN has been detected mainly in statesof wakefulness (Atienza, Cantero, & Dominguez-Marin, 2002; Friederici et al., 2002). Studieshave roughly divided the sleep state into different stages (such as REM (rapid eye movement)and non-REM (NREM) sleep) and demonstrated the adult MMN during REM sleep but not inNREM sleep (Atienza et al., 2002; Nashida et al., 2000). However, the MMN amplitude obtainedin REM sleep is considerably more attenuated as compared with that obtained during wakeful-ness (Atienza et al., 2002). By contrast, MMN in newborns can be recorded during wakefulness,and quiet (equivalent to NREM sleep in adults) and active (equivalent to REM sleep in adults)sleep, and its amplitude does not vary across these states (Cheour, Leppänen, & Kraus, 2000;Martynova et al., 2003). This implies that active (REM) and quiet (NREM) sleep in newbornsmay serve different functional processes as compared with the respective sleep stages in adults.
The maturation of the neural structure, as indicated by fewer neurons, sparser dendritic branch-ing, fewer axon collaterals, and incomplete myelination of axons in primary and secondarycortical regions, may also contribute to the morphological difference between developmentalgroups. Previous studies have suggested that neonates are not capable of blocking or inhibitingafferent or efferent systems during sleep because of an immature thalamic inhibitory system ininfants (Winkler, 2007). The thalamus is incapable of efficiently inhibiting information duringsleeping before the second or third month of life. Therefore, it is reasonable to expect MMRsare elicited in different states of infant sleep, indicating that newborns are able to process andlearn auditory input while sleeping (Weber, Hahne, Friedrich, & Friederici, 2004). It would bemuch more relevant to use MMRs to determine whether infants can develop sufficient ability todiscriminate a set of speech units at certain ages, than to examine the morphological differences(such as MMR latency and amplitude) across different age groups.
CONCLUSION
This study demonstrated how maturation and stimulus-dependent factors affect the mismatchresponses to the Mandarin lexical tone in adults and early infancy. In adults, typical MMN wereobtained for both the T1/T3 pair and the T2/T3 pair. The acoustically distinct T1/T3 pair elicitedan earlier and larger MMN than the acoustically similar T2/T3 did. In the infants, T1/T3 pairelicited a left frontal-distributed P-MMR in the newborns, whereas it elicited an adult-like MMNin the 6-month-old infants. However, the T2/T3 did not elicit any MMRs in the newborns butelicited a P-MMR in the 6-month-old infants only when they were awake. Apparently, the P-MMR surfaced in newborns when the change sufficed in salience. The P-MMR switched tothe adult-like MMN with infants’ growth, but the trajectory of the polarity transition dependedon the degree of deviance. The coexistence of P-MMR and adult-like MMN suggested that thechange detection mechanism in 6-month-old infants may not be as automatic as those in adults.Additional studies are required to determine when the automatic change detection indexed byadult-like MMN for T2/T3 discrimination can be shown in later ages. In addition, more detailed

298 CHENG ET AL.
study of how responses to different sound features develop across ages in normal developingchildren would be crucial for early identification of children with language impairments.
REFERENCES
Ahmmed, A. U., Clarke, E. M., & Adams, C. (2008). Mismatch negativity and frequency representational width inchildren with specific language impairment. Developmental Medicine and Child Neurology, 50, 938–944.
Alho, K., Sainio, K., Sajaniemi, N., Reinikainen, K., & Naatanen, R. (1990). Event-related brain potential of humannewborns to pitch change of an acoustic stimulus. Electroencephalography and Clinical Neurophysiology, 77,151–155.
Atienza, M., Cantero, J. L., & Dominguez-Marin, E. (2002). Mismatch negativity (MMN): An objective measure ofsensory memory and long-lasting memories during sleep. International Journal of Psychophysiology, 46, 215–225.
Brannon, E. M., Libertus, M. E., Meck, W. H., & Woldorff, M. G. (2008). Electrophysiological measures of timeprocessing in infant and adult brains: Weber’s Law holds. Journal of Cognitive Neuroscience, 20, 193–203.
Brannon, E. M., Roussel, L. W., Meck, W. H., & Woldorff, M. (2004). Timing in the baby brain. Cognitive Brain Research,21, 227–233.
Chandrasekaran, B., Krishnan, A., & Gandour, J. T. (2007a). Mismatch negativity to pitch contours is influenced bylanguage experience. Brain Research, 1128, 148–156.
Chandrasekaran, B., Gandour, J. T., & Krishnan, A. (2007b). Neuroplasticity in the processing of pitch dimensions: Amultidimensional scaling analysis of the mismatch negativity. Restorative Neurology and Neuroscience, 25, 195–210.
Chandrasekaran, B., Krishnan, A., & Gandour, J. T. (2009). Sensory processing of linguistic pitch as reflected by themismatch negativity. Ear and Hearing, 30, 552–558.
Cheour, M., Leppänen, P. H., & Kraus, N. (2000). Mismatch negativity (MMN) as a tool for investigating auditorydiscrimination and sensory memory in infants and children. Clinical Neurophysiology, 111, 4–16.
Cheour, M., Kushnerenko, E., Ceponiene, R., Fellman, V., & Näätänen, R. (2002). Electric brain responses obtained fromnewborn infants to changes in duration in complex harmonic tones. Developmental Neuropsychology, 22, 471–479.
Cheour, M., Alho, K., Ceponiene, R., Reinikainen, K., Sainio, K., Pohjavuori, M., & Näätänen, R. (1998a). Maturationof mismatch negativity in infants. International Journal of Psychophysiology, 29, 217–226.
Cheour, M., Ceponiene, R., Lehtokoski, A., Luuk, A., Allik, J., Alho, K., & Näätänen, R. (1998b). Development oflanguage-specific phoneme representations in the infant brain. Nature Neuroscience, 1, 351–353.
Cheour, M., Ceponiene, R., Leppanen, P., Alho, K., Kujala, T., Renlund, M., . . . Naatanen, R. (2002). The auditorysensory memory trace decays rapidly in newborns. Scandinavian Journal of Psychology, 43, 33–39.
Cheour-Luhtanen, M., Alho, K., Kujala, T., Sainio, K., Reinikainen, K., Renlund, M., . . . Näätänen, R. (1995). Mismatchnegativity indicates vowel discrimination in newborns. Hearing Research, 82, 53–58.
Clumeck, H. (1980). The acquisition of tone. In G. H. Yeni-Komshian, J. F. Kavanagh, & C. A. Ferguson (Eds.), Childphonology (Vol. 1, pp. 257–275). New York, NY: Academic Press.
Dehaene-Lambertz, G., & Baillet, S. (1998). A phonological representation in the infant brain. Neuroreport, 9,1885–1888.
Dehaene-Lambertz, G., & Dehaene, S. (1994). Speed and cerebral correlates of syllable discrimination in infants. Nature,370, 292–295.
Escera, C., Alho, K., Winkler, I., & Näätänen, R. (1998). Neural mechanisms of involuntary attention to acoustic noveltyand change. Journal of Cognitive Neuroscience, 10, 590–604.
Friederici, A. D., Friedrich, M., & Christophe, A. (2007). Brain responses in 4-month-old infants are already languagespecific. Current Biology, 17, 1208–1211.
Friederici, A. D., Friedrich, M., & Weber, C. (2002). Neural manifestation of cognitive and precognitive mismatchdetection in early infancy. Neuroreport, 13, 1251–1254.
Friedman, D., Cycowicz, Y. M., & Gaeta, H. (2001). The novelty P3: An event-related brain potential (ERP) sign of thebrain’s evaluation of novelty. Neuroscience and Biobehavioral Reviews, 25, 355–373.
Gandour, J. (1983). Tone perception in far eastern-languages. Journal of Phonetics, 11, 149–175.Gandour, J. T., & Harshman, R. A. (1978). Crosslanguage differences in tone perception: A multidimensional scaling
investigation. Language and Speech, 21, 1–33.

MISMATCH RESPONSE TO LEXICAL TONES IN INFANCY 299
Guttorm, T. K., Leppänen, P. H., Hamalainen, J. A., Eklund, K. M., & Lyytinen, H. J. (2010). Newborn event-relatedpotentials predict poorer pre-reading skills in children at risk for dyslexia. Journal of Learning Disabilities, 43,391–401.
Guttorm, T. K., Leppänen, P. H., Poikkeus, A. M., Eklund, K. M., Lyytinen, P., & Lyytinen, H. (2005). Brain event-relatedpotentials (ERPs) measured at birth predict later language development in children with and without familial risk fordyslexia. Cortex, 41, 291–303.
He, C., Hotson, L., & Trainor, L. J. (2007). Mismatch responses to pitch changes in early infancy. Journal of CognitiveNeuroscience, 19, 878–892.
He, C., Hotson, L., & Trainor, L. J. (2008). Maturation of cortical mismatch responses to occasional pitch change in earlyinfancy: Effects of presentation rate and magnitude of change. Neuropsychologia, 47, 218–229.
He, C., Hotson, L., & Trainor, L. J. (2009a). Development of infant mismatch responses to auditory pattern changesbetween 2 and 4 months old. European Journal of Neuroscience, 29, 861–867.
He, C., Hotson, L., & Trainor, L. J. (2009b). Maturation of cortical mismatch responses to occasional pitch change inearly infancy: Effects of presentation rate and magnitude of change. Neuropsychologia, 47, 218–229.
Hua, Z. (2002). Phonological development in specific contexts: Studies of Chinese-speaking children. Clevedon, England:Wayside Book.
Hua, Z., & Dodd, B. (2000). The phonological acquisition of Putonghua (Modern Standard Chinese). Journal of ChildLanguage, 27, 3–42.
Huang, T., & Johnson, K. (2010). Language specificity in speech perception: Perception of Mandarin tones by native andnonnative listeners. Phonetica, 67, 243–267.
Jing, H., & Benasich, A. A. (2006). Brain responses to tonal changes in the first two years of life. Brain and Development,28, 247–256.
Kushnerenko, E., Ceponiene, R., Balan, P., Fellman, V., & Näätänen, R. (2002). Maturation of the auditory changedetection response in infants: A longitudinal ERP study. Neuroreport, 13, 1843–1848.
Kushnerenko, E., Cheour, M., Ceponiene, R., Fellman, V., Renlund, M., Soininen, K., . . . Näätänen, R. (2001). Centralauditory processing of durational changes in complex speech patterns by newborns: An event-related brain potentialstudy. Developmental Neuropsychology, 19, 83–97.
Kushnerenko, E., Winkler, I., Horvath, J., Näätänen, R., Pavlov, I., Fellman, V., & Huotilainen, M. (2007). Processingacoustic change and novelty in newborn infants. European Journal of Neuroscience, 26, 265–274.
Lee, C.-Y., Yen, H.-L., Yeh, P.-W., Lin, W.-H., Cheng, Y.-Y., Tzeng, Y.-L., & Wu, H.-C. (2012). Mismatch responses tolexical tone, initial consonant, and vowel in Mandarin-speaking preschoolers. Neuropsychologia, 50, 3228–3239.
Leppänen, P. H., Eklund, K. M., & Lyytinen, H. (1997). Event-related brain potentials to change in rapidly presentedacoustic stimuli in newborns. Developmental Neuropsychology, 13, 175–204.
Leppänen, P. H., Guttorm, T. K., Pihko, E., Takkinen, S., Eklund, K. M., & Lyytinen, H. (2004). Maturational effects onnewborn ERPs measured in the mismatch negativity paradigm. Experimental Neurology, 190(Suppl 1), S91–S101.
Leppänen, P. H., Hamalainen, J. A., Salminen, H. K., Eklund, K. M., Guttorm, T. K., Lohvansuu, K., . . . Lyytinen, H.(2010). Newborn brain event-related potentials revealing atypical processing of sound frequency and the subsequentassociation with later literacy skills in children with familial dyslexia. Cortex, 46, 1362–1376.
Li, C. N., & Tompson, S. A. (1977). The acquisition of tone in Mandarin-speaking children. Journal of Child Language,4, 185–199.
Luo, H., Ni, J. T., Li, Z. H., Li, X. O., Zhang, D. R., Zeng, F. G., & Chen, L. (2006). Opposite patterns of hemispheredominance for early auditory processing of lexical tones and consonants. Proceedings of the National Academy ofSciences of the United States of America, 103, 19558–19563.
Martynova, O., Kirjavainen, J., & Cheour, M. (2003). Mismatch negativity and late discriminative negativity in sleepinghuman newborns. Neuroscience Letters, 340, 75–78.
Maurer, U., Bucher, K., Brem, S., & Brandeis, D. (2003). Development of the automatic mismatch response: From frontalpositivity in kindergarten children to the mismatch negativity. Clinical Neurophysiology, 114, 808–817.
McBride-Chang, C., Tong, X., Shu, H., Wong, A. M. Y., Leung, K. W., & Tardif, T. (2008). Syllable, phoneme, and tone:Psycholinguistic units in early Chinese and English word recognition. Scientific Studies of Reading, 12, 171–194.
Meng, X., Sai, X., Wang, C., Wang, J., Sha, S., & Zhou, X. (2005). Auditory and speech processing and readingdevelopment in Chinese school children: Behavioural and ERP evidence. Dyslexia, 11, 292–310.
Molfese, D. L. (2000). Predicting dyslexia at 8 years of age using neonatal brain responses. Brain and Language, 72,238–245.

300 CHENG ET AL.
Molfese, D. L., & Molfese, V. J. (1985). Electrophysiological indices of auditory discrimination in newborn infants: Thebases for predicting later language development? Infant Behavior and Development, 8, 197–211.
Molfese, D. L., & Molfese, V. J. (1997). Discrimination of language skills at five years of age using event-relatedpotentials recorded at birth. Devlopmental Neuropsychology, 13, 135–156.
Moore, J. K. (2002). Maturation of human auditory cortex: Implications for speech perception. Annals of Otology,Rhinology, and Laryngology. Supplement, 189, 7–10.
Moore, J. K., & Linthicum, F. H., Jr. (2007). The human auditory system: A timeline of development. InternationalJournal of Audiology, 46, 460–478.
Morr, M. L., Shafer, V. L., Kreuzer, J. A., & Kurtzberg, D. (2002). Maturation of mismatch negativity in typicallydeveloping infants and preschool children. Ear and Hearing, 23, 118–136.
Näätänen, R., Kujala, T., & Winkler, I. (2011). Auditory processing that leads to conscious perception: A unique windowto central auditory processing opened by the mismatch negativity and related responses. Psychophysiology, 48, 4–22.
Naatanen, R., Lehtokoski, A., Lennes, M., Cheour, M., Huotilainen, M., Iivonen, A., . . . Alho, K. (1997). Language-specific phoneme representations revealed by electric and magnetic brain responses. Nature, 385, 432–434.
Näätänen, R., Paavilainen, P., Rinne, T., & Alho, K. (2007). The mismatch negativity (MMN) in basic research of centralauditory processing: A review. Clinical Neurophysiology, 118, 2544–2590.
Näätänen, R., Pakarinen, S., Rinne, T., & Takegata, R. (2004). The mismatch negativity (MMN): Towards the optimalparadigm. Clinical Neurophysiology, 115, 140–144.
Nashida, T., Yabe, H., Sato, Y., Hiruma, T., Sutoh, T., Shinozaki, N., & Kaneko, S. (2000). Automatic auditory informationprocessing in sleep. Sleep, 23, 821–828.
Novitski, N., Huotilainen, M., Tervaniemi, M., Näätänen, R., & Fellman, V. (2007). Neonatal frequency discriminationin 250-4000-Hz range: Electrophysiological evidence. Clinical Neurophysiology, 118, 412–419.
Pulvermuller, F., Kujala, T., Shtyrov, Y., Simola, J., Tiitinen, H., Alku, P., . . . Naatanen, R. (2001). Memory traces forwords as revealed by the mismatch negativity. Neuroimage, 14, 607–616.
Pulvermuller, F., Shtyrov, Y., Kujala, T., & Naatanen, R. (2004). Word-specific cortical activity as revealed by themismatch negativity. Psychophysiology, 41, 106–112.
Ren, G. Q., Yang, Y., & Li, X. (2009). Early cortical processing of linguistic pitch patterns as revealed by the mismatchnegativity. Neuroscience, 162, 87–95.
Shafer, V. L., Yu, Y. H., & Datta, H. (2011). The development of English vowel perception in monolingual and bilingualinfants: Neurophysiological correlates. Journal of Phonetics, 39, 527–545.
Shestakova, A., Huotilainen, M., Ceponiene, R., & Cheour, M. (2003). Event-related potentials associated with secondlanguage learning in children. Clinical Neurophysiology, 114, 1507–1512.
Trainor, L., McFadden, M., Hodgson, L., Darragh, L., Barlow, J., Matsos, L., & Sonnadara, R. (2003). Changes inauditory cortex and the development of mismatch negativity between 2 and 6 months of age. International Journal ofPsychophysiology, 51, 5–15.
Trainor, L. J., Samuel, S. S., Desjardins, R. N., & Sonnadara, R. R. (2001). Measuring temporal resolution in infantsusing mismatch negativity. Neuroreport, 12, 2443–2448.
Tsao, F. M. (2008). The effect of acoustic similarity on lexical-tone perception of one-year-old Mandarin-learning infants.Chinese Journal of Psychology, 50, 111–124.
Weber, C., Hahne, A., Friedrich, M., & Friederici, A. D. (2004). Discrimination of word stress in early infant perception:Electrophysiological evidence. Cognitive Brain Research, 18, 149–161.
Weber, C., Hahne, A., Friedrich, M., & Friederici, A. D. (2005). Reduced stress pattern discrimination in 5-month-olds asa marker of risk for later language impairment: Neurophysiologial evidence. Cognitive Brain Research, 25, 180–187.
Winkler, I. (2007). Interpreting the mismatch negativity. Journal of Psychophysiology, 21, 147–163.Wong, P., Schwartz, R. G., & Jenkins, J. J. (2005). Perception and production of lexical tones by 3-year-old Mandarin-
speaking children. Journal of Speech, Language, and Hearing Research, 48, 1065–1079.Xi, J., Zhang, L., Shu, H., Zhang, Y., & Li, P. (2010). Categorical perception of lexical tones in Chinese revealed by
mismatch negativity. Neuroscience, 170, 223–231.
Related Documents











