The Desmoglein-Specific Cytoplasmic Region Is Intrinsically Disordered in Solution and Interacts with Multiple Desmosomal Protein Partners Keiichiro Kami 1 , Martyn Chidgey 1 , Timothy Dafforn 2 and Michael Overduin 1 ⁎ 1 School of Cancer Sciences, University of Birmingham, Birmingham B15 2TT, UK 2 School of Biosciences, University of Birmingham, Birmingham B15 2TT, UK Received 3 November 2008; received in revised form 18 December 2008; accepted 19 December 2008 Available online 30 December 2008 The desmoglein-specific cytoplasmic region (DSCR) is a conserved region of unknown structure and function that uniquely defines the desmoglein family of cell adhesion molecules. It is the site of caspase cleavage during apoptosis, and its mutation is linked to cardiomyopathy. Here, we reveal that a 276-residue DSCR construct of human desmoglein 1 is intrinsically disordered and forms an interaction hub for desmosomal proteins. In solution, it contains 6.5% helical and 10.3% β-strand structure based on circular dichroism spectroscopy. A single monomeric state with a predominantly unfolded structure is found by size-exclusion chromatogra- phy and analytical ultracentrifugation. Thermal stability assays and nuclear magnetic resonance spectroscopy reveal a nonglobular structure under a range of solution conditions. However, the introduction of detergent micelles increases structure to 18% helical and 16% β-strand character, suggesting an inducible structure. The DSCR exhibits weak but specific interactions with plakoglobin, the plakin domain of desmoplakin, plako- philin 1, and the cytoplasmic domain of desmocollin 1. The desmoglein 1 membrane proximal region also interacts with all four DSCR ligands, strongly with plakoglobin and plakophilin and more weakly with des- moplakin and desmocollin 1. Thus, the DSCR is an intrinsically disordered functional domain with an inducible structure that, along with the mem- brane proximal region, forms a flexible scaffold for cytoplasmic assembly at the desmosome. © 2009 Elsevier Ltd. All rights reserved. Edited by P. Wright Keywords: desmoglein; desmosome; cadherin; intrinsically disordered protein; membrane proximal region Introduction Desmosomal cadherins are cell adhesion molecules found in desmosomes, which are strong intercellular junctions that connect epithelial and cardiac muscle cells. The two major subtypes, the desmogleins and desmocollins, are distinguished by unique cytoplas- mic regions. Humans express four desmogleins, Dsg1, Dsg2, Dsg3, and Dsg4, 1,2 which exhibit tissue- specific patterns of expression, with Dsg1 largely restricted to suprabasal cell layers of the epidermis. 3 All desmogleins share a similar architecture. A set of extracellular cadherin domains (ECs; EC1–EC4) mediate cell–cell adhesion in the intercellular space. These domains are linked via an extracellular anchor and membrane-spanning domain to the intracellular anchor (IA) and intracellular cadherin-typical se- quence (ICS) motifs, which are also found in desmo- collins and classical cadherins. Together, the IA and ICS domains constitute the membrane proximal region (MPR) of Dsg1 (Fig. 1a). In addition, all des- mogleins contain a unique, conserved motif, the *Corresponding author. E-mail address: [email protected]. Abbreviations used: DSCR, desmoglein-specific cytoplasmic region; EC, extracellular cadherin domain; IA, intracellular anchor; ICS, intracellular cadherin-typical sequence; MPR, membrane proximal region; PL, proline-rich linker; RUD, repeated unit domain; DTD, desmoglein-specific terminal domain; PKC, protein kinase C; PG, plakoglobin; DP, desmoplakin; PKP1, plakophilin 1; Dsc1, desmocollin 1; GST, glutathione S-transferase; AUC, analytical ultracentrifugation; TFE, trifluoroethanol; TCEP, Tris(2-chloroethyl) phosphate. doi:10.1016/j.jmb.2008.12.054 J. Mol. Biol. (2009) 386, 531–543 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2009 Elsevier Ltd. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1016/j.jmb.2008.12.054 J. Mol. Biol. (2009) 386, 531–543
Available online at www.sciencedirect.com
The Desmoglein-Specific Cytoplasmic Region IsIntrinsically Disordered in Solution and Interacts withMultiple Desmosomal Protein Partners
Keiichiro Kami1, Martyn Chidgey1, Timothy Dafforn2
and Michael Overduin1⁎
1School of Cancer Sciences,University of Birmingham,Birmingham B15 2TT, UK2School of Biosciences,University of Birmingham,Birmingham B15 2TT, UKReceived 3 November 2008;received in revised form18 December 2008;accepted 19 December 2008Available online30 December 2008
*Corresponding author. E-mail [email protected] used: DSCR, desm
cytoplasmic region; EC, extracellularintracellular anchor; ICS, intracellulasequence; MPR, membrane proximaproline-rich linker; RUD, repeated udesmoglein-specific terminal domainC; PG, plakoglobin; DP, desmoplaki1; Dsc1, desmocollin 1; GST, glutathAUC, analytical ultracentrifugation;TCEP, Tris(2-chloroethyl) phosphate
0022-2836/$ - see front matter © 2009 E
The desmoglein-specific cytoplasmic region (DSCR) is a conserved region ofunknown structure and function that uniquely defines the desmogleinfamily of cell adhesion molecules. It is the site of caspase cleavage duringapoptosis, and its mutation is linked to cardiomyopathy. Here, we revealthat a 276-residue DSCR construct of human desmoglein 1 is intrinsicallydisordered and forms an interaction hub for desmosomal proteins. Insolution, it contains 6.5% helical and 10.3% β-strand structure based oncircular dichroism spectroscopy. A single monomeric state with apredominantly unfolded structure is found by size-exclusion chromatogra-phy and analytical ultracentrifugation. Thermal stability assays and nuclearmagnetic resonance spectroscopy reveal a nonglobular structure under arange of solution conditions. However, the introduction of detergentmicelles increases structure to 18% helical and 16% β-strand character,suggesting an inducible structure. The DSCR exhibits weak but specificinteractions with plakoglobin, the plakin domain of desmoplakin, plako-philin 1, and the cytoplasmic domain of desmocollin 1. The desmoglein 1membrane proximal region also interacts with all four DSCR ligands,strongly with plakoglobin and plakophilin and more weakly with des-moplakin and desmocollin 1. Thus, the DSCR is an intrinsically disorderedfunctional domain with an inducible structure that, along with the mem-brane proximal region, forms a flexible scaffold for cytoplasmic assembly atthe desmosome.
© 2009 Elsevier Ltd. All rights reserved.
Keywords: desmoglein; desmosome; cadherin; intrinsically disorderedprotein; membrane proximal region
Edited by P. WrightIntroduction
Desmosomal cadherins are cell adhesionmoleculesfound in desmosomes, which are strong intercellular
ess:
oglein-specificcadherin domain; IA,r cadherin-typicall region; PL,nit domain; DTD,; PKC, protein kinasen; PKP1, plakophilinione S-transferase;TFE, trifluoroethanol;.
lsevier Ltd. All rights reserve
junctions that connect epithelial and cardiac musclecells. The two major subtypes, the desmogleins anddesmocollins, are distinguished by unique cytoplas-mic regions. Humans express four desmogleins,Dsg1, Dsg2, Dsg3, and Dsg4,1,2 which exhibit tissue-specific patterns of expression, with Dsg1 largelyrestricted to suprabasal cell layers of the epidermis.3All desmogleins share a similar architecture. A set
of extracellular cadherin domains (ECs; EC1–EC4)mediate cell–cell adhesion in the intercellular space.These domains are linked via an extracellular anchorandmembrane-spanning domain to the intracellularanchor (IA) and intracellular cadherin-typical se-quence (ICS) motifs, which are also found in desmo-collins and classical cadherins. Together, the IA andICS domains constitute the membrane proximalregion (MPR) of Dsg1 (Fig. 1a). In addition, all des-mogleins contain a unique, conserved motif, the
d.
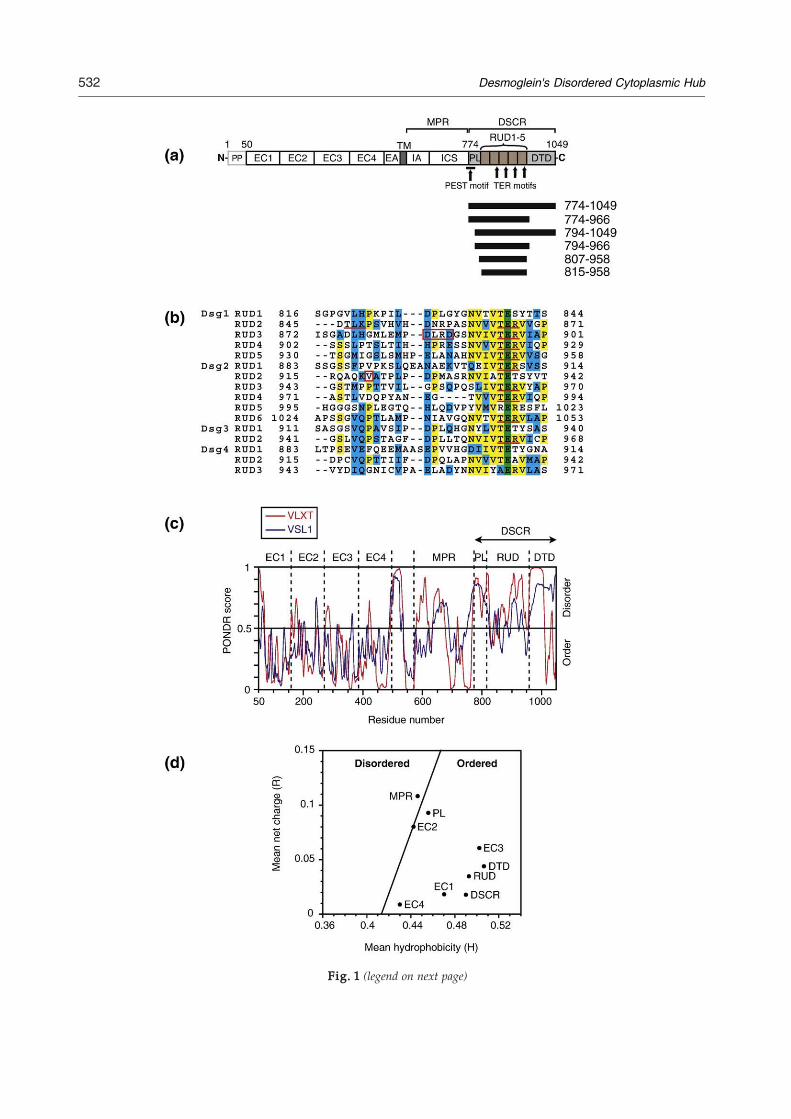
Fig. 1 (legend on next page)
532 Desmoglein's Disordered Cytoplasmic Hub

533Desmoglein's Disordered Cytoplasmic Hub
desmoglein-specific cytoplasmic region (DSCR). TheDSCR of desmogleins includes a proline-rich linker(PL), repeated unit domain (RUD), and a desmo-glein-specific terminal domain (DTD). The numberof repeats within the RUD varies from one desmo-glein to another; there are 5, 6, 2, and 3 in Dsg1,Dsg2, Dsg3, and Dsg4, respectively (Fig. 1b). TheRUD conformation remains unsolved and is notsimilar to known folds. However, a Dsg1 fragmentencompassing the DSCR appears to form multimerswith globular heads and thin tails when analyzed byelectron microscopy.5 The role of the conservedRUD sequences is not known, although consensussites for protein kinase C (PKC) phosphorylation6
and a caspase-3 cleavage site7 have been identifiedand could contribute to signaling and dismantling ofdesmosomes during apoptosis.Desmogleins are at the heart of the desmosomal
complex. The ICS motif is known to interact with thearmadillo family member plakoglobin (PG).8–10 Indesmosomes, PG may act as a linker between thedesmosomal cadherins and desmoplakin (DP), amember of the plakin family of cytolinkers, which,in turn, interacts with intermediate filaments. How-ever, a PG-independent interaction between full-length Dsg1 and DP has also been reported.11 Otherarmadillo family members found in desmosomesinclude the plakophilins,12 which could form lateralinteractions between linear chains of desmosomalcadherin–PG–DP molecules.13 Interactions betweenthe desmoglein cytoplasmic tail and plakophilinshave been reported.13–17 Whether the DSCR and itsconstituent PL, RUD, and DTD regions have directroles in these interactions remains unknown.Desmogleins play a crucial role in cell adhesion
andmaintaining epidermal tissue integrity, with lossof desmoglein-mediated cell adhesion resulting indisintegration of skin and cardiacmuscle. Dsg1 is thetarget of autoantibodies in the skin blistering diseasepemphigus foliaceus and is specifically cleaved byexfoliative toxins in staphylococcal scalded skinsyndrome.18 Mutations in Dsg1 cause the epidermaldisease striate palmoplantar keratoderma.19,20 Lossof expression of DSG2 is associated with gastric
Fig. 1. Bioinformatics analysis of the Dsg1 sequence. (a)protein including leader and pro-peptides (PP) is shown. Thethrough EC4), an extracellular anchor (EA), transmembrane (Tmembrane proximal region (MPR), which contains intracellul(ICS) motifs, and the desmoglein-specific cytoplasmic regionunit domain (RUD), and a desmoglein-specific terminal dpreproprotein sequence. The position of the PESTmotif and pu(b) Alignment of all the RUD sequences found in humanhighlighted in green, identical residues in yellow, and similarDsg1 and V920 in RUD2 of Dsg2 (which is mutated to Gldysplasia) are boxed in red. The positions of the phosphorylusing ClustalWwith Gonnet weightings and shaded with Boxsorder and disorder regions in the entire sequence of the madisorder prediction values are plotted for each residue using thEC1–EC4 (residues 50–158, 159–270, 271–385, and 386–497 reand DTD (959–1049) domains are indicated. (d) The net chacharge (R) and mean hydrophobicity (H) values are plotted,window size of 5. The black line represents the consensus bodefined by the equation R=2.785×H−1.151.4
cancer,21 and its RUD and ECs are mutated inpatients with arrhythmogenic right ventricularcardiomyopathy/dysplasia,22 indicating that bothregions are essential for mediating cell adhesion indesmosomes.In this study, we report the first structural
characterization of the DSCR. Results from nuclearmagnetic resonance (NMR) and circular dichroism(CD) spectroscopy demonstrate that the DSCR ofhuman Dsg1 is intrinsically disordered in solution,having little intrinsically stable structure but withinducible conformations that can be stabilized by co-solvents and micelles. Analysis of its hydrodynamicbehavior demonstrates that the protein exists as amonomer and in an unfolded conformation ratherthan a globular form. This molecular state enables theprotein to interactwith other proteins, and pull-downassays show that the DSCR binds to a number ofdesmosomal proteins including PG,DP, plakophilin 1(PKP1), and desmocollin 1 (Dsc1). These proteins alsointeract with the Dsg1 MPR. We conclude that theDSCR forms an intrinsically disordered state insolution with inducible structure that, together withthe MPR, forms a hub for multiple protein interac-tions during desmosomal cell adhesion.
Results
Prediction of structured and disordered regionsin the DSCR
In order to predict structured and disorderedregions in the mature Dsg1 protein (amino acids 50–1049), we used the PONDR algorithms VL-XT23,24
and VSL1.25 As expected, both PONDR algorithmspredicted that the ECs are structured (Fig. 1c). TheMPR contains sequences of predicted disorder andorder, reflecting its role as a complex docking hubfor desmosomal proteins. The DSCR sequence ispredicted to be primarily disordered with theexception of the conserved C-termini of some RUDsequences according to VSL1, which discriminatesregular structure from disorder. The C-terminus of
Schematic representation of human Dsg1. The precursormature protein consists of four extracellular domains (EC1M) region, and cytoplasmic domain. The latter consists of aar anchor (IA) and intracellular cadherin-typical sequence(DSCR), consisting of a proline-rich linker (PL), repeatedomain (DTD). The residue numbering is based on thetative PKC phosphorylation sites (TERmotifs) is indicated.desmogleins. The absolutely conserved Glu residue isresidues in blue. The caspase-3 cleavage site in RUD3 ofy in arrhythmogenic right ventricular cardiomyopathy/ation sites are underlined. The alignment was performedhade in the BiologyWorkbench server. (c) Prediction of theture Dsg1 by the PONDR neural network program. Thee algorithms VL-XT (red) and VLS1 (blue). The positions ofspectively), MPR (570–772), PL (773–815), RUD (816–958),rge–hydrophobicity plot of Dsg1 domains. The mean nethaving been calculated using Kyte–Doolittle values and aundary between disordered and ordered proteins and is

534 Desmoglein's Disordered Cytoplasmic Hub
the DTD is predicted as ordered by VL-XT, which ismore sensitive to local propensity for order, such asprotein elements, which become ordered whenbound.Analysis of the net charge versus hydrophobicity
within the DSCR sequences predicts the presence ofordered elements (Fig. 1d). Independently foldedglobular proteins exhibit high mean net hydro-phocity,4 as evidenced by Dsg1's ECs, which possessimmunoglobulin-like folds.26 On the other hand,natively disordered proteins possess low meanhydrophobicity and high net charge, as exemplifiedby the E-cadherin cytoplasmic domain, whichbecomes structured when bound by partnerproteins.27,28 Order is predicted in several regionsof the DSCR, which lacks significant sequencesimilarity to any known structural domains.Hence, we initiated an experimental investigationof whether it contains independently folded, stable,or inducible structures.
Purification of the DSCR
A series of Dsg1 constructs encompassing the fiveRUD repeats and adjacent PL and DTD regions(Fig. 1a) were expressed as glutathione S-transfer-ase (GST) fusions in Escherichia coli in order toexperimentally identify structural domains withinthe DSCR. Of these, the construct spanning theentire human Dsg1 DSCR (residues 774–1049) wassoluble after cell lysis and readily purified (Fig. 2).A yield of 6.3 mg of purified DSCR protein per literof culture was obtained following GST tag cleavageand glutathione affinity and anion-exchange andcation-exchange chromatography, providing suffi-cient pure material for characterization of itsstructural and functional properties.
Fig. 2. Purification of the recombinant Dsg1 DSCRprotein. Samples were resolved on a 4–12% SDS-PAGEgel. S, supernatant following lysis of bacteria andcentrifugation; E, partially purified GST-tagged DSCR;D, partially purified DSCR following digestion of GST-DSCR by PreScission Protease; P1, partially purified DSCRafter anion-exchange chromatography; P2, fully purifiedDSCR after cation-exchange chromatography. The posi-tion of the 28-kDa DSCR is indicated.
Hydrodynamic properties of DSCR
The degree of compactness and the oligomericstate of the DSCR were investigated by size-exclusion chromatography and analytical ultracen-trifugation (AUC), respectively. The DSCR proteineluted as a single peak by gel filtration with anapparent molecular mass of 121 kDa based oncalibration against known molecular mass stan-dards (Fig. 3a). Given its predicted monomer size of28.6 kDa, this suggests the presence of either anunfolded conformation or a folded tetramer. Themonomeric state of the DSCR was confirmed bysedimentation equilibrium analyses carried out atthree different rotor speeds (30,000, 33,000, and36,000 rpm) and three different protein concentra-tions (116, 58, and 19 μM), yielding nine differentdata sets (Fig. 3b). The best fit from global analysis toall nine data sets indicated a molecular mass of27,398±309 Da, which is close to the theoreticalmonomeric value of 28.6 kDa. Based on its gel-filtration elution profile (Fig. 3a), the Stokes radius(Rs) of DSCR was estimated to be 41.8 Å.29 Thiscompares more favorably with the theoretical valueof a fully unfolded protein of 28.6 kDa (47.6 Å) thanthat of a globular monomer (24.6 Å). Together, theseresults indicate that the DSCR has the dimensions ofa largely unfolded monomer.
DSCR is intrinsically disordered
To detect the presence of the tertiary structure inthe DSCR, we measured one-dimensional protonNMR spectra in solution conditions that resemblethe physiological environment. In a stably foldedprotein, the 1H NMR signals are well dispersedbecause each proton group experiences a uniquechemical environment. In contrast, the NMR signalsof the DSCR were not well dispersed, with narrow,intense, and overlapped methyl proton resonancesat random-coil values of 0.8 to 1.0 ppm. The back-bone amide protons are almost entirely clusteredbetween 7.9 and 8.5 ppm, with only two downfieldpeaks discernable at 8.70 and 9.21 ppm. The DSCR'ssole Trp appeared disordered, exhibiting random-coil NɛH chemical shifts of 10.05 and 10.24 ppm,presumably due to the apparent cis–trans isomeriza-tion of its neighboring proline residues (Fig. 4).Together, this indicates a disordered region lackingin stable tertiary structure and is consistent with thelargely unfolded conformation of the DSCR basedon its hydrodynamic properties.We tested whether folded structure could be
induced in the purified DSCR by screening a widevariety of buffers and additives known to stabilizeother proteins. Thermal shift assays have beendeveloped for high-throughput ligand screeningand identification of optimal solution conditionsfor globular proteins.30 This method distinguishesfolded structures from unfolded states, whichinteract preferentially with hydrophobic fluorop-robes. The probe is quenched in aqueous solutionbut preferentially binds to the exposed hydrophobic
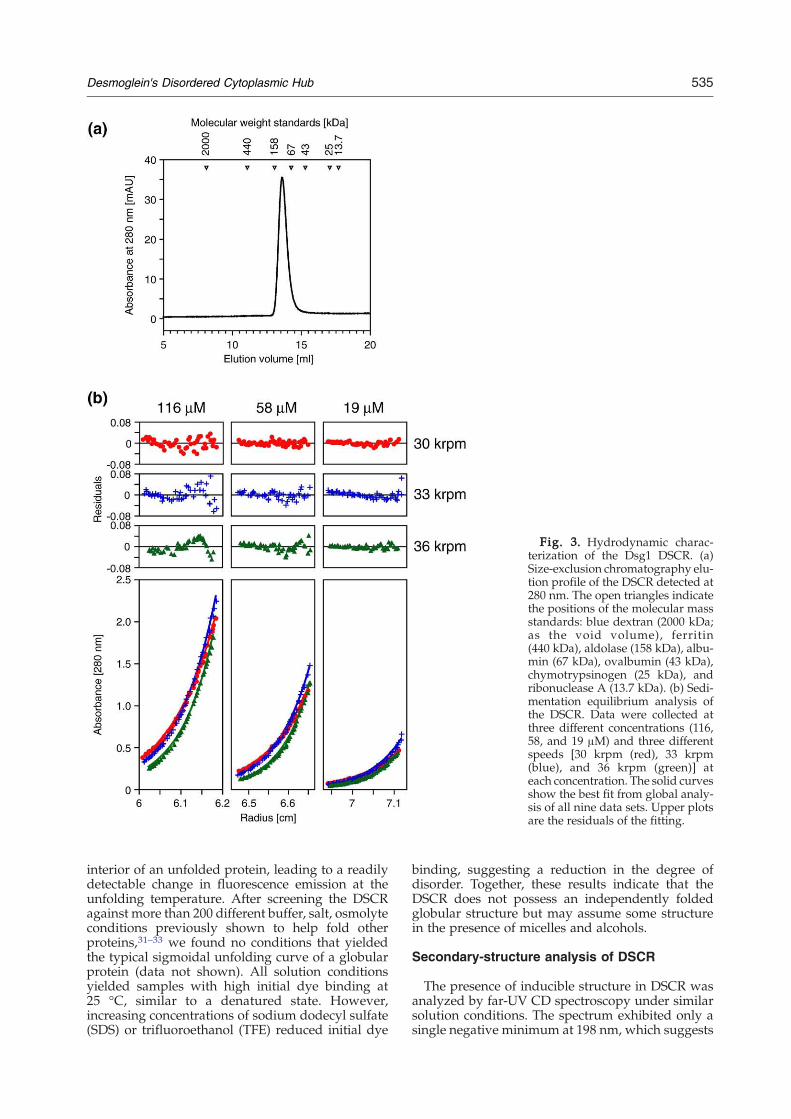
Fig. 3. Hydrodynamic charac-terization of the Dsg1 DSCR. (a)Size-exclusion chromatography elu-tion profile of the DSCR detected at280 nm. The open triangles indicatethe positions of the molecular massstandards: blue dextran (2000 kDa;as the void volume), ferritin(440 kDa), aldolase (158 kDa), albu-min (67 kDa), ovalbumin (43 kDa),chymotrypsinogen (25 kDa), andribonuclease A (13.7 kDa). (b) Sedi-mentation equilibrium analysis ofthe DSCR. Data were collected atthree different concentrations (116,58, and 19 μM) and three differentspeeds [30 krpm (red), 33 krpm(blue), and 36 krpm (green)] ateach concentration. The solid curvesshow the best fit from global analy-sis of all nine data sets. Upper plotsare the residuals of the fitting.
535Desmoglein's Disordered Cytoplasmic Hub
interior of an unfolded protein, leading to a readilydetectable change in fluorescence emission at theunfolding temperature. After screening the DSCRagainst more than 200 different buffer, salt, osmolyteconditions previously shown to help fold otherproteins,31–33 we found no conditions that yieldedthe typical sigmoidal unfolding curve of a globularprotein (data not shown). All solution conditionsyielded samples with high initial dye binding at25 °C, similar to a denatured state. However,increasing concentrations of sodium dodecyl sulfate(SDS) or trifluoroethanol (TFE) reduced initial dye
binding, suggesting a reduction in the degree ofdisorder. Together, these results indicate that theDSCR does not possess an independently foldedglobular structure but may assume some structurein the presence of micelles and alcohols.
Secondary-structure analysis of DSCR
The presence of inducible structure in DSCR wasanalyzed by far-UV CD spectroscopy under similarsolution conditions. The spectrum exhibited only asingle negative minimum at 198 nm, which suggests
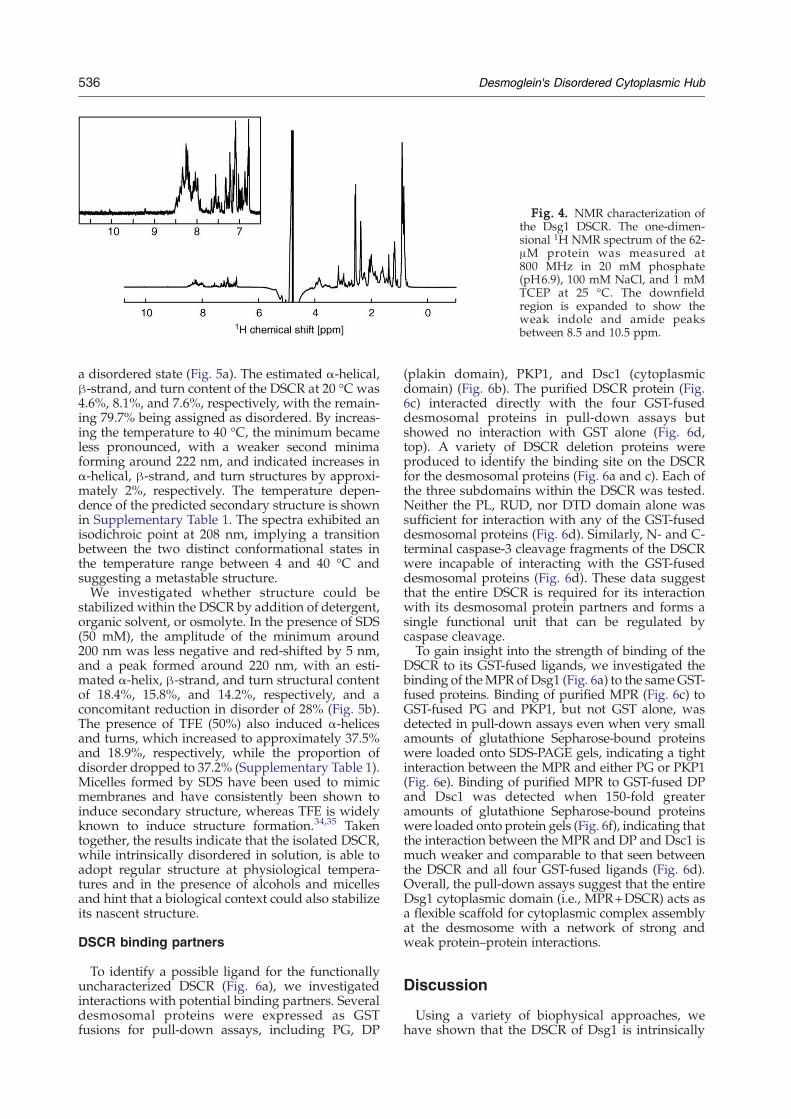
Fig. 4. NMR characterization ofthe Dsg1 DSCR. The one-dimen-sional 1H NMR spectrum of the 62-μM protein was measured at800 MHz in 20 mM phosphate(pH6.9), 100 mM NaCl, and 1 mMTCEP at 25 °C. The downfieldregion is expanded to show theweak indole and amide peaksbetween 8.5 and 10.5 ppm.
536 Desmoglein's Disordered Cytoplasmic Hub
a disordered state (Fig. 5a). The estimated α-helical,β-strand, and turn content of the DSCR at 20 °C was4.6%, 8.1%, and 7.6%, respectively, with the remain-ing 79.7% being assigned as disordered. By increas-ing the temperature to 40 °C, the minimum becameless pronounced, with a weaker second minimaforming around 222 nm, and indicated increases inα-helical, β-strand, and turn structures by approxi-mately 2%, respectively. The temperature depen-dence of the predicted secondary structure is shownin Supplementary Table 1. The spectra exhibited anisodichroic point at 208 nm, implying a transitionbetween the two distinct conformational states inthe temperature range between 4 and 40 °C andsuggesting a metastable structure.We investigated whether structure could be
stabilized within the DSCR by addition of detergent,organic solvent, or osmolyte. In the presence of SDS(50 mM), the amplitude of the minimum around200 nm was less negative and red-shifted by 5 nm,and a peak formed around 220 nm, with an esti-mated α-helix, β-strand, and turn structural contentof 18.4%, 15.8%, and 14.2%, respectively, and aconcomitant reduction in disorder of 28% (Fig. 5b).The presence of TFE (50%) also induced α-helicesand turns, which increased to approximately 37.5%and 18.9%, respectively, while the proportion ofdisorder dropped to 37.2% (Supplementary Table 1).Micelles formed by SDS have been used to mimicmembranes and have consistently been shown toinduce secondary structure, whereas TFE is widelyknown to induce structure formation.34,35 Takentogether, the results indicate that the isolated DSCR,while intrinsically disordered in solution, is able toadopt regular structure at physiological tempera-tures and in the presence of alcohols and micellesand hint that a biological context could also stabilizeits nascent structure.
DSCR binding partners
To identify a possible ligand for the functionallyuncharacterized DSCR (Fig. 6a), we investigatedinteractions with potential binding partners. Severaldesmosomal proteins were expressed as GSTfusions for pull-down assays, including PG, DP
(plakin domain), PKP1, and Dsc1 (cytoplasmicdomain) (Fig. 6b). The purified DSCR protein (Fig.6c) interacted directly with the four GST-fuseddesmosomal proteins in pull-down assays butshowed no interaction with GST alone (Fig. 6d,top). A variety of DSCR deletion proteins wereproduced to identify the binding site on the DSCRfor the desmosomal proteins (Fig. 6a and c). Each ofthe three subdomains within the DSCR was tested.Neither the PL, RUD, nor DTD domain alone wassufficient for interaction with any of the GST-fuseddesmosomal proteins (Fig. 6d). Similarly, N- and C-terminal caspase-3 cleavage fragments of the DSCRwere incapable of interacting with the GST-fuseddesmosomal proteins (Fig. 6d). These data suggestthat the entire DSCR is required for its interactionwith its desmosomal protein partners and forms asingle functional unit that can be regulated bycaspase cleavage.To gain insight into the strength of binding of the
DSCR to its GST-fused ligands, we investigated thebinding of theMPR of Dsg1 (Fig. 6a) to the sameGST-fused proteins. Binding of purified MPR (Fig. 6c) toGST-fused PG and PKP1, but not GST alone, wasdetected in pull-down assays even when very smallamounts of glutathione Sepharose-bound proteinswere loaded onto SDS-PAGE gels, indicating a tightinteraction between the MPR and either PG or PKP1(Fig. 6e). Binding of purified MPR to GST-fused DPand Dsc1 was detected when 150-fold greateramounts of glutathione Sepharose-bound proteinswere loaded onto protein gels (Fig. 6f), indicating thatthe interaction between the MPR and DP and Dsc1 ismuch weaker and comparable to that seen betweenthe DSCR and all four GST-fused ligands (Fig. 6d).Overall, the pull-down assays suggest that the entireDsg1 cytoplasmic domain (i.e., MPR+DSCR) acts asa flexible scaffold for cytoplasmic complex assemblyat the desmosome with a network of strong andweak protein–protein interactions.
Discussion
Using a variety of biophysical approaches, wehave shown that the DSCR of Dsg1 is intrinsically
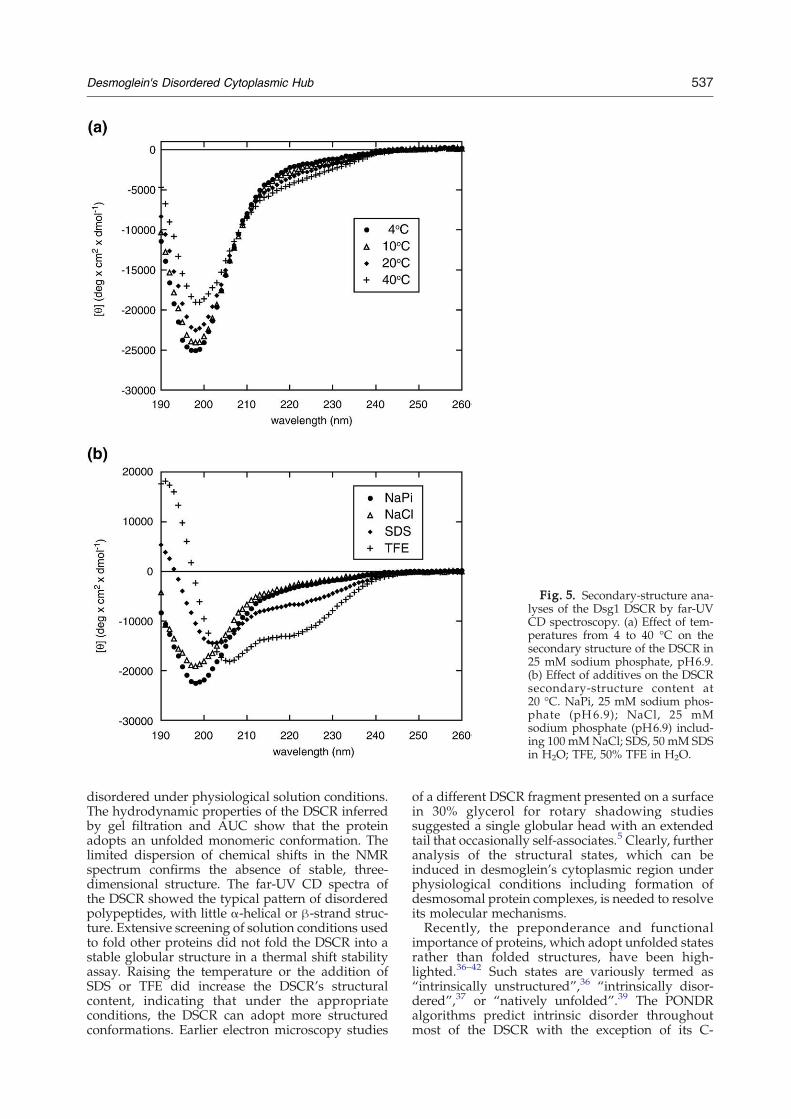
Fig. 5. Secondary-structure ana-lyses of the Dsg1 DSCR by far-UVCD spectroscopy. (a) Effect of tem-peratures from 4 to 40 °C on thesecondary structure of the DSCR in25 mM sodium phosphate, pH6.9.(b) Effect of additives on the DSCRsecondary-structure content at20 °C. NaPi, 25 mM sodium phos-phate (pH6.9); NaCl, 25 mMsodium phosphate (pH6.9) includ-ing 100 mMNaCl; SDS, 50 mM SDSin H2O; TFE, 50% TFE in H2O.
537Desmoglein's Disordered Cytoplasmic Hub
disordered under physiological solution conditions.The hydrodynamic properties of the DSCR inferredby gel filtration and AUC show that the proteinadopts an unfolded monomeric conformation. Thelimited dispersion of chemical shifts in the NMRspectrum confirms the absence of stable, three-dimensional structure. The far-UV CD spectra ofthe DSCR showed the typical pattern of disorderedpolypeptides, with little α-helical or β-strand struc-ture. Extensive screening of solution conditions usedto fold other proteins did not fold the DSCR into astable globular structure in a thermal shift stabilityassay. Raising the temperature or the addition ofSDS or TFE did increase the DSCR's structuralcontent, indicating that under the appropriateconditions, the DSCR can adopt more structuredconformations. Earlier electron microscopy studies
of a different DSCR fragment presented on a surfacein 30% glycerol for rotary shadowing studiessuggested a single globular head with an extendedtail that occasionally self-associates.5 Clearly, furtheranalysis of the structural states, which can beinduced in desmoglein's cytoplasmic region underphysiological conditions including formation ofdesmosomal protein complexes, is needed to resolveits molecular mechanisms.Recently, the preponderance and functional
importance of proteins, which adopt unfolded statesrather than folded structures, have been high-lighted.36–42 Such states are variously termed as“intrinsically unstructured”,36 “intrinsically disor-dered”,37 or “natively unfolded”.39 The PONDRalgorithms predict intrinsic disorder throughoutmost of the DSCR with the exception of its C-
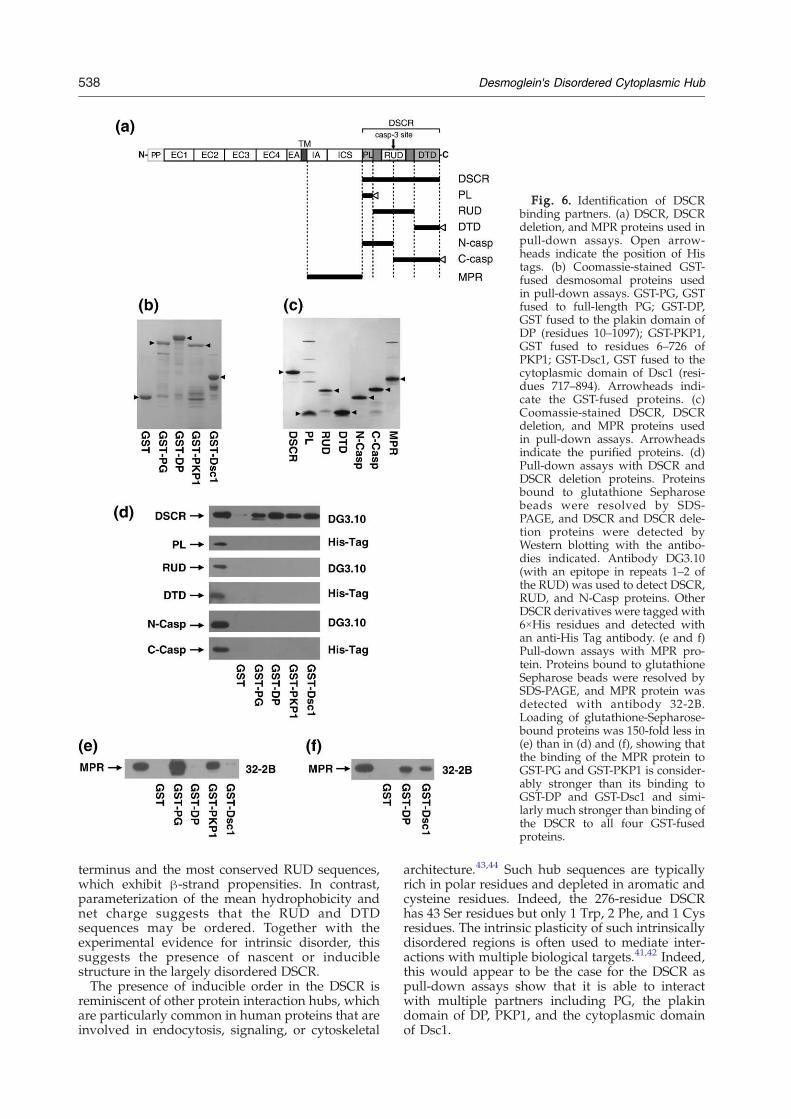
Fig. 6. Identification of DSCRbinding partners. (a) DSCR, DSCRdeletion, and MPR proteins used inpull-down assays. Open arrow-heads indicate the position of Histags. (b) Coomassie-stained GST-fused desmosomal proteins usedin pull-down assays. GST-PG, GSTfused to full-length PG; GST-DP,GST fused to the plakin domain ofDP (residues 10–1097); GST-PKP1,GST fused to residues 6–726 ofPKP1; GST-Dsc1, GST fused to thecytoplasmic domain of Dsc1 (resi-dues 717–894). Arrowheads indi-cate the GST-fused proteins. (c)Coomassie-stained DSCR, DSCRdeletion, and MPR proteins usedin pull-down assays. Arrowheadsindicate the purified proteins. (d)Pull-down assays with DSCR andDSCR deletion proteins. Proteinsbound to glutathione Sepharosebeads were resolved by SDS-PAGE, and DSCR and DSCR dele-tion proteins were detected byWestern blotting with the antibo-dies indicated. Antibody DG3.10(with an epitope in repeats 1–2 ofthe RUD) was used to detect DSCR,RUD, and N-Casp proteins. OtherDSCR derivatives were tagged with6×His residues and detected withan anti-His Tag antibody. (e and f)Pull-down assays with MPR pro-tein. Proteins bound to glutathioneSepharose beads were resolved bySDS-PAGE, and MPR protein wasdetected with antibody 32-2B.Loading of glutathione-Sepharose-bound proteins was 150-fold less in(e) than in (d) and (f), showing thatthe binding of the MPR protein toGST-PG and GST-PKP1 is consider-ably stronger than its binding toGST-DP and GST-Dsc1 and simi-larly much stronger than binding ofthe DSCR to all four GST-fusedproteins.
538 Desmoglein's Disordered Cytoplasmic Hub
terminus and the most conserved RUD sequences,which exhibit β-strand propensities. In contrast,parameterization of the mean hydrophobicity andnet charge suggests that the RUD and DTDsequences may be ordered. Together with theexperimental evidence for intrinsic disorder, thissuggests the presence of nascent or induciblestructure in the largely disordered DSCR.The presence of inducible order in the DSCR is
reminiscent of other protein interaction hubs, whichare particularly common in human proteins that areinvolved in endocytosis, signaling, or cytoskeletal
architecture.43,44 Such hub sequences are typicallyrich in polar residues and depleted in aromatic andcysteine residues. Indeed, the 276-residue DSCRhas 43 Ser residues but only 1 Trp, 2 Phe, and 1 Cysresidues. The intrinsic plasticity of such intrinsicallydisordered regions is often used to mediate inter-actions with multiple biological targets.41,42 Indeed,this would appear to be the case for the DSCR aspull-down assays show that it is able to interactwith multiple partners including PG, the plakindomain of DP, PKP1, and the cytoplasmic domainof Dsc1.

539Desmoglein's Disordered Cytoplasmic Hub
The Dsg1 MPR is able to bind the same fourdesmosomal proteins. It is well established that PGbinds to the ICS subdomain of the MPR.8,9 There issome disagreement in the literature regarding thebinding of PKP1 to the cytoplasmic domain ofDsg1. PKP1 has been shown to bind to Dsg1 in invitro overlay assays14 and in the yeast two-hybridsystem,13,15 but it does not immunoprecipitate withDsg1 following transfection into COS cells.13 How-ever, both PKP2 and PKP3 do coimmunoprecipitatewith Dsg1.16,17 In pull-down assays (Fig. 6e), astrong interaction between purified MPR proteinand GST-fused PKP1 was detected, thus mapping amajor PKP1 binding site to the MPR of Dsg1 andsupporting the view that PKP1 interacts directlywith Dsg1.The E-cadherin cytoplasmic domain is intrinsi-
cally disordered in the absence of its ligand β-catenin but, in its presence, binds in an extendedconformation that forms a large interface.27,28 It islikely that the DSCR also forms a large interface withits ligands as the entire DSCR is required for binding(Fig. 6). Whether the DSCR can bind all four ligandssimultaneously is unclear; it is possible that itsinducible conformation may be able to accommo-date single or multiple ligands. Our preliminaryanalysis (Fig. 1c and d, and unpublished data)suggests that the MPR is intrinsically disordered inthe absence of its ligands. Hence, we propose thatthe intrinsic disorder of both the DSCR and MPRendows the entire Dsg1 cytoplasmic domain withthe unique ability to bind desmosomal partners,thus enabling it to act as a hub for multiple proteininteractions at the cytoplasmic face of the desmo-some. We suggest that strong interactions betweensome cytoplasmic desmosomal proteins (e.g., MPRand PG/PKP1) are supported by a network ofweaker interactions between others (MPR and DP/Dsc1; DSCR and PG/PKP1/DP/Dsc1). This web ofinteractions places the desmoglein cytoplasmicdomain at the heart of the desmosomal complex,thus enabling it to play important role in desmo-some assembly and intercellular adhesion.The conformational flexibility of the DSCR may
facilitate posttranslational modifications such asphosphorylation. Interestingly, the Dsg1 RUD con-tains consensus PKC phosphorylation sites in themost conserved and hydrophobic sequences (Sup-plementary Fig. 1), suggesting a direct means ofregulating binding interactions. During woundhealing, PKC relocalizes from the cytoplasm to thedesmosomal plaque,45,46 and it has been suggestedthat phosphorylation of one or more cytoplasmicdesmosomal components by PKC may generate atransmembrane signal that alters the conformationof desmosomal cadherin extracellular domains andreduces their adhesive binding.46 It is tempting tospeculate that phosphorylation within the RUD,followed by a conformational change within theDSCR, could be involved in the transmission of asignal to the extracellular space. Such events couldbe abrogated by the Dsg2 V920G mutation, whichremoves conserved hydrophobicity in RUD repeat 2
and, thus, may compromise its interactions, leadingto the development of arrhythmogenic right ven-tricular cardiomyopathy/dysplasia.22The DSCR may provide a regulatory mechanism
for the regulation of desmosomal adhesion throughproteolytic degradation. There is a caspase-3 clea-vage site in RUD3 of Dsg1 that is responsible forregulating apoptosis in keratinocytes.7 Crucially,neither of the N- or C-terminal caspase-3 cleavagefragments of Dsg1 is able to bind other desmosomalproteins, suggesting that the DSCR could play acritical role in the dismantling of desmosomesfollowing caspase-3 cleavage. Thus, it may be thatcaspase-3 cleavage results in dissociation of otherdesmosomal components from the DSCR, a vitaland necessary first step in the disassembly ofdesmosomes. In addition, the DSCR also possessesa PEST motif between the ICS and PL domains(Supplementary Fig. 1). PEST sequences areenriched in Pro, Glu, Ser, and Thr residues and areflanked by, but do not contain, basic residues. Theyare correlated with rapid protein turnover in vivo.47Thus, in addition to having a potential role indesmosome assembly, the DSCR is well placed tomake important contributions to the regulation andloss of desmosomal adhesion in processes such asapoptosis and desquamation.
Materials and Methods
Expression and purification of the DSCR of Dsg1
DNA encoding the human Dsg1 DSCR (amino acidresidues 774–1049) was isolated by reverse-transcriptase-mediated PCR. Total RNAwas prepared from cultures ofprimary human keratinocytes (TCS CellWorks) by guani-dinium isothiocyanate extraction. RNA was reversetranscribed using random primers and a first-strandcDNA synthesis kit (Roche). Amplification was thenperformed using primers MC262/MC264 (SupplementaryTable 2). Amplified cDNA was cloned into BamHI andEcoRI sites of pGEX-6P-1 (GE Healthcare) and sequenced.The GST-DSCR fusion protein was expressed in E. colistrain BL21(DE3). Bacterial cells were grown initially at37 °C in 2xYTmedium and then induced for 4 h at 20 °C byaddition of 1 mM isopropyl β-D-thiogalactopyranoside.Cells were harvested by centrifugation, resuspended in50 mM phosphate (pH7.2) buffer and 150 mM KCl, andfrozen at −80 °C.Cells were thawed and lysed by lysozyme in the
presence of 1% Triton X-100 and complete proteaseinhibitor cocktail (Roche). Lysed cells were sonicatedand cleared by centrifugation. After filtering the super-natant, glutathione Sepharose 4B resin (GE Healthcare)was added. Following equilibration, the resin was washedand the fusion protein was eluted with 30 mM glu-tathione, 200 mM Tris–HCl (pH8.0), and 250 mM NaCl.The GST-DSCR protein was incubated overnight at 4 °Cwith PreScission Protease (GE Healthcare), and thecleaved GST was removed by binding to glutathioneSepharose resin. The DSCR protein was then passedsequentially through HiTrap Q and HiTrap SP columns(GE Healthcare) to remove any impurities and degrada-tion products.

540 Desmoglein's Disordered Cytoplasmic Hub
Expression and purification of DSCR deletion proteinsand the Dsg1 MPR
DNA encoding DSCR deletion constructs (PL, RUD,DTD, N-Casp, and C-Casp) was amplified from DSCRDNA using the primers listed in Supplementary Table 2.PL, DTD, and C-Casp DNAwere tagged at the 3′ end withDNA encoding 6×His residues. DNA encoding the Dsg1MPR (amino acid residues 574–773) was amplified by RT-PCR using human keratinocyte RNA as starting materialand primers MC292/MC293 (Supplementary Table 1). Ineach case, DNA was cloned into pGEX-6P-1, and theproteins were expressed and purified as described for theDSCR except that following PreScission protease diges-tion, PL, RUD, N-Casp, and MPR proteins were purifiedusing glutathione agarose and HiTrap Q columns and theDTD and C-Casp proteins were purified with a HisTrapcolumn (GE Healthcare). Concentration of deletion pro-teins was estimated by SDS-PAGE using fully purifiedDSCR as standard.
Expression and purification of GST-fused desmosomalproteins
A full-length human PG cDNA (obtained from P.Cowin, New York University School of Medicine, NewYork) was cloned into the SalI and HindIII sites of pGEX-6P-1. The 5′ end of the PG cDNA was amplified withprimers MC288 and MC289 to ensure that DNA encodingGST and PG was in frame (Supplementary Table 2). ThePCR product was then cloned into BamHI- and SphI-cutpGEX-6P-1/PG. A plasmid encoding GST fused to theplakin head domain (amino acids 10–1097) of human DPwas obtained by cloning a 3.3-kb SacII (blunt end)-XbaI(blunt end) fragment of the full-length cDNA (obtainedfrom K. Green, Northwestern University Feinberg Schoolof Medicine, Chicago) into the SmaI site of pGEX-4T-3.DNA encoding GST fused to residues 6–726 of humanPKP1 (i.e., full length but missing the five N-terminalamino acids) was obtained by cloning a BsrBI (blunt end)-XhoI fragment of the full-length human cDNA (obtainedfrom Andrew South, Queen Mary's School of Medicineand Dentistry, London) into SalI (blunt end)-XhoI-cutpGEX-6P-1. A plasmid encoding GST fused to thecytoplasmic domain of human Dsc1a (residues 717–894)was obtained by PCR with primers MC290 and MC291(Supplementary Table 2) using a full-length Dsc1a cDNAclone (obtained from Takashi Hashimoto, Kurume Uni-versity School of Medicine) as template. The PCR productwas cloned into BamHI- and EcoRI-cut pGEX-6P-1. AllGST fusion proteins were expressed in E. coli and purifiedusing glutathione agarose. Concentrations of GST fusionproteins were estimated by SDS-PAGE using fully purifiedGST as a standard.
Protein sequence analyses and structural predictions
Secondary-structure prediction was performed usingthe DomPred48 or PredictProtein.49,50 The PEST-FINDprogram was used to identify PEST motifs.47 Disorderprediction was carried out using the PONDR VL-XT andVSL1 neural network programs (http://www.pondr.com).23–25 Access to the PONDR server was provided byMolecular Kinetics (Indianapolis, IN). In order to predictwhether conserved regions were sufficiently hydrophobicto fold, we determined the mean net charge (R) of the Dsg1protein regions as the absolute value of the differencebetween the numbers of positively and negatively charged
residues divided by the total number of amino acidresidues, as calculated with the program ProtParam. Themean hydrophobicity (H) was calculated as the sum of thenormalized hydrophobicities of all residues divided by thetotal number of residues minus 4.4 The normalizedhydrophobicities were calculated using the programProtscale,51 using the default Hphob/Kyte & Doolittlevalues and a window size of 5 and normalizing the scalefrom 0 to 1.
Size-exclusion chromatography
The DSCR protein (29 μM) in 100 μl of a buffercontaining 20 mM phosphate (pH7.2), 100 mM KCl, and1 mM DTT was applied onto a Superdex 200 HR10/30column (GE Healthcare) on an AKTA Purifier instrument(GE Healthcare), running with the same buffer at a flowrate of 0.5 ml/min at room temperature. Protein standardsfrom high- or low-molecular-mass gel-filtration kits(Amersham Biosciences) were used to calibrate thecolumn as follows: blue dextran (2000 kDa) to determinethe void volume, ferritin (440 kDa; Rs=61 Å), aldolase(158 kDa; Rs=48.1 Å), albumin (67 kDa; Rs=35.5 Å),ovalbumin (43 kDa; Rs =30.5 Å), chymotrypsinogen(25 kDa; Rs=20.9 Å), and ribonuclease A (13.7 kDa;Rs =16.4 Å). Preparation of a calibration curve forestimation of apparent molecular mass and Stokes radiuswas carried out with retention volumes of the standardproteins. The theoretical Stokes radii of native and fullyunfolded proteins were calculated by standard methods.29
Sedimentation equilibrium analyses
A Beckman XL-I analytical ultracentrifuge (BeckmanCoulter, Palo Alto, CA, USA) using an 8-cell 50Ti rotor wasused for the AUC studies. Samples of the DSCR at 19, 58,and 116 μM were prepared in 20 mM phosphate (pH7.2)buffer including 100 mM NaCl and 1 mM DTT and werecentrifuged at 30, 33, and 36 krpm for 20 h at 4 °C. Theabsorbance of the sample was measured at a wavelengthof 280 nm throughout the cell. A total of three measure-ments were taken at 1-h intervals at the end of each run.These measurements were compared to ensure thatequilibrium had been reached. The partial specific volumeof the protein and the buffer density were calculated usingSEDNTERP.52 Data from each experiment were analyzedusing SEDPHAT.53
One-dimensional 1H NMR spectroscopy
Spectra were recorded at 25 °C on a Varian Inova800MHz spectrometer equippedwith a 5-mmpulsed-fieldgradient cryogenic probe. The NMR sample contained62 μM DSCR protein in 20 mM phosphate (pH6.9) buffer,100 mM NaCl, 1 mM Tris(2-chloroethyl) phosphate(TCEP), 90% H2O, and 10% 2H2O. The number of scanswas 256. The spectra were processed with NMRLABsoftware.54
Thermal shift assays
ThermoFluor® experiments were carried out with areal-time PCRmachine Mx3005P (Stratagene). The protein(2 μl) was mixed with a fluorescent dye, SYPRO Orange(Molecular Probes), and 18 μl of each folding buffer in aThermo-Fast 96-well PCR plate (ABgene), resulting in

541Desmoglein's Disordered Cytoplasmic Hub
final protein concentrations of 2 μM. The folding condi-tions included Tris–HCl, sodium phosphate, Bis–Tris, 4-morpholineethanesulfonic acid, Hepes, or cacodylate as abuffer salt at 50 mM (pH range from 6.0 to 8.0), and 100–150 mMNaCl, 150 mMKCl, 150 mMNH4Cl, 2 mMCaCl2,2 mM ZnCl2, 1–50 mM TCEP, 1 M NDSB201, 200–500 mMsucrose, 0.3 mM DDM, 800 mM L-arginine, 1–2 M TMAO,0.05% polyethylene glycol 4000 or 1–2 M sarcosine, 10–100% TFE, 2–50 mM SDS, 10–200 mM FOS-CHOLINE-ISO-9 (Anatrace), and 4–40 mM DPC or 4–40 mM 3-[(3-cholamidopropyl)dimethylammonio]propanesulfonicacid as additives. The plate was heated at a rate of 1 °C/min from 25 to 95 °C, and fluorescence was measured at1 °C increments. Fluorescence was filtered throughcustom interference excitation (492 nm) and emission(568 nm) filters.
Far-UV CD spectroscopy
The CD spectra were measured on a Jasco J-810spectropolarimeter using a 0.02-cm path-length cuvetteand protein concentration of 13 μM (0.37 mg/ml). Thescanned wavelength range was 185–300 nm. The second-ary-structure content was estimated from the CD spectrausing the CONTINLL algorithms.55,56 The DSCR spectrawere compared with reference data set 7 (which containsspectra from 48 proteins including 5 that are denatured) atthe DICHROWEB server.57,58 The back-calculated spectraand experimental spectra were compared to estimate thenormalized root-mean-square deviation values, whichwere below 0.1.
Pull-down assays
Binding assays were performed with purified DSCR,DSCR deletion, and MPR proteins (1 μM concentration) in25 mM phosphate (pH7.2) including 100 mMNaCl, 5 mMDTT, and 0.2% Triton X-100 (400 μl). The GST control andGST fusions of desmosomal proteins were added (1 μMconcentration), and the mixtures were rotated for 1 h at4 °C. A 50% slurry of glutathione Sepharose 4B (GEHealthcare) (40 μl) was then added, and the mixture wasrotated for a further 1 h at 4 °C. After centrifugation andremoval of the supernatant, beads were washed threetimes with 1 ml assay buffer. After the final wash, beadswere suspended in SDS-PAGE sample buffer and boiledfor 5 min. The mixture was then centrifuged and thesupernatant was removed. Proteins were resolved by SDS-PAGE and blotted onto polyvinylidene fluoride mem-brane. Blotted proteins were detected using antibodyDG3.10 against Dsg1+2 (Progen) (epitope in repeats 1–2 ofthe RUD), 32-2B against Dsg1+359 (epitope in MPR), or ananti-His Tag monoclonal antibody (Novagen) against6×His residues.
Acknowledgements
We thank the Birmingham Biophysical Character-ization Facility and the HenryWellcome Building forBiomolecular NMR Spectroscopy, which is sup-ported by the Wellcome Trust. This research wasfunded by Biotechnology and Biological SciencesResearch Council, EU PRISM, and Cancer ResearchUK (M.O.) grants. K.K. was supported in part by a
fellowship of Yamanouchi Foundation for Researchon Metabolic Disorders.
Supplementary Data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.jmb.2008.12.054
References
1. Dusek, R. L., Godsel, L. M. & Green, K. J. (2007).Discriminating roles of desmosomal cadherins: beyonddesmosomal adhesion. J. Dermatol. Sci. 45, 7–21.
2. Garrod, D. & Chidgey, M. (2008). Desmosomestructure, composition and function. Biochim. Biophys.Acta, 1778, 572–587.
3. Garrod, D., Chidgey, M. & North, A. (1996). Desmo-somes: differentiation, development, dynamics anddisease. Curr. Opin. Cell Biol. 8, 670–678.
4. Uversky, V. N., Gillespie, J. R. & Fink, A. L. (2000).Why are “natively unfolded” proteins unstructuredunder physiologic conditions? Proteins, 41, 415–427.
5. Rutman, A. J., Buxton, R. S. & Burdett, I. D. J. (1994).Visualization by electron microscopy of the uniquepart of the cytoplasmic domain of a desmoglein, acadherin-like protein of the desmosome type of celljunction. FEBS Lett. 353, 194–196.
6. Nilles, L. A., Parry, D. A. D., Powers, E. E., Angst, B.D., Wagner, R. M. & Green, K. J. (1991). Structuralanalysis and expression of human desmoglein: acadherin-like component of the desmosome. J. CellSci. 99, 809–821.
7. Dusek, R. L., Getsios, S., Chen, F., Park, J. K.,Amargo, E. V., Cryns, V. L. & Green, K. J. (2006).The differentiation-dependent desmosomal cadherindesmoglein 1 is a novel caspase-3 target thatregulates apoptosis in keratinocytes. J. Biol. Chem.281, 3614–3624.
8. Mathur, M., Goodwin, M. L. & Cowin, P. (1994).Interactions of the cytoplasmic domain of the desmo-somal cadherin with plakoglobin. J. Biol. Chem. 269,14075–14080.
9. Troyanovsky, S. M., Troyanovsky, R. B., Eshkind,L. G., Krutovskikh, V. A., Leube, R. E. & Franke,W. W. (1994). Identification of the plakoglobin-binding domain in desmoglein and its role in plaqueassembly and intermediate filament anchorage. J. CellBiol. 127, 151–160.
10. Troyanovsky, S. M., Troyanovsky, R. B., Eshkind,L. G., Leube, R. E. & Franke, W. W. (1994). Identifica-tion of amino acid sequence motifs in desmocollin, adesmosomal glycoprotein, that are required forplakoglobin binding and plaque formation. Proc.Natl Acad. Sci. USA, 91, 10790–10794.
11. Bornslaeger, E. A., Godsel, L. M., Corcoran, C. M.,Park, J. K., Hatzfeld, M., Kowalczyk, A. P. & Green,K. J. (2001). Plakophilin 1 interferes with plakoglobinbinding to desmoplakin, yet together with plako-globin promotes clustering of desmosomal plaquecomplexes at cell–cell borders. J. Cell Sci. 114, 727–738.
12. Hatzfeld, M. (2007). Plakophilins: multifunctionalproteins or just regulators of desmosomal adhesion?Biochim. Biophys. Acta, 1773, 69–77.
13. Kowalczyk, A. P., Hatzfeld, M., Bornslaeger, E. A.,Kopp, D. S., Borgwardt, J. E., Corcoran, C. M. et al.(1999). The head domain of plakophilin-1 binds to

542 Desmoglein's Disordered Cytoplasmic Hub
desmoplakin and enhances its recruitment to desmo-somes. J. Biol. Chem. 274, 18145–18148.
14. Smith, E. A. & Fuchs, E. (1998). Defining the interac-tions between intermediate filaments and desmo-somes. J. Cell Biol. 141, 1229–1241.
15. Hatzfeld, M., Haffner, C., Schulze, K. & Vinzens, U.(2000). The function of plakophilin 1 in desmosomeassembly and actin filament organization. J. Cell Biol.149, 209–222.
16. Chen, X., Bonne, S., Hatzfeld, M., van Roy, F. &Green, K. J. (2002). Protein binding and functionalcharacterization of plakophilin 2. J. Biol. Chem. 277,10512–10522.
17. Bonne, S., Gilbert, B., Hatzfeld, M., Chen, X., Green,K. J. & van Roy, F. (2003). Defining desmosomalplakophilin-3 interactions. J. Cell Biol. 161, 403–416.
18. Stanley, J. R. & Amagai, M. (2006). Pemphigus,bullous impetigo, and the staphylococcal scalded-skin syndrome. N. Engl. J. Med. 355, 1800–1810.
19. Rickman, L., Simrak, D., Stevens, H. P., Hunt, D. M.,King, I. A., Bryant, S. P. et al. (1999). N-terminaldeletion in a desmosomal cadherin causes the auto-somal dominant skin disease striate palmoplantarkeratoderma. Hum. Mol. Genet. 8, 971–976.
20. Hunt, D. M., Rickman, L., Whittock, N. V., Eady, R. A.,Simrak, D., Dopping-Hepenstal, P. J. et al. (2001).Spectrum of dominant mutations in the desmosomalcadherin desmoglein 1, causing the skin disease striatepalmoplantar keratoderma. Eur. J. Hum. Genet. 9,197–203.
21. Yashiro, M., Nishioka, N. & Hirakawa, K. (2006).Decreased expression of the adhesion moleculedesmoglein-2 is associated with diffuse-type gastriccarcinoma. Eur. J. Cancer, 42, 2397–2403.
22. Syrris, P., Ward, D., Asimaki, A., Evans, A.,Sen-Chowdhry, S., Hughes, S. E. et al. (2007).Desmoglein-2 mutations in arrhythmogenic rightventricular cardiomyopathy: a genotype-phenotypecharacterization of familial disease. Eur. Heart J. 28,581–588.
23. Li, X., Romero, P., Rani, M., Dunker, A. K. &Obradovic, Z. (1999). Predicting protein disorder forN-, C-, and internal regions. Genome Inform. Ser.Workshop Genome Inform. 10, 30–40.
24. Romero, P., Obradovic, Z., Li, X., Garner, E. C., Brown,C. J. & Dunker, A. K. (2001). Sequence complexity ofdisordered protein. Proteins, 42, 38–48.
25. Obradovic, Z., Peng, K., Vucetic, S., Radivojac, P. &Dunker, A. K. (2005). Exploiting heterogeneoussequence properties improves prediction of proteindisorder. Proteins, 61, 176–182.
26. Overduin, M., Harvey, T. S., Bagby, S., Tong, K. I., Yau,P., Takeichi, M. L. et al. (1995). Solution structure of theepithelial cadherin domain responsible for selectivecell adhesion. Science, 267, 386–389.
27. Huber, A. H., Stewart, D. B., Laurents, D. V., Nelson,W. J. & Weis, W. I. (2001). The cadherin cytoplasmicdomain is unstructured in the absence of β-catenin.J. Biol. Chem. 276, 12301–12309.
28. Huber, A. H. & Weis, W. I. (2001). The structure of theβ-catenin/E-cadherin complex and the molecularbasis of diverse ligand recognition by β-catenin. Cell,105, 391–402.
29. Uversky, V. N. (1993). Use of fast size-exclusionliquid chromatography to study the unfolding ofproteins which denature through the molten globule.Biochemistry, 32, 13288–13298.
30. Mezzasalma, T.M., Kranz, J. K., Chan,W., Struble,G. T.,Schalk-Hihi, C., Deckman, I. C. et al. (2007). Enhancing
recombinant protein quality and yield by proteinstability profiling. J. Biomol. Screening, 12, 418–428.
31. Baskakov, I. & Bolen, D. W. (1998). Forcing thermo-dynamically unfolded proteins to fold. J. Biol. Chem.273, 4831–4834.
32. Vincentelli, R., Canaan, S., Campanacci, V., Valencia,C., Maurin, D., Frassinetti, F. et al. (2004). High-throughput automated refolding screening of inclu-sion bodies. Protein Sci. 13, 2782–2792.
33. Willis,M. S.,Hogan, J. K., Prabhakar, P., Liu, X., Tsai, K.,Wei, Y. et al. (2005). Investigation of protein refoldingusing a fractional factorial screen: a study of reagenteffects and interactions. Protein Sci. 14, 1818–1826.
34. Song, J., Lee,M. S., Carlberg, I., Vener, A. V. &Markley,J. L. (2006). Micelle-induced folding of spinachthylakoid soluble phosphoprotein of 9 kDa and itsfunctional implications. Biochemistry, 45, 15633–15643.
35. Longhi, S., Receveur-Brechot, V., Karlin, D., Johansson,K., Darbon, H., Bhella, D. et al. (2003). The C-terminaldomain of the measles virus nucleoprotein isintrinsically disordered and folds upon binding tothe C-terminal moiety of the phosphoprotein. J. Biol.Chem. 278, 18638–18648.
36. Wright, P. E. & Dyson, H. J. (1999). Intrinsicallyunstructured proteins: re-assessing the protein struc-ture–function paradigm. J. Mol. Biol. 293, 321–331.
37. Dunker, A. K., Lawson, J. D., Brown, C. J., Williams,R. M., Romero, P., Oh, J. S. et al. (2001). Intrinsicallydisordered protein. J. Mol. Graphics Modell. 19, 26–59.
38. Tompa, P. (2002). Intrinsically unstructured proteins.Trends Biochem. Sci. 27, 527–533.
39. Uversky, V. N. (2002). Natively unfolded proteins: apoint where biology waits for physics. Protein Sci. 11,739–756.
40. Fink, A. L. (2005). Natively unfolded proteins. Curr.Opin. Struct. Biol. 15, 35–41.
41. Dyson, H. J. & Wright, P. E. (2005). Intrinsicallyunstructured proteins and their functions. Nat. Rev.,Mol. Cell Biol. 6, 197–208.
42. Tompa, P., Szasz, C. & Buday, L. (2005). Structuraldisorder throws new light on moonlighting. TrendsBiochem. Sci. 30, 484–489.
43. Haynes, C., Oldfield, C. J., Ji, F., Klitgord, N., Cusick,M. E., Radivojac, P. et al. (2006). Intrinsic disorder is acommon feature of hub proteins from four eukaryoticinteractomes. PLoS Comput. Biol. 2, e100.
44. Dafforn, T. R. & Smith, C. J. (2004). Natively unfoldeddomains in endocytosis: hooks, lines and linkers.EMBO Rep. 5, 1046–1052.
45. Wallis, S., Lloyd, S., Wise, I., Ireland, G., Fleming, T. P.& Garrod, D. (2000). The alpha isoform of proteinkinase C is involved in signaling the response ofdesmosomes to wounding in cultured epithelial cells.Mol. Biol. Cell, 11, 1077–1092.
46. Garrod, D. R., Berika, M. Y., Bardsley, W. F., Holmes,D. & Tabernero, L. (2005). Hyper-adhesion in desmo-somes: its regulation in wound healing and possiblerelationship to cadherin crystal structure. J. Cell Sci.118, 5743–5754.
47. Rechsteiner,M. &Rogers, S.W. (1996). PESTsequencesand regulation by proteolysis. Trends Biochem. Sci. 21,267–271.
48. Marsden, R. L., McGuffin, L. J. & Jones, D. T. (2002).Rapid protein domain assignment from amino acidsequence using predicted secondary structure. ProteinSci. 11, 2814–2824.
49. Rost, B. & Sander, C. (1993). Prediction of proteinsecondary structure at better than 70% accuracy.J. Mol. Biol. 232, 584–599.

543Desmoglein's Disordered Cytoplasmic Hub
50. Rost, B., Yachdav, G. & Liu, J. (2004). The PredictPro-tein server. Nucleic Acids Res. 32, 321–326.
51. Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S.,Wilkins, M. R., Appel, R. D. & Bairoch, A. (2005).Protein identification and analysis tools on theExPASy server. In The Proteomics Protocols Handbook(Walker, J. M., ed.), pp. 571–607, Humana Press, NJ.
52. Laue, T., Shah, B., Ridgeway, T. & Pelletier, S.(1992). In Analytical ultracentrifugation in biochemistryand polymer science (Harding, S., Rowe, A. &Horyon, J., eds), pp. 90–125, Royal Society ofChemistry, Cambridge, UK.
53. Vistica, J., Dam, J., Balbo, A., Yikilmaz, E., Mariuzza,R. A., Rouault, T. A. & Schuck, P. (2004). Sedimenta-tion equilibrium analysis of protein interactions withglobal implicit mass conservation constraints andsystematic noise decomposition. Anal. Biochem. 326,234–256.
54. Gunther, U. L., Ludwig, C. & Ruterjans, H. (2000).NMRLAB—advanced NMR data processing inMatlab. J. Magn. Reson. 145, 201–208.
55. Provencher, S. W. & Glockner, J. (1981). Estimation ofglobular protein secondary structure from circulardichroism. Biochemistry, 20, 33–37.
56. van Stokkum, I. H. M., Spoelder, H. J. W.,Bloemendal, M., van Grondelle, R. & Groen, F. C. A.(1990). Estimation of protein secondary structureand error analysis from circular dichroism spectra.Anal. Biochem. 191, 110–118.
57. Lobley, A., Whitmore, L. & Wallace, B. A. (2002).DICHROWEB: an interactive website for the analysisof protein secondary structure from circular dichroismspectra. Bioinformatics, 18, 211–212.
58. Whitmore, L. & Wallace, B. A. (2004). DICHROWEB,an online server for protein secondary structureanalyses from circular dichroism spectroscopic data.Nucleic Acids Res. 32, 668–673.
59. Vilela, M. J., Hashimoto, T., Nishikawa, T., North, A. J.& Garrod, D. R. (1995). A simple epithelial cell line(MDCK) shows heterogeneity of desmoglein iso-forms, one resembling pemphigus vulgaris antigen.J. Cell Sci. 108, 1743–1750.
Related Documents