Kenneth C. Hayes a Pramod Khosla b a Foster Biomedical Research Lab, Brandeis University, Waltham, USA b Department of Nutrition and Food Science, Wayne State University, Detroit, USA The complex interplay of palm oil fatty acids on blood lipids The cholesterol-raising potential of saturated fats has been noted for 50 years, but how they function and which among the saturated fats are most to blame is only now becoming apparent. The focus has shifted from saturated fats to individual fatty acids and the total composite of fatty acids (SFA, MUFA, PUFA) that comprise our daily fat intake. An adequate intake of both PUFA and SFA may be needed for the ideal LDL/ HDL ratio in blood, as both contribute to the regulatory balance in lipoprotein metab- olism. Palm oil, by virtue of its fatty acid composition equally balanced between satu- rated and unsaturated fatty acids, impacts blood lipids in different ways, depending on an individual’s lipid profile. Recently, the realization that partially hydrogenated fats (containing trans fatty acids) have adverse health effects has necessitated finding alternative fat sources that provide specific functional characteristics without com- promising health. The immediate solution would seem to be blending appropriate nat- ural saturated and polyunsaturated fats to meet the combined requirements for food technology and energy metabolism. In technical situations where a higher-melting point fat is needed, as in margarines, shortenings, and frying oil, palm oil – or more specifically palm olein – would appear to be the fat of choice for blending with unsa- turated oils. Keywords: Palm oil, palm olein, lipid metabolism, lipoproteins, trans fat, interesterified fat. Eur. J. Lipid Sci. Technol. 109 (2007) 453–464 DOI 10.1002/ejlt.200700005 453 1 Introduction After soybean oil (SBO), oil from the fruit of the oil palm (Elaeis guineensis) is the most widely traded edible oil in the world, accounting for about one quarter of the world fats and oils supply. Furthermore, palm oil (PO), a palmi- tic-oleic acid-rich vegetable oil grown in the tropical cli- mates of Southeast Asia, Africa, and Latin America, is the most cost-effective edible oil to produce and is projected to overtake SBO production in the near future. An inter- esting aspect of PO is its versatility and functionality. Unlike most other edible oils that come as a single entity, PO is readily fractionated to provide a variety of oils of different solid fat content and melting points that can serve different food applications. In addition to PO itself, these oils include a liquid palm olein (POL) and the more solid palm stearin (Pst), separated as the upper and lower fractions during dry (melt) fractionation of PO itself. On a lesser scale, POL again can be dry fractionated to pro- duce superpalm olein and palm mid fraction, a softer stearin. Finally, the seed kernel within the fruit gives rise to palm kernel oil (PKO), which has a much more saturated fatty acid (SFA) composition than PO, very similar to coconut oil. These more saturated oils serve different nutritional applications, being primarily used for con- fectionaries or as hardstock in margarines and some shortenings. A thorough review of the chemistry and bio- chemistry of PO has been published [1]. When initially extracted, crude PO gives rise to red palm oil (RPO), which contains the many carotenoids (vitamin A precursors) and tocols. From the nutritional perspective, it is noteworthy and of public health significance that this crude RPO now serves as an inexpensive cooking oil in areas of the world where vitamin A deficiency is endemic. Its supplementation is making great strides in eradicating growth and blindness problems associated with that deficiency [2–6]. The bulk of PO and its fractions are directed to general food applications, as in margarines, shortenings, and fry- ing oil; and their nutritional properties have been exten- sively studied. Nutrition research related to PO has focused primarily on two major aspects: (1) the fatty acid composition affecting blood cholesterol and (2) enhanc- ing functionality of food products requiring a stable shelf life or heat stability during frying. The objective of this review is to consider the nutritional aspects of PO within the context of the major edible oils in the world, including the interrelated aspects of the fatty acids between PO Correspondence: Kenneth C. Hayes, Foster Biomedical Research Lab, Brandeis University, Waltham, MA 02453, USA. Phone: 11 781 736 2051, Fax: 11 781 736 2451, e-mail: [email protected] © 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com Review Article

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Kenneth C. Hayesa
Pramod Khoslab
a Foster Biomedical ResearchLab,Brandeis University,Waltham, USA
b Department of Nutrition andFood Science,Wayne State University,Detroit, USA
The complex interplay of palm oil fatty acids onblood lipids
The cholesterol-raising potential of saturated fats has been noted for 50 years, but howthey function and which among the saturated fats are most to blame is only nowbecoming apparent. The focus has shifted from saturated fats to individual fatty acidsand the total composite of fatty acids (SFA, MUFA, PUFA) that comprise our daily fatintake. An adequate intake of both PUFA and SFA may be needed for the ideal LDL/HDL ratio in blood, as both contribute to the regulatory balance in lipoprotein metab-olism. Palm oil, by virtue of its fatty acid composition equally balanced between satu-rated and unsaturated fatty acids, impacts blood lipids in different ways, depending onan individual’s lipid profile. Recently, the realization that partially hydrogenated fats(containing trans fatty acids) have adverse health effects has necessitated findingalternative fat sources that provide specific functional characteristics without com-promising health. The immediate solution would seem to be blending appropriate nat-ural saturated and polyunsaturated fats to meet the combined requirements for foodtechnology and energy metabolism. In technical situations where a higher-meltingpoint fat is needed, as in margarines, shortenings, and frying oil, palm oil – or morespecifically palm olein – would appear to be the fat of choice for blending with unsa-turated oils.
Keywords: Palm oil, palm olein, lipid metabolism, lipoproteins, trans fat, interesterifiedfat.
Eur. J. Lipid Sci. Technol. 109 (2007) 453–464 DOI 10.1002/ejlt.200700005 453
1 Introduction
After soybean oil (SBO), oil from the fruit of the oil palm(Elaeis guineensis) is the most widely traded edible oil inthe world, accounting for about one quarter of the worldfats and oils supply. Furthermore, palm oil (PO), a palmi-tic-oleic acid-rich vegetable oil grown in the tropical cli-mates of Southeast Asia, Africa, and Latin America, is themost cost-effective edible oil to produce and is projectedto overtake SBO production in the near future. An inter-esting aspect of PO is its versatility and functionality.Unlike most other edible oils that come as a single entity,PO is readily fractionated to provide a variety of oils ofdifferent solid fat content and melting points that canserve different food applications. In addition to PO itself,these oils include a liquid palm olein (POL) and the moresolid palm stearin (Pst), separated as the upper and lowerfractions during dry (melt) fractionation of PO itself. On alesser scale, POL again can be dry fractionated to pro-duce superpalm olein and palm mid fraction, a softerstearin. Finally, the seed kernel within the fruit gives rise topalm kernel oil (PKO), which has a much more saturated
fatty acid (SFA) composition than PO, very similar tococonut oil. These more saturated oils serve differentnutritional applications, being primarily used for con-fectionaries or as hardstock in margarines and someshortenings. A thorough review of the chemistry and bio-chemistry of PO has been published [1].
When initially extracted, crude PO gives rise to red palmoil (RPO), which contains the many carotenoids (vitamin Aprecursors) and tocols. From the nutritional perspective, itis noteworthy and of public health significance that thiscrude RPO now serves as an inexpensive cooking oil inareas of the world where vitamin A deficiency is endemic.Its supplementation is making great strides in eradicatinggrowth and blindness problems associated with thatdeficiency [2–6].
The bulk of PO and its fractions are directed to generalfood applications, as in margarines, shortenings, and fry-ing oil; and their nutritional properties have been exten-sively studied. Nutrition research related to PO hasfocused primarily on two major aspects: (1) the fatty acidcomposition affecting blood cholesterol and (2) enhanc-ing functionality of food products requiring a stable shelflife or heat stability during frying. The objective of thisreview is to consider the nutritional aspects of PO withinthe context of the major edible oils in the world, includingthe interrelated aspects of the fatty acids between PO
Correspondence: Kenneth C. Hayes, Foster BiomedicalResearch Lab, Brandeis University, Waltham, MA 02453, USA.Phone: 11 781 736 2051, Fax: 11 781 736 2451, e-mail:[email protected]
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Rev
iew
Art
icle

454 K. C. Hayes and P. Khosla Eur. J. Lipid Sci. Technol. 109 (2007) 453–464
and polyunsaturated oils during metabolism. The goodnews is that in combination with SBO or other poly-unsaturated oils the resulting two-oil blend has thepotential to provide the world with a healthy balance of allrequired fatty acids needed for metabolism and energy inthe 21st century and beyond.
The main concern for fats, particularly PO, when con-sidered from a general nutritional perspective, has to dowith their degree of saturation and the effect they have onblood lipids, i.e. blood cholesterol and its distributionamong its transport particles. Circulating cholesterol is ahigh-profile, readily measured biological response utiliz-ing clinically available assays linked to heart disease, so itis a highly relevant index of our nutritional wellbeing that issensitive to fat intake and composition. Furthermore, theresponse in cholesterol likely reflects how fats and fattyacids function at the cellular level, in general. Manyeffects of fats on other systems are now known, includingthose on immune function, nervous system developmentand function, and insulin dynamics and diabetes, tomention a few. But these are beyond the scope of thisreview, where our intent is more fundamental and focusedon basic questions concerning fats and nutrition, allowingthe reader to better comprehend the complexity of POnutrition. Our focus on heart disease illustrates the meta-bolic effects of PO, and is well documented in terms ofunderstanding the PO-nutrition relationship.
The discussion concerning PO consumption and heartdisease is complex [7, 8], but it is generally agreed bymost in the field that the cholesterol response to dietaryfat is best evaluated by measurements of total bloodcholesterol (TC), and its two main lipoprotein (LP) sub-components, LDL and HDL, with the lowest TC and LDL-C/HDL-C ratio being considered ideal.
2 The LP profile and the LDL/HDL ratio
How the LP profile of someone with a normal cholesterolvalue versus someone with high plasma cholesterol fitsinto this story is an important consideration. It appearsthat the fundamental blood cholesterol response to thecurrent American Heart Association (AHA) balanced fattyacid diet (see below for details) is similar in both cate-gories of individuals. That is, the approximately 1 : 1 : 1balance in SFA to monounsaturated fatty acids (MUFA orM) to polyunsaturated fatty acids (PUFA or P), S : M : P,generally recommended appears to be important to bothgroups for generating the ideal LDL/HDL ratio. In absoluteterms, the response by the high-cholesterol individual tochanges in the diet fatty acid profile is more dramatic, butthe person with a normal cholesterol value responds inthe same manner, just not to the same degree.
It is true that elevated cholesterol (TC .180 mg/100 mL,LDL .110 mg/100 mL) begins to increase the risk forcoronary heart disease (CHD). Any increase above180 mg/100 mL occurs mostly in the LDL pool, and thisLP is the one that is deposited during arterial cholesterolbuild-up. On the other hand, people (and essentially allanimals) with naturally high levels of HDL do not developCHD, primarily because this LP transports cholesterolback to the liver for excretion in bile. HDL in the arterialwall also blocks LDL deterioration, thereby preventing thelocal damage induced by LDL accumulation. Thus, the“bad” and “good” connotation for these two LP becomesapparent, and it is easy to understand why one wishes tohave the lowest LDL and highest HDL (i.e. lowest LDL/HDL ratio) for any given TC value. It is also easy toappreciate the desire to identify the best dietary fat com-position to make it happen.
3 Basic fats, basic facts
In the consideration of dietary fat and heart disease, sev-eral questions arise. First, how much fat is “healthy”, bothin terms of the total dietary amount (in grams) and as apercent of total daily calorie intake (abbreviated as %en),for example, 20, 30 or 40%en? Second, once these fatintake parameters are ascertained, what is the properbalance of fatty acids, i.e. the ratio of SFA to MUFA toPUFA that one should consume each day? And, ulti-mately, where does PO and its fractions fit into the pic-ture? Does it matter whether we consume 2 g or 20 g ofPO per day? If so, what sets the limits? Is one fraction ofPO different in its impact from another? And within eachof the SFA, MUFA, and PUFA categories, which specificSFA and PUFA are most healthy? Third, how does onemeasure a “healthy response” to changes in these dietaryfat components to know whether the fat consumed isnutritious and healthy? Fourth, what influence does diet-ary cholesterol have in this scenario, and what implica-tions does this have for PO, which is cholesterol free?Finally, how important is the underlying LP profile of anindividual when considering their response to dietary fat?
4 How much fat?
The current United States National Cholesterol EducationProgram (NCEP) and AHA Dietary Guidelines encompassthe best and most relevant guide for fat and cholesterolintake [9]. These heart-healthy programs recommend lim-iting fat to 25–35%en, with prudence favoring the low endsince 35%en from fat, which is common in the Americandiet, tends to have the undesirable consequence of raisingTC and LDL. Decreasing fat intake to 20%en also can be
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Eur. J. Lipid Sci. Technol. 109 (2007) 453–464 455
troublesome because, even though TC and LDL maydecline, HDL may also fall, while triglycerides (TG) tend torise. This combination typically leads to smaller and den-ser atherogenic LDL particles that induce CHD [10]. Thisadverse LP profile can often be traced to the dietary fattyacid imbalance between S : M : P, which is typically dis-torted at 20%en. That is, the essential fatty acid pool ofPUFA can easily become limited, thereby distorting the LPmetabolism and the LP profile (see below).
5 Fats versus fatty acids
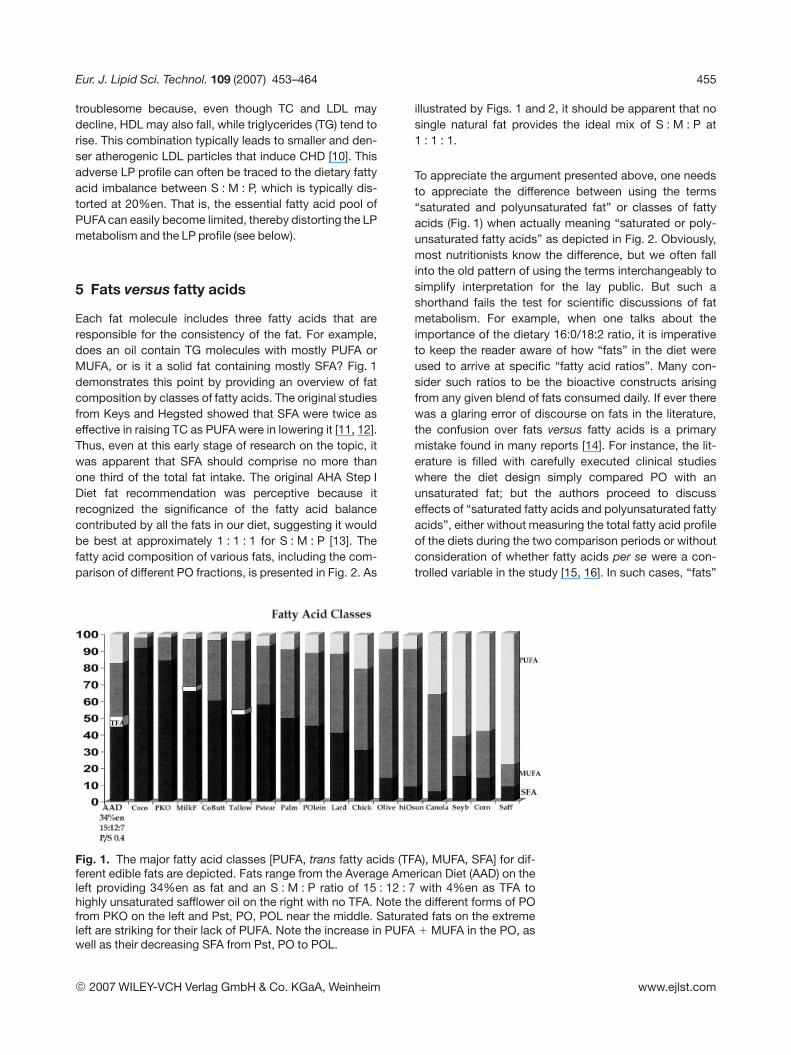
Each fat molecule includes three fatty acids that areresponsible for the consistency of the fat. For example,does an oil contain TG molecules with mostly PUFA orMUFA, or is it a solid fat containing mostly SFA? Fig. 1demonstrates this point by providing an overview of fatcomposition by classes of fatty acids. The original studiesfrom Keys and Hegsted showed that SFA were twice aseffective in raising TC as PUFA were in lowering it [11, 12].Thus, even at this early stage of research on the topic, itwas apparent that SFA should comprise no more thanone third of the total fat intake. The original AHA Step IDiet fat recommendation was perceptive because itrecognized the significance of the fatty acid balancecontributed by all the fats in our diet, suggesting it wouldbe best at approximately 1 : 1 : 1 for S : M : P [13]. Thefatty acid composition of various fats, including the com-parison of different PO fractions, is presented in Fig. 2. As
illustrated by Figs. 1 and 2, it should be apparent that nosingle natural fat provides the ideal mix of S : M : P at1 : 1 : 1.
To appreciate the argument presented above, one needsto appreciate the difference between using the terms“saturated and polyunsaturated fat” or classes of fattyacids (Fig. 1) when actually meaning “saturated or poly-unsaturated fatty acids” as depicted in Fig. 2. Obviously,most nutritionists know the difference, but we often fallinto the old pattern of using the terms interchangeably tosimplify interpretation for the lay public. But such ashorthand fails the test for scientific discussions of fatmetabolism. For example, when one talks about theimportance of the dietary 16:0/18:2 ratio, it is imperativeto keep the reader aware of how “fats” in the diet wereused to arrive at specific “fatty acid ratios”. Many con-sider such ratios to be the bioactive constructs arisingfrom any given blend of fats consumed daily. If ever therewas a glaring error of discourse on fats in the literature,the confusion over fats versus fatty acids is a primarymistake found in many reports [14]. For instance, the lit-erature is filled with carefully executed clinical studieswhere the diet design simply compared PO with anunsaturated fat; but the authors proceed to discusseffects of “saturated fatty acids and polyunsaturated fattyacids”, either without measuring the total fatty acid profileof the diets during the two comparison periods or withoutconsideration of whether fatty acids per se were a con-trolled variable in the study [15, 16]. In such cases, “fats”
Fig. 1. The major fatty acid classes [PUFA, trans fatty acids (TFA), MUFA, SFA] for dif-ferent edible fats are depicted. Fats range from the Average American Diet (AAD) on theleft providing 34%en as fat and an S : M : P ratio of 15 : 12 : 7 with 4%en as TFA tohighly unsaturated safflower oil on the right with no TFA. Note the different forms of POfrom PKO on the left and Pst, PO, POL near the middle. Saturated fats on the extremeleft are striking for their lack of PUFA. Note the increase in PUFA 1 MUFA in the PO, aswell as their decreasing SFA from Pst, PO to POL.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

456 K. C. Hayes and P. Khosla Eur. J. Lipid Sci. Technol. 109 (2007) 453–464
Fig. 2. Refer to Fig. 1 for orien-tation, but note the inclusion ofspecific fatty acids here. Thelack of 18:2 in coconut oil andPKO is conspicuous comparedto the more generous amount inPOL, which has the greatest18:2 content among all thesaturated fats. This fact aloneplays a major role in its biologi-cal effectiveness. See text fordetails. Only canola oil and SBOhave appreciable 18:3n-3, mak-ing them the preferred fats forblending with POL, which alsohas the least amount of SFAamong the PO.
may have elicited differences, but it is not possible to at-tribute differences to “specific fatty acids” because two ormore fatty acids were altered simultaneously during theexchange. Thus, the question becomes an issue of toofew PUFA or too many SFA? The answer implies differentsolutions to correct the problem, which is what nutrition isall about.
The ambiguity surrounding SFA relates to their beinglumped together within a single class, even though differ-ent SFA, as presented in whole fats, exert disparateeffects on LP and hemostatic parameters, such thateffects of 12:0 1 14:0 are greater than 16:0 greater than18:0 when isolated by regression analysis for individualfatty acids [12, 17, 18]. As pointed out below, a focus onindividual fatty acids is probably too simplistic. Histori-cally, most nutritionists, and certainly the lay press, haveused the terms “saturated fats” and “saturated fattyacids” interchangeably without distinguishing betweenindividual SFA, as well. But PKO (like coconut oil and milkfat) is not a group of SFA; they represent saturated fatsrich in certain SFA, poor in PUFA, with each fatty acidsituated on a specific carbon of glycerol, the total mix ofwhich comprises the fat. Most importantly, PKO (andcoconut oil and milk fat) have almost no PUFA, so onecannot say whether the cholesterol-raising responseobserved when consuming them is due to too many SFAor not enough PUFA unless studies are specificallydesigned to test the point.
6 SFA, PUFA, and trans fatty acids
Within the concept of a “balance” among classes ofsaturates, monounsaturates, and polyunsaturates (i.e. thewhole fats and classes of fatty acids) is the issue of whichspecific fatty acids, among the SFA or PUFA, are most
suitable for consumption, and whether PO can reason-ably contribute to the pool. For example, four SFA typi-cally constitute the major pool of fatty acids in saturatedfat TG molecules (remember that each fat molecule con-tains three fatty acids). These are lauric (12:0), myristic(14:0), palmitic (16:0), and stearic (18:0) acids. Saturatesin PO are mostly 16:0 with a small amount of 18:0, whilePKO is particularly rich in 12:0 and 14:0 (Tabs. 1, 2).Similarly, several specific PUFA of the omega 3 (n-3) oromega 6 (n-6) families are the major contributors to thepool of fatty acids in a polyunsaturated fat. In vegetableoils these are linoleic (18:2n-6, the most prevalent of allPUFA) and linolenic (18:3n-3) acids. These two represent
Tab. 1. TG composition of PO, POL and Pst.
TG-MS percent
PO POL§ Pst
OLL 0.5PLL 1.9 2.6 1.0OOL 1.6 2.2 0.8POL 10.3 13.8 6.6PPL 11.2 11.6 8.7? 0.5 – 1.0OOO 3.9 5.4 2.4POO 25.1 31.1 18.2POP 34.1 26.2 45.4PPP 6.3 – 10.4SOO 2.5 1.7 1.7POS 2.0 4.7 2.9PPS 0.6 – 0.9
Source: Kalyana Sundram, unpublished data (2007).L = linoleic; O = oleic; P = palmitic; S = stearic.§ Triglyceride species for POL indicate that .85% of theTG-MS have sn2 as 18:1. Nor does POL contain tripalmi-tin (PPP), present only in PO and Pst. Most of the 18:2 (L)is at sn1,3 in all three fats.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Eur. J. Lipid Sci. Technol. 109 (2007) 453–464 457
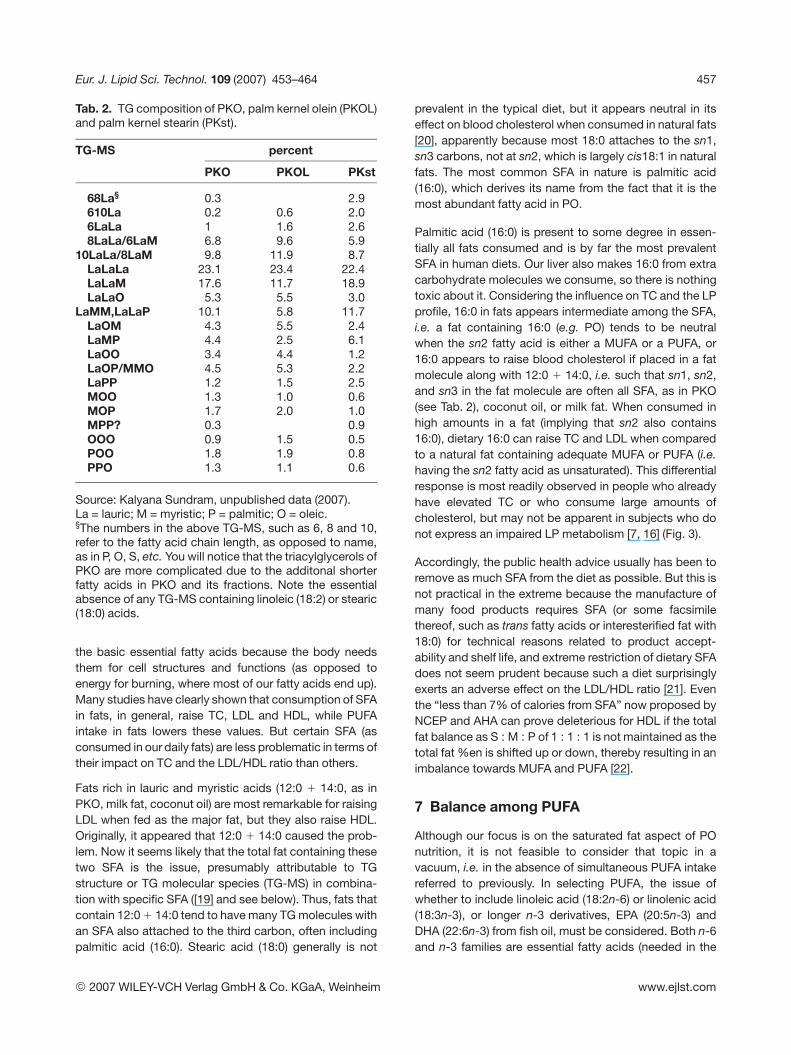
Tab. 2. TG composition of PKO, palm kernel olein (PKOL)and palm kernel stearin (PKst).
TG-MS percent
PKO PKOL PKst
68La§ 0.3 2.9610La 0.2 0.6 2.06LaLa 1 1.6 2.68LaLa/6LaM 6.8 9.6 5.9
10LaLa/8LaM 9.8 11.9 8.7LaLaLa 23.1 23.4 22.4LaLaM 17.6 11.7 18.9LaLaO 5.3 5.5 3.0
LaMM,LaLaP 10.1 5.8 11.7LaOM 4.3 5.5 2.4LaMP 4.4 2.5 6.1LaOO 3.4 4.4 1.2LaOP/MMO 4.5 5.3 2.2LaPP 1.2 1.5 2.5MOO 1.3 1.0 0.6MOP 1.7 2.0 1.0MPP? 0.3 0.9OOO 0.9 1.5 0.5POO 1.8 1.9 0.8PPO 1.3 1.1 0.6
Source: Kalyana Sundram, unpublished data (2007).La = lauric; M = myristic; P = palmitic; O = oleic.§The numbers in the above TG-MS, such as 6, 8 and 10,refer to the fatty acid chain length, as opposed to name,as in P, O, S, etc. You will notice that the triacylglycerols ofPKO are more complicated due to the additonal shorterfatty acids in PKO and its fractions. Note the essentialabsence of any TG-MS containing linoleic (18:2) or stearic(18:0) acids.
the basic essential fatty acids because the body needsthem for cell structures and functions (as opposed toenergy for burning, where most of our fatty acids end up).Many studies have clearly shown that consumption of SFAin fats, in general, raise TC, LDL and HDL, while PUFAintake in fats lowers these values. But certain SFA (asconsumed in our daily fats) are less problematic in terms oftheir impact on TC and the LDL/HDL ratio than others.
Fats rich in lauric and myristic acids (12:0 1 14:0, as inPKO, milk fat, coconut oil) are most remarkable for raisingLDL when fed as the major fat, but they also raise HDL.Originally, it appeared that 12:0 1 14:0 caused the prob-lem. Now it seems likely that the total fat containing thesetwo SFA is the issue, presumably attributable to TGstructure or TG molecular species (TG-MS) in combina-tion with specific SFA ([19] and see below). Thus, fats thatcontain 12:0 1 14:0 tend to have many TG molecules withan SFA also attached to the third carbon, often includingpalmitic acid (16:0). Stearic acid (18:0) generally is not
prevalent in the typical diet, but it appears neutral in itseffect on blood cholesterol when consumed in natural fats[20], apparently because most 18:0 attaches to the sn1,sn3 carbons, not at sn2, which is largely cis18:1 in naturalfats. The most common SFA in nature is palmitic acid(16:0), which derives its name from the fact that it is themost abundant fatty acid in PO.
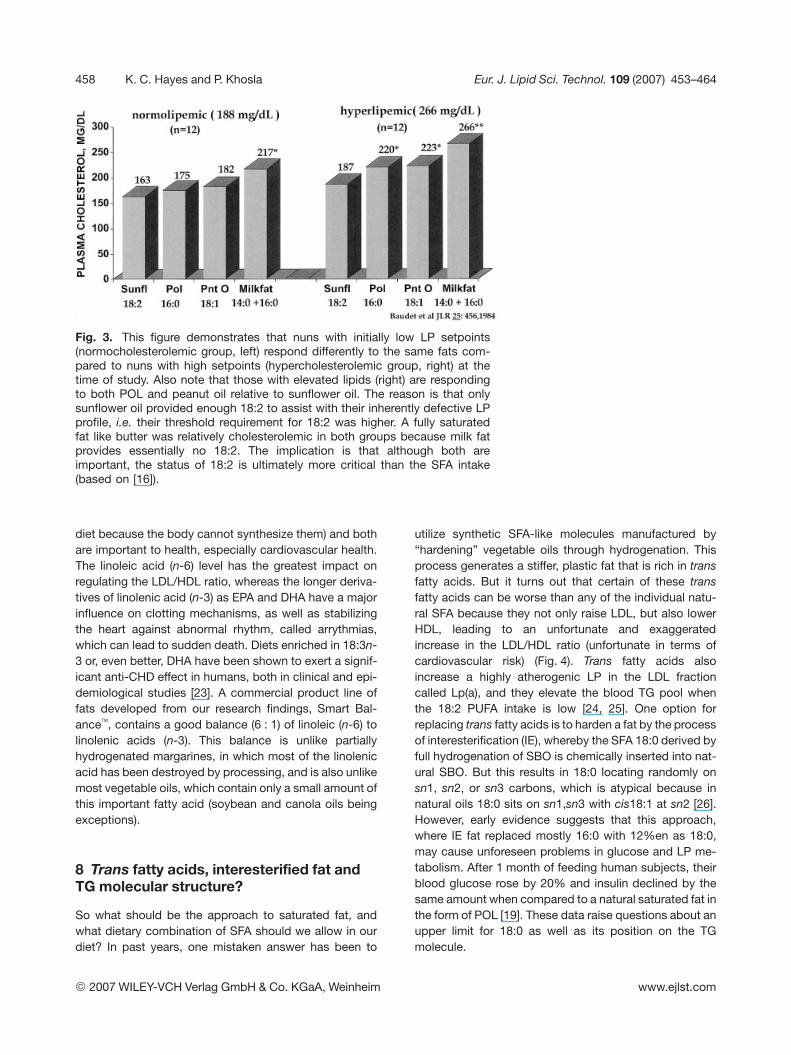
Palmitic acid (16:0) is present to some degree in essen-tially all fats consumed and is by far the most prevalentSFA in human diets. Our liver also makes 16:0 from extracarbohydrate molecules we consume, so there is nothingtoxic about it. Considering the influence on TC and the LPprofile, 16:0 in fats appears intermediate among the SFA,i.e. a fat containing 16:0 (e.g. PO) tends to be neutralwhen the sn2 fatty acid is either a MUFA or a PUFA, or16:0 appears to raise blood cholesterol if placed in a fatmolecule along with 12:0 1 14:0, i.e. such that sn1, sn2,and sn3 in the fat molecule are often all SFA, as in PKO(see Tab. 2), coconut oil, or milk fat. When consumed inhigh amounts in a fat (implying that sn2 also contains16:0), dietary 16:0 can raise TC and LDL when comparedto a natural fat containing adequate MUFA or PUFA (i.e.having the sn2 fatty acid as unsaturated). This differentialresponse is most readily observed in people who alreadyhave elevated TC or who consume large amounts ofcholesterol, but may not be apparent in subjects who donot express an impaired LP metabolism [7, 16] (Fig. 3).
Accordingly, the public health advice usually has been toremove as much SFA from the diet as possible. But this isnot practical in the extreme because the manufacture ofmany food products requires SFA (or some facsimilethereof, such as trans fatty acids or interesterified fat with18:0) for technical reasons related to product accept-ability and shelf life, and extreme restriction of dietary SFAdoes not seem prudent because such a diet surprisinglyexerts an adverse effect on the LDL/HDL ratio [21]. Eventhe “less than 7% of calories from SFA” now proposed byNCEP and AHA can prove deleterious for HDL if the totalfat balance as S : M : P of 1 : 1 : 1 is not maintained as thetotal fat %en is shifted up or down, thereby resulting in animbalance towards MUFA and PUFA [22].
7 Balance among PUFA
Although our focus is on the saturated fat aspect of POnutrition, it is not feasible to consider that topic in avacuum, i.e. in the absence of simultaneous PUFA intakereferred to previously. In selecting PUFA, the issue ofwhether to include linoleic acid (18:2n-6) or linolenic acid(18:3n-3), or longer n-3 derivatives, EPA (20:5n-3) andDHA (22:6n-3) from fish oil, must be considered. Both n-6and n-3 families are essential fatty acids (needed in the
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
458 K. C. Hayes and P. Khosla Eur. J. Lipid Sci. Technol. 109 (2007) 453–464
Fig. 3. This figure demonstrates that nuns with initially low LP setpoints(normocholesterolemic group, left) respond differently to the same fats com-pared to nuns with high setpoints (hypercholesterolemic group, right) at thetime of study. Also note that those with elevated lipids (right) are respondingto both POL and peanut oil relative to sunflower oil. The reason is that onlysunflower oil provided enough 18:2 to assist with their inherently defective LPprofile, i.e. their threshold requirement for 18:2 was higher. A fully saturatedfat like butter was relatively cholesterolemic in both groups because milk fatprovides essentially no 18:2. The implication is that although both areimportant, the status of 18:2 is ultimately more critical than the SFA intake(based on [16]).
diet because the body cannot synthesize them) and bothare important to health, especially cardiovascular health.The linoleic acid (n-6) level has the greatest impact onregulating the LDL/HDL ratio, whereas the longer deriva-tives of linolenic acid (n-3) as EPA and DHA have a majorinfluence on clotting mechanisms, as well as stabilizingthe heart against abnormal rhythm, called arrythmias,which can lead to sudden death. Diets enriched in 18:3n-3 or, even better, DHA have been shown to exert a signif-icant anti-CHD effect in humans, both in clinical and epi-demiological studies [23]. A commercial product line offats developed from our research findings, Smart Bal-ance™, contains a good balance (6 : 1) of linoleic (n-6) tolinolenic acids (n-3). This balance is unlike partiallyhydrogenated margarines, in which most of the linolenicacid has been destroyed by processing, and is also unlikemost vegetable oils, which contain only a small amount ofthis important fatty acid (soybean and canola oils beingexceptions).
8 Trans fatty acids, interesterified fat andTG molecular structure?
So what should be the approach to saturated fat, andwhat dietary combination of SFA should we allow in ourdiet? In past years, one mistaken answer has been to
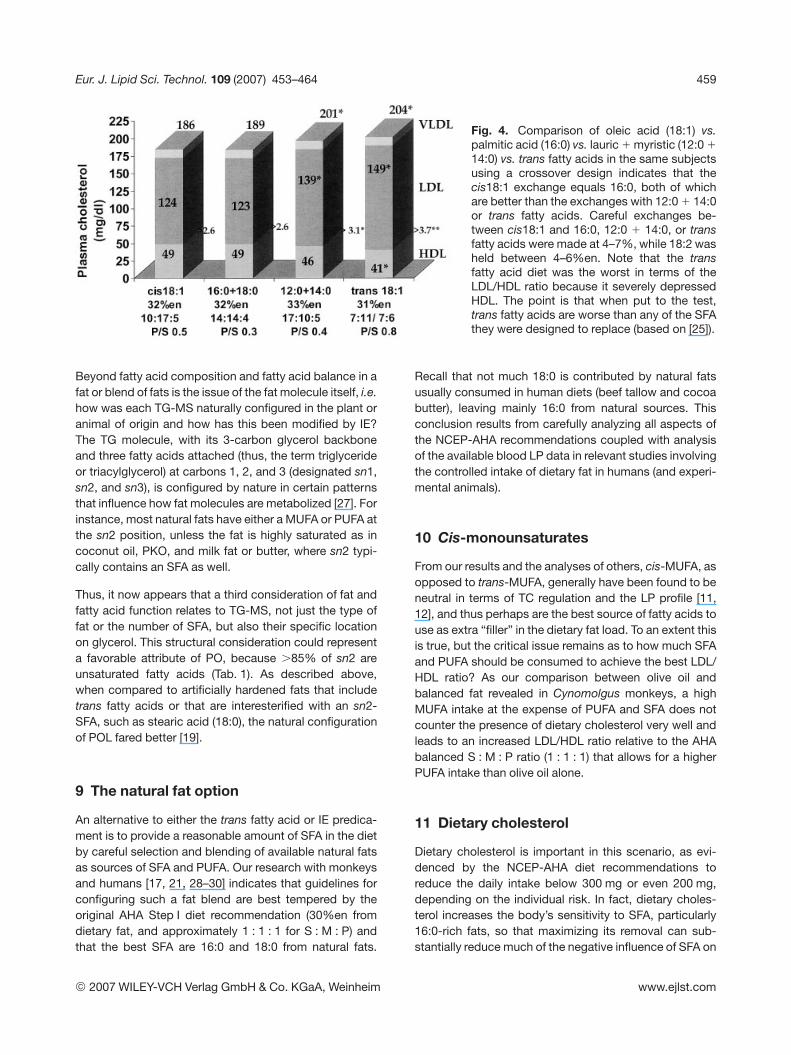
utilize synthetic SFA-like molecules manufactured by“hardening” vegetable oils through hydrogenation. Thisprocess generates a stiffer, plastic fat that is rich in transfatty acids. But it turns out that certain of these transfatty acids can be worse than any of the individual natu-ral SFA because they not only raise LDL, but also lowerHDL, leading to an unfortunate and exaggeratedincrease in the LDL/HDL ratio (unfortunate in terms ofcardiovascular risk) (Fig. 4). Trans fatty acids alsoincrease a highly atherogenic LP in the LDL fractioncalled Lp(a), and they elevate the blood TG pool whenthe 18:2 PUFA intake is low [24, 25]. One option forreplacing trans fatty acids is to harden a fat by the processof interesterification (IE), whereby the SFA 18:0 derived byfull hydrogenation of SBO is chemically inserted into nat-ural SBO. But this results in 18:0 locating randomly onsn1, sn2, or sn3 carbons, which is atypical because innatural oils 18:0 sits on sn1,sn3 with cis18:1 at sn2 [26].However, early evidence suggests that this approach,where IE fat replaced mostly 16:0 with 12%en as 18:0,may cause unforeseen problems in glucose and LP me-tabolism. After 1 month of feeding human subjects, theirblood glucose rose by 20% and insulin declined by thesame amount when compared to a natural saturated fat inthe form of POL [19]. These data raise questions about anupper limit for 18:0 as well as its position on the TGmolecule.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Eur. J. Lipid Sci. Technol. 109 (2007) 453–464 459
Fig. 4. Comparison of oleic acid (18:1) vs.palmitic acid (16:0) vs. lauric 1 myristic (12:0 1
14:0) vs. trans fatty acids in the same subjectsusing a crossover design indicates that thecis18:1 exchange equals 16:0, both of whichare better than the exchanges with 12:0 1 14:0or trans fatty acids. Careful exchanges be-tween cis18:1 and 16:0, 12:0 1 14:0, or transfatty acids were made at 4–7%, while 18:2 washeld between 4–6%en. Note that the transfatty acid diet was the worst in terms of theLDL/HDL ratio because it severely depressedHDL. The point is that when put to the test,trans fatty acids are worse than any of the SFAthey were designed to replace (based on [25]).
Beyond fatty acid composition and fatty acid balance in afat or blend of fats is the issue of the fat molecule itself, i.e.how was each TG-MS naturally configured in the plant oranimal of origin and how has this been modified by IE?The TG molecule, with its 3-carbon glycerol backboneand three fatty acids attached (thus, the term triglycerideor triacylglycerol) at carbons 1, 2, and 3 (designated sn1,sn2, and sn3), is configured by nature in certain patternsthat influence how fat molecules are metabolized [27]. Forinstance, most natural fats have either a MUFA or PUFA atthe sn2 position, unless the fat is highly saturated as incoconut oil, PKO, and milk fat or butter, where sn2 typi-cally contains an SFA as well.
Thus, it now appears that a third consideration of fat andfatty acid function relates to TG-MS, not just the type offat or the number of SFA, but also their specific locationon glycerol. This structural consideration could representa favorable attribute of PO, because .85% of sn2 areunsaturated fatty acids (Tab. 1). As described above,when compared to artificially hardened fats that includetrans fatty acids or that are interesterified with an sn2-SFA, such as stearic acid (18:0), the natural configurationof POL fared better [19].
9 The natural fat option
An alternative to either the trans fatty acid or IE predica-ment is to provide a reasonable amount of SFA in the dietby careful selection and blending of available natural fatsas sources of SFA and PUFA. Our research with monkeysand humans [17, 21, 28–30] indicates that guidelines forconfiguring such a fat blend are best tempered by theoriginal AHA Step I diet recommendation (30%en fromdietary fat, and approximately 1 : 1 : 1 for S : M : P) andthat the best SFA are 16:0 and 18:0 from natural fats.
Recall that not much 18:0 is contributed by natural fatsusually consumed in human diets (beef tallow and cocoabutter), leaving mainly 16:0 from natural sources. Thisconclusion results from carefully analyzing all aspects ofthe NCEP-AHA recommendations coupled with analysisof the available blood LP data in relevant studies involvingthe controlled intake of dietary fat in humans (and experi-mental animals).
10 Cis-monounsaturates
From our results and the analyses of others, cis-MUFA, asopposed to trans-MUFA, generally have been found to beneutral in terms of TC regulation and the LP profile [11,12], and thus perhaps are the best source of fatty acids touse as extra “filler” in the dietary fat load. To an extent thisis true, but the critical issue remains as to how much SFAand PUFA should be consumed to achieve the best LDL/HDL ratio? As our comparison between olive oil andbalanced fat revealed in Cynomolgus monkeys, a highMUFA intake at the expense of PUFA and SFA does notcounter the presence of dietary cholesterol very well andleads to an increased LDL/HDL ratio relative to the AHAbalanced S : M : P ratio (1 : 1 : 1) that allows for a higherPUFA intake than olive oil alone.
11 Dietary cholesterol
Dietary cholesterol is important in this scenario, as evi-denced by the NCEP-AHA diet recommendations toreduce the daily intake below 300 mg or even 200 mg,depending on the individual risk. In fact, dietary choles-terol increases the body’s sensitivity to SFA, particularly16:0-rich fats, so that maximizing its removal can sub-stantially reduce much of the negative influence of SFA on
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
460 K. C. Hayes and P. Khosla Eur. J. Lipid Sci. Technol. 109 (2007) 453–464
the LP profile. PUFA, on the other hand, are the major fattyacids able to actually offset the negative impact of dietarycholesterol because linoleic acid (18:2n-6) increases theremoval of plasma LDL, the main LP that is increased bydietary cholesterol [7, 17].
12 PO and its fractions
With the above primer on fats and cholesterol metabo-lism, we can now turn our attention to PO and it fractionsas sources of fatty acids and their metabolic effects on LPwhen consumed.
13 Fatty acid balance affects the LDL/HDLratio
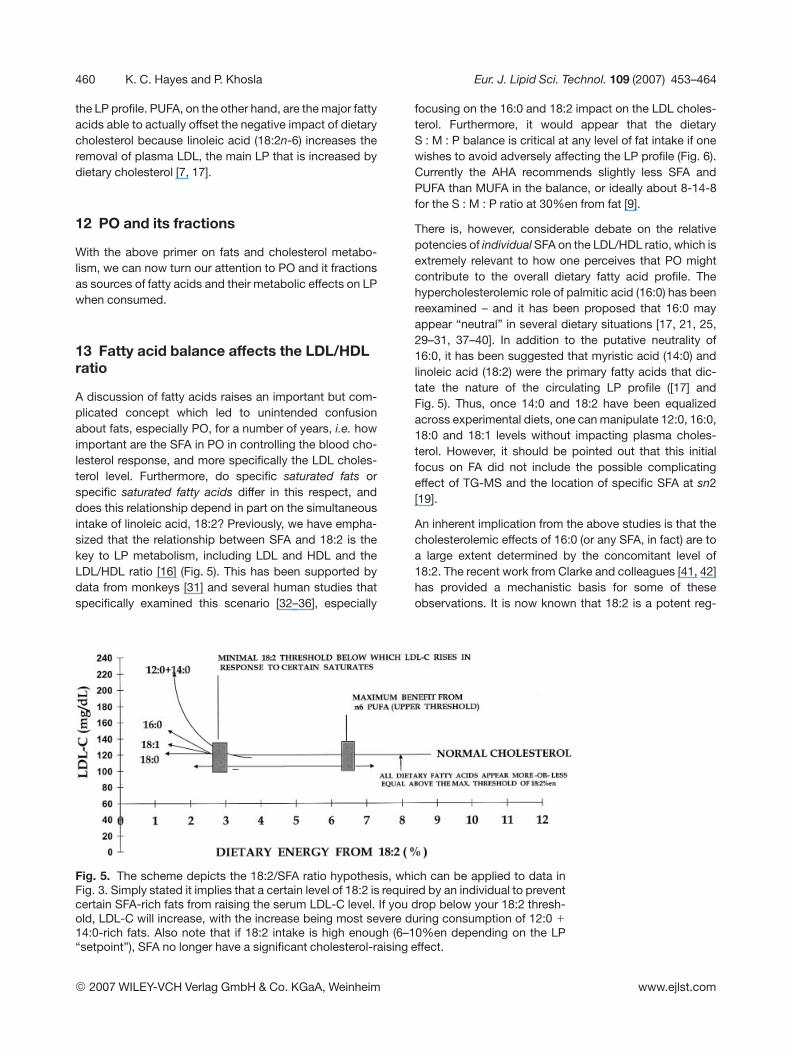
A discussion of fatty acids raises an important but com-plicated concept which led to unintended confusionabout fats, especially PO, for a number of years, i.e. howimportant are the SFA in PO in controlling the blood cho-lesterol response, and more specifically the LDL choles-terol level. Furthermore, do specific saturated fats orspecific saturated fatty acids differ in this respect, anddoes this relationship depend in part on the simultaneousintake of linoleic acid, 18:2? Previously, we have empha-sized that the relationship between SFA and 18:2 is thekey to LP metabolism, including LDL and HDL and theLDL/HDL ratio [16] (Fig. 5). This has been supported bydata from monkeys [31] and several human studies thatspecifically examined this scenario [32–36], especially
focusing on the 16:0 and 18:2 impact on the LDL choles-terol. Furthermore, it would appear that the dietaryS : M : P balance is critical at any level of fat intake if onewishes to avoid adversely affecting the LP profile (Fig. 6).Currently the AHA recommends slightly less SFA andPUFA than MUFA in the balance, or ideally about 8-14-8for the S : M : P ratio at 30%en from fat [9].
There is, however, considerable debate on the relativepotencies of individual SFA on the LDL/HDL ratio, which isextremely relevant to how one perceives that PO mightcontribute to the overall dietary fatty acid profile. Thehypercholesterolemic role of palmitic acid (16:0) has beenreexamined – and it has been proposed that 16:0 mayappear “neutral” in several dietary situations [17, 21, 25,29–31, 37–40]. In addition to the putative neutrality of16:0, it has been suggested that myristic acid (14:0) andlinoleic acid (18:2) were the primary fatty acids that dic-tate the nature of the circulating LP profile ([17] andFig. 5). Thus, once 14:0 and 18:2 have been equalizedacross experimental diets, one can manipulate 12:0, 16:0,18:0 and 18:1 levels without impacting plasma choles-terol. However, it should be pointed out that this initialfocus on FA did not include the possible complicatingeffect of TG-MS and the location of specific SFA at sn2[19].
An inherent implication from the above studies is that thecholesterolemic effects of 16:0 (or any SFA, in fact) are toa large extent determined by the concomitant level of18:2. The recent work from Clarke and colleagues [41, 42]has provided a mechanistic basis for some of theseobservations. It is now known that 18:2 is a potent reg-
Fig. 5. The scheme depicts the 18:2/SFA ratio hypothesis, which can be applied to data inFig. 3. Simply stated it implies that a certain level of 18:2 is required by an individual to preventcertain SFA-rich fats from raising the serum LDL-C level. If you drop below your 18:2 thresh-old, LDL-C will increase, with the increase being most severe during consumption of 12:0 1
14:0-rich fats. Also note that if 18:2 intake is high enough (6–10%en depending on the LP“setpoint”), SFA no longer have a significant cholesterol-raising effect.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Eur. J. Lipid Sci. Technol. 109 (2007) 453–464 461
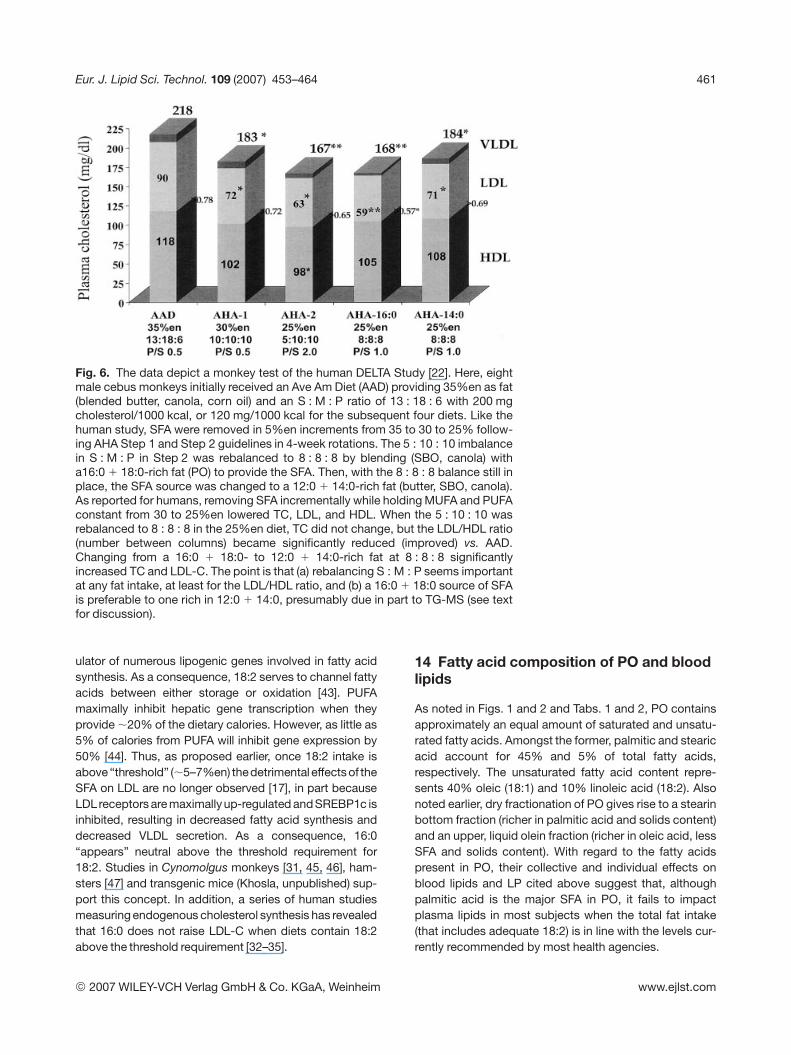
Fig. 6. The data depict a monkey test of the human DELTA Study [22]. Here, eightmale cebus monkeys initially received an Ave Am Diet (AAD) providing 35%en as fat(blended butter, canola, corn oil) and an S : M : P ratio of 13 : 18 : 6 with 200 mgcholesterol/1000 kcal, or 120 mg/1000 kcal for the subsequent four diets. Like thehuman study, SFA were removed in 5%en increments from 35 to 30 to 25% follow-ing AHA Step 1 and Step 2 guidelines in 4-week rotations. The 5 : 10 : 10 imbalancein S : M : P in Step 2 was rebalanced to 8 : 8 : 8 by blending (SBO, canola) witha16:0 1 18:0-rich fat (PO) to provide the SFA. Then, with the 8 : 8 : 8 balance still inplace, the SFA source was changed to a 12:0 1 14:0-rich fat (butter, SBO, canola).As reported for humans, removing SFA incrementally while holding MUFA and PUFAconstant from 30 to 25%en lowered TC, LDL, and HDL. When the 5 : 10 : 10 wasrebalanced to 8 : 8 : 8 in the 25%en diet, TC did not change, but the LDL/HDL ratio(number between columns) became significantly reduced (improved) vs. AAD.Changing from a 16:0 1 18:0- to 12:0 1 14:0-rich fat at 8 : 8 : 8 significantlyincreased TC and LDL-C. The point is that (a) rebalancing S : M : P seems importantat any fat intake, at least for the LDL/HDL ratio, and (b) a 16:0 1 18:0 source of SFAis preferable to one rich in 12:0 1 14:0, presumably due in part to TG-MS (see textfor discussion).
ulator of numerous lipogenic genes involved in fatty acidsynthesis. As a consequence, 18:2 serves to channel fattyacids between either storage or oxidation [43]. PUFAmaximally inhibit hepatic gene transcription when theyprovide ,20% of the dietary calories. However, as little as5% of calories from PUFA will inhibit gene expression by50% [44]. Thus, as proposed earlier, once 18:2 intake isabove “threshold” (,5–7%en) thedetrimental effects of theSFA on LDL are no longer observed [17], in part becauseLDLreceptors are maximallyup-regulated and SREBP1c isinhibited, resulting in decreased fatty acid synthesis anddecreased VLDL secretion. As a consequence, 16:0“appears” neutral above the threshold requirement for18:2. Studies in Cynomolgus monkeys [31, 45, 46], ham-sters [47] and transgenic mice (Khosla, unpublished) sup-port this concept. In addition, a series of human studiesmeasuring endogenous cholesterol synthesis has revealedthat 16:0 does not raise LDL-C when diets contain 18:2above the threshold requirement [32–35].
14 Fatty acid composition of PO and bloodlipids
As noted in Figs. 1 and 2 and Tabs. 1 and 2, PO containsapproximately an equal amount of saturated and unsatu-rated fatty acids. Amongst the former, palmitic and stearicacid account for 45% and 5% of total fatty acids,respectively. The unsaturated fatty acid content repre-sents 40% oleic (18:1) and 10% linoleic acid (18:2). Alsonoted earlier, dry fractionation of PO gives rise to a stearinbottom fraction (richer in palmitic acid and solids content)and an upper, liquid olein fraction (richer in oleic acid, lessSFA and solids content). With regard to the fatty acidspresent in PO, their collective and individual effects onblood lipids and LP cited above suggest that, althoughpalmitic acid is the major SFA in PO, it fails to impactplasma lipids in most subjects when the total fat intake(that includes adequate 18:2) is in line with the levels cur-rently recommended by most health agencies.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
462 K. C. Hayes and P. Khosla Eur. J. Lipid Sci. Technol. 109 (2007) 453–464
Numerous dietary intervention studies designed to eval-uate the ability of PO to influence blood cholesterol havebeen reported. The results from these studies have oftenbeen conflicting, primarily because some were carefullydesigned to “evaluate the effects of individual SFA” andused PO “as the primary source of palmitic acid”, whileothers compared PO per se with other dietary oils or fats(without controlling all fatty acids being consumed). Thus,it has not always been possible to separate the effect offat from specific fatty acids. Early studies evaluated POwithin the context of typical Western diets prevalent at thetime (,40% of total calories from fat). For example, whenmale subjects were fed liquid-formula diets with 40% ofcalories supplied exclusively by a single fat, PO washypercholesterolemic relative to high-oleic safflower,high-linoleic safflower oil, and a high-stearate inter-esterified fat, but hypocholesterolemic relative to coconutoil. In all instances concern for the simultaneous intake ofPUFA was ignored, and the primary effect of PO (withrelatively less 18:2) was to raise LDL levels [15, 48, 49].
Recent studies [21, 25, 29, 32–35, 40] have evaluated POwithin the context of prudent diets both in terms of totalfat (,30% of total calories) and SFA content (,10% oftotal calories). In these studies, using mildly hypercholes-terolemic to normocholesterolemic younger subjects, thecholesterol-raising attribute of PO was either muted ordisappeared. Under such conditions, stable-isotopestudies have revealed no effects on endogenous choles-terol synthesis [32–35]. In contrast to the older studies,recent trials have used POL, the liquid fraction of PO,rather than PO itself. In such scenarios, direct compar-isons of POL with some of the MUFA-rich vegetable oils(olive and canola oils) have revealed comparable effectson plasma lipids and LP [25, 36, 40]. It is unclear whetherthe switch to POL, with its higher unsaturated fatty acidcomposition (reduced palmitic, increased oleic and lino-leic acids), or the removal of tripalmitin (PPP, see Tab. 1)resulted in the muted cholesterol response in the sub-jects.
The neutral effect of POL typically observed in studies inwhich total fat represented ,30% of total calories may inpart be explained by the studies of Hayes and coworkers[17, 30, 50–53] described above. That is, individual SFAdo not appear equal in their ability to induce hypercho-lesterolemia, with saturated fats containing lauric andmyristic acid (12:0 and 14:0) being the most cholester-olemic, followed by fats containing palmitic (16:0) andstearic (18:0) acids.
Thus, POL, which is essentially devoid of 12:0 and 14:0(Tab. 1), appears to have sufficient 18:2 to counter mostdeleterious effects of the 16:0 present. In the abovemodel, the monounsaturated oleic acid (18:1) is believed
to be neutral. The above concept of using an amount of18:2 to essentially balance SFA in the diet is ingrained intomost current dietary recommendations, which essentiallyadvocate equivalent amounts of SFA and PUFA, with theremaining fatty acids being derived from MUFA.
To illustrate the above concept further, Tab. 3 represents atheoretical calculation revealing the maximum amount ofPOL allowable to supply ,10%en as SFA in a diet with 20,25, 30, or 40%en from fat. In these situations, POL couldrepresent 100, 87, 72, 62, and 54% of the fat, respec-tively. For example, when total fat contributes 30% oftotal calories, 72% of the dietary fat as POL would resultin an SFA intake of 10% of total calories. On the otherhand, it is apparent that in comparisons where total fatwas 40% of calories and POL was the sole fat utilized, theSFA content of the diet would greatly exceed the recom-mended levels of 10% of calories. Furthermore, the SFAwould also exceed any 18:2 in the diet. In the studieswhere total fat was set at 30–35% of total calories, even ifPOL represented up to 60% of the total dietary fat, theSFA content of the diet would still be within the recom-mended levels, allowing for the presence of adequate18:2 to balance the 16:0 derived from POL. Thus, asTab. 2 suggests, POL can be used as the major dietary fatat essentially any fat level (20 to 35% of total calories),and the SFA would still be within the recommendedlevels.
Tab. 3. Maximizing POL in fat formulations.
% calories % calories from total fatfrom SFA 20 25 30 35 40
Maximum % POL allowed
5 54 43 36 31 276 65 52 43 37 327 76 61 51 43 388 87 69 58 49 439 97 78 65 56 49
10 100§ 87 72 62 54
Tab. 3 depicts the maximum percentage of total dietaryfat that can be provided as POL (in italics) to achieve theindicated intakes for total SFA (as % calories on left),assuming that POL is the only source of SFA. For exam-ple, when total fat contributes 25% of total calories, 43%of the dietary fat as POL will result in an SFA intake below5% of total calories, whereas 61% of the dietary fat asPOL will result in SFA intake below 7% of total calories,etc. The calculation assumes that the remaining fat willhave a low SFA content, e.g. soybean, canola, safflower,corn oil, which would invariably provide significant linoleicacid. Note that at total fat intake of 20% of daily calories,100%§ of the dietary fat as POL would provide only 9.2%of calories from SFA.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

Eur. J. Lipid Sci. Technol. 109 (2007) 453–464 463
The above concept of balancing different fatty acids, hastaken on added significance since optimal fatty acidblends are believed to be important for maintaining bothplasma LDL and HDL levels (see [54] for the latest review).Since SFA raise both LDL and HDL, cis-MUFA lower LDL,trans-MUFA raise LDL and lower HDL, while PUFA lowerLDL and HDL (at high intakes), teleologically it is apparentthat optimal blends of fatty acids can be selected tomaximally lower LDL while maintaining or raising HDL.Sundram et al. [20] have presented evidence that suchblends are indeed possible.
Finally, with regard to PO and its fatty acid composition,the recent rush to remove trans fatty acids from foodproducts has led to a scramble to find replacement-suit-able fats. Legislation by various countries worldwide hasadvocated elimination of trans fatty acids from the marketplace. In the United States, the Food and Drug Adminis-tration called for mandatory labeling of trans fatty acidswhich came into effect in 2006. Thus, the food industry isbeing forced to find alternatives to trans fatty acids for usein solid-fat formulations. Various options are being con-sidered, including genetic engineering of oil crops, fullhydrogenation of liquid vegetable oils followed by enzy-matic or chemical interesterification with a suitable liquidvegetable oil, as well as the use of naturally occurring fatsand oils to provide a source of SFA, needed for productsrequiring a certain amount of solid fat content. In the latteroption, natural PO should find an even broader accep-tance due to its 50% content of SFA.
15 Conclusions
PO is a nutritious fat that has been consumed for cen-turies. Although a saturated fat by natural design withabout 50% of its fatty acids as SFA, it is possible to blendunsaturated oils with such saturated fats to developexcellent products that can be beneficial to lipid metabo-lism, even improving the LP profile. Alternatives to naturalsaturated fats, such as trans fatty acids and IE fat havepotentially negative metabolic effects above certainintakes and will require additional scrutiny if they are to befound acceptable substitutes to natural fat blends.
References
[1] R. Sambanthamurthi, K. Sundram, Y. Tan: Chemistry andbiochemistry of palm oil. Prog Lipid Res. 2000, 39, 507–558.
[2] R. Manorama, G. N. Brahmam, C. Rukmini: Red palm oil as asource of beta-carotene for combating vitamin A deficiency.Plant Foods Hum Nutr. 1996, 49, 75–82.
[3] M. E. van Stuijvenberg, M. A. Dhansay, C. J. Lombard, M.Faber, A. J. Benadé: The effect of a biscuit with red palm oil asa source of beta-carotene on the vitamin A status of primary
school children: A comparison with beta-carotene from asynthetic source in a randomised controlled trial. Eur J ClinNutr. 2001, 55, 657–662.
[4] Y. Sivan, Y. Alwin Jayakumar, C. Arumughan, A. Sundaresan,A. Jaylekshmy, K. P. Suja, D. R. Soban Kumar, S. S. Deepa,M. Damodaran, C. R. Soman, V. Raman Kutty, P. SankarSarma: Impact of vitamin A supplementation through differ-ent dosages of red palm oil and retinol palmitate on pre-school children. J Trop Pediatr. 2002, 48, 24–28.
[5] E. Hedrén, G. Mulokozi, U. Svanberg: In vitro accessibility ofcarotenes from green leafy vegetables cooked with sun-flower oil or red palm oil. Int J Food Sci Nutr. 2002, 53, 445–453.
[6] N. M. Zagré, F. Delpeuch, P. Traissac, H. Delisle: Red palm oilas a source of vitamin A for mothers and children: Impact ofa pilot project in Burkina Faso. Public Health Nutr. 2003, 6,733–742.
[7] K. C. Hayes: Saturated fats and blood lipids: New slant onan old story. Can J Cardiol. 1995, 11 Suppl G, 39G–46G.
[8] P. Khosla: Palm oil: A nutritional overview. J Agro Food Ind.2006, 17, 21–23.
[9] A. H. Lichtenstein, L. J. Appel, M. Brands, M. Carnethon, S.Daniels, H. A. French, B. Franklin, P. Kris-Etherton, W. S.Harris, B. Howard, N. Karanja, M. Lefevre, L. Rudel, F.Sacks, L. Van Horn, M. Winston, J. Wylie-Rosett: Summaryof American Heart Association diet and lifestyle recommen-dations revision 2006. Arterioscler Thromb Vasc Biol. 2006,26, 2186–2191.
[10] B. Lamarche, I. Lemieux, J. P. Després: The small, denseLDL phenotype and the risk of coronary heart disease: Epi-demiology, patho-physiology and therapeutic aspects. Dia-betes Metab. 1999, 25, 199–211.
[11] A. Keys, J. T. Anderson, F. Grande: Serum cholesterol re-sponse to changes in the diet. IV. Particular saturated fattyacids in the diet. Metabolism 1965, 14, 776–787.
[12] D. M. Hegsted, R. B. McGandy, M. L. Myers, F. J. Stare:Quantitative effects of dietary fat on serum cholesterol inman. Am J Clin Nutr. 1965, 17, 281–295.
[13] R. M. Krauss, R. H. Eckel, B. Howard, L. J. Appel, S. R.Daniels, R. J. Deckelbaum, J. W. Jr. Erdman, P. Kris-Ether-ton, I. J. Goldberg, T. A. Kotchen, A. H. Lichtenstein, W. E.Mitch, R. Mullis, K. Robinson, J. Wylie-Rosett, S. St. Jeor, J.Suttie, D. L. Tribble, T. L. Bazzarre: AHA Dietary Guidelines:Revision 2000: A statement for healthcare professionalsfrom the Nutrition Committee of the American Heart Asso-ciation. Circulation 2000, 102, 2284–2299.
[14] F. B. Hu, M. J. Stampfer, J. E. Manson, E. Rimm, G. A. Col-ditz, B. A. Rosner, C. H. Hennekens, W. C. Willett: Dietary fatintake and the risk of coronary heart disease in women. NEngl J Med. 1997, 337, 1491–1499.
[15] F. H. Mattson, S. M. Grundy: Comparison of effects of die-tary saturated, monounsaturated, and polyunsaturated fattyacids on plasma lipids and lipoproteins in man. J Lipid Res.1985, 26, 194–202.
[16] M. F. Baudet, C. Dachet, M. Lasserre, O. Esteva, B. Jacotot:Modification in the composition and metabolic properties ofhuman low density and high density lipoproteins by differentdietary fats. J Lipid Res. 1984, 25, 456–468.
[17] K. C. Hayes, P. Khosla: Dietary fatty acid thresholds andcholesterolemia. FASEB J. 1992, 6, 2600–2607.
[18] P. Khosla, K. Sundram: Effects of dietary fatty acid compo-sition on plasma cholesterol. Prog Lipid Res. 1996, 35, 93–132.
[19] K. Sundram, T. Karupaiah, K. C. Hayes: Stearic acid-richinteresterified fat and trans-rich fat raise the LDL/HDL ratioand plasma glucose relative to palm olein in humans. NutrMetab. 2007, 4, 3.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

464 K. C. Hayes and P. Khosla Eur. J. Lipid Sci. Technol. 109 (2007) 453–464
[20] S. Yu, J. Derr, T. D. Etherton, P. M. Kris-Etherton: Plasmacholesterol-predictive equations demonstrate that stearicacid is neutral and monounsaturated fatty acids are hypo-cholesterolemic. Am J Clin Nutr. 1995, 61, 1129–1139.
[21] K. Sundram, K. C. Hayes, O. H. Siru: Both dietary 18:2 and16:0 may be required to improve the serum LDL/HDL cho-lesterol ratio in normocholesterolemic men. J Nutr Biochem.1995, 6, 179–187.
[22] H. N. Ginsberg, P. Kris-Etherton, B. Dennis, P. J. Elmer, A.Ershow, M. Lefevre, T. Pearson, P. Roheim, R. Ramakrish-nan, K. Stewart, P. Stewart, K. Phillips, N. Andersen: Effectsof reducing dietary saturated fatty acids on plasma lipidsand lipoproteins in healthy subjects: The DELTA Study, pro-tocol 1. Arterioscler Thromb Vasc Biol. 1998, 18, 441–449.
[23] A. Leaf, J. X. Kang, Y. F. Xiao: Omega-3 fatty acids and ven-tricular arrhythmias. World Rev Nutr Diet. 2005, 94, 129–138.
[24] M. B. Katan, P. L. Zock, R. P. Mensink: Trans fatty acids andtheir effects on lipoproteins in humans. Annu Rev Nutr. 1995,15, 473–493.
[25] K. Sundram, A. Ismail, K. C. Hayes, R. Jeyamalar, R. Path-manathan: Trans (elaidic) fatty acids adversely affect thelipoprotein profile relative to specific saturated fatty acids inhumans. J Nutr. 1997, 127, 514S–520S.
[26] F. H. Mattson, E. S. Lutton: The specific distribution of fattyacids in the glycerides of animal and vegetable fats. J BiolChem. 1958, 233, 868–871.
[27] D. Kritchevsky, S. A. Tepper, S. C. Chen, G. W. Meijer, R. M.Krauss: Cholesterol vehicle in experimental atherosclerosis.23. Effects of specific synthetic triglycerides. Lipids 2000,35, 621–625.
[28] P. Khosla, T. Hajri, A. Pronczuk, K. C. Hayes: Replacingdietary palmitic acid with elaidic acid (t-C18:1 delta9)depresses HDL and increases CETP activity in cebus mon-keys. J Nutr. 1997, 127, 531S–536S.
[29] K. Sundram, K. C. Hayes, O. H. Siru: Dietary palmitic acidresults in lower serum cholesterol than does a lauric-myristicacid combination in normolipemic humans. Am J Clin Nutr.1994, 59, 841–846.
[30] P. Khosla, K. C. Hayes: Comparison between the effects ofdietary saturated (16:0), monounsaturated (18:1), and poly-unsaturated (18:2) fatty acids on plasma lipoprotein metab-olism in cebus and rhesus monkeys fed cholesterol-freediets. Am J Clin Nutr. 1992, 55, 51–62.
[31] S. V. Gupta, N. Yamada, T. V. Fungwe, P. Khosla: Replacing40% of dietary animal fat with vegetable oil is associatedwith lower HDL cholesterol and higher cholesterol estertransfer protein in Cynomolgus monkeys fed sufficient lino-leic acid. J Nutr. 2003, 133, 2600–2606.
[32] M. T. Clandinin, S. L. Cook, S. D. Konrad, Y. K. Goh, M. A.French: The effect of palmitic acid on lipoprotein cholesterollevels and endogenous cholesterol synthesis in hyperlipi-demic subjects. Lipids 1999, 34 Suppl., S121–S124.
[33] M. T. Clandinin, S. L. Cook, S. D. Konrad, M. A. French: Theeffect of palmitic acid on lipoprotein cholesterol levels. Int JFood Sci Nutr. 2000, 51 Suppl., S61–S71.
[34] K. Sundram, M. A. French, M. T. Clandinin: Exchanging par-tially hydrogenated fat for palmitic acid in the diet increasesLDL-cholesterol and endogenous cholesterol synthesis innormocholesterolemic women. Eur J Nutr. 2003, 42, 188–194.
[35] M. T. French, K. Sundram, M. T. Clandinin: Cholesterolaemiceffect of palmitic acid in relation to other dietary fatty acids.Asia Pac J Clin Nutr. 2002, 11 Suppl. 7, S401–S407.
[36] N. Choudhury, L. Tan, A. S. Truswell: Comparison of palmo-lein and olive oil: Effects on plasma lipids and vitamin E inyoung adults. Am J Clin Nutr. 1995, 61, 1043–1051.
[37] P. Khosla, K. C. Hayes: Dietary fat saturation in rhesusmonkeys affects LDL concentrations by modulating the in-dependent production of LDL apolipoprotein B. BiochimBiophys Acta 1991, 1083, 46–56.
[38] P. Khosla, K. C. Hayes: Dietary palmitic acid raises plasmaLDL cholesterol relative to oleic acid only at a high intake ofcholesterol. Biochim Biophys Acta 1993, 1210, 13–22.
[39] P. Khosla, T. Hajri, A. Pronczuk, K. C. Hayes: Decreasingdietary lauric and myristic acids improves plasma lipidsmore favorably than decreasing dietary palmitic acid in rhe-sus monkeys fed AHA step 1 type diets. J Nutr. 1997, 127,525S–530S.
[40] T. K. Ng, K. C. Hayes, G. F. DeWitt, M. Jegathesan, N. Sat-gunasingan, A. S. Ong, D. Tan: Dietary palmitic and oleicacids exert similar effects on serum cholesterol and lipo-protein profiles in normocholesterolemic men and women. JAm Coll Nutr. 1992, 11, 383–390.
[41] D. B. Jump, S. D. Clarke: Regulation of gene expression bydietary fat. Annu Rev Nutr. 1999, 19, 63–90.
[42] S. D. Clarke: Polyunsaturated fatty acid regulation of genetranscription: A molecular mechanism to improve the meta-bolic syndrome. J Nutr. 2001, 131, 1129–1132.
[43] J. D. Horton, J. L. Goldstein, M. S. Brown: SREBPs: Activa-tors of the complete program of cholesterol and fatty acidsynthesis in the liver. J Clin Invest. 2002, 109, 1125–1131.
[44] S. D. Clarke, D. B. Jump: Regulation of gene transcription bypolyunsaturated fatty acids. Prog Lipid Res. 1993, 32, 139–149.
[45] S. V. Gupta, P. Khosla: Pork fat and chicken fat similarlyaffect plasma lipoprotein metabolism in Cynomolgus mon-keys fed diets with adequate levels of linoleic acid. J Nutr.2000, 130, 1217–1224.
[46] S. V. Gupta, P. Khosla: Palmitic and stearic acids similarlyaffect plasma lipoprotein metabolism in Cynomolgus mon-keys fed diets with adequate levels of linoleic acid. J Nutr.2001, 131, 2115–2120.
[47] L. Ramamoorthy, S. V. Gupta, P. Khosla: Effects ofexchanging 4%en between dietary stearic and palmitic acidon hamster plasma lipoprotein metabolism. Int J Food SciNutr. 2000, 51 Suppl., S51–S59.
[48] A. Bonanome, S. M. Grundy: Effect of dietary stearic acid onplasma cholesterol and lipoprotein levels. N Engl J Med.1988, 318, 1244–1248.
[49] S. M. Grundy, G. L. Vega: Plasma cholesterol responsive-ness to saturated fatty acids. Am J Clin Nutr. 1988, 47, 822–824.
[50] A. Pronczuk, P. Khosla, K. C. Hayes: Dietary myristic, pal-mitic, and linoleic acids modulate cholesterolemia in gerbils.FASEB J. 1994, 8, 1191–1200.
[51] K. C. Hayes, A. Pronczuk, S. Lindsey, D. Diersen-Schade:Dietary saturated fatty acids (12:0, 14:0, 16:0) differ in theirimpact on plasma cholesterol and lipoproteins in nonhumanprimates. Am J Clin Nutr. 1991, 53, 491–498.
[52] A. Pronczuk, G. Patton, Z. Stephan, K. C. Hayes: Speciesvariation in the atherogenic profile of monkeys: Relationshipbetween dietary fats, lipoproteins, and platelet aggregation.Lipids 1991, 26, 213–222.
[53] S. Lindsey, J. Benattar, A. Pronczuk, K. C. Hayes: Dietarypalmitic acid (16:0) enhances high density lipoprotein cho-lesterol and low density lipoprotein receptor mRNA abun-dance in hamsters. Proc Soc Exp Biol Med. 1990, 195, 261–269.
[54] M. S. Wilke, M. T. Clandinin: Influence of dietary saturatedfatty acids on the regulation of plasma cholesterol con-centration. Lipids 2005, 40, 1207–1213.
[Received: January 8, 2007; accepted: March 5, 2007]
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Related Documents











