The calcareous nannofossil response to the end-Cretaceous warm event in the Tropical Pacific Nicolas Thibault a, ⁎, Silvia Gardin b a Department of Geography and Geology, University of Copenhagen, Øster Voldgade 10, 1350 Copenhagen C., Denmark b CR2P «Centre de Recherche sur la Paléobiodiversité et les Paléoenvironnements», UMR CNRS 7207, Université Pierre et Marie Curie Paris 6, case 104, 4, place Jussieu, 75252 Paris cedex 05, France abstract article info Article history: Received 15 September 2009 Received in revised form 3 February 2010 Accepted 26 February 2010 Available online 6 March 2010 Keywords: Late Maastrichtian Calcareous nannofossils Deccan Shatsky Rise Tropical Pacific The response of calcareous nannofossils to the end-Cretaceous warming is investigated in the Tropical Pacific DSDP Site 577A (Shatsky Rise) through the record of species richness, Shannon diversity, distribution patterns and statistical treatments. This event is marked by a strong acme of the tropical species Micula murus and is accompanied by a decrease of Placozygus spiralis, the disappearance of Biscutum constans and a decrease in bulk δ 13 C, indicative of a decrease in primary productivity. An increase in abundance of other Micula species at Site 577A and a drop in species richness recorded in all sites of Shatsky Rise, suggest stressed environmental conditions coincident with the end-Cretaceous warming. The acme of M. murus at Shatsky Rise and in the Atlantic Ocean is recorded within magnetochron C29r and correlates (1) with the intense warming as expressed in intermediate waters through the δ 18 O of benthic foraminifera, and in surface waters through poleward migration, reduced diversity and dwarfism in planktonic foraminifer assemblages, (2) with a sharp decline in marine 187 Os/ 188 Os, chemical marker of Deccan volcanic activity, and (3) with a rise in the atmospheric pCO 2 record of terrestrial plants in Texas, USA, probably triggered by Deccan volcanic degassing. However, a drop of calcareous nannofossil cool- water taxa in the Equatorial and South Atlantic, as well as a first rise in abundance of M. murus in the South Atlantic and Tropical Pacific occur in the topmost part of chron C30n, and correlates with the rise in palaeotemperature record of terrestrial plants in North Dakota, USA. This suggests that initiation of the warming in the atmosphere and surface waters may predate the striking warming of intermediate water masses by 150/200 kyr. The coincidence and the links between climate change, volcanism, geochemical and biotic events at the end of the Maastrichtian thus remain to be fully elucidated. © 2010 Elsevier B.V. All rights reserved. 1. Introduction The well-documented long-term cooling trend that started in the late Campanian and continued into the early Palaeogene (Douglas and Savin, 1975; Savin, 1977; Boersma and Schackleton, 1981) was interrupted by a short warming event between ∼65.6 and 66 Ma. 1 Cooling resumed in the last 100 kyr of the Maastrichtian (Li and Keller, 1998a). During the warm event, temperature of intermediate waters (IWs) increased globally by 2–4 °C, as inferred from the record of benthic foraminiferal oxygen stable isotopes of numerous deep-sea sites in the North Atlantic, South Atlantic, Tropical Pacific and Indian Ocean (Barrera and Savin, 1999 and references therein). A warm pulse of 3–4 °C is well-recorded in surface waters in southern high-latitude Sites 689 and 690 (Stott and Kennett, 1990; Barrera, 1994) but is poorly resolved through planktonic foraminiferal stable isotopes in tropical and temperate latitudes (Li and Keller, 1998a; Barrera and Savin, 1999). The response of the planktonic community to this event suggests that surface waters were also strongly affected by a temperature rise. In South Atlantic Site 525A, the warming pulse is accompanied by reduced species richness and Shannon diversity, dwarfing and reduced photosymbiotic activity of planktonic forami- nifera (Abramovich and Keller, 2003). Poleward migration of low- latitude planktonic foraminifer species and an increase in abundance of Contusotruncana contusa and Pseudotextularia elegans are associat- ed with this end-Maastrichtian warming (Huber and Watkins, 1992; Kucera and Malmgren, 1998; Olsson et al., 2001). In calcareous nannofossil assemblages of Sites 1258A (western equatorial Atlantic) and 525A, the warming event is marked by (1) the temporary disappearance of the high-fertility species Biscutum constans, (2) a demise of cool-water indicators and (3) an acme of tropical species Micula murus (Thibault and Gardin, 2006, 2007). Whether this warm event is linked or not to the onset of Deccan volcanic activity remains controversial. Though several authors sug- gested that flood basalt eruptions were likely to cause greenhouse warming via CO 2 degassing (Wignall, 2001; Cohen and Coe, 2002), results of palaeotemperature models based on the estimation of gas emissions Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252 ⁎ Corresponding author. E-mail address: [email protected] (N. Thibault). 1 Absolute ages of Maastrichtian climate trends described in this paper refer to the age of the K/Pg boundary given in Gradstein and Ogg (2004). All ages given in the literature prior to this publication may therefore differ by at least 0.5 Ma. 0031-0182/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2010.02.036 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
The calcareous nannofossil response to the end-Cretaceous warm eventin the Tropical Pacific
Nicolas Thibault a,⁎, Silvia Gardin b
a Department of Geography and Geology, University of Copenhagen, Øster Voldgade 10, 1350 Copenhagen C., Denmarkb CR2P «Centre de Recherche sur la Paléobiodiversité et les Paléoenvironnements», UMR CNRS 7207, Université Pierre etMarie Curie Paris 6, case 104, 4, place Jussieu, 75252 Paris cedex 05, France
⁎ Corresponding author.E-mail address: [email protected] (N. Thibault).
1 Absolute ages of Maastrichtian climate trends descrage of the K/Pg boundary given in Gradstein and Oggliterature prior to this publication may therefore differ
0031-0182/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.palaeo.2010.02.036
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 September 2009Received in revised form 3 February 2010Accepted 26 February 2010Available online 6 March 2010
Keywords:Late MaastrichtianCalcareous nannofossilsDeccanShatsky RiseTropical Pacific
The response of calcareous nannofossils to the end-Cretaceouswarming is investigated in the Tropical Pacific DSDPSite 577A (Shatsky Rise) through the record of species richness, Shannon diversity, distribution patterns andstatistical treatments. This event is marked by a strong acme of the tropical species Micula murus and isaccompanied by a decrease of Placozygus spiralis, the disappearance of Biscutum constans and a decrease in bulkδ13C, indicative of a decrease in primary productivity. An increase in abundance of otherMicula species at Site 577Aand a drop in species richness recorded in all sites of Shatsky Rise, suggest stressed environmental conditionscoincident with the end-Cretaceous warming. The acme of M. murus at Shatsky Rise and in the Atlantic Ocean isrecorded within magnetochron C29r and correlates (1) with the intense warming as expressed in intermediatewaters through the δ18O of benthic foraminifera, and in surface waters through poleward migration, reduceddiversity and dwarfism in planktonic foraminifer assemblages, (2) with a sharp decline in marine 187Os/188Os,chemicalmarker of Deccan volcanic activity, and (3)with a rise in the atmospheric pCO2 record of terrestrial plantsin Texas, USA, probably triggered by Deccan volcanic degassing. However, a drop of calcareous nannofossil cool-water taxa in the Equatorial and South Atlantic, aswell as a first rise in abundance ofM.murus in the South Atlanticand Tropical Pacific occur in the topmost part of chron C30n, and correlates with the rise in palaeotemperaturerecord of terrestrial plants inNorthDakota, USA. This suggests that initiationof thewarming in the atmosphere andsurface waters may predate the striking warming of intermediate water masses by 150/200 kyr. The coincidenceand the links between climate change, volcanism, geochemical and biotic events at the end of the Maastrichtianthus remain to be fully elucidated.
ibed in this paper refer to the(2004). All ages given in theby at least 0.5 Ma.
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The well-documented long-term cooling trend that started in thelate Campanian and continued into the early Palaeogene (Douglas andSavin, 1975; Savin, 1977; Boersma and Schackleton, 1981) wasinterrupted by a short warming event between ∼65.6 and 66Ma.1
Cooling resumed in the last 100 kyr of the Maastrichtian (Li andKeller, 1998a). During the warm event, temperature of intermediatewaters (IWs) increased globally by 2–4 °C, as inferred from the recordof benthic foraminiferal oxygen stable isotopes of numerous deep-seasites in the North Atlantic, South Atlantic, Tropical Pacific and IndianOcean (Barrera and Savin, 1999 and references therein). A warmpulseof 3–4 °C is well-recorded in surface waters in southern high-latitudeSites 689 and 690 (Stott and Kennett, 1990; Barrera, 1994) but ispoorly resolved through planktonic foraminiferal stable isotopes in
tropical and temperate latitudes (Li and Keller, 1998a; Barrera andSavin, 1999). The response of the planktonic community to this eventsuggests that surface waters were also strongly affected by atemperature rise. In South Atlantic Site 525A, the warming pulse isaccompanied by reduced species richness and Shannon diversity,dwarfing and reduced photosymbiotic activity of planktonic forami-nifera (Abramovich and Keller, 2003). Poleward migration of low-latitude planktonic foraminifer species and an increase in abundanceof Contusotruncana contusa and Pseudotextularia elegans are associat-ed with this end-Maastrichtian warming (Huber and Watkins, 1992;Kucera and Malmgren, 1998; Olsson et al., 2001). In calcareousnannofossil assemblages of Sites 1258A (western equatorial Atlantic)and 525A, the warming event is marked by (1) the temporarydisappearance of the high-fertility species Biscutum constans, (2) ademise of cool-water indicators and (3) an acme of tropical speciesMicula murus (Thibault and Gardin, 2006, 2007).
Whether this warm event is linked or not to the onset of Deccanvolcanic activity remains controversial. Though several authors sug-gested that flood basalt eruptions were likely to cause greenhousewarmingviaCO2degassing (Wignall, 2001;CohenandCoe, 2002), resultsof palaeotemperature models based on the estimation of gas emissions

240 N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
suggest a small impact on temperature rise or even cooling through therelease of SO2 (Chenet et al., 2005; Jones et al., 2005; Self et al., 2006).
Here we present new data on calcareous nannofossil assemblagesat DSDP Site 577A in an attempt to decipher their response to the end-Cretaceous warming in the Tropical Pacific. These data are comparedto previous results obtained on the nannofossil assemblages of Sites1258A and 525A. The correlation of calcareous nannofossil distribu-tion patterns with chemical markers of Deccan volcanism andpreviously published markers of climate change is tested.
2. Shatsky Rise: setting and previous work
Shatsky Rise (Fig. 1) is a medium-sized large igneous province(LIP), in the west-central Pacific which has been the target of threeDeep Sea Drilling Project (DSDP) expeditions—Legs 6, 32, and 86—andtwo Ocean Drilling Program (ODP) expeditions—Legs 132 and 198(Bralower et al., 2002a). A high quality palaeoceanographic recordwasobtained during Leg 86 at Site 577A, and limited to the Paleogene anduppermost Maastrichtian (Gerstel et al., 1986; Zachos and Arthur,1986; Zachos et al., 1989; D'Hondt and Keller, 1991). This site has agood magnetostratigraphic record (Bleil, 1985) that allows precisedating and solid correlations of palaeoclimatic eventswith other areas.
Fig. 1.Map of Shatsky Rise showing bathymetry of the rise, location of DSDP Site 577A and oLarge Igneous Provinces in the Pacific Ocean. mbsl: meters below seafloor.After Bralower et al. (2002a, modified).
More recent drillings of Shatsky Rise, such as ODP Leg 198 (Sites1207 to 1214) gave rise to numerous palaeoceanographic investiga-tions regarding the early Cretaceous (Dumitrescu et al., 2006), the K–Pg boundary (Bown, 2005), the Paleocene–Eocene (Zachos et al., 2003;Gibbs et al., 2006a, 2006b; Kaiho et al., 2006), or synthetic overviewsover longer time-intervals (Bralower et al., 2002b; Thomas, 2004;Bralower et al., 2006). A palaeoceanographic investigation of Franket al. (2005) was mainly focused on the early and mid-Maastrichtian.
However, these sites lack of a good magnetostratigraphic controlthat DSDP Site 577A has, thus providing precise dating and soundcorrelations of palaeoclimatic events with other areas.
The majority of Shatsky Rise sites were situated at relativelyshallow palaeodepths, mostly b2000 m, thus above the calcitecompensation depth throughout the Cretaceous. Palaeoreconstruc-tions have a great deal of uncertainty but all sites should lay between10°N and 20°N during the Late Cretaceous (Bralower et al., 2002a).
3. Materials and methods
DSDP Site 577A has been intensively studied for palaeoceanographicpurposes and material is limited. Nineteen samples of the upperMaastrichtian of Site 577A were analyzed for calcareous nannofossil
ther DSDP/ODP drill sites and (inset) the location of Shatsky Rise relative to other major

Table 1Depths of magnetic reversals and calculated sedimentation rates at Sites 1258A, 577A and 525A.
Sites Magnetochrons Top depth(mbsf)
Bottom depth(mbsf)
Mean length(cm)
Duration(ka)a
Sedimentation rate(cm/ka)
1258A Cretaceous C29r 252.55±0.05b 262.17±0.6b 962 361 2.66C30n–C31n 262.17±0.6b 294.65±2.3b 3248 2871 1.13
577A Cretaceous C29r 109.62c 113.015±0.075c 339.5 361 0.94C30n 113.015±0.075c 121.18±0.09c 816.5 1835 0.44
525A Cretaceous C29r 451.71d 456.39±0.3d 468 361 1.3C30n 456.39±0.3d 475.45±0.3d 1906 1835 1.04
a Ogg and Smith (2004).b Suganuma and Ogg (2006).c Bleil (1985).d Chave (1984).
Table 3Main nannofossil bio-events recorded and inferred biozonations.
Core, section,interval(cm)
Depth(mbsf)
Nannofossilevents
Nannofossilzones afterBurnett (1998)
Nannofossil zonesafter Sissingh (1977)and Perch-Nielsen(1985)
12-4_69-70 109.59 UC20dTP CC26b12-4_78-79 109.68
241N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
assemblages. Samples were processed following the method given inThibault and Gardin (2006). Relative species abundances werecalculated as the percentage of at least 500 specimens randomlycounted at a magnification of ×1575 (×63 oil objective with a ×2.5additional lens). Absolute abundanceswere calculated for one species asthe ratio between the number of the counted specimens and thenumber of fields of view examined to count them. After measuring thearea of one field of view (0.0172 mm2), abundances were transformedinto number of specimens permm2. As the grain density is similar for allsmear-slides (1 g/L), abundances per unit area are approximatelyproportional to the quantity of a given species per gram of sediment.This method is comparable to that used in the works of Backman andShackleton (1983), Henriksson (1993) and Koch and Young (2007). Forcomparisons of nannofossil data between Sites 577A, 1258A and525A, absolute abundances were transformed into accumulation rates(N/cm2/ka) by taking into account the density of thematerial (2.1 g/cm3
for chalk) and estimated mean sedimentation rates for each magneto-chron (Table 1). The range chart for nannofossil data of Site 577A isarchived in Appendix A. Species from the same genus and with similarbehaviours, as well as species that were too rare for an accuratequantitative analysis were grouped into higher taxonomic levels(Table 2). Three traverses of the slide were examined for speciesrichness. Species not considered during the counting for relativeabundances were noted as rare (R), few (F) and Common (C) in therange chart (Appendix A). For the calculation of Shannon diversityindex, these annotations were replaced by one and the total number ofspecimens encountered subsequently corrected. Three states of preser-vation (bad, moderate and good) were considered in this study, on thebasis of features of etching and overgrowth given by Roth (1983). Allsamples show a moderate preservation.
The range charts published by Lees and Bown (2005) were used togenerate records of nannofossil species richness at Sites 1209A,1209C, 1210A, 1210B and 1211A. Other sites of Leg 198 were notconsidered because of stratigraphical incompleteness in the lateMaastrichtian.
Carbon isotopic composition of bulk carbonates were measured on17 samples at the Laboratoire Biominéralisations et Paléoenvironne-ments (Université Pierre et Marie Curie, Paris 6, France). The
Table 2Composition of the main groups of taxa quoted in the text.
Group Species
Ceratolithoides spp. C. indiensis, C. kamptneri, C. ultimusCylindralithus spp. C. nudus (dominant), C. scuptus, C. serratusEiffellithus spp. E. gorkae, E. parallelus, E. turriseiffeliiMicula spp. M. decussata (dominant), M. concava, M. cubiformis,
M. swasticaPrediscosphaeracretacea s.l.
P. cretacea (dominant), P. grandis, P. majungae
Retecapsa spp. Retecapsa angustiforata (dominant), R. crenulata, R. ficula,R. schizobrachiata, R. surirella
Rhagodiscus spp. R. indistinctus, R. plebeius, R. reniformis, R. spendens
extraction of CO2 was done by reaction with anhydrous orthopho-sphoric acid at 50 °C. Analysis was performed with a mass spectrom-eter Finnigan Delta E. The oxygen and carbon isotope values areexpressed in per mil relative to the V-PDB standard reference. Theanalytical precision is estimated at 0.1‰ for oxygen and 0.05‰ forcarbon.
4. Results
4.1. Biostratigraphy
The calcareous nannofossil biostratigraphy formerly establishedby Monechi (1985) was refined, based on the First Occurrences (FO)ofMicula murus, Ceratolithoides kamptneri andMicula prinsii (Table 3).Biozonation of Burnett (1998) was applied (Table 3). In addition, 4secondary bio-events are documented: the Last Occurrence (LO) ofTegumentum stradneri, the FO of Ceratolithoides ultimus, the LO ofPseudomicula quadrata and the Last Consistent Occurrence (LCO) ofBiscutum constans (Table 3). The LO of T. stradneri and the FO of C.ultimus appear in the same nannofossil zones in Hole 577A as in Holes1209A, 1210A, 1210B and 1211A (Lees and Bown, 2005). The LCO of B.constans recorded in the top of chron C30n correlates well with its LOat sites 1258A and 525A (Thibault and Gardin, 2006, 2007). Mixing ofUpper Cretaceous and early Danian assemblages of planktonicforaminifera was noted for the last meter of the Maastrichtian atSite 577A (D'Hondt and Keller, 1991). No Danian nannofossil speciesnor apparent mixing was found in the 3 samples examined in thisinterval.
12-4_82-83 109.7212-4_126-127 110.1612-5_6-7 110.46 LO P. quadrata12-5_86-87 111.26 FO C. ultimus12-5_146-147 111.8612-6_26-27 112.1612-6_86-87 112.76 FO M. prinsii12-6_146-147 113.36 UC20cTP CC26a12-7_14-15 113.54 LCO B. constans13-1_44-45 114.3413-2_106-107 116.4613-3_44-45 117.34 FO C. kamptneri13-4_106-107 119.46 LO T. stradneri UC20bTP CC25c13-5_44-45 120.3413-6_106-107 122.4613-7_4-5 122.94 FO M. murus13CC_18-19 123.38 UC20aTP CC25b
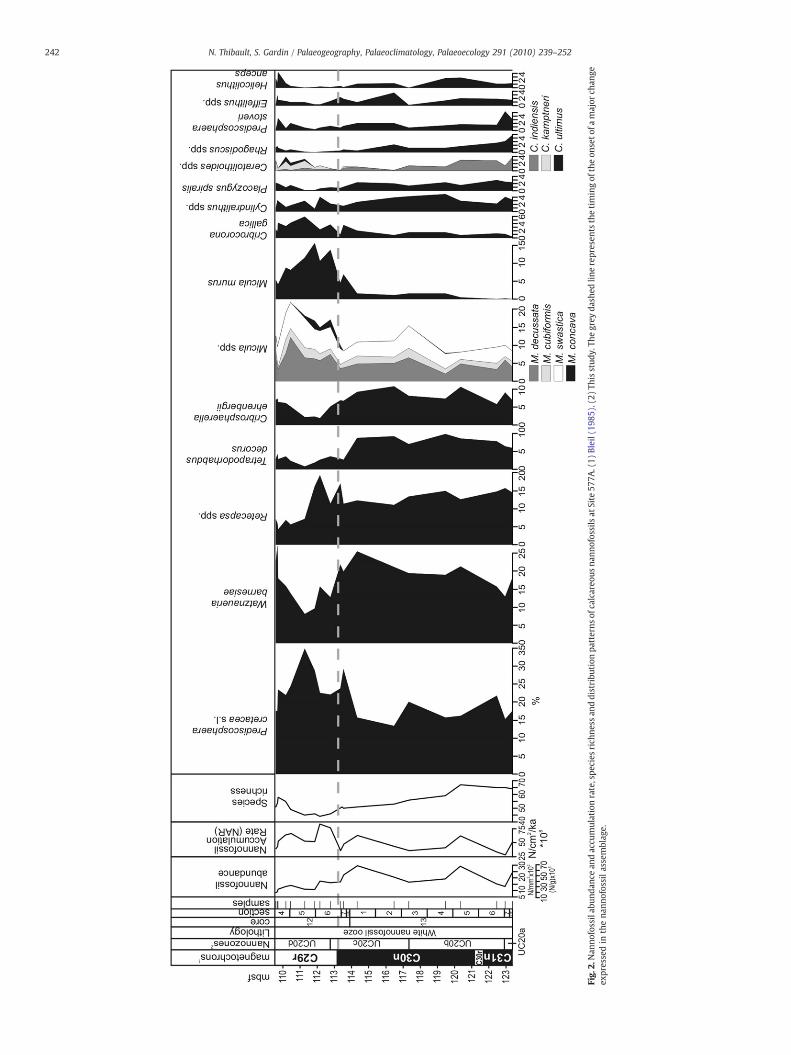
Fig.
2.Nan
nofossilab
unda
ncean
daccu
mulationrate,spe
cies
rich
ness
anddistribu
tion
patterns
ofcalcareo
usna
nnofossilsat
Site
577A
.(1)
Bleil(19
85).(2
)Th
isstud
y.Th
egrey
dash
edlin
erepresen
tsthetimingof
theon
seto
famajor
chan
geex
pressedin
thena
nnofossila
ssem
blag
e.
242 N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252

243N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
4.2. Calcareous nannofossil assemblages at Site 577A
4.2.1. Absolute abundance, accumulation rate and species richnessThe total nannofossil abundance varies between 2000 and
3000 specimens/mm2 between 123.5 and 113.5 mbsf (meters belowseafloor). A first drop to around 1600 specimens/mm2 is observed at113.5 mbsf and a second drop down to 1000 specimens/mm2 occursaround 112 mbsf. It then remains relatively low up to the K–Pgboundary (Fig. 2). This drop in the absolute abundance is probablydue to an increase of the sedimentation rate in chron C29r by theaddition of other calcitic contributors. Indeed, it appears that thenannofossil accumulation rate (NAR) did not actually decrease at theend of the Cretaceous (Fig. 2). Species richness is about 65 between123.5 and 120.5 mbsf. It progressively decreases down to 50 at113.5 mbsf. It drops to 45 in the interval between 113.5 and110.5 mbsf and then increases again to reach a mean value of 55 inthe last meter of the Maastrichtian (Fig. 2). Shannon diversity, a morerobust index of diversity whose calculation takes into account therelative abundance of each species, exhibits more or less the samefeatures (Fig. 3). Moreover, the LO of Tegumentum stradneri (1), the FOof Ceratolithoides kamptneri (2) and the FO ofMicula prinsii (3) allow acorrelation of the patterns of species richness at Site 577A with that ofSites 1209A, 1209C, 1210A, 1210B and 1211A (Fig. 4), calculated fromthe range charts of Lees and Bown (2005). Species richness displaysthe same patterns in all these sites, regardless of the preservationstate of the assemblage (Lees and Bown, 2005, Tables 4, 5, 6, 7 and 8).
4.2.2. Composition of the assemblageExamination of the calcareous nannofossil data indicates that the
assemblages of Site 577A are dominated by four main taxa (Predisco-sphaera cretacea s.l.,Watznaueria barnesiae, Retecapsa spp. andMiculaspp.), each of which has an average abundance of more than 10%. Afifth additional taxon, Micula murus, becomes dominant in the latestMaastrichtian, with abundance reaching more than 10% in chron C29r(Fig. 2). Two other dominant taxa are characterized by averageabundances higher than 5% and sometimes reaching up to 10%(Tetrapodorhabdus decorus and Cribrosphaerella ehrenbergii) and
Fig. 3.Nannofossil accumulation rate, species richness, Shannon–Weaver index, PCA1 scores,(1) Bleil (1985). (2) This study.
8 secondary taxa are characterized by having average values ofmore than 2% (Fig. 2). In total, these taxa represent at least 90% of thenannofossil assemblages.
The seven dominant taxa exhibit a major change of abundance ataround 113.5 mbsf and a return to the values that prevail before thismajor event from around 110.5 mbsf to the K–Pg boundary (Fig. 2).High-latitude taxa (Ahmuellerella octoradiata, Gartnerago spp., Kampt-nerius magnificus and Nephrolithus frequens, Thierstein, 1981; Wind,1979; Pospichal and Wise, 1990; Lees, 2002) are absent at Site 577Aduring the late Maastrichtian. Biscutum constans, a high-fertilityspecies (Erba, 1990; Erba et al., 1992; Williams and Bralower, 1995;Fisher and Hay, 1999) is very rare at this site. It shows patchyoccurrences from 123.5 to 113.5 mbsf and is then completely absentin the interval from 113.5 mbsf up to the K–Pg boundary, except forone sample at 109.7 mbsf (Appendix A).
Prediscosphaera cretacea remains stable around 17% between 123.5and 114 mbsf, exhibits higher abundances (23 to more than 30%) inthe interval between 113.5 and 110.5 mbsf and tends to return toprevious values in the last meter of the Maastrichtian (Fig. 2).
Watznaueria barnesiae tends to increase from 15 to 25% between123.5 and 114 mbsf. It progressively decreases down to 8% between114 and 111.3 mbsf and increases again from 111.3 mbsf up to the K–Pg boundary (Fig. 2). The drop in relative abundance of W. barnesiaebetween 114 and 111 mbsf cannot be attributed to any close-sumeffect since its accumulation rate shows the same trends (Fig. 3).
The abundance of Micula decussata is quite low at Site 577A ascompared to previous studies in the Maastrichtian of the tropicalrealm (Thierstein, 1981; Eshet and Almogi-Labin, 1996; Tantawy,2003; Thibault and Gardin, 2006, 2007). Micula swastica, anotherspecies of the genus with a morphology close to that of M. decussata,has a noticeably high abundance at Shatsky Rise (Fig. 2). Both speciesare around 5% between 123.5 and 113 mbsf as well as in the last meterof the Maastrichtian and show a 2 to 7% increase in the intervalbetween 113.5 and 110.5 mbsf (Fig. 2).
Retecapsa spp. remains around 12% between 123.5 and 113.5 mbsf.It shows a 5% excursion between 113.5 and 111.5 mbsf, drops down to8% at 111.5 mbsf and slightly decreases up to the K–Pg boundary.
accumulation rates of selected individual nannofossil species and bulk δ13C at Site 577A.
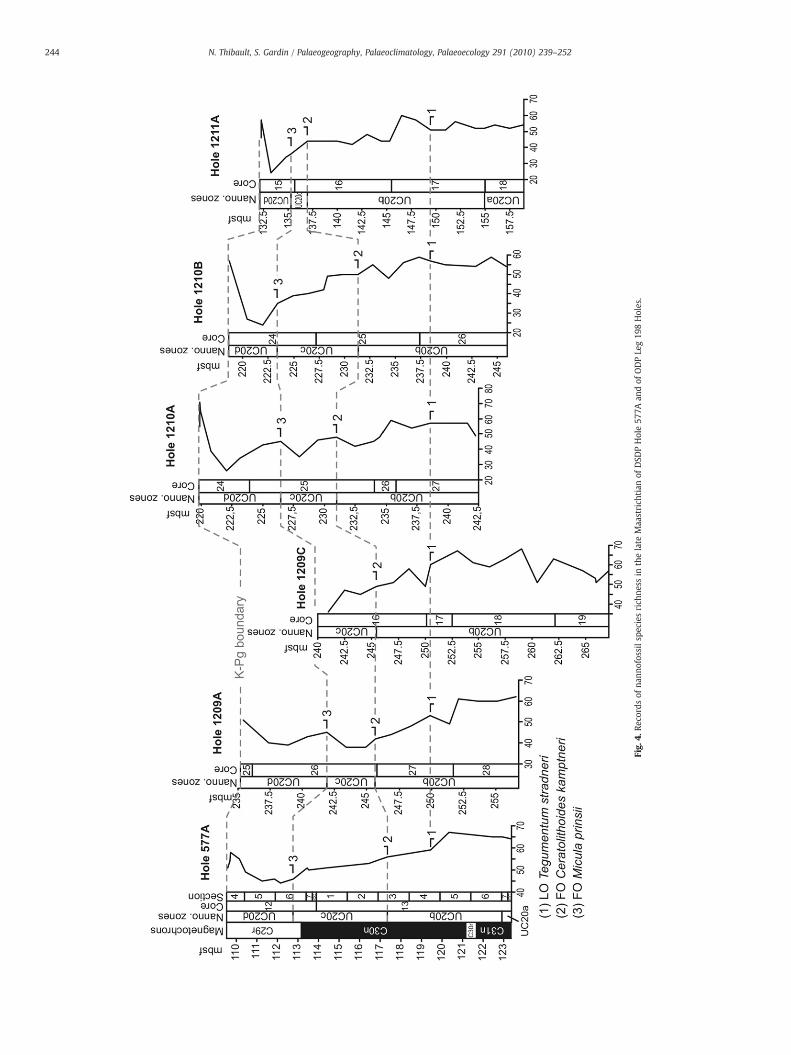
Fig.
4.Re
cordsof
nann
ofossils
pecies
rich
ness
inthelate
Maa
strich
tian
ofDSD
PHole57
7Aan
dof
ODPLe
g19
8Holes.
244 N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252

Table 4Significant species, Factor loadings and their correlations to PCA1.
Taxa Component loadings Correlation to PCA1
Ceratolithoides spp. 0.02 0.16C. gallica 0.14 0.86C. ehrenbergii −0.24 −0.87Cylindralithus spp. −0.06 −0.48Eiffelithus spp. −0.05 −0.50H. anceps −0.06 −0.48Micula spp. 0.36 0.78M. murus 0.48 0.90P. spiralis −0.07 −0.79P. cretacea s.l. 0.53 0.89P. stoveri −0.06 −0.44Rhagodiscus spp. −0.08 −0.61Retecapsa spp. −0.08 −0.17T. decorus −0.25 −0.82W. barnesiae −0.42 −0.77
245N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
Cribrosphaerella ehrenbergii is around 7.5% between 123.5 and113.5 mbsf, then drops down to values of 2% and shows a 7% increasein the last 1.5 m of the Maastrichtian (Fig. 2).
Tetrapodorhabdus decorus remains stable around 7.5% between123.5 and 114 mbsf, then suddenly drops down to 3% and remains lowuntil the K–Pg boundary (Fig. 2).
After its first appearance, Micula murus is less than 2% abundantbelow 113.5 mbsf where it starts to increase to reach abundances upto more than 15% around 112 mbsf. Then it decreases but remainshigher than 8% until 110.5 mbsf. It drops down to slightly less than 5%in the last meter of the Maastrichtian (Fig. 2).
Cribrocorona gallica is less than 2% abundant between 123.5 and114 mbsf. Then it increases and fluctuates between 3 and 6% between113.5 mbsf and the K–Pg boundary (Fig. 2).
Placozygus spiralis (synonyms: Zeugrhabdotus spiralis and Placozy-gus fibuliformis of some authors) shows very low abundances as com-pared to previous studies in the Maastrichtian of the Tropical Realm(Thierstein, 1981; Eshet and Almogi-Labin, 1996; Tantawy, 2003;Thibault andGardin, 2006, 2007). Its abundance remains stable around2.5% between 123.5 and 114 mbsf. It suddenly decreases at 113.5 mbsfand remains less than1% abundantuntil 110.5 mbsfwhere it returns toprevious values which persist up to the K–Pg boundary.
4.2.3. Preservation of the assemblageThe preservation of the samples considered in this study is mode-
rate. No change of the preservation state is observed in samples fromthe interval between 113.5 and 110.5 mbsf where the major change isobserved in the nannofossil assemblage. In theMaastrichtian, the mostresistant taxon to dissolution is Micula decussata with a ratio ofresistance to dissolution which is more than 10 times that of othernannofossil taxa (Thierstein, 1980). Therefore an independent test ofthe preservation of the assemblage is the comparison of its relativeabundance with species richness and total nannofossil absoluteabundance. At Site 577A, M. decussata, Micula concava and Miculaswasticawere grouped together asMicula spp. because of similar sizesand morphologies which make these species equally resistant todissolution.Miculamuruswasnot includedbecause this species ismuchsmaller and has a much lower ratio of resistance to dissolution(Thierstein, 1980). The abundance of Micula spp. does not show anysignificant correlationwith nannofossil absolute abundance (r2=0.21)and species richness (r2=0.4). The abundance of Watznaueriabarnesiae, also considered as a solution-resistant species, though witha lower ratio of resistance to dissolution thanM. decussata (Thierstein,1980), does not decrease and shows on the contrary a strong decreasein the intervalwhereMicula peaks (Fig. 2).We conclude that diageneticprocesses cannot be involved to explain the strong increase ofM.murusand other changes observed in the nannofossil assemblage. Patterns ofabundances as well as diversity indices can be interpreted as areflection of original palaeoecological conditions.
4.2.4. Principal component analysis (PCA)Principal component analysis (PCA) was performed on the
variance–covariance matrix including all taxa considered in Fig. 2and accounting in total for at least 90% of the assemblage. Miculacubiformis,Micula concava,Micula decussata andMicula swasticaweregrouped together as Micula spp. because they generally showed thesame behaviour at site 577A. The first principal component (PCA1)accounts for 59.3% of the total variance. Species that show positiveloadings and significant positive correlations to PCA1 are the threedominant taxaMicula murus, Prediscosphaera cretacea, andMicula spp.and the secondary species Cribrocorona gallica (Table 4). Species thatshow negative loadings and significant negative correlations to PCA1are the dominant taxa Watznaueria barnesiae, Cribrosphaerellaehrenbergii and Tetrapodorhabdus decorus, and the secondary speciesPlacozygus spiralis (Table 4). Scores of PCA1 reveal a strong positiveexcursion in the interval between 113.5 and 110.5 mbsf (Fig. 3).
4.3. Stable carbon isotopes from Site 577A
Results of stable isotopes from bulk chalks are commonly hinderedby diagenesis overprinting because of the high porosity of the chalk(Schrag et al., 1995; Mitchell et al., 1997). A correlation test of carbon-and oxygen-isotope values shows no significant trends and lacks thepronounced covariance (r2=0.22) seen in many mixing linesproduced by the addition of variable quantities of isotopicallyhomogeneous diagenetic cement to isotopically homogeneous skel-etal calcite (Jenkyns et al., 1995; Mitchell et al., 1997). The pattern ofδ13C values generally conforms to trends observed in planktonic andbenthic foraminifera at Sites 525A (South Atlantic Ocean, Fig. 3) (Liand Keller, 1998a) and 761 (Indian Ocean) (Barrera and Savin, 1999).Given that chalk sediments of Site 577A are mainly composed ofcalcareous nannofossils, the characteristic form of this curve likelyreflects primary seawater values with any diagenetic effects affectingthe section in generally consistent manner.
Carbon isotope values range between 2.8 and 3.2‰. The δ13Cprofile displays quite stable values around a mean of 3‰ between123.5 and 113.5 mbsf and a slight negative excursion down to valuesof 2.8–2.9‰ in the interval between 113.5 and 110.5 mbsf. An increaseup to a value of 3.15‰ is recorded in the last sample of theMaastrichtian (Fig. 3).
5. Discussion
5.1. Ecological affinities of selected Upper Cretaceous nannofossil taxa
Prediscosphaera cretacea does not show any clear latitudinalpreferences (Wind, 1979; Thierstein, 1981; Henriksson and Mal-mgren, 1997; Lees, 2002). Several authors suggested an affinity of thistaxon to low-productivity conditions (Watkins, 1989; Eshet et al.,1992; Eshet and Almogi-Labin, 1996).
Watznaueria barnesiae is ubiquitous in Cretaceous assemblages butabsent or rare in northern and southern high latitudes (Bukry, 1973;Wind, 1979; Thierstein, 1981; Watkins et al., 1996) during someintervals of the Maastrichtian where its varying abundances andpatterns of migration were linked to temperature changes (Stott andKennett, 1990; Huber and Watkins, 1992). Several studies showedthat this is also a low-nutrient indicator (Roth and Krumbach, 1986;Erba et al., 1992; Lamolda et al., 1992; Williams and Bralower, 1995;Fisher and Hay, 1999).
Micula decussata (the dominant species of the genus) is a majorcomponent of Late Cretaceous assemblages. Its biological affinity andits palaeoecological significance are not yet well-understood. Sincethe studies of Hill (1975) and Thierstein (1980), its distribution haslargely been interpreted as the result of preservation patterns.However, several authors reported very high abundances of this

246 N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
species in well-preserved uppermost Maastrichtian samples showingno evidence of dissolution or overgrowth (Eshet et al., 1992; Tantawy,2003). Different palaeoecological significances of this species areproposed in the literature: (1) affinity for sub-tropical to intermediatelatitudes (Wind, 1979; Thierstein, 1981; Henriksson and Malmgren,1997), (2) affinity for low latitudes (Shafik, 1990), (3) cosmopolitan(Lees, 2002), (4) low-nutrient indicator and high-stress marker(Eshet et al., 1992; Tantawy, 2003; Thibault and Gardin, 2006).
Cribrosphaerella ehrenbergii does not show any latitudinal prefer-ences (Thierstein, 1981; Henriksson andMalmgren, 1997; Lees, 2002)nor was related to surface water fertility changes. The significance ofits distribution patterns remains unknown.
Retecapsa ficula increases concomitantly with high-fertility speciesBiscutum spp. and Zeugrhabdotus erectus during Oceanic anoxic event2 at Demerara Rise and was thus interpreted as another high-fertilityspecies (Hardas and Mutterlose, 2007). In the Cenomanian/Turonianboundary interval of Wunstorf section, Retecapsa spp. does not reactlike the high-fertility taxon Biscutum spp. (Linnert et al., 2010). Highfrequencies of Retecapsa spp. are linked to palaeoceanographicconditions which are in favour of black shale deposition,corresponding to short-term pulses of intense fertilisation. Thegenus is thus interpreted as an opportunistic stress resistant formwhose increased flourishment is linked to highly elevated primaryproduction (Linnert et al., 2010).
Tetrapodorhabdus decorus was more abundant in southern highlatitudes than low latitudes in the Maastrichtian (Lees, 2002).
Micula murus is one of the rare Maastrichtian species that is clearlyrestricted to low latitudes (Worsley and Martini, 1970; Thierstein,1981; Lees, 2002). It has an unequivocal preference for warm surfacewaters but is generally less than 5% in tropical assemblages(Thierstein, 1981; Tantawy, 2003; Thibault and Gardin, 2006, 2007).
Cribrocorona gallica was noted a preference for low latitudes(Shafik, 1990). Lees (2002) noticed its absence in the subantarcticpalaeobiogeographical zone.
Placozygus spiralis does not show any clear biogeographicpreferences (Thierstein, 1981; Lees, 2002) but is considered as agood indicator of mid- and high-fertility conditions (Roth andBowdler, 1981; Watkins, 1989; Erba, 1992; Lees et al., 2005).
5.2. The nannofossil record of the end-Maastrichtian warming at ShatskyRise
PCA scores and diversity indices point to a striking event in thelatest Maastrichtian at Site 577A (Fig. 3). This event cannot be theresult of an artifact due to a possible mixing of Cretaceous and earlyDanian assemblages because the interval where mixing of planktonicforaminifera was observed corresponds to the last meter of theMaastrichtian whereas the most important changes observed in thenannofossil assemblage occur 4 m before the K–Pg boundary (Figs. 2and 3). The highest component loadings for PCA1, Prediscosphaeracretacea and Micula murus, show respective correlations of 0.89 and0.90 with the component factor (Table 4). They thus indicate low-fertility conditions and warming during the positive excursion of thisfactor (Fig. 3). Warming is further supported by a correlation of 0.85between PCA1 and Cribrocorona gallica, another warm-water species,and by a negative correlation (−0.82) between PCA1 and Tetra-podorhabdus decorus, which showed a preference for high southernlatitudes in the Maastrichtian. Low-fertility conditions during thewarming event are further supported by (1) a negative correlation(−0.79) between the mid-fertility species Placozygus spiralis andPCA1, (2) the absence of the high-fertility species Biscutum constans,and (3) a decrease of bulk δ13C (Fig. 3). The progressive decrease ofspecies richness before the warming event at Site 577A, followed by adrop during the event itself, are reliable patterns that can becorrelated throughout ODP sites at Shatsky Rise (Fig. 4). This decrease,along with the drop of total nannofossil abundance, suggests low-
fertility and stressed environmental conditions as a response to thewarm event. The increase of Micula spp. (excluding M. murus) duringthe event interval as well as its positive correlation to PCA1 (0.78,Table 4) also support this interpretation, as already suggested byTantawy (2003) and Thibault and Gardin (2006).
The warm event terminated about 100 kyr before the K–Pg andwas followed by a short cooling episode (Li and Keller, 1998a; Wilfet al., 2003), which is indicated at Site 577A by the decrease in theabundance of Micula murus, as well as by the decrease of PCA1. Thereturn of all parameters (relative abundances, PCA1, diversity indices,and bulk δ13C) to values that prevailed before the warming event(Fig. 3) suggests a recovery of the nannofossil assemblage during thecooling event.
5.3. Comparison of calcareous nannofossil and oxygen-isotope records inthe Tropical Atlantic and Pacific Oceans
An acme of Micula murus, coincident with the end-Maastrichtianwarming, was previously recorded in the Equatorial Atlantic (ODP Site1258A) and in the South Atlantic (DSDP Site 525A) (Thibault andGardin, 2006, 2007). Such high relative abundances of M. murus,however, were never recorded before (N15% at Site 577A during theacme event, as compared to 3 and 4% only at Sites 1258A and 525A,respectively). Thierstein (1981) also found anomalously high abun-dances of this taxon in the latest Maastrichtian of Tropical DSDP Sites317 and 47.2. No quantitative data are available from ODP Leg 198 butM. murus is frequently recorded as common in the latest Maastrich-tian of several sites of this Leg (Lees and Bown, 2005). Thispronounced acme of M. murus thus seems to be a peculiar feature ofthe Tropical Pacific Ocean that could be explained by the conjunctionof warming and ultra-oligotrophic environmental conditions. In theLate Cretaceous, these sites were indeed located very far away fromthe continental coastlines within the large oligotrophic gyres of thePacific Ocean. In this region, the decrease of productivity associated tothe warming event would have resulted in ultra-oligotrophicconditions, or at least in more oligotrophic conditions than in theTropical Atlantic, which would explain the differences observed in therelative abundance of M. murus between the two oceans.
The decrease of Watznaueria barnesiae observed at Site 577Aduring the end-Cretaceous warming is difficult to reconcile with thesupposed ecological affinities of this species. During the lateMaastrichtian, this species reacted positively to warming only insouthern and northern high latitudes (Huber and Watkins, 1992;Sheldon, 2008). In the Equatorial Atlantic, no significant change isnoticed in the abundance of W. barnesiae during the end-Cretaceouswarming (Thibault and Gardin, 2006) whereas in the South Atlantic, aslight increase of its abundance, concomitant to the acme of Miculamurus, was interpreted as lowered fertility (Thibault and Gardin,2007). Ultra-oligotrophic and stressed environmental conditionsassociated to the warming event seem to have favoured the lower-fertility species Prediscosphaera cretacea over W. barnesiae at ShatskyRise.
To date, at Tropical latitudes, the end-Maastrichtian warming iswell-recorded in intermediate waters through the δ18O of benthicforaminifera whereas the record of surface waters based onplanktonic foraminiferal δ18O exhibits a very slight increase withhighly variable fluctuations (Abramovich and Keller, 2003). This canbe explained by the fact that most planktonic foraminifer species ofthe Campanian–Maastrichtian occupied a subsurface mixed layerrather than surface waters (Abramovich et al., 2003). This is notablythe case for Rugoglobigerina rugosa, from which were inferred sea-surface temperatures at DSDP Site 525A (Abramovich and Keller,2003). In contrast, most nannofossil species of the Maastrichtian wererestricted to surface waters. For the Maastrichtian, bulk-rock δ18Osignals of pure nannofossil chalks and nannofossil fractions that didnot undergo intense diagenetic alteration might actually better reflect

247N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
sea-surface temperatures than the existing record of planktonicforaminifera. Such analyses are lacking for the latest Maastrichtianand are greatly needed to better constrain the impact of the end-Maastrichtian warming on global SSTs.
The acme ofMicula murus in the Tropical Pacific and in the Atlanticcorrelates well with the end-Maastrichtian warming as expressed inthe δ18O of benthic foraminifera and with the dwarfism observed inplanktonic foraminifera within chron C29r (Fig. 5). However, thecomparison of the records of calcareous nannofossil assemblagesbetween the Tropical Pacific and the Atlantic Ocean suggests thatwarming of the surface waters might have initiated at the top of chronC30n. This view is supported by a drop of cool-water taxa in theuppermost part of chron C30n in the Equatorial and South Atlantic(Fig. 6) and by the first increase of M. murus at the top of C30n at Site577A (Figs. 2 and 6) and in the last sample of chron C30n at Site 525A(Fig. 6). The disappearance of Biscutum constans occurs right after thedrop of cool-water taxa at the top of chron C30n and can be correlatedthroughout Sites 1258A, 577A and 525A. This event supports adecrease of global productivity coincident with the onset of warmingin surface waters.
5.4. End-Maastrichtian warming and climatic impact of Deccanvolcanism
Several authors have stated that Deccan volcanism induced theend-Maastrichtian warming through the addition of greenhouse gasCO2 to the atmosphere (Dessert et al., 2001;Wignall, 2001; Cohen andCoe, 2002). Caldeira and Rampino (1990), Rampino and Self (2000)and Self et al. (2006, 2008) have challenged this hypothesis,suggesting that CO2 emission triggered by Deccan volcanism wouldhave caused a rise of less than 2 °C in global mean temperatures, incontrast to the 4 °C warming in intermediate water masses of theocean derived from the δ18O of benthic foraminifera (Li and Keller,1998a; Barrera and Savin, 1999) and 4 °C on land derived from leaf-margin analyses of terrestrial plants (Wilf et al., 2003). Recent studiessuggested that the impact of SO2 degassing related to flood basaltvolcanism would rather trigger climatic cooling (Chenet et al., 2005;Jones et al., 2005; Self et al., 2008). Estimations of CO2 and SO2
degassing and subsequent palaeotemperature models are somewhatconflicting with a cause-and-effect scenario between Deccan volca-nism and the end-Cretaceous warming.
A coincidence of the main Deccan volcanic activity and the end-Maastrichtian warming has been further suggested by Ravizza andPeucker-Ehrenbrink (2003) and Robinson et al. (2009). The latterdocumented a major drop in the marine 187Os/188Os record in chronC29r at Sites 525A (South Atlantic), 577A (Tropical Pacific), 690(Southern Ocean) and at the Bottacione section (Gubbio, Italy).According to these authors, this decline witnesses the initiation of theparoxysmal activity of Deccan volcanism.
Recent 40 K/40Ar dating (Chenet et al., 2007), paleomagneticinvestigations of Deccan lavas (Chenet et al., 2008, 2009) andpalaeontological investigations of intertrappean sediments in theRajahmundry area (Keller et al., 2008) suggest that ∼80% of theDeccan trap eruptions would have occurred in two distinct majorphases across the K–Pg boundary (one in the late Maastrichtian andone in the early Danian), within less than a total of 0.6 Myr in chronC29r. Chenet et al. (2009) consider that the main Cretaceous phase ofDeccan volcanism began less than 400 kyr before the K–Pg boundary.
The warming event, as recorded (1) in intermediate waters of theAtlantic by the δ18O of benthic foraminifera (Fig. 5, Li and Keller,1998a), (2) in surface waters by poleward migration of low-latitudeplanktonic foraminifer species (Huber and Watkins, 1992), reduceddiversity and dwarfism in planktonic foraminifera (Abramovich andKeller, 2003), and by the increase in abundance of Contusotruncanacontusa and Pseudotextularia elegans in the Atlantic ocean (Kucera andMalmgren, 1998; Olsson et al., 2001), initiates 100/150 kyr after the
C30n/C29r boundary (Abramovich and Keller, 2003), and wouldtherefore correlates well with the onset of the main Cretaceous pulseof Deccan volcanism. In addition, a record of increasing atmosphericpCO2 (Fig. 5), calculated from the δ13C of palaeosol carbonates atDawson Creek (Texas), argues for greenhouse conditions within chronC29r (Nordt et al., 2003). In contrast, warming in oceanic surfacewatersand in the atmosphere, as recorded in the nannofossil assemblage(Fig. 6), and by the record of terrestrial plants in North Dakota (Fig. 5),could have initiated in the topmost part of chron C30n, thus predatingthe striking warming of intermediate waters and, possibly, the mainCretaceous pulse of Deccan volcanism, by 150/200 kyr.
5.5. A multi-proxy based scenario for the end-Cretaceous warming
The correlation of biotic and abiotic events in the South Atlanticand the central Pacific, with the palaeotemperature record ofterrestrial plants and the atmospheric pCO2 record in North America(Fig. 5) suggests the following scenario: warming initiated in theatmosphere and in surface waters of the Atlantic and Pacific oceans inthe uppermost part of chron C30n around 66/66.1 Ma as indicated bythe record of terrestrial plants and nannofossil assemblages (Figs. 5and 6). At that time, waters were probably well stratified because thethermohaline circulation operated in a mode more similar to thepresent circulation during the preceding cool climate mode of theMaastrichtian (Barrera and Savin, 1999). Therefore, this progressivewarming did not expand to the underlying layers of the world oceanwhich would explain why it is not recorded in the δ18O of benthicforaminifera. During this progressive warming, fertility of surfacewaters started to decrease, as recorded by the disappearance ofBiscutum constans (Fig. 6) and the drop of Placozygus spiralis at Site577A. This is also indicated by the decrease of planktonic foraminiferalδ13C in the uppermost part of C30n at Site 525A and a slightly delayeddecrease of bulk δ13C at the C30n/C29r boundary at Site 577A (Fig. 5).At ∼65.8 Ma, within chron C29r, a huge amount of CO2 was added tothe atmosphere as indicated by the record of paleosol carbonate inTexas. The large output of CO2 to the atmosphere greatly intensifiedthe warming in the atmosphere and in oceanic surface waters asindicated by maximum temperatures in the terrestrial plant recordand by maximum abundances of the warm-water nannofossil speciesMicula murus at Sites 1258A, 577A and 525A (Fig. 5). Ecological stresswas triggered in the planktonic community by this intensifiedwarming and/or by the release of gases associated to Deccanvolcanism. This stress is recorded in the planktonic foraminiferalassemblage by the appearance of dwarfed specimens and a drop in theShannon–Weaver index at Site 525A (Fig. 5, Abramovich and Keller,2003). In the Tropical Pacific, ecological stress is also recorded innannofossil assemblages byminimum values of the Shannon diversityat Site 577A (Fig. 3) and by a minimum in the record of speciesrichness at all sites of Shatsky Rise (Fig. 4). This intensified warmingwithin chron C29r was probably sufficient to suppress stratificationbetween surface and intermediate water layers of the Atlantic andPacific oceans. The thermocline may have lowered significantly andthe surface-to-deep temperature gradient decreased abruptly, whichwould well explain the abrupt warming recorded in the δ18O ofbenthic foraminifera (Fig. 5).
It is tempting to link this large CO2 output to the Deccan volcanismon the basis of the aforementioned correlation between biotic andabiotic sedimentary proxies. A cause-and-effect scenario betweenDeccan volcanic activity and greenhouse warming, however, is beingdisputed both by the modelling of Deccan environmental impacts andrecent datings of the lava pile. The onset of a climatic trend towardshigher temperatures shown in this study in the top of C30n remainsan open question and even the link between greenhouse warmingwithin chron C29r and Deccan degassing remains to be betterdocumented. The succession, timing and duration of each volcanicphase and their correlation with the sedimentary record (Os isotopes,
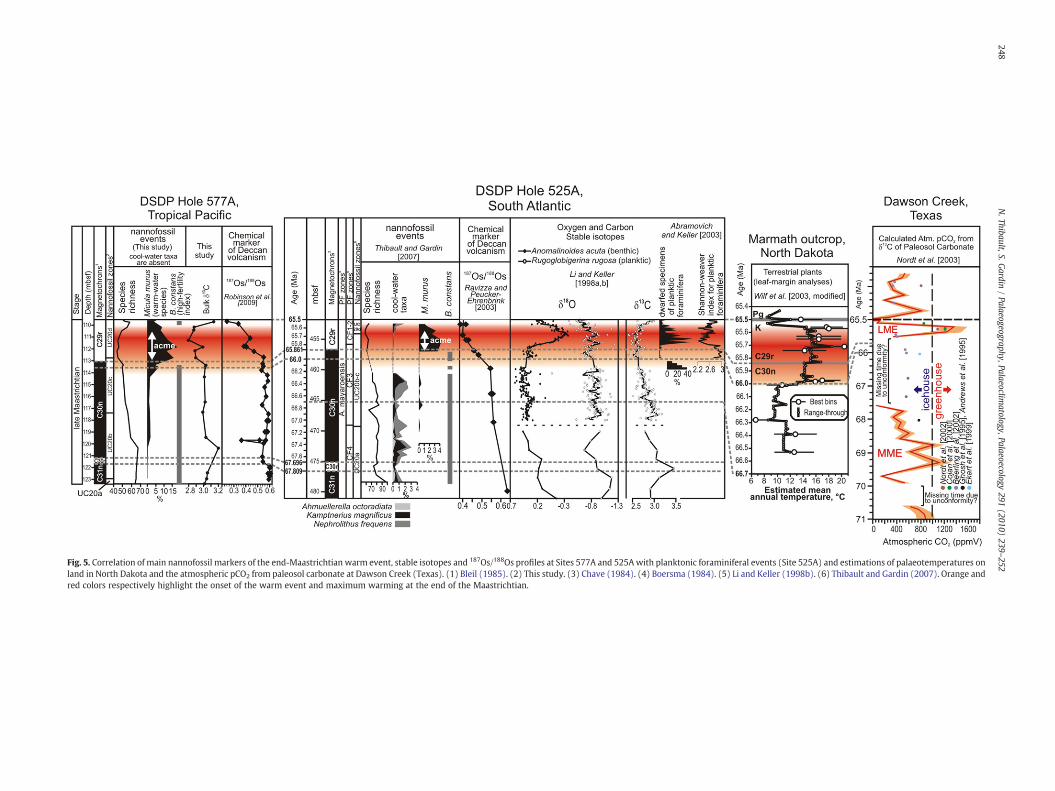
Fig. 5. Correlation of main nannofossil markers of the end-Maastrichtian warm event, stable isotopes and 187Os/188Os profiles at Sites 577A and 525Awith planktonic foraminiferal events (Site 525A) and estimations of palaeotemperatures onland in North Dakota and the atmospheric pCO2 from paleosol carbonate at Dawson Creek (Texas). (1) Bleil (1985). (2) This study. (3) Chave (1984). (4) Boersma (1984). (5) Li and Keller (1998b). (6) Thibault and Gardin (2007). Orange andred colors respectively highlight the onset of the warm event and maximum warming at the end of the Maastrichtian.
248N.Thibault,S.G
ardin/Palaeogeography,Palaeoclim
atology,Palaeoecology291
(2010)239
–252
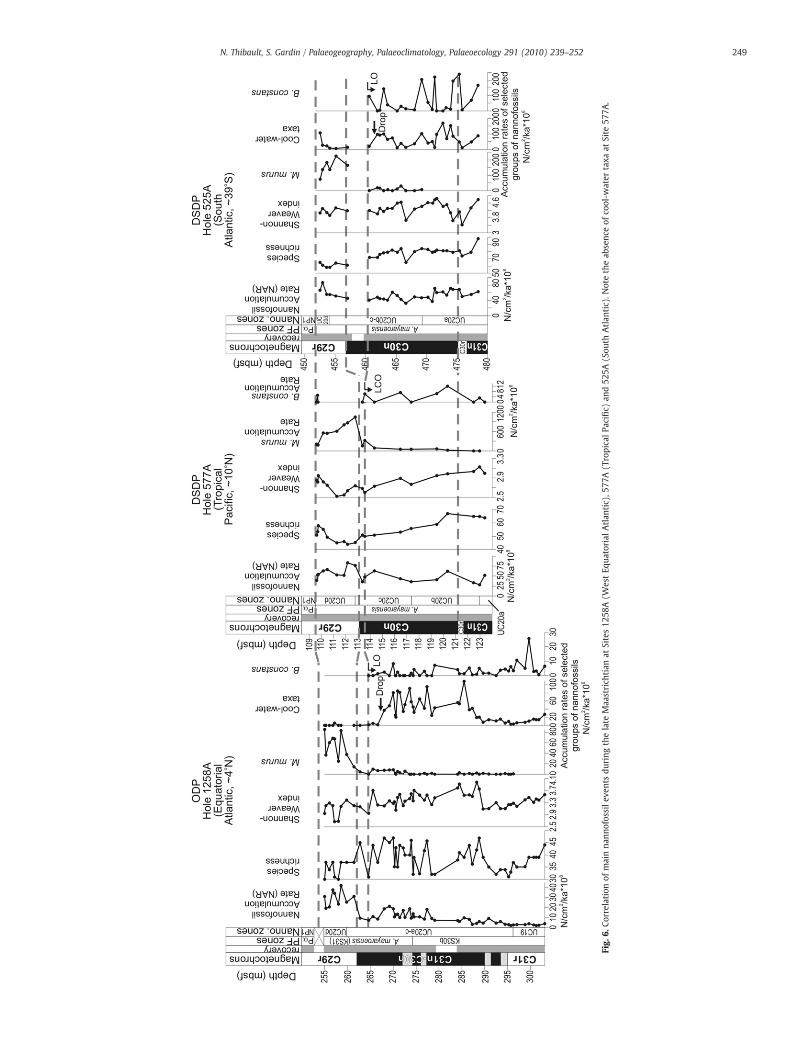
Fig.
6.Co
rrelationof
mainna
nnofossile
ventsdu
ring
thelate
Maa
strich
tian
atSites12
58A(W
estEq
uatorial
Atlan
tic),5
77A(TropicalP
acific)
and52
5A(Sou
thAtlan
tic).N
otetheab
senc
eof
cool-w
ater
taxa
atSite
577A
.
249N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252

250 N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
stable isotopes and biotic events) requires a more accurate calibrationbetween absolute and relative ages of biological and chemical eventsin the last 700 kyr of the Maastrichtian. Further high resolutionanalysis of the record of Deccan volcanic activity, climate change andbiological assemblages is necessary in order to establish sound linksbetween volcanic, biotic and abiotic events and to shed light on theCO2 vs SO2 environmental effects of Deccan degassing.
6. Conclusions
At Shatsky Rise, the end-Maastrichtian warming event wasremarkably recorded by calcareous nannofossil assemblages:
(1) An important acme of the warm-water species Micula murusmarks this event. The highest abundance of this species in theTropical Pacific, as compared to that of the Atlantic Ocean, isinterpreted as the conjunction of warming and ultra-oligotro-phic conditions.
(2) The disappearance of the high-fertility species Biscutumconstans together with lower abundances of the fertility speciesPlacozygus spiralis, higher abundances of the low-fertilityspecies Prediscosphaera cretacea and lower δ13C values suggestsa lowering of fertility levels during this warm event.
(3) An important drop of species richness recorded at all sites ofShatsky Rise and an increase of other Micula species at DSDPSite 577A suggest that this intense warming episode wascoincident with an environmental stress in the Tropical Pacific.
The comparison of nannofossil events, planktonic foraminiferalevents, stable isotope records and the marine Osmium record in theSouth Atlantic and the central Pacific, and their correlation to thepalaeotemperature record of terrestrial plants and to the atmosphericpCO2 record in North America, suggests the following:
(4) The warm event could have initiated in the atmosphere andsurface waters within the topmost part of chron C30n, andwould thus predate thewarming of intermediate watermasses.Whether its onset coincides or not with the beginning of theDeccan volcanic activity remains to be explored.
(5) Maximumwarming tookplacewithin chronC29r andwasdue tothe addition of greenhouse gas CO2 to the atmosphere, probablytriggered by the main Cretaceous pulse of Deccan volcanism.
Acknowledgements
We warmly thank Simonetta Monechi for providing us the sam-ples of Site 577A and Greg Ravizza who provided us the data on 187Os/188Os from Sites 525A and 577A. We are grateful to NathalieLabourdette, Damien Huygues and Vincent Gressier for technicalsupport and analytical help. Funding for this study was provided byEclipse II Program. We are grateful to Gerta Keller and an anonymousreviewer for critical review, many helpful suggestions and discussionsof the manuscript.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.palaeo.2010.02.036.
References
Abramovich, S., Keller, G., 2003. Planktonic foraminiferal response to the latestMaastrichtian abrupt warm event: a case study from South Atlantic DSDP Site525A. Mar. Micropaleontol. 48, 225–249.
Abramovich, S., Keller, G., Stüben, D., Berner, Z., 2003. Characterization of lateCampanian foraminiferal depth habitats and vital activities based on stableisotopes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 202, 1–29.
Backman, J., Shackleton, N.J., 1983. Quantitative biochronology of Pliocene and EarlyPleistocene calcareous nannofossils from the Atlantic, Indian and Pacific Oceans.Mar. Micropaleontol. 8, 141–170.
Barrera, E., 1994. Global environmental changes preceding the Cretaceous–Tertiaryboundary: Early-late Maastrichtian transition. Geology 22, 877–880.
Barrera, E., Savin, S.M., 1999. Evolution of Campanian–Maastrichtian marine climatesand oceans. In: Barrera, E., Johnson, C.C. (Eds.), Evolution of the Cretaceous Ocean-Climate System: Spec. Publ. Geol. Soc. Am., 332, pp. 245–282.
Bleil, U., 1985. The magnetostratigraphy of northwest Pacific sediments. Init. Rep. DeepSea Drill. Proj. 86, 441–458.
Boersma, A., 1984. Cretaceous–Tertiary planktic foraminifers from the southeasternAtlantic, Walvis Ridge area. Init. Rep. Deep Sea Drill. Proj. 74, 501–523.
Boersma, A., Schackleton, N.J., 1981. Oxygen and carbon isotope variations andplanktonic foraminiferal depth habitats: Late Cretaceous to Paleocene, CentralPacific, DSDP Sites 463 and 465, Leg 65. Init. Rep. Deep Sea Drill. Proj. 65,513–526.
Bown, P.R., 2005. Selective calcareous nannoplankton survivorship at the Cretaceous–Tertiary boundary. Geology 33 (8), 653–656.
Bralower, T.J., Premoli Silva, I., Malone, M.J., et al., 2002a. Proc. ODP, Init. Repts., 198,College Station, TX (Ocean Drilling Program). doi:10.2973/odp.proc.ir.198.2002.
Bralower, T.J., Premoli Silva, I., Malone, M.J., Scientific Participants of Leg 198, 4–10,2002b. New evidence for abrupt climate change in the Cretaceous and Paleogene:an Ocean Drilling Program expedition to Shatsky Rise, northwest Pacific. GSA Today12 (11). doi:10.1130/1052-5173(2002)012b0004:NEFACCN2.0.CO;2.
Bralower, T.J., Premoli Silva, I., Malone, M.J., 2006. Leg 198 synthesis: a remarkable 120-Ma record of climate and oceanography from Shatsky Rise, northwest PacificOcean. Proc. Ocean Drill. Program Sci. Results 198, 1–47. doi:10.2973/odp.proc.sr.198.101.2006.
Bukry, D., 1973. Coccolith and silicoflagellate stratigraphy, Tasman Sea and southwest-ern Pacific Ocean, Deep Sea Drilling Project Leg 21. Init. Rep. Deep Sea Drill. Proj. 21,885–893.
Burnett, J.A., 1998. Upper Cretaceous. In: Bown, P.R. (Ed.), Calcareous NannofossilBiostratigraphy, Chapman and Hall, Cambridge Univ. Press, Cambridge, U.K.,pp. 132–199.
Caldeira, K., Rampino, M.R., 1990. Carbon dioxide emissions fromDeccan volcanism anda K/T boundary greenhouse effect. Geophys. Res. Lett. 17 (9), 1299–1302.
Chave, A.D., 1984. Lower Paleocene–upper Cretaceous magnetostratigraphy, sites 525,527, 528 and 529, Deep Sea Drilling Project Leg 74. Init. Rep. Deep Sea Drill. Proj. 74,525–531.
Chenet, A.L., Fluteau, F., Courtillot, V., 2005. Modelling massive sulphate aerosolpollution, following the large 1783 Laki basaltic eruption. Earth Planet. Sci. Lett.236, 721–731.
Chenet, A.L., Quidelleur, X., Fluteau, F., Courtillot, V., Bajpai, S., 2007. 40K–40Ar dating ofthe Main Deccan large igneous province: Further evidence of KTB age and shortduration. Earth Planet. Sci. Lett. 263, 1–15.
Chenet, A.L., Fluteau, F., Courtillot, V., Gérard, M., Subbarao, K.V., 2008. Determination ofrapid Deccan eruptions across the Cretaceous-Tertiary boundary using paleomag-netic secular variation: results from a 1200-m-thick section in the Mahabaleshwarescarpment. J. Geophys. Res. 113, B4. doi:10.1029/2006JB004635.
Chenet, A.L., Courtillot, V., Fluteau, F., Gérard, M., Quidelleur, X., Khadri, S.F.R., Subbarao,K.V., Thordarson, T., 2009. Determination of rapid Deccan eruptions across theCretaceous-Tertiary boundary using paleomagnetic secular variation: 2. Con-straints from analysis of eight new sections and synthesis for a 3500-m-thickcomposite section. J. Geophys. Res. 114, B6. doi:10.1029/2008JB005644.
Cohen, A.S., Coe, A.L., 2002. New geochemical evidence for the onset of volcanism in theCentral Atlantic magmatic province and environmental change at the Triassic–Jurassic boundary. Geology 30 (3), 267–270.
Dessert, C., Dupré, B., François, L.M., Schott, J., Gaillardet, J., Chakrapani, G., Bajpai, S.,2001. Erosion of Deccan Traps determined by river geochemistry: impact on theglobal climate and the 87Sr/86Sr ratio of seawater. Earth Planet. Sci. Lett. 188,469–474.
D'Hondt, S., Keller, G., 1991. Some patterns of planktic foraminiferal assemblageturnover at the Cretaceous–Tertiary boundary. Mar. Micropaleontol. 17, 77–118.
Douglas, R.G., Savin, S.M., 1975. Oxygen and carbon isotope analyses of Tertiary andCretaceous microfossils from Shatsky Rise and other sites in the North PacificOcean. Init. Rep. Deep Sea Drill. Proj. 32, 509–520.
Dumitrescu, M., Brassell, S.C., Schouten, S., Hopmans, E.C., Sinninghe Damsté, J.S., 2006.Instability in tropical Pacific sea-surface temperatures during the early Aptian.Geology 34 (10), 833–836. doi:10.1130/G22882.1.
Erba, E., 1990. Middle Cretaceous calcareous nannofossils from the western Pacific (Leg129): evidence for paleoequatorial crossings. Proc. Ocean Drill. Program Sci Results129, 189–196.
Erba, E., 1992. Calcareous nannofossil distribution in pelagic rhythmic sediments(Aptian–Albian Piobicco core, central Italy). Rivista Ital. Paleontol. Stratigr. 97,455–484.
Erba, E., Castradori, F., Guasti, G., Ripepe, M., 1992. Calcareous nannofossils andMilankovitch cycles: the example of the Gault Clay Formation (southern England).Palaeogeogr., Palaeoclimatol. Palaeoecol. 93, 47–69.
Eshet, Y., Almogi-Labin, A., 1996. Calcareous nannofossils as paleoproductivityindicators in Upper Cretaceous organic-rich sequences in Israel. Mar. Micropa-leontol. 29 (1), 37–61.
Eshet, Y., Moshkovitz, S., Habib, D., Benjamini, C., Margaritz, M., 1992. Calcareousnannofossil and dinoflagellate stratigraphy across the Cretaceous/Tertiary bound-ary at Hor Hahar, Israel. Mar. Micropaleontol. 18, 199–228.
Fisher, C.G., Hay, W.W., 1999. Calcareous nannofossils as indicators of mid-Cretaceouspaleofertility along an ocean front, U.S.Western Interior. In: Barrera, E., Johnson, C.C.

251N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
(Eds.), Evolution of the Cretaceous Ocean-climate System: Spec. Publ. Geol. Soc. Am.,332, pp. 161–180.
Frank, T.D., Thomas, D.J., Leckie, R.M., Arthur, M.A., Bown, P.R., Jones, K., Lees, J.A., 2005.The Maastrichtian record from Shatsky Rise (northwest Pacific): a tropicalperspective on global ecological and oceanographic changes. Paleoceanography20, PA1008. doi:10.1029/2004PA001052.
Gerstel, J., Thunell, R.C., Zachos, J.C., Arthur, M.A., 1986. The Cretaceous/Tertiaryboundary event in the North Pacific: planktonic foraminiferal results from Deep SeaDrilling Project Site 577, Shatsky Rise. Paleoceanography 1, 97–117.
Gibbs, S.J., Bralower, T.J., Bown, P.R., Zachos, J.C., Bybell, L.M., 2006a. Shelf and open-ocean calcareous phytoplankton assemblages across the Paleocene–Eocene Ther-mal Maximum: implications for global productivity gradients. Geology 34 (4),233–236. doi:10.1130/G22381.1.
Gibbs, S.J., Bown, P.R., Sessa, J.A., Bralower, T.J., Wilson, P.A., 2006b. Nannoplanktonextinction and origination across the Paleocene–Eocene Thermal Maximum.Science 314 (5806), 1770–1773. doi:10.1126/science.1133902.
Gradstein, F.M., Ogg, J.G., 2004. Geologic Time Scale 2004, why, how, and where next.Lethaia 37, 175–181.
Hardas, P., Mutterlose, J., 2007. Calcareous nannofossil assemblages of Oceanic AnoxicEvent 2 in the equatorial Atlantic: evidence of an eutrophication event. Mar.Micropaleontol. 66, 52–69.
Henriksson, A.S., 1993. A quick and easy method for estimating absolute abundances ofcalcareous nannofossils. Int. Nannoplankton Assoc. Newsl. 15 (2), 68.
Henriksson, A.S., Malmgren, B.A., 1997. Biogeographic and Ecologic Patterns incalcareous nannoplankton in the Atlantic and Pacific Oceans during the TerminalCretaceous. Studia Geologica Salmanticensia 33, 17–40.
Hill, M.E., 1975. Selective dissolution of mid-Cretaceous (Cenomanian) calcareousnannofossils. Micropaleontology 21 (2), 227–235.
Huber, B.T., Watkins, D.K., 1992. Biogeography of Campanian–Maastrichtian calcareousplankton in the region of the Southern Ocean: Paleogeographic and Paleoclimaticimplications. In: Kennett, J.P., Warnke, D.A. (Eds.), The Antarctic Paleoenvironment: APerspective on Global Change. : Antarctic Research Series, 56. AGU, Washington, D.C,pp. 31–60.
Jenkyns, H.C., Mutterlose, J., Sliter, W.V., 1995. Upper Cretaceous carbon- and oxygen-isotope stratigraphy of deep-water sediments from the North-Central Pacific (Site869, Flank of Pikini-Wodejebato, Marshall Islands). Proc. Ocean Drill. Program Sci.Results 143, 105–108.
Jones, G.S., Gregory, J.M., Stott, P.A., Tett, S.F.B., Thorpe, R.B., 2005. AnAOGCM simulation ofthe climate response to a volcanic super-eruption. Clim. Dynam. 25 (7–8), 671–880.
Kaiho, K., Takeda, K., Petrizzo, M.R., Zachos, J.C., 2006. Anomalous shifts in tropicalPacific planktonic and benthic foraminiferal test size during the Paleocene–Eocenethermal maximum. Palaeogeogr. Palaeoclimatol. Palaeoecol. 237, 456–464.doi:10.1016/j.palaeo.2005.12.017.
Keller, G., Adatte, T., Gardin, S., Bartolini, A., Bajpai, S., 2008. Main Deccan volcanismphase ends near the K–T boundary: evidence from the Krishna-Godavari Basin, SEIndia. Earth Planet. Sci. Lett. 268, 293–311. doi:10.1016/j.epsl.2008.01.015.
Koch, C., Young, J.R., 2007. A simple weighing and dilution technique for determiningabsolute abundances of coccoliths from sediment samples. J. Nannoplankton Res.29 (1), 67–69.
Kucera, M., Malmgren, B.A., 1998. Terminal Cretaceous warming event in the mid-latitude South Atlantic Ocean: evidence from poleward migration of Contuso-truncana contusa (planktonic foraminifera) morphotypes. Palaeogeogr. Palaeocli-matol. Palaeoecol. 138 (1–3), 1–15.
Lamolda, M.A., Gorostidi, A., Paul, R.C., 1992. Quantitative estimates of calcareousnannofossil changes across the Plenus Marls (latest Cenomanian), Dover,England: implications for the generation of the Cenomanian–Turonianboundary event. Cretaceous Res. 15, 143–164.
Lees, J.A., 2002. Calcareous nannofossils biogeography illustrates palaeoclimate changein the Late Cretaceous Indian Ocean. Cretaceous Res. 23, 537–634.
Lees, J.A., Bown, P.R., 2005. Upper Cretaceous calcareous nannofossil biostratigraphy,ODP Leg 198 (Shatsky Rise, northwest Pacific Ocean). Proc. Ocean Drill. ProgramSci. Results 198, 1–60. doi:10.2973/odp.proc.sr.198.114.2005.
Lees, J.A., Bown, P.R.,Mattioli, E., 2005. Problemswithproxies?Cautionary tales of calcareousnannofossil paleoenvironmental indicators. Micropaleontology 51 (4), 333–345.
Li, L., Keller, G., 1998a. Abrupt deep-sea warming at the end of the Cretaceous. Geology26 (11), 995–998.
Li, L., Keller, G., 1998b. Maastrichtian climate, productivity and faunal turnovers inplanktic foraminifera in South Atlantic DSDP sites 525A and 21. Mar. Micro-paleontol. 33, 55–86.
Linnert, C., Mutterlose, J., Erbacher, J., 2010. Calcareous nannofossils of the Cenomanian/Turonian boundary interval from the Boreal Realm (Wunstorf, northwestGermany). Mar. Micropaleontol. 74, 38–58.
Mitchell, S.F., Ball, J.D., Crowley, S.F., Marshall, J.D., Paul, C.R.C., Veltkamp, C.J., Samir, A.,1997. Isotope data from cretaceous chalks and foraminifera: environmental ordiagenetic signals? Geology 25, 691–694.
Monechi, S., 1985. Campanian to Pleistocene calcareous nannofossil stratigraphy fromthe Northwest Pacific Ocean, Deep Sea Drilling Project Leg 86. Init. Rep. Deep SeaDrill. Proj. 86, 301–336.
Nordt, L., Atchley, S., Dworkin, S., 2003. Terrestrial evidence for two greenhouse eventsin the latest Cretaceous. GSA Today 13 (12), 4–9.
Ogg, J.G., Smith, A.G., 2004. The geomagnetic polarity time scale. In: Gradstein, F.M., Ogg, J.G., Smith, A.G. (Eds.), A Geologic Time Scale 2004. Cambridge Univ. Press, Cambridge,U.K., pp. 63–86.
Olsson, R.K., Wright, J.D., Miller, K.G., 2001. Paleobiogeography of Pseudotextulariaelegans during the latest Maastrichtian global warming event. J. Foraminiferal Res.31 (3), 275–282.
Perch-Nielsen, K., 1985. Mesozoic calcareous nannofossils. In: Bolli, H.M., Saunders, J.B.,Perch-Nielsen, K. (Eds.), Plankton Stratigraphy. CambridgeUniv. Press, Cambridge, U.K.,pp. 329–426.
Pospichal, J.J.,Wise Jr., S.W.,1990.Calcareousnannofossils across theK–Tboundary,ODPHole690C, Maud Rise, Weddell Sea. Proc. Ocean Drill. Program Sci. Results 113, 515–532.
Rampino, M.R., Self, S., 2000. Volcanism and biotic extinctions. In: Sigurdsson, H. (Ed.),Encyclopedia of Volcanoes. Academic Press, New-York, pp. 1083–1091.
Ravizza, G., Peucker-Ehrenbrink, B., 2003. Chemostratigraphic evidence of Deccanvolcanism from the marine osmium isotope record. Science 302, 1392–1395.
Robinson, N., Ravizza, G., Coccioni, R., Peucker-Ehrenbrink, B., Norris, R., 2009. A highresolution marine 187Os/188Os record for the late Maastrichtian: distinguishingthe chemical fingerprints of Deccan volcanism and the KP impact event. EarthPlanet. Sci. Lett. 281, 159–168.
Roth, P.H., 1983. Jurassic and Lower Cretaceous calcareous nannofossils in thewestern North Atlantic (Site 534): biostratigraphy, preservation, and someobservations on biogeography and paleoceanography. Init. Rep. Deep SeaDrill. Proj. 76, 587–621.
Roth, P.H., Bowdler, J.L., 1981. Middle Cretaceous calcareous nannoplankton biostra-tigraphy and oceanography of the Atlantic Ocean. In: Warme, J.E., Douglas, R.G.,Winterer, E.L. (Eds.), The Deep Sea Drilling Project: A Decade of Progress: Spec.Publ. Soc. Econ. Paleontol. Mineral., 32, pp. 517–546.
Roth, P.H., Krumbach, K.R., 1986. Middle Cretaceous nannofossil biogeography andpreservation in the Atlantic and Indian Oceans: implications for palaeoceanogra-phy. Mar. Micropaleontol. 10, 235–266.
Savin, S.M., 1977. The history of the Earth's surface temperature during the past100 million years. Annu. Rev. Earth Planet. Sci. 5, 319–355.
Schrag, D.P., DePaolo, D.J., Richter, F.M., 1995. Reconstructing past sea surfacetemperatures: correcting for diagenesis of bulk marine carbon. Geochim.Cosmochim. Acta 59, 2265–2278.
Self, S., Widdowson, M., Thordarson, T., Jay, A.E., 2006. Volatile fluxes during floodbasalt eruptions and potential effects on the global environment: a Deccanperspective. Earth Planet. Sci. Lett. 248 (1–2), 518–532.
Self, S., Blake, S., Sharma, K., Widdowson, M., Sephton, S., 2008. Sulfur and chlorine inlate Cretaceous Deccan magmas and eruptive gas release. Science 319, 1654–1657.
Shafik, S., 1990. Late Cretaceous nannofossil biostratigraphy and biogeography of theAustralian western margin. Bureau of Mineral Resources, Geology and Geophysics,Australia. Rep., 295. 164 pp.
Sheldon, E., 2008. Late Maastrichtian short-term warming events in the Boreal Realm:nannofossil evidence from Denmark. 12th INA Conference, Lyon 2008, France,Abstracts, p. 105.
Sissingh, W., 1977. Biostratigraphy of Cretaceous Calcareous Nannoplankton. Geologieen Mijnbouw 56, 37–65.
Stott, L.D., Kennett, J.P., 1990. The paleoceanographic and climatic signature of theCretaceous/Paleogene boundary in the Antarctic: stable isotopic results from ODPLeg 113. Proc. Ocean Drill. Program Sci. Results 113, 829–848.
Suganuma, Y., Ogg, J.G., 2006. Campanian through Eocene Magnetostratigraphy of Sites1257–1261, ODP Leg 207, Demerara Rise (Western Equatorial Atlantic). Proc.Ocean Drill. Program Sci. Results 207. doi:10.2973/odp.proc.sr.207.102.2006.
Tantawy, A.A.A.M., 2003. Calcareous nannofossil biostratigraphy and palaeoecology ofthe Cretaceous–Tertiary transition in the central eastern desert of Egypt. Mar.Micropaleontol. 47, 323–356.
Thibault, N., Gardin, S., 2006. Maastrichtian calcareous nannofossil biostratigraphy andpaleoecology in the Equatorial Atlantic (Demerara Rise, ODP Leg 207 Hole 1258A).Rev. Micropal. 49, 199–214.
Thibault, N., Gardin, S., 2007. The late Maastrichtian nannofossil record of climatechange in the South Atlantic DSDP Hole 525A. Mar. Micropaleontol. 65, 163–184.
Thierstein, H.R., 1980. Selective dissolution of Late Cretaceous and Earliest Tertiarycalcareous nannofossils: experimental evidence. Cretaceous Res. 2, 165–176.
Thierstein, H.R., 1981. Late Cretaceous nannoplankton and the change at theCretaceous–Tertiary boundary. In: Warme, J.E., Douglas, R.G., Winterer, E.L.(Eds.), The Deep Sea Drilling Project: A Decade of Progress: Spec. Publ. Soc. Econ.Paleontol. Mineral., 32, pp. 355–394.
Thomas, D.J., 2004. Evidence for deep-water production in the North Pacific Oceanduring the early Cenozoic warm interval. Nature 430, 65–68. doi:10.1038/nature02639.
Watkins, D.K., 1989. Nannoplankton productivity fluctuations and rhythmically-bedded pelagic carbonates of the Greenhorn Limestone (Upper Cretaceous).Palaeogeogr. Palaeoclimatol. Palaeoecol. 74, 75–86.
Watkins, D.K., Wise Jr., S.W., Pospichal, J.J., Crux, J., 1996. Upper Cretaceous calcareousnannofossil biostratigraphy and paleoceanography of the Southern Ocean. In:Moguilevsky, A., Whatley, R. (Eds.), Microfossils and Oceanic Environments.Aberystwyth Press, University of Wales, pp. 355–381.
Wignall, P.B., 2001. Large igneous provinces and mass extinction. Earth-Sci. Rev. 53 (1–2),1–33.
Wilf, P., Johnson, K.R., Huber, B.T., 2003. Correlated terrestrial and marine evidence forglobal climate changes before mass extinction at the Cretaceous–Paleogeneboundary. Proc. Natl. Acad. Sci. U. S. A. 100 (2), 599–604.
Williams, J.R., Bralower, T.J., 1995. Nannofossil assemblages, fine-fraction stableisotopes, and the paleoceanography of the Valanginian-Barremian (Early Creta-ceous) North Sea Basin. Paleoceanography 10 (4), 815–839.
Wind, F.H., 1979. Maestrichtian–Campanian nannofloral provinces of the southernAtlantic and Indian Oceans. In: Talwani, M., et al. (Ed.), Deep Sea Drilling Results inthe Atlantic Ocean: Continental Margins and Paleoenvironment. : Maurice EwingSer., 3. AGU, Washington, D.C., pp. 123–137.
Worsley, T., Martini, E., 1970. Late Maastrichtian nannoplankton provinces. Nature 225,1242–1243.

252 N. Thibault, S. Gardin / Palaeogeography, Palaeoclimatology, Palaeoecology 291 (2010) 239–252
Zachos, J.C., Arthur, M.A., 1986. Paleoceanography of the Cretaceous/Tertiary boundaryevent: inferences from stable isotopic and other data. Paleoceanography 1, 5–26.
Zachos, J.C., Arthur, M.A., Dean, W.E., 1989. Geochemical evidence for suppression ofpelagic marine productivity at the Cretaceous/Tertiary boundary. Nature 337,61–64.
Zachos, J.C., Wara, M.W., Bohaty, S., Delaney, M.L., Petrizzo, M.R., Brill, A., Bralower, T.J.,Premoli-Silva, I., 2003. A transient rise in tropical sea surface temperature duringthe Paleocene–Eocene Thermal Maximum. Science 302, 1551–1554. doi:10.1126/science.1090110.
Related Documents











