The agglutinated foraminifera from the SW Pacific bathyal sediments of the last 550 kyr: Relationship with the deposition of tephra layers Nicoletta Mancin a, ⁎, Elena Basso a , Claudia Lupi a , Miriam Cobianchi a , Bruce W. Hayward b a Department of Earth and Environment Sciences, University of Pavia, via Ferrata 1, 27100 Pavia, Italy b Geomarine Research, 49 Swainston Rd, St Johns, Auckland, New Zealand abstract article info Article history: Received 12 May 2014 Received in revised form 11 November 2014 Accepted 25 December 2014 Available online 2 January 2015 Keywords: Agglutinated foraminifera Test microstructure SEM–EDS analysis Grain selectivity Tephra deposition The agglutinated foraminiferal content from the last 550 kyr record of the IMAGES core MD 97-2114 (Chatham Rise, New Zealand) was analysed in order to detect the possible linkage existing between the composition of the grains forming the agglutinated tests and the deposition of tephras. The core was collected east of New Zealand, about 680 km from the active Taupo Volcanic Zone (TVZ) located on the North Island, thus it contains numerous macro- and microscopic tephra layers. Scanning Electron Microscopy and Energy Dispersive Spectroscopy analyses were carried out on entire aggluti- nated foraminifera as well as on sectioned specimens, sampled around and within the tephra layers. The analyses show that the studied foraminifera built structurally complex tests picking and selecting mineral and biogenic particles on the basis of their availability and abundance in the substratum, as well as their composition, size and shape. In most of the studied species, belonging to the order Textulariida, the composition of the agglutinated grains does not change when the deposition of the tephra layer strongly enriched the substratum in volcanic glass shards. Only the species Karreriella novangliae changed significantly its grain composition, mostly selecting volcanic glass fragments to cover the test surface. Nevertheless, the tephra deposition seems to influence the wall microstructure of the agglutinated tests. Textulariid specimens coming from the volcanoclastic layers have a thin- ner wall which is also characterised by a less abundant calcareous matrix with respect to the specimens sampled above or below the tephra layer. We hypothesise that the volcanic ash deposition probably interfered with the normal agglutinating process by causing the development of more aggressive waters at the sea floor which, in turn, could have induced carbonate dissolution. Our observations therefore suggest that the sediment type of the substratum is not the only control- ling factor in the construction of the agglutinated foraminiferal test and grain selection, which appears to be species-dependent. © 2015 Elsevier B.V. All rights reserved. 1. Introduction The ability of recent and fossil agglutinated foraminifera to select grains of particular composition, size or shape, sometimes organising these particles within the test wall, has been investigated for over a cen- tury (e.g. Heron-Allen and Earland, 1909; Heron-Allen, 1915; Gooday et al., 1995; Allen et al., 1998; Tuckwell et al., 1999; Mancin, 2001; Thomsen and Rasmussen, 2008; Makled and Langer, 2010; Rothe et al., 2011; Mancin et al., 2012; Armynot du Châtelet et al., 2013a, b). These studies show that the sedimentary environment determines and controls which types of grains are available and thereby influences the nature and composition of agglutinated foraminiferal tests (e.g. Armynot du Châtelet et al., 2013a and references therein). However there are still questions about how much influence the sediment has on grain selection and construction of the agglutinated test and this is further investigated here. In open-ocean bathyal environments, deep-sea sediments are main- ly carbonate pelagic biogenic mud with a relatively homogeneous com- position. In some geographic areas, such as around New Zealand, large explosive eruptions are capable of wide dispersal offshore of abundant volcanic ash. These ash falls produce sudden and important perturba- tions in the geochemical environment of both deep-sea sediments and sea-surface waters, affecting the living biota (Hess and Kuhnt, 1996; Hess et al., 2001). In the deep-sea environment, thicker tephra layers prevent oxygen from penetrating into the underlying sediments, partly modifying the complex system of deep-sea carbonate dissolution (Haeckel et al., 2001). On the other hand, in the ocean surface environ- ment, airborne volcanic particulate matter and gases can enrich shallow waters with a wide suite of chemical elements, which in turn, can influ- ence the sea-surface geochemistry and ecosystems (Frogner et al., 2001; Frogner Kockum et al., 2006; Jones and Gislason, 2008; Wall-Palmer et al., 2011). Marine Micropaleontology 115 (2015) 39–58 ⁎ Corresponding author. Tel.: +39 0382985894; fax: +39 0382985890. E-mail address: [email protected] (N. Mancin). http://dx.doi.org/10.1016/j.marmicro.2014.12.004 0377-8398/© 2015 Elsevier B.V. All rights reserved. Contents lists available at ScienceDirect Marine Micropaleontology journal homepage: www.elsevier.com/locate/marmicro

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Marine Micropaleontology 115 (2015) 39–58
Contents lists available at ScienceDirect
Marine Micropaleontology
j ourna l homepage: www.e lsev ie r .com/ locate /marmicro
The agglutinated foraminifera from the SW Pacific bathyal sediments ofthe last 550 kyr: Relationship with the deposition of tephra layers
Nicoletta Mancin a,⁎, Elena Basso a, Claudia Lupi a, Miriam Cobianchi a, Bruce W. Hayward b
a Department of Earth and Environment Sciences, University of Pavia, via Ferrata 1, 27100 Pavia, Italyb Geomarine Research, 49 Swainston Rd, St Johns, Auckland, New Zealand
⁎ Corresponding author. Tel.: +39 0382985894; fax: +E-mail address: [email protected] (N. Mancin
http://dx.doi.org/10.1016/j.marmicro.2014.12.0040377-8398/© 2015 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 12 May 2014Received in revised form 11 November 2014Accepted 25 December 2014Available online 2 January 2015
Keywords:Agglutinated foraminiferaTest microstructureSEM–EDS analysisGrain selectivityTephra deposition
The agglutinated foraminiferal content from the last 550 kyr record of the IMAGES core MD 97-2114 (ChathamRise, New Zealand) was analysed in order to detect the possible linkage existing between the composition of thegrains forming the agglutinated tests and the deposition of tephras. The core was collected east of New Zealand,about 680 km from the active Taupo Volcanic Zone (TVZ) located on the North Island, thus it contains numerousmacro- and microscopic tephra layers.Scanning Electron Microscopy and Energy Dispersive Spectroscopy analyses were carried out on entire aggluti-nated foraminifera aswell as on sectioned specimens, sampled around andwithin the tephra layers. The analysesshow that the studied foraminifera built structurally complex tests picking and selecting mineral and biogenicparticles on the basis of their availability and abundance in the substratum, as well as their composition, sizeand shape. Inmost of the studied species, belonging to the order Textulariida, the composition of the agglutinatedgrains does not change when the deposition of the tephra layer strongly enriched the substratum in volcanicglass shards. Only the species Karreriella novangliae changed significantly its grain composition, mostly selectingvolcanic glass fragments to cover the test surface. Nevertheless, the tephra deposition seems to influence thewallmicrostructure of the agglutinated tests. Textulariid specimens coming from the volcanoclastic layers have a thin-ner wall which is also characterised by a less abundant calcareousmatrix with respect to the specimens sampledabove or below the tephra layer.We hypothesise that the volcanic ash deposition probably interfered with the normal agglutinating process bycausing the development ofmore aggressivewaters at the sea floor which, in turn, could have induced carbonatedissolution. Our observations therefore suggest that the sediment type of the substratum is not the only control-ling factor in the construction of the agglutinated foraminiferal test and grain selection, which appears to bespecies-dependent.
© 2015 Elsevier B.V. All rights reserved.
1. Introduction
The ability of recent and fossil agglutinated foraminifera to selectgrains of particular composition, size or shape, sometimes organisingthese particleswithin the test wall, has been investigated for over a cen-tury (e.g. Heron-Allen and Earland, 1909; Heron-Allen, 1915; Goodayet al., 1995; Allen et al., 1998; Tuckwell et al., 1999; Mancin, 2001;Thomsen and Rasmussen, 2008; Makled and Langer, 2010; Rotheet al., 2011; Mancin et al., 2012; Armynot du Châtelet et al., 2013a, b).These studies show that the sedimentary environment determinesand controls which types of grains are available and thereby influencesthe nature and composition of agglutinated foraminiferal tests (e.g.Armynot du Châtelet et al., 2013a and references therein). Howeverthere are still questions about how much influence the sediment has
39 0382985890.).
on grain selection and construction of the agglutinated test and this isfurther investigated here.
In open-ocean bathyal environments, deep-sea sediments aremain-ly carbonate pelagic biogenic mudwith a relatively homogeneous com-position. In some geographic areas, such as around New Zealand, largeexplosive eruptions are capable of wide dispersal offshore of abundantvolcanic ash. These ash falls produce sudden and important perturba-tions in the geochemical environment of both deep-sea sediments andsea-surface waters, affecting the living biota (Hess and Kuhnt, 1996;Hess et al., 2001). In the deep-sea environment, thicker tephra layersprevent oxygen from penetrating into the underlying sediments, partlymodifying the complex system of deep-sea carbonate dissolution(Haeckel et al., 2001). On the other hand, in the ocean surface environ-ment, airborne volcanic particulatematter and gases can enrich shallowwaters with a wide suite of chemical elements, which in turn, can influ-ence the sea-surface geochemistry and ecosystems (Frogner et al., 2001;Frogner Kockum et al., 2006; Jones and Gislason, 2008; Wall-Palmeret al., 2011).

40 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
This work aims to provide new evidence on the response of deep-sea agglutinated foraminifera to tephra deposition recorded in anupper Pleistocene core (IMAGES MD 97-2114), recovered east of NewZealand in the SW Pacific Ocean (Fig. 1). The target is to determine ifthe agglutinated grain composition of selected foraminiferal specieschanged as a result of tephra deposition, in response to the enrichmentof volcanic glass shards within the substratum. Scanning Electron Mi-croscopy (SEM) and Energy Dispersive Spectroscopy (EDS) analyses,carried out on the agglutinated tests picked around and within themajor tephra layers, were used to address the question.
2. Pleistocene rhyolitic tephra layers recorded offshore ofNew Zealand
The Taupo Volcanic Zone (TVZ) in New Zealand is one of the mostactive and productive rhyolitic systems on Earth and it is part of a south-ward propagating rifted arc, which forms the southern portion ofthe Kermadec subduction zone (Fig. 1). This area has been characterisedby voluminous and highly explosive eruptions, starting from ca. 1.6 Ma,which dispersed rhyolitic and andesitic ashes widely offshore aroundNewZealand (e.g. Carter et al., 1995;Wilson et al., 1995). The regionallywidespread occurrence of volcanic ash in deep-sea sediment drift de-posits east of New Zealand, wasmostly driven by the prevailingwester-ly wind field coupled with the relatively small size of the New Zealandmicro-continent (Carter et al., 1995).
Fig. 1. Synthetic map of the area around New Zealandwith the studied Images Site MD 97-2114the main water masses, fronts and currents as well as the location of the active Taupo Volcanicfigure legend, the reader is referred to the web version of this article.)Partly modified after Hayward et al. (2010).
Tephras, sourced from the TVZ, form important marker beds in theoffshore sedimentary successions, as documented in the deep-sea coreODP 1123 (Alloway et al., 2004, 2005; Carter et al., 2004). In this core,drilled on the northern side of the Chatham Rise close to the studiedsite IMAGES MD 97-2114 (Fig. 1), at least 12 tephra layers post-datingthe magnetostratigraphic Brunhes/Matuyama boundary (ca. 780 ka)were identified by Alloway et al. (2005). These tephras, 1 to 10 cmthick, mainly consist of microscopic glass shards with a chemical com-position matching the rhyolitic compositional field (ranges for SiO2 of71–78 wt.% and K2O + Na2O of 6.5–9.0 wt.%). Some of them, such asthe uppermost Pleistocene Kawakawa and Omataroa tephras, can beeasily characterised and regionally distinguished on the basis of a fewmajor elements in the glass shards, e.g. FeO and CaO (varying respec-tively, from 1.0 to 1.3 wt.% and from 0.9 to 1.3 wt.%) (Alloway et al.,2005 and references therein).
The studied IMAGES core (42°22′27″S; 171°20′42″W), collected at1936 m depth and about 680 km from the TVZ (Fig. 1), displays a con-tinuous record of the last 1.07 Ma, with an average accumulation rateof ca. 2.6 cm/ka (Cobianchi et al., 2012). The studied portion (ca. 16 mlong, recording the last 550 kyr) consists mostly of carbonate pelagicand hemipelagic biogenic mud intercalated with micro- and macro-scopic tephra layers characterised by abrupt bases and normal grading(Fig. 2). Eleven tephra levels are preserved throughout the studied in-terval, while others are only present as crypto-tephra. The majorvolcaniclastic layers were identified in the studied washed residualson the basis of both a strong enrichment in glass shards and a marked
and the neighbour site ODP 1123. The surface oceanographywith the relative position ofZone (TVZ in yellow) are indicated. (For interpretation of the references to colour in this

A
B
Fig. 2. A: Stratigraphic correlation of the major tephras occurring within cores ODP 1123and MD 97-2114 (in orange). The correlation is based on a combination of orbitally-tuned stable-isotope data coupledwith the increased abundance of glass fragments docu-mented in both cores. Yellow lines (labelled as T1 to T4)mark the tephras recognised onlyin the core MD 97-2114. Dating of tephra marker beds follows Carter et al. (2004) partlyupdated by Alloway et al. (2005). The standard LR04 curve (on the left) of Lisiecki andRaymo (2005) is also indicated. Interglacials are shaded. B: Image of a macroscopic tephralayer recorded in the studied core; note the typical abrupt base followed by normalgrading. (For interpretation of the references to colour in this figure legend, the reader isreferred to the web version of this article.)
41N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
decrease in abundance of foraminiferal tests (Fig. 2). The stratigraphiccorrelation of the tephra layers in the sites MD 97-2114 and in the adja-cent ODP 1123 is mainly based on a combination of orbitally-tuned sta-ble oxygen-isotope data and on the abundance peaks of glass fragments
(Fig. 2). The isotope data and the counting of glass fragments in site ODP1123 are, respectively, from Hall et al. (2001) and Crundwell et al.(2008).
3. Methods
A total of 130 samples (about 3 cm3 of mud, every 10 cm downcoreand corresponding to a time-resolution of ca. 4 kyr) were analysed fortheir agglutinated foraminiferal content. The analysis developed withan increasing resolution through four subsequent steps, following themethodology recently described by Mancin et al. (2014).
The first step consisted of standard micropaleontological analysesusing a stereomicroscope to determine and quantify the total abun-dance of the agglutinated foraminifera. The whole (and not aliquots)N150 μm and 63–150 μm fractions of each sample were analysed; allthe recognised agglutinated foraminifera, together with the calcareousimperforate genus Sigmoilopsis, were picked, identified (to specieslevel when possible) and counted. Benthic foraminiferal census countshave been made on approximately 200 benthic specimens, whereverpossible (the census count dataset was already available from Mancinet al., 2013).
Quantitative data are synthesised in Fig. 3 and Table 1 (available on-line as Supplementary data). Agglutinated foraminiferal taxonomymostly follows Hayward et al. (2010).
In the second step, selected agglutinated specimens were mountedon stubs using carbon conductive adhesive tape and gold-coated foranalyses by SEM (inBeam technique) at the CISRiC-Arvedi Laboratory.The SEM (Tescan FESEM, series Mira 3XMU) observations were madewith increasingmagnification, at 15mmworking distance, using an ac-celerating voltage of 15 kV. The topographic images resulting from sec-ondary electrons were used to provide high quality three-dimensionaldetails of the agglutinated test surface; some of the collected imagesare presented in Plates 1 and 2. In order to ascertain the chemical–min-eralogical variability of the grains forming the agglutinated test surfaceof some selected species, two-dimensional X-ray maps, which permitthe scanning by the electron beam of a selected area by the SEM controlsystem, were also collected. The elemental maps were provided using aresolution of 256 × 256 pixels at 300–500 × magnification using theGenesis software. Results are reported in Plate 3.
In the third step, representative agglutinated specimens were sam-pled from around and within the major tephra layers, using the follow-ing strategy: 1 to 3 individuals sampled under the tephra layer (wherepossible, ca. 10 cm below), 1 to 3 individuals within and 1 to 3 individ-uals just above (where possible ca. 10 cm above). A total of 25 foraminif-eral specimenswere picked fromaround the tephras AT-390, AT-334, T2andT3 (Fig. 3 and Table 1, available in the data repository). The collectedspecimens were then oriented and embedded in epoxy resin, cut andpolishedwith diamond pastes from0.25 to 6 μm in grain-size and finallyanalysed through the SEM equipped with an X-ray EDS. Back ScatteredElectron (BSE) images of cross-sectioned specimens highlighted compo-sitional dissimilarities among grains throughout the wall thickness, onthe basis of the mean atomic number of each grain forming the test.
The last step consisted of standardless spot microanalyses fordetermining the elemental composition of the single grains formingthe agglutinated wall. The measurements were collected for 100 s at15.8 mm working distance and using an accelerating voltage of 20 kv;chemical data are synthesised in Table 2 (available as Supplementarydata). In order to check the chemical variability of the agglutinatedgrains within the sectioned test wall of each specimen, two-dimensional X-ray maps of selected elements were also provided.Each elemental map was collected simultaneously, since the colour in-tensity results proportional to the element concentration in everypixel of the image (in black areas the element is lacking) the comparisonof the maps collected in the same area of the sample provided an over-view of the mineralogical distribution in that area. The results are re-ported in Plates 4–8.

Fig. 3. Distribution curves reporting, from left to right, the relative abundance of glass fragments and of benthic foraminifera (benthos %), the relative abundance of agglutinated forami-nifera (%) and of the calcareous imperforate species Sigmoilopsis schlumbergeri (%) (these last percentages calculated with respect to the total benthic foraminiferal fauna) together withthe distribution curves showing the total number of agglutinated specimens belonging to the most common foraminiferal taxa counted in the whole N150 and 150–63 μm fractions. Theidentified tephra layers are also reported (on the left). Red points indicate the agglutinated specimens picked around and within some key tephra layers. Interglacials are shaded.(For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
42 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
The term “cement” is commonly used to refer to thematerial secret-ed by the organism during the agglutination process (e.g. Wood, 1949;Bender andHemleben, 1988; Loeblich and Tappan, 1989). Thismaterial,binding the grains together, can be calcareous (as in Textulariida), or-ganic (as in Astrorhizida) or of a mixed nature (as in Lituolida andLoftusiida). In fossil specimens, the organic parts are not preservedthus it is quite difficult to recognise the secondary calcite cement depos-ited within the cavities previously filled by organic material, hence weprefer to use the termmatrix to indicate the fine material that enclosesthe agglutinated grains.
4. Results
4.1. Abundance/distribution patterns of the recordedagglutinated foraminifera
In all the samples the relative abundance of agglutinated taxa usuallydid not exceed 4% of the benthic foraminiferal assemblages (Fig. 3).
Plate 1. SEM pictures of some agglutinated foraminifera from the last 550 kyr record of the coregrainsweremostly coccoliths belonging to the genera Coccolithus (Co), Calcidiscus (Ca),Helicospeach agglutinated species seemingly selected a particular nannofossil genus and/or species. The10.92 mbsf (metres below sea floor); a–b) details of the test surface made of coccoliths of thecalcareous matrix; and c) detail of the crushed test wall which internally displays a coccolith.coccoliths of the genus Helicosphaera. 3. Textularia aff. pseudogramen; at 0.50 mbsf; a) detail oPontosphaera. 4. Siphotextularia rolshauseni Phleger & Parker, 1951, at 11.12 mbsf; a–b) detailNote that the coccoliths are regularly arranged with their proximal shields oriented towardVella, 1957, at 14.94 mbsf; a) detail of the test surface made of coccoliths of two different nanat 12.41 mbsf; a) detail of the test surface formed by mineral fragments regularly arranged. 7.equidimensional mineral fragments.
More than 1500 agglutinated specimens, including specimens of themiliolid Sigmoilopsis schlumbergeri, were isolated and identified fromthe studied record (Table 1, available as Supplementary data). Theorder Textulariida was the most abundant and diverse (Plates 1 and 2and Table 1, in the data repository); rare and sporadically distributedspecimens belonging to the orders Astrorhizida (Rhabdammina antarcti-ca and Saccorhiza sp.) and Lituolida (Reophax sp.) were also found(Plate 2, images 4–5). The studied specimens were usually quite wellpreserved although some elongate taxa (e.g. Martinottiella communisand R. antarctica) were mostly fragmented tests.
The graphs of the total abundance of themost common agglutinatedtaxa (Fig. 3) show that all species were sparsely and discontinuouslyrecorded throughout the studied record, with the exception ofS. schlumbergeri that has higher average abundances, particularly inthe upper part of the core. Only Eggerella bradyi, Karreriella bradyi,Karreriella novangliae, Siphotextularia rolshauseni and S. schlumbergerioccurring around and within some tephra layers were represented bysufficient specimens to be picked and sectioned for SEM–EDS analyses(Fig. 3; Table 1, Supplementary data).
MD-97-2114which carefully selected the grains to form the agglutinated test surface. Thehaera (He) and Pontosphaera (Po) aswell as equidimensionalmineral fragments. Note thatimages are in secondary electrons (SE). 1. Eggerella bradyi (Cuschman, 1911), sampled at
species Coccolithus pelagicus and Calcidiscus leptoporus embedded within the fine-grained2. Karreriella bradyi (Cushman, 1911), at 15.18 mbsf; a) detail of the test surface made off the test surface made of coccoliths of two different nannofossil genera, Coccolithus ands of the test surface exclusively formed by coccoliths of the species Coccolithus pelagicus.s the outside to form a homogeneous external pavement. 5. Siphotextularia mestayeraenofossil genera, Coccolithus and Helicosphaera. 6. Siphotextularia flintii (Cuschman, 1911),Karreiella novangliae (Cushman, 1922), at 9.82 mbsf; a) detail of the test surface made of

43N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

44 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
4.2. Characterisation of the agglutinated test surface
Most of the studied agglutinated species (over 60 specimens ran-domly sampled throughout the whole record) selected the grains tobuild their test surfaces (Plates 1 and 2). Some of them, such asE. bradyi (Plate 1, images 1, 1a–b), K. bradyi (2, 2a), Siphotextulariapseudogramen (3, 3a), S. rolshauseni (4, 4a–b) and Siphotextulariamestayerae (5, 5a), picked exclusively coccoliths, sometimes evenchoosing a particular nannofossil genus or species. K. bradyi mostlytook elliptical coccoliths belonging to the genus Helicosphaera (2a),whereas S. rolshauseni exclusively picked discoidal coccoliths of thespecies Coccolithus pelagicus (4a–b). On the other hand, E. bradyi,S. pseudogramen and S.mestayerae did not select a particular nannofossilgenus or species making their test surface with different coccolith taxa(e.g. the genera Coccolithus, Calcidiscus, Pontosphaera, Helicospaera)which co-occur in the same agglutinated specimen (e.g. Plate 1, 1b;3a; 5a).
Moreover, the arrangement of the nannofossils at the surface of theagglutinated test is not always fortuitous. In both K. bradyi (Plate 1, 2a)and S. rolshauseni (Plate 1, 4b), the coccoliths were embedded intoa very fine-grained matrix and arranged in an orderly way, withthe proximal shields oriented towards the outside to form a veryregular test surface. This coccolith disposition created an external pave-ment made of equidimensional calcite particles. Conversely, in theS. pseudogramen and S. mestayerae tests, the coccoliths were differentin size and shape and arranged randomly (Plate 1, 3a; 5a).
In other agglutinated species, such as Siphotextularia flintii andK. novangliae, the grains forming the agglutinated test surface wereexclusively composed of mineral fragments, mostly equidimensionaland placed in an orderlyway (Plate 1, 6a; 7a). Even R. antarctica selectedthe agglutinated grains, mostly glass shards, on the basis of theirchemical–mineralogical composition, but both the grain size and thegrain arrangement on the test surface are quite chaotic (Plate 2, 4a).
These characteristics occur constantly in all the studied specimensbelonging to the same species, even when they were picked withinsamples from different stratigraphic intervals (e.g. within glacial or in-terglacial phases). The test homogeneity persisted during ontogeny, asno significant changes in grain composition and arrangement wereobserved within the same specimen (Plate 1, images 1–7).
In spite of the particular grain selection and arrangement document-ed in some of the studied species, other agglutinated foraminifera builttests without selecting the grains. Within the textulariid group, the spe-ciesM. communis, Textularia lythostrota, and Spiroplectammina sagittulapicked heterogeneous grains, composed of both mineral fragmentsand biogenic remains and characterised by different sizes and shapes(Plate 2, 1a, 2a and 3a). Likely, Saccorhiza sp. (order Astrorhizida) andS. schlumbergeri (order Miliolida) did not select any particular grain;they probably picked what the substratum contained (Pl. 2, 5a and 6a,respectively).
In order to better document the chemical–mineralogical variability(or homogeneity) of the test surface of those specieswhich agglutinatedmostly mineral grains (e.g. S. flintii, Karreriella novangliae, R. antarticaand S. schlumbergeri), X-ray maps were collected (Plate 3). AlthoughS. flintii, K. novangliae and S. schlumbergeri were characterised by asimilar test surface composed of mineral fragments (Plates 3, 1,2,4),the chemical–mineralogical composition of the selected grains wasquite diverse. S. flintii mostly agglutinated grains of albite and quartz;K. novangliae mostly selected volcanic glass fragments and plagioclase;and S. schlumbergeri picked heterogeneous grains made of quartz,volcanic glass, plagioclase and rarely k-feldspar. Spot-microanalysesperformed on single grains in each species as well as the reported ele-mental maps of Si, Na and Ca confirm these results (Plate, 3; Table 2,Supplementary data). For R. antarctica, no elemental mapswere collect-ed because the grain composition was homogeneous and the testswere exclusively made of coarse-grained shards of volcanic glass(Plates 3, 3a–b).
Our results also show that, among the studied agglutinated forami-nifera, only the species R. antarctica and K. novangliae selected glassfragments to build their agglutinated test surface (Plate 3, images 2and 3a). Therefore these species could be the best candidates to testthe possible linkage between the composition of the agglutinated grainsand the deposition of tephras. Unfortunately, the rare abundance ofR. antarctica (only 3 fragmented specimens was isolated throughoutthewhole studied record; Table 1, Supplementary data) made it impos-sible to collect this species with the same sampling strategy used for theother selected species (Fig. 3).
4.3. Test wall microstructure and composition of the agglutinated grains
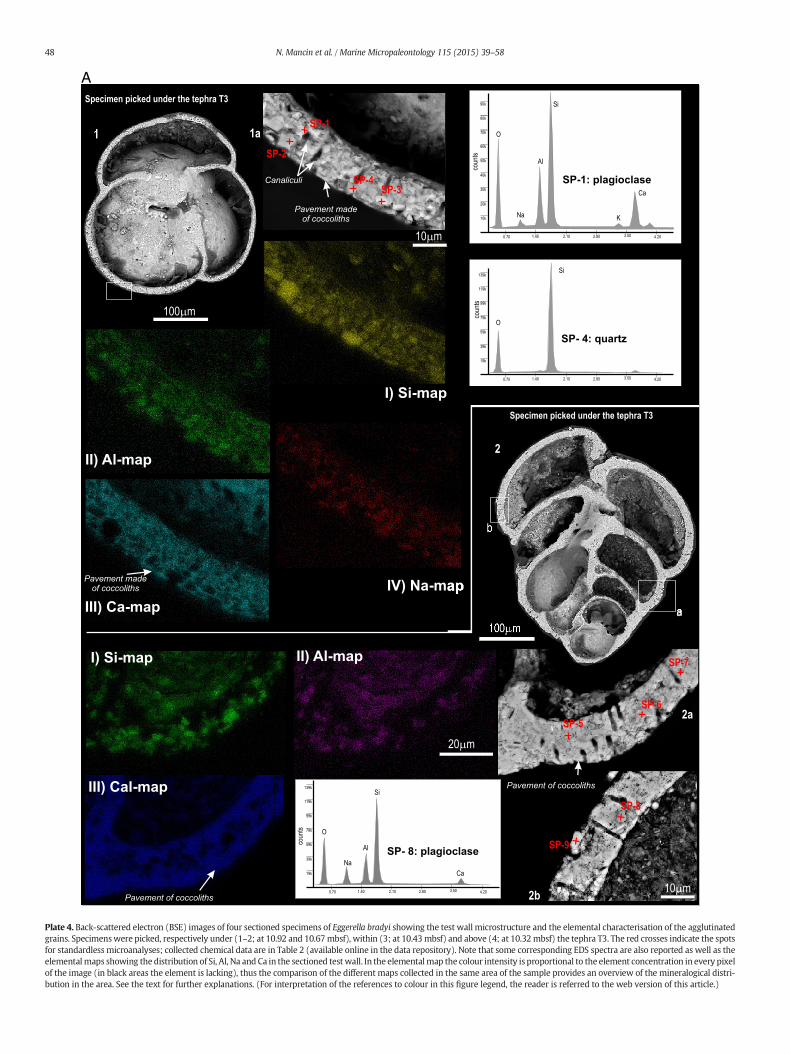
4.3.1. E. bradyi and K. bradyi (Plates 4A–B; 5A–B)Sectioned specimens document that in both species, under the sur-
face made of coccoliths (Plates 1, 1a–b, 2a), the test wall microstructurewas more complex and characterised by some recurring features. Theyshowed: i) a layered structure of the caniculate wall, with the largestgrains arranged close to the external surface and progressively smallergrains towards the inside; ii) a wall internally composed of mineralgranules and externally by the thin calcareous pavement made ofcoccoliths and, iii) agglutinated grains which were always embeddedinto a homogeneous fine-grained calcareous matrix (Plate 4, images1a, 2a–b, 3a, 4a; Plate 5, images 1a, 2a, 3a–b, 4a–b).
Spot microanalyses carried out on single grains (Plates 4, 1a, 2a–b,3a; Plates 5, 1a, 2a, 3b, 4b) showed that they were mostly silicate min-erals, such as: plagioclase, k-feldspar, rarely quartz, and rhyolitic volca-nic glass (Table 2 in the data repository). Limited to E. bradyi, the silicategranules co-occurredwith somenannofossil remains (e.g. Plates 4B, 3a).
The complexity of the test wall is more evident if we compare theelemental maps of Si, Al, Ca and Na collected for each of the sectionedspecimens (e.g. Plate 4A, specimen 2; Plate 5B, specimen 4). Si, Al andNa, considered as distinctive elements of Na-plagioclase, were mostlyconcentrated within the internal portion of the wall, while Ca occurredmainly towards the surface of the test (the external pavement ofcoccoliths) and was also widespread within the wall (the matrix). Afurther result is that the chemical–mineralogical composition of theagglutinated grains did not manifestly change if we compare E. bradyiand K. bradyi specimens sampled, respectively, below (Plate 4A, images1a, 2a–b; Plate 5, 1a), within (Plate 4B, 3a; Plate 5A, 2a) and above(Plate 4B, 4a; Plate 5B, 3b, 4b) the tephra layers. Although the tephra de-position strongly enriched the substratum in volcanic glass shards, noneof the specimens sampled within the tephra layers (Plate 4B, specimen3; Plate 5A, specimen 2) increased significantly the content of glass frag-ments in their test wall; this evidence is well supported by the Si-maps.However, the tephra deposition seems to influence much more themicrostructure of the agglutinated wall. The specimens of K. bradyifrom the tephra layer (Plate 5A; 2, 2a) developed a thinner test wall(b20 μm thick), with less abundant calcite matrix and characterisedby the absence of the external pavementmade of coccoliths. These char-acteristics are documented by the open entrance of canaliculi on bothsides of thewall (Plate 5A; 2a) and by the Ca-map (Plate 5A).Moreover,the grainswere still arranged to form the typical layered structurewith-in thewall but the grain arrangementwas less evident than in the spec-imens sampled below (Plate 5A, 1a) or above (Plate 5B, 3a–b, 4a–b) thetephra layer.
4.3.2. K. novangliae (Plates 6A–B)The sectioned specimens of this species show again a complex wall
microstructure characterised by an external surface of the test exclu-sively composed of larger mineral granules (Plates 6A–B, images 1a,1c, 2a, 2c, 3a, 3b) and towards the inside, calcareous nannofossilremains which co-occurred with very-fine grained silicate minerals(images 1b, 2b). Spot microanalyses carried out on the single grainsshow that they were mostly plagioclase (albite or Na–plagioclase),k-feldspar, volcanic glass and quartz (Table 2, Supplementary data).

Plate 2. SEM pictures of some agglutinated foraminifera from the last 550 kyr record of the core MD-97-2114 which did not select the grains to form the agglutinated test surface. Thegrains were biogenic remains as well as mineral fragments and characterised by heterogeneous sizes and shapes. The images are in secondary electrons (SE). 1.Martinottiella communis(d'Orbigny, 1846), at 8.47mbsf; a) detail of the test surface formed bymineral fragments of different sizes and shapes. 2. Textularia lythostrota (Schwager, 1866), at 12.41mbsf; a) detail ofthe test surface formed by both mineral fragments and planktonic foraminiferal test remains (white arrow) of different sizes and shapes. 3. Spiroplectammina (ex Textularia) sagittulaDefrance, 1824, at 0.74 mbsf; a) detail of the test surface mostly formed by biogenic remains of different sizes and shapes. 4. Rhabdammina antarctica Saidowa, 1975, at 3.02 mbsf;a) detail of the test surface formed by mineral fragments of different sizes and shapes; the grains are exclusively coarse-grained volcanic glass shards. 5. Saccorhiza cf. ramosa (Brady,1879), at 15.18 mbsf; a) detail of the agglutinated test surface formed by heterogeneous grains. 6. Sigmoilopsis schlumbergeri (Silvestri, 1904), at 15.74 mbsf; a) detail of the test surfacemade of mineral fragments of different sizes and shapes.
45N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

Plate 3. Back-scattered electron (BSE) images of the agglutinated test surface of the species Siphotextularia flintii (1), Karreriella novangliae (2), Rhabdammina antarctica (3) and Sigmoilopsisschlumbergeri (4) and the associated combination of elemental analysis mappings (Si, Na and Ca). In the elemental map the colour intensity is proportional to the element concentration inevery pixel of the image (in black areas the element is lacking), thus the comparison of thedifferentmaps collected in the same area of the sample provides an overviewof themineralogicaldistribution in the area. The spots for standardless microanalyses are also indicated (blue crosses); as an example one EDS spectrum (volcanic glass) is also indicated. Collected chemicaldata are in Table 2 (available online in the data repository). (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
46 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

47N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
This evidence is supported by the elemental maps of Si, Al, Na and Cacollected for each of the sectioned specimens (Plates 6A–B). Notewor-thy is the fact that specimens of K. novangliae sampledwithin the tephralayer (Plate 6A, 2, 2a–c), showed a significant increase of glass shards inthe wall and particularly on the surface of the test (2a). This evidence issupported by the Si-map, which displays amassive concentration of sil-icon in most of the grains forming the outer portion of the wall.
In contrast, the internal part of the wall is made of smaller plagio-clase grains, whichwere internally covered by a calcite surface probablyof coccoliths (Plate 6A, Ca-map). Moreover, the results show that thetephra deposition caused the variation of the chemical–mineralogicalcomposition of the selected grains and also influenced the microstruc-ture of the agglutinated wall. It became thinner (b20 μm thick) andwas characterised by less abundant calcite matrix (Plate 6A, 2a–c).Both of these characters changed in the specimens sampled fromabove the tephra (Plate 6B); here the specimens had a thicker wall(Plate 6B, 3a–b), with a more developed calcareous matrix (Ca-map),and with the wall of the test formed mainly of plagioclase grains (Aland Na map), and with a minor amount of glass fragments (Si-map).
4.3.3. S. rolshauseni (Plates 7A–B)Among the studied textulariids, S. rolshauseni is the only species that
did not select mineral grains to build the test wall. Instead it exclusivelyconsisted of a single layer of coccoliths (C. pelagicus), embedded in avery-fine grained calcareous matrix (Plate 7, images 1a–b; 2a, 3a, 4a–b). The composition of the agglutinated particles did not change in thespecimens whether sampled beneath (1a–b), within (2a) or above (3aand 4a–b) the tephra layer. None of these specimens selected glass frag-ments or other silicate minerals to build the test wall.
4.3.4. S. schlumbergeri (Plates 8A–B)The test wall of this species typically had two distinct portions: an
inner calcareous layer secreted by the foraminifer and an outer andthicker agglutinated portion (Plate 8 1a, 2a, 3a–b). Within the aggluti-nated portion, the grains were never selected in function of size andshape, thus resulting in a heterogeneous arrangement. Even from acompositional point of view, the grains were different and composedof silicate minerals, such as k-feldspar, plagioclase, quartz, and morerarely volcanic glass and pyroxene; the matrix was formed by calcite.The grain compositional variability is confirmed by both spot micro-analyses (Plate 8, 1a, 2a, 3b; Table 2, in the data repository) and elemen-tal maps. It is noteworthy that these features persisted unchanged in allthe studied specimens, even in those sampled within the tephra layers(image 2a).
5. Discussion
5.1. Agglutinated foraminiferal test construction
The substratum on or within which the agglutinated foraminiferalive is the source of the material with which they build their test. How-ever, numerous papers report that the agglutinated grains of recent andfossil foraminifera do not exclusively depend on the sediment composi-tion, because the foraminiferal cell is able to sort the grains by composi-tion, size and shape (e.g. Bowser and Bernhard, 1993; Gooday et al.,1995; Allen et al., 1998; Bertram and Cowen, 1998; Tuckwell et al.,1999; Thomsen and Rasmussen, 2008; Makled and Langer, 2010;Rothe et al., 2011; Mancin et al., 2012; Armynot du Châtelet et al.,2013a). Two different modes of grain selection appear mainly usedby the agglutinated foraminifera when they build their test in threedimensions: selection according to specific chemical–mineralogicalcomposition (e.g. Gooday et al., 1995; Mancin, 2001; Armynot duChâtelet et al., 2008; Makled and Langer, 2010) and selection accordingto grain size and/or shape (e.g. Bowser and Bernhard, 1993; Allan et al.,1998; Tuckwell et al., 1999; Thomsen and Rasmussen, 2008; Armynotdu Châtelet et al., 2009; Mancin et al., 2012).
Our results document that most of the studied foraminifera builtstructurally complex tests selecting grains from the substratum by com-position, size and shape. The species E. bradyi,K. bradyi, S. pseudogramen,S. rolshauseni and S. mestayerae, selected calcite coccoliths (Plate 1,images 1–5), sometimes even choosing a specific nannofossil speciesor genus. The sediment samples, picked from the core IMAGES MD97-2114, mostly consist of carbonate nannofossil and foraminiferalooze (up to 90% by volume, Lupi et al., 2008; Lupi, 2009) with a minoramount of silicate minerals (b4%) sourced from volcanic eruptions orland-derived (Orpin et al., 2008). Thus we could reasonably supposethat the foraminifera picked coccoliths because of their high abundanceand availability on the sea floor. Other authors have argued that afterblooms of coccolithophores, the bloom species become the dominantparticles available for use by agglutinated foraminifera (Henrikssonet al., 1998). It must be noted that some of the selected nannofossil spe-cies were also among the major nannofossil components of the studiedassemblages (e.g. C. pelagicus and Calcidiscus leptoporus, Lupi, 2009;Cobianchi et al., 2012). Nevertheless, the species K. bradyi (Table 1,Supplementary data), which co-occurred with E. bradyi, S. rolshauseni,S. pseudogramen and S. mestayerae in the same foraminiferal assem-blage, selected mainly Helicospharea coccoliths (Plate 1, 2a), althoughthis genus was not very abundant (average relative abundance ofca. 6%; Lupi, 2009). It appears that other nannofossil taxa, such asthe small and medium Gephyrocapsa, that are particularly abundant inthe studied core (more than 50% of the assemblage, Lupi, 2009),were generally avoided by the studied agglutinated species. Startingfrom these observations, we can prudently argue that the studiedspecies did not pick coccoliths just on the basis of their availabilityand abundance in the substratum. They seem to have preferredand generally to have selected quite robust and moderately-sized coccoliths, with smooth concave or convex surfaces andcharacterised by elliptical or discoid shapes. At the same time, smallercoccoliths with larger holes and/or with very delicate structures (likeGephyrocapsa) and/or particular morphologies (like Rhabdospaheraclavigera)were avoided.
In spite of the great abundance of coccoliths in the substratum, thestudied species picked coccoliths mostly in order to form the calcitepavement that covers the external surface of the agglutinated wallmade of mineral grains (e.g. Plate 5, 1a, 3a, 4a).
Under the carbonate test surface the other grainswere almost exclu-sively silicates even if the silicate component of the sediment bulk doesnot exceed 4% (Orpin et al., 2008). This could suggest that a test wallmostly composed of silicate grains was probably more functional (pos-sibly more resistant) than a wall exclusively composed of calcareousbiogenic remains but, at the same time, an agglutinated surface of thetest composed of calcite particles could be advantageous on a carbonatesea floor. We do not know what is the real benefit in possessing an ex-ternal pavement made of calcareous particles, but we could hazard thatin a “calcareous environment”, like the seafloor of the Chatham Rise, itcould be useful to build a calcite surface which makes the agglutinatedshell greatly resemble a calcareous-hyaline shell (e.g. Plate 1, 4a), per-haps as camouflage against predators.
Conversely, K. novangliae mostly picked silicate minerals in place ofcoccoliths to make its test surface (Plates 3, 2; Plates 6A–B), althoughit belongs to the same genus and is morphologically similar (Plate 1,images 2 and 7) and co-occurs in the same samples as K. bradyi. It isdifficult to explain this difference: possibly the two Karreriella speciesagglutinated different grains to avoid inter-species competition or prob-ably because grain selection is firstly driven by genetics, thus species-dependent. Few studies have demonstrated preferential selection forcoccoliths in natural modern environments (e.g. Young and Giesen,2002; Gooday et al., 2010) or in the fossil record (e.g. Almogi-Labinet al., 1996; Holbourn and Kaminski, 1997; Thomsen and Rasmussen,2008) and none of the test wall structures described in these papers ap-pear to be identical to thewall structure displayed by the studied agglu-tinated specimens.
A
Plate 4. Back-scattered electron (BSE) images of four sectioned specimens of Eggerella bradyi showing the test wall microstructure and the elemental characterisation of the agglutinatedgrains. Specimenswere picked, respectively under (1–2; at 10.92 and 10.67mbsf), within (3; at 10.43mbsf) and above (4; at 10.32mbsf) the tephra T3. The red crosses indicate the spotsfor standardless microanalyses; collected chemical data are in Table 2 (available online in the data repository). Note that some corresponding EDS spectra are also reported as well as theelementalmaps showing the distribution of Si, Al, Na and Ca in the sectioned test wall. In the elementalmap the colour intensity is proportional to the element concentration in every pixelof the image (in black areas the element is lacking), thus the comparison of the different maps collected in the same area of the sample provides an overview of the mineralogical distri-bution in the area. See the text for further explanations. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
48 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

B
Plate 4 (continued).
49N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

A
Plate 5. Back-scattered electron (BSE) images of four sectioned specimens of Karreriella bradyi showing the test wall microstructure and the elemental characterisation of the agglutinatedgrains. Specimens were picked, respectively under (1; at 10.04mbsf), within (2, at 9.94mbsf) and above (3–4; at 9.82 and 9.02 mbsf) the tephra T2. The red crosses indicate the spots forstandardless microanalyses; collected chemical data are in Table 2 (available online in the data repository). Some corresponding EDS spectra are also reported as well as the elementalmaps showing the distribution of Si, Al, Na and Ca in the sectioned test wall. In the elemental map the colour intensity is proportional to the element concentration in every pixel ofthe image (in black areas the element is lacking), thus the comparison of the differentmaps collected in the same area of the sample provides an overviewof themineralogical distributionin the area. See the text for further explanations. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
50 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

B
Plate 5 (continued)
51N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
Our results also document grain selection on the basis of size andshape. The largest grains were generally placed close to the externalwall edge whereas the smaller particles were arranged progressively
towards the inside in order to form a kind of layered wall structure (e.g.Plates 4–6). Thiswallmicrostructure is a recurrent feature in bothmodernand fossil foraminifera, particularly in the textulariids (e.g. Allen et al.,

A
Plate 6. Back-scattered electron (BSE) images of three sectioned specimens of Karreriella novangliae showing the test wall microstructure and the elemental characterisation of the agglu-tinated grains. Specimenswere picked, respectively under (1; at 6.97mbsf),within (2; at 6.84mbsf) and above (3; at 6.48mbsf) the tephra AT-390.Note that the specimen from the tephralayermostly selects volcanic glass fragments to build the testwall. The red crosses indicate the spots for standardlessmicroanalyses; collected chemical data are in Table 2 (available onlinein the data repository). Some corresponding EDS spectra are also reported as well as the elemental maps showing the distribution of Si, Al, Na and Ca in the sectioned test wall. In theelemental map the colour intensity is proportional to the element concentration in every pixel of the image (in black areas the element is lacking), thus the comparison of the differentmaps collected in the same area of the sample provides an overview of themineralogical distribution in the area. See the text for further explanations. (For interpretation of the referencesto colour in this figure legend, the reader is referred to the web version of this article.)
52 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

B
Plate 6 (continued)
53N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
1998; Tuckwell et al., 1999; Mancin, 2001; Kaminski, 2004; Mancin et al.,2012). Tuckwell et al. (1999) tentatively explained this grain selectionand arrangement using numerical models and hypothesised a particularfractal grain distribution within the agglutinated wall. In this model, thelayered grain arrangement could limit the need for biologically producedcementing material and increased the overall strength of the test wall.
If we assume that the prime purpose of the foraminiferal test in adeep-sea environment is protection, what would be the adaptive benefitof using different grain shapes, sizes and compositions? We could pru-dently suppose that a layered test with different grains and orientationsin different layers is probably stronger than an unlayered one, thus it ismore able to prevent a potential predator from breaking into the testby crunching it or by pulling it apart. The smoother regular outer surfaceformed by equidimensional particlesmight help in this function bymak-ing the test more resistant to being grabbed and pulled apart.
Among our studied agglutinated foraminifera, however, othertextulariid species did not sort particular grains (e.g. T. lythostrota andS. sagittula; Plate 2) and in this case it is more difficult to explain whatmight be the possible advantage of this choice. We tentatively suggestthat another form of protection against predation might be to have acamouflaged test, but the camouflage would be tactile (touch) or olfac-tory (chemosensitivity) and not visual as the deep-sea is dark. Havingthe test composed of the dominant sediment grains arranged randomlyas in the substratum could deceive a potential predator as it might notrecognise the test so readily as having food inside.
5.2. Impact of volcanic ash on agglutinated test construction
Explosive volcanic eruptions often form imposing columns of air-borne particles and ash, which then fall out over a wide area downwind

Plate 7. Back-scattered electron (BSE) images of four sectioned specimens of Siphotextularia rolshauseni showing the test wall microstructure and the elemental characterisation of theagglutinated grains. Specimens were picked, respectively under (1; at 8.70 mbsf), within (2; at 8.45 mbsf) and above (3–4; at 8.24 and 8.00 mbsf) the tephra layer AT-334. Note thatin all the collected specimens the test wall exclusively consisted of a single layer of coccoliths (Cn pelagicus) embedded into a very-fine grained calcareous matrix.
54 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

A
Plate 8. Back-scattered electron (BSE) images of three sectioned specimens of Sigmoilopsis schlumbergeri showing the test wall microstructure and the elemental characterisation of theagglutinated grains. Specimenswere picked, respectively under (1; at 6.97mbsf), within (2; at 6.84mbsf) and above (3; at 6.48mbsf) the tephra layer AT-390. The red crosses indicate thespots for standardless microanalyses; collected chemical data are in Table 2 (available online in the data repository). Some corresponding EDS spectra are also reported as well as the el-ementalmaps showing thedistribution of Si, Al, Na and Ca in the sectioned testwall. In the elementalmap the colour intensity is proportional to the element concentration in every pixel ofthe image (in black areas the element is lacking), thus the comparison of the differentmaps collected in the same area of the sample provides an overviewof themineralogical distributionin the area. See the text for further explanations. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
55N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58

Plate 8 (continued)
56 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
of the volcano. The ash fall usually causes important perturbations inboth surface and deep-sea environments (Haeckel et al., 2001; Hesset al., 2001). These perturbations can directly affect deep-sea organisms,that are buried and decimated, or induce geochemical changes in thedeep-sea environment. Only a few studies have dealt with the responseof the deep-sea environment to the massive deposition of volcanic ash,andmost primarily focused on the re-establishment of deep-sea ecosys-tems after the perturbing event (e.g. Hess and Kuhnt, 1996; Hess et al.,2001; Kuhnt et al., 2005).
We believe that agglutinated foraminifera, which use the substra-tum as the source of material for building their test, would reflect the
enrichment of the substratum with enormous quantities of volcanicglass shards.
Our results derived from the study of a limited number of mostlytextulariid species (E. bradyi, K. bradyi, K. novangliae, S. rolshauseni andS. schlumbergeri) document, however, that the composition of the agglu-tinated grains did not change when the tephra layer was deposited(Plate 4B, specimen 3; Plate 5A; 2; Plate 7, 2 and Plate 8A, 2). The resultis quite curious because abrupt changes in wall composition were com-monly observed in recent/stained agglutinated foraminiferawhichwereaffected by the 1991 Mount Pinatubo tephra in the South China Sea(Hess et al., 2001). Many of these taxa, mostly using organic cements,

57N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
are subject to bacterial decay during early diagenesis, and are thus notcommonly found in Pleistocene sedimentary records (e.g. Haywardet al., 2001). Early re-colonizers such as various Reophax species alsocommonly select the newly available volcanic material to constructtheir test (Kuhnt et al., 2005). It must be noted that the analysed speciesof the present paper are mostly represented by calcareous-cementedtextulariid specimens that were probably not among those early re-colonizers and possibly, when they re-appeared on the sea floor afterthe tephra event, may have found a substratum probably already com-posed of hemipelagic and ashmaterial, mixed by bioturbational activity.
Only the species K. novangliae significantly changed the compositionof its grains by selecting mainly volcanic glass fragments (Plate 6A,specimen 2). This observation, although based on a limited number ofanalysed species, seems to suggest the concept of genetic control ongrain selection (to the species level). Nevertheless, tephra depositionappears to have influenced the wall microstructure of agglutinatedtests. The textulariid specimens, collected from the volcanoclasticlayers, have a thinner test wall lacking the external pavement formedby coccoliths;moreover thewall is also characterised by a less abundantcalcareous matrix with respect to the specimens sampled above orbelow the tephra layer (e.g. Plate 5A, 2a). This evidence seems to sug-gest that the tephra deposition is able to induce some kind of perturba-tion of the normal agglutinating process.
Possibly the miliolid species S. schlumbergeri did not record anysignificant change within its test microstructure because it is not atrue agglutinated foraminifera, thus with a diverse genetic plain.
An alternative hypothesis to explain the loss of calcareous partsin the studied agglutinated tests from the tephra layers is that thevolcanoclastic deposition induced local carbonate dissolution. Increasedcalcite dissolution as a result of tephra deposition has been recorded bydissolution proxies from the same samples. Probably, a lowering of pHvalues at the sea-floor and in the pore water, accompanied by acidifica-tion of surface water inhibiting bio-calcification, could be responsiblefor a reduced availability of calcareous remains, mainly coccoliths, inthe sediments.
6. Conclusions
The studied agglutinated species built complex tests, picking andselecting particles on the basis of their availability and abundance inthe substratum, as well as of the grain size, shape and composition.
The composition of the agglutinated grains, however, did not changewhen tephra deposition strongly enriched the substratumwith volcanicglass shards; a result consistentwith the fact that the selection of the ag-glutinated grains is probably controlled mostly by genetics. Instead thetephra deposition influenced the microstructure of the agglutinatedwall; the textulariid specimens coming from the volcanoclastic layerswere characterised by a thinner test wall lacking the external pavementof coccoliths, andwith a less abundant calcareousmatrixwith respect tothe specimens sampled above or below the tephra layer. The tephra de-position probably disturbed the normal agglutination process possiblyfavouring the development of undersaturated conditions at the seafloor, which in turn induced calcite dissolution.
In conclusion, our results demonstrate that there are more ways ofconstructing the test wall in deep-sea agglutinated foraminifera thanhas previously been described. This ismostly a result of the handling ca-pabilities of the foraminiferal cell to sort the grains by composition, sizeand shape while only partly dependent on the sediment composition.Clearly more modern and fossil agglutinated foraminifera need to beanalysed before we may fully understand this complex phenomenon.
Acknowledgements
The authors acknowledge the staff of the CISRiC-Arvedi Laboratoryfor their technical assistance and SEM–EDS analyses. The helpfulcomments and suggestions of Prof. F. Jorissen, Dr. F. Frontalini and an
anonymous reviewer are also gratefully acknowledged. This projectwas funded by the "Fondi di Ateneo per la Ricerca- FAR 2010-2012" ofthe University of Pavia (Italy).
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.marmicro.2014.12.004.
References
Allen, K., Roberts, S., Murray, J.W., 1998. Fractal grain distribution in agglutinated forami-nifera. Paleobiology 24 (3), 349–358.
Alloway, B.V., Westgate, J.A., Pillans, B., Pearce, N., Newnham, R., Byrami, M., Aarburg, S.,2004. Stratigraphy, age and correlation of middle Pleistocene silicic tephras in theAuckland region. New Zealand: a prolific distal record of TVZ silicic volcanism. NZJ. Geol. Geophys. 24, 39–63.
Alloway, B.V., Pillans, B.J., Carter, L., Naish, T.R.,Westgate, J.A., 2005. Onshore–offshore cor-relation of Pleistocene rhyolitic eruptions from New Zealand: implications for TVZeruptive history and paleoenvironmental construction. Quat. Sci. Rev. 24, 1601–1622.
Almogi-Labin, A., Hemleben, C., Meischner, D., Erlenkeuser, H., 1996. Response of Red Seadeep-water agglutinated foraminifera to water-mass changes during Late Quaterna-ry. Mar. Micropaleontol. 28, 283–297.
Armynot du Châtelet, E., Bout-Roumazeilles, V., Coccioni, R., Frontalini, F., Guillot, F.,Kaminski, M.A., Recourt, P., Riboulleau, A., Trentesaux, A., Tribovillard, N., Ventalon,S., 2013a. Environmental control on shell structure and composition of agglutinatedforaminifera along a proximal–distal transect in the Marmara Sea. Mar. Geol. 335,114–128.
Armynot du Châtelet, E., Frontalini, F., Guillot, F., Recourt, P., Ventalon, S., 2013b. Surfaceanalysis of agglutinated benthic foraminifera through SEM–EDS and Raman analyses:an expeditious approach for tracing mineral diversity. Mar. Micropaleontol. 105,18–29.
Armynot du Châtelet, E., Recourt, P., Chopin, V., 2008. Mineralogy of agglutinated benthicforaminifera; implications for paleoenvironmental reconstructions. Bull. Soc. Geol. Fr.179 (6), 583–593.
Bender, H., Hemleben, C., 1988. Calcitic cement secreted by agglutinated foraminifersgrown in laboratory culture. J. Foraminifer. Res. 3, 60–68.
Bertram, M.A., Cowen, J.P., 1998. Biomineralization in agglutinating foraminifera: an ana-lytical SEM investigation of external wall composition in three small test forms.Aquat. Geochem. 4, 455–468.
Bowser, S.S., Bernhard, J.M., 1993. Structure, bioadhesive distribution and elastic proper-ties of the agglutinated test of Astrammina rara (Protozoa: Foraminiferida).J. Eukaryot. Microbiol. 40 (2), 121–131.
Carter, L., Nelson, C.S., Neil, H.L., Froggatt, P.C., 1995. Correlation, dispersal and preserva-tion of the Kawakawa tephra and other late Quaternary tephra layers in theSouthwest Pacific Ocean. NZ J. Geol. Geophys. 38, 29–46.
Carter, L., Alloway, B.V., Shane, P., Westgate, J.A., 2004. Late Cenozoic major rhyolitic erup-tions and dispersal-deep ocean records from off New Zealand. NZ J. Geol. Geophys. 47(3), 481–500.
Cobianchi, M., Luciani, V., Lupi, C., Mancin, N., Lirer, F., Pelosi, N., Trattenero, I., Bordiga, M.,Hall, I.R., Sprovieri, M., 2012. Pleistocene biogeochemical record in the south-westPacific Ocean (Images Site MD 97-2114, Chatham Rise). J. Quat. Sci. 27, 519–530.
Crundwell, M., Scott, G., Naish, T., Carter, L., 2008. Glacial–interglacial ocean climate vari-ability from planktonic foraminifera during the Mid-Pleistocene transition in thetemperate Southwest Pacific, ODP Site 1123. Palaeogeogr. Palaeoclimatol. Palaeoecol.260, 202–229.
du Châtelet, Armynot, Bout-Roumazeilles, V., Riboulleau, A., Trentesaux, A., 2009. Sedi-ment (grain size and clay mineralogy) and organic matter quality control on livingbenthic foraminifera. Rev. Micropaleontol. 52, 75–84.
Frogner Kockum, P.C., Herbert, R.B., Gislason, S.R., 2006. A diverse ecosystem response tovolcanic aerosols. Chem. Geol. 231, 57–66.
Frogner, P., Gislason, S.R., Oskarsson, S.R., 2001. Fertilizing potential of volcanic ash inocean surface water. Geology 29, 487–490.
Gooday, A.J., Nott, J.A., Davis, S., Mann, S., 1995. Apatite particles in the test wall of thelarge agglutinated foraminifer Bathysiphon major (Protista). J. Mar. Biol. Assoc. U. K.75, 469–481.
Gooday, A.J., Uematsu, K., Kitazato, H., Toyofuku, T., Young, J.R., 2010. Traces of dissolvedparticles, including coccoliths, in the tests of agglutinated foraminifera from theChallenger Deep (10,897 m water depth, western equatorial Pacific). Deep-Sea Res.I 57, 239–247.
Haeckel, M., van Beusekom, J., Wiesner, M.G., Konig, I., 2001. The impact of the 1991Mount Pinatubo tephra fallout on the geochemical environment of deep-sea sedi-ments in the South China. Earth Planet. Sci. Lett. 193, 151–166.
Hall, I.R., McCave, I.N., Shackleton, N.J., Graham, P.W., Harris, S.E., 2001. Intensified deepPacific inflow and ventilation in Pleistocene glacial times. Nature 412, 809–812.
Hayward, B.W., Carter, R., Grenfell, H.R., Hayward, J.J., 2001. Depth distribution of Recentdeep-sea benthic foraminifera east of New Zealand, and their potential for improvingpaleobathymetric assessments of Neogene microfaunas. N. Z. J. Geol. Geophys. 44,555–587.
Hayward, B.W., Grenfell, H.R., Sabaa, A.T., Neil, H.L., Buzas, M.A., 2010. Recent NewZealand deep-water benthic foraminifera: taxonomy, ecologic distribution, biogeog-raphy and use in paleoenvironmental assessment. GNS Monogr. 26, 363.

58 N. Mancin et al. / Marine Micropaleontology 115 (2015) 39–58
Henriksson, A.S., Widmark, J.G.V., Holbourn, A.E.L., Thies, A., Kuhnt,W., 1998. Coccoliths astest building material for foraminifera (“Coccolithofera”). J. Nannoplank. Res. 20 (1),15–19.
Heron-Allen, E., 1915. A short statement upon the theory, and phenomena of purpose andintelligence exhibited by the protozoa, as illustrated by selection and behaviour in theforaminifera. J.R. Microsc. Soc. 547–557.
Heron-Allen, E., Earland, A., 1909. On a new species of Technitella from the North Sea, withsome observation upon selective power as exercised by certain species of arenaceousforaminifera. J. Queckett Microsc. Club, Ser 210 (64), 402–412.
Hess, S., Kuhnt, W., 1996. Deep-sea benthic foraminiferal recolonization of the 1991Mt. Pinatubo ash layer in the South China Sea. Mar. Micropaleontol. 28, 171–197.
Hess, S., Kuhnt, W., Hill, S., Kaminski, M.A., Holbourn, A., de Leon,M., 2001.Monitoring therecolonization of the Mt. Pinatubo 1991 ash layer by benthic foraminifera. Mar.Micropaleontol. 43, 119–142.
Holbourn, A.E., Kaminski, M.A., 1997. Lower Cretaceous deep-water benthic foraminiferaof the Indian Ocean. Grzybowski Found. Spec. Publ. 4, 1–172.
Jones, M.T., Gislason, S.R., 2008. Rapid releases of metal salts and nutrients following thedeposition of volcanic ash into aqueous environments. Geochim. Cosmochim. Acta72, 3661–3680.
Kaminski, M.A., 2004. The year 2000 classification of agglutinated foraminifera. In: Bubik,M., Kaminski, M.A. (Eds.), Proceedings of the Sixth International Workshop on Agglu-tinated Foraminifera. Grzybowski Found., Spec. Pub. 8, pp. 237–255.
Kuhnt, W., Hess, S., Holbourn, A., Paulsen, H., Salomon, B., 2005. The impact of the 1991Mt. Pinatubo eruption on deep-sea foraminiferal communities: a model for theCretaceous/Tertiary (K/T) boundary? Palaeogeog. Palaeoclim. Palaeoecol 224,83–107.
Lisiecki, L.E., Raymo, M.E., 2005. A Plio-Pleistocene stack of 57 globally distributed benthicδ18O records. Paleoceanography 600, 20. http://dx.doi.org/10.1029/2004PA001071.
Loeblich, A.R., Tappan, H., 1989. Implications of wall composition and structure in agglu-tinated foraminifera. J. Paleontol. 63, 769–777.
Lupi, C., 2009. Biostratigraphic correlation and paleoceanographic interpretation ofPleistocene calcareous nannofossils from the Subtropical Front to the Antarctic Diver-gence. Micropaleontol. 55, 383–395.
Lupi, C., Luciani, V., Cobianchi, M., 2008. Integrated calcareous nannofossil and planktonicforaminiferal bioevents of the last 1.07 Ma: a case study from the East New ZealandPacific Ocean. Micropaleontol. 54, 463–476.
Makled, W.A., Langer, M.R., 2010. Preferential selection of titanium-bearing minerals inagglutinated Foraminifera: ilmenite (FeTiO3) in Textularia hauerii d'Orbigny fromthe Bazaruto Archipelago Mozambique. Rev. Micropaleontol. 53 (3), 163–173.
Mancin, N., 2001. Agglutinated foraminifera from the Epiligurian succession (Middle Eocene/Lower Miocene, Northern Apennines, Italy): scanning electron microscopic characteriza-tion and paleoenvironmental implications. J. Foraminifer. Res. 31 (4), 294–308.
Mancin, N., Basso, E., Pirini, C., Kaminski, M.A., 2012. Selective mineral composition, func-tional test morphology and paleoecology of the agglutinated foraminiferal genusColominella Popescu, 1998 in the Mediterranean Pliocene (Liguria, Italy). Geol.Carpath. 63 (6), 491–502.
Mancin, N., Basso, E., Kaminski, M.A., Dogan, U., 2014. A standard SEM–EDSmethodology todetermine the test microstructure of fossil agglutinated foraminifera. Micropaleontol.60 (1), 13–26.
Mancin, N., Hayward, B.W., Trattenero, I., Cobianchi, M., Lupi, C., 2013. Can the morphol-ogy of deep−sea benthic foraminifera reveal what caused their extinction during themid−Pleistocene Transition? Mar. Micropaleontol. 104, 53–70.
Orpin, A., Carter, L., Goh, A., Mackay, E., Pallentin, A., Verdier, A.L., Chiswell, S., Sutton, P.,2008. New Zealand's diverse seafloor sediments. NIWA Chart, Miscellaneous Ser. N°86.FCA.
Rothe, N., Gooday, A.J., Pearce, R.B., 2011. Intracellular mineral grains in thexenophyophore Nazareammina tenera (Rhizaria, Foraminifera) from Nazaré Canyon(Portuguese margin, NE Atlantic). Deep-Sea Res. I 58, 1189–1195.
Thomsen, E., Rasmussen, T.L., 2008. Coccolith-agglutinating foraminifera from the EarlyCretaceous and how they constructed their tests. J. Foraminifer. Res. 38 (3), 193–214.
Tuckwell, G.W., Allen, K., Roberts, S., Murray, J.W., 1999. Simple models of agglutinatedforaminifera test construction. J. Eukaryot. Microbiol. 46 (3), 248–253.
Wall−Palmer, D., Jones, M.T., Hart, M.B., Fisher, J.K., Smart, C.W., Hembury, D.J., Palmer,M.R., Fones, G.R., 2011. Explosive volcanism as cause for mass mortality of pteropods.Mar. Geol. 282, 231–239.
Wilson, C.J.N., Houghton, B.F., Kamp, P.J.J., McWilliams, M.O., 1995. An exceptionally wide-spread ignimbrite with implications for pyroclastic flow emplacement. Nature 378,605–607.
Wood, A., 1949. The structure of the wall of the test in the Foraminifera; its value in clas-sification. Q. J. Geol. Soc. Lond. 104, 229–255.
Young, J.R., Giesen, M., 2002. Xenospheres — associations of coccoliths resemblingcoccospheres. J. Nannoplank. Res. 24, 27–35.
Related Documents











