Send Orders for Reprints to [email protected] 446 Current Cancer Drug Targets, 2014, 14, 446-457 The Activation of β 1 -integrin by Type I Collagen Coupling with the Hedgehog Pathway Promotes the Epithelial-Mesenchymal Transition in Pancreatic Cancer Wanxing Duan 1,† , Jiguang Ma 2,† , Qingyong Ma 1, * , Qinhong Xu 1 , Jianjun Lei 1 , Liang Han 1 , Xuqi Li 3 , Zheng Wang 1 , Zheng Wu 1 , Shifang Lv 1 , Zhenhua Ma 1 , Mouzhu Liu 4 , Fengfei Wang 5 and Erxi Wu 5, * 1 Department of Hepatobiliary Surgery, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 2 Department of Oncology, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 3 Department of General Surgery, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 4 Department of Operating Room, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 5 Department of Pharmaceutical Sciences, North Dakota State University, Fargo, ND 58105, USA Abstract: Pancreatic ductal adenocarcinoma (PDAC) is characterized by the excessive deposition of extracellular matrix (ECM), which is thought to contribute to this tumor’s malignant behavior. However, the detailed mechanism and the contribution of excessive deposition of ECM in PDAC progression remain unclear. A better understanding of the mechanism involved in this process is essential for the design of new effective therapies. In this study, we demonstrated that pancreatic cancer cells exhibited increased proliferation and decreased apoptosis in response to type I collagen. In addition, PDAC cells exposed to type I collagen lost the expression of E-cadherin and increased expression of mesenchymal markers, including N-cadherin and vimentin. This epithelial- mesenchymal transition (EMT) was correlated with enhanced cell migration and invasiveness. Knockdown of β 1 -integrin abolished the effects induced by type I collagen, and further investigation revealed that type I collagen activates β 1 -integrin (marked by phosphorylation of β 1 integrin downstream effectors, focal adhesion kinase [FAK], AKT, and ERK) accompanied by markedly up-regulation of Gli-1, a component of the Hedgehog (HH) pathway. Knockdown of Gli-1 reversed the effects of type I collagen on PDAC invasion and EMT. These results suggest that there is cross-talk between the β 1 -integrin signaling pathway and the HH pathway in pancreatic cancer and that activation of the HH pathway plays a key role in the type I collagen-induced effects on pancreatic cancer. Keywords: Desmoplastic reaction, epithelial–mesenchymal transition, extracellular matrix, hedgehog pathway, β 1 -integrin, pancreatic cancer. INTRODUCTION Pancreatic ductal adenocarcinoma (PDAC) is one of the most lethal human cancers, with a 5-year survival rate of only approximately 6% [1]. The extremely poor clinical prognosis of PDAC is due to its early stage extensive invasion into the surrounding tissues and the formation of distant metastases, which makes disease management very difficult. The molecular mechanisms involved in the early aggressive malignant behavior of this cancer remain poorly understood. One of the most significant pathological characteristics of PDAC is the desmoplastic reaction, defined as the excessive *Address correspondence to these authors at the Department of Hepatobiliary Surgery, First Affiliated Hospital, Medical College, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, China; Tel: 0086-29-85323899; E-mail: [email protected] and Department of Pharmaceutical Sciences, North Dakota State University, Fargo, ND 58105, USA; Tel: 001-701-231-7250; Email: [email protected] † These authors contributed equally to this work. deposition of extracellular matrix (ECM) around the tumor cells [2]. Increasing evidence indicates that the desmoplastic reaction is a process of creating a tumor-supportive microenvironment for tumor growth and invasion [3]. Type I collagen is an important component of ECM, and is mainly generated by activated pancreatic stellate cells [4, 5]. When PDAC cells are seeded onto type I collagen, they exhibit reduced E-cadherin expression, increased proliferation and migration [6, 7]. Integrins are a large family of heterodimers composed of non-covalently bound α and β subunits, consisting of the receptors corresponding to various ECM components. The β 1 subunit, in particular, is found in 12 dierent integrin complexes that constitute the largest subgroup of the integrin family [8]. These integrins play critical roles in the induction and maintenance of cell dierentiation and are involved in various physiological functions through binding to collagen, fibronectin, or other ECM components. Several studies have provided a wealth of evidence that β 1 -integrin induces a malignant phenotype in PDAC cells in vitro by interacting with ECM proteins in the tumor microenvironment [6, 7]. In 1873-5576/14 $58.00+.00 © 2014 Bentham Science Publishers

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Send Orders for Reprints to [email protected] 446 Current Cancer Drug Targets, 2014, 14, 446-457
The Activation of β1-integrin by Type I Collagen Coupling with the Hedgehog Pathway Promotes the Epithelial-Mesenchymal Transition in Pancreatic Cancer
Wanxing Duan1,†, Jiguang Ma2,†, Qingyong Ma1,*, Qinhong Xu1, Jianjun Lei1, Liang Han1, Xuqi Li3, Zheng Wang1, Zheng Wu1, Shifang Lv1, Zhenhua Ma1, Mouzhu Liu4, Fengfei Wang5 and Erxi Wu5,*
1Department of Hepatobiliary Surgery, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 2Department of Oncology, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 3Department of General Surgery, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 4Department of Operating Room, First Affiliated Hospital, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, Shaanxi, China; 5Department of Pharmaceutical Sciences, North Dakota State University, Fargo, ND 58105, USA
Abstract: Pancreatic ductal adenocarcinoma (PDAC) is characterized by the excessive deposition of extracellular matrix (ECM), which is thought to contribute to this tumor’s malignant behavior. However, the detailed mechanism and the contribution of excessive deposition of ECM in PDAC progression remain unclear. A better understanding of the mechanism involved in this process is essential for the design of new effective therapies. In this study, we demonstrated that pancreatic cancer cells exhibited increased proliferation and decreased apoptosis in response to type I collagen. In addition, PDAC cells exposed to type I collagen lost the expression of E-cadherin and increased expression of mesenchymal markers, including N-cadherin and vimentin. This epithelial- mesenchymal transition (EMT) was correlated with enhanced cell migration and invasiveness. Knockdown of β1-integrin abolished the effects induced by type I collagen, and further investigation revealed that type I collagen activates β1-integrin (marked by phosphorylation of β1 integrin downstream effectors, focal adhesion kinase [FAK], AKT, and ERK) accompanied by markedly up-regulation of Gli-1, a component of the Hedgehog (HH) pathway. Knockdown of Gli-1 reversed the effects of type I collagen on PDAC invasion and EMT. These results suggest that there is cross-talk between the β1-integrin signaling pathway and the HH pathway in pancreatic cancer and that activation of the HH pathway plays a key role in the type I collagen-induced effects on pancreatic cancer.
Keywords: Desmoplastic reaction, epithelial–mesenchymal transition, extracellular matrix, hedgehog pathway, β1-integrin, pancreatic cancer.
INTRODUCTION
Pancreatic ductal adenocarcinoma (PDAC) is one of the most lethal human cancers, with a 5-year survival rate of only approximately 6% [1]. The extremely poor clinical prognosis of PDAC is due to its early stage extensive invasion into the surrounding tissues and the formation of distant metastases, which makes disease management very difficult. The molecular mechanisms involved in the early aggressive malignant behavior of this cancer remain poorly understood.
One of the most significant pathological characteristics of PDAC is the desmoplastic reaction, defined as the excessive
*Address correspondence to these authors at the Department of Hepatobiliary Surgery, First Affiliated Hospital, Medical College, Xi'an Jiaotong University, 277 West Yanta Road, Xi’an 710061, China; Tel: 0086-29-85323899; E-mail: [email protected] and Department of Pharmaceutical Sciences, North Dakota State University, Fargo, ND 58105, USA; Tel: 001-701-231-7250; Email: [email protected] †These authors contributed equally to this work.
deposition of extracellular matrix (ECM) around the tumor cells [2]. Increasing evidence indicates that the desmoplastic reaction is a process of creating a tumor-supportive microenvironment for tumor growth and invasion [3]. Type I collagen is an important component of ECM, and is mainly generated by activated pancreatic stellate cells [4, 5]. When PDAC cells are seeded onto type I collagen, they exhibit reduced E-cadherin expression, increased proliferation and migration [6, 7].
Integrins are a large family of heterodimers composed of non-covalently bound α and β subunits, consisting of the receptors corresponding to various ECM components. The β1 subunit, in particular, is found in 12 di�erent integrin complexes that constitute the largest subgroup of the integrin family [8]. These integrins play critical roles in the induction and maintenance of cell di�erentiation and are involved in various physiological functions through binding to collagen, fibronectin, or other ECM components. Several studies have provided a wealth of evidence that β1-integrin induces a malignant phenotype in PDAC cells in vitro by interacting with ECM proteins in the tumor microenvironment [6, 7]. In
1873-5576/14 $58.00+.00 © 2014 Bentham Science Publishers

The Activation of β1-Integrin Induces EMT Current Cancer Drug Targets, 2014, Vol. 14, No. 5 447
an experiment model of metastasis, the abnormal activation of β1-integrin correlated with the ability of PDAC to metastasize [9].
EMT is a critical biological process that is involved in multiple aspects of normal embryonic morphogenesis [10]. A large body of evidence indicates that tumor cells can achieve heightened invasiveness through a similar process [11]. During EMT, epithelial cells undergo a series of changes to the transition from an epithelial phenotype to a mesenchymal phenotype. These include dissolution of tight junctions, loss of cell polarity, reduction of epithelial markers (e.g., E-cadherin), and the acquisition of markers (e.g., N-cadherin and vimentin) that are associated with a mesenchymal state. Patients whose pancreatic tumors bear mesenchymal features have worse survival than those patients whose tumors lack markers of EMT [12]. Pancreatic cancer cells with a mesenchymal phenotype readily develop metastatic lesions to liver compared with those cells without EMT [13].
The HH signaling pathway, which acts as a crucial regulator of proliferation and differentiation during embryo- genesis, is aberrantly activated in multiple human solid tumors, including pancreatic cancer [14]. In the canonical sonic Hedgehog (sHH) pathway, the binding of sHH ligands to Patched (Ptch) which is a 12-pass transmembrane receptor, results in the release of Smoothened (Smo), a 7-pass transmembrane spanning protein. Activated Smo triggers an intracellular signaling cascade, resulting in the transcription of HH-specific target genes, including Gli-1 [15]. Other pathways can activate the downstream components of the HH signaling pathway in an HH-independent manner [16]. As a result, the constitutive activation of the HH pathway contributes to tumor formation and tumor progression. Our previous work demonstrated that the aberrant activation of the HH pathway in pancreatic cancer induces EMT through a non-canonical signaling pathway and thus results in enhanced invasion and metastasis [17].
In the present study, we studied the role of β1-integrin in the interaction between tumor cells and their ECM in pancreatic cancer. We found that pancreatic cancer cells exhibited increased proliferation, enhanced invasion, and decreased apoptosis as well as up-regulation of Gli-1 in response to type I collagen. These effects were abolished by β1-integrin knockdown. The ability of type I collagen to stimulate EMT was reversed by knockdown of Gli-1. Taken together, these results suggest a critical role of β1-integrin-Gli-1 pathway in mediating the ability of type I collagen to induce EMT in pancreatic cancer.
MATERIALS AND METHODS
Cell Culture and Reagents
The human pancreatic cancer cell lines ASPC-1, BxPC-3, CFPAC-1, Panc-1, and SW1990 with different metastatic and differentiation stages [18] were obtained from the Chinese Academy of Sciences Cell Bank of Type Culture Collection (CBTCCCAS) (Shanghai, China). The cells were cultured in Dulbecco's modified Eagle medium (DMEM) (HyClone, Logan, UT, USA) or RPMI 1640 medium containing 10% fetal bovine serum (FBS), 100 μg/ml ampicillin, and
100 μg/ml streptomycin. Pancreatic stellate cells (PSCs) were isolated from an adenocarcinoma (T2N0M0, moderate histological grade) found in the head of the pancreas of a 57-year-old male who underwent pancreaticoduodenectomy and were cultured as previously described [19]. The use of pancreatic cancer surgical specimens was approved by the relevant Ethical Committee of the First Affiliated Hospital of Medical College, Xi'an Jiaotong University. The cells were maintained in humidified air with 5% CO2 at 37°C. For the induction experiments, culture plates were incubated with or without 15 μg/cm2 of type I collagen overnight (Sigma-Aldrich, c3867) before the cells were seeded. Antibodies were purchased from different sources: anti-β1-integrin subunit, anti-ERK (#9102), anti-phospho-ERK (Thr202/ Tyr204, #4370), anti-AKT(#9272), and anti-phospho-AKT (S473, #4060), anti-FAK (#3285), anti-phospho-FAK (Y577, #3281) antibodies were obtained from Cell Signaling Technology; anti-sHH (sc-9024), anti-Smo (sc-166685), anti-Gli-1 (sc-20687), anti-E-cadherin (sc-7870), anti-β-actin (sc-47778), anti-uPA (sc-14019), and anti-MMP-9 (sc-13520) antibodies were obtained from Santa Cruz Biotechnology; anti-N-cadherin (#610921), and anti-vimentin (#550513) antibodies were obtained from BD Biosciences. The antibodies were stored as recommended by the manufacturer.
PROLIFERATION ASSAY
The 96-well culture plates were either uncoated or coated with 15 μg/cm2 type I collagen overnight before use and then, cells (8 × 103 cells in 200 μl) were seeded in 96- well plates. Cell proliferation was assessed at the indicated time points by the MTT (3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide) assay and the absorbance was measured at 490 nm using a multi-well microplate reader (BIO-TEC Inc, VA).
APOPTOSIS ASSAY
One milliliter of a signal cell suspension (containing 1 × 105 cells) was plated onto type I collagen-coated or uncoated 24-well plates. Following incubation at 37°C in a humidified atmosphere containing 5% CO2 for 24 hours, apoptosis was quantified by staining with annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) followed by flow cytometry analysis using a FACSCalibur (BD Bioscience, CA) instrument.
WOUND-HEALING ASSAY
The cells were grown to 80% confluence in a 6-well plate, and a wound was created using a 200-μl sterile pipette tip. The wounded area was photographed at 0 hour and 48 hour time points. The distance travelled by the cells at the migrating edge after 48 hours was calculated by measuring 3 separate points within each field of view from at least 3 separate experiments. Images were acquired by microscope (Nikon Instruments Inc.) at a magnification of 40×.
MATRIGEL INVASION ASSAY
The invasiveness of PDAC cells was examined by using a chamber-based invasion assay. In brief, the upper surface of a filter (pore size, 8.0 μm; Millipore, Billerica, MA, USA) was coated with Matrigel (BD Biosciences, Franklin

448 Current Cancer Drug Targets, 2014, Vol. 14, No. 5 Duan et al.
Lakes, NJ, USA). The cells were suspended in DMEM containing 1% FBS. Thereafter, the cell suspensions (100 μl containing 2 × 104 cells) were added to the upper chamber and concurrently 500 μl of DMEM containing 20% FBS were placed in the lower chamber. The cells were allowed to migrate for 48 hours at 37°C. The non-migratory cells were removed from the upper surface by scraping with a wet cotton swab. Invading cells on the bottom surface of the filter were fixed in methanol and stained with crystal violet. The invasive ability of cancer cells was determined by counting the stained cells under microscope (Nikon Instruments Inc.) at a magnification of 100×. The experiments were performed in triplicate.
qRT-PCR ASSAY
qRT-PCR was performed to examine the messenger RNA (mRNA) levels of β1-integrin, Gli-1, E-cadherin, N-cadherin, vimentin, snail, and GAPDH. Total RNA was prepared using TRIzol reagent (Invitrogen, CA, USA), and then reverse transcription was performed using a PrimeScript RT reagent Kit (TaKaRa, Dalian, China). The qRT-PCR experiments were conducted on an iQ5 Multicolor Real-Time PCR Detection System (Bio-Rad, Hercules, CA) using a SYBR Green Real-time PCR Master Mix (TaKaRa). The PCR primers used for each target are shown in Supplementary Table 1. The comparative CT method was used to quantitate the expression of each target gene, with GAPDH as the normalization control [20].
WESTERN BLOTTING ANALYSIS
Equivalent amounts of proteins were subjected to separation by sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis. The proteins were then transferred to Immobilon membranes (Millipore). Following incubation with the primary antibodies at 4°C overnight, the membranes were hybridized with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibody (Santa Cruz Bio- technology) for 1 hour at room temperature. The immuno- reactive bands were visualized using an enhanced chemiluminescence kit (Millipore).
IMMUNOFLUORESCENCE
Glass coverslips placed (2 × 105 cells/ml) in a 6-well plate were left untreated or coated overnight with type I collagen prior to cell seeding. After 48 hours, the cells were fixed with 4% paraformaldehyde in PBS for 20 minutes at room temperature and the endogenous peroxidase activity was quenched with 3% hydrogen peroxide. The cells were then permeabilized with 0.2% Triton X-100 plus 1% normal goat serum (NGS) in PBS for 20 minutes on ice. The coverslips were incubated with primary antibodies at 1:100 dilutions overnight at 4°C in a humidified chamber. Following incubation, the cells were rinsed with PBS 3 times, followed by incubation with fluorescein-conjugated secondary antibodies at 1:100 dilutions for 2 hours. The cells were then rinsed 4 times with PBS, mounted with medium containing 4’,6-diamidino-2-phenylindole (DAPI, Vector Laboratories), and imaged using a fluorescence microscope (Nikon Instruments Inc.) at a magnification of 200×. The experiments were performed in triplicate.
siRNA TRANSFECTION
The cells were seeded in 6-well plates at 50% to 70% confluence and then transfected with 100 nM siRNA using RNAi MAX Reagent (Invitrogen, CA, USA) in accordance to the manufacturer’s instructions. Fresh media were added 12 hours after the beginning of the transfection. The cells were used for further experiments at 24 hours after the transfection. The sequences of the control siRNA, anti-β1-integrin siRNA, and anti-Gli-1 siRNA are shown in Supplementary Table 2. The siRNAs were purchased from GenePharm (Shanghai, China).
STATISTICAL ANALYSES
The data are presented as the mean ± standard deviation. Significant differences between groups in the assays were calculated using one-way ANOVA, with the LSD post hoc test for multiple comparisons, using SPSS version 13.0 software. Significance was defined as p < 0.05.
RESULTS
Expression of the β1-integrin Subunit in Pancreatic Cancer Cells
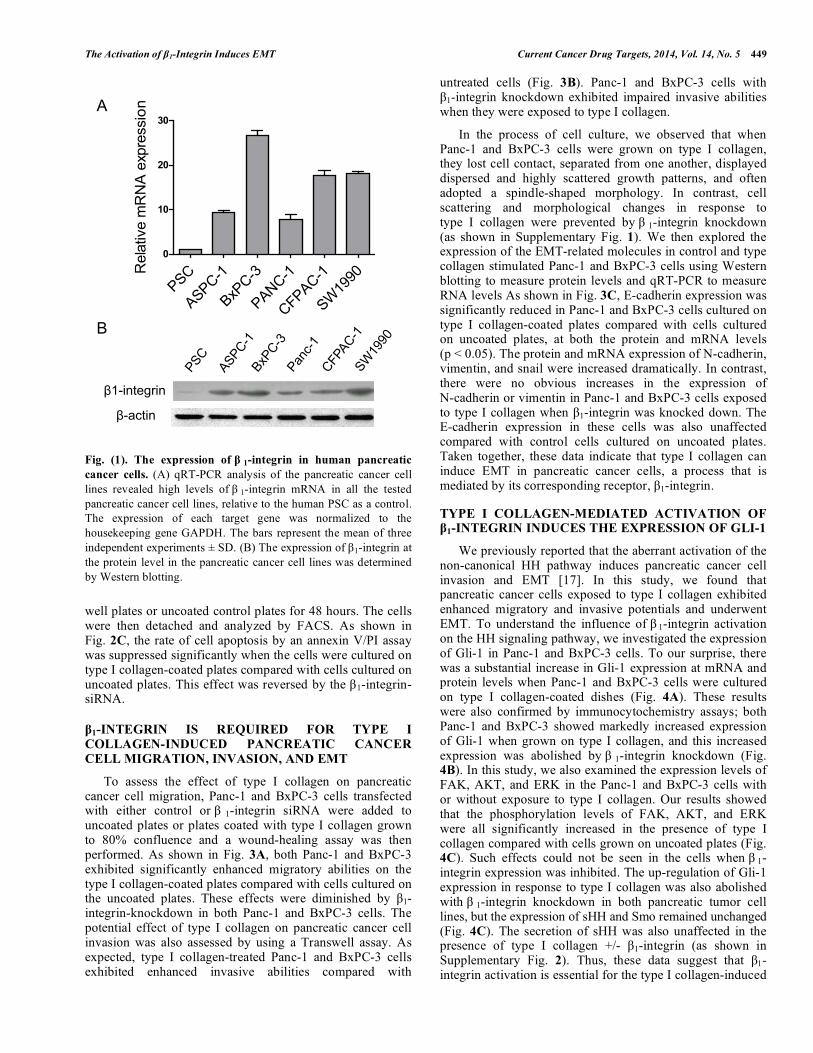
To assess the expression levels of β1-integrin subunit in pancreatic cancer cells, five pancreatic cancer cell lines, including ASPC-1, BxPC-3, CFPAC-1, Panc-1, and SW1990 (human PSC was also used as a control), were examined for β1-integrin subunit expression with qRT-PCR and immunoblotting. As shown in Fig. 1, all tested pancreatic cancer cell lines expressed high levels of β1-integrin subunit at both the mRNA (Fig. 1A) and the protein levels (Fig. 1B) relative to PSC. Among the five tested pancreatic cancer cell lines, BxPC-3 exhibited the highest level of β1-integrin expression, whereas Panc-1 and ASPC-1 showed the lowest level of tumor cell β1-integrin expression. Therefore, Panc-1 and BxPC-3 were employed for further investigation in this study.
β1-INTEGRIN IS INVOLVED IN TYPE I COLLAGEN-INDUCED EFFECTS ON PANCREATIC CANCER CELL PROLIFERATION AND APOPTOSIS
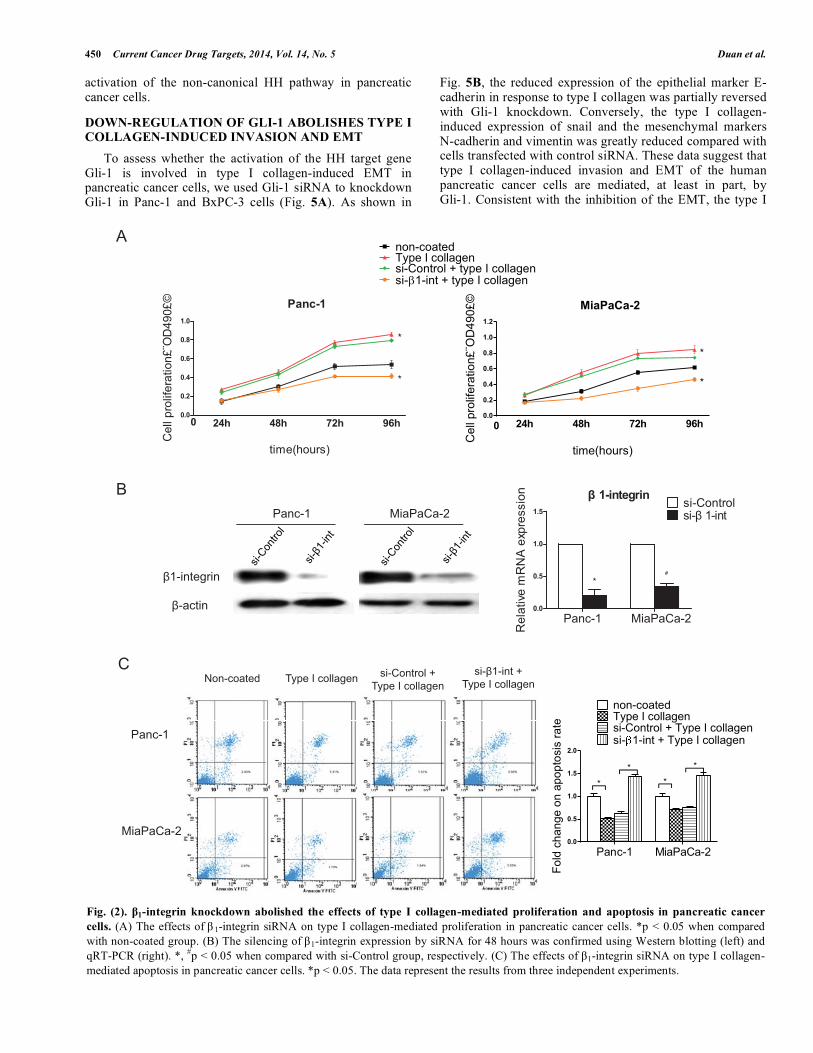
Next, we examined the effects of type I collagen on the biological behavior of Panc-1 and BxPC-3 pancreatic cancer cells. As shown in Fig. 2A, when the pancreatic cancer cells Panc-1 and BxPC-3 were seeded on type I collagen-coated plates, they exhibited enhanced cell proliferation rates. To study whether the type I collagen-mediated promotion of cell proliferation is dependent on β1-integrin, a specific siRNA targeting β1-integrin was used to knockdown β1-integrin in Panc-1 and BxPC-3 cells (Fig. 2B). The knockdown of β1-integrin in Panc-1 and BxPC-3 cells delayed the time required to reach confluence when equal number of cells was seeded on type I collagen-coated plates, as compared with control cells under the same culture conditions. Taken together, our data indicated that type I collagen plays a role in the promotion of pancreatic cancer cell proliferation and this effect is mediated by β1-integrin.
To determine the effect of type I collagen on pancreatic cancer cell apoptosis, cells transfected with either control or β1-integrin siRNA were cultured on type I collagen-coated 6-
The Activation of β1-Integrin Induces EMT Current Cancer Drug Targets, 2014, Vol. 14, No. 5 449
well plates or uncoated control plates for 48 hours. The cells were then detached and analyzed by FACS. As shown in Fig. 2C, the rate of cell apoptosis by an annexin V/PI assay was suppressed significantly when the cells were cultured on type I collagen-coated plates compared with cells cultured on uncoated plates. This effect was reversed by the β1-integrin-siRNA.
β1-INTEGRIN IS REQUIRED FOR TYPE I COLLAGEN-INDUCED PANCREATIC CANCER CELL MIGRATION, INVASION, AND EMT
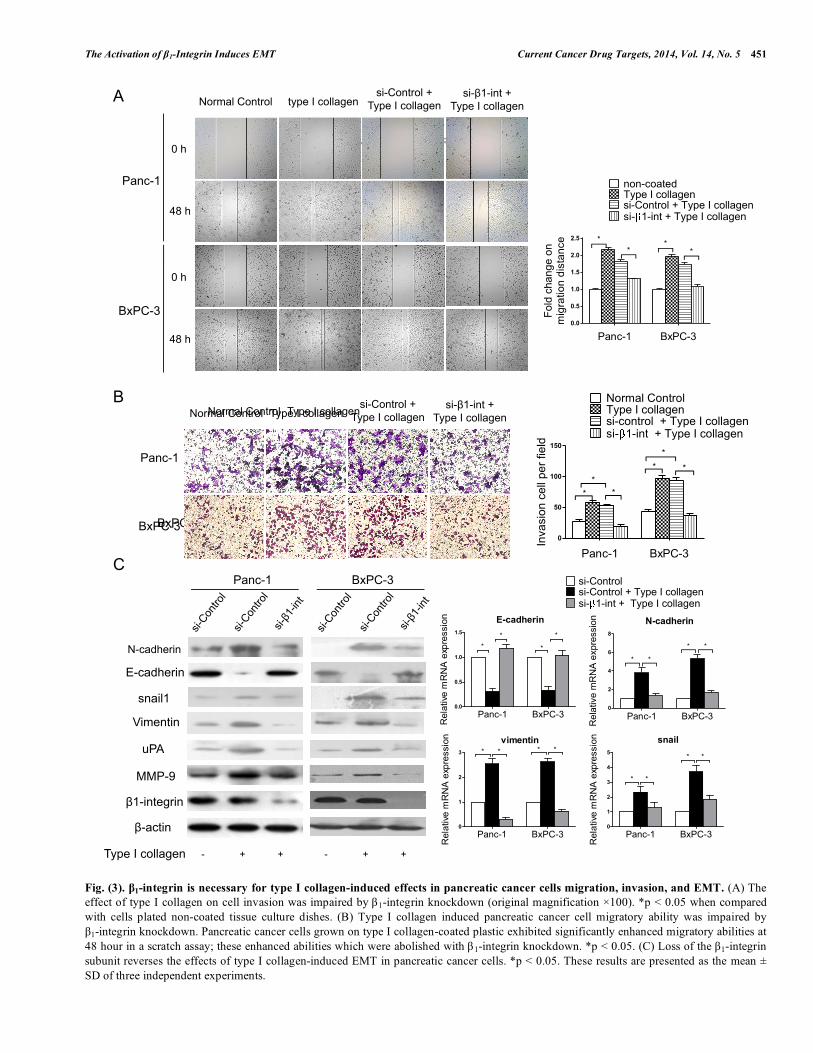
To assess the effect of type I collagen on pancreatic cancer cell migration, Panc-1 and BxPC-3 cells transfected with either control or β 1-integrin siRNA were added to uncoated plates or plates coated with type I collagen grown to 80% confluence and a wound-healing assay was then performed. As shown in Fig. 3A, both Panc-1 and BxPC-3 exhibited significantly enhanced migratory abilities on the type I collagen-coated plates compared with cells cultured on the uncoated plates. These effects were diminished by β1-integrin-knockdown in both Panc-1 and BxPC-3 cells. The potential effect of type I collagen on pancreatic cancer cell invasion was also assessed by using a Transwell assay. As expected, type I collagen-treated Panc-1 and BxPC-3 cells exhibited enhanced invasive abilities compared with
untreated cells (Fig. 3B). Panc-1 and BxPC-3 cells with β1-integrin knockdown exhibited impaired invasive abilities when they were exposed to type I collagen.
In the process of cell culture, we observed that when Panc-1 and BxPC-3 cells were grown on type I collagen, they lost cell contact, separated from one another, displayed dispersed and highly scattered growth patterns, and often adopted a spindle-shaped morphology. In contrast, cell scattering and morphological changes in response to type I collagen were prevented by β 1-integrin knockdown (as shown in Supplementary Fig. 1). We then explored the expression of the EMT-related molecules in control and type collagen stimulated Panc-1 and BxPC-3 cells using Western blotting to measure protein levels and qRT-PCR to measure RNA levels As shown in Fig. 3C, E-cadherin expression was significantly reduced in Panc-1 and BxPC-3 cells cultured on type I collagen-coated plates compared with cells cultured on uncoated plates, at both the protein and mRNA levels (p < 0.05). The protein and mRNA expression of N-cadherin, vimentin, and snail were increased dramatically. In contrast, there were no obvious increases in the expression of N-cadherin or vimentin in Panc-1 and BxPC-3 cells exposed to type I collagen when β1-integrin was knocked down. The E-cadherin expression in these cells was also unaffected compared with control cells cultured on uncoated plates. Taken together, these data indicate that type I collagen can induce EMT in pancreatic cancer cells, a process that is mediated by its corresponding receptor, β1-integrin.
TYPE I COLLAGEN-MEDIATED ACTIVATION OF β1-INTEGRIN INDUCES THE EXPRESSION OF GLI-1
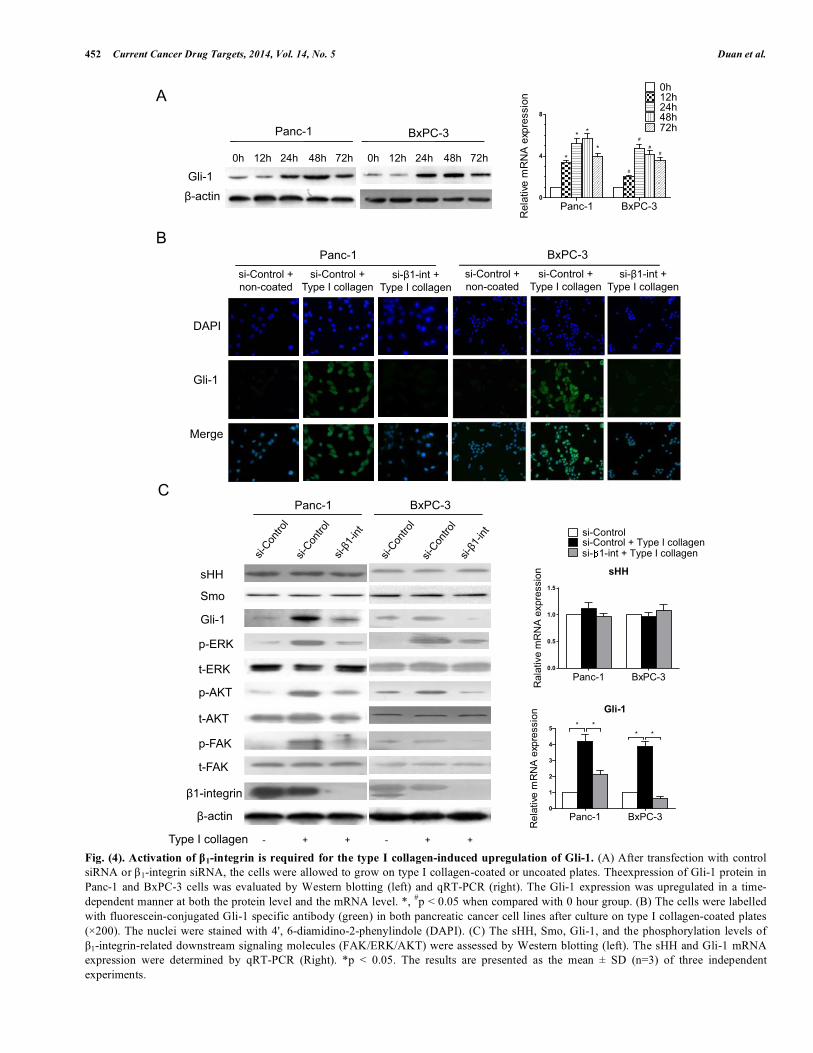
We previously reported that the aberrant activation of the non-canonical HH pathway induces pancreatic cancer cell invasion and EMT [17]. In this study, we found that pancreatic cancer cells exposed to type I collagen exhibited enhanced migratory and invasive potentials and underwent EMT. To understand the influence of β 1-integrin activation on the HH signaling pathway, we investigated the expression of Gli-1 in Panc-1 and BxPC-3 cells. To our surprise, there was a substantial increase in Gli-1 expression at mRNA and protein levels when Panc-1 and BxPC-3 cells were cultured on type I collagen-coated dishes (Fig. 4A). These results were also confirmed by immunocytochemistry assays; both Panc-1 and BxPC-3 showed markedly increased expression of Gli-1 when grown on type I collagen, and this increased expression was abolished by β 1-integrin knockdown (Fig. 4B). In this study, we also examined the expression levels of FAK, AKT, and ERK in the Panc-1 and BxPC-3 cells with or without exposure to type I collagen. Our results showed that the phosphorylation levels of FAK, AKT, and ERK were all significantly increased in the presence of type I collagen compared with cells grown on uncoated plates (Fig. 4C). Such effects could not be seen in the cells when β 1-integrin expression was inhibited. The up-regulation of Gli-1 expression in response to type I collagen was also abolished with β 1-integrin knockdown in both pancreatic tumor cell lines, but the expression of sHH and Smo remained unchanged (Fig. 4C). The secretion of sHH was also unaffected in the presence of type I collagen +/- β1-integrin (as shown in Supplementary Fig. 2). Thus, these data suggest that β1-integrin activation is essential for the type I collagen-induced
Fig. (1). The expression of β 1-integrin in human pancreatic cancer cells. (A) qRT-PCR analysis of the pancreatic cancer cell lines revealed high levels of β 1-integrin mRNA in all the tested pancreatic cancer cell lines, relative to the human PSC as a control. The expression of each target gene was normalized to the housekeeping gene GAPDH. The bars represent the mean of three independent experiments ± SD. (B) The expression of β1-integrin at the protein level in the pancreatic cancer cell lines was determined by Western blotting.
PSC
ASPC-1
BxPC-3
PANC-1
CFPAC-1
SW19
900
10
20
30
Rel
ativ
e m
RN
A e
xpre
ssio
n
�1-integrin
�-actin
A
B
450 Current Cancer Drug Targets, 2014, Vol. 14, No. 5 Duan et al.
activation of the non-canonical HH pathway in pancreatic cancer cells.
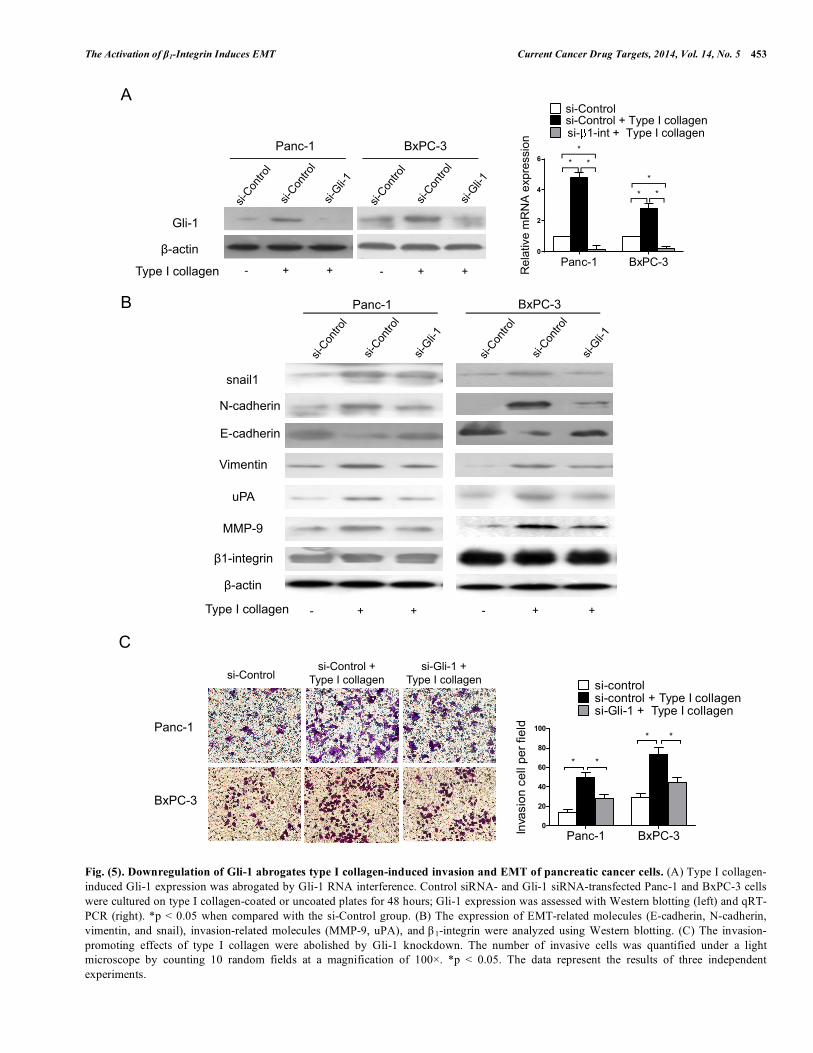
DOWN-REGULATION OF GLI-1 ABOLISHES TYPE I COLLAGEN-INDUCED INVASION AND EMT
To assess whether the activation of the HH target gene Gli-1 is involved in type I collagen-induced EMT in pancreatic cancer cells, we used Gli-1 siRNA to knockdown Gli-1 in Panc-1 and BxPC-3 cells (Fig. 5A). As shown in
Fig. 5B, the reduced expression of the epithelial marker E-cadherin in response to type I collagen was partially reversed with Gli-1 knockdown. Conversely, the type I collagen-induced expression of snail and the mesenchymal markers N-cadherin and vimentin was greatly reduced compared with cells transfected with control siRNA. These data suggest that type I collagen-induced invasion and EMT of the human pancreatic cancer cells are mediated, at least in part, by Gli-1. Consistent with the inhibition of the EMT, the type I
Fig. (2). β1-integrin knockdown abolished the effects of type I collagen-mediated proliferation and apoptosis in pancreatic cancer cells. (A) The effects of β 1-integrin siRNA on type I collagen-mediated proliferation in pancreatic cancer cells. *p < 0.05 when compared with non-coated group. (B) The silencing of β1-integrin expression by siRNA for 48 hours was confirmed using Western blotting (left) and qRT-PCR (right). *, #p < 0.05 when compared with si-Control group, respectively. (C) The effects of β1-integrin siRNA on type I collagen-mediated apoptosis in pancreatic cancer cells. *p < 0.05. The data represent the results from three independent experiments.
A
C
B
Non-coated si-Control + Type I collagen
Type I collagen si-�1-int +
Type I collagen
Panc-1
MiaPaCa-2
Panc-1 MiaPaCa-2
�1-integrin
�-actin
� 1-integrin
Panc-1 MiaPaCa-20.0
0.5
1.0
1.5si-Controlsi-� 1-int
*#
Rel
ativ
e m
RN
A e
xpre
ssio
n
00.0
0.2
0.4
0.6
0.8
1.0
24h 48h 72h 96h
*
*
Panc-1
time(hours)
Cel
l pro
lifer
atio
n£¨O
D49
0£©
The Activation of β1-Integrin Induces EMT Current Cancer Drug Targets, 2014, Vol. 14, No. 5 451
Fig. (3). β1-integrin is necessary for type I collagen-induced effects in pancreatic cancer cells migration, invasion, and EMT. (A) The effect of type I collagen on cell invasion was impaired by β1-integrin knockdown (original magnification ×100). *p < 0.05 when compared with cells plated non-coated tissue culture dishes. (B) Type I collagen induced pancreatic cancer cell migratory ability was impaired by β1-integrin knockdown. Pancreatic cancer cells grown on type I collagen-coated plastic exhibited significantly enhanced migratory abilities at 48 hour in a scratch assay; these enhanced abilities which were abolished with β1-integrin knockdown. *p < 0.05. (C) Loss of the β1-integrin subunit reverses the effects of type I collagen-induced EMT in pancreatic cancer cells. *p < 0.05. These results are presented as the mean ± SD of three independent experiments.
Normal Control Type I collagen
BxPC-3
Normal Control Type I collagen
BxPC-3
Panc-1
A
0 h
48 h
Non-coated si-Control +
type I collagen type I collagen
si-β1-int + type I collagen
BxPC-3
Panc-1
B
C
β-actin
β1-integrin
snail1
MMP-9
uPA
E-cadherin
Vimentin
N-cadherin
Type I collagen
E-cadherin
Panc-1 BxPC-30.0
0.5
1.0
1.5
*
* *
*
Re
lativ
e m
RN
A e
xpre
ssio
n
vimentin
Panc-1 BxPC-30
1
2
3 * * * *
Rel
ativ
e m
RN
A e
xpre
ssio
n snail
Panc-1 BxPC-30
1
2
3
4
5
* *
* *
Re
lativ
e m
RN
A e
xpre
ssio
n
0 h
48 h
BxPC-3
- + +
Panc-1
- + +
si-Control + Type I collagen
si-β1-int + Type I collagen
Normal Control type I collagen si-Control +
Type I collagen si-β1-int +
Type I collagen
452 Current Cancer Drug Targets, 2014, Vol. 14, No. 5 Duan et al.
Fig. (4). Activation of β1-integrin is required for the type I collagen-induced upregulation of Gli-1. (A) After transfection with control siRNA or β1-integrin siRNA, the cells were allowed to grow on type I collagen-coated or uncoated plates. Theexpression of Gli-1 protein in Panc-1 and BxPC-3 cells was evaluated by Western blotting (left) and qRT-PCR (right). The Gli-1 expression was upregulated in a time-dependent manner at both the protein level and the mRNA level. *, #p < 0.05 when compared with 0 hour group. (B) The cells were labelled with fluorescein-conjugated Gli-1 specific antibody (green) in both pancreatic cancer cell lines after culture on type I collagen-coated plates (×200). The nuclei were stained with 4', 6-diamidino-2-phenylindole (DAPI). (C) The sHH, Smo, Gli-1, and the phosphorylation levels of β1-integrin-related downstream signaling molecules (FAK/ERK/AKT) were assessed by Western blotting (left). The sHH and Gli-1 mRNA expression were determined by qRT-PCR (Right). *p < 0.05. The results are presented as the mean ± SD (n=3) of three independent experiments.
A
B
BxPC-3 Panc-1
Gli-1 �-actin
0h 12h 24h 48h 72h
C
�-actin
�1-integrin
Type I collagen
t-AKT
t-ERK
p-ERK
p-AKT
Gli-1
t-FAK
p-FAK
- + +
BxPC-3 Panc-1
sHH
Panc-1 BxPC-30
4
8
0h12h24h48h72h
#
** *
*
#
##
Rel
ativ
e m
RN
A e
xpre
ssio
n
Gli-1
Panc-1 BxPC-30
1
2
3
4
5 * ***
Rel
ativ
e m
RN
A e
xpre
ssio
n
Smo
0h 12h 24h 48h 72h
DAPI
Gli-1
Merge
Panc-1 si-�1-int +
Type I collagen si-Control +
Type I collagen si-Control + non-coated
BxPC-3 si-�1-int +
Type I collagen si-Control +
Type I collagen si-Control + non-coated
- + +
The Activation of β1-Integrin Induces EMT Current Cancer Drug Targets, 2014, Vol. 14, No. 5 453
Fig. (5). Downregulation of Gli-1 abrogates type I collagen-induced invasion and EMT of pancreatic cancer cells. (A) Type I collagen-induced Gli-1 expression was abrogated by Gli-1 RNA interference. Control siRNA- and Gli-1 siRNA-transfected Panc-1 and BxPC-3 cells were cultured on type I collagen-coated or uncoated plates for 48 hours; Gli-1 expression was assessed with Western blotting (left) and qRT-PCR (right). *p < 0.05 when compared with the si-Control group. (B) The expression of EMT-related molecules (E-cadherin, N-cadherin, vimentin, and snail), invasion-related molecules (MMP-9, uPA), and β 1-integrin were analyzed using Western blotting. (C) The invasion-promoting effects of type I collagen were abolished by Gli-1 knockdown. The number of invasive cells was quantified under a light microscope by counting 10 random fields at a magnification of 100×. *p < 0.05. The data represent the results of three independent experiments.
si-Control + Type I collagen
si-Gli-1 + Type I collagen si-Control
BxPC-3
Panc-1
A
B
β-actin
BxPC-3 Panc-1
β1-integrin
snail1
MMP-9
uPA
E-cadherin
Vimentin
N-cadherin
Type I collagen
- + +
BxPC-3 Panc-1
- + +
β-actin
Gli-1
C
Panc-1 BxPC-30
20
40
60
80
100
si-controlsi-control + Type I collagensi-Gli-1 + Type I collagen
* *
**
Inva
sio
n c
ell
pe
r fie
ld
- + +
Type I collagen
- + +
- + +

454 Current Cancer Drug Targets, 2014, Vol. 14, No. 5 Duan et al. collagen-induced up-regulation of the invasion-relevant indicators MMP-9 and uPA was partially abolished by Gli-1 knockdown in both the Panc-1 and BxPC-3 cells (Fig. 5B). A cell invasion assay revealed that the enhanced invasiveness of pancreatic cancer in the presence of type I collagen was significantly abolished by Gli-1 knockdown and the number of migratory cells was reduced by 50% after Gli-1 knockdown compared with control siRNA-treated cells (Fig. 5C). These data indicate that the activation of the HH pathway plays a pivotal role in type I collagen-induced pancreatic cancer cell invasion and EMT.
DISCUSSION In this study, we detected higher expression of β1-integrin
in six pancreatic cancer cell lines, relative to non-malignant cells. BxPC-3 cells exhibited the highest expression and Panc-1 cells had the lowest expression, compared with human PSC. Growth of the human pancreatic cancer cell lines Panc-1 and BxPC-3 on type I collagen resulted in a more malignant phenotype, characterized by enhanced proliferation, migration and invasion, and a depressed degree of apoptosis. These results suggest that both cell lines gained an EMT phenotype in response to the exposure to type I collagen. We demonstrated that the changes of phenotype were accompanied with activation of β1-integrin, the downstream of sHH signaling, ERK and AKT signaling. We found that the elevated expression of Gli-1, a downstream target of the sHH pathway, is partially responsible for type I collagen-induced EMT of pancreatic cancer cells. We further revealed that the activation of β1-integrin is critical for type I collagen induced cancer cells to become a more malignant phenotype through a non-classical sHH signaling pathway.
PDAC is characterized by its dense desmoplastic stroma, which is formed through the increased deposition of various collagens and other ECM proteins that are synthesized and secreted mainly by pancreatic stellate cells [21]. The increased expression of type I collagen, the major ECM component in the desmoplastic reaction, is detected in the PDAC stroma [22]. This remodeled ECM interacts with these tumor cells, promoting the malignant behavior of the tumor cells [7, 23-26]. The α2β1-integrin has been demonstrated to be the dominant receptor responsible for cell binding to type I collagen. The knockdown of the β1-integrin subunit but not of the α2-integrin subunit significantly inhibits pancreatic cancer progression and metastasis in vivo, suggesting a more prominent role for the β1-integrin subunit in the interactions between tumor cells and type I collagen.
Goel et al. [27] showed that β1-integrin downregulation reduces activation of AKT as well as expression of the sonic hedgehog pathway effector protein, Gli-1, resulting in inhibition of cell proliferation in human prostate cancer cells. Our study suggests that type I collagen mediates β1-integrin activation induced EMT in pancreatic cancer through up-regulation of Gli-1 in pancreatic cancer cells. Therefore, there is a close link between β1-integrin signaling and HH Pathway in cancer.
There is a wealth of evidence supporting the paracrine role of HH signaling in which HH produced by the pancreatic cancer cells acts in the adjacent stroma and
contributes to the desmoplastic reaction in PDAC [28, 29]. We found that type I collagen from stroma cells could induce a more malignant phenotype in cancer cells through a non-classical sHH signaling pathway. This raises a concern about the efficacy for cancer therapy of only blocking the classical paracrine sHH pathway. New drug combinations to block the downstream pathway involved in both classical and non-classical sHH signaling might prove more effective than current approaches [30].
FAK, ERK, and AKT mediate signaling pathways thought to be involved in the ECM-integrin related behaviors of cells and to participate in the regulation of cell proliferation, survival, and differentiation. In our study, the phosphorylation level of FAK was increased when Panc-1 and BxPC-3 were grown on type I collagen-coated dishes. The activation levels of ERK and AKT were also elevated; while specific siRNA targeting β1-integrin significantly inhibited the phosphorylation levels of FAK, ERK, and AKT. These findings are consistent with those of previous studies, suggesting that the type I collagen-induced activation of FAK, ERK, and AKT is dependent on integrin [31, 32]. In addition to the activation of these downstream effectors of β1-integrin signaling, we also observed a dramatic increase in Gli-1 expression. Several lines of evidence suggest that HH signaling could be regulated by additional parallel intracellular signaling pathways, such as RAS/MEK and PI3K/AKT [16]. The endogenous MEK and AKT signaling cascade plays a positive regulatory role in Gli-1 transcriptional activity in melanoma [33]. In our study, the type I collagen-induced up-regulation of Gli-1 was mediated by β1-integrin. The potential involvement of ERK and AKT signaling in mediating the link between β1-integrin activation and Gli-1 up-regulation requires further investigation.
The activation of β1-integrin-PI3K-Rac1-JNK signaling by type I collagen can effectively induce E-cadherin/N-cadherin switching, which is thought to be an important hallmark of EMT, leading to a more motile invasive phenotype of mouse mammary epithelial cells [34]. Furthermore, numerous studies demonstrated that, in response to type I collagen, pancreatic cancer cells display a scattered phenotype, becoming highly motile and undergoing a cadherin switch, in which decreased E-cadherin and increased N-cadherin expression was detected [6, 7, 23, 35]. In our studies, Gli-1 was also involved in the type I collagen-induced phenotypic reversion of PDAC cells (as shown in Fig. 6). Based on these observations, the activation of multiple signaling pathways may potentially contribute to the effects induced by type I collagen.
The aberrant activation of HH pathways has been implicated in pancreatic cancer, and is thought to contribute to lymph node and distant metastases [36]. EMT is increasingly acknowledged as an important process contributing to rapid tumor progression [37]. Through EMT, the cancer cells acquire enhanced motility and invasive features. In this study, we found that the elevated expression of Gli-1 is partially responsible for type I collagen-induced EMT of pancreatic cancer cells. Both the ERK and AKT signaling were activated during collagen treatment, and Gli-1 siRNA was used to rule out the possibility that type I collagen-induced EMT changes are mediated by ERK and AKT

The Activation of β1-Integrin Induces EMT Current Cancer Drug Targets, 2014, Vol. 14, No. 5 455
signaling. The knockdown of Gli-1 prevented type I collagen-induced pancreatic cancer cell EMT. Our recent published data also indicate an important role for Gli-1 in hypoxia-induced EMT changes in pancreatic cancer cells, since the down-regulation of Gli-1 effectively abrogated hypoxia-induced EMT [38]. This finding is in agreement with the evidence that the transcriptional silencing of Gli-1 compromises the ability of cancer cells to undergo migration and invasion in vitro and impairs their ability to metastasize in nude mice [39]. Moreover, the over-expression of Gli-1 promotes MUC5AC expression, leading to decreased E-cadherin-dependent cell-cell adhesion and facilitating the migration and invasion of pancreatic cancer cells [40]. However, a previous work by Joost and colleagues found that the RNAi-mediated knockdown of Gli-1 resulted in repressive effects on E-cadherin expression [41]. In contrast, the over-expression of Gli-1 increased E-cadherin expression. This group cited differing cell densities as the main explanation for these discrepancies [42]. The inconsistent effect of the up-regulation of Gli-1 on EMT in PDAC may also be explained by the presence or intermediates whose expression or activation might be induced by the stimulation of other intracellular signaling pathways, including the AKT and ERK signaling cascades. Thus, further study is required to clarify the inner link between Gli-1 and EMT.
CONCLUSIONS
The desmoplastic reaction that surrounds the malignant epithelial cells is one of the main histologically characters of PDAC, and has been closely linked to the enhanced aggressiveness nature of this disease. Our data suggest that
the aberrant activation of the HH pathway plays an important role in type I collagen-induced EMT in pancreatic cancer. In the process of type I collagen-induced EMT, the cross-talk between the β1-integrin signaling pathway and the Hedgehog (HH) pathway in pancreatic cancer is especially important. This study provides a further understanding the mechanisms involved whereby the remodeled tumor stroma promotes tumor growth and invasion in PDAC.
CONFLICT OF INTEREST
The authors declare that they have no competing interests.
ACKNOWLEDGEMENTS
This study was supported by grants from the National Natural Scientific Foundation of China (No. 81172360 to QM, No. 81201824 to XL), and a pilot project grant (EW) from a Program Project Grant from the National Center for Research Resources (NCRR; P20 RR020151) and the National Institute of General Medical Sciences (NIGMS; P20 GM103505) from the National Institutes of Health (NIH). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The authors would like to extend their thanks to Dr. Richard M. Niles (Marshall University) and Ronald E. Vincent (BIOCON Scientific) for their thoughtful reading.
SUPPLEMENTARY MATERIAL
Supplementary material is available on the publisher’s web site along with the published article.
Fig. (6). Schematic diagram of the relationship among the β1-integrin, type I collagen in the Hedgehog pathway and the EMT. The type I collagen induced β1-integrin activation could modulate the non-canonical HH pathway by increasing the transcription of Gli-1 without the alteration of sHH and Smo. Accumulated Gli-1 translocates to nuclear, results in activation of EMT-related genes, leading to the EMT switch of cancer cells. In addition, the activation of β1-integrin may also trigger other pathways (e.g., ERK, AKT) involved in EMT.

456 Current Cancer Drug Targets, 2014, Vol. 14, No. 5 Duan et al.
LIST OF ABBREVIATIONS
ECM = extracellular matrix
EMT = epithelial-mesenchymal transition
Gli-1 = glioma-associated oncogene family zinc finger 1
PDAC = pancreatic ductal adenocarcinoma
Ptch = Patched
PSCs = pancreatic stellate cells
sHH = sonic Hedgehog
Smo = Smoothened
REFERENCES [1] Siegel R.; Naishadham D.; Jemal, A. Cancer statistics, 2013. CA
Cancer J Clin. 2013, 63(1): 11-30. [2] Shields, M. A.; Dangi-Garimella, S.; Redig, A. J.; Munshi, H. G.
Biochemical role of the collagen-rich tumour microenvironment in pancreatic cancer progression. Biochem. J. 2012, 441(2), 541-552.
[3] Shields, M. A.; Dangi-Garimella, S.; Krantz, S. B.; Bentrem, D. J.; Munshi, H. G. Pancreatic cancer cells respond to type I collagen by inducing snail expression to promote membrane type 1 matrix metalloproteinase-dependent collagen invasion. J. Biol. Chem. 2011, 286(12), 10495-10504.
[4] Bachem, M. G.; Schunemann, M.; Ramadani, M.; Siech, M.; Beger, H.; Buck, A.; Zhou, S.; Schmid-Kotsas, A.; Adler, G. Pancreatic carcinoma cells induce fibrosis by stimulating proliferation and matrix synthesis of stellate cells. Gastroenterology 2005. 128(4), 907-921.
[5] Masamune, A.; Kikuta, K.; Watanabe, T.; Satoh, K.; Hirota, M.; Hamada, S.; Shimosegawa, T. Fibrinogen induces cytokine and collagen production in pancreatic stellate cells. Gut 2009, 58(4), 550-559.
[6] Imamichi, Y.; Konig, A.; Gress, T.; Menke, A. Collagen type I-induced Smad-interacting protein 1 expression downregulates E-cadherin in pancreatic cancer. Oncogene 2007, 26(16), 2381-2385.
[7] Koenig, A.; Mueller, C.; Hasel, C.; Adler, G.; Menke, A. Collagen type I induces disruption of E-cadherin-mediated cell-cell contacts and promotes proliferation of pancreatic carcinoma cells. Cancer Res. 2006, 66(9), 4662-4671.
[8] Brakebusch, C.; Fassler, R. beta 1 integrin function in vivo: adhesion, migration and more. Cancer Metastasis Rev. 2005, 24(3), 403-411.
[9] Shintani, Y.; Hollingsworth, M. A.; Wheelock, M. J.; Johnson, K. R. Collagen I promotes metastasis in pancreatic cancer by activating c-Jun NH(2)-terminal kinase 1 and up-regulating N-cadherin expression. Cancer Res. 2006, 66(24), 11745-53.
[10] Thiery, J. P.; Acloque, H.; Huang, R. Y.; Nieto, M. A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139(5), 871-890.
[11] Yilmaz, M.; Christofori, G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009, 28(1-2), 15-33.
[12] Rasheed, Z. A.; Yang, J.; Wang, Q.; Kowalski, J.; Freed, I.; Murter, C.; Hong, S. M.; Koorstra, J. B.; Rajeshkumar, N. V.; He, X.; Goggins, M.; Iacobuzio-Donahue, C.; Berman, D. M.; Laheru, D.; Jimeno, A.; Hidalgo, M.; Maitra, A.; Matsui, W. Prognostic significance of tumorigenic cells with mesenchymal features in pancreatic adenocarcinoma. J. Natl. Cancer Inst. 2010, 102(5): 340-51.
[13] Rhim, A. D.; Mirek, E. T.; Aiello, N. M.; Maitra, A, Bailey, J. M.; McAllister, F.; Reichert, M.; Beatty, G. L.; Rustgi, A. K.; Vonderheide, R. H.; Leach, S. D.; Stanger, B. Z. EMT and dissemination precede pancreatic tumor formation. Cell 2012, 148(1-2), 349-61.
[14] Kelleher, F. C. Hedgehog signaling and therapeutics in pancreatic cancer. Carcinogenesis 2011, 32(4), 445-451.
[15] Hooper, J. E.; Scott, M. P. Communicating with Hedgehogs. Nat. Rev. Mol. Cell Biol. 2005, 6(4), 306-317.
[16] Stecca, B.; Ruiz, I. A. A. Context-dependent regulation of the GLI code in cancer by HEDGEHOG and non-HEDGEHOG signals. J. Mol. Cell Biol., 2010, 2(2), 84-95.
[17] Li, X.; Ma, Q.; Xu, Q.; Liu, H.; Lei, J.; Duan, W.; Bhat, K.; Wang, F.; Wu, E.; Wang, Z. SDF-1/CXCR4 signaling induces pancreatic cancer cell invasion and epithelial-mesenchymal transition in vitro through non-canonical activation of Hedgehog pathway. Cancer Lett. 2012, 322(2), 169-176.
[18] Deer, E. L.; Gonzalez-Hernandez, J.; Coursen, J. D. Shea, J.E.; Ngatia, J.; Scaife, C.L.; Firpo, M.A.; Mulvihill, S.J. Phenotype and genotype of pancreatic cancer cell lines. Pancreas 2010, 39(4), 425-35.
[19] Bachem, M. G.; Schneider, E.; Gross, H.; Weidenbach, H.; Schmid, R. M.; Menke, A.; Siech, M.; Beger, H.; Grunert, A.; Adler, G. Identification, culture, and characterization of pancreatic stellate cells in rats and humans. Gastroenterology 1998, 115(2), 421-432.
[20] Schmittgen, T. D.; Livak, K. J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3(6), 1101-1108.
[21] Erkan, M.; Adler, G.; Apte, M. V.; Bachem, M. G.; Buchholz, M.; Detlefsen, S.; Esposito, I.; Friess, H.; Gress, T. M.; Habisch, H. J.; Hwang, R. F.; Jaster, R.; Kleeff, J.; Kloppel, G.; Kordes, C.; Logsdon, C. D.; Masamune, A.; Michalski, C. W.; Oh, J.; Phillips, P. A.; Pinzani, M.; Reiser-Erkan, C.; Tsukamoto, H.; Wilson, J. StellaTUM: current consensus and discussion on pancreatic stellate cell research. Gut 2012, 61(2), 172-178.
[22] Linder, S.; Castanos-Velez, E.; von, R. A.; Biberfeld, P. Immuno- histochemical expression of extracellular matrix proteins and adhesion molecules in pancreatic carcinoma. Hepatogastroenterology 2001, 48(41), 1321-1327.
[23] Shintani, Y.; Hollingsworth, M. A.; Wheelock, M. J.; Johnson, K. R. Collagen I promotes metastasis in pancreatic cancer by activating c-Jun NH(2)-terminal kinase 1 and up-regulating N-cadherin expression. Cancer Res. 2006, 66(24), 11745-11753.
[24] Medici, D.; Nawshad, A. Type I collagen promotes epithelial-mesenchymal transition through ILK-dependent activation of NF-kappaB and LEF-1. Matrix Biol. 2010, 29(3), 161-165.
[25] Shintani, Y.; Maeda, M.; Chaika, N.; Johnson, K. R.; Wheelock, M. J. Collagen I promotes epithelial-to-mesenchymal transition in lung cancer cells via transforming growth factor-beta signaling. Am. J. Respir. Cell Mol. Biol. 2008, 38(1), 95-104.
[26] Eble, J. A.; Haier J. Integrins in cancer treatment. Curr. Cancer Drug Targets 2006, 6(2), 89-105.
[27] Goel, H. L.; Underwood, J. M.; Nickerson, J. A.; Hsieh, C. C.; Languino, L. R. Beta1 integrins mediate cell proliferation in three-dimensional cultures by regulating expression of the sonic hedgehog effector protein, GLI1. J. Cell Physiol. 2010, 224(1), 210-217.
[28] Li, X.; Ma, Q.; Duan, W.; Liu, H.; Xu, H.; Wu, E. Paracrine sonic hedgehog signaling derived from tumor epithelial cells: a key regulator in the pancreatic tumor microenvironment. Crit. Rev. Eukaryot. Gene Expr. 2012, 22(2), 97-108.
[29] Theunissen, J. W.; de Sauvage, F. J. Paracrine Hedgehog signaling in cancer. Cancer Res. 2009, 69(15), 6007-6010.
[30] Drenkhahn, S.K.; Jackson, G. A.; Slusarz, A.; Starkey, N. J.; Lubahn, D. B. Inhibition of hedgehog/Gli signaling by botanicals: a review of compounds with potential hedgehog pathway inhibitory activities. Curr Cancer Drug Targets 2013, 13(5), 580-95.
[31] Grzesiak, J. J.; Ho, J. C.; Moossa, A. R.; Bouvet, M. The integrin-extracellular matrix axis in pancreatic cancer. Pancreas 2007, 35(4), 293-301.
[32] Stagge, V.; Seufferlein, T.; Duerschmied, D.; Menke, A.; Adler, G.; Beil, M. Integrin-mediated differentiation of a pancreatic carcinoma cell line is independent of FAK or MAPK activation levels. Pancreas 2001, 23(3), 236-245.
[33] Stecca, B.; Mas, C.; Clement, V.; Zbinden, M.; Correa, R.; Piguet, V.; Beermann, F.; Ruiz, I. A. A. Melanomas require HEDGEHOG-GLI signaling regulated by interactions between GLI1 and the RAS-MEK/AKT pathways. Proc. Natl. Acad. Sci. U.S.A. 2007, 104(14), 5895-5900.
[34] Shintani, Y.; Wheelock, M.; J.Johnson, K. R. Phosphoinositide-3 kinase-Rac1-c-Jun NH2-terminal kinase signaling mediates collagen I-induced cell scattering and up-regulation of N-cadherin expression in mouse mammary epithelial cells. Mol. Biol. Cell 2006, 17(7), 2963-2975.
[35] Shintani, Y.; Fukumoto, Y.; Chaika, N.; Svoboda, R.; Wheelock, M. J.; Johnson, K. R. Collagen I-mediated up-regulation of N-

The Activation of β1-Integrin Induces EMT Current Cancer Drug Targets, 2014, Vol. 14, No. 5 457
cadherin requires cooperative signals from integrins and discoidin domain receptor 1. J. Cell Biol. 2008, 180(6), 1277-1289.
[36] Feldmann, G.; Dhara, S.; Fendrich, V.; Bedja, D.; Beaty, R.; Mullendore, M.; Karikari, C.; Alvarez, H.; Iacobuzio-Donahue, C.; Jimeno, A.; Gabrielson, K. L.; Matsui, W.; Maitra, A. Blockade of hedgehog signaling inhibits pancreatic cancer invasion and metastases: a new paradigm for combination therapy in solid cancers. Cancer Res. 2007, 67(5), 2187-2196.
[37] Rhim, A. D.; Mirek, E. T.; Aiello, N. M.; Maitra, A.; Bailey, J. M.; McAllister, F.; Reichert, M.; Beatty, G. L.; Rustgi, A. K.; Vonderheide, R. H.; Leach, S. D.; Stanger, B. Z. EMT and dissemination precede pancreatic tumor formation. Cell 2012, 148(1-2), 349-361.
[38] Lei, J.; Ma, J.; Ma, Q.; Li, X.; Liu, H.; Xu, Q.; Duan, W.; Sun, Q.; Xu, J.; Wu, Z.; Wu, E. Hedgehog signaling regulates hypoxia induced epithelial to mesenchymal transition and invasion in pancreatic cancer cells via a ligand-independent manner. Mol. Cancer 2013, 12, 66.
[39] Das, S.; Harris, L. G.; Metge, B. J.; Liu, S.; Riker, A. I.; Samant, R. S.; Shevde, L. A. The hedgehog pathway transcription factor GLI1 promotes malignant behavior of cancer cells by up-regulating osteopontin. J. Biol. Chem. 2009, 284(34), 22888-22897.
[40] Inaguma, S.; Kasai, K.; Ikeda, H. GLI1 facilitates the migration and invasion of pancreatic cancer cells through MUC5AC-mediated attenuation of E-cadherin. Oncogene 2011, 30(6), 714-723.
[41] Joost, S.; Almada, L. L.; Rohnalter, V.; Holz, P. S.; Vrabel, A. M.; Fernandez-Barrena, M. G.; McWilliams, R. R.; Krause, M.; Fernandez-Zapico, M. E.; Lauth, M. GLI1 inhibition promotes epithelial-to-mesenchymal transition in pancreatic cancer cells. Cancer Res. 2012, 72(1), 88-99.
[42] Joost, S.; Almada, L. L.; Rohnalter, V.; Holz, P. S.; Fernandez-Barrena, M. G.; Fernandez-Zapico, M.; E.Lauth, M. GLI1 Modulates EMT in Pancreatic Cancer--Response. Cancer Res. 2012, 72(14), 3704-3705.
Received: August 20, 2013 Revised: January 26, 2014 Accepted: April 01, 2014
Related Documents











