Ž . Molecular Brain Research 75 2000 225–236 www.elsevier.comrlocaterbres Research report The 5 X 2 promoter of the neuronal nitric oxide synthase dual promoter complex mediates inducibility by nerve growth factor Terrie K. Rife a,b , Jinling Xie a,b , Chanelle Redman b , Anthony P. Young a,b,c, ) a Ohio State Biochemistry Program, The Ohio State UniÕersity, Columbus, OH, USA b Neurobiotechnology Center, The Ohio State UniÕersity, Columbus, OH, USA c DiÕision of Pharmacology, College of Pharmacy, The Ohio State UniÕersity, Columbus, OH, USA Accepted 5 October 1999 Abstract Ž . Ž . Neuronal nitric oxide synthase nNOS is induced by nerve growth factor NGF in pheochromocytoma PC12 cells. Previous studies Ž X X . from our laboratory identified two closely linked promoters designated 5 1 and 5 2 that mediate transcription of the human nNOS gene w in the brain J. Xie, P. Roddy, T.K. Rife, F. Murad, A.P. Young, Two closely linked but separable promoters for human neuronal nitric Ž . x oxide synthase gene transcription, Proc. Natl. Acad. Sci. U. S. A. 92 1995 1242–1246 . In this report, we demonstrate that luciferase fusion genes under transcriptional control by the 5 X 1 and 5 X 2 dual promoter complex are inducible by NGF in stably transformed PC12 Ž . Ž . cells. In sharp contrast, neither epidermal growth factor EGF nor fibroblast growth factor 2 FGF2 are able to significantly enhance the expression of NOS–luciferase fusion genes. Deletion studies indicate that the 5 X 2 promoter plays a major role in mediating NGF inducibility. The 5 X 2 promoter contains six potential Ets binding sites as well as four potential AP1 binding sites. Thus, it is possible that activation of Ets andror AP1 transcription factors by the Ras–Raf–MAP kinase cascade contributes to the NGF-mediated induction of nNOS. q 2000 Published by Elsevier Science B.V. All rights reserved. Keywords: Nitric oxide synthase; Gene expression; Nerve growth factor inducibility; Signal transduction mechanism; Ets factor 1. Introduction Ž . Nitric oxide NO is a signaling molecule that acts in a wx vast array of physiological processes 7 . In the central Ž . nervous system CNS , NO is thought to function as a w x mediator of long-term potentiation 41 and synaptic plas- w x ticity and remodeling 21 . NO is also a major contributor w x to neuronal cell death during ischemia 40 and possibly w x neurodegenerative disease 38 , most likely through its wx reaction with superoxide anion to form peroxynitrite 1 . AbbreÕiations: DMEM, Dulbecco’s modified Eagle’s medium; EGF, epidermal growth factor; FGF2, fibroblast growth factor 2; NGF, nerve growth factor; NO, nitric oxide; NOS, nitric oxide synthase; nNOS, neuronal NOS; PBS, phosphate-buffered saline; PCR, polymerase chain reaction; RLU, relative light unit; RT, reverse transcription; S.D., stan- dard deviation ) Corresponding author. Neurobiotechnology Center, The Ohio State University, 182 Rightmire Hall, 1060 Carmack Rd., Columbus, OH 43210, USA. Fax: q 1-614-292-5379; e-mail: [email protected] Ž . Three distinct isoforms of nitric oxide synthase NOS , the enzyme responsible for NO biosynthesis, are expressed wx in different cells and tissues 8 . NOS1 or neuronal NOS Ž . wx nNOS is the predominant isoform found in neurons 6. Because of the pivotal roles played by NO in CNS physiol- ogy and pathophysiology, achieving a more profound un- derstanding of the control of NOS1 gene expression in neurons is an important objective. The human NOS1 gene contains 29 exons that span w x X over 150 kb 19 . Moreover, the 5 UTR of NOS1 mRNA is heterogeneous, with several different first exons ex- w x w x pressed in the CNS 53 and other tissues 47 . Our labora- tory has established that human NOS1 mRNAs with at least four different 5 X terminal exons are present in the Ž CNS G. Hartt, S. Pierson, A.P. Young, manuscript in . w x X preparation 53 . Two of these 5 terminal exons, desig- nated 5 X 1 and 5 X 2, are transcribed from closely linked but w x X X separable promoters 53 . Interestingly, exons 5 2 and 5 1 bear strong sequence similarity to the 5 X and 3 X ends, respectively, of a single rodent NOS1 exon 1 that has been w x wx designated exon 1a in rats 26 and mice 9 . 0169-328Xr00r$ - see front matter q 2000 Published by Elsevier Science B.V. All rights reserved. Ž . PII: S0169-328X 99 00293-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ž .Molecular Brain Research 75 2000 225–236www.elsevier.comrlocaterbres
Research report
The 5X 2 promoter of the neuronal nitric oxide synthase dual promotercomplex mediates inducibility by nerve growth factor
Terrie K. Rife a,b, Jinling Xie a,b, Chanelle Redman b, Anthony P. Young a,b,c,)
a Ohio State Biochemistry Program, The Ohio State UniÕersity, Columbus, OH, USAb Neurobiotechnology Center, The Ohio State UniÕersity, Columbus, OH, USA
c DiÕision of Pharmacology, College of Pharmacy, The Ohio State UniÕersity, Columbus, OH, USA
Accepted 5 October 1999
Abstract
Ž . Ž .Neuronal nitric oxide synthase nNOS is induced by nerve growth factor NGF in pheochromocytoma PC12 cells. Previous studiesŽ X X .from our laboratory identified two closely linked promoters designated 5 1 and 5 2 that mediate transcription of the human nNOS gene
win the brain J. Xie, P. Roddy, T.K. Rife, F. Murad, A.P. Young, Two closely linked but separable promoters for human neuronal nitricŽ . xoxide synthase gene transcription, Proc. Natl. Acad. Sci. U. S. A. 92 1995 1242–1246 . In this report, we demonstrate that luciferase
fusion genes under transcriptional control by the 5X1 and 5X2 dual promoter complex are inducible by NGF in stably transformed PC12Ž . Ž .cells. In sharp contrast, neither epidermal growth factor EGF nor fibroblast growth factor 2 FGF2 are able to significantly enhance the
expression of NOS–luciferase fusion genes. Deletion studies indicate that the 5X2 promoter plays a major role in mediating NGFinducibility. The 5X2 promoter contains six potential Ets binding sites as well as four potential AP1 binding sites. Thus, it is possible thatactivation of Ets andror AP1 transcription factors by the Ras–Raf–MAP kinase cascade contributes to the NGF-mediated induction ofnNOS. q 2000 Published by Elsevier Science B.V. All rights reserved.
Keywords: Nitric oxide synthase; Gene expression; Nerve growth factor inducibility; Signal transduction mechanism; Ets factor
1. Introduction
Ž .Nitric oxide NO is a signaling molecule that acts in aw xvast array of physiological processes 7 . In the central
Ž .nervous system CNS , NO is thought to function as aw xmediator of long-term potentiation 41 and synaptic plas-
w xticity and remodeling 21 . NO is also a major contributorw xto neuronal cell death during ischemia 40 and possibly
w xneurodegenerative disease 38 , most likely through itsw xreaction with superoxide anion to form peroxynitrite 1 .
AbbreÕiations: DMEM, Dulbecco’s modified Eagle’s medium; EGF,epidermal growth factor; FGF2, fibroblast growth factor 2; NGF, nervegrowth factor; NO, nitric oxide; NOS, nitric oxide synthase; nNOS,neuronal NOS; PBS, phosphate-buffered saline; PCR, polymerase chainreaction; RLU, relative light unit; RT, reverse transcription; S.D., stan-dard deviation
) Corresponding author. Neurobiotechnology Center, The Ohio StateUniversity, 182 Rightmire Hall, 1060 Carmack Rd., Columbus, OH43210, USA. Fax: q1-614-292-5379; e-mail: [email protected]
Ž .Three distinct isoforms of nitric oxide synthase NOS ,the enzyme responsible for NO biosynthesis, are expressed
w xin different cells and tissues 8 . NOS1 or neuronal NOSŽ . w xnNOS is the predominant isoform found in neurons 6 .Because of the pivotal roles played by NO in CNS physiol-ogy and pathophysiology, achieving a more profound un-derstanding of the control of NOS1 gene expression inneurons is an important objective.
The human NOS1 gene contains 29 exons that spanw x Xover 150 kb 19 . Moreover, the 5 UTR of NOS1 mRNA
is heterogeneous, with several different first exons ex-w x w xpressed in the CNS 53 and other tissues 47 . Our labora-
tory has established that human NOS1 mRNAs with atleast four different 5X terminal exons are present in the
ŽCNS G. Hartt, S. Pierson, A.P. Young, manuscript in. w x Xpreparation 53 . Two of these 5 terminal exons, desig-
nated 5X1 and 5X 2, are transcribed from closely linked butw x X Xseparable promoters 53 . Interestingly, exons 5 2 and 5 1
bear strong sequence similarity to the 5X and 3X ends,respectively, of a single rodent NOS1 exon 1 that has been
w x w xdesignated exon 1a in rats 26 and mice 9 .
0169-328Xr00r$ - see front matter q 2000 Published by Elsevier Science B.V. All rights reserved.Ž .PII: S0169-328X 99 00293-4

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236226
Ž .Nerve growth factor NGF enhances survival and dif-w xferentiation of certain neurons 45 . Progress in understand-
ing the molecular mechanisms that produce these neu-rotrophic effects has been dramatically accelerated by thedevelopment and study of rat PC12 pheochromocytomacells. PC12 cells differentiate and assume a neuronal phe-
w xnotype in response to NGF 18 . Moreover, treatment ofw xPC12 cells 26,36,37,42 or cholinergic neurons of the
w xbasal forebrain 22 with NGF induces production of NOS1.Analysis by RTrPCR demonstrates that exon 1a is the 5X
terminal exon present in NGF-stimulated NOS1 mRNA inw xPC12 cells 26 . These data suggest that the NGF-mediated
differentiation of PC12 cells might provide a biologicalrelevant cell culture system to examine mechanisms oftranscriptional control by the human NOS1 5X1 and 5X 2promoter complex.
This report describes the expression of NOS1-luciferasefusion genes in stably transformed PC12 cells. Our datademonstrate that the 5X1 and 5X 2 dual promoter complexconfers inducibility by NGF. Moreover, deletion studiesindicate that cis elements comprising the 5X 2 promoter aresufficient to mediate this induction. This system providesan experimentally tractable model to elucidate mechanismsof transcriptional control of NOS1 gene expression.
2. Materials and methods
2.1. Plasmids and DNA cloning
All procedures were performed using standard molecu-w x w xlar biologic protocols 29 . The plasmids p4.3NOS1-L 55 ,
w x Ž . Žp2.3NOS1-L 53 , pNOS1 1880r2187 -L akaX . w x Ž . w xp5 1ANOS1-L 53 and pGS y436rq34 -L 27 have
been previously described. The additional NOS1–luciferasefusion genes described in this report were constructedusing PCR-based methodology with oligonucleotides de-signed to generate NOS1 promoter fragments with KpnIand BglII restriction sites at the 5X and 3X ends, respec-
w xtively, followed by subcloning into pXP2 33 . TheseŽ .fusion genes are labeled as pNOS1 xry -L where x and y
reflect the nucleotide position of GenBank Accession No.U15666 and refer to the 5X and 3X boundaries, respectively,of the NOS1 promoter. Structures of all fusion genes wereconfirmed by DNA sequencing.
2.2. Cell culture
HeLa and PC12 cells were grown at 378C under anatmosphere of 5% CO . HeLa cells were grown in Dul-2
Ž . Žbecco’s modified Eagle’s medium DMEM wrsodium. Ž .pyruvate supplemented with 1 grl glucose, 10% vrv
fetal bovine serum, 50 unitsrml penicillin, and 50 unitsrmlstreptomycin. PC12 cells were grown in DMEM supple-
Ž .mented with 4.5 grl glucose, 5% vrv fetal bovineŽ .serum, 5% vrv horse serum, 50 unitsrml penicillin, and
Ž .50 unitsrml streptomycin PC12 cell medium . NGFŽ .mouse, natural , FGF-2, and EGF were purchased fromAustral Biochemicals.
2.3. Generation of stably transformed PC12 cells
Transfection of PC12 cells was performed using Super-Ž .fect reagent Qiagen according to the manufacturer’s rec-
ommendation. Briefly, 27 mg of test plasmid, 3 mg ofŽ .pCDNA neo Invitrogen , and 900 ml of DMEM high
Ž .glucose medium serum- and antibiotic-free were mixedŽ .and centrifuged. Superfect reagent 120 ml was added,
followed by light vortexing for 10 s and incubation atŽ .room temperature for 10 min. PC12 cell medium 9 ml
was added and one-third of this transfection solution wasapplied to 5=105 PC12 cells that had been previouslyplated onto a 100-mm polystyrene dish, incubatedovernight, and washed once with PBS. After 3 h, the plateswere washed four times with PBS followed by incubationin PC12 cell medium for 24 h. The medium was thenreplaced with PC12 cell medium supplemented with 400
Ž .mgrml G418 GibcorBRL and the cells were grown for18–21 days in the presence of the drug with mediumchanges every fourth day. Individual colonies were iso-lated from PBS-washed plates using cloning wells andpropagated in PC12 cell medium supplemented with 200mgrml G418.
As an initial screen, extracts derived from individualcolonies were assayed for luciferase activity. Cells thatfailed to show luciferase activity above background levelswere discarded. Approximately, two-third of the coloniesderived from cells transfected with NOS1- or glutamine
Ž .synthetase GS –luciferase fusion genes survived this ini-tial screening procedure. In contrast, only one-sixth of thecolonies transfected with pXP2 survived the initial screen.
2.4. Enzyme assays
Calcium phosphate-mediated gene transfer of HeLa cellsw xwas performed as published 53 . HeLa or PC12 cells were
harvested by suspension in 1 ml of PBS, followed by twoŽ .cycles of centrifugation using a microfuge 12 000=g
and resuspension in 1 ml PBS. Pellets were stored aty808C. Extracts were prepared from the frozen pellets by
Ž .adding 150–300 ml of ice cold 0.25 mM Tris pH 7.8 andplacing them on ice. The pellets were then sonicatedŽ .seven pulses using a Branson Sonifier 450 equipped witha microtip and set at 50% duty, 6.5 output. Sonicated
Ž .samples were then centrifuged 12 000=g in a microfugefor 10 min and the supernatants utilized for assays of
w x w x w xluciferase 13 , b-galactosidase 35 , andror protein 5using published procedures.
For NOS enzyme assays, PC12 cells were harvested asdescribed above, placed in 150 ml of 200 mM Tris bufferŽ .pH 7.4 , and supernatants prepared by sonication and
w14 xcentrifugation as described above. Conversion of C -

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236 227
w14 xlabeled arginine to C -labeled citrulline was then moni-w xtored as described by Bredt and Synder 6 .
2.5. RTrPCR
To isolate RNA, PC12 cells grown in 10 mm wellpolystyrene culture dishes were rinsed with PBS and incu-
Ž .bated for 5 min with 500 ml Trizol reagent GibcorBRL .The lysates were added to a microfuge tube, vortexedvigorously with 100 ml of chloroform and centrifuged. Theaqueous phase was transferred to a new microfuge tubeand RNA precipitated by centrifugation after a 10-minincubation with 250 ml of isopropyl alcohol. After anadditional suspension and precipitation with isopropyl al-cohol, the RNA pellets were rinsed with 70% ethanol,dried, and dissolved in DEPC-treated H O. RNA concen-2
tration was estimated by measuring the A of an aliquot.260Ž .Total RNA 5 mg was reverse-transcribed in the pres-
ence of 100 ng random hexamers using 0.2 units ofŽ .Superscript II reverse transcriptase GibcorBRL accord-
ing to the enzyme manufacturer’s instructions. One-fifth ofthe reaction mixture was denatured for 2 min at 948C
Žfollowed by 40 cycles of PCR denaturation at 948C for 30s; annealing at 588C for 1 min; and elongation at 728C for
.2 min using enzyme, primers, and buffer componentsaccording to the Taq DNA polymerase manufacturer’s
Ž .instructions GibcorBRL . After the 40th cycle, the reac-tion mix was incubated at 728C for 10 min and stored at48C. Rat NOS1 and b-actin primers were those specified
w xby Lee et al. 26 .
2.6. Statistical treatment of data
Statistical comparisons were conducted using SigmaStatŽ .software Jandel Scientific . Because the data collected
were not normally distributed, non-parametric comparisons
w xwere made 56 . For each promoter sequence, associationsbetween basal gene expression and NGF inducibility were
Ževaluated statistically using Spearman rank order non-.parametric correlation analysis. Sensitivity to NGF induc-
tion was compared among various promoters at similarŽbasal luciferase expression less than 15 RLUs of lu-
.ciferase activityrmg protein for each cell line includedŽ .using non-parametric analysis of variance ANOVA
Ž .Kruskal–Wallis , followed by Student–Newman–Keulsw xpost tests 56 . In all tests statistical significance was
assigned at p-0.05.
3. Results
3.1. NGF- and FGF2-mediated induction of NOS1 inPC12 cells
PC12 cells treated with NGF or FGF2 leave the cellw xcycle and produce an extensive network of neurites 24 .
The induction of NOS1 has been suggested to play animportant role in this process, by producing an NO-media-
w xted block in PC12 cell proliferation 37 . EGF stimulatesŽ . w xproliferation and not differentiation of PC12 cells 30 .
However, one previous report indicated that NOS1 proteinwas induced in PC12 cells treated with NGF, FGF2, or
w xEGF 42 . The PC12 cells obtained by our laboratoryextend neurites in response to treatment with NGF or
Ž .FGF2 but not EGF data not shown . Moreover, theRTrPCR-based semi-quantitative analysis shown in Fig. 1demonstrates that NGF or FGF2 treatment stimulates accu-mulation of NOS1 mRNA while EGF treatment does not.Thus, our data indicate that the growth factor-mediatedinduction of NOS1 is correlated to differentiation of PC12cells. Whether the difference in EGF effects reflects a
Ždifference in culture conditions e.g., serum-containing vs.
Fig. 1. RTrPCR analysis of NOS1 mRNA in PC12 cells treated with growth factors. RNA was isolated from PC12 cells cultured without growth factorsŽ . Ž . Ž . Ž .no treatment or cultured for the indicated number of days with NGF 100 ngrml , FGF2 50 ngrml , or EGF 100 ngrml . RT was then performed usingrandom primers, followed by PCR of NOS1 and b-actin using primers discussed in the text that amplify products of 328 and 736 bp, respectively.
Ž .RTrPCR was also performed in the absence of template RNA no template . PCR products were analyzed by 2% agarose gel electrophoresis. The size ofelectrophoresis markers are indicated.

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236228
.serum-free medium , a clonal difference in the PC12 cells,Žthe difference in the method of assay RTrPCR vs. im-
.munochemical , or an as of yet unappreciated mechanismis not clear.
3.2. PC12 cells that express NGF-inducible NOS1–luciferase fusion genes
The effect of NGF on the human NOS1 5X1 and 5X 2promoter complex was analyzed using colonies of PC12cells that had been stably transformed with NOS1–luciferase fusion genes. As controls, analyses were per-formed using colonies of PC12 cells that were transformedwith either the promoterless luciferase-encoding plasmid
w x w xpXP2 34 or with a GS–luciferase fusion gene 27 . Datasummarized in Fig. 2 shows the effects of 8-day treatmentwith 100 ngrml NGF on the luciferase specific activitiesof individual colonies. For each data point, three plates ofcells were treated with NGF and three plates treatedwithout NGF. Basal luciferase specific activities reflectmean values obtained after analysis of the cells treatedwithout NGF. The fold induction by NGF was defined asthe ratio of mean values obtained with and without NGFtreatment.
Basal luciferase specific activities are low and vary byless that an order of magnitude in colonies transformed
Ž .with the promoterless luciferase vector pXP2 Fig. 2A .Ž .Moreover, only one colony gave an appreciable G4-fold
NGF-mediated induction of luciferase. Basal specific activ-ities vary by nearly four orders of magnitude in coloniestransformed with the GS–luciferase fusion gene,
Ž . w x Ž .pGS y436rq34 -L 27 Fig. 2B , suggesting that theGS promoter is sensitive to positional effects. However,NGF treatment produced a G4-fold induction of lu-
Ž .ciferase in only a small fraction of these colonies 3 of 12 .ŽBased on analysis using Spearman rank order non-para-
. w xmetric correlation analysis 56 , there was not a statisti-cally significant relationship between basal expression ofthe reporter and NGF inducibility for either pXP2 or
Ž .pGS y436rq34 -L.Data obtained with the luciferase-encoding plasmid un-
der transcriptional control by the NOS1 dual promoterŽ w x.complex p2.3NOS1-L, 53 are shown in Fig. 2C. As was
Ž .the case with pGS y436rq34 -L, basal luciferase activi-ties vary by nearly four orders of magnitude. However, insharp contrast to the results obtained with the GS–luciferase fusion gene, there is a clear and statistically
Ž .significant p-0.001 inverse relationship between NGFinducibility and basal expression among lines transformedwith p2.3NOS1-L. Among the seven colonies transfectedwith p2.3NOS1-L and showing a basal specific activityF15 RLUsrmg protein, six show an induction by NGFthat is G4-fold. However, none of the eight colonies withbasal expression levels that exceed 15 RLUsrmg proteinshow an NGF-mediated induction that reaches 4-fold.
PC12 cells stably transformed with the NOS1–luciferaseŽ . Ž .fusion genes pNOS1 1r1842 -L, pNOS1 1613r1842 -L,
Ž .and pNOS1 1880r2187 -L have also been analyzed. TheNOS1 promoter fragments contained in these fusion genes
Ž .are shown schematically in Fig. 3. pNOS1 1r1842 -L andŽ . Ž X .pNOS1 1613r1842 -L include only the upstream 5 2
promoter of the dual promoter complex. The promoterfragments within these fusion genes terminate at a com-
X X Žmon 3 boundary within exon 5 2 nucleotide position1842; nucleotide positions discussed in this report all refer
.to GenBank accession number U15666 . However, theŽ . Xpromoter within pNOS1 1r1842 -L has a 5 boundary
Ž .identical to that of p2.3NOS1-L nucleotide position 1Ž .while the pNOS1 1613r1842 -L promoter is truncated
with a 5X boundary at nucleotide position 1613. Con-Ž .versely, pNOS1 1880r2187 -L contains only the down-
Ž X . X Xstream 5 1 promoter, with 5 and 3 boundaries at posi-tions 1880 and 2187, respectively. The data obtained withthese fusion genes are summarized in Fig. 2D–F, respec-tively.
Ž . Ž .The results obtained with pNOS1 1r1842 -L Fig. 2Dclosely resemble those obtained with p2.3NOS1-L. Basalexpression levels are distributed over a wide range and
Ž .there is a statistically significant p-0.001 inverse rela-tionship between NGF inducibility and basal expression.Among the nine colonies transfected with NOS1Ž .1r1842 luc and showing a basal specific activity F15RLUsrmg protein, eight show an induction by NGF that isG4-fold. In contrast, none of the nine colonies with basalexpression levels that exceed 15 RLUsrmg protein showan NGF-mediated induction that reaches 4-fold.
Ž . Ž .Results obtained with pNOS1 1613r1842 -L Fig. 2Eindicate that truncation of the 5X 2 promoter attenuatesNGF-inducibility. Of the twelve colonies with basal ex-pression levels F15 RLUsrmg protein, only four showan NGF-mediated induction G4-fold. This frequency isclearly diminished relative to p2.3NOS1-L and, moreover,
Ž .pNOS1 1r1842 -L. Consistent with this observation, thereis not a statistically significant relationship between in-ducibility by NGF and basal expression among colonies
Ž .transformed with pNOS1 1613r1842 -L.NGF-inducibility G4-fold was observed in approxi-
Ž .mately one-half four of nine of the colonies showing lowŽ .basal expression specific activities F15 RLUsrmg of
Ž . XpNOS1 1880r2187 -L, a plasmid containing only the 5 1Ž .promoter Fig. 2F . High basal expression of this fusion
gene was accompanied by a general lack of NGF-induci-bility. An inverse relationship exists between NGF in-ducibility and basal expression but reaches statistical sig-
Ž .nificance at a lower confidence level p-0.01 than thatobtained from the data for p2.3NOS1-L or
Ž . Ž .pNOS1 1r1842 -L p-0.001 .The differences in NGF inducibility among the various
fusion genes were compared using non-parametric ANOVAŽ .Kruskal–Wallis , followed by Student–Newman–Keuls
w xpost tests 56 . Because NGF-inducibility is linked to low
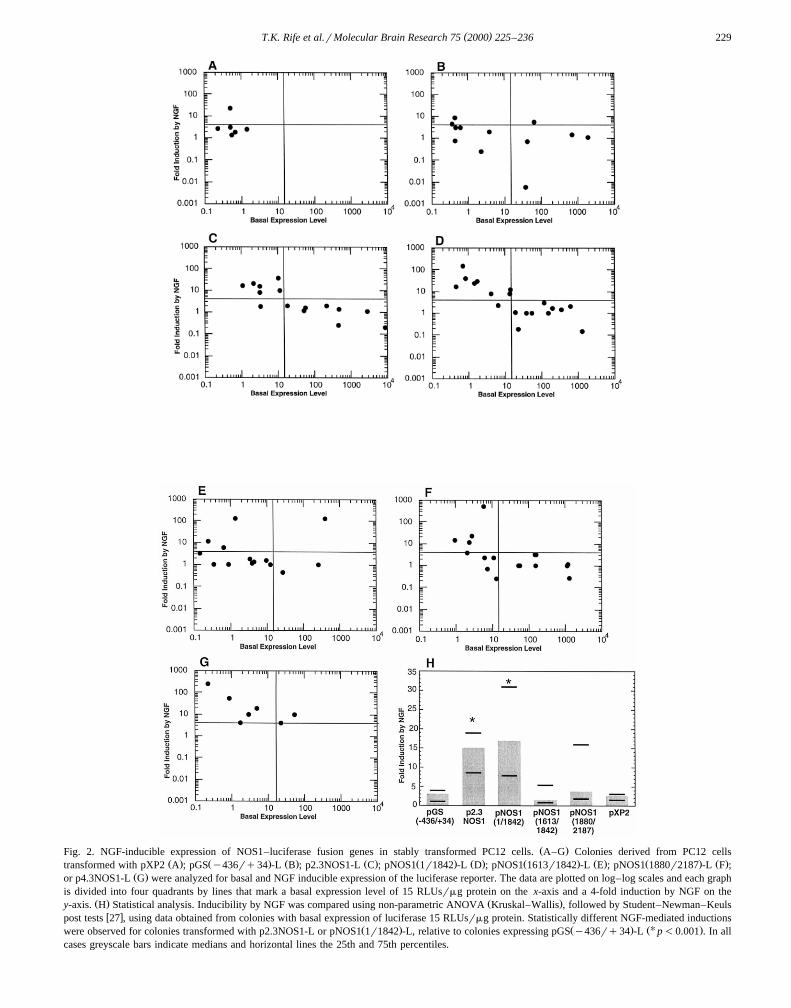
( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236 229
Ž .Fig. 2. NGF-inducible expression of NOS1–luciferase fusion genes in stably transformed PC12 cells. A–G Colonies derived from PC12 cellsŽ . Ž . Ž . Ž . Ž . Ž . Ž . Ž . Ž . Ž .transformed with pXP2 A ; pGS y436rq34 -L B ; p2.3NOS1-L C ; pNOS1 1r1842 -L D ; pNOS1 1613r1842 -L E ; pNOS1 1880r2187 -L F ;
Ž .or p4.3NOS1-L G were analyzed for basal and NGF inducible expression of the luciferase reporter. The data are plotted on log–log scales and each graphis divided into four quadrants by lines that mark a basal expression level of 15 RLUsrmg protein on the x-axis and a 4-fold induction by NGF on the
Ž . Ž .y-axis. H Statistical analysis. Inducibility by NGF was compared using non-parametric ANOVA Kruskal–Wallis , followed by Student–Newman–Keulsw xpost tests 27 , using data obtained from colonies with basal expression of luciferase 15 RLUsrmg protein. Statistically different NGF-mediated inductions
Ž . Ž . ŽU .were observed for colonies transformed with p2.3NOS1-L or pNOS1 1r1842 -L, relative to colonies expressing pGS y436rq34 -L p-0.001 . In allcases greyscale bars indicate medians and horizontal lines the 25th and 75th percentiles.
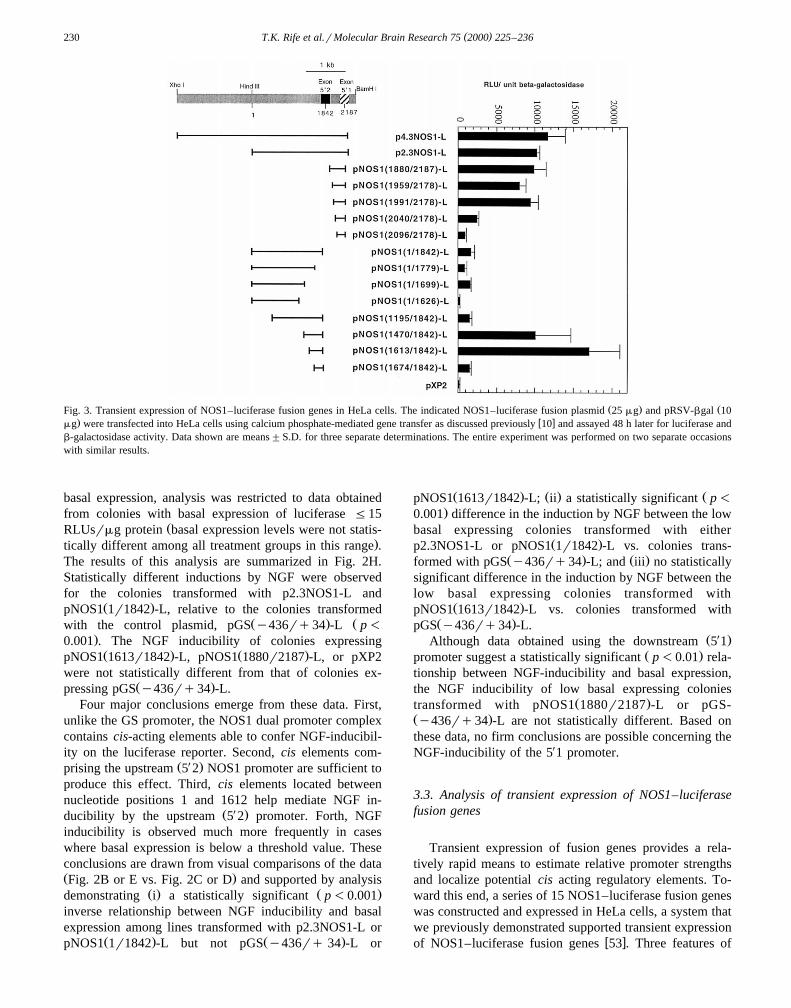
( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236230
Ž . ŽFig. 3. Transient expression of NOS1–luciferase fusion genes in HeLa cells. The indicated NOS1–luciferase fusion plasmid 25 mg and pRSV-bgal 10. w xmg were transfected into HeLa cells using calcium phosphate-mediated gene transfer as discussed previously 10 and assayed 48 h later for luciferase and
b-galactosidase activity. Data shown are means"S.D. for three separate determinations. The entire experiment was performed on two separate occasionswith similar results.
basal expression, analysis was restricted to data obtainedfrom colonies with basal expression of luciferase F15
ŽRLUsrmg protein basal expression levels were not statis-.tically different among all treatment groups in this range .
The results of this analysis are summarized in Fig. 2H.Statistically different inductions by NGF were observedfor the colonies transformed with p2.3NOS1-L and
Ž .pNOS1 1r1842 -L, relative to the colonies transformedŽ . Žwith the control plasmid, pGS y436rq34 -L p-
.0.001 . The NGF inducibility of colonies expressingŽ . Ž .pNOS1 1613r1842 -L, pNOS1 1880r2187 -L, or pXP2
were not statistically different from that of colonies ex-Ž .pressing pGS y436rq34 -L.
Four major conclusions emerge from these data. First,unlike the GS promoter, the NOS1 dual promoter complexcontains cis-acting elements able to confer NGF-inducibil-ity on the luciferase reporter. Second, cis elements com-
Ž X .prising the upstream 5 2 NOS1 promoter are sufficient toproduce this effect. Third, cis elements located betweennucleotide positions 1 and 1612 help mediate NGF in-
Ž X .ducibility by the upstream 5 2 promoter. Forth, NGFinducibility is observed much more frequently in caseswhere basal expression is below a threshold value. Theseconclusions are drawn from visual comparisons of the dataŽ .Fig. 2B or E vs. Fig. 2C or D and supported by analysis
Ž . Ž .demonstrating i a statistically significant p-0.001inverse relationship between NGF inducibility and basalexpression among lines transformed with p2.3NOS1-L or
Ž . Ž .pNOS1 1r1842 -L but not pGS y436rq 34 -L or
Ž . Ž . ŽpNOS1 1613r1842 -L; ii a statistically significant p-.0.001 difference in the induction by NGF between the low
basal expressing colonies transformed with eitherŽ .p2.3NOS1-L or pNOS1 1r1842 -L vs. colonies trans-
Ž . Ž .formed with pGS y436rq34 -L; and iii no statisticallysignificant difference in the induction by NGF between thelow basal expressing colonies transformed with
Ž .pNOS1 1613r1842 -L vs. colonies transformed withŽ .pGS y436rq34 -L.
Ž X .Although data obtained using the downstream 5 1Ž .promoter suggest a statistically significant p-0.01 rela-
tionship between NGF-inducibility and basal expression,the NGF inducibility of low basal expressing colonies
Ž .transformed with pNOS1 1880r2187 -L or pGS-Ž .y436rq34 -L are not statistically different. Based onthese data, no firm conclusions are possible concerning theNGF-inducibility of the 5X1 promoter.
3.3. Analysis of transient expression of NOS1–luciferasefusion genes
Transient expression of fusion genes provides a rela-tively rapid means to estimate relative promoter strengthsand localize potential cis acting regulatory elements. To-ward this end, a series of 15 NOS1–luciferase fusion geneswas constructed and expressed in HeLa cells, a system thatwe previously demonstrated supported transient expression
w xof NOS1–luciferase fusion genes 53 . Three features of

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236 231
these data, shown in Fig. 3, are notable. First, expressionŽ .of pNOS1 1613r1842 -L is well above background levels.
Hence, the attenuated NGF inducibility of this constructdoes not appear to result from a loss of core promoter
Ž .function. Second, a negative acting regulatory element sappears to be located between nucleotides 1196 and 1469.This interpretation is supported by results indicating that
Ž .pNOS1 1470r1842 -L is expressed at a level approxi-Ž .mately 7-fold higher than either pNOS1 1r1842 -L or
Ž .pNOS1 1195r1842 -L. Third, a positive acting regulatoryelement appears to be located between nucleotides 1627and 1698, as evidenced by the relative expression levels of
Ž . Ž .pNOS1 1r1699 -L and pNOS1 1r1626 -L. The potentialcontribution of these elements to the NGF-mediated induc-tion of the NOS1 promoter complex are discussed below.
3.4. Expression of NOS1–luciferase fusion genes duringNGF-mediated differentiation of PC12 cells
We determined whether the induction of an NOS1–luciferase fusion gene during NGF-mediated PC12 celldifferentiation truly recapitulated that of the endogenousNOS1 gene. Three parameters of the response were con-
Ž . Ž .sidered: i the timing, ii the concentration dependence,Ž .and iii the effects of glucocorticoid hormones. For these
Fig. 4. Time course of the NGF-mediated induction of NOS1–luciferasefusion gene expression in stably transformed PC12 cells. The p4.3NOS1-L
Ž . Ž .expressing lines 4.3 C top and 4.3 F bottom were cultured for theindicated time in the presence of 100 ngrml NGF with medium changesevery second day followed by harvesting and assay for luciferase specificactivity. Data shown are means"S.D. for three separate determinations.
Fig. 5. Concentration dependence of the NGF-mediated induction ofNOS1–luciferase fusion gene expression in stably transformed PC12cells. The p4.3NOS1-L expressing line 4.3F was cultured for 8 days inthe presence of the indicated concentration of NGF with medium changesevery second day followed by harvesting and assay for luciferase specificactivity. Data shown are means based on three separate determinations.An EC of 6.5 ngrml was calculated from these data using Graph Pad50
software.
studies, colonies of PC12 cells stably transformed withp4.3NOS1-L were examined. In this plasmid, transcrip-tional control is mediated by an NOS1 promoter with a 5X
boundary extending an additional 2 kb upstream that ofŽ .p2.3NOS1-L see Fig. 3 . As shown in Fig. 2G,
p4.3NOS1-L expressing colonies showed basal specificactivities ranging from 0.2 to 68 RLUsrmg and NGFinductions ranging from 4- to 200-fold. Four of thesep4.3NOS1-L-expressing colonies were selected for addi-tional analysis.
Differentiation of PC12 cells, as evidenced by an exten-sive network of neurites, requires approximately 1 week of
w xcontinuous exposure to NGF 18 . However, kinase signal-ing cascades are initiated within minutes of NGF treatment
Fig. 6. The effect of dexamethasone on the NGF-mediated induction ofNOS1–luciferase fusion gene expression in stably transformed PC12cells. The p4.3NOS1-L expressing line 4.3C was cultured for the indi-cated number of days in the presence of 100 ngrml NGF with mediumchanges every second day. After 4 days, the incubation medium was
Ž .supplemented with either vehicle solution alone NGF, solid circles orŽ . Ždexamethasone final concentration 1 mM in the vehicle solution NGF
.qDEX, open circles . Cells were harvested on the indicated days andassayed for luciferase specific activity. Data shown are means"S.D. forthree separate determinations. For this experiment, all serum was charcoalstripped prior to use.

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236232
w x24,54 , resulting in the enhanced transcription of immedi-w xate early gene products 43 . Transcription of other gene
products, the delayed-early genes, is initiated within sev-eral hours of NGF treatment, via a protein synthesis-de-pendent mechanism that is likely to require immediate
w x w xearly gene expression 16 . Finally, late genes 43 arew xtranscribed hours to days after NGF treatment 46 . As
shown in Fig. 1, NOS1 mRNA accumulates slowly overthe first week of treatment with NGF. Thus, the inductionof NOS1 appears to be a very late event in NGF-mediatedsignaling. In two different sublines of p4.3NOS1-L-ex-
Ž .pressing cells lines 4.3C and 4.3F , luciferase expression
continues to rise for approximately 1 week during expo-Ž .sure to 100 ngrml NGF Fig. 4 . Similar results were
Ž .obtained using sublines 4.3B and 4.3D data not shown .The profile of NOS1–luciferase fusion gene expressionclosely resembles that of the endogenous NOS1 gene andsupports the hypothesis that common molecular mecha-nisms mediate both responses.
NGF binds to two classes of receptors that are presentŽ .on the surface of PC12 cells, the high affinity trk A
Ž .receptor and the low affinity p75 receptor. Dissociationw125 xconstants calculated based on I -labeled NGF binding
to these two classes of receptors are approximately 7.5 and
Fig. 7. Growth factor-mediated inductions of NOS1–luciferase fusion gene expression and NOS1 enzyme activity in stably transformed PC12 cells. TheŽ .indicated p4.3NOS1-L expressing sublines were incubated for eight days without growth factors no treatment or with 100 ngrml NGF, 50 ngrml FGF2,
Ž . Ž .or 100 ngrml EGF as indicated with medium changes every 2 days followed by assay for luciferase A or NOS B specific activity. Data shown aremeans"S.D. for three separate determinations.

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236 233
w x100 ngrml, respectively 2 . Moreover, the concentrationdependence of NGF-mediated differentiation of PC12 cells
Žclosely resembles that of binding to the high affinity trk.A receptor. Data shown in Fig. 5 demonstrate that the
NGF-mediated induction of luciferase in line 4.3F is satu-ratable and mediated through a receptor with an apparentdissociation constant of 6.5 ngrml. Thus, like differentia-tion of PC12 cells, the induction of NOS1–luciferasefusion gene expression appears to be mediated throughNGF binding to the high affinity trk A receptor.
Glucocorticoid hormones are well known to opposeNGF-mediated inductions of a variety of marker genes in
w xPC12 cells 14,15,17 . Moreover, glucocorticoids have beenreported to diminish NOS1 gene expression in rat hip-
w x w xpocampus 28,39,50 and cerebellum 50 . Therefore, weexamined the effects of the synthetic glucocorticoid dexa-methasone on expression of the p4.3NOS1-L in the 4.3Csubline of PC12 cells. Dexamethasone treatment of naive
Ž .cells elicits a modest induction 1.75-fold of luciferaseŽ .activity data not shown . To test the effects of dexametha-
sone and NGF cotreatment on NOS1 expression, the 4.3CŽ .subline of PC12 cells was treated with NGF 100 ngrml
for 4 days followed by an treatment for an additional 1–4days with NGF in the presence or absence of 0.1 mMdexamethasone. Data summarized in Fig. 6 demonstratethat dexamethasone treatment attenuates the induction byNGF.
3.5. EGF or FGF2 do not substantially induce p4.3NOS1-Lin stably transformed PC12 cells
Our findings demonstrate that the NOS1 dual promotercomplex contains cis elements sufficient to confer NGF-mediated inducibility on an appropriately linked reportervia a mechanism that is likely to parallel that of theendogenous NOS1 gene. We questioned whether the re-sponses of the endogenous NOS1 gene and the NOS1–luciferase fusion gene to treatment with FGF2 and EGFwould also correlate. Data shown in Fig. 7a summarize theresults obtained after 8-day treatment of four p4.3NOS1-L
Ž . Ž .expressing sublines with i no growth factor, ii NGF,Ž . Ž .iii EGF, or iv FGF2. As expected, treatment with NGFincreased luciferase gene expression in all four sublines.Treatment with EGF had no major effect, a result inconcert with the inability of EGF to induce endogenous
Ž .NOS1 in our system Fig. 1 . Surprisingly, treatment withFGF2 produced only modest inductions of luciferase in
Ž . Ž . Ž .lines 4.3C 2.6-fold , 4.3D 1.5-fold , and 4.3F 1.4-foldand failed to affect expression of the reporter in line 4.3BŽ .1.0-fold .
Since FGF2 did not produce a robust induction ofp4.3NOS1-L, it is possible that the plasmid lacks an
Ž .essential FGF2 response element s . If so, distinct ciselements must mediate the response to NGF and FGF2,because p4.3NOS1-L is clearly NGF responsive. However,it is also possible that clonal alterations in the amplitude of
FGF2 signaling are responsible for the limited inducibilityof p4.3NOS1-L. To provide a quantitative index of signal-ing, the 4.3C and 4.3F sublines were subjected to growthfactor treatment for 8 days followed by harvesting of cellsand assay for NOS enzyme activity. The results obtainedare summarized in Fig. 7b. Basal NOS enzyme activitieswere similar for both sublines and NGF treatment pro-duced inductions of 12.1- and 4.2-fold in sublines 4.3Cand 4.3F, respectively. EGF treatment did not induce NOSenzyme activity in either subline. FGF2 treatment pro-duced inductions of 5.7- and 2.5-fold in sublines 4.3C and4.3F, respectively. Thus, although there was clonal varia-tion, the FGF2-mediated induction of NOS enzyme activ-ity was approximately one-half that mediated by NGF ineach case. Subline 4.3C shows an FGF2-mediated induc-tion of luciferase that approaches one-half of that mediatedby NGF. However, the other three sublines show sharplyattenuated FGF2-mediated inductions. These data suggestthat p4.3NOS1-L lacks cis elements essential to mediatean induction by FGF2.
4. Discussion
4.1. NGF-mediated inductions of NOS1–luciferase fusiongenes are restricted to PC12 cells that express low basalleÕels of the reporter
This report demonstrates that the human NOS1 dualpromoter complex confers NGF-inducible expression onthe luciferase reporter in lines of stably transformed PC12cells, provided that the basal level of expression of thereporter is low. Luciferase is not generally NGF-induciblein lines that exhibit high basal expression. Why is this thecase?
The variation in inducibility does not reflect clonaldifferences in NGF-signaling. To test this, endogenousNOS enzyme activity was assayed in six cell lines thatexpressed p2.3NOS1-L. Three lines showed high basal
Ž .expression of luciferase 18.5–463 RLUsrmg that wasnot NGF-inducible and three lines showed low basal ex-
Ž .pression of luciferase 1.1–10.5 RLUsrmg that wasNGF-inducible. The basal levels of NOS enzyme activitywere similar in all six lines and inducibility by NGF
Ž .ranged only from 4- to 10-fold data not shown .Consequently, the differences in reporter gene expres-
sion are more likely to reflect differences in the numberŽ .andror positioning of the NOS1–luciferase fusion gene s
in different sublines. It is possible that cell lines showinghigh basal expression contain a large number of copies ofthe NOS1–luciferase fusion gene. Moreover, increasedcopy number might be linked to low NGF inducibility.Although we do not favor this model, it should be notedthat amplification of the metallothionein I gene in tissueculture cells results in a loss of inducibility by glucocorti-
w xcoids 31 . Alternatively, integration of the NOS1–

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236234
luciferase fusion gene near powerful endogenous enhancerelements might lead to high basal expression and overridethe NGF-mediated activation of the NOS1 promoter. Sup-port for this model is provided by studies showing thatpositional effects produce aberrant patterns of fusion gene
w xexpression in transgenic animals 52 .Two additional points should be considered with respect
to the relationship between basal expression and NGFinducibility of NOS1–luciferase fusion genes. First, ex-pression of endogenous NOS1 in naive PC12 cells is weak,as evidenced by the modest signal obtained by RTrPCRŽw x .26 , Fig. 1 . Hence, low basal expression coupled withNGF inducibility reflect properties of the endogenousNOS1 gene. Second, our study predicts that expression ofthe relevant NOS1–luciferase fusion genes will not beNGF-inducible if assayed after pooling colonies of stablytransformed cells. This is because the enzyme activitymeasured for the pool will disproportionately reflect the
Ž .activities of the highest expressing non-induciblecolonies. Consequently, although it is more tedious, theanalysis of individual colonies was necessary in order toreveal NGF inducibility of fusion gene expression.
4.2. Molecular mechanisms mediating NGF inducibility
Luciferase fusion genes under transcriptional control byX X Žeither the 5 1 and 5 2 dual promoter complex p4.3 NOS1-L
. Xand p2.3NOS1-L or the 1.8-kb 5 2 promoter aloneŽ Ž .pNOS1 1r1842 -L are generally NGF-inducible in stablytransformed PC12 cells that express low basal levels of the
Ž .reporter. pNOS1 1613r1842 -L, a truncated version of the5X 2–luciferase fusion gene lacking 1.6 kb of 5X flankingDNA, is less likely to be inducible. Inspection of therelevant sequence as well as analysis of transient expres-sion data suggest possible mechanisms that underlie NGFinducibility by the NOS1 promoter.
Fig. 8 shows the distribution of consensus binding sitesfor transcription factors located within the 5X 2 promoter,
w xbased on analysis using the program TF search 20 . It isstriking that six potential binding sites for members of theEts or TCF subfamilies of Ets transcription factors as wellas four potential AP1 binding sites are revealed. Moreover,five of the six Ets sites and all four AP1 sites lie betweennucleotides 1 and 1613. Ets proteins represent a largefamily of transcription factors that serve as phosphoryla-tion targets of growth factor-mediated signaling through
w xthe Ras–Raf–MAP kinase cascade 48 and appear to actw xin conjunction with other transcription factors 4 . Thus,
Ž .ras-responsive elements RREs from the polyoma en-w x w xhancer 10 and several cellular genes 49 have been
shown to consist of juxtaposed Ets and AP1 sites. RREshave also been shown to contain binding sites for Etsproteins that function in conjunction with cell type-specific
w xtranscription factors 3 . Consequently, it is intriguing tospeculate that NGF treatment stimulates NOS1 transcrip-tion via ras-mediated activation of RREs that require Ets
Fig. 8. Transcription factor binding sites in the NOS1 5X 2 promoter.Potential binding sites for the transcription factors are indicated, based on
Ž .analysis using the program Tfsearch 42 : Ets and CTF subfamilies of Etsfactors, highlighted greyscale; AP1, boxed; ATFrCREB, single under-lined; E2F, double underlined; Oct-1, overlined; MZF, double overlined.
factors functioning in concert with AP1 andror neuron-specific transcription factors.
Transient expression data shown in Fig. 3 suggest anadditional potential target on the 5X 2 promoter. Deletion ofcis elements located between nucleotides 1195 and 1428enhances NOS1–luciferase fusion gene expression in HeLa
Ž .cells, suggesting that a negative regulator s of NOS1transcription maps to this region. Although this sequencelacks similarity to previously identified non-neuronal si-
w xlencers 25,32,51 , it remains possible that NGF ultimatelyacts by relieving transcriptional constraints that map to thisor other regions of the 5X 2 promoter.
Comparison of transient expression of pNOS1Ž . Ž .1r1626 -L with that of pNOS1 1r1699 -L and
Ž . Ž .pNOS1 1674r1842 -L with that of pNOS1 1613r1842 -LŽ .Fig. 3 is also of interest. These data demonstrate thatdeletion of nucleotides 1626 to 1699 from the 3X end of thepromoter or nucleotides 1613 to 1674 from the 5X end ofthe promoter sharply attenuates expression, respectively,suggesting that nucleotides 1626 through 1674 play acritical role in 5X 2 core promoter function. Hence, it ispossible that cis elements located between nucleotides1626 and 1674 are necessary but not sufficient to achieveNGF inducibility. Interestingly, sites for the transcription
w x w x w xfactors Oct1 44 , MZF 23 , and E2F 12 map to thisregion.
GAP-43, a gene that is NGF-inducible in PC 12 cellsw x14 , contains a non-neuronal silencer distinct from that of

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236 235
the SCG10 and sodium channel type II genes. The silenceris located immediately downstream of the GAP-43 TATA
w xbox, an arrangement that is essential to its functioning 51 .The NOS1 5X 2 promoter contains a putative TATA box
w xlocated at nts 1600–1609 19 . Moreover, the region im-mediately downstream of this TATA box is homologous to
w xthe GAP-43 silencer 51 . Hence, it is possible that NGFacts to relieve negative regulation at this site and induceboth GAP-43 and NOS1. However, it should be noted that
Ž .the transient expression data Fig. 3 do not provide exper-imental support for negative regulation at this locus of the5X 2 promoter.
Finally, it is interesting that the NOS1 dual promotercomplex does not confer inducibility by EGF or FGF2 onthe luciferase reporter. EGF, FGF2, and NGF all activate
w xRAS–RAF–MAP kinase cascades in PC12 cells 30 andinduce the expression of similar immediate early gene
w xproducts 11 . However, with the culture conditions usedhere, EGF neither elicits neurite outgrowth from PC12
Ž .cells data not shown nor induces endogenous NOS1 geneŽ .expression Fig. 1 . Hence, by failing to respond to EGF,
human NOS1–luciferase fusion genes are recapitulating aŽ .property of the endogenous rat NOS1 gene. FGF2 medi-
w xates neurite outgrowth from PC12 cells 24 and inducesŽw x .expression of the endogenous NOS1 gene 42 , Fig. 1 .
However, despite the overall similarities between FGF2and NGF action on PC12 cells, subtle differences mustexist in their signaling, since the human NOS1 dual pro-moter complex is able to discriminate between the twoneurotrophic factors.
Acknowledgements
We wish to thank Dr. John A. Bauer for lending hisexpertise in statistical analysis of these data. This researchhas been supported by NIH grant NS34524.
References
w x1 J.S. Beckman, W.H. Koppenol, Nitric oxide, superoxide, and perox-Ž .ynitrite: the good, the bad, and the ugly, Am. J. Physiol. 271 1996
C1424–C1437.w x 1252 P. Bernd, L.A. Greene, Association of I-nerve growth factor with
PC12 pheochromocytoma cells. Evidence for internalization viahigh-affinity receptors only and for long-term regulation by nervegrowth factor of both high- and low-affinity receptors, J. Biol.
Ž .Chem. 259 1984 15509–15516.w x3 A.P. Bradford, K.E. Conrad, P.H. Tran, M.C. Ostrowski, A. Gutier-
rez-Hartmann, GHF-1rPit-1 functions as a cell-specific integrator ofras signaling by targeting the ras pathway to a composite Ets-
Ž .1rGHF-1 response element, J. Biol. Chem. 271 1996 24639–24648.
w x4 A.P. Bradford, C. Wasylyk, B. Wasylyk, A. Gutierrez-Hartmann,Interaction of Ets-1 and the POU-homeodomain protein GHF-1rPit-1reconstitutes pituitary-specific gene expression, Mol. Cell. Biol. 17Ž .1997 1065–1074.
w x5 M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye
Ž .binding, Anal. Biochem. 72 1976 248–256.w x6 D.S. Bredt, S.H. Synder, Isolation of nitric oxide synthase, a
calmodulin-requiring enzyme, Proc. Natl. Acad. Sci. U. S. A. 87Ž .1990 682–685.
w x7 D.S. Bredt, S.H. Synder, Nitric oxide, a novel neuronal messenger,Ž .Neuron 8 1992 3–11.
w x8 D.S. Bredt, S.H. Synder, Nitric oxide: a physiologic messengerŽ .molecule, Annu. Rev. Biochem. 63 1994 175–195.
w x9 J.E. Brenman, D.S. Chao, S.H. Gee, A.W. McGee, S.E. Craven,D.R. Santillano, Z. Wu, F. Huang, H. Xia, M.F. Peters, S.C.Froehner, D.S. Bredt, Interaction of nitric oxide synthase with thepostsynaptic density protein PSD-95 and alpha 1-syntrophin medi-
Ž .ated by PDZ domains, Cell 84 1996 757–767.w x10 J.T. Bruder, G. Heidecker, U.R. Rapp, Serum-, TPA-, and ras-in-
duced expression from AP-1rEts driven promoters requires Raf-1Ž .kinase, Genes Dev. 6 1992 545–556.
w x11 M.V. Chao, Growth factor signaling: where is the specificity?, CellŽ .68 1992 995–997.
w x12 S.P. Chellappan, S. Hiebert, M. Mudryj, J.M. Horowitz, J.R. Nevins,The E2F transcription factor is a cellular target for the RB protein,
Ž .Cell 65 1991 1053–1061.w x13 J.R. DeWet, K.W. Wood, M. DeLuca, D.R. Helinski, S. Subramani,
Firefly luciferase gene: structure and expression in mammalian cells,Ž .Mol. Cell. Biol. 7 1987 725–737.
w x14 H.J. Federoff, E. Grabczyk, M.C. Fishman, Dual regulation ofGAP-43 gene expression by nerve growth factor and glucocorti-
Ž .coids, J. Biol. Chem. 263 1988 19290–19295.w x15 P.J. Foreman, G. Taglialatela, G.R. Jackson, J.R. Perez-Polo, Dex-
amethasone blocks nerve growth factor induction of nerve growthŽ .factor receptor mRNA in PC12 cells, J. Neurosci. Res. 31 1992
52–57.w x16 E. Gizang-Ginsberg, E.B. Ziff, Nerve growth factor regulates tyro-
sine hydroxylase gene transcription through a nucleoprotein complexŽ .that contains c-fos, Genes Dev. 4 1990 477–491.
w x17 N.J. Grant, T. Caludepierre, D. Aunis, K. Langley, Glucocorticoidsand nerve growth factor differentially modulate cell adhesion
Ž .molecule L1 expression in PC12 cells, J. Neurochem. 66 19961400–1408.
w x18 L.A. Greene, A.S. Tischler, Establishment of a noradrenergic clonallines of rat adrenal pheochromocytoma cells which respond to nerve
Ž .growth factor, Proc. Natl. Acad. Sci. U. S. A. 73 1976 2424–2428.w x19 A.V. Hall, H. Antoniou, Y. Wang, A.H. Cheung, A.M. Arbus, S.L.
Olson, W.C. Lu, C.-L. Kau, P.A. Marsden, Structural organizationŽ .of the human neuronal nitric oxide synthase gene NOS1 , J. Biol.
Ž .Chem. 269 1994 33082–33090.w x20 T. Heinemeyer, E. Wingender, I. Reuter, H. Hermjakob, A.E. Kel,
O.V. Kel, E.V. Ignatieva, E.A. Ananko, O.A. Podkolodnaya, F.A.Kolpakov, L. Kolpakov, P.N. Kolpakov, N.A. Kolchanov, Databaseson transcriptional regulation: TRANSFAC, TRRD, and COMPEL,
Ž .Nucleic Acids Res. 26 1998 364–370.w x21 D.T. Hess, S.I. Patterson, D.S. Smith, J.H.P. Skene, Neuronal
growth cone collapse and inhibition of protein fatty acylation byŽ .nitric oxide, Nature 366 1993 562–565.
w x22 E.M. Holtzman, J. Kilbridge, D.S. Bredt, S.M. Black, Y. Li, D.O.Clary, L.F. Reichardt, W.C. Mobley, NOS induction by NGF inbasal forebrain cholinergic neurons: evidence for regulation of brain
Ž .NOS by a neurotrophin, Neurobiol. Dis. 1 1994 51–60.w x23 R. Hromas, S.J. Collins, D. Hickstein, W. Raskind, L.L. Deaven, P.
O’Hara, F.S. Hagen, K. Kaushansky, A retinoic acid-responsivehuman zinc finger gene, MZF-1, preferentially expressed in myeloid
Ž .cells, J. Biol. Chem. 266 1991 14183–14187.w x24 T. Karlsson, K. Kullander, M. Welsh, The Src homology 2 domain
protein Shb transmits basic fibroblast growth factor- and nervegrowth factor-dependent differentiation signals in PC12 cells, Cell
Ž .Growth Differ. 9 1998 757–766.

( )T.K. Rife et al.rMolecular Brain Research 75 2000 225–236236
w x25 S.D. Kraner, J.A. Chong, H.J. Tsay, G. Mandel, Silencing the typeII sodium channel gene: a model for neural-specific gene regulation,
Ž .Neuron 9 1992 37–44.w x26 M.A. Lee, L. Cai, N. Hubner, Y.A. Lee, K. Lindpainter, Tissue- and
development-specific expression of multiple alternatively splicedtranscripts of rat neuronal nitric oxide synthase, J. Clin. Invest. 100Ž .1997 1507–1512.
w x27 Y.C. Li, D. Beard, S. Hayes, A.P. Young, A transcriptional enhancerof the glutamine synthetase gene that is selective for retinal Muller¨
Ž .glial cells, J. Mol. Neurosci. 6 1996 169–183.w x28 M.O. Lopez-Figueroa, K. Itoi, S.J. Watson, Regulation of nitric
oxide synthase messenger RNA expression in the rat hippocampusŽ .by glucocorticoids, Neuroscience 87 1998 439–446.
w x29 T. Maniatis, E.F. Fritsch, J. Sambrook, Molecular Cloning: a Labo-ratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor,NY, 1982.
w x30 C.J. Marshall, Specificity of receptor tyrosine kinase signaling:transient veresus sustained extracellular signal-regulated kinase acti-
Ž .vation, Cell 80 1995 179–185.w x31 K.E. Mayo, R.D. Palmiter, Glucocorticoid regulation of the mouse
metallothionein I gene is selectively lost following amplification ofŽ .the gene, J. Biol. Chem. 257 1982 3061–3067.
w x32 N. Mori, C. Schoenherr, D.J. Vandenbergh, D.J. Anderson, Acommon silencer element in the SCG10 and type II Naq channelgenes binds a factor present in nonneuronal cells but not in neuronal
Ž .cells, Neuron 9 1992 45–54.w x33 S.K. Nordeen, Luciferase reporter gene vectors for analysis of
Ž .promoters and enhancers, BioTechniques 6 1988 454–457.w x34 S.K. Nordeen, B.J. Suh, B. Kuhnel, C.A.I. Hutchison, Structural¨
determinants of a glucorticoid receptor recognition element, Mol.Ž .Endocrinol. 4 1990 1866–1873.
w x35 P.A. Norton, J.M. Coffin, Bacterial b-galactosidase as a marker ofRous sarcoma virus gene expression and replication, Mol. Cell. Biol.Ž .5 1985 281–290.
w x36 N. Peunova, G. Enikolopov, Nitric oxide triggers a switch to growthŽ .arrest during differentiation of neuronal cells, Nature 375 1995
68–73.w x37 W. Poluha, C. Schonhoff, K. Harrington, M.B. Lachyankar, N.E.
Crosbie, D.A. Bulseco, A.H. Ross, A novel, nerve growth factor-activated pathway involving nitric oxide, p53, and p21WAF1 regu-lates neuronal differentiation of PC12 cells, J. Biol. Chem. 273Ž .1997 24002–24007.
w x38 S. Prezedborski, V. Jackson-Lewis, R. Yokoyam, T. Shibata, V.L.Dawson, T.M. Dawson, Role of neuronal nitric oxide in 1-methyl-
Ž .4-phenyl-1,2,3,6-tetrahydropyridine MPTP -induced dopaminergicŽ .neurotoxicity, Proc. Natl. Acad. Sci. U. S. A. 93 1996 4565–4571.
w x39 L.P. Reagan, C.R. McKittrick, B.S. McEwen, Corticosterone andphenytoin reduce neuronal nitric oxide synthase messenger RNA
Ž .expression in rat hippocampus, Neuroscience 91 1999 211–219.
w x40 A.F. Samdani, T.M. Dawson, V.L. Dawson, Nitric oxide synthase inŽ .models of focal ischemia, Stroke 28 1997 1283–1288.
w x41 E.M. Schuman, D.V. Madison, A requirement for the intercellularŽ .messenger nitric oxide in long-term potentiation, Science 254 1991
1503–1506.w x42 A.M. Sheehy, Y.T. Phung, R.K. Riemer, S.M. Black, Growth factor
induction of nitric oxide synthase in rat pheochromocytoma cells,Ž .Mol. Brain Res. 52 1997 71–77.
w x43 M. Sheng, M.E. Greenberg, The regulation and function of c-fos andŽ .other immediate early genes in the nervous system, Neuron 4 1990
477–485.w x44 R.A. Sturm, G. Das, W. Herr, The ubiquitous octamer-binding
protein contains a POU domain with a homeo box subdomain,Ž .Genes Dev. 2 1988 1582–1599.
w x45 H. Thoenen, Neurotrophins and neuronal plasticity, Science 270Ž .1995 593–598.
w x46 M.A. Thompson, E. Lee, D. Lawe, E. Gizang-Ginsberg, E.B. Ziff,Nerve growth factor-induced depression of peripherin gene expres-sion is associated with alterations in proteins binding to a negative
Ž .regulatory element, Mol. Cell. Biol. 12 1992 2501–2513.w x47 Y. Wang, P.A. Marsden, Nitric oxide synthases: gene structure and
Ž .regulation, Adv. Pharmacol. 34 1995 71–90.w x48 B. Wasylyk, J. Hagman, A. Gutierrez-Hartmann, Ets transcription
factors: nuclear effectors of the Ras–MAP-kinase signaling pathway,Ž .Trends Biochem. Sci. 23 1998 .
w x49 B. Wasylyk, S.H. Hahn, A. Giovane, The Ets family of transcriptionŽ .factors, Eur. J. Biochem. 211 1993 7–18.
w x50 C.M. Weber, B.C. Eke, M.D. Maines, Corticosterone regulates hemeoxygenase-2 and NO synthase transcription and protein expression
Ž .in rat brain, J. Neurochem. 63 1994 953–962.w x51 J.R. Weber, J.H.P. Skene, Identification of a novel repressive ele-
ment that contributes to neuron-specific gene expression, J. Neu-Ž .rosci. 17 1997 7583–7593.
w x52 C. Wilson, H.G. Bellen, W.J. Gehring, Position effects on eukary-Ž .otic gene expression, Annu. Rev. Cell Biol. 6 1990 679–714.
w x53 J. Xie, P. Roddy, T.K. Rife, F. Murad, A.P. Young, Two closelylinked but separable promoters for human neuronal nitric oxidesynthase gene transcription, Proc. Natl. Acad. Sci. U. S. A. 92Ž .1995 1242–1246.
w x54 R.D. York, H. Yao, T. Dillon, C.L. Ellig, S.P. Eckert, E.W. Mc-Cleskey, P.J. Stork, Rap1 mediates sustained MAP kinase activation
Ž .induced by nerve growth factor, Nature 392 1998 622–626.w x55 A.P. Young, F. Murad, H. Vaessin, J. Xie, T.K. Rife, Transcription
of the human neuronal nitric oxide synthase gene in the CNS isŽ .mediated by multiple promoters, in: L. Ignarro, F. Murad Eds. ,
Nitric oxide: Biochemistry, Molecular Biology, and TherapeuticImplications, Academic Press, San Diego, 1995, 91–112.
w x56 J.H. Zar, Biostatistical Analysis, 3rd edn., Prentice-Hall, UpperSaddle River, NJ, 1996.
Related Documents











