UNIVERSIDAD CENTRAL “MARTA ABREU” DE LAS VILLAS UNIVERSIDAD ESTATAL AMAZÓNICA Caracterización de Moniliophthora roreri Evans et al. y evaluación de alternativas de control biológico en cacao, para la Amazonía ecuatoriana TESIS PRESENTADA EN OPCIÓN AL GRADO CIENTÍFICO DE DOCTOR EN CIENCIAS AGRÍCOLAS KARINA MARÍA ELENA CARRERA SÁNCHEZ SANTA CLARA, CUBA 2016

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

UNIVERSIDAD CENTRAL “MARTA ABREU”
DE LAS VILLAS
UNIVERSIDAD ESTATAL AMAZÓNICA
Caracterización de Moniliophthora roreri Evans et al. y evaluación de
alternativas de control biológico en cacao, para la Amazonía ecuatoriana
TESIS PRESENTADA EN OPCIÓN AL GRADO CIENTÍFICO DE
DOCTOR EN CIENCIAS AGRÍCOLAS
KARINA MARÍA ELENA CARRERA SÁNCHEZ
SANTA CLARA, CUBA
2016

UNIVERSIDAD CENTRAL “MARTA ABREU”
DE LAS VILLAS
UNIVERSIDAD ESTATAL AMAZÓNICA
Caracterización de Moniliophthora roreri Evans et al. y evaluación de
alternativas de control biológico en cacao, para la Amazonía ecuatoriana
TESIS PRESENTADA EN OPCIÓN AL GRADO CIENTÍFICO DE
DOCTOR EN CIENCIAS AGRÍCOLAS
Autora: Ing. Karina María Elena Carrera Sánchez, M.Sc.
Tutor: Prof. Tit., Ing. Manuel Rafael Díaz Castellanos, Dr. C.
SANTA CLARA, CUBA
2016

DEDICATORIA
A Dios porque sin él nada soy.
A Pedro Carrera y Yolanda Sánchez mi ejemplo de vida.
A Byron Loaiza por su infinita paciencia, mi pilar fundamental y
compañero de vida.
A Sebastián y Emilia Loaiza Carrera mis adorados hijos, por ser mi
inspiración, por soportar mi ausencia y ser mi fortaleza.
A mi hermano Marcelo Carrera por darme ánimos y sostenerme siempre.
A toda mi familia.

AGRADECIMIENTOS
A la Universidad Central Marta Abreu de Las Villas, de manera especial al cuerpo docente
y científico, quienes contribuyeron en mi formación académica.
A las autoridades y docentes de la Universidad Estatal Amazónica quienes oportunamente
respaldaron el desarrollo de mi investigación doctoral.
A mi tutor de tesis, Dr. C. Manuel Díaz Castellanos por su amistad, experiencia, paciencia
y motivación para que pueda concluir mi investigación con éxito.
A los Dr. C. Alexander Bernal, Leonides Castellanos, Cristóbal Ríos, Luis Bravo y Javier
Domínguez, por la revisión del documento científico y su gran amistad.
A la Dra. C. Novisel Veita por ayudar en el procesamiento estadístico de la investigación.
A los Dr. C. Roldan Torres y Orelvis Portal, por su guía oportuna en la identificación
molecular del presente trabajo.
A mis grandes colaboradores Daisy Changoluisa, Laura Mosquera, Lizbeth Cevallos, Tirsa
Kuja y Henrry Tuqueres por su incondicional apoyo en la realización de la investigación.
Al Dr. C. Michel Leiva Mora por sus valiosos aportes en el montaje de los experimentos.
Al Ing. Henry Navarrete por su apoyo en la interpretación de la información geográfica.
Al personal técnico y docente del Instituto Biotecnológico de Plantas por todo el apoyo y
conocimiento brindado.
A todos los pequeños productores amazónicos de cacao, por engrandecer con su trabajo
diario a mi país Ecuador, de manera especial a la Asociación KALLARI, por todo el apoyo
brindado.
A Susy, María Victoria, Diana, Lorena, Yuri, Duby, Amanda y Carmita mis entrañables
amigas y soporte, gracias por estar siempre conmigo.
A Mayrita y mi familia cubana, los llevo en mi corazón.
A todos mis familiares y amigos, quienes pusieron un granito de arena para la realización
de mi sueño.
A todos mil gracias.

Mira que te mando que te esfuerces y seas valiente;
no temas ni desmayes,
porque Jehová tu Dios estará contigo
en donde quiera que vayas.
Josué 1:9

SÍNTESIS
El presente trabajo tuvo como objetivo caracterizar aislados de Moniliophthora roreri
procedentes de la Amazonía ecuatoriana para evaluar la respuesta de genotipos de cacao y
alternativas de control biológico de la Moniliasis en plantaciones orgánicas de cacao. A
partir de mazorcas colectadas en seis fincas de producción orgánica de cacao cultivar
Nacional, ubicadas en la provincias de Napo, Pastaza y Morona Santiago, se realizaron
aislamientos del hongo patógeno, que posteriormente fueron identificados mediante
caracterizaciones culturales, morfológicas, fisiológicas, moleculares y patogénicas.
Además, se determinó la respuesta de genotipos de cacao frente a la inoculación artificial
de M. roreri en ensayo monocíclico, utilizando mazorcas sanas y cinco genotipos de cacao
inoculados con una mezcla de los aislados más agresivos; se evaluaron las variables
Severidad Interna, Severidad Externa, Índice de Infección, Área Bajo la Curva del Progreso
de la Enfermedad, y susceptibilidad de los genotipos. Finalmente se evaluaron in vitro e in
vivo alternativas de control biológico de la Moniliasis del cacao mediante la determinación
del Porcentaje de Inhibición del Crecimiento Radial, y la eficacia técnica de aislados de
Trichoderma spp. caracterizados cultural y molecularmente. Como resultado, se obtuvieron
20 aislados de M. roreri cuyas características culturales, morfológicas, fisiológicas,
moleculares y patogénicas se correspondieron con la especie. Todos los genotipos de cacao
inoculados artificialmente con M. roreri, manifestaron los síntomas característicos de la
Moniliasis en mazorcas, siendo el cultivar Nacional el más susceptible. Los mejores
resultados en cuanto al efecto in vitro e in vivo de los tratamientos con Trichoderma spp.
sobre M. roreri se obtuvieron con los aislados UEA-Th1 (Trichoderma harzianum) y UEA-
Tv3 (Trichoderma viride) este último mostró la mayor eficacia técnica en el control de
M. roreri en condiciones de campo.

INDICE
INTRODUCCIÓN .................................................................................................................. 1
2. REVISIÓN BIBLIOGRÁFICA ...................................................................................... 6
2.1. Origen, importancia y clasificación del cacao................................................................6
2.2 Descripción botánica del cacao...................................................................................... 9
2.3 Variedades de cacao......................................................................................................11
2.4 Principales enfermedades fúngicas del cultivo............................................................13
2.4.1 Escoba de bruja ..................................................................................................... 14
2.4.2 Mal de machete ..................................................................................................... 14
2.4.3 Mazorca negra ....................................................................................................... 15
2.4.4 Moniliasis .............................................................................................................. 15
2.5. Área bajo la curva del progreso de la enfermedad (ABCPE)......................................30
3. MATERALES Y MÉTODOS ..................................................................................... 31
3.1 Caracterización cultural, morfológica, fisiológica, molecular y patogénica de aislados
de M. roreri...........................................................................................................................32
3.1.1 Caracterización cultural de M. roreri ..................................................................... 32
3.1.2 Caracterización morfológica de M. roreri .............................................................. 34
3.1.3. Caracterización fisiológica de M. roreri ................................................................. 34
3.1.4. Identificación molecular de aislados de M. roreri .................................................. 37
3.1.5. Caracterización patogénica y agresividad de los aislados de M. roreri .................. 39
3.2 Respuesta de genotipos de cacao frente a M. roreri en un ensayo monocíclico.........41
3.3 Caracterización cultural y molecular de Trichoderma spp. procedentes de
plantaciones de cacao orgánico............................................................................................. 42
3.4 Evaluación de la actividad antagónica in vitro de Trichoderma spp. frente a
M. roreri................................................................................................................................44
3.5. Evaluación de la eficacia de aislados nativos de Trichoderma spp. y biopreparados
comerciales en el control de la Moniliasis del cacao en condiciones de campo..................48
4. RESULTADOS Y DISCUSIÓN ................................................................................ 51

4.1. Caracterización cultural, morfológica, fisiológica, molecular y patogénica de aislados
de M. roreri........................................................................................................................... 56
4.1.1. Caracterización cultural de M. roreri ....................................................................... 56
4.1.2. Caracterización morfológica de M. roreri ................................................................ 60
4.1.3. Caracterización fisiológica de M. roreri ................................................................... 63
4.1.4. Identificación molecular de aislados de M. roreri .................................................... 67
4.1.5. Caracterización patogénica y agresividad de los aislados de M. roreri .................... 69
4.2. Respuesta de genotipos de cacao frente a M. roreri en un ensayo monocíclico...........73
4.3. Caracterización cultural y molecular de Trichoderma spp. procedentes de plantaciones
de cacao orgánico..................................................................................................................82
4.4. Evaluación de la actividad antagónica in vitro de Trichoderma spp. frente a
M. roreri............................................................................................................................... 86
4.5. Evaluación de la eficacia de aislados nativos de Trichoderma spp. y biopreparados
comerciales en el control de la Moniliasis del cacao en condiciones de campo...................92
5. CONCLUSIONES ........................................................................................................ 98
6. RECOMENDACIONES ............................................................................................... 99
7. REFERENCIAS BIBLIOGRÁFICAS
ANEXOS

Introducción

Introducción
1
1. INTRODUCCIÓN
El cacao (Theobroma cacao L.) es una planta oriunda de los trópicos húmedos de América
del Sur, cuyo centro de origen se ubica en el noroeste, en la zona alta amazónica (Enríquez,
2010).
Desde 1830 Ecuador produce cacao fino de aroma, con el uso del cultivar Nacional, que
ocupa aproximadamente 243 146 ha, de las cuales solo se manejan 2 294 ha. Ecuador
exporta anualmente entre 250 000 y 300 000 t, por lo que este cultivo constituye una de las
principales fuentes generadoras de divisas y trabajo. El año 2015 cerró las exportaciones
con 260 000 t en productos semielaborados equivalentes a su peso en gramos de cacao, de
esta cifra el 87% corresponde a granos, 12% a productos semielaborados y 1% a productos
terminados de cacao. Según el Servicio Mensual de Estadísticas de Exportación de cacao en
Ecuador, en la última década se ha incrementado en un 110% la exportación y producción
de cacao. Para 2016 se piensa cerrar con 300 000 t de granos elaborados de cacao
(ANECACAO, 2015a; ANECACAO, 2015b).
En Ecuador se produce cacao en 16 provincias de las 24 que posee, la mayor producción se
encuentra localizada en la región Litoral, Amazonía y en aquellas ubicadas en las
estribaciones de la Cordillera de los Andes (INIAP, 2012). De la superficie nacional, el
8,86% corresponde a la región norte de la Amazonía con 44 300 ha, de las cuales el 83% de
la superficie corresponde a cacao cultivar Nacional y el restante 17% a otros tipos de
cacaos trinitarios (Moreno y Flores, 2011).
El cacao constituye uno de los principales rubros desde el inicio de la colonia en la Región
Amazónica Ecuatoriana (RAE) por su importancia en la generación de ingresos para las
familias de los productores; sin embargo, el cultivo no está exento de problemas
fitosanitarios como: Moniliasis (Moniliophthora roreri Evans et al.), Mazorca negra
(Phytophthora palmivora (E.J. Butler)) y Escoba de bruja (Moniliophthora perniciosa
Aime & Phillis-Mora), los cuales constituyen los principales factores limitantes de la
producción y donde el manejo integrado del cultivo es la estrategia más exitosa
(INIAP, 2012).

Introducción
2
La Moniliasis es una de las enfermedades más destructivas del cacao, registrada en 11
países latinoamericanos (Phillips-Mora, 2006a). La severidad del ataque del hongo
patógeno varía según la zona y época del año, de acuerdo con las condiciones climáticas
que prevalezcan (PROAMAZONÍA, 2003). Sánchez-Mora et al. (2011) informaron que en
Ecuador las pérdidas pueden llegar hasta un 60%.
Frente a este panorama, se han llevado a cabo varios estudios sobre el organismo patógeno
con el fin de identificar su origen, formas de diseminación, diversidad genética y
caracterización de la Moniliasis. Evans et al. (1978), realizaron estudios de la taxonomía de
M. roreri, y describieron los caracteres morfológicos de este hongo fitopatógeno, lo cual
constituyó la base fundamental para su estudio y clasificación.
Phillips-Mora (2003), en su trabajo Origen, biogeografía, diversidad genética y taxonomía
de M. roreri, realizó una descripción de aislados procedentes de Centroamérica, Venezuela,
Colombia, Ecuador y Perú, cultivó el hongo fitopatógeno en medio V8 modificado a 20 ºC
durante 20 días, y determinó que la velocidad de crecimiento entre los aislados de los
distintos países fue diferente. Las esporas variaron respecto a su forma (globosa,
subglobosa y elipsoidal).
Villavicencio (2010), por vez primera en Ecuador, caracterizó morfológica, fisiológica y
patogénicamente aislados de M. roreri en cinco provincias de la costa ecuatoriana, donde
determinó que no se encontraron diferencias en cuanto al borde y la textura de las colonias.
El color de las colonias varió en dependencia de la edad del cultivo. Se presentaron formas
de esporas globosas, subglobosas y elipsoidales. No existió relación entre el crecimiento de
las colonias y la patogenicidad para la mayoría de los aislados, aunque en algunos de ellos
se observó relación entre esas variables.
Según INIAP (2012), el Manejo Integrado de las Enfermedades de Cacao comienza con la
aplicación combinada de varias prácticas: resistencia genética del material de siembra,
prácticas culturales (adecuado balance nutricional, control oportuno de malezas, podas de
mantenimiento y sanitarias, remoción de frutos enfermos, deschuponado), control biológico

Introducción
3
con el uso de agentes antagonistas (Trichoderma spp.), y control químico con productos de
baja toxicidad.
En los últimos años existe un cambio de hábito en los consumidores de cacao, quienes
prefieren alimentos sanos, seguros, orgánicos, étnicos y productos de especialidad, y debido
a esto cada vez es más creciente la demanda de cacao de un sabor y origen determinados.
En Ecuador, y en particular en la Amazonía, el cacao posee un aroma y gusto especiales,
una historia, una organización social específica, lo que ha puesto en altos sitiales la
producción de cacao orgánico amazónico (Isla y Andrade, 2009; FAO, 2010; INIAP,
2011).
La Región Amazónica Ecuatoriana (RAE) comprende las provincias de Orellana, Pastaza,
Napo, Sucumbíos, Morona Santiago y Zamora Chinchipe. En ellas la temperatura media
anual es de 24 y 25 °C. Las lluvias se distribuyen uniformemente durante todo el año, por
lo cual se considera una de las zonas más lluviosas con totales anuales que fluctúan entre
los 3 000 a 4 000 mm y 88% de humedad relativa (INAMHI, 2014).
Según Matamoros (2007), el ecosistema amazónico, en especial el bosque tropical lluvioso,
se extiende sobre un área de 123 000 km² de exuberante vegetación, propia de los bosques
húmedo-tropicales, lo que representa el 45% del territorio nacional. La RAE contiene una
importante variedad de ecosistemas, por lo cual se considera como uno de los hábitats
vegetales y animales más ricos y complejos del mundo, donde alberga aproximadamente el
80% de la biodiversidad del país.
Pérez (2013) señaló que las comunidades indígenas amazónicas manejan sus cultivos
mediante el sistema de chakra, sistema que asegura el sustento familiar, genera excedentes
y rescata valores del Sumak Kawsay (Buen Vivir). Asimismo, es fuente de valores sociales,
económicos, culturales y ambientales. El valor de las tradiciones, de la medicina ancestral,
de las artesanías, el entorno, que no explota a la gente. Las prácticas agrícolas en las
comunidades son sustentables y amigables con el medio ambiente. Preparan el suelo,
cultivan, cosechan y mantienen el suelo rotando e intercalando con especies que incorporan

Introducción
4
elementos nitrogenados al suelo aumentando su fertilidad. En el proceso de cultivo utilizan
solo sus manos y machetes, y así evitan dañar la superficie cultivable.
Las plantas cultivadas en su área incluyen: maíz, plátano, papaya, frijol, piña, papa,
malanga, café, cacao, palmas, árboles frutales, madera dura y plantas de uso medicinal y
artesanal. Estas prácticas se han visto afectadas por plagas y enfermedades lo que reduce
significativamente la economía de los productores, debido a las pérdidas que ocasionan,
especialmente la Moniliasis (INIAP, 2011).
La Amazonía del Ecuador reviste gran importancia en la producción de cacao orgánico, sin
embargo no se ha caracterizado la Moniliasis, enfermedad responsable de las pérdidas
económicas para los productores, ni se dispone de alternativas de control biológico de la
misma con aislados nativos de antagonistas para el cultivar Nacional.
Con todas las consideraciones, el presente trabajo se propone la siguiente hipótesis:
La caracterización de aislados de Moniliophthora roreri procedentes de la Amazonía
ecuatoriana, la evaluación de los niveles de resistencia de los diferentes genotipos al
patógeno y la caracterización de aislados nativos de antagonistas, posibilitará disponer de
nuevas alternativas de control biológico de la Moniliasis en plantaciones orgánicas de
cacao. Para comprobar esta hipótesis, se plantean como objetivos los siguientes:
Objetivo general
Determinar las características de los aislados de Moniliophthora roreri, la respuesta de
genotipos de cacao y la eficacia del control biológico contra la Moniliasis en plantaciones
orgánicas de cacao en la Amazonía ecuatoriana.
Objetivos específicos
1. Caracterizar cultural, morfológica, fisiológica, molecular y patogénicamente
aislados de M. roreri.
2. Evaluar la respuesta de genotipos de cacao frente a la inoculación artificial de
M. roreri, en un ensayo monocíclico.

Introducción
5
3. Caracterizar cultural y molecularmente aislados de Trichoderma spp. procedentes
de plantaciones de cacao orgánico.
4. Evaluar la actividad antagónica in vitro de Trichoderma spp. frente a M. roreri.
5. Evaluar la eficacia de aislados nativos de Trichoderma spp. y biopreparados
comerciales en el control de la Moniliasis del cacao en condiciones de campo.
Este estudio presenta NOVEDAD CIENTÍFICA, puesto que por primera vez en Ecuador se
cuenta con un banco de 20 aislados de M. roreri, de la Amazonía ecuatoriana, debidamente
caracterizados. Asimismo, se logró a través de un ensayo monocíclico utilizar el Área Bajo
la Curva del Progreso de la Enfermedad (ABCPE) para determinar la respuesta diferencial
de aislados de M. roreri, al usar como unidad experimental mazorcas de cacao cultivar
Nacional. Se caracterizó por vez primera a Trichoderma harzianum Rifai y Trichoderma
viride Pers ex S.F Gray. en condiciones amazónicas vinculadas a cacao, y se demostró su
efectividad in vitro y en campo, frente a este hongo fitopatógeno.
Desde el punto de vista PRÁCTICO se pone a disposición de la comunidad científica una
colección de cultivo de M. roreri, así como tres cepas nativas de Trichoderma
adecuadamente identificadas y caracterizadas. Asimismo, se logró demostrar la efectividad
de T. harzianum y T. viride sobre M. roreri, lo cual ayudará a reducir en campo las pérdidas
ocasionadas por este hongo fitopatógeno.
Este trabajo además tiene IMPORTANCIA TEÓRICA puesto que contribuye a un mejor
entendimiento de la interacción M. roreri-Trichoderma spp. en condiciones in vitro y
campo, y debido a su valor metodológico, podrá usarse para futuras investigaciones con
hongos fitopatógenos y sus interacciones.

2
Revisión Bibliográfica

Revisión Bibliográfica
6
2. REVISIÓN BIBLIOGRÁFICA
2.1. Origen, importancia y clasificación del cacao
Algunos autores indican que el cultivo del cacao se inició en México y América Central y
señalan al mismo tiempo que los españoles no lo vieron cultivado en América del Sur
cuando arribaron al continente, aunque lo encontraron en forma natural en muchos bosques
a lo largo de los ríos Amazonas, Orinoco y sus afluentes, donde aún hoy existen tipos
genéticos de mucho valor (Batista, 2009).
En México se encontró que los aztecas usaban las almendras del cacao no sólo para la
preparación de bebidas sino también como moneda. Creían que el árbol del cacao era de
origen divino y que su bebida confería discreción y sabiduría. Por eso Linneo asignó a la
especie el nombre de Theobroma, que significa alimento de los dioses. En los tiempos de
Cristóbal Colón, los mayas eran los verdaderos cultivadores de cacao; perfeccionaron su
cultivo, aprendieron a curar y conservar las semillas y a hacer una bebida agradable. Ricos
y pobres consumían la bebida en su dieta diaria y comerciaban el producto con los aztecas,
quienes muy pronto llegaron a apreciar sus cualidades (Batista, 2009; Jaimes y Aranzazu,
2010).
El cultivo del cacao tiene su origen en el noreste de América del Sur (Alto Amazonas)
luego se dispersó hacia el norte por la costa pacífica que comprende países como Colombia,
Ecuador, Perú y Brasil, donde se ha encontrado una alta variabilidad genética. Desde este
lugar de origen, las especies se fueron difundiendo y evolucionando en dos grupos de cacao
con características fenotípicas y genotípicas bien definidas, las cuales corresponden a los
cacaos Criollo y Forastero (Enríquez, 2010).
Desde la colonización española, el cultivo se ha extendido a otros países, que hoy son
líderes en la producción mundial; se cultiva principalmente en 13 países, como Costa de
Marfil, Ghana, Nigeria y Camerún en el continente africano, con cerca del 60% de la
cosecha mundial, que significa 1,6 millones t. En el continente americano, el mayor

Revisión Bibliográfica
7
productor es Brasil con el 18%; le siguen Ecuador con el 6%, Colombia y México
contribuyen con el 1% de la producción mundial. Se estima que más de 20 millones de
personas dependen directamente de este cultivo para subsistir y que el 90% de la
producción se cosecha en minifundios con menos de 5 ha (Ramírez, 2008).
Ecuador es el primer productor de cacao fino de aroma a nivel mundial, con
aproximadamente el 62% del volumen global; por lo tanto, es esencial preservar este legado
cultural, social y económico. Además, se considera como el producto de exportación con
mayor historia en la economía de Ecuador, que involucra alrededor de 100 000 familias de
pequeños y medianos productores. Se le reconoce a nivel mundial por sus marcadas
características de aroma y color, sumamente apreciadas en la preparación de chocolates
finos (ACEPROCACAO, 2012).
La región amazónica constituye uno de los principales centros de origen del cacao. Se
encuentra de manera silvestre en varias zonas de las seis provincias amazónicas de
Ecuador, donde las precipitaciones fluctúan entre 2 000 y 4 000 mm/año, en altitudes de
200 a 1 100 msnm, temperaturas entre 19 a 25 °C y luminosidad entre 1 050 a 1 350
horas/luz. En suelos de los tipos Inseptisoles, Ultisoles y Entisoles, cuya producción está
vinculada a especies maderables y frutales que aportan sombra necesaria para el sistema
agroforestal (chakra), lo cual constituye un valor agregado en los mercados que valoran las
contribuciones ambientales que genera la producción de cacao (ECORAE, 2001; Moreno y
Flores, 2011; Ordoñez et al., 2011; Pérez, 2013).
Con el propósito de conocer el lugar de origen del cacao Nacional se han realizado
investigaciones a través de la comparación de huellas genéticas de ADN del material nativo
con los representantes de la variedad nativa Nacional, lo cual demostró el gran parentesco
existente con algunos árboles colectados hace más de 30 años en la región amazónica de
Zamora Chinchipe, en el sur de Ecuador. A pesar de que en la actualidad su cultivo se
encuentra más desarrollado en la región costera del Pacífico, la región oriental de Zamora
Chinchipe resulta ser el centro de origen de la domesticación probable del cultivar Nacional
(ANECACAO, 2015a).

Revisión Bibliográfica
8
En Ecuador existen 500 000 ha de cacao, la mayoría asociadas con otras especies,
establecidas en más de 100 000 fincas, principalmente de pequeños productores de diversas
culturas de las nacionalidades indígenas de la Amazonía, siendo el tercer rubro agrícola
más importante del país (ANECACAO, 2015c).
El valor de la producción de cacao en el año 2008 aumentó de USD (dólares americanos)
94,3 millones a 229 millones en el año 2013, es decir existió un alza de USD 205 millones
en la producción global, casi el equivalente al valor de la producción anual de Brasil; la
“pepa de oro” obedece también a los buenos precios que se pagan en el mercado
internacional. Durante el año 2012 el precio promedio por tonelada fue de USD 2 392, en el
2013 fue de USD 2 439 y en el 2014 este precio subió a USD 3 126, según datos de Inside
Future, y para el segundo semestre de 2015 fue de USD 3 165 (ANECACAO, 2015c).
El cacao básicamente se produce en las regiones Costa y Amazonía de Ecuador,
principalmente en las provincias de Los Ríos, Guayas, Manabí y Sucumbíos, donde se
cultivan dos tipos de cacao: cacao Nacional y cacao CCN-51 (Colección Castro Naranjal,
este último producto de un injerto de varios clones, realizado por el ecuatoriano Homero
Castro en el año 1965 con el objetivo de combatir plagas), aproximadamente 500 000 ha,
de las cuales 110 000 ha se encuentran en manos de pequeños agricultores, con parcelas no
mayores a 5 ha. Este producto representó el 7% del Producto Interno Bruto (PIB) del país,
lo que benefició a pequeños y medianos productores agrícolas (ANECACAO, 2015b).
La clasificación botánica más aceptada para el cacao según Ayala (2008) es:
Reino: Plantae
Subreino: Tracheobionta
División: Magnoliophyta
Clase: Magnoliopsida
Subclase: Dilleniidae
Orden: Malvales
Familia: Sterculiaceae
Subfamilia: Byttnerioideae

Revisión Bibliográfica
9
Género: Theobroma
Especie: T. cacao L.
2.2 Descripción botánica del cacao
El cacao es una especie diploide (2n=20 cromosomas). Es alógama, ya que su polinización
es cruzada, ubicándose por encima del 95%. Crece y se desarrolla usualmente bajo sombra
en los bosques tropicales húmedos de América del Sur (Enríquez, 2010). Normalmente
alcanza alturas de 6 a 8 m, con excepción del cacao Nacional de Ecuador y el amelonado
de África Occidental que alcanzan alturas de hasta 12 m (Batista, 2009).
Según Enríquez (2010) en la primera fase de crecimiento es ortotrópico (vertical), que
perdura por 12-15 meses. Luego este crecimiento se interrumpe para dar lugar a la
formación de 4-5 ramas secundarias (“horqueta”), que son de crecimiento plagiotrópico
(horizontal), el cual puede variar su ángulo de desarrollo. Este tipo de árbol alcanza hasta
4 m de altura.
Raíz: Tiene una raíz principal pivotante, la cual sirve de medio de anclaje; especialmente
en los primeros meses de vida de la planta puede crecer hasta 200 cm, el sistema de raíces
secundarias absorbe los nutrientes y agua disponibles en el suelo. Su longitud y forma
varían mucho de acuerdo principalmente con la estructura, textura y consistencia del suelo
(Hardy, 1961).
Hojas: Las hojas adultas son simples y enteras, membranosas de tamaño medio, base
obtuso-atenuada, margen entero, ápice largamente acuminado, coriáceas de color verde.
Cuando jóvenes su coloración es variable de verde pálido a café claro, hasta rojizas. Las
hojas son caducas, cada dos o tres meses se presentan picos de brotación de nuevas hojas,
que reemplazan a las que se caen (Rondón y Cumana, 2005).
Flores: Las flores de cacao son órganos complejos ya que poseen características que las
hacen únicas, los órganos de reproducción son realmente importantes el momento de

Revisión Bibliográfica
10
estimar la diferenciación genotípica de los clones (Bartley, 2005). Estas producen y
desarrollan cojinetes florales, de color rosado, hermafroditas, de 10-20 mm de largo,
pentámeras; es decir que están constituidas de cinco sépalos, pétalos, estaminodios,
estambres, y lóculos por ovario, que crecen en simetría radiada. Ovario 2-3 mm de largo,
oblongo-ovoide, pentagonal, pubescente. Las flores tienen una viabilidad de hasta dos días;
luego de lo cual si no son fecundadas caen (Rondón y Cumana, 2005; Enríquez, 2010).
Frutos: Los frutos más conocidos como mazorcas, son drupas bastante grandes que pueden
variar ampliamente de forma, espesor, rugosidad, color y tamaño, según el origen genético,
el medio ambiente donde crece y se desarrolla el árbol, así como el manejo de la
plantación. Se observa toda una gama de colores, que en estado inmaduro va de tonos
verdes, rojizos y cafés; con surcos y lomos pigmentados, mientras que en estado maduro
varían de amarillo, café amarillento a rojizo anaranjados. Las mazorcas de cacao por su
forma están clasificadas como: amelonado, calabacillo, angoleta y cundeamor, variando
según el genotipo (Batista, 2009).
Según Enríquez (2010), las semillas presentan una forma oblonga y varían mucho de
tamaño según el tipo de cacao, tal es el caso del cacao Criollo y Nacional de Ecuador;
otros son aplanados en el caso de los Forasteros. La cantidad de semillas en cada mazorca
depende del número de óvulos en cada ovario. Tienen un recubrimiento o cutícula que
protege los cotiledones. En la parte exterior se encuentra el mucílago, que es la pulpa dulce
de la semilla, lo que permite diferenciar algunos genotipos de cacao. El color de la semilla
también varía de acuerdo con el genotipo, desde blanco ceniciento, blanco puro, hasta un
morado oscuro y todas las tonalidades intermedias. El color interno de las almendras es
violeta pálido o lila, aunque en algunas ocasiones se observan semillas blancas.
La semilla está constituida por dos cotiledones que contienen grasa, alcaloides, taninos y
otras sustancias que al alterarse dan origen al sabor y olor del chocolate manufacturado.
Las semillas de cacao no requieren de reposo para germinar, mueren al poco tiempo

Revisión Bibliográfica
11
cuando sufren deshidratación. El tiempo desde la fecundación hasta la madurez fisiológica
de una mazorca es de alrededor de 180 días (Hardy, 1961).
2.3 Variedades de cacao
Ecuador es uno de los países donde existe la mayor diversidad genética de T. cacao. Por su
origen y características genéticas, en cada zona específica se desarrollaron cultivares
diferentes de cacao (Ayala, 2008; Paredes, 2009).
Según Paredes (2009), el cacao está clasificado en cuatro variedades:
Criollo. Son árboles bajos y poco robustos, su copa es de color verde, gruesa, redonda con
hojas pequeñas, y sus almendras son de color blanco marfil con sabor a nuez y fruta. La
principal característica de este tipo de cacao está en la mazorca que es alargada, de colores
verde y rojizo en estado inmaduro, y amarillo y anaranjado al alcanzar la madurez (Figura
1a). El cacao Criollo ha sido domesticado y adaptado a diferentes regiones del planeta, lo
que hace que sea más delicado y de poca productividad, y por ende más susceptible a
enfermedades.
Forastero amazónico. Se les conoce como amazónicos al encontrarse distribuidos en la
cuenca del río Amazonas y sus afluentes. Las mazorcas en estado inmaduro son de colores
verde y amarillo cuando han alcanzado su madurez, presentan forma de cuello de botella,
las almendras son aplanadas con cotiledones de color morado (Figura 1b). De este tipo de
cacao se obtiene chocolate con sabor básico de cacao que proporciona el 80% de la
producción mundial.
Trinitario. Es un resultado del cruce entre cacao Criollo de Trinidad (de ahí deriva su
nombre) y Forastero multiplicado en la cuenca del río Orinoco. Este tipo de cacao abastece
del 10 al 15% de la producción mundial. Es más resistente a enfermedades y ha podido
adaptarse mejor a muchos ambientes, presenta sabor a cacao de medio a alto, usualmente
asociado además con sabor a frutas y nueces. Este tipo de cacao es el que más se cultiva en

Revisión Bibliográfica
12
Ámérica, presenta diversas formas intermedias de mazorcas al igual que su coloración
desde tonos verdes y rojizos, e incluso una mezcla de ambos (Figura 1c).
Cacao Nacional de Ecuador. Es el único grupo natural de cacao que se cultiva en el
occidente de Ecuador. Por su fina calidad de almendra, este grupo está más relacionado con
el grupo Criollo que con el grupo Forastero. Las mazorcas son amelonadas, presentan
estrangulaciones en el ápice y la base, con surcos y lomos poco profundos, el color interno
de las almendras es violeta pálido o lila, aunque también se pueden observar semillas
blancas (Figura 1d). De este tipo de cacao se obtienen los mejores chocolates del mundo
por su sabor y aroma floral.
Figura 1. Tipos de cacao a) Criollo, b) Forastero, c) Trinitario y d) Nacional. Fuente:
Paredes, 2009
Entre los principales clones que se producen en Ecuador según INIAP (2000) y García
(2010), se encuentran:
CCN-51. Pertenece al Grupo genético (IMc-67 x ICS-95) x forastero desconocido; este
tipo de cacao actualmente cubre parte de las plantaciones de la Amazonía ecuatoriana. Las
mazorcas en estado inmaduro son de color rojizo-morado y maduras son rojizo-anaranjado.
Moderadamente susceptible a la Moniliasis. Tienen sabor a cacao de medio a bajo, el
potencial de esta variedad es la manteca de cacao, con rendimiento de semilla de 2 760 kg
ha-1 (Figura 2a).
EET-95. Pertenece al Grupo genético Nacional x desconocido, cacao que se desarrolla en
suelos franco-arcilloso-limosos con buen drenaje y ricos en materia orgánica.
a
b
c
d

Revisión Bibliográfica
13
Moderadamente susceptible a la Moniliasis. Las mazorcas se caracterizan por tener forma
de cuello de botella, en estado inmaduro son de color verde y en la madurez de color
amarillo. De calidad arriba. El rendimiento de semilla es de 1 564 kg ha-1 (Figura 2b).
EET-96. Grupo genético Nacional x Venezolano amarillo, se adapta a una temperatura de
21 a 28 °C con poca variación entre el día y la noche, suelo franco-arcilloso-limoso con
buen drenaje. Susceptible a Moniliasis. Las mazorcas presentan una base ancha, surcos
profundos, superficie rugosa, en estado inmaduro son de color verde y maduro verde
amarillento, semillas cilíndricas. El rendimiento de semilla es de 1 656 kg ha-1 (Figura 2c).
EET-103. Grupo genético Nacional x venezolano amarillo, adaptable a una temperatura de
21 a 28 °C con poca variación entre el día y la noche, suelo franco-arcilloso-limoso con
buen drenaje. Sus frutos son amelonados; en estado inmaduro son de color verde y en la
madurez amarillo. Moderadamente susceptible a la Moniliasis. El rendimiento de semilla
es de 1 518 kg ha-1 (Figura 2d).
Figura 2. Características de las mazorcas de los clones de cacao a) CCN-51, b) EET-95,
c) EET-96 y d) EET-103. Fuente: El productor 2012
2.4 Principales enfermedades fúngicas del cultivo
El cacao es atacado por microorganismos que afectan su producción y calidad, lo cual
constituye un ambiente favorable para el desarrollo de enfermedades fúngicas, y por
consiguiente se incrementan los costos de producción (Fulton, 1989; Porras y Sánchez,
1991). En Ecuador las enfermedades del cacao causan más pérdidas al agricultor que los
insectos, hasta en un 80% (Phillips-Mora y Wilkinson, 2007). Según Jaimes y Aranzazu
a b d c

Revisión Bibliográfica
14
(2010), las principales enfermedades en los cacaotales del Ecuador son Escoba de bruja,
Mal de machete, Mazorca negra y Moniliasis.
2.4.1 Escoba de bruja
La Escoba de bruja es causada por el hongo basidiomicete Moniliophthora perniciosa
(Aime & Phillis-Mora), (Aime y Phillips-Mora, 2005) puede ocasionar pérdidas del 50 al
90% (Meinhardt et al., 2008). Se caracteriza por la proliferación de yemas apicales y
axilares en ramas de cacao a modo de escobas de bruja. El hongo afecta todos los órganos
en crecimiento activo, principalmente los brotes nuevos, cojinetes florales, flores y frutos,
en los cuales producen hipertrofias y crecimientos anormales. Según con el órgano y la
edad de la planta, cuando el hongo infecta, se manifiesta una amplia diversidad de síntomas
a los cuales se les asigna una denominación particular, lo que genera hipertrofias de los
brotes vegetativos y reproductivos que ocasionan un continuo debilitamiento del árbol. Los
rebrotes jóvenes son los más susceptibles, pues cuando son afectados, abortan (Esquivel
et al., 2012).
2.4.2 Mal de machete
Según Suárez (2008), el Mal de machete es una enfermedad que se presenta en el tronco y
las ramas de cacao, producida por el hongo Ceratocystis cacaofunesta (Engelbrecht &
Harrington), la cual es transmitida por un insecto del género Xileborus spp., que es un
coleóptero perforador del tronco, y por las herramientas sin desinfectar. Los síntomas de la
enfermedad consisten en perforaciones y la presencia de aserrín en los sitios donde hizo las
perforaciones el insecto, los cuales, por lo general, se encuentran en el tronco y las ramas
primarias del árbol. El síntoma inicial de la enfermedad es un amarillamiento de las hojas
que mueren rápidamente y quedan adheridas a las ramas, aun después de muerto el árbol.
Esta enfermedad produce una muerte súbita del árbol. Para el control, lo mejor es la
prevención, por lo que se recomienda no utilizar las herramientas que hayan sido utilizadas
en labores realizadas a plantas enfermas (Alarcón et al., 2012).

Revisión Bibliográfica
15
2.4.3 Mazorca negra
La enfermedad Mazorca negra es producida por Phytophthora sp., y se encuentra con más
frecuencia en la región de Centroamérica. Sin embargo, en otras regiones del mundo la
enfermedad la producen otras especies como Phytophthora capsici (Leonian), Phytopthora
citrophthora (R.E. Sm. & E.H. Sm.), que se presentan mayormente en algunas regiones de
América del Sur y el Caribe, y P. megakarya (Brasier & Griffin), que solo está presente en
ciertos países de África. Avances sobre el estudio de esta enfermedad en Ecuador la asocian
con Phytophthora palmivora (E.J. Butler), (Mfegue, 2012; Akrofi, 2015). Diferentes
especies de este Oomicete han sido reportadas dando lugar a daños en el cacaotero. Atacan
los frutos principalmente, y producen daños de importancia económica, ya que reducen la
producción de semillas, sin embargo, también puede afectar otras partes de los árboles
como brotes tiernos, tronco, cojinetes florales, raíces y plántulas de cacao en vivero
(Urdaneta y Delgado, 2007; Pérez, 2009).
2.4.4 Moniliasis
2.4.4.1 Origen de la Moniliasis
Durante muchos años se dijo que la Moniliasis se detectó en el año de 1916 en Ecuador
(Rorer, 1918; Ampuero, 1967; Najar y Thomas, 2001). Estudios posteriores mencionan a
Colombia como el centro de origen de esta grave enfermedad, en el año 1800. Desde
entonces se ha dispersado a 12 países de Sur y Centroamérica productores de cacao
(Colombia, Ecuador, Perú, Bolivia, Venezuela, Panamá, Costa Rica, Honduras, Guatemala,
Belice, México y El Salvador), y ha ocasionado pérdidas superiores al 90% de la
producción, y el abandono de los cultivos a lo largo del continente americano. Esta
situación ha causado efectos negativos en los cacaoteros, sus comunidades, y en los
agroecosistemas en que se produce cacao (Phillips-Mora, 2006a; Phillips-Mora et al.,
2015).

Revisión Bibliográfica
16
La Moniliasis constituye un factor muy importante de infección en las plantaciones de
cacao, debido a que el hombre es el agente más efectivo en la dispersión a largas distancias,
dado que el hongo está muy bien adaptado a este método indirecto de diseminación (Evans,
1986). Sobre las plantaciones afectadas, existe una presencia permanente de esporas de
M. roreri, suspendidas en el aire, debido al alto nivel de adaptación a diferentes ambientes,
por lo que la infección del fruto puede ocurrir en cualquier ambiente favorable (Schmitz,
1985). La regulación del microclima ha sido una de las prácticas más recomendables para
el combate de la Moniliasis (Meléndez y Somarriba, 1999).
2.4.4.2 Clasificación taxonómica, según Bioscience (2004)
Reino: Fungi
Phylum: Basidiomycota
Subclase: Agaricomycetidae
Orden: Agaricales
Familia: Marasmiaceae
Género: Moniliophthora
Especies: M. roreri
Los nombres comunes de la enfermedad, según el Instituto Dominicano de Investigaciones
Agropecuarias y Forestales (IDIAF), son: Moniliasis del cacao, Pudrición de fruto,
Pudrición acuosa del fruto, Helada, Mancha ceniza y Enfermedad de Quevedo
(IDIAF, 2012).
2.4.4.3. Etiología
El agente causal fue inicialmente llamado Monilla roreri por Ciferri y Parodi (1933) y
clasificado dentro del Phylum Ascomicota, describiéndolo como un hongo anamorfo
debido a la aparente ausencia de un estado meiótico o de estructuras sexuales, y a sus
similitudes morfológicas con otros fitopatógenos del género (Evans et al., 2003a). Sin
embargo, Evans et al. (1978), mediante estudios de microscopia electrónica, encontraron la

Revisión Bibliográfica
17
presencia de septo dolíporo (característico de hongos basidiomicetos) y un evento único de
esporogénesis basipetal, resultado que motivó la creación del nuevo género
Moniliophthora. Phillips-Mora (2003), mediante el uso de técnicas moleculares confirmó
que el hongo es un Basidiomycete. Posteriormente, Aime y Phillips-Mora (2005)
reafirmaron la ubicación de M. roreri dentro de la familia Tricholomataceae.
2.4.4.4 Ubicación de los grupos genéticos
A nivel internacional se han identificado cinco grupos genéticos principales del hongo, tres
son endémicos y están localizados en las cordilleras central y oriental de Colombia, y en
Ecuador. Los otros dos están ampliamente distribuidos y se encuentran en una fase
invasiva, uno de ellos se localiza en el Oriente de Colombia, en la periferia de Ecuador,
Venezuela y Perú; y el otro en el Occidente de Colombia, Centro de Ecuador y América
Central (Phillips-Mora et al., 2007).
Las investigaciones han permitido establecer que los síntomas varían con la edad del fruto,
pero la velocidad de desarrollo depende de las condiciones ambientales, básicamente de la
temperatura, humedad relativa, susceptibilidad del clon o variedad de cacao y agresividad
de los aislados (Porras y Sánchez, 1991).
2.4.4.5. Diseminación
Las esporas de M. roreri son los únicos propágulos infectivos que se diseminan fácilmente
por acción del viento y de la lluvia (Campuzano, 1976; Barros, 1977; Oliveira y Luz,
2005). Phillips-Mora (2006b) señaló que la época crítica para la infección de M. roreri son
los cuatro primeros meses desde la floración a la formación de frutos.
2.4.4.6. Hospedantes
M. roreri ha sido reportado solamente en especies de los géneros Theobroma y Herrania
(Jaimes y Aranzazu, 2010). Rorer (1918) reportó en Ecuador el ataque a frutos de

Revisión Bibliográfica
18
T. bicolor Humb. & Bonpl. y H. balaensis P. Preuss. Además, Evans (1981) identificó
infecciones de M. roreri en T. mammosum Cuatrec. & J. León, T. simiarum Donn. Sm.,
T. sylvestre Mart., T. angustifolium K. Schum., H. nítida (Poepp.) R.E. Schult.,
H. pulcherrima Goudot. Posteriormente se identificó el hongo patógeno en T. grandiflorum
(Willd. ex Spreng.) K. Schum. y H. purpurea (Pittier) R.E. Schult. (Cuhna, 2006).
2.4.4.7. Epifitiología
Esta enfermedad ha sido descrita en un amplio rango altitudinal entre 0 y 1 520 msnm,
donde existen precipitaciones con fluctuaciones que van desde 780-5 500 mm por año y
una temperatura de 18 a 28 °C (Phillips-Mora, 2006a). En sitios donde la pluviometría es
igual o mayor a 2 500 mm/año y la humedad relativa está sobre el 80%, la incidencia de
Moniliasis puede superar el 90% (Desrosiers et al., 1955; Evans, 1981; Porras, 1983).
Meléndez (1993) encontró que existe una estrecha relación entre la humedad relativa y el
movimiento de esporas del hongo, lo que indica que la liberación es realizada entre el 71 y
74% de humedad relativa y en un horario que va desde las 08:00-14:00 h
aproximadamente.
Según Torres (2010), las esporas permanecen en el aire durante todo el año. Las
condiciones secas, humedad relativa baja y temperatura mayor de 26 °C, favorecen su
liberación y dispersión; y las lluvias intensas y frecuentes ayudan a la presencia de agua
libre, lo que permite la penetración y germinación de las esporas en los frutos (Phillips-
Mora, 2006b).
La gran diversidad y adaptabilidad que ha demostrado el hongo patógeno, combinado con
la longevidad de sus esporas y las condiciones agroclimáticas que ofrecen los países
productores convierten a Moniliasis en un invasor formidable aun en ecosistemas donde no
se encuentre (Phillips-Mora et al., 2005).

Revisión Bibliográfica
19
2.4.4.8. Ciclo de vida del hongo patógeno
De acuerdo con Evans et al. (1978), M. roreri representó el estado mitótico (anaformo) de
un basidiomycete desconocido. Posteriormente, Evans (1981) especuló que el teleomorfo
podría ser una especie de Crinipellis, dado el inusual ciclo vital pleomórfico
hemibiotrófico, que presenta M. roreri, el cual también es observado en otro de los hongos
patógenos importantes del cacao: C. perniciosa.
Evans et al. (2002) encontraron evidencias de que la meiosis ocurre dentro de los conidios,
un fenómeno que tiene coherencia con el contenido nuclear variable de estos; por lo que
proponen que la fase reproductiva observada en M. roreri es sexual, y que este hongo
patógeno corresponde a un teleomorfo modificado.
Durante la esporogénesis y germinación ocurre la meiosis, la cual da lugar a la formación
de hifas infectivas monocarióticas. La fase parasítica haploide se da solo en el tejido vivo
(biotrófico), y en ella el hongo crece y se alimenta intercelularmente sin activar los
mecanismos de defensa del hospedante. Una señal no identificada (asociada con la edad del
fruto) estimula la transición a la fase diploide necrotrófica, la cual induce los síntomas
característicos de la Moniliasis. De acuerdo con Griffith et al. (2003), lo anterior parece
indicar que un antepasado de M. roreri perdió la capacidad de formar basidiocarpos, pero
no la capacidad de experimentar división nuclear meiótica.
La sobrevivencia del hongo patógeno empieza en los residuos de cosecha (mazorcas
contaminadas). Luego, las esporas son diseminadas por el viento y la lluvia, lo que produce
contaminación de frutos o mazorcas con Moniliasis de una plantación a otra (Navarro y
Mendoza, 2009). Oliveira y Luz (2005), plantean que la diseminación de las esporas se
realiza por el viento, y que el agua de lluvia tiene un papel importante en las infecciones a
corta distancia en la copa del cacao. Además, debido al movimiento producido por las
labores de cosecha, las esporas se dispersan en el aire, y bajo condiciones propicias de

Revisión Bibliográfica
20
humedad y temperatura, infectan los frutos que recién están formándose (Amores et al.,
2009).
Según Meléndez (1993), en su investigación sobre microambiente, la mayor cantidad de
esporas de M. roreri se encuentran a 1 m de altura en las plantas de cacao. Las esporas se
depositan sobre el fruto, germinan si hay agua, o mueren por la radiación/desecación; estos
al germinar pueden penetrar directamente en la epidermis del fruto a través de los estomas,
donde crecen entre las células del córtex, lo que produce esporas dentro y en la superficie
de los frutos. Existen dos formas de germinación: a través del tubo germinativo o
directamente por su epidermis (Oliveira y Luz, 2005).
El tiempo de infección puede ser de tres a ocho semanas, que varía según la edad del fruto,
severidad del patógeno, susceptibilidad del hospedante y condiciones del clima,
principalmente la presencia de lluvias, mientras que en frutos tiernos, en días lluviosos y
calurosos, el período de incubación se acorta a 21 días (Fundación Hondureña para
Investigación Agrícola, 2003); sin embargo, Cruz (1993) reportó que el período de
incubación fluctúa entre 30 y 70 días.
2.4.4.9. Descripción morfológica del cultivo de M. roreri in vivo
Las esporas en germinación, son capaces de penetrar la epidermis de las mazorcas de cacao
en alguna etapa del desarrollo. Estas hifas invaden el tejido intercelularmente, y crecen
entre las células del parénquima cortical en forma de características hifas hinchadas de
4-7 µm de diámetro enroscadas o grumosas. Las mazorcas jóvenes pueden desarrollar
hinchamientos pronunciados, pero los síntomas pueden estar totalmente ausentes hasta la
formación de la lesión, que va de 45 a 90 días después de la penetración. Esta puede ser
considerada como la fase biotrófica (Evans et al., 1977).
La fase necrotrófica (la cual puede ser precedida por maduración organoléptica prematura e
irregular), desarrolla rápidamente lesiones irregulares de apariencia chocolate a café
oscuro, las que se unen gradualmente hasta cubrir la superficie entera de las mazorcas

Revisión Bibliográfica
21
infectadas. En una etapa tardía, predominan hundimientos limitados y lesiones de color
café oscuro. Un pseudostroma de color blanco a crema, crece sobre la superficie de la
mazorca de 3 a 8 días después del inicio de la lesión y pronto se cubre con una densa capa
polvorienta de esporas florecientes, que gradualmente cambian a gris, canela o café
(Evans et al., 1978). El cambio de coloración, al parecer se asocia con el engrosamiento de
la pared conidial, lo que habilita a la espora para sobrevivir a la desecación y retener la
infectividad por períodos prolongados (Evans et al., 1977).
2.4.4.10. Signos y síntomas de la enfermedad
Una de las características del hongo M. roreri es su largo período de incubación, el cual
puede ser de tres a ocho semanas, según la edad de la mazorca, severidad del ataque,
susceptibilidad del genotipo de cacao, y las condiciones del clima, principalmente la
presencia de lluvias (Hernández et al., 1991; Johnson et al., 2008; Villavicencio, 2010).
Los frutos son los únicos órganos que pueden ser infectados naturalmente por M. roreri. La
infección, normalmente ocurre cuando estos son jóvenes, ya que cuando maduran son más
resistentes. Las esporas se depositan sobre el fruto, germinan si hay agua, o mueren por la
radiación/desecación. El período de incubación es largo, generalmente mayor a un mes. El
daño se extiende internamente conforme los frutos crecen, y posteriormente el organismo
emerge a la superficie para esporular. El hongo patógeno se desarrolla intracelularmente e
invade las células del parénquima cortical. Algunos frutos que están muy dañados
internamente, a veces no muestran evidencias externas de la enfermedad (infecciones
ocultas), pero pueden ser reconocidos porque pesan más, debido a que están llenos de agua
por la descomposición interna que sufren (Phillips-Mora, 2004; Correa et al., 2014).
Los síntomas externos de la enfermedad, inician con la aparición de pequeños puntos
aceitosos sobre los frutos de 30-45 días de edad. Estos usualmente no son detectados hasta
que incrementan su tamaño, formando manchas necróticas (manchas de color chocolate)
aproximadamente a los 75 días después de la infección. Otros síntomas iniciales son la
deformación o muerte de frutos pequeños, lo cual en algunos casos se puede confundir con

Revisión Bibliográfica
22
la muerte fisiológica de los mismos. También es frecuente que algunas mazorcas en
diferentes estados de desarrollo, presenten una maduración prematura en toda o en una
parte de su superficie (Phillips-Mora, 2004). Luego de 4 a 5 días después de la aparición de
las manchas chocolate, aparece micelio blancuzco que se va tornando un poco más oscuro,
a medida que las esporas maduran. En las semanas posteriores, los frutos se deshidratan y
van momificándose gradualmente (Phillips-Mora, 2004; Correa et al., 2014).
En el caso de los síntomas internos, el hongo produce una necrosis progresiva de los tejidos
internos. En los estados avanzados de la enfermedad, el interior de los frutos (los tejidos
centrales, la pulpa, las semillas), forma una sola masa compacta acuosa en la que es difícil
distinguir sus partes originales. Sin embargo, en dependencia del momento en que los
frutos fueron infectados, las semillas podrían presentar un mayor o menor nivel de
deterioro (Evans et al., 1978; Phillips-Mora, 2004) (Figura 3).
Figura 3. a) Mazorca con pequeños abultamientos o gibas; b) fruto con manchas chocolate
a café oscuro, con bordes irregulares c) mazorca con capa polvorienta de color blanco
d) fruto con infecciones ocultas, almendras, cáscara, tejidos centrales formando una
pudrición acuosa. Fuente: Propiedad del autor
2.4.4.11. Descripción del cultivo de M. roreri in vitro
M. roreri, crece lentamente sobre agar extracto de malta. Las colonias alcanzan un diámetro
de 8-15 mm después de dos semanas, y muestran un borde levemente levantado. El
crecimiento en forma de tapete de aspecto lanoso a fieltro, de color salmón pálido a rosa
beige, finalmente se vuelve de color beige canela a arcilla, o localmente café lanoso a
chabacano pálido. Ocasionalmente el tapete es de inicio suave a farináceo, de color blanco
a b c d

Revisión Bibliográfica
23
a crema con matices ocráceos. No produce olor. En la zona avanzada, la hifa es hialina, de
pared delgada, septada y algunas veces es levemente irregular con hinchamientos de
1,5-5,0 µm de ancho. La hifa aérea es de dos tipos: a) como en la zona avanzada, pero con
paredes levemente engrosadas, y b) hialina densa de color pardusco pálido, no septado de
1-1,5 y 2-3 µm de ancho y el esqueleto raramente ramificado (Evans et al., 1978; Phillips-
Mora, 2003).
Los conidióforos ramificados dan lugar a cadenas maduras de esporas. Las esporas son
fácilmente separables, de paredes gruesas, de color amarillo pálido y de color café en masa,
típicamente globosa o subglobosa, de 6,5-8,0 y de 15-25 µm de diámetro, algunas veces
elipsoidales de 8-20 x 5-14 µm, paredes de 2 µm de grosor. Las esporas cilíndricas de
paredes delgadas también están presentes y probablemente se derivan de cadenas
inmaduras (Evans et al., 1978; Evans, 1986 y Phillips-Mora, 2003) (Figura 4).
Figura 4. Estructuras morfológicas de M. roreri que muestran esporas con paredes gruesas
y micelio septado. Fuente: Evans et al. (1978)
Las hifas se sumergen como en la zona avanzada pero con hinchamientos más frecuentes.
El hongo crece y esporula bien sobre tallos leñosos de cacao esterilizados con pérdidas
significativas de peso del sustrato; grumos de color crema compactos, de micelio estéril con
gotitas de exudado amarillo que se forman frecuentemente y los parches rojo púrpura
compuestos de hifas rojas pesadamente incrustadas que están presentes en los cultivos
viejos. Las temperaturas de crecimiento óptimo son de 25 a 26 °C y máximo 33 °C
(Evans et al., 1978; Phillips-Mora, 2003).

Revisión Bibliográfica
24
2.4.4.12. Medidas de control
Control cultural. Evitar el excesivo crecimiento del árbol de cacao, ya que el autosombreo
reduce su actividad fotosintética, y crea un microclima interno que estimula la infección y
el desarrollo de la enfermedad en los frutos (Amores et al., 2009), lo que también ayuda a
una adecuada recolección de frutos en el momento de la cosecha. Las podas sanitarias en la
cosecha constituyen otra buena práctica cultural para eliminar las partes afectadas por
insectos o enfermedades (Enríquez, 2010).
La poda frecuente, regulación del estrato superior, buen drenaje, densidades de plantación
apropiadas, control de malezas y correcto programa de fertilización, ayudan al óptimo
desarrollo del árbol pues hace que los patógenos que incidan en las plantaciones tengan
pocas probabilidades de establecerse y desarrollarse; en caso de que la enfermedad llegue a
establecerse, con un manejo cultural adecuado puede ser controlada económicamente y se
convivir con ella (IICA, 2006; Johnson et al., 2008). También es importante realizar
semanalmente la recolección y destrucción de frutos con signos del hongo fitopatógeno: se
deben remover en las primeras horas de la mañana o en la tarde después de las 16:00 h,
para evitar que la humedad propicie el desprendimiento de las esporas, y por último, en
dependencia de la cantidad de frutos enfermos, pueden enterrarse o trasladarse a otro sitio
(Krauss et al., 2003; Sánchez et al., 2003; FEDECACAO, 2004).
Control biológico. Se define como la reducción de la densidad de inóculo o sus
estructuras reproductoras de un hongo patógeno o parásito en estado activo o de
dormancia, por uno o más organismos obtenidos naturalmente (Uquillas, 2004). Aunque
con menor aplicación a nivel de campo, el manejo biológico surge como una alternativa
promisoria para el control del fitopatógeno, con resultados exitosos contra un amplio
rango de ellos. En los últimos años se ha demostrado el gran potencial de hongos y
bacterias endófitos para el control de la Moniliasis. Entre los microorganismos más
importantes se encuentran las bacterias de los géneros Pseudomonas y Bacillus, así como
hongos de los géneros Clonostachys y Trichoderma (Evans et al., 2003a; Suárez y
Rangel, 2014; Sun et al., 2015).

Revisión Bibliográfica
25
Trichoderma
El género Trichoderma, se presenta en forma natural en diversas condiciones ambientales y
sistemas agrícolas, principalmente en aquellos que contienen materia orgánica o desechos
en descomposición (Harman, 2006; Martínez et al., 2013).
Según Infante et al. (2009), las colonias del género Trichoderma presentan color blanco en
un inicio de su desarrollo, que se torna verde oscuro o amarillento, con esporulación densa.
El micelio se halla disperso, fino al microscopio, los conidióforos son ramificados, similar
a un árbol pequeño. Estos terminan en fiálides donde se forman las esporas asexuales o
conidios, aspecto importante en la identificación taxonómica a nivel de especies. Son
haploides y su pared está compuesta por quitina y glucanos.
Mecanismos de acción directos de Trichoderma
Trichoderma ha mostrado efecto sobre hongos patógenos a través de mecanismos de
acción, los que no son mutuamente excluyentes y pueden variar según el patógeno y la cepa
a utilizar (Martínez et al., 2013). Se han descrito diferentes mecanismos de acción para
controlar hongos patógenos, entre los cuales se encuentran competencia (por espacio y
nutrientes), antibiosis, micoparasitismo, y desactivación de enzimas de los patógenos
(Harman, 2000).
La competencia se define como el comportamiento desigual de dos o más organismos ante
un mismo requerimiento, siempre y cuando la utilización de este por uno de los organismos
reduzca la cantidad o espacio disponible para los demás (Infante et al., 2009). Según
Hernández et al. (2011), se da principalmente por nutrientes y espacio; de esta forma la
manera más común de muerte de un microorganismo es por inanición, debido a que
requiere de nutrientes exógenos como carbono y hierro para germinar, penetrar e infectar el
tejido, así si los sitios de infección están ocupados por el organismo benéfico el hongo
fitopatógeno tendrá dificultades para prosperar.

Revisión Bibliográfica
26
La antibiosis es considerada como el mecanismo de acción directa de antibióticos o
metabolitos tóxicos producidos por un microorganismo con acción sobre otro. Los
metabolitos de acción antifúngica secretados por Trichoderma constituyen un grupo de
compuestos volátiles y no volátiles muy diversos en cuanto a su estructura y función.
Trichoderma produce metabolitos secundarios, los que inhiben el crecimiento de otros
microorganismos con los cuales no llegan a establecer contacto físico (Martínez et al.,
2013). Las especies de Trichoderma secretan enzimas hidrolíticas como glucanasas,
quitinasas y proteasas que hidrolizan componentes de la pared celular de hongos como
Sclerotium rolfsii Sacc, Rhizoctonia solani Kühn y Pythium sp. (González et al., 2011).
El micoparasitismo se define como la interacción antagónica (directa) entre dos hongos, es
decir el parasitismo de un hongo (hospedante) por otro hongo (micoparásito) (Lecuona,
1996). Según Martínez et al. (2013) el proceso complejo del micoparasitismo ocurre en
cuatro etapas:
Crecimiento quimiotrófico: donde Trichoderma puede detectar a distancia a sus
posibles hospedantes.
Reconocimiento: Se considera que existe una alta especificidad del antagonista por
su sustrato.
Adhesión y enrollamiento: Ocurre por la asociación de un azúcar de la pared del
antagonista con una lectina presente en la pared del patógeno.
Actividad lítica: Producción de enzimas líticas extracelulares, fundamentalmente
quitinasas, glucanasas y proteasas, que degradan las paredes celulares del patógeno
y posibilitan la penetración de las hifas de Trichoderma.
El micoparasitismo concluye con la pérdida del contenido citoplasmático de la célula
hospedante, que muestra síntomas de disgregación.
Diferentes interacciones hifales están involucradas en el micoparasitismo, tales como:
enrollamiento, penetración, vacuolización, granulación, coagulación, desintegración y lisis.
A nivel microscópico no todas esas interacciones son siempre observadas, pues al parecer

Revisión Bibliográfica
27
dependen del aislado de Trichoderma, del patógeno y de las condiciones del ambiente
(Chet et al., 1981).
Mecanismos de acción indirectos de Trichoderma spp. como agente de biocontrol
Harman (2000, 2006); Harman y Shoresh (2007) y Vinale et al. (2008) refirieron varios
mecanismos indirectos de Trichoderma sobre los patógenos, ya que su acción es elicitar o
impulsar mecanismos de defensa fisiológicos o bioquímicos en la planta, como son:
Aceleración del desarrollo del sistema radical que posibilita la tolerancia al estrés
por parte de la planta.
Solubilización y absorción de nutrientes inorgánicos.
Estimulación del crecimiento vegetal.
Inducción de resistencia.
Trichoderma spp. es un microorganismo con capacidad de solubilizar nutrientes y
minerales del suelo no disponibles para las plantas. La forma en que lo hace es mediante
tres mecanismos: acidificación del medio mediante la liberación de ácidos orgánicos que
secuestran cationes y acidifican el medio alrededor de las raíces, producción de metabolitos
quelantes (sideróforos) que quelatizan Fe, Fe3+ y Cu2+, y actividad redox (Olivera Costa y
Rodríguez, 2014).
El género Trichoderma además de controlar enfermedades producidas por diferentes
patógenos, tiene la capacidad de promover el crecimiento de las plantas inoculadas con él
(Negrone et al., 2008).
Según Agrios (2005), la resistencia se define como la capacidad de un organismo para
excluir o superar, por completo o en algún grado, el efecto de un agente patógeno o factor
perjudicial de otro tipo. Las plantas presentan mecanismos de defensa, uno de ellos es
denominado constitutivo, que es el que actúa de forma pasiva, ya que la resistencia la
otorga mediante deposición de cera y cutícula sobre células de la epidermis, modificación
de la estructura de las paredes celulares por la acumulación de lignina, calosa, suberina,

Revisión Bibliográfica
28
gomas, cutina, glicósidos fenólicos, fenoles, quinonas, esteroides, glicoalcaloides,
terpenoides y tioninas, paredes celulares más gruesas, tamaño, localización y forma de
estomas, y presencia de tricomas, conocidos como mecanismos físicos; los bioquímicos son
fitoanticipinas, lectinas, proteínas inhibidoras de poligalacturonasas, quitinasas y
glucanasas. En cambio, la inducción de mecanismos de resistencia a enfermedades, se
activa por la mediación de sistemas de reconocimiento específicos mediante los cuales la
planta reconoce el ataque de un patógeno (Rossi, 2010).
Bacterias como Pseudomona aeruginosa (Schroeter, Migula), Leuconostoc mesenteroides
(Tsenkovkii, Van Tieghem), Bacillus subtilis (Conh), y Burkholderia cepacia (Burkholder),
han sido evaluadas ampliamente en el control de Moniliasis (Jiménez 1986; Sandoval et al.,
1987; Jiménez et al., 1988; Macagman et al., 2005; Solís y Suárez-Capello, 2006; Peralvo y
Saavedra, 2006).
Bacillus subtilis (Conh) pertenece a la familia Bacillaceae, género Bacillus, y sus
características son: Gram positivo, catalasa positiva, aerobia estricta (aunque puede crecer
en vía anaerobia), productor de esporas, antibióticos y matriz extracelular que comúnmente
se encuentra en el suelo. El potencial de B. subtilis se basa en la capacidad de producir una
amplia gama de moléculas bioactivas, que muestran fuertes propiedades antifúngicas, junto
con una baja toxicidad, alta biodegradabilidad, y características amigables con el medio
ambiente, en comparación con plaguicidas químicos. Adicionalmente la capacidad de
formar endoesporas, que le proporciona un nivel alto de resistencia a condiciones
ambientales extremas, hace que estas bacterias sean buenas candidatas para el desarrollo de
bioproductos. Debido a su eficiente control de Moniliasis en cacao, existen productos
registrados a base de esta bacteria (Errington, 2003; Ongena et al., 2009; Bettiol et al.,
2014).
Control químico. Control que trata básicamente de evitar la entrada del hongo patógeno a
la plantación y si está presente impedir que encuentre las condiciones favorables de
infección, multiplicación y diseminación (Sánchez y Garcés, 2012).

Revisión Bibliográfica
29
Para el control químico de M. roreri se emplean tradicionalmente fungicidas protectantes,
aunque con cuestionable eficacia. Sin embargo, el uso de cobre y protectantes orgánicos ha
permitido reducir la incidencia y severidad de la enfermedad. Aunque los sistemas de
aplicación mejorados con fungicidas sistémicos benefician la eficiencia en el control del
hongo patógeno, existe una baja adopción por el incremento en los costos de producción
(Flood y Murphy, 2004).
En Colombia, el control químico de M. roreri ha sido, después del método cultural, el más
investigado. Los resultados en plantaciones híbridas han sido erráticos o contradictorios y
en algunos de ellos antieconómicos, por lo que los fungicidas a base de cobre, son los
únicos productos o ingredientes activos con mejor respuesta. Después de varios ensayos, en
Santander, Argüello (2000) concluyó que el mejor control se obtiene con óxido cuproso
(Oxicloruro de cobre al 35%). Sin embargo, con el fin de minimizar los efectos adversos
que presentan sobre los agroecosistemas y sus pobladores, es necesario desarrollar
productos nuevos y aceptables para el control de fitopatógenos. Uno de los efectos que se
presenta es la emergencia de fitopatógenos resistentes a los fungicidas usados, otro es la
intoxicación aguda y general de humanos y otros organismos (Barrera-Necha et al., 2008).
Con la aplicación de productos químicos se consigue controlar la enfermedad de manera
significativa, lo que demuestra los beneficios de los fungicidas en la protección de los
frutos; los fungicidas sistémicos muestran un mejor control de M. roreri en comparación
con los de contacto, lo cual implica reducir el número y los costos de aplicaciones; al
combinar las labores culturales con los controles químicos, se logran incrementos de cacao
sano en alrededor del 20% (Ayala, 2008).
Control genético. Entre los cultivares de la especie T. cacao hay diferencias en la
susceptibilidad a M. roreri, lo cual muestra que en esta especie existen fuentes de
resistencia al hongo. Aún no se ha descubierto un material inmune a M. roreri, pero de las
pruebas en Ecuador, Colombia, Costa Rica y Honduras, se conoce que hay cultivares
(clones o híbridos) que consistentemente muestran menor número de mazorcas infectadas.

Revisión Bibliográfica
30
Ejemplo de esos materiales son: UF-273, EET-75, EET- 233, UF-296, CC-210, IMC-67 y
el CC-266. (González y Roble, 2014).
2.5. Área bajo la curva del progreso de la enfermedad (ABCPE)
Van der Plank (1963) propuso una modificación al modelo epifitiológico de puntos
múltiples, donde se usó el ABCPE como descriptor de la epifitia, basada en la
determinación de las pérdidas de diferentes cultivos afectados por agentes fitopatógenos.
Esta variable relaciona las pérdidas de los cultivos con las sumas de enfermedades, durante
un período de tiempo dado en un período específico del crecimiento del cultivo. Shaner y
Finney (1977), a su vez lograron definir matemáticamente la fórmula que actualmente se
utiliza para su cálculo.
Para determinar el ABCPE, se puede utilizar el método del punto medio o el método de
integración trapezoidal en dos intervalos de tiempo dados (Campbell y Madden, 1990).
Generalmente se utiliza en epifitias de corta duración, que se presentan de forma tardía
durante el crecimiento del cultivo. A medida que los intervalos de tiempo en los cuales se
evalúa la epifitia son menores, se logra mayor exactitud del ABCPE. Esta variable asume
que el daño al hospedante es proporcional a la cantidad de tejido enfermo, y a la duración
de la enfermedad.

Materiales y Métodos

Materiales y Métodos
31
3. MATERIALES Y MÉTODOS
La investigación se desarrolló en la Universidad Estatal Amazónica, cantón Pastaza,
parroquia Puyo, provincia de Pastaza, Ecuador; en el período comprendido de enero de
2013 a enero de 2016.
Recolección de mazorcas con síntomas de Moniliasis
La recolección de mazorcas del cultivar Nacional que presentaban síntomas de Moniliasis
se realizó en pequeñas fincas de producción orgánica en Napo, Pastaza y Morona Santiago
(Anexos 1, 2 y 3), provincias de la Región Amazónica Ecuatoriana, mediante un muestreo
sistemático, de acuerdo con los criterios de Sánchez et al. (2003) (Anexo 4).
Se recolectaron 50 mazorcas por finca y se colocaron en bolsas de papel, que fueron
trasladadas al laboratorio; se tomaron fotos con la cámara digital (Nikon AF-S NIKKOR).
Dichas mazorcas se separaron según sus síntomas y fueron codificadas de acuerdo con la
finca de procedencia con vistas a su fácil manejo e identificación.
Aislamiento y obtención de cultivos puros de M. roreri
El trabajo experimental se realizó en el Laboratorio de Microbiología de la Universidad
Estatal Amazónica. Con un sacabocado de 5 mm de diámetro, se tomaron segmentos de
mazorca con partes de tejido enfermo, y sano, estos se colocaron en un Erlenmeyer de
500 mL y se lavaron con agua corriente durante 5 min con el uso de un tamiz metálico. Los
fragmentos se desinfectaron en agitación continua con alcohol al 70% durante 3 min y
nuevamente se agitaron en Erlenmeyer con agua estéril durante 2 min. Finalmente se
colocaron en cámara húmeda.
Asimismo, se tomaron segmentos de mazorca que presentaban crecimiento micelial de
color blanco y esporulación de color crema, los cuales se ubicaron directamente en la
cámara húmeda sin ser previamente desinfectados.

Materiales y Métodos
32
Los segmentos desinfectados se colocaron en placas de Petri (90 mm Ø) Marca Senna
Modelo 3-61015; conformadas las cámaras húmedas se incubaron a 26±2 °C en
condiciones de luz alterna (12 h luz más 12 h oscuridad) durante 48 h. Transcurrido este
tiempo se aislaron los hongos filamentosos por medio del examen bajo el microscopio
óptico (MEIJI, aumento 800 X) y la ayuda de un asa previamente esterilizada. Bajo el
microscopio estereoscopio binocular (Kruss, aumento 200X), se realizó la siembra directa
en placas de Petri (90 mm Ø), que contenían 16 mL de medio de cultivo V8 modificado
(Anexo 5). Una vez realizada la siembra se incubaron los cultivos a 26±2 °C durante 15
días. La identificación se realizó teniendo en cuenta los criterios morfológicos (esporas y
conidióforos) acorde a lo referido por Johnson (1946). La observación de las estructuras
morfológicas se hizo con la ayuda del microscopio óptico (MEIJI, aumento 800 X).
Reaislamiento de M. roreri
Para el reaislamiento, se tomaron esporas individuales, las cuales fueron colocadas en
placas de Petri (90 mm Ø) que contenían 10 mL del medio de cultivo V8 modificado y se
incubaron a 26±2 °C durante 15 días; transcurrido este tiempo se seleccionaron las
colonias típicas de M. roreri.
Registro y conservación de M. roreri
Los aislados monospóricos de M. roreri fueron conservados en glicerol al 15% y en agua
estéril a una temperatura de -4±2 °C en refrigerador (Indurama), y en el ultra congelador
(Daihan Scientific) a una temperatura de -80±2 °C hasta su empleo.
3.1 Caracterización cultural, morfológica, fisiológica, molecular y patogénica de
aislados de M. roreri
3.1.1 Caracterización cultural de M. roreri
Cada aislado monospórico de M. roreri se subcultivó en placas de Petri (90 mm Ø)
contentivas de 10 mL de V8 modificado, y se incubaron a 26±2 °C en condiciones de
oscuridad durante dos semanas.

Materiales y Métodos
33
La caracterización cultural se realizó por medio de análisis cualitativo y cuantitativo. Se
utilizó un diseño completamente aleatorizado donde los aislados fueron los tratamientos
con 10 repeticiones (placas de Petri).
Se evaluó el crecimiento micelial (Ø de la colonia mm), color del anverso y reverso de la
colonia, bordes y textura de la colonia, presencia o no de líquidos de transpiración,
presencia de cristales, presencia de pigmentos difusibles en los medios de cultivo y
presencia de sectores. El crecimiento de los aislados de M. roreri se determinó con la
ayuda del calibrador digital Truper®; mediante observación visual se definió el color del
anverso y reverso de las colonias según lo especificado en trabajos realizados por Villamil
et al. (2012), borde y textura, presencia o no de líquidos de transpiración, presencia de
cristales, presencia de pigmentos difusibles en los medios de cultivo, presencia de sectores
de las colonias por cada aislado, bajo el microscopio estereoscopio.
Para ello se usaron placas de Petri (90 mm Ø) contentivas de 10 mL de medio de cultivo
V8 modificado (Evans et al., 1978 y Phillips et al., 2006), las cuales se inocularon en el
centro con un disco de micelio de 5 mm de cada uno de los aislados. Las condiciones de
incubación fueron a 26±2 °C en condiciones de oscuridad. Las mediciones fueron
registradas diariamente, hasta que la placa de Petri estuvo completamente cubierta.
Para procesar la información de las variables color del anverso y reverso de la colonia,
bordes, textura, presencia de líquido de transpiración, cristales, presencia de pigmentos
difusibles en el medio de cultivo y sectores se utilizó la estadística descriptiva, y se
realizaron gráficos de distribución de frecuencia.
La variable crecimiento micelial de las colonias (mm), se comparó entre los tratamientos
mediante una prueba no paramétrica de Kruskal Wallis complementada con la prueba de
Mann Whitney (P≤0,05), previa comprobación de los supuestos de normalidad de los datos
y homogeneidad de varianzas. Para todos los análisis se utilizó el paquete estadístico IBM
SPSS Statistic versión 21,0 para Windows.

Materiales y Métodos
34
3.1.2 Caracterización morfológica de M. roreri
Previamente se realizó la inducción de formación de esporas en los cultivos monospóricos
en medio V8 modificado, según lo recomendado por Phillips et al. (2006) y Villavicencio
(2010).
Para la caracterización morfológica de los aislados se realizaron microcultivos (Riddel,
1950) y se midió el largo y ancho (µm) de las esporas por la zona más gruesa, en 100
estructuras tomadas al azar de cada uno de los aislados.
Se consideró un diseño completamente aleatorizado para los aislados (tratamientos) y las
100 esporas como repeticiones.
La observación se realizó con la ayuda del microscopio óptico (aumento 400 X). Se
tomaron imágenes digitales con la cámara acoplada (DINOLITE). Además se describió la
forma de las esporas según los criterios descritos por Evans et al. (1978).
Para el análisis de los datos correspondientes al ancho y largo de las esporas se utilizó la
prueba de C de Dunnett (P≤0,05), previa comprobación de los supuestos de normalidad de
los datos y homogeneidad de las varianzas. Las variables fueron evaluadas con el paquete
estadístico IBM SPSS Statistic versión 21,0 para Windows.
3.1.3. Caracterización fisiológica de M. roreri
Para la caracterización fisiológica se utilizó el aislado que presentó mayor crecimiento
micelial en el acápite 3.1.1.
3.1.3.1. Influencia del pH en el crecimiento micelial
Se utilizó el medio de cultivo Caldo papa dextrosa (PDB Difco ®), y esterilizó en
autoclave a 121 °C y 1 atm por 15 min; el medio fue vertido en Erlenmeyer de 100 mL de
capacidad, bajo la cámara de flujo laminar. En este medio se inocularon cuatro discos de
micelio con la ayuda de un sacabocado de 5 mm de diámetro y una aguja de inoculación, el

Materiales y Métodos
35
ajuste del pH se realizó antes de esterilizarlo: se ajustó con NaOH con una concentración
de 0,001 M, en un intervalo de pH desde 6 a 7,5 con una escala de 0,5 entre pH, y se
mantuvieron en la incubadora a temperatura de 26±2 °C durante 21 días.
Transcurrido este tiempo el micelio se separó del medio de cultivo mediante filtración con
filtros de papel (Whatman de 125 mm Ø), luego se desecó en estufa (Memmert) a 60 °C
hasta mantener el peso constante y se determinó su masa seca (g) en una balanza analítica
digital (22 ADAM ).
Los diferentes tratamientos de pH (6, 6,5, 7 y 7,5) se dispusieron en un diseño
completamente aleatorizado con 10 repeticiones (placas de Petri).
La masa del micelio de los distintos tratamientos se comparó por medio de una prueba no
paramétrica de Kruskal Wallis complementada con la prueba de Mann Whitney (P≤0,05),
previa comprobación de los supuestos de normalidad de los datos y homogeneidad de
varianza. Las variables fueron evaluadas con el paquete estadístico IBM SPSS Statistic
versión 21,0 para Windows.
3.1.3.2 Influencia de la temperatura en el crecimiento micelial
El rango de temperaturas se ensayó desde 15±2 °C hasta 35±2 °C a intervalos térmicos de
5 °C. La inoculación del hongo patógeno se realizó en medio V8 modificado, donde se
colocó un disco de micelio de 5 mm Ø de M. roreri en el centro de la placa de Petri
(90 mm Ø). La incubación fue realizada en ausencia de luz a las temperaturas en estudio.
La temperatura de 15±2 °C se aseguró en un refrigerador, mientras que el intervalo térmico
de 20, 25, 30, 35±2 °C fue logrado en incubadora.
Las temperaturas determinadas en cada equipo fueron monitoreadas por un período de 15 a
20 días, con la utilización de termómetro de mercurio (Scientific). Se evaluó el crecimiento
micelial mediante la observación diaria hasta los 12 días.

Materiales y Métodos
36
Los diferentes tratamientos de temperatura (15, 20, 25, 30 y 35 °C) se dispusieron en un
diseño completamente aleatorizado con 10 repeticiones (placas de Petri).
La variable crecimiento micelial (mm), se comparó mediante la prueba no paramétrica de
Kruskal Wallis complementada con la prueba de Mann Whitney (P≤0,05), previa
comparación de los supuestos de distribución normal y homogeneidad de las varianzas.
Las variables fueron evaluadas con el paquete estadístico IBM SPSS Statistic versión 21,0
para Windows.
3.1.3.3 Influencia de diferentes regímenes de luz en el crecimiento micelial
Se evaluaron cuatro tratamientos (regímenes de luz): luz alterna (12 h luz y 12 h
oscuridad), luz blanca, luz ultravioleta (longitud de onda 404 nm) y oscuridad continua,
con diez repeticiones (placas de Petri). Se empleó un diseño completamente aleatorizado.
La variante de luz alterna fue provocada en condiciones naturales de laboratorio. En el
caso de la variante de luz blanca se logró en una caja de madera estéril con una lámpara de
200 W colocada a 50 cm de distancia de las superficies de las placas de Petri. La variante
de luz ultravioleta fue provocada en un aparato de inducción de esporulación en el cual se
colocaron dos lámparas negras (BLB40W 120 V con 173.88 lx) separadas a una distancia
de 50 cm de los cultivos de los hongos. Finalmente, la variante de oscuridad continua se
aseguró en una estufa (Memmert).
La inoculación de M. roreri se realizó en medio V8 modificado de la misma forma en que
fue descrita en el experimento anterior. La incubación se hizo a temperatura de 26±2 °C.
Las evaluaciones y los procesamientos estadísticos se realizaron del mismo modo que los
descritos en el acápite 3.1.3.2.

Materiales y Métodos
37
3.1.4. Identificación molecular de aislados de M. roreri
La extracción de ADN de aislados de M. roreri se realizó a partir de cultivos monospóricos
de cada uno de los aislados objeto de estudio. Cultivos esporulados de cada uno de los
aislados se sembraron en Erlenmeyer de 250 mL, que contenían medio caldo de papa
dextrosa. Para la siembra, se depositaron dos discos de 5 mm de diámetro por cada
Erlenmeyer, los cuales se incubaron en agitación (Thermo Scientific, EE.UU.) a 250 rpm y
26±2 °C durante 15 días. Posterior al período de crecimiento, los aislados se filtraron
usando papel de filtro (Whatman #1, EE.UU.), y los micelios se depositaron en placas de
Petri (90 mm Ø) durante 2 h en cabina de flujo laminar para garantizar el secado total.
La extracción de ADN genómico se llevó a cabo mediante el uso del sistema comercial
Purelink Plant Extraction Kit (Invitrogen, EE.UU.), según las instrucciones del fabricante.
El producto de la extracción se analizó por espectrofotometría UV/VIS en un Nanodrop
2000 (Thermo Scientific, EE.UU.) y por electroforesis en gel de agarosa al 1% en tampón
TBE 1 x (890 mM Tris-borato, 890 mM Ácido bórico y 20 mM EDTA), con una intensidad
de 100 V durante 45 min. Los productos fueron teñidos con bromuro de etidio (Sigma-
Aldrich, EE.UU.) y visualizados en luz ultravioleta (Sambrook y Rusell, 2001).
La amplificación de la subregión 18S ADNr se realizó mediante la Reacción en cadena de
la polimerasa (PCR), con la utilización de los cebadores universales NS1
(5´-GTAGTCATATGCTTGTCTC-3´) y NS8 (5´-TCCGCAGGTTCACCTACGGA-3´)
(White et al., 1990). En la Figura 5 se observa la posición de los oligonucleótidos NS1 y
NS8 en el gen que codifica para los ARNr en hongos.
Figura 5. Posición de los cebadores NS1 y NS8 en el gen que codifica para los ARNr en los hongos

Materiales y Métodos
38
La amplificación por PCR se llevó a cabo en el termociclador Veriti 9902 (Applied
Biosystems, EE.UU.), en un volumen final de 50 µL, que contenía: 37 µL de agua MiliQ,
5 µL de dNTPs 2mM, 5 µL de tampón Platinum 10 x, 1 µL de cada uno de los cebadores
(NS1 y NS8), 1,8 µL de MgCl2, 0,2 µL de Taq ADN polimerasa Platinum (Thermo
Scientific, EE.UU.) y 1 µL de ADN. En la reacción se adicionó un control para determinar
si existió contaminación o no en las muestras de ADN.
Las condiciones para las reacciones de PCR consistieron en una desnaturalización inicial de
94 °C durante 85 s, seguido de 35 ciclos de amplificación con una desnaturalización a
95 °C durante 35 s, hibridación a 55 °C por 55 s, y extensión a 72 °C por 45 s.
Adicionalmente, una extensión final a 72 °C durante 10 min. Los productos de PCR se
verificaron mediante electroforesis en gel de agarosa al 1%, según se describió
anteriormente. La observación de los productos de PCR se realizó en un fotodocumentador
ENDUROTM GDS TOUCH (Labnet, EE.UU.).
La secuenciación de los aislados se realizó en las instalaciones de Macrogen (EE.UU.), en
un secuenciador automático ABI 3730xl (Applied Biosystems, EE.UU.), para lo cual se
envió el ADN genómico, extraído con concentraciones y purezas adecuadas, en lugar del
producto de PCR para garantizar la re-secuenciación de las muestras en caso de fallos en el
proceso.
Las secuencias nucleotídicas obtenidas se limpiaron con la utilización del programa DNA
Baser Sequence Assembler v.4 (2013) y la herramienta Convertix. Para la búsqueda de
homología en las bases de datos GenBank se utilizó el programa Blastn
(Altschul et al., 1990), disponible en: http://www.ncbi.nlm.nih.gov/BLAST/. El
alineamiento múltiple de los nucleótidos se generó en MUSCLE (Edgar, 2004) en el
programa MEGA v.6 (Tamura et al., 2013).

Materiales y Métodos
39
3.1.5. Caracterización patogénica y agresividad de los aislados de M. roreri
El estudio se realizó con el fin de determinar la capacidad infectiva de los aislados de
M. roreri sobre mazorcas sanas del cultivar Nacional de cacao, así como para comprobar
los síntomas, y su correspondencia con lo observado en condiciones naturales (Phillips
et al., 2006).
Las mazorcas fueron colectadas en el Centro de Investigación, Posgrado y Conservación
Amazónica (CIPCA) de la Universidad Estatal Amazónica, procedentes de un sistema
agroforestal con manejo orgánico. Las mismas fueron envueltas con papel kraft
humedecido, y colocadas en un recipiente plástico, para su transportación. En el
laboratorio las mazorcas fueron lavadas con agua corriente durante 5 min, luego se
desinfectaron con alcohol al 70% durante 3 min, y posteriormente fueron enjuagadas con
agua destilada estéril por 2 min.
Para el crecimiento micelial de M. roreri, se empleó medio de cultivo PDB, y se procedió a
sembrar un disco de micelio con la ayuda de un sacabocado de 5,0 mm de diámetro. Los
Erlenmeyers se colocaron en una incubadora a 26±2 °C y se mantuvieron durante 12 días.
Se colectó el micelio dentro de la cámara de flujo laminar. Se juntó el micelio colectado de
cada aislado y se batió el contenido en una licuadora marca Oster®, durante 2 min. El
homogeneizado micelial, se filtró por cuatro capas en gasa estéril. Posteriormente se
determinó la concentración del filtrado de micelio en la cámara de Neubauer (Celeromics),
que fue ajustada a una concentración aproximada de 1,2 x 105 ufc mL-1.
La inoculación de las mazorcas de dos meses de edad (aproximadamente 10 cm de largo)
se realizó en la parte media, dentro de la cámara de flujo laminar, con la ayuda de una
micropipeta (SOCOREX SWISS 10-100 µl) con 0,5 mL de suspensión micelial. Una vez
secas, las mazorcas fueron colocadas en bolsas transparentes de polietileno herméticas
(16,5 cm x 14,9 cm) con papel filtro (Macherey Nagel) humedecido con 25 mL de agua
destilada estéril. Las bolsas fueron colocadas en la incubadora a 26±2 ºC durante tres

Materiales y Métodos
40
semanas. El papel de filtro se retiró 48 h después de la inoculación, con la ayuda de una
pinza (Phillips, 1986; Phillips-Mora et al., 2005).
Para evaluar la patogenicidad y agresividad de los aislados de M. roreri se determinó la
presencia o ausencia de síntomas sobre las mazorcas inoculadas, así como la magnitud
del daño para cada aislado. Se utilizaron diez mazorcas sanas de cacao del cultivar
Nacional por aislado e igual número sin inocular como control. Se evaluaron las
variables:
Severidad externa: basada en la apariencia externa del fruto, los síntomas y signos
inducidos por la incubación de los aislados sobre las mazorcas de cacao; para la evaluación
se utilizó la escala propuesta por Brenes (1983) (Anexo 6).
Severidad interna: basada en el porcentaje de necrosis interna observada en un corte
longitudinal del fruto y medido en relación con la escala desarrollada por Sánchez et al.
(1987) (Anexo 7).
Índice de infección: Ii= 1001 TNnb (Townsend y Heuberger, 1943), donde
n= número de porciones de cada mazorca en cada nivel; b= valor de severidad acorde con
la escala evaluativa, N= número de niveles de la escala, T= número total de porciones
(parte basal, parte media y parte distal de la mazorca) por cada mazorca inoculada.
Área bajo la curva del progreso de la enfermedad (ABCPE): Se calculó mediante la
fórmula propuesta por Shaner y Finney (1977):
Donde yi representa el índice de infección (severidad), en diferentes momentos de
evaluación (ti).

Materiales y Métodos
41
Los tratamientos (aislados) se dispusieron en un diseño completamente aleatorizado. Se
utilizaron diez repeticiones por cada aislado. Las evaluaciones se realizaron diariamente,
hasta los 12 días.
Los datos del ABCPE se analizaron estadísticamente previa comprobación de los supuestos
de normalidad de los datos y homogeneidad de varianza. Las comparaciones de las medias
se realizaron mediante la prueba no paramétrica de Kruskal Wallis complementada con la
prueba de Mann Whitney (P≤0,05). Las variables fueron evaluadas con el paquete
estadístico IBM SPSS Statistic versión 21.0 para Windows.
3.2 Respuesta de genotipos de cacao frente a M. roreri en un ensayo monocíclico
El experimento se realizó con la finalidad de determinar la respuesta de genotipos de cacao
ante la mezcla microbiana de los aislados de M. roreri de mayor agresividad en el acápite
3.1.5. Para ello se utilizaron mazorcas sanas de dos meses de edad de aproximadamente
10 cm, de cinco genotipos (cultivar Nacional y los clones CCN-51, EET-95, EET-96 y
EET-103), los cuales constituyeron los tratamientos.
La preparación del inóculo de la mezcla microbiana de M. roreri y la metodología de
inoculación empleada fueron similares a las referidas en el acápite 3.1.5.
Se inocularon con la mezcla microbiana 10 mazorcas por cada genotipo de cacao.
Se evaluó la respuesta de los genotipos de cacao frente a la mezcla microbiana de M. roreri
a través de las variables Severidad Externa, Severidad Interna, Índice de infección y
ABCPE, de la misma forma que fue descrito en el acápite 3.1.5. Las evaluaciones se
realizaron diariamente.
Para la clasificación de la respuesta de los genotipos, se empleó lo referido por Phillips-
Mora et al. (2005), quienes utilizan los valores de media de Severidad Interna (Sánchez
et al., 1987) donde:

Materiales y Métodos
42
0 – 1,25 Resistente
1,26 – 2,50 Moderadamente resistente
2,51 – 3,75 Moderadamente susceptible
3,76 – 5,0 Susceptible
El experimento se dispuso bajo un diseño completamente aleatorizado con cinco
tratamientos (genotipos) y 10 repeticiones (mazorcas).
Los datos del ABCPE se analizaron estadísticamente, previa comprobación de los
supuestos de normalidad de los datos y homogeneidad de las varianzas. Las comparaciones
de las medias se realizaron mediante la prueba de Kruskal Wallis complementada con la
prueba de Mann Whitney (P≤0,05). Se utilizó el paquete estadístico IBM SPSS Statistic
versión 21,0 para Windows.
3.3 Caracterización cultural y molecular de Trichoderma spp. procedentes de
plantaciones de cacao orgánico
El aislamiento de Trichoderma spp. se realizó a partir de muestras de suelo procedentes de
tres sistemas agroforestales (cacao orgánico) de la provincia de Napo.
Se colectaron muestras de suelo de los siguientes sectores:
1. Borja, ubicado en el Valle de los Quijos-Napo, Latitud: 0°18’49,046’’ (m),
Longitud: 77°47’0,458’’ (m). Altura: 1 455 (msnm). Sistema agroforestal que
incluyó cacao.
2. Ahuano, Napo. Latitud: 1°3’42,302’’ (m), Longitud: 77°32’52,433’’ (m). Altura:
355 (msnm). Sistema agroforestal que incluye cacao, de siete años de edad.
3. Centro de Investigación, Posgrados y Conservación Amazónica (CIPCA) que se
encuentra ubicado en el km 42 vía Puyo-Tena, Latitud: 1°15’39,708’’ (m),
Longitud: 77°53’23,490’’ (m). Altura: 575 (msnm). Plantación de cacao de cinco
años de edad.

Materiales y Métodos
43
Para la toma de muestras cada finca se dividió en cinco transeptos, donde se tomaron
veinte muestras de suelo rizosférico, a 20 cm de profundidad con un muestreador manual.
Las muestras se combinaron y homogeneizaron en un recipiente para obtener una muestra
única de 500 g de suelo por plantación. Se envasaron en bolsas plásticas codificadas, se
colocaron en nevera portátil para su traslado al laboratorio de Microbiología de la
Universidad Estatal Amazónica, y se secaron 24 h a temperatura ambiente.
Para aislar Trichoderma de las muestras de suelo, se realizó una dilución al 10% p/v de 1 g
de muestra de suelo en 10 mL de agua destilada, se dejó reposar durante 5 min, y se tomó
una alícuota de 200 µL de solución con la que se inoculó en placas de Petri (90 mm Ø) con
medio V8 modificado y se incubaron a 28±2 °C en oscuridad. Las colonias de
Trichoderma se identificaron según su morfología con la ayuda de un microscopio
trilocular óptico con cámara acoplada.
Para la purificación del cultivo se extrajo cuidadosamente el micelio en pequeñas
proporciones de Agar de cada colonia identificada como Trichoderma, y se sembraron en
medio V8 modificado.
La caracterización cultural de los aislados de Trichoderma spp. se realizó según la forma,
color y tipo de micelio, color de esporas, olor, presencia de fiálides y crecimiento de las
colonias teniendo en cuenta los criterios establecidos por Samuels et al. (2006b) y Samuels
et al. (2012a). Se sembraron discos de 5 mm en el centro de las placas de Petri (90 mm Ø),
contentivas con medio V8 modificado. Las placas se incubaron durante 72 h a 28±2 ºC.
Se determinó el crecimiento de los aislados de Trichoderma spp., con la ayuda del
calibrador digital; mediante observación visual se definió forma, color y tipo de micelio,
color de esporas, olor, presencia de fiálides de las colonias por cada aislado, bajo el
microscopio estereoscopio. Las mediciones se realizaron diariamente, hasta que la placa de
Petri (90 mm Ø) estuvo completamente cubierta.

Materiales y Métodos
44
El experimento se dispuso en un diseño completamente aleatorizado con tres tratamientos
(aislados) y 10 repeticiones (placas de Petri).
Se utilizó un diseño completamente aleatorizado. Para procesar la información de las
variables forma, color y tipo de micelio, color de esporas, olor, presencia de fiálides de las
colonias se hizo uso de la estadística descriptiva, mediante gráficos de distribución de
frecuencia.
La variable velocidad del crecimiento micelial (mm), se analizó estadísticamente aplicando
un análisis de varianza simple. Las medias se compararon según la prueba de rangos
múltiples de Duncan (P≤0,05). Se utilizó el paquete estadístico IBM SPSS Statistic versión
21,0 para Windows.
La identificación molecular de los aislados se realizó según el acápite 3.1.4.
3.4 Evaluación de la actividad antagónica in vitro de Trichoderma spp. frente a
M. roreri
Se evaluó el antagonismo in vitro del aislado de M. roreri que presentó mayor velocidad de
crecimiento en el acápite 3.1.1, frente a los aislados de Trichoderma spp. obtenidos en cada
uno de los sectores (acápite 3.3), cuyo código aparece debajo y que fueron considerados
como tratamientos :
UEA-T1: Aislado Trichoderma spp. (Borja)
UEA-T2: Aislado Trichoderma spp. (CIPCA)
UEA-T3: Aislado Trichoderma spp. (Ahuano)
Se sembraron discos del patógeno de 5,0 a 20 mm del extremo de la placa de Petri
(90 mm Ø) con medio de cultivo V8 modificado, y fueron incubados a 26±2 °C. El
antagonista Trichoderma se obtuvo con 72 h de crecimiento en medio de cultivo V8
modificado.

Materiales y Métodos
45
Cuando M. roreri alcanzó un desarrollo de 30 mm de diámetro en la placa de Petri (5 días
después de la siembra) (Solís, 1999), los aislados de Trichoderma se enfrentaron a
M. roreri mediante cultivo dual, para lo cual se colocó en el extremo opuesto de la placa de
Petri (90 mm Ø), un disco de 4,3 mm de diámetro de micelio del antagonista (de 6 días de
edad) con 50 mm de separación entre ellos (Howell, 2003). Los cultivos se incubaron a 26
±2 °C y oscuridad, y cada 24 h se midió el crecimiento radial del micelio de los hongos
hasta los siete días de inoculados.
El experimento se condujo en un diseño completamente aleatorizado con diez repeticiones
de cada tratamiento de Trichoderma spp. y diez placas de Petri (90 mm Ø) inoculadas con
Trichoderma spp. y M. roreri como controles.
Los mecanismos de acción de competencia se evaluaron teniendo en cuenta espacio,
micoparasitismo, antibiosis ejercidos por Trichoderma spp. sobre M. roreri.
La competencia por espacio se realizó mediante la comparación de la velocidad del
crecimiento. Se hicieron las siguientes evaluaciones 1. Interacción Trichoderma-hongo
patógeno y 2. Control (hongo patógeno), se midieron los radios de crecimiento del
patógeno (RCP) y del antagonista (RCA) con la ayuda de un calibrador digital, a partir de
las 24 horas, hasta que uno de los microorganismos en estudio cubriera la placa totalmente.
Conjuntamente, se calculó el porcentaje de inhibición de crecimiento radial (PICR), con el
empleo de la fórmula de Samaniego et al. (1989), donde R1 y R2 son los radios mayor y
menor de crecimiento radial del hongo patógeno, respectivamente.
De las 24 a 72 h se evaluó la tasa de crecimiento micelial en los aislados de Trichoderma
spp. según la fórmula de Lilly y Barnet (1951), donde CL1 y CL2 son los crecimientos en la

Materiales y Métodos
46
primera y segunda mediciones, y T1 y T2 son los momentos de la primera y segunda
mediciones.
A los días cuarto y quinto se determinó la capacidad antagónica del Trichoderma spp.,
frente a M. roreri de acuerdo con las clases de antagonismo descritas en la escala de
Bell et al. (1982):
Clase 1. Trichoderma sobrecrece completamente al patógeno y cubre totalmente la
superficie del medio de cultivo.
Clase 2. Trichoderma sobrecrece las dos terceras partes de la superficie del medio de
cultivo.
Clase 3. Trichoderma y el patógeno colonizan cada uno aproximadamente la mitad de la
superficie del medio de cultivo y ningún organismo parece dominar al otro.
Clase 4. El patógeno coloniza las dos terceras partes de la superficie del medio de cultivo y
parece resistir a la invasión por Trichoderma.
Clase 5. El patógeno sobrecrece completamente a Trichoderma y ocupa la superficie total
del medio de cultivo.
A su vez la zona de antagonismo se evaluó con la escala propuesta por Punja y Grogan
(1983), que consta de los niveles siguientes.
0. No hay zona de antagonismo.
1. Aversión ligera (Zona de antagonismo de +/- 1 mm de ancho).
2. Aversión moderada (Zona de antagonismo de 2 a 4 mm de ancho).
3. Aversión fuerte (Zona de antagonismo de más de 4 mm de ancho).
Para el mecanismo de acción micoparasitismo, se evaluaron cinco muestras tomando
fragmentos de micelio de la zona de interacción a partir de que existiera contacto entre las
hifas de M. roreri y el antagonista desde las 24 hasta las 240 h y se observó al microscopio

Materiales y Métodos
47
óptico con aumento de 400X el tipo de interacción hifal (enrollamiento, penetración,
vacuolización y/o lisis).
La antibiosis de Trichoderma se evaluó a través del efecto de metabolitos volátiles; se
realizó la prueba según el protocolo propuesto por Dennis y Webster (1971). En una tapa
de placa de Petri (90 mm Ø) que contenía medio de cultivo V8 modificado, se inoculó
micelio de M. roreri de 5,0 mm de diámetro y se dejó incubar a 26±2 °C hasta alcanzar un
diámetro de 30 mm (cuatro días después de la siembra). En la tapa de otra placa de Petri
(90 mm Ø) se añadió medio de cultivo V8 modificado con un disco de micelio de
Trichoderma spp. de 5,0 mm de diámetro (de seis días de edad). Se colocaron las dos tapas
una frente a la otra y se sellaron con parafilm. Las placas se incubaron a 26±2 ºC durante
siete días. Pasado el tiempo de incubación se observó el proceso de inhibición de M. roreri
y se comparó con el control en ausencia del antagonista. Se emplearon diez repeticiones
por cada tratamiento.
Al finalizar la prueba, a los 12 días, se cuantificó la esporulación de M. roreri, en cada uno
de los tratamientos. Para el efecto, sobre cada uno de los ejes horizontal y vertical, a 1 mm
del centro de la placa de Petri (90 mm Ø) se cortó un disco en el medio de cultivo, con la
ayuda de un sacabocados de 5,0 mm de diámetro. Los cuatro discos obtenidos por placa,
fueron suspendidos en un tubo de ensayo que contenía 10 mL de agua destilada estéril, con
una gota de Tween 20 (0,05%) y se agitó vigorosamente durante 10 min, para facilitar el
desprendimiento de las esporas del micelio. El conteo de las esporas se realizó en un
microscopio trinocular óptico, utilizando una cámara de Neubauer. Se calculó el porcentaje
de inhibición de esporulación (% I.E.) (Vásquez, et al., 2014) de la siguiente manera:
Los datos del potencial antagónico in vitro de Trichoderma spp. y el porcentaje de
inhibición de esporulación de M. roreri, fueron sometidos al análisis de varianza. Las

Materiales y Métodos
48
medias se compararon mediante la prueba Duncan (P≤0,05), utilizando el paquete
estadístico IBM SPSS Statistic versión 21,0 para Windows.
3.5. Evaluación de la eficacia de aislados nativos de Trichoderma spp. y biopreparados
comerciales en el control de la Moniliasis del cacao en condiciones de campo
La evaluación de la eficacia de aislados nativos de Trichoderma spp. sobre M. roreri se
realizó en el CIPCA, en áreas experimentales de cacao, cultivar Nacional de seis años de
edad, al inicio de la producción del árbol de cacao en el período comprendido de
septiembre de 2015 a enero de 2016, sobre suelo Inseptisol.
Los biopreparados a base de los aislados nativos de Trichoderma spp., se realizaron
artesanalmente en el Laboratorio de Microbiología de la Universidad Estatal Amazónica, a
partir de un sustrato sólido de cabecilla de arroz, al que se le añadió agua destilada estéril;
se distribuyó homogéneamente en frascos Roux, posteriormente se esterilizó en autoclave a
121 ºC durante 30 min, después de 2 h de reposo se inoculó el sustrato con una suspensión
obtenida de cultivos puros esporulados de siete días, a la concentración de
108 conidios mL-1, seguidamente se incubó a 28 ± 2ºC en posición horizontal hasta
colonizar y esporular todo el sustrato, y finalmente se llevó al área de secado. Se calculó la
concentración por gramo y se realizaron los cálculos correspondientes para determinar la
dosis de aplicación en campo. Respecto al control de la calidad se tuvieron en cuenta
aspectos referidos por Suárez-Castellá et al. (2009) para sistemas de control de la calidad
de laboratorios en la producción de plantas in vitro, aplicables en este caso.
Las suspensiones de Trichoderma spp. se mantuvieron por separado en botellas plásticas de
8 L a 5 ºC, desde su preparación hasta el momento de ser transportadas al campo dentro de
una caja térmica. La germinación de las esporas fue confirmada con siembras de las
suspensiones finales en medio V8 modificado, para de esta forma garantizar la viabilidad
de las esporas que fueron llevadas al campo.

Materiales y Métodos
49
El experimento de campo se desarrolló bajo un diseño de bloques completos al azar, con
seis tratamientos: los dos mejores aislados en condiciones de laboratorio, dos biopreparados
comerciales (Trichotic líquido y Rhapsody), un tratamiento químico y un testigo sin
tratamiento con tres réplicas (Tabla 1). Cada réplica estuvo conformada por una parcela de
cuatro árboles plantados a 4 x 4 m. Se dejó un área de cuatro hileras de plantas para evitar
el efecto de borde entre parcelas, tanto entre los tratamientos como entre los bloques. Antes
de las aplicaciones de los tratamientos, se realizó una poda sanitaria de los árboles para
reducción de la copa, junto con una eliminación de chupones y malezas.
Las aspersiones de los tratamientos se realizaron con atomizadores manuales (Magic,
500 mL), para asegurar la descarga exacta por árbol. Para cada tratamiento se utilizaron
equipos y materiales diferentes, con la finalidad de evitar la contaminación entre especies
de Trichoderma. Las aplicaciones se dirigieron a todo el árbol. Se hicieron cuatro
aplicaciones con una frecuencia mensual.
Tabla 1. Concentración de las suspensiones de aislados de Trichoderma, biopreparados
comerciales y fungicida (Óxido cuproso)
TRATAMIENTO CONCENTRACIÓN
DE LA SUSPENSIÓN
1. UEA T3 10⁸ esp mL-1
2. UEA T2 + UEA T3 10⁸ esp mL-1
3. Trichotic (Trichoderma Comercial ) 0,15 mL P.C árbol
4. Rhapsody 1.34 SC (B. subtillis) 1 mL P.C árbol
5. Óxido cuproso (Cobre Nordox 50) 3,0 g i.a árbol
6. Control Sin aplicación Se calculó un gasto de 200 mL de agua por árbol
Trichotic líquido (Trichoderma comercial, cuyo ingrediente activo son esporas de los hongos
Trichoderma harzianum,T. viride y T. koningii) con una concentración de 5 x 1010 esporas g-1,
200 g por 4 000 mL de agua.
Rhapsody (Bacillus subtilis cepa QST 713 con una concentración de 13,4 g L-1)
Quincenalmente se registró el número de mazorcas sanas y mazorcas con síntomas de
Moniliasis. Las mazorcas muertas que presentaron una longitud de hasta diez cm se
consideraron como Cherelle Wilt (marchitamiento prematuro); todas las mazorcas enfermas

Materiales y Métodos
50
se eliminaron en cada fecha de evaluación. Toda esta información fue registrada para cada
árbol de la parcela útil.
La eficacia técnica para cada uno de los tratamientos se calculó mediante la fórmula de
Abbott (1925), donde:
Los datos de porcentaje de infección y eficacia técnica fueron transformados mediante
2arcosen según Lerch (1977). Estas variables se analizaron estadísticamente, previa
comprobación de los supuestos de normalidad de los datos y la homogeneidad de las
varianzas. La prueba de comparación entre medias se realizó por la prueba de rangos
múltiples de Duncan (P≤0,05). Se empleó el paquete estadístico IBM SPSS Statistic versión
21,0 para Windows.

Resultados y Discusión
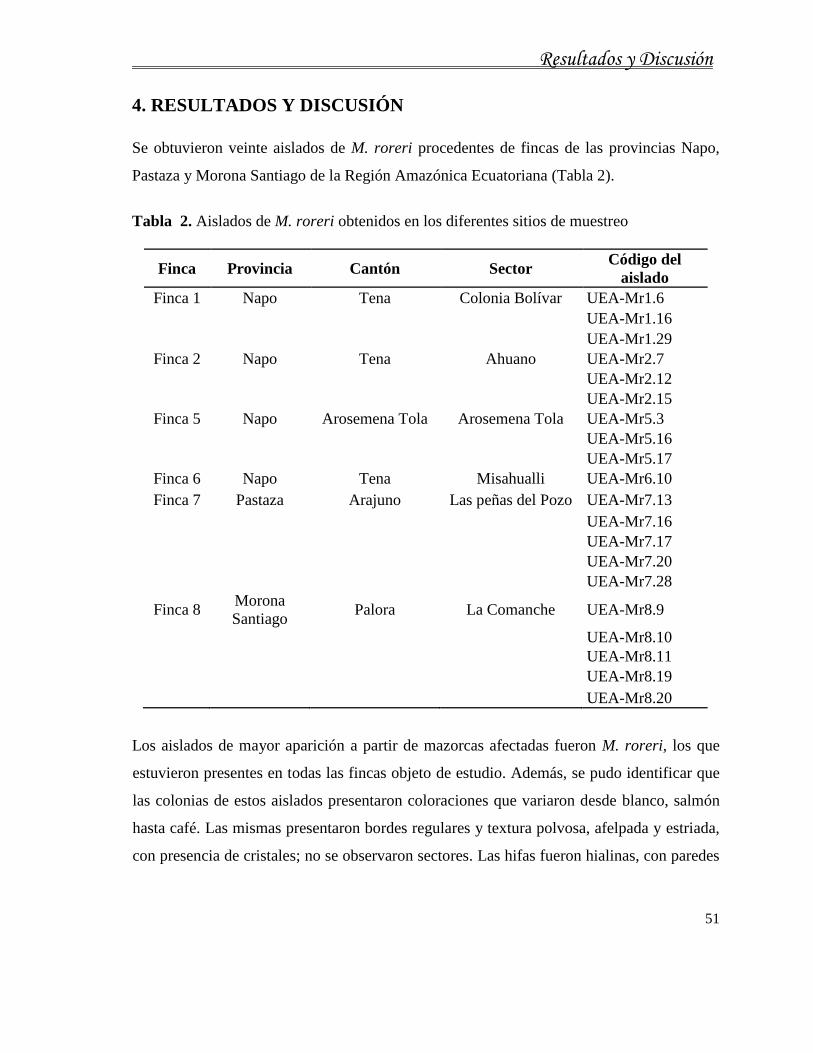
Resultados y Discusión
51
4. RESULTADOS Y DISCUSIÓN
Se obtuvieron veinte aislados de M. roreri procedentes de fincas de las provincias Napo,
Pastaza y Morona Santiago de la Región Amazónica Ecuatoriana (Tabla 2).
Tabla 2. Aislados de M. roreri obtenidos en los diferentes sitios de muestreo
Finca Provincia Cantón Sector Código del
aislado
Finca 1 Napo Tena Colonia Bolívar UEA-Mr1.6
UEA-Mr1.16
UEA-Mr1.29
Finca 2 Napo Tena Ahuano UEA-Mr2.7
UEA-Mr2.12
UEA-Mr2.15
Finca 5 Napo Arosemena Tola Arosemena Tola UEA-Mr5.3
UEA-Mr5.16
UEA-Mr5.17
Finca 6 Napo Tena Misahualli UEA-Mr6.10
Finca 7 Pastaza Arajuno Las peñas del Pozo UEA-Mr7.13
UEA-Mr7.16
UEA-Mr7.17
UEA-Mr7.20
UEA-Mr7.28
Finca 8 Morona
Santiago Palora La Comanche UEA-Mr8.9
UEA-Mr8.10
UEA-Mr8.11
UEA-Mr8.19
UEA-Mr8.20
Los aislados de mayor aparición a partir de mazorcas afectadas fueron M. roreri, los que
estuvieron presentes en todas las fincas objeto de estudio. Además, se pudo identificar que
las colonias de estos aislados presentaron coloraciones que variaron desde blanco, salmón
hasta café. Las mismas presentaron bordes regulares y textura polvosa, afelpada y estriada,
con presencia de cristales; no se observaron sectores. Las hifas fueron hialinas, con paredes

Resultados y Discusión
52
delgadas, septadas y algunas veces levemente irregulares, raramente ramificadas. Se
observaron conidióforos ramificados, dando lugar a cadenas maduras de esporas (Figura 6).
Las esporas fueron fácilmente separables y con paredes gruesas, de forma globosa,
subglobosas y algunas elipsoidales, con un largo de 8,76 µm y un ancho promedio de
8,18 µm. Los aislados de M. roreri cumplieron los postulados de Koch, lo que evidenció su
identidad.
Figura 6. Cadenas de esporas de M. roreri
Durante el aislamiento de M. roreri, se identificó cuatro géneros de hongos anamorfos,
asociados a mazorcas enfermas con síntomas de Moniliasis, correspondientes a los géneros:
Cylindrocladium, Dichobotrys, Colletotrichum y Phytophthora, procedentes de la provincia
de Napo (Anexo 8).
Se identificó al género Cylindrocladium asociado a mazorcas enfermas con síntomas de
Moniliasis. Se observaron conidióforos erectos, penicilados, con ramificaciones primarias,
vesícula terminal, y la presencia de masas de conidios monocelulares cilíndricos en sus
extremos. No se observó la producción de clamidosporas. Los conidióforos tuvieron un
largo promedio de 160 µm y 7 µm de ancho. Las ramas primarias, secundarias y terciarias
mostraron una longitud promedio de 14,01 µm, 18,24 µm y 10,4 µm, respectivamente. Las
fiálides tuvieron un largo promedio de 13,4 µm y 3,2 µm de ancho. Los conidios mostraron
un largo promedio de 40,5 µm y 3,5 µm de ancho (Figura 7).

Resultados y Discusión
53
Figura 7. Cylindrocladium sp. asociado a mazorcas de cacao
En relación con estos resultados Feitosa et al. (1986), demostraron que Cylindrocladium
floridanus (Sober et Seymour), fue patogénico en cacao, sin embargo estos autores no
describieron las dimensiones de las estructura asexuales. Por otra parte, Bastos y Bezerra
(2010) refirieron la presencia de Cylindrocladium scoparium Morg., lo que provocó la
pudrición de frutos de cacao, pero no efectuaron una caracterización morfológica de este
agente causal.
Asimismo, Evans et al. (2003b) informaron la presencia de aislados presuntamente
micoparasíticos pertenecientes al género Cylindrocladium que afectaba mazorcas de
Theobroma gileri Cuatrec. infectadas con Crinipellis roreri (Ciferri), sin embargo, no
describieron la morfología ni las dimensiones de las estructuras asexuales. Disponer de una
correcta caracterización morfológica de esta especie, aislada a partir de frutos de cacao con
síntomas de Moniliasis, permitirá discriminarla del resto de la micobiota existente en frutos
de cacao y evaluar en un futuro su papel ecológico en ese nicho.
Se aislaron hongos del género Dichobotrys asociados a mazorcas de cacao con síntomas de
Moniliasis. Se observó la presencia de conidióforos largos, ramificaciones dicotómicas
regulares, delgadas, presencia de células fértiles algo infladas, globosas, productoras de
conidios simultáneamente, con la presencia de conidios del tipo botryoblastosporas casi
globosos, hialinos, monocelulares unidos a dentículos cortos (Figura 8). Los conidióforos
tuvieron un largo promedio de 4,8 µm, sin embargo no se determinaron las dimensiones de
los conidios ni de sus hifas. Este es el primer informe de este género en cacao.

Resultados y Discusión
54
Figura 8. Dichobotrys sp. asociado a mazorcas de cacao
En la micobiota asociada a mazorcas de cacao se pudo observar la presencia de
Colletotrichum, pues se identificaron acérvulos similares a puntos negros incluidos en el
medio de cultivo PDA, en las placas de Petri. En los microcultivos se observó la presencia
de esporodoquios no bien diferenciados.
Los conidióforos fueron hialinos, simples, erectos, sustentando un conidio en cada ápice de
la fiálides. Se determinó la presencia de conidios hialinos, cilíndricos, ovoides y
monocelulares (Figura 9). En los cultivos realizados en medios sintéticos no se observaron
setas, sin embargo en lesiones ocasionadas en frutos sí se pudieron apreciar. Las hifas
tuvieron un ancho promedio de 2,47 µm, los conidióforos mostraron un largo promedio de
9,56 µm y ancho de 3,82 µm. El largo promedio de los conidios fue de 7,07 µm y un ancho
de 2,37 µm. En la descripción de las características del género, estas fueron similares a las
referidas por Rojas et al. (2010), a pesar de que en el presente trabajo las dimensiones de
los conidios y conidióforos fueron menores.
Figura 9. Colletotrichum sp. asociado a mazorcas de cacao

Resultados y Discusión
55
Se observó la presencia de P. palmivora, con micelio cenocítico, sin la formación de
estructuras reproductivas sexuales, presencia de esporangios diferenciados que contenían
zoosporas. Se observaron abundantes esporangios papilados de forma ovoide que tenían
como promedio 40,65 µm de largo y 23,62 µm de ancho (Figura 10). Estas dimensiones
fueron similares a las encontradas por Stamps (1985), quien denotó que dichas estructuras
se forman fácilmente en medios de cultivo y que los esporangios presentan paredes bien
definidas prevaleciendo las formas ovoides a elipsoidales.
Figura 10. Phytophthora sp. asociado a mazorcas de cacao
Mfegue et al. (2012) determinaron que en frutos de cacao en África, la principal especie
causante de la pudrición de la mazorca de cacao fue P. megakarya en lugar de
P. palmivora que prevalece en América. Sin embargo, lo anterior sugiere la posibilidad de
la existencia de varias especies de este género, causando una misma sintomatología a nivel
de mazorcas de cacao, por lo que realizar la caracterización sobre la base de caracteres
morfológicos resulta de interés para discriminar los daños que puedan provocar estas
especies en la producción de cacao en la Amazonía ecuatoriana.
Las características de M. roreri y Colletotrichum sp. coincidieron con las descritas por
Maridueña (2011), en un estudio similar; sin embargo este autor identificó una mayor
diversidad de especies de hongos filamentosos asociados a mazorcas de cacao. Lo
informado en esta investigación se corresponde con lo descrito por Urdaneta y Delgado
(2007), quienes identificaron los géneros Moniliophthora, Cylindrocladium, Colletotrichum
y Phytophthora en la micobiota del filoplano del cacao en Venezuela.

Resultados y Discusión
56
4.1. Caracterización cultural, morfológica, fisiológica, molecular y patogénica de
aislados de M. roreri
4.1.1. Caracterización cultural de M. roreri
Todas las colonias de los aislados fueron capaces de crecer de forma radial y sus
características culturales fueron similares en medio de cultivo V8 modificado. De los 20
aislados identificados de M. roreri, UEA-Mr8.9 y UEA-Mr8.10 procedentes de Morona
Santiago, La Comanche, fueron los únicos capaces de cubrir la placa de Petri (90 mm Ø) a
los 12 días de incubación a 26±2 °C, mostrando así la mayor velocidad de crecimiento
micelial en medio de cultivo V8 modificado. A su vez los aislados UEA-Mr5.3,
UEA-Mr5.16, UEA-Mr5.17 provenientes de Arosemena Tola, Napo; UEA-Mr7.28,
UEA-Mr7.13 Palora, Morona Santiago y UEA-Mr8.11 Arajuno, Pastaza lo hicieron a los
15 días, en las mismas condiciones (Figura 11).
Figura 11. Crecimiento micelial de los aislados más destacados de M. roreri por su
velocidad de crecimiento en medio de cultivo V8 modificado a 26±2 ºC a los 12 y 15 días
de incubación. Barras con letras diferentes, indican diferencias significativas por la prueba
de Kruskal-Wallis/Mann Whitney P ≤0,05 (n=10).

Resultados y Discusión
57
Estos resultados están en concordancia con los de Villavicencio (2010), quien informó un
crecimiento promedio del micelio de M. roreri de 90,00 mm a los 15 días, en aislados
procedentes de la costa ecuatoriana. Por otra parte, Arbeláez (2010) obtuvo valores de
41,51 mm de rango promedio al día 13 de evaluación en aislados colombianos de
M. roreri.
Los aislados UEA-Mr7.16, UEA-Mr7.17, UEA-Mr7.20, UEA-Mr8.19 y UEA-Mr8.20
cubrieron las placas de Petri (90 mm Ø) a los 16-17 días, mientras que UEA-Mr1.6, UEA-
Mr1.16, UEA-Mr1.29, UEA-Mr2.7, UEA-Mr2.12, UEA-Mr2.15 y UEA-Mr6.10 lo
hicieron en 18 días.
La velocidad de crecimiento radial de los aislados de M. roreri fue de 7,30 mm/día. Estos
resultados son cercanos a los informados por Uquillas (2004), quien determinó que la
velocidad del crecimiento radial de M. roreri fue de 9,81 mm/día al estudiar la inducción de
la germinación para mejorar la eficacia de dos agentes antagonistas para el control de la
Moniliasis.
Se apreció una considerable diversidad en las tonalidades de los colores observados en los
aislados, los que fueron característicos de M. roreri, variando según los días de
crecimiento, detectándose colores como: blanco, salmón y café, debido a la formación
masiva de esporas con masa conidial pulverulenta.
Las tonalidades de colores del micelio de las colonias de los aislados, variaron según la
edad de las mismas; en el caso de los aislados procedentes de Napo, se apreció una
coloración blanca en todo su desarrollo. Las colonias de los aislados procedentes de las
provincias de Pastaza y Morona Santiago presentaron inicialmente una coloración blanca,
pasando por tonalidad salmón y terminaron su desarrollo con coloración café, lo cual
coincide con lo informado por Evans et al. (1978), Phillips et al. (2006) y Arbeláez (2010).
El reverso de las colonias de los aislados de M. roreri presentó diferentes tonalidades de
color, a medida que las colonias fueron creciendo, pasando por blanco, salmón y café.

Resultados y Discusión
58
Todas las colonias de los aislados mostraron color salmón en el reverso, excepto
UEA-Mr8.11 y UEA-Mr8.19, que presentaron coloración café, lo que coincide con lo
informado por Evans et al. (1978).
Las tonalidades de colores observados en las colonias son características de M. roreri,
según lo manifestado por Suárez (2006). Las colonias de este hongo fitopatógeno tienen
una considerable variabilidad en cuanto a tonos de coloración del micelio que varían del
blanco, salmón, hasta el café (Phillips et al., 2006), por lo que este parámetro no permite
asegurar con certeza que los fenotipos de los aislados obtenidos pertenezcan a una misma
especie (Anexo 9).
Las colonias de todos los aislados presentaron bordes regulares, similar a lo informado por
Phillips et al. (2006) y que difiere de lo referido por Arbeláez (2010), quien determinó en
Colombia que el borde más representativo fue el plumoso.
Todos los aislados presentaron crecimiento superficial y compacto; características
informadas por Phillips et al. (2006). No existió presencia de pigmentos difusibles en el
medio de cultivo. Los sectores estuvieron ausentes en la mayoría de las colonias (90%),
excepto en los aislados UEA-Mr.8.19 y UEA-Mr.1.6 (Tabla 3), siendo más notorios a
medida que envejeció el cultivo en los aislados, lo cual coincide con lo informado por
Arbeláez (2010).
En los aislados procedentes de Napo (Colonia Bolívar) se observó textura estriada 15%, en
los aislados de Napo (Ahuano, Arosemena Tola y Misahualli) textura afelpada 35%, y en
los aislados de Pastaza (Arajuno) y Morona Santiago (Palora), textura polvosa que
correspondió al 50% del total de los aislados (Tabla 3). Estos resultados difieren con lo
referido por Arbeláez (2010), quien obtuvo una textura afelpada para el 79,3% de los
aislados en un trabajo similar.
No se observó la presencia de líquido de transpiración en los aislados de M. roreri, excepto
en los aislados UEA-Mr1.6, UEA-Mr1.16 y UEA-Mr1.29, donde se presentaron en forma

Resultados y Discusión
59
de pequeñas gotas (muy frecuentes) y gotas de mayor tamaño (menos frecuentes)
(Tabla 3). En todos los casos los líquidos transpirados fueron incoloros.
Tabla 3. Características culturales de Moniliophthora roreri en medio de cultivo V8
modificado
Aislados Sectores Textura Líquido de
transpiración Cristales
UEA.Mr1.6 Presentes Estriada Con líquido Ausentes
UEA.Mr1.16 Ausentes Estriada Con líquido Presentes
UEA.Mr1.29 Ausentes Estriada Con líquido Presentes
UEA.Mr2.7 Ausentes Afelpada Sin líquido Presentes
UEA.Mr2.12 Ausentes Afelpada Sin líquido Presentes
UEA.Mr2.15 Ausentes Afelpada Sin líquido Presentes
UEA.Mr5.3 Ausentes Afelpada Sin líquido Presentes
UEA.Mr5.16 Ausentes Afelpada Sin líquido Presentes
UEA.Mr5.17 Ausentes Afelpada Sin líquido Presentes
UEA.Mr6.10 Ausentes Afelpada Sin líquido Presentes
UEA.Mr7.13 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr7.16 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr7.17 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr7.20 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr7.28 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr8.9 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr8.10 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr8.11 Ausentes Polvosa Sin líquido Ausentes
UEA.Mr8.19 Presentes Polvosa Sin líquido Ausentes
UEA.Mr8.20 Ausentes Polvosa Sin líquido Ausentes
Se observó presencia de cristales en los aislados procedentes de Napo, excepto en el aislado
UEA-Mr1.6. Para los aislados procedentes de Arajuno, Pastaza y Palora, Morona Santiago,
no se observó la presencia de cristales durante su desarrollo (Tabla 3).
En relación con estas últimas variables descritas, la literatura científica consultada no
mostró informe previo donde se haya evaluado este aspecto, con vistas a caracterizar las
colonias de M. roreri.

Resultados y Discusión
60
M. roreri es un hongo variable respecto a sus características culturales. El predominio de
las esporas como estructuras infectivas en condiciones naturales (Evans et al., 2003b)
implica una amplia variabilidad genética debido a que dichas estructuras son el resultado
del proceso de reproducción sexual donde tienen lugar eventos recombinatorios (Tiburcio
et al., 2010; Meinhardt et al., 2014). Todos estos eventos de variación genética pueden
explicar las diferencias observadas respecto a los caracteres culturales en los aislados de la
Amazonía ecuatoriana de M. roreri.
4.1.2. Caracterización morfológica de M. roreri
Todos los aislados presentaron hifas, conidios y conidióforos que se correspondieron con
las características descritas por Evans et al. (1978) para el estado asexual de M. roreri.
Las características morfológicas de los aislados de M. roreri en medio V8 modificado
fueron similares. En este medio de cultivo se desarrollaron tanto micelio como esporas para
todos los aislados; similar a lo reportado por otros autores que han usado este medio de
cultivo; el crecimiento fue en forma de tapete de aspecto lanoso (Evans et al., 1978;
Phillips et al., 2006; García, 2007; Arbeláez, 2010; Villavicencio, 2010; Cuervo-Parra
et al., 2011). Las hifas tuvieron paredes delgadas, septadas y levemente irregulares,
ramificadas ocasionalmente, con hinchamientos, hialinas. Los conidióforos fueron
ramificados dando lugar a cadenas maduras de esporas (Figura 12).
Figura 12. Cadenas maduras de esporas pertenecientes al aislado UEA-Mr8.9.
Microfotografía con un aumento de 400X

Resultados y Discusión
61
Las esporas presentaron coloración amarillo pálido y en masa color café oscuro, fácilmente
separables de las colonias, heteromórficas y con paredes gruesas (Figura 13).
Figura 13. Estructuras basipetalas conoidiales pertenecientes al aislado UEA-Mr8.9.
Microfotografía con un aumento de 400X
Las formas de las esporas obtenidas en medio de cultivo V8 modificado fueron globosas,
subglobosas y elipsoidales (Figura 14).
Figura 14. Esporas con formas globosas, subglobosas y elipsoidales, pertenecientes al
aislado UEA-Mr8.9. Microfotografía con un aumento de 1000X
La mayoría de los aislados produjeron esporas de formas globosas (7-9 µm de diámetro),
sin embargo se encontraron formas subglobosas (8-14 µm de diámetro) y elipsoidales
(8-20 x 6-17 µm), obtenidos a partir del medio de cultivo V8 modificado.
Resultados informados por Holliday (1970), Evans et al. (1978), Suárez (2006), Phillips-
Mora et al. (2015) describieron formas de esporas de M. roreri desde globosas,
subglobosas a elipsoidales.

Resultados y Discusión
62
Este planteamiento es confirmado por Evans et al. (1978), quienes describen dimensiones
cercanas en la descripción taxonómica de M. roreri. González y Roble (2014), en una
caracterización de M. roreri, en El Salvador, describieron esporas globosas (4-5 µm),
subglobosas (7-8 µm) y elípticas (9-11 µm); lo que difiere con las dimensiones de las
esporas encontradas en esta investigación.
En la Tabla 4 se muestran las dimensiones de ancho y largo promedio de las esporas de los
aislados obtenidos de M. roreri, donde se aprecia que el aislado que presentó el mayor
valor de ancho de esporas promedio fue UEA-Mr5.3 con un valor de 8,93 µm, y el aislado
que presentó el menor valor fue UEA-Mr.2.7 con 7,34 µm. El aislado que presentó un
mayor valor promedio para el largo de las esporas fue UEA-Mr8.9 con 10,78 µm; mientras
el aislado de menor valor promedio fue UEA-Mr2.7 de 7,42 µm de largo.
Tabla 4. Características morfológicas de las esporas producidas por aislados de M. roreri
Aislados Ancho de las esporas
(µm)
Largo de las esporas
(µm)
UEA-Mr1.6 7,527 ± 0,161 efg 7,771 ± 0,152 efg
UEA-Mr1.16 7,834 ± 0,133 cdefg 7,967 ± 0,133 efg
UEA-Mr1.29 7,666 ± 0,167 defg 7,679 ± 0,175 fg
UEA-Mr2.7 7,340 ± 0,137 g 7,428 ± 0,148 g
UEA-Mr2.12 7,419 ± 0,137 fg 7,537 ± 0,136 g
UEA-Mr2.15 7,342 ± 0,137 g 7,458 ± 0,128 g
UEA-Mr5.3 8,930 ± 0,206 a 9,811 ± 0,337 abc
UEA-Mr5.16 8,587 ± 0,175 abcd 9,750 ± 0,411 abc
UEA-Mr5.17 8,687 ± 0,206 abc 9,460 ± 0,331 abcd
UEA-Mr6.10 7,407 ± 0,156 fg 7,657 ± 0,169 fg
UEA-Mr7.13 7,922 ± 0,220 bcdefg 8,196 ± 0,246 defg
UEA-Mr7.16 8,477 ± 0,192 abcde 8,840 ± 0,218 cdefg
UEA-Mr7.17 8,704 ± 0,171 abc 9,027 ± 0,187 cdef
UEA-Mr7.20 8,330 ± 0,212 abcdef 8.731 ± 0,206 cdefg
UEA-Mr7.28 8,564 ± 0,235 abcd 9.453 ± 0,274 abcd
UEA-Mr8.9 8,663 ± 0,218 abc 10,781 ± 0,605 a
UEA-Mr8.10 8,786 ± 0,256 ab 10,557 ± 0,603 ab
UEA-Mr8.11 8,719 ± 0,213 abc 9,229 ± 0,266 bcde
UEA-Mr8.19 8,112 ± 0,158 abcdefg 8,803 ± 0,219 cdefg
UEA-Mr8.20 8,678 ± 0,201 abc 9,110 ± 0,185 bcdef Medias con letras diferentes en el sentido de la columna difieren para P≤0,05 por la prueba de C de
Dunnett

Resultados y Discusión
63
Evans et al. (2003a), obtuvieron valores medios para las esporas de M. roreri muy
próximos a los del presente trabajo, en un aislado procedente de la provincia de Napo,
Ecuador. Así mismo las características de las esporas de los aislados se correspondieron
con las descritas por Phillips-Mora et al. (2015) para la especie de M. roreri, quienes
describieron valores entre 7-10 µm de largo y 8-11 µm de µm de ancho, al informar por
primera vez la presencia de M. roreri en Bolivia.
Contrario a estos resultados Cuervo-Parra et al. (2011) en México, verificaron que las
esporas de M. roreri presentaron 8-19 x 5-11 µm en la caracterización morfológica del
hongo patógeno en Tabasco, México.
4.1.3. Caracterización fisiológica de M. roreri
El aislado UEA-Mr8.9 correspondiente a la finca 8 (Sector La Comanche) fue el que
presentó mayor crecimiento micelial en la caracterización cultural de M. roreri, y fue con el
que se realizaron todos los ensayos en esta caracterización.
4.1.3.1. Influencia del pH en el crecimiento micelial
Se observó crecimiento micelial del aislado UEA-Mr8.9 en todos los valores de pH
evaluados (desde 6,0 hasta 7,5). El pH 6,5 fue el que más favoreció el crecimiento micelial
del hongo patógeno con 8,2 mg de masa seca del micelio, a diferencia del pH 7,5 que fue
donde obtuvieron los menores valores con 0,35 mg, y que a su vez no difirió
estadísticamente del pH 6,0 con 0,42 mg. Valores intermedios se obtuvieron a pH 7,0 con
0,51 mg de masa seca del micelio (Figura 15).
En condiciones controladas M. roreri crece fácilmente en un intervalo de pH entre 4,5 a
7,5. Para el desarrollo micelial el pH está entre 5,0 y 6,5; mientras que para la formación de
esporas requiere pH 7,5 (Arbeláez, 2010).
En este sentido Villegas (1979), en un estudio desarrollado en la Estación Experimental
Tulio Ospina del ICA en Colombia, evaluó la influencia del pH en el crecimiento micelial
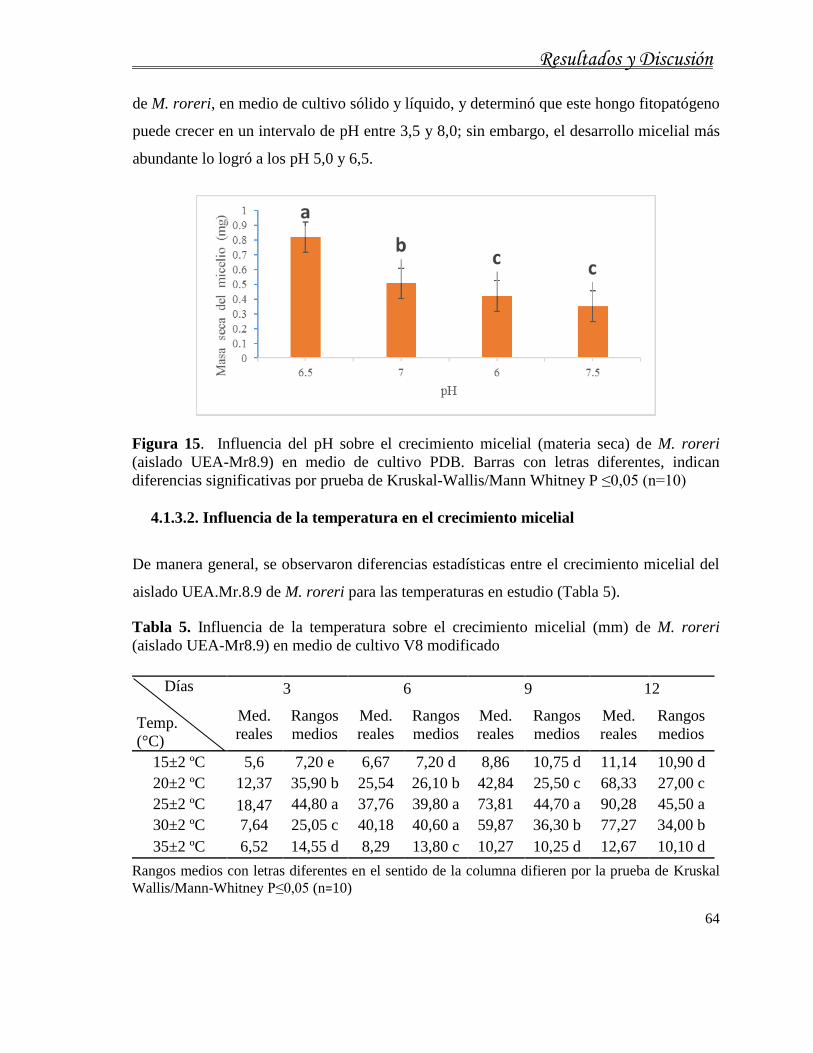
Resultados y Discusión
64
de M. roreri, en medio de cultivo sólido y líquido, y determinó que este hongo fitopatógeno
puede crecer en un intervalo de pH entre 3,5 y 8,0; sin embargo, el desarrollo micelial más
abundante lo logró a los pH 5,0 y 6,5.
Figura 15. Influencia del pH sobre el crecimiento micelial (materia seca) de M. roreri
(aislado UEA-Mr8.9) en medio de cultivo PDB. Barras con letras diferentes, indican
diferencias significativas por prueba de Kruskal-Wallis/Mann Whitney P ≤0,05 (n=10)
4.1.3.2. Influencia de la temperatura en el crecimiento micelial
De manera general, se observaron diferencias estadísticas entre el crecimiento micelial del
aislado UEA.Mr.8.9 de M. roreri para las temperaturas en estudio (Tabla 5).
Tabla 5. Influencia de la temperatura sobre el crecimiento micelial (mm) de M. roreri
(aislado UEA-Mr8.9) en medio de cultivo V8 modificado
Días
Temp.
(°C)
3 6 9 12
Med.
reales
Rangos
medios
Med.
reales
Rangos
medios
Med.
reales
Rangos
medios
Med.
reales
Rangos
medios
15±2 ºC 5,6 7,20 e 6,67 7,20 d 8,86 10,75 d 11,14 10,90 d
20±2 ºC 12,37 35,90 b 25,54 26,10 b 42,84 25,50 c 68,33 27,00 c
25±2 ºC 18,47 44,80 a 37,76 39,80 a 73,81 44,70 a 90,28 45,50 a
30±2 ºC 7,64 25,05 c 40,18 40,60 a 59,87 36,30 b 77,27 34,00 b
35±2 ºC 6,52 14,55 d 8,29 13,80 c 10,27 10,25 d 12,67 10,10 d
Rangos medios con letras diferentes en el sentido de la columna difieren por la prueba de Kruskal
Wallis/Mann-Whitney P≤0,05 (n=10)

Resultados y Discusión
65
La temperatura de 25±2 °C fue la que mostró los mayores valores de crecimiento micelial
del hongo patógeno en todos los tiempos de evaluación, a diferencia de las temperaturas de
15 y 35±2 °C, que fue donde se hallaron los menores valores para esta variable. Este
resultado resalta la importancia del conocimiento de la temperatura de incubación óptima
en el crecimiento micelial de M. roreri en condiciones controladas, catalogando así a este
como un patógeno psicrófilo facultativo.
Estos resultados coincidieron con los reportados por Villamil et al. (2012), quienes
obtuvieron un crecimiento micelial favorable con el empleo del medio de cultivo PDA a
25 °C, para la producción in vitro de micelio de M. roreri.
Por otra parte, Evans et al. (1978) en la descripción de las características morfológicas de
M. roreri, informaron que las temperaturas de crecimiento óptimo fueron de 25 a 26 °C
con un máximo de 33 °C.
Phillips-Mora (2003), evaluó el efecto in vitro de tres temperaturas (18, 24 y 30 °C) sobre
el crecimiento micelial de 15 aislados de M. roreri procedentes de los grupos genéticos
más importantes de este hongo fitopatógeno en América, y determinó que las temperaturas
tuvieron un efecto significante sobre el crecimiento de los aislados de M. roreri, asimismo
sitúo el valor óptimo para crecer y esporular a 24 °C.
4.1.3.3 Influencia de diferentes regímenes de luz en el crecimiento micelial
De manera general, se observaron diferencias estadísticas entre el crecimiento micelial del
aislado UEA.Mr.8.9 de M. roreri para las condiciones de iluminación en estudio
(Tabla 6). La condición de oscuridad fue la que mostró los mayores valores de
crecimiento micelial del hongo patógeno en todos los tiempos de evaluación a diferencia
de las otras variantes en estudio, en las cuales existieron variaciones en la respuesta según
el tiempo de evaluación. En condiciones de luz ultravioleta se apreció el crecimiento
micelial más lento a los 12 y 15 días de evaluación.
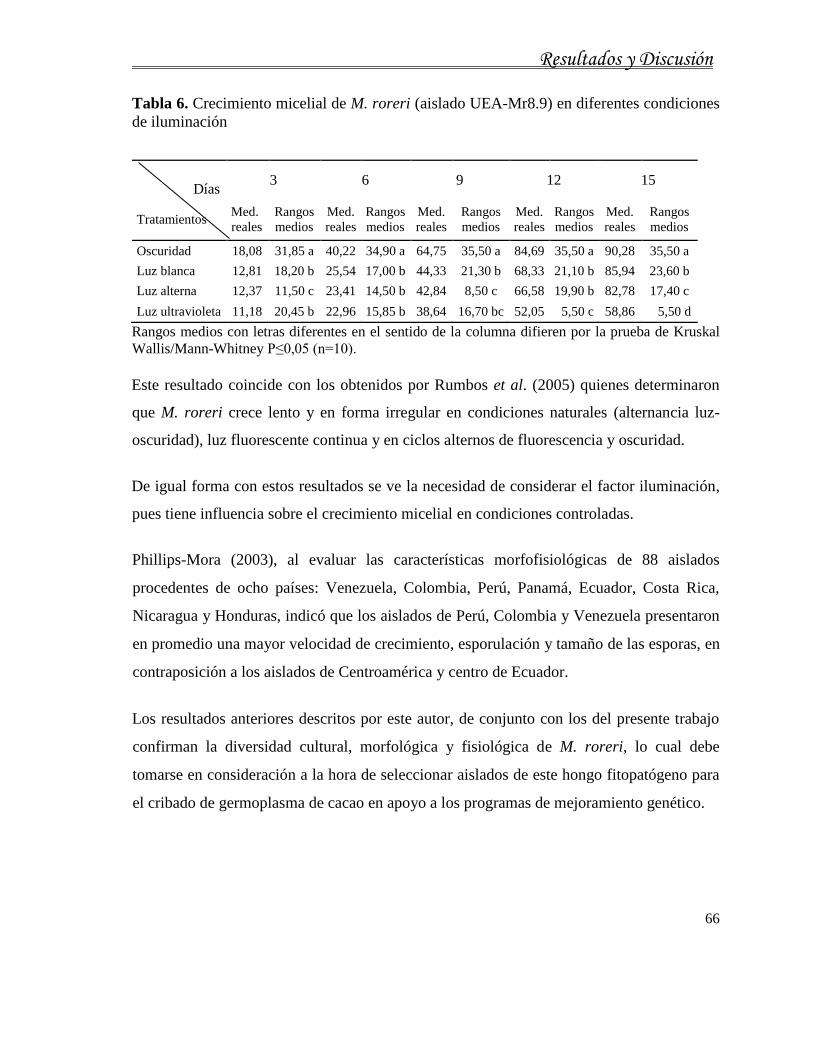
Resultados y Discusión
66
Tabla 6. Crecimiento micelial de M. roreri (aislado UEA-Mr8.9) en diferentes condiciones
de iluminación
Días
3 6 9 12 15
Tratamientos Med.
reales
Rangos
medios
Med.
reales
Rangos
medios
Med.
reales
Rangos
medios
Med.
reales
Rangos
medios
Med.
reales
Rangos
medios
Oscuridad 18,08 31,85 a 40,22 34,90 a 64,75 35,50 a 84,69 35,50 a 90,28 35,50 a
Luz blanca 12,81 18,20 b 25,54 17,00 b 44,33 21,30 b 68,33 21,10 b 85,94 23,60 b
Luz alterna 12,37 11,50 c 23,41 14,50 b 42,84 8,50 c 66,58 19,90 b 82,78 17,40 c
Luz ultravioleta 11,18 20,45 b 22,96 15,85 b 38,64 16,70 bc 52,05 5,50 c 58,86 5,50 d
Rangos medios con letras diferentes en el sentido de la columna difieren por la prueba de Kruskal
Wallis/Mann-Whitney P≤0,05 (n=10).
Este resultado coincide con los obtenidos por Rumbos et al. (2005) quienes determinaron
que M. roreri crece lento y en forma irregular en condiciones naturales (alternancia luz-
oscuridad), luz fluorescente continua y en ciclos alternos de fluorescencia y oscuridad.
De igual forma con estos resultados se ve la necesidad de considerar el factor iluminación,
pues tiene influencia sobre el crecimiento micelial en condiciones controladas.
Phillips-Mora (2003), al evaluar las características morfofisiológicas de 88 aislados
procedentes de ocho países: Venezuela, Colombia, Perú, Panamá, Ecuador, Costa Rica,
Nicaragua y Honduras, indicó que los aislados de Perú, Colombia y Venezuela presentaron
en promedio una mayor velocidad de crecimiento, esporulación y tamaño de las esporas, en
contraposición a los aislados de Centroamérica y centro de Ecuador.
Los resultados anteriores descritos por este autor, de conjunto con los del presente trabajo
confirman la diversidad cultural, morfológica y fisiológica de M. roreri, lo cual debe
tomarse en consideración a la hora de seleccionar aislados de este hongo fitopatógeno para
el cribado de germoplasma de cacao en apoyo a los programas de mejoramiento genético.

Resultados y Discusión
67
4.1.4. Identificación molecular de aislados de M. roreri
La implementación del método de extracción de ADN genómico utilizado demostró ser
eficiente para la obtención de adecuadas concentraciones y pureza de ADN de los aislados
objeto de estudio (Anexo 10). Aunque en algunos aislados las concentraciones fueron algo
bajas, la pureza fue adecuada, por lo que se decidió el envío del ADN para su
amplificación.
Los cebadores NS1 y NS8 demostraron que amplifican un fragmento de aproximadamente
1,8 kb dentro de la subregión 18S ARNr. Esta subregión de conjunto con las regiones
intergénicas ITS1 e ITS2 están informadas como las más conservadas dentro del genoma de
los eucariontes, y es un fragmento diana para la identificación molecular (Gardes y Bruns,
1993).
Todas las secuencias tuvieron un porcentaje de similitud con la base de datos GenBank
entre 99 y 100%, lo que demuestra lo efectivo de esta región para la identificación hasta el
nivel de especie.
Para definir una especie utilizando este marcador se aplica la regla universal descrita por
Stackebrandt y Goebel (1994). Aquellos aislados que tengan una diferencia mayor a 97%
en la secuencia de nucleótidos de este gen serán considerados como especies diferentes.
El 100% de los aislados analizados fueron identificados molecularmente como M. roreri,
resultado esperado teniendo en cuenta la caracterización morfo-cultural realizada.
No obstante, los aislados en estudio mostraron similitud por Blastn con tres cepas
diferentes de este hongo descritas con anterioridad, y predominio de las cepas MRO1
(36,4%) y MROCP (36,4%), como se describe en la Tabla 7.

Resultados y Discusión
68
Tabla 7. Identificación molecular de aislados de M. roreri
Código de
aislado FASTA más cercana N° Accesión
% de
identidad Referencia
UEA-Mr1.6 Moniliophthora roreri aislado MROCP KM998972.1 99 Concepción-Brindis et al. (2015)
UEA-Mr2.7 Moniliophthora roreri cepa MRO1 JF730693.1 99 Sánchez et al. (2011)
UEA-Mr5.3 Moniliophthora roreri aislado MROCP KM998972.1 99 Concepción-Brindis et al. (2015)
UEA-Mr5.17 Moniliophthora roreri aislado MROCP KM998972.1 99 Concepción-Brindis et al. (2015)
UEA-Mr6.10 Moniliophthora roreri cepa C21 AY916745.1 100 Aime y Phillips-Mora (2005)
UEA-Mr7.13 Moniliophthora roreri cepa MRO1 JF730693.1 99 Sánchez et al. (2011)
UEA-Mr7.20 Moniliophthora roreri cepa MRO1 JF730693.1 99 Sánchez et al. (2011)
UEA.Mr8.9 Moniliophthora roreri cepa C21 AY916745.1 100 Aime y Phillips-Mora (2005)
UEA-Mr8.10 Moniliophthora roreri cepa MRO1 JF730693.1 99 Sánchez et al. (2011)
UEA-Mr8.19 Moniliophthora roreri aislado MROCP KM998972.1 99 Concepción-Brindis et al. (2015)
Existe gran diversidad de métodos moleculares usados para la identificación de M. roreri.
Aime y Phillips-Mora (2005) indican que la identificación de M. roreri se ha realizado a
través de la amplificación por PCR de la secuencia de la subregión ITS, así como el 28S
ARNr. Sánchez (2011) demuestra que los marcadores moleculares del fragmento del 18S
ARNr y la región ITS1-5,8S-ITS2 son eficaces y eficientes para la identificación molecular
de cepas del hongo M. roreri. En este sentido, Phillips-Mora et al. (2015) y Suárez (2016)
identificaron molecularmente aislados de M. roreri extraídos de T. cacao en Bolivia y
Colombia respectivamente, mediante marcadores moleculares de ITS.
Por otra parte, White et al. (1990) manifiestan que las regiones codificantes de los genes
nucleares 18S, 5,8S y 28S ARNr evolucionan lentamente y están altamente conservadas,
por lo que se utilizan para obtener información sobre las relaciones filogenéticas. No
obstante, la subregión ITS es en la actualidad la región más secuenciada del ADN del
genoma en hongos, y ha sido usada para explicar o inferir afinidades filogenéticas de
grupos relacionados (Lee y Taylor, 1992).
Mediante la secuenciación de la subregión 18S ARNr se logró la identificación de los
aislados monospóricos objetos de estudio, los cuales pertenecen a la especie M. roreri. Por
vez primera en la Amazonía ecuatoriana se usan herramientas moleculares para el

Resultados y Discusión
69
diagnóstico de este hongo, lo que significa un aporte metodológico para futuras
investigaciones.
4.1.5. Caracterización patogénica y agresividad de los aislados de M. roreri
Los 20 aislados de M. roreri fueron patogénicos, capaces de reproducir los síntomas
característicos de la Moniliasis, en los frutos inoculados de cacao Nacional.
Estos fueron similares a los observados en condiciones naturales de campo, donde el
período de incubación varió de tres a ocho semanas de acuerdo con la edad del fruto,
mientras que a nivel de laboratorio los síntomas se desarrollaron con mayor velocidad (en
un período de 12 días), lo cual coincide con lo informado por Ampuero (1967); Suárez
(1971); Barros (1977) y Villavicencio (2010).
La evaluación diaria de la severidad externa de las mazorcas inoculadas evidenció los
siguientes síntomas: al día dos de la inoculación se manifestó el grado 1 (puntos aceitosos),
el día cinco presentó grado 2 (tumefacciones o clorosis). En el día siete se observó grado 3
(mancha necrótica).
En la evaluación realizada al día 11 se observó el grado 4 (micelio presente en un 25% de la
mancha), y al día 12 grado 5 (micelio en más del 25% de la mancha) en la mayoría de los
aislados, excepto para UEA-Mr1.6, UEA-Mr2.7, UEA-Mr6.10, UEA-Mr7.13, quienes
manifestaron los síntomas de grado 5 a los ocho días.
Este último resultado evidenció la capacidad de agresividad de estos aislados, al reproducir
la enfermedad y síntomas más severos (necrosis más esporulación en un área mayor de la
cuarta parte de la superficie necrótica) en un menor período de tiempo, de manera similar a
Villavicencio (2010) quien informó de un período de incubación de ocho días al inocular
frutos de cacao con diez aislados virulentos de M. roreri procedentes de la costa
ecuatoriana.

Resultados y Discusión
70
Por otro lado, luego de los 15 días, no se observaron diferencias en los síntomas de las
mazorcas inoculadas, siendo su comportamiento uniforme en el transcurso de los días
(Figura 16).
Los primeros síntomas internos se manifestaron a los cinco días de la inoculación, cuando
se observó grado 1 (1-20% área necrosada), a los siete días se manifestó el grado 2
(21-40% área necrosada), a los nueve días se presentó el grado 3 (41-60% área necrosada),
a los 11 días se observó grado 4 (61-80% área necrosada) y a los 15 días la mazorca se
encontró infectada completamente hasta grado 5 (100% área necrosada). En relación con
esta variable en la literatura científica no se han encontrado referencias previas al uso de la
misma para determinar la severidad interna de aislados de M. roreri, lo cual implica un
aporte del presente trabajo con vistas a enriquecer la caracterización patogénica de este
hongo (Figura 17).
Figura 17. Severidad interna acorde a la escala de Sánchez et al. (1987) del daño ocasionado por
aislados de M. roreri en ensayo monocíclico a) grado 0 b) grado 1 c) grado 2 d) grado 3 e) grado 4
f) grado 5
a
b c
d e f
Figura 16. Severidad externa acorde con la escala de Brenes (1983) del daño ocasionado por
aislados de M. roreri en ensayo monocíclico a) puntos aceitosos (dos días) b) tumefacción o
clorosis (cinco días) c) mancha necrótica (siete días) d) micelio en un 25% de la mancha (once
días) e) micelio en más del 25% de la mancha (doce días)
a
b
c
d
e

Resultados y Discusión
71
En el caso de los aislados UEA-Mr6-10, UEA-Mr1.16, UEA-Mr1.6, UEA-Mr7.13, UEA-
Mr7.17 y UEA-Mr2.7 no se encontraron diferencias estadísticas en cuanto a su agresividad
a los seis días, a pesar del aislado UEA-Mr6-10 presentara el mayor valor de ABCPE, y
UEA-Mr7.20 el menor valor (Tabla 8).
Tabla 8. Área Bajo la Curva del Progreso de la Enfermedad en el período de 1 hasta 6 días
posteriores a la inoculación
Aislados ABCPE Rangos medios
UEA-Mr6.10 202 174,7 a
UEA-Mr1.16 132 166,7 ab
UEA-Mr1.6 134 163,5 ab
UEA-Mr7.13 196 161,1 ab
UEA-Mr7.17 128 130,5 ab
UEA-Mr2.7 132 125,7 ab
UEA-Mr8.11 114 115,1 b
UEA-Mr5.17 94 112,5 b
UEA-Mr8.19 98 112,3 b
UEA-Mr2.12 90 102,5 b
UEA-Mr8.10 96 102,5 b
UEA-Mr8.20 78 99,7 b
UEA-Mr8.9 76 95,1 b
UEA-Mr5.3 96 93,7 b
UEA-Mr7.16 128 87,9 b
UEA-Mr1.29 78 83,1 b
UEA-Mr7.28 64 82,7 bc
UEA-Mr5.16 62 75,5 bc
UEA-Mr215 58 70,9 bc
UEA-Mr7.20 36 47,3 c
Rangos medios con letras diferentes difieren por la prueba de Kruskal Wallis/Mann-Whitney
P≤0,05 (n=10).
El Área Bajo la Curva del Progreso de la Enfermedad en el intervalo continuo de 1 a 17
días permitió encontrar una respuesta diferencial en la agresividad de los aislados de
M.roreri, encontrándose en los aislados UEA-Mr6.10, UEA-Mr1.6, UEA-Mr7.13 la mejor
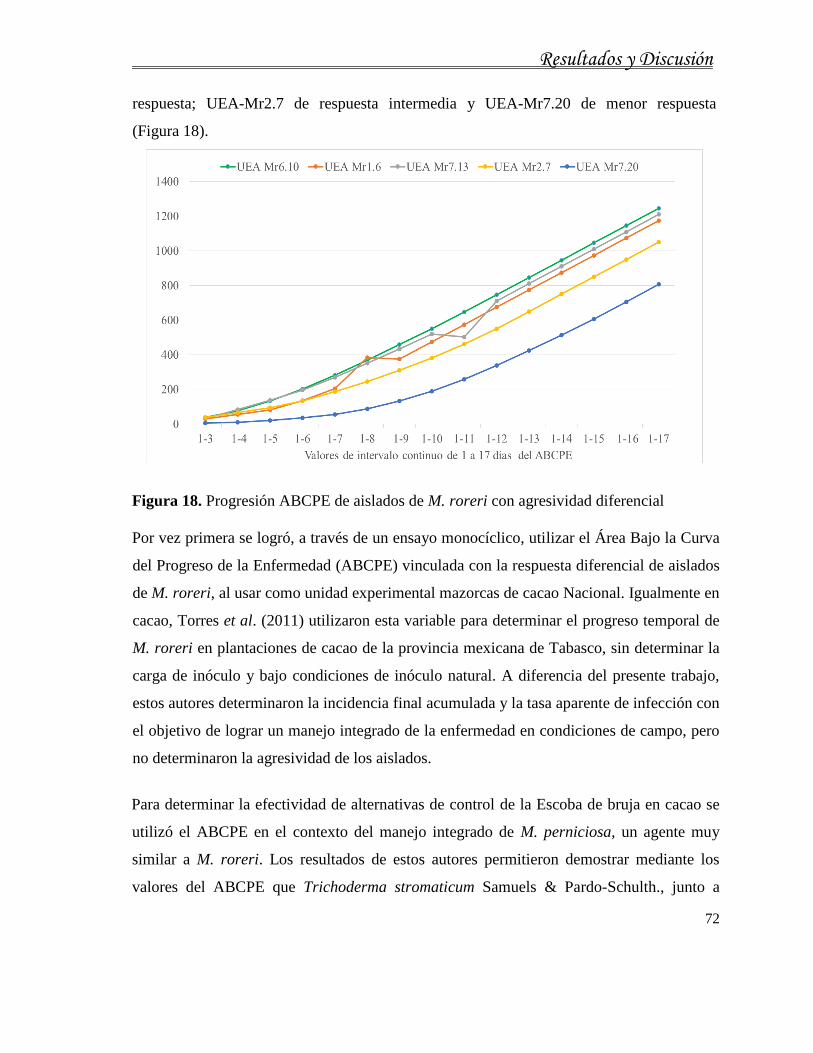
Resultados y Discusión
72
respuesta; UEA-Mr2.7 de respuesta intermedia y UEA-Mr7.20 de menor respuesta
(Figura 18).
Figura 18. Progresión ABCPE de aislados de M. roreri con agresividad diferencial
Por vez primera se logró, a través de un ensayo monocíclico, utilizar el Área Bajo la Curva
del Progreso de la Enfermedad (ABCPE) vinculada con la respuesta diferencial de aislados
de M. roreri, al usar como unidad experimental mazorcas de cacao Nacional. Igualmente en
cacao, Torres et al. (2011) utilizaron esta variable para determinar el progreso temporal de
M. roreri en plantaciones de cacao de la provincia mexicana de Tabasco, sin determinar la
carga de inóculo y bajo condiciones de inóculo natural. A diferencia del presente trabajo,
estos autores determinaron la incidencia final acumulada y la tasa aparente de infección con
el objetivo de lograr un manejo integrado de la enfermedad en condiciones de campo, pero
no determinaron la agresividad de los aislados.
Para determinar la efectividad de alternativas de control de la Escoba de bruja en cacao se
utilizó el ABCPE en el contexto del manejo integrado de M. perniciosa, un agente muy
similar a M. roreri. Los resultados de estos autores permitieron demostrar mediante los
valores del ABCPE que Trichoderma stromaticum Samuels & Pardo-Schulth., junto a

Resultados y Discusión
73
fungicidas cúpricos, permitieron lograr un manejo integrado eficiente de la Escoba de bruja,
así como incrementar los rendimientos agrícolas y el retorno económico del cultivo
(Medeiros et al., 2010; Samuels et al., 2012b).
En un estudio realizado en Santo Domingo, Ecuador, se utilizó el ABCPE con el fin de
mitigar la Moniliasis en cacao híbrido nacional x trinitario con el uso de biopreparados,
resultados que fueron evaluados mediante la escala propuesta por Sánchez y González
(1989). Estos valores fueron la base para calcular el Área Bajo la Curva del Progreso de la
Enfermedad, donde se demostró que el tratamiento Cepacide (Fungicida biológico cuyo
ingrediente activo es Pseudomona cepacia) presentó un área menor en relación con la
incidencia de la Moniliasis, lo que evidenció que este tratamiento fue el que mejor controló
el patógeno en el tiempo, con 1 211,0 unidades (Estrella y Cedeño, 2012).
Con los resultados del presente trabajo, se concluye que el ABCPE puede ser un descriptor
de la agresividad de aislados de M. roreri, si se tiene en cuenta que permitió determinar el
efecto cuantitativo negativo de los aislados del hongo fitopatógeno sobre los frutos de
cacao, lo cual refleja la cantidad relativa de la magnitud de los síntomas sobre las mazorcas
inoculadas.
A pesar de que en esta investigación solo se caracterizaron 20 aislados, se apreció que
existieron diferencias respecto a la agresividad de los mismos, lo cual es un aspecto
importante para la investigación, aplicada al mejoramiento genético del cacao en la
Amazonía ecuatoriana, una vez que se desee seleccionar aislados de M. roreri para realizar
evaluaciones del germoplasma de cacao en condiciones controladas y semicontroladas y
colaborar con programas de mejoramiento genético de este cultivo en la selección de
cultivares resistentes.
4.2. Respuesta de genotipos de cacao frente a M. roreri en un ensayo monocíclico
La Figura 19 muestra la respuesta del cultivar Nacional a la inoculación artificial por
M. roreri a través de un ensayo monocíclico. Los síntomas de severidad externa de las
mazorcas al tercer día de la inoculación manifestaron grado 1 (puntos aceitosos) y grado 2

Resultados y Discusión
74
(tumefacción o clorosis). A partir del día cuatro se observó grado 3 (mancha necrótica). En
la evaluación realizada del día nueve al día 11 se observó grado 4 (micelio hasta en un 25%
de la mancha), y al día 13 grado 5 (micelio en más del 25% de la mancha) en la mayoría de
las repeticiones.
Figura 19. Severidad externa acorde con la escala de Brenes (1983) del daño ocasionado por
aislados de M. roreri en ensayo monocíclico en el cultivar Nacional a) fruto sano b) puntos
aceitosos c) tumefacción o clorosis d) mancha necrótica e) micelio en un 25% de la mancha
f) micelio en más del 25% de la mancha
En el caso de las mazorcas de clon CCN-51 al día seis de la inoculación manifestaron grado
1 (puntos aceitosos), a partir del día nueve se observó grado 3 (mancha necrótica), el grado
4 (micelio hasta en un 25% de la mancha) se manifestó al día 18, y el grado 5 (micelio en
más del 25% de la mancha) se observó al día 19 en la mayoría de las repeticiones. No se
pudo evidenciar la presencia de grado 2 (tumefacción o clorosis), debido al color rojo que
presenta este tipo de clon (Figura 20).
Figura 20. Severidad externa acorde con la escala de Brenes (1983) del daño ocasionado por
aislados de M. roreri en ensayo monocíclico en el clon CCN-51 a) fruto sano b) puntos aceitosos
c) mancha necrótica d) micelio en un 25% de la mancha e) micelio en más del 25% de la mancha
b
c
d
e
f
a
b
c
d
e
a

Resultados y Discusión
75
Estos resultados coinciden con lo referido por Rodríguez (2004); García (2010) y
Avendaño et al. (2010), quienes manifestaron que el índice de severidad externa no permite
identificar infecciones ocultas; lo que hace muy difícil identificar síntomas anteriores a la
formación de la mancha sobre todo en genotipos de color rojo (Ampuero, 1967;
Evans, 1981).
En mazorcas del clon EET-95, se observó grado 1 (puntos aceitosos) a los tres días después
de la inoculación, grado 2 (tumefacción o clorosis) se manifestó al día siete. A partir del
día ocho presentó grado 3 (mancha necrótica), grado 4 (micelio hasta en un 25% de la
mancha) se evidenció al día 13 y al día 16 se observó grado 5 (micelio en más del 25% de
la mancha) (Figura 21).
Figura 21. Severidad externa acorde con la escala de Brenes (1983) del daño ocasionado por
aislados de M. roreri en ensayo monocíclico en el clon EET-95 a) fruto sano b) puntos aceitosos
c) tumefacción o clorosis d) mancha necrótica e) micelio en un 25% de la mancha f) micelio en
más del 25% de la mancha
Las mazorcas de clon EET-96 manifestaron grado 1 (puntos aceitosos) al día tres después
de la inoculación artificial, al día ocho se observó grado 2 (tumefacción o clorosis). A partir
del día nueve las mazorcas manifestaron grado 3 (mancha necrótica), al día 13 presentó
grado 4 (micelio hasta en un 25% de la mancha) y grado 5 (micelio en más del 25% de la
mancha) manifestó al día 16 (Figura 22). A los 17 días los síntomas expresados en las
mazorcas por la mezcla de aislados inoculados, se comportaron uniformemente en el
transcurso de los días y no se observó diferencia entre ellos.
b
c
d
e
f
a

Resultados y Discusión
76
Figura 22. Severidad externa acorde con la escala de Brenes (1983) del daño ocasionado por
aislados de M. roreri en ensayo monocíclico en el clon EET-96 a) fruto sano b) puntos aceitosos
c) tumefacción o clorosis d) mancha necrótica e) micelio en un 25% de la mancha f) micelio en
más del 25% de la mancha
Mazorcas de clon EET-103, al día tres de la inoculación artificial manifestaron grado 1
(puntos aceitosos), al día siete grado 2 (tumefacción o clorosis). A partir del día ocho se
observó grado 3 (mancha necrótica), grado 4 (micelio hasta en un 25% de la mancha) al día
14, y grado 5 (micelio en más del 25% de la mancha) al día 16 en la mayoría de las
repeticiones (Figura 23). A los 17 días los síntomas expresados en las mazorcas por la
mezcla de los aislados inoculados, se comportaron uniformemente en el transcurso de los
días y no se observaron diferencias entre ellos.
Figura 23. Severidad externa acorde con la escala de Brenes (1983) del daño ocasionado por
aislados de M. roreri en ensayo monocíclico en el clon EET-103 a) fruto sano b) puntos aceitosos
c) tumefacción o clorosis d) mancha necrótica e) micelio en un 25% de la mancha f) micelio en
más del 25% de la mancha
b
c
d
e
f
a
b
c
d
e
f
a

Resultados y Discusión
77
Al analizar la severidad interna, se observaron los siguientes síntomas en los cinco
genotipos de cacao:
Cultivar Nacional.
Figura 24. Severidad interna acorde a la escala de Sánchez et al. (1987) del daño ocasionado por la
mezcla de los cuatro aislados de M. roreri en ensayo monocíclico cultivar Nacional a) grado 0
(cero área necrosada) b) grado 1 (1-20% del área necrosada) c) grado 2 (21-40% del área necrosada)
d) grado 3 (41-60% del área necrosada) e) grado 4 (61-80% del área necrosada) f) grado 5 (100%
del área necrosada)
Clon CCN-51
Figura 25. Severidad interna acorde a la escala de Sánchez et al. (1987) del daño ocasionado por la
mezcla de los cuatro aislados de M. roreri en ensayo monocíclico en el clon CCN-51 a) grado 0
(cero área necrosada) b) grado 1 (1-20% del área necrosada) c) grado 2 (21-40% del área necrosada)
d) grado 3 (41-60% del área necrosada) e) grado 4 (61-80% del área necrosada) f) grado 5 (100%
del área necrosada)
b c
d
e f
a
a
b c
d e f

Resultados y Discusión
78
Clon EET-95
Figura 26. Severidad interna acorde a la escala de Sánchez et al. (1987) del daño ocasionado por la
mezcla de los cuatro aislados de M. roreri en ensayo monocíclico en el clon EET-95 a) grado 0
(cero área necrosada) b) grado 1 (1-20% del área necrosada) c) grado 2 (21-40% del área necrosada)
d) grado 3 (41-60% del área necrosada) e) grado 4 (61-80% del área necrosada) f) grado 5 (100%
del área necrosada)
Clon EET-96
Figura 27. Severidad interna acorde a la escala de Sánchez et al. (1987) del daño ocasionado por la
mezcla de los cuatro aislados de M. roreri en ensayo monocíclico en el clon EET-96 a) grado 0
(cero área necrosada) b) grado 1 (1-20% del área necrosada) c) grado 2 (21-40% del área necrosada)
d) grado 3 (41-60% del área necrosada) e) grado 4 (61-80% del área necrosada) f) grado 5 (100%
del área necrosada)
a
b c
d e f
a
b c
d e f

Resultados y Discusión
79
Clon EET-103
Figura 28. Severidad interna acorde a la escala de Sánchez et al. (1987) del daño ocasionado por la
mezcla de los cuatro aislados de M. roreri en ensayo monocíclico en el clon EET-103 a) grado 0
(cero área necrosada) b) grado 1 (1-20% del área necrosada) c) grado 2 (21-40% del área necrosada)
d) grado 3 (41-60% del área necrosada) e) grado 4 (61-80% del área necrosada) f) grado 5 (100%
del área necrosada)
En relación con esta variable en la literatura científica no se han encontrado referencias
previas del uso de la misma para determinar la severidad interna de aislados de M. roreri en
variedades de cacao en condiciones in vitro, lo cual constituye un aporte del presente
trabajo para enriquecer la determinación de la respuesta de genotipos de cacao a la
inoculación artificial de M. roreri.
A partir de este estudio se pudo concluir que el cultivar Nacional resultó susceptible
(4,0 SI) en relación con CCN-51 (3,0 SI), EET-95 (3,7 SI), EET-96 (3,6 SI), EET-103
(3,3 SI), los que según los valores de la escala de severidad interna resultaron
moderadamente susceptibles.
Los valores de SI de la investigación fueron similares a los informados por Phillips-Mora
et al. (2005) y Porras (1985) quienes obtuvieron valores de SI que oscilaron entre 0,8 a 4,6
es decir con un necrosamiento interno de 0 a 100% al evaluar diferentes genotipos de cacao
con inoculaciones de M. roreri en condiciones de campo.
a
b c
d e f

Resultados y Discusión
80
Estos resultados reafirman lo reportado por Saquicela (2010), quien determinó la alta
susceptibilidad que presenta el cultivar Nacional ante la Moniliasis al evaluar los
componentes de manejo integrado en campo.
García (2010) en el catálogo de cultivares de cacao del Perú manifiesta que los clones
EET-95, EET-96, EET-103 son moderadamente susceptibles frente al ataque de M. roreri,
información que coincide con los resultados obtenidos en el presente trabajo.
De la misma manera Sterling et al. (2015) al evaluar in situ 50 materiales genéticos de
T. cacao, T. bicolor y T. grandiflorum con inoculación controlada de aislados autóctonos de
M. roreri en Caquetá, Colombia, determinaron que el 44% de genotipos evaluados presentó
resistencia completa o moderada a M. roreri, y el 26% no presentó signos ni síntomas de la
enfermedad (dos materiales genéticos de T. cacao, cinco de T. bicolor y seis de
T. grandiflorum), la incidencia y severidad de la enfermedad entre materiales genéticos
estuvieron influenciados por la agresividad del aislado.
Sin embargo, Phillips-Mora et al. (2005) realizaron pruebas biológicas de resistencia en
condiciones de campo, para lo cual usaron ensayos en inoculaciones artificiales de
suspensión de esporas (1,2 x 105 esporas mL-1) de siete aislados de M. roreri en cinco
cultivares de cacao de la Finca Suiza Experimental, Colombia, y obtuvieron variaciones en
los niveles de resistencia en los diferentes genotipos.
En este mismo sentido, CATIE (2012) ha desarrollado métodos de inoculación artificial
eficientes para evaluar la respuesta de diferentes genotipos a la Moniliasis (Sánchez et al.,
1987 y Phillips-Mora y Galindo, 1988), en condiciones de campo. Estos identificaron dos
clones resistentes (CATIE-R4 y CATIE-R6), un clon moderadamente resistente
(CATIE-R1), y tres clones moderadamente susceptibles (CC-137, ICS-95 T1 y PMCT-58),
al emplear altas concentraciones de inóculo (1,2 x 105 esporas mL-1) y cámara húmeda para
favorecer la infección. Los resultados obtenidos por ese método artificial son similares a los
observados en forma natural.

Resultados y Discusión
81
El cultivar Nacional fue el que mostró mayor valor de ABCPE, en relación con CCN-51,
EET-95, EET-96, EET-103 (Tabla 9). Este genotipo a pesar de su susceptibilidad es el que
se recomienda para la región amazónica, por sus características productivas y
organolépticas, distintivas del cacao ecuatoriano.
Tabla 9. Respuesta in vitro de cinco genotipos de cacao según el valor del ABCPE
Genotipos ABCPE 1-10
Medias reales
Rangos
medios
Nacional 350 41,60 a
CCN-51 279 24,10 b
EET-95 279 24,40 b
EET-96 252 18,70 b
EET-103 252 18,70 b
Rangos medios con letras diferentes, difieren por la prueba de Kruskal Wallis/Mann Whitney
P ≤0,05 (n=10)
De manera general se encontró que todos los genotipos de cacao inoculados artificialmente
con M. roreri fueron capaces de reproducir los síntomas característicos de la Moniliasis en
frutos. Estos fueron similares a los observados en campo donde el período de incubación es
de tres a ocho semanas de acuerdo con la edad del fruto (Ampuero, 1967; Suárez, 1971;
Barros, 1977; Campuzano, 1980 y Villavicencio, 2010), mientras que a nivel de laboratorio
los síntomas se desarrollaron con mayor velocidad, en un período de hasta 19 días.
Si se considera que combatir a la Moniliasis con variedades resistentes aumenta la
producción y reduce el uso de productos químicos, siendo de esta forma amigable con el
medio ambiente y atractivo para los productores, se propone como una alternativa la
obtención de nuevos genotipos resistentes a la Moniliasis desde los puntos de vista técnico,
económico y de manejo para los pequeños productores de cacao en la Amazonía
ecuatoriana.

Resultados y Discusión
82
Por vez primera, se logró determinar la agresividad de aislados de M. roreri en un ensayo
monocíclico de cacao, lo que demostró que con ensayos de inoculación artificial de este
hongo fitopatógeno se pueden diferenciar genotipos de cacao sin tener que realizar
inoculaciones en condiciones de campo, y así evitar la liberación de inóculo que pueda
impactar negativamente sobre el agro ecosistema cacaotero de producción orgánica en la
Amazonía ecuatoriana.
4.3. Caracterización cultural y molecular de Trichoderma spp. procedentes de
plantaciones de cacao orgánico
Se obtuvieron tres aislados del género Trichoderma procedentes de la rizosfera de tres
plantaciones de cacao, que de acuerdo con las características culturales en medio de cultivo
PDA (Figura 29) y moleculares, se correspondieron a T. harzianum y T. viride, procedentes
de la provincia del Napo (Anexo 11).
Figura 29. Características culturales en medio de cultivo PDA de los aislados de
Trichoderma spp. a) UEA-Th1; b) UEA-Th2; c) UEA-Tv3.
Para los aislados UEA-Th1 y UEA-Th2, el análisis de secuencia de los nucleótidos
mediante Blastn mostró una homología de 98 y 99% respectivamente, para T. harzianum
con número de accesión KP133166 según GenBank (Pozzo et al., 2015). El análisis Blastn
indicó que la secuencia parcial del gen 18S ADNr amplificada a partir del ADN del aislado
UEA-Th3, correspondió con un 99% de similitud a T. viride (Li et al., 2002), con respecto
a la secuencia AF525230 depositada en la base de datos de GenBank de la NCBI
(Tabla 10).
a
b
c

Resultados y Discusión
83
Tabla 10. Identificación molecular de aislados de Trichoderma spp.
Código de
aislado FASTA más cercana N° Accesión
% de
identidad Referencia
UEA-Th1 Trichoderma harzianum
aislado BLT4A KP133166.1 98 Pozzo et al. (2015)
UEA-Th2 Trichoderma harzianum
aislado BLT4A KP133166.1 99 Pozzo et al. (2015)
UEA-Tv3 Trichoderma viride AF525230.1 99 Li et al. (2002)
Los tres aislados obtenidos difirieron en cuanto a sus características morfológicas y
culturales. Trichoderma presentó diversas formas de crecimiento en relación con el
micelio, el color de sus colonias y la esporulación (Figura 30). Se observó que de acuerdo
con sus características macroscópicas, UEA-Th1 y UEA-Th2 demostraron similitud con lo
descrito por Rifai (1969), Fernández y Suárez (2009) y Romero et al. (2015) quienes
describieron los caracteres de T. harzianum. UEA-Tv3 presentó similitud a lo descrito por
Leickfeldt et al. (1999) para los caracteres morfológicos y moleculares de T. viride.
Figura 30. Microfotografía de las características morfológicas de los aislados de
Trichoderma, a) UEA-Th1; b) UEA-Th2; c) UEA-Tv3 con un aumento de 400X.
Entre las características relevantes que se observaron en los tres aislados de
Trichoderma spp. se pudo determinar que el color del micelio fue blanco, y que este era
retorcido y plano, las fiálides estuvieron presentes y la esporulación fue de color verde
(Tabla 11). Este resultado es coincidente con lo descrito por Rifai (1969) y Olivera Costa y
Rodríguez (2014), quienes al referirse a las colonias de Trichoderma mencionan esta
coloración como la típica para este género.
a
b
c

Resultados y Discusión
84
El aroma a coco que emanó fue claramente perceptible en el aislado UEA-Tv3 debido a su
efecto antagónico, similar a lo observado por Dennis y Wester (1971) y Lieckfeld et al.
(1999). Este resultado se corrobora con lo descrito por Samuels et al. (2006b) quien al
estudiar aislados de Trichoderma de Perú y Ecuador determinó el fuerte olor a coco que
emanan estos biocontroladores, hecho distintivo del clado "viride" debido a la presencia del
ácido nonanoico conocido por inhibir el crecimiento y la germinación de las esporas
(Garrett y Robinson, 1969).
Tabla 11. Características culturales y morfológicas de los tres aislados de Trichoderma
spp. procedentes de la rizosfera de plantaciones de cacao
Aislados Forma del
micelio
Color del
micelio
Tipo de
micelio
Color de
esporas Olor
Presencia
de Fiálides
UEA-Th1 Retorcido Blanco Plano Verde Ninguno Si
UEA-Th2 Retorcido Blanco Plano Verde Ninguno Si
UEA-Tv3 Retorcido Blanco Aéreo Verde Aroma a coco Si
En relación con la presencia de Trichoderma asociada a cacao en la Amazonía del
Ecuador, Evans et al. (2003), describieron la presencia de Trichoderma hamatum Rifai,
T. harzianum, T. virens (J.H. Mill., Giddens & A.A. Foster) y T. viride asociados a
Theobroma gileri árbol amazónico, en Esmeraldas y Pichincha, provincias de la Costa y
Sierra ecuatoriana, respectivamente.
De la misma forma Evans (1999), descrito por Samuels et al. (2006a), aisló Trichoderma
ovalisporum Samuels & Schroers aislado (DIS 70a) de una liana (Banisteropsis caapi
Mar.) procedente de la provincia de Sucumbíos, región Amazónica del Ecuador en la
ribera de los ríos Panacocha y Yanayacu.
En la literatura científica consultada no se encontró hallazgo previo de informe, de
T. harzianum y T. viride en la Amazonía ecuatoriana, asociada a cacao, por lo que este
trabajo constituye el primer reporte de estas especies antagonistas en plantaciones
orgánicas amazónicas de cacao.

Resultados y Discusión
85
Velocidad de crecimiento micelial de aislados de Trichoderma spp.
Se pudo evidenciar que los promedios de crecimiento micelial de Trichoderma spp. fueron
similares en los tres aislados, cubriendo la placa de Petri (90 mm Ø) a las 72 h de
inoculación, por lo tanto no presentaron diferencias significativas en cada una de las
evaluaciones (Tabla 12).
Tabla 12. Crecimiento micelial (mm) de tres aislados de Trichoderma spp. a las 24, 48 y
72 h
Aislados Crecimiento micelial (mm)
24 h 48 h 72 h
UEA-Th1 21,26 71,58 90,05
UEA-Th2 19,89 68,76 90,05
UEA-Tv3 17,22 71,90 90,05
Datos por la prueba de Duncan P<0,05 (n=10)
Dennis y Webster (1971), y Cook y Baker (1983) afirmaron que la velocidad de
crecimiento de Trichoderma sirve como guía para su utilización como antagonistas en el
control de fitopatógenos, demostrando así la variabilidad de la actividad antagónica de las
especies de Trichoderma; lo cual constituye una ventaja en el momento de colonizar el área
de crecimiento del hongo fitopatógeno.
Los resultados obtenidos coinciden con lo descrito por Párraga y Zambrano (2012),
quienes al estudiar la tasa de crecimiento de cepas de Trichoderma spp. in vitro
determinaron que las cepas C1, C10, C4 y C9 cubrieron la totalidad de las placas de Petri
(90 mm Ø) a los tres días, después de incubadas.
Estos resultados difieren de lo descrito por Arcos (2011), quien al evaluar cepas nativas de
Trichoderma spp. en el biocontrol de Botrytis cinerea Pers. en rosas, determinó que el
crecimiento del diámetro de la colonia de Trichoderma spp. fue de 89,3 mm a los cuatro
días.

Resultados y Discusión
86
Entre los tres aislados de Trichoderma spp. no se encontraron diferencias estadísticas en
cuanto a la tasa de crecimiento micelial, a pesar de que el aislado UEA-Tv3, mostró un
valor relativo ligeramente superior (Tabla 13).
Estos resultados difieren de los de Solís (1999), quien al evaluar organismos antagónicos
asociados a M. roreri determinó que la velocidad de crecimiento de Trichoderma spp. a las
96 h fue de 22,50 mm día.
Tabla 13. Tasa de crecimiento micelial de aislados de Trichoderma a los tres días de
incubación y al cuarto día para los biopreparados
Aislados Tasa de crecimiento micelial (mm día)
UEA-Tv3 36,42
UEA-Th2 35,08
UEA-Th1 34,40
Datos por la prueba de Duncan P<0,05 (n=10)
4.4. Evaluación de la actividad antagónica in vitro de Trichoderma spp. frente a
M. roreri
Competencia por espacio
Al evaluar el enfrentamiento en cultivo dual de Trichoderma harzianum y T. viride, frente a
M. roreri, se determinó que al primer día los aislados UEA-Th1, UEA-Th2 y UEA-Tv3,
presentaron crecimientos promedios de 18,17; 17,75 y 16,45 mm respectivamente.
M. roreri tuvo un crecimiento promedio de 29,58 mm. En el segundo día de evaluación
Trichoderma UEA-Tv3 presentó mayor crecimiento con un promedio de 48,00 mm,
UEA-Th2 y UEA-Th1 alcanzaron promedios de 44,72 y 43,36 mm respectivamente. Los
aislados de M. roreri presentaron crecimiento promedio de 36,38 mm. Al tercer día de
evaluación, el aislado UEA-Tv3 obtuvo el mayor crecimiento con un promedio de 61,79
mm, seguido de UEA-Th1 y UEA-Th2 con promedios de 56,37 y 56,13 mm
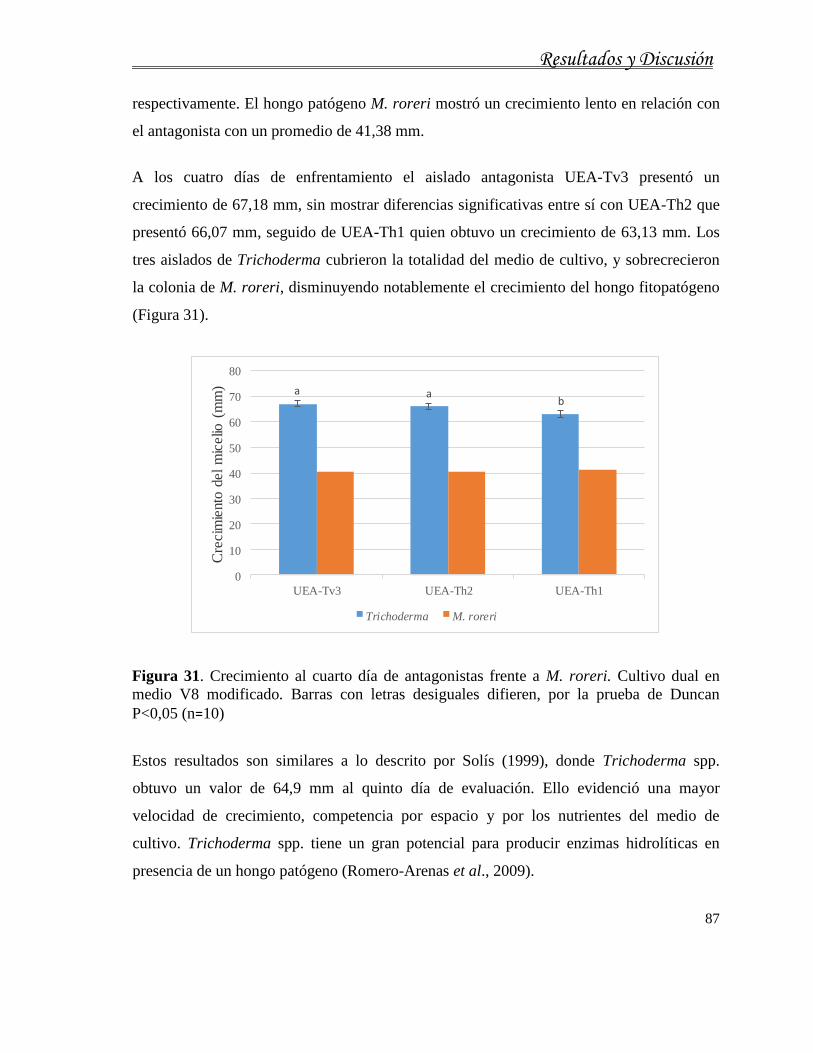
Resultados y Discusión
87
respectivamente. El hongo patógeno M. roreri mostró un crecimiento lento en relación con
el antagonista con un promedio de 41,38 mm.
A los cuatro días de enfrentamiento el aislado antagonista UEA-Tv3 presentó un
crecimiento de 67,18 mm, sin mostrar diferencias significativas entre sí con UEA-Th2 que
presentó 66,07 mm, seguido de UEA-Th1 quien obtuvo un crecimiento de 63,13 mm. Los
tres aislados de Trichoderma cubrieron la totalidad del medio de cultivo, y sobrecrecieron
la colonia de M. roreri, disminuyendo notablemente el crecimiento del hongo fitopatógeno
(Figura 31).
a ab
0
10
20
30
40
50
60
70
80
UEA-Tv3 UEA-Th2 UEA-Th1
Cre
cim
iento
del
mic
elio
(m
m)
Trichoderma M. roreri
Figura 31. Crecimiento al cuarto día de antagonistas frente a M. roreri. Cultivo dual en
medio V8 modificado. Barras con letras desiguales difieren, por la prueba de Duncan
P<0,05 (n=10)
Estos resultados son similares a lo descrito por Solís (1999), donde Trichoderma spp.
obtuvo un valor de 64,9 mm al quinto día de evaluación. Ello evidenció una mayor
velocidad de crecimiento, competencia por espacio y por los nutrientes del medio de
cultivo. Trichoderma spp. tiene un gran potencial para producir enzimas hidrolíticas en
presencia de un hongo patógeno (Romero-Arenas et al., 2009).

Resultados y Discusión
88
El comportamiento del enfrentamiento dual de los microorganismos antagonistas frente
M. roreri en medio V8 modificado, en relación con la reducción del hongo patógeno,
evidenció que al quinto día de evaluación, los aislados UEA-Tv3, UEA-Th2 y UEA-Th1
mostraron el mayor porcentaje de inhibición, sin diferencias significativas entre sí con
PICR de 47,66; 47,64 y 46,45 respectivamente.
Este planteamiento es confirmado por Samuels et al. (2006b), quienes obtuvieron 44,93%
de PICR al evaluar M. roreri en caldo extracto de malta con filtrados de T. theobromicola
Samuels & H.C. Evans procedentes de Perú.
La capacidad antagónica según la escala de Bell et al. (1982), de los aislados de
Trichoderma spp. sobre M. roreri fue de clase 2 (Tabla 14), donde a través de
observaciones macroscópicas, se evidenció que los antagonistas colonizaron las dos
terceras partes de la superficie de la placa de Petri (90 mm Ø) en la que inhibieron el
crecimiento de M. roreri.
Tabla 14. Clase y zonas de antagonismo de aislados de Trichoderma spp. en medio V8
modificado
Antagonista Clase de
antagonismo
Zonas de
antagonismo
UEA-Th1 2 3
UEA-Th2 2 3
UEA-Tv3 2 3
Estos resultados son semejantes a los obtenidos por Solís (1999), quien al evaluar a
Trichoderma spp. sobre M. roreri, determinó que se ubicó en clase 1.
Otros autores tales como Harman (2003), Bernal et al. (2004), Hernández et al. (2006),
Martinez et al. (2008), Infante et al. (2009) y Martínez et al. (2013), afirmaron que
Trichoderma spp. tiene una alta capacidad de competencia por nutrientes y espacio, dado
fundamentalmente porque su velocidad de crecimiento es superior a la del hongo patógeno.

Resultados y Discusión
89
De conformidad a la escala de Punja y Grogan (1983), la zona de antagonismo de los tres
aislados de Trichoderma spp. se situó en la zona 3, presentando aversión fuerte (Tabla 14).
Micoparasitismo
Los resultados de las observaciones microscópicas de las interacciones entre antagonista y
fitopatógeno dependieron del aislado. UEA-Tv3 mostró un efecto marcado sobre el micelio
y las esporas de M. roreri, en los que se evidenciaron mecanismos de control como
degradación parcial o total (lisis), ruptura y fragmentación. Se observó penetración y
formación de haustorios dentro de las hifas de M. roreri (Figura 32). Los resultados
encontrados en el presente trabajo coinciden con los mecanismos de penetración
informados por Howell (2003) en otras interacciones antagonistas-hongos fitopatógenos.
Figura 32. Micoparasitismo de Trichoderma viride aislado UEA-Tv3, en M. roreri, con
aumento de 400X.
Estos resultados se relacionan con lo encontrado por Gonzales (2001), quien al evaluar
T. viride y Trichoderma sp. cepa C-66 detectó micoparasitismo sobre aislados de
Rhizoctonia solani Kühn, en frijol (Phaseolus vulgaris L.) con reducción del crecimiento
micelial del hongo patógeno. Otros autores como Guédez et al. (2009) comprobaron el
fenómeno de micoparasitismo por enrollamiento, penetración y lisis en interacción con
hongos que afectan el cultivo de la fresa (Fragaria x ananassa Duch.), mientras que Pérez
et al. (2013), al evaluar mecanismos de interacción hifal de T. harzianum sobre Bipolaris

Resultados y Discusión
90
oryzae Breda de Haan, observaron enrollamiento, penetración hifal, lisis, y vacuolización
en hifas.
Además, los tres aislados del género de Trichoderma mostraron la propiedad de agrupar
esporas sobre el micelio M. roreri (Figura 33). UEA-Th1 y UEA-Th2 fueron capaces de
desarrollar estructuras de reproducción.
Figura 33. Agrupamiento de esporas de Trichoderma harzianum aislado UEA-Th2, sobre
micelio de M. roreri, con aumento de 400X
No se observó enrollamiento, lo que coincide con lo descrito por Acosta-Suárez et al.
(2013), quienes determinaron que T. harzianum no causó enrollamiento sobre
Mycosphaerella fijiensis Morelet en condiciones in vitro.
Antibiosis
En la prueba de metabolitos volátiles, el aislado UEA-Tv3 fue el que mostró la mayor
actividad en la reducción del crecimiento relativo de M. roreri a las 168 h con un promedio
de inhibición de 21,12%, pero sin diferencias significativas con UEA-Th2, UEA-Th1 que
presentaron una actividad de 15,89 y 13,65% respectivamente, así como reducción del
crecimiento sobre el fitopatógeno (Tabla 15).

Resultados y Discusión
91
Tabla 15. Crecimiento de M. roreri en presencia de metabolitos volátiles de antagonistas,
a los siete días de evaluación
Crecimiento radial (mm h-1) de M. roreri PICR
(%) Antagonista 24 h 48 h 72 h 96 h 120 h 144 h 168 h
UEA-Th1 27,39 ab 36,62 45,68 ab 57,16 a 61,09 66,97 71,39 13.65
UEA-Th2 28,41 a 38,72 48,55 a 57,17 a 59,24 66,69 69,54 15.89
UEA-Tv3 25,46 b 35,03 43,67 b 51,61 b 56,39 63,42 65,22 21.12
EE (±) 0,70 1,15 1,21 1,59 2,71 2,27 2,45
Medias con letras diferentes, en el sentido de las columnas difieren por la prueba de Duncan
P ≤0,05 (n=10)
Los aislados UEA-Tv3 y UEA-Th2 tuvieron influencia en la reducción de la esporulación
de M. roreri. Las afectaciones de este antagonista se observaron en las características
culturales del fitopatógeno, y provocaron variaciones en los bordes y textura del micelio,
mientras el aislado UEA-Th1 inhibió la esporulación con menor eficacia (Tabla 16).
Tabla 16. Porcentaje de Inhibición de Esporulación (% I.E) de M. roreri en presencia de
antagonistas
Samuels et al. (2006b), al evaluar la actividad micoparasítica de Trichoderma
theobromicola obtenida a partir de árboles sanos de cacao de la Amazonía peruana y
T. paucisporum Samuels, C. Suarez & Solís aislada a partir de mazorcas de cacao
parcialmente infectadas con M. roreri en la provincia de Los Ríos, Ecuador, demostraron
que los dos antagonistas fueron capaces de producir antibióticos difusibles volátiles que
inhibieron el desarrollo de M. roreri in vitro y en campo.
Los resultados de esta investigación se corresponden además con lo informado por otros
autores respecto a la capacidad antagónica de Trichoderma. León et al. (2012)
Antagonista Esporulación
(conidias mL-¹) % IE
UEA-Th1 8,87 x 104 47,82
UEA-Th2 4,75 x 104 72,06
UEA-Tv3 3,70 x 104 78,24
Control 1,70 x 105

Resultados y Discusión
92
determinaron micoparasitismo sobre Sclerotium rolfsii Sacc., Rhizoctonia sp. y
Fusarium sp. que provoca estrangulamiento o lisis de las paredes de las hifas, y con ello la
desintegración y degradación de las paredes celulares y el debilitamiento o muerte de los
patógenos. Inhibe por competencia e hiperparasitismo el crecimiento de Macrophomina
phaseolina Tassi (Goid) (Hernández et al., 2011). Alvindia (2012) determinó inhibición del
crecimiento de M. fijensis en la cepa de T. harzianum DGA01, aislada de frutos de banano
después de 10 días de incubación.
Estos resultados corroboraron el alto potencial antagonista de Trichoderma sobre M. roreri
lo que permite seleccionar aislados que se destacan por su amplia variabilidad de
mecanismos frente a este fitopatógeno.
Es de gran importancia contar con aislados nativos que controlen M. roreri por ser
considerada la principal enfermedad del cacao en la Amazonía ecuatoriana, ocasionando
grandes pérdidas a los productores.
Los resultados obtenidos al utilizarse los aislados de T. harzianum y T. viride como agentes
de control biológico de M. roreri, constituyen los primeros informados para la Amazonía
del Ecuador.
4.5. Evaluación de la eficacia de aislados nativos de Trichoderma spp. y biopreparados
comerciales en el control de la Moniliasis del cacao en condiciones de campo
Los aislados de Trichoderma spp. (UEA-Th2 y UEA-Tv3), seleccionados por su
comportamiento in vitro frente a M. roreri, presentaron resultados muy alentadores para los
ensayos en campo. El efecto que presentaron los biocontroladores (UEA-Tv3 y UEA-Th2 +
UEA-Tv3), en la reducción del porcentaje de mazorcas infectadas con hongos
fitopatógenos y otras enfermedades mostraron diferencias significativas con los
biopreparados comerciales Trichotic y Rhapsody, siendo estadísticamente diferentes del
control. Se determinó que los aislados UEA-Tv3 y UEA-Th2 tuvieron un efecto

Resultados y Discusión
93
significativo sobre las enfermedades de cacao, sin embargo UEA-Tv3 tuvo niveles muy
cercanos a Óxido cuproso (Tabla 17).
Tabla 17. Porcentaje de mazorcas enfermas (M. roreri, M. perniciosa, y otras
enfermedades) registradas cada quince días de evaluación en parcelas aplicadas con
fungicidas químicos y biológicos
Medias con letras diferentes, en el sentido de las columnas difieren por la prueba de Duncan
P ≤0,05
Los biocontroladores ejercieron el mejor control del porcentaje de mazorcas enfermas con
respecto al control, pero fueron inferiores al tratamiento químico, sin embargo el
tratamiento con UEA-Tv3 no difirió estadísticamente del Óxido cuproso en las
evaluaciones 5, 6 y 7, lo cual no ocurrió para los demás biocontroladores.
El tratamiento con UEA-Tv3 superó al resto de los tratamientos biológicos al final de las
evaluaciones, mientras que el tratamiento UEA-Th2 + UEA-Tv3 fue superior al tratamiento
con Trichotic y Rhapsody en la última evaluación.
Esto concuerda con Solís y Suárez-Capello (2006), quienes determinaron que la reducción
de enfermedades en parcelas de cacao, tratadas con T. koningiopsis (Samuel, Súarez y
H.C. Evans) y T. stromaticum fueron similares a las obtenidas con fungicidas.
Evaluaciones
(días)
Tratamientos
Mazorcas Infectadas (%)
15 30 45 60 75 90 105
1. UEA-Tv3 47,20 b 42,9 b 35,25 b 31,5 c 29,44 cd 19,89 c 18,14 de
2. UEA-Th2 + UEA-Tv3 52,05 b 41,35 b 38,66 b 38,21 bc 35,16 bc 29,43 b 23,41 cd
3. Trichotic 52,40 b 43,90 b 40,63 b 38,9 bc 37,90 b 34,05 b 30,22 b
4. Rhapsody 50,96 b 42,58 b 42,03 b 43,10 ab 36,05 bc 30,49 b 28,16 bc
5. Óxido cuproso 50,59 b 38,74 b 34,84 b 29,86 c 25,60 d 18,9 c 15,56 e
6. Control 67,55 a 57,69 a 52,83 a 53,19 a 55,14 a 57,41 a 59,08 a
EE (±) 3,21 2,72 2,53 3,63 2,73 2,70 2,05

Resultados y Discusión
94
Resultados similares fueron descritos por Holmes et al. (2004); Benito et al. (2007); Bailey
et al. (2008); Osorio (2010) y Villamil et al. (2015), quienes al evaluar T. viride,
T. harzianum y T. hamatum, encontraron antagonismo sobre M. roreri in vitro e in situ.
Todos los tratamientos biológicos y el químico fueron mejores que el control en relación
con mazorcas infectadas con M. roreri (%) en las evaluaciones quincenales realizadas en el
ensayo de campo. Los tratamientos UEA-Tv3 y UEA-Th2 + UEA-Tv3 no difirieron del
tratamiento químico a los 45, 90 y 105 días. El tratamiento con el aislado del antagonista
UEA-Tv3 fue mejor que los productos comerciales Trichotic y Rhapsody en las
evaluaciones realizadas a los 75, 90 y 105 días (Tabla 18).
Tabla 18. Porcentaje de mazorcas enfermas con M. roreri a los cuatro meses de
evaluación en condiciones de campo
Evaluaciones
(días)
Tratamientos
Mazorcas infectadas con M. roreri (%)
15 30 45 60 75 90 105
1. UEA Tv3 40,30 b 38,38 b 31,52 bc 30,52 b 25,05 bc 13,04 d 10,75 c
2. UEA Th2 + UEA Tv3 44,19 ab 40,19 b 34,27 bc 33,27 b 29,75 b 16,80 cd 15,85 bc
3. Trichotic 45,94 ab 45,94 ab 39,94 b 35,94 b 31,54 b 25,85 b 23,13 b
4. Rhapsody 45,34 ab 43,34 ab 39,25 b 37,08 b 29,14 b 24,46 bc 21,78 b
5. Óxido cuproso 40,15 b 34,65 b 30,02 c 29,02 b 15,30 c 12,99 d 9,39 c
6. Control 56,63 a 53,47 a 56,51 a 56,51 a 54,18 a 51,51 a 53,18 a
EE (±) 4,16 4,17 2,87 2,84 3,80 3,03 2,49
Medias con letras diferentes, en el sentido de las columnas, difieren por la prueba de Duncan
P ≤0,05
Estos resultados coinciden con lo descrito por Osorio (2010), quien determinó la eficiencia
de T. harzianum como biocontrolador sobre M. roreri en la Costa ecuatoriana a partir de los
62 días de la primera aspersión. De la misma forma Guerrero y Arias (2006), determinaron
que T. koningiopsis resultó efectivo en el control de Moniliasis en la misma magnitud que
los fungicidas químicos. No así Robles (2008), quien al usar productos biológicos basados
en bacterias, determinó su producción por encima de los productos químicos.
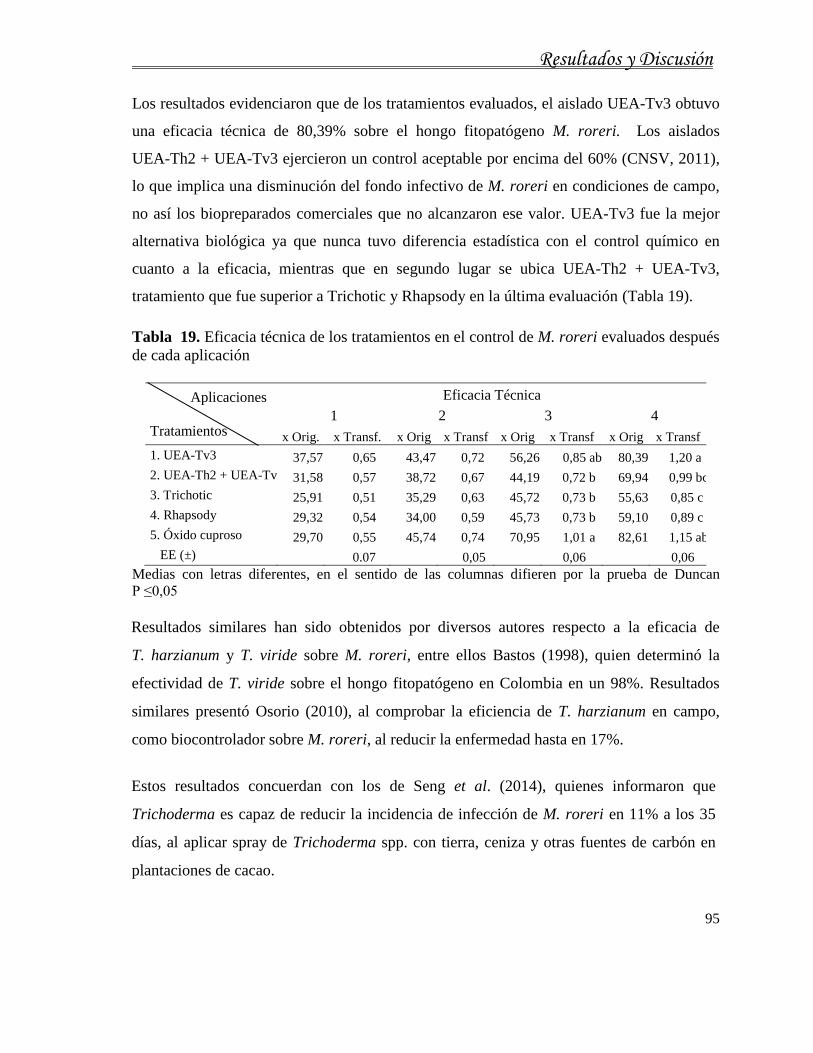
Resultados y Discusión
95
Los resultados evidenciaron que de los tratamientos evaluados, el aislado UEA-Tv3 obtuvo
una eficacia técnica de 80,39% sobre el hongo fitopatógeno M. roreri. Los aislados
UEA-Th2 + UEA-Tv3 ejercieron un control aceptable por encima del 60% (CNSV, 2011),
lo que implica una disminución del fondo infectivo de M. roreri en condiciones de campo,
no así los biopreparados comerciales que no alcanzaron ese valor. UEA-Tv3 fue la mejor
alternativa biológica ya que nunca tuvo diferencia estadística con el control químico en
cuanto a la eficacia, mientras que en segundo lugar se ubica UEA-Th2 + UEA-Tv3,
tratamiento que fue superior a Trichotic y Rhapsody en la última evaluación (Tabla 19).
Tabla 19. Eficacia técnica de los tratamientos en el control de M. roreri evaluados después
de cada aplicación
Aplicaciones
Tratamientos
Eficacia Técnica
1 2 3 4
x Orig. x Transf. x Orig x Transf x Orig x Transf x Orig x Transf
1. UEA-Tv3 37,57 0,65 43,47 0,72 56,26 0,85 ab 80,39 1,20 a
2. UEA-Th2 + UEA-Tv3 31,58 0,57 38,72 0,67 44,19 0,72 b 69,94 0,99 bc
3. Trichotic 25,91 0,51 35,29 0,63 45,72 0,73 b 55,63 0,85 c
4. Rhapsody 29,32 0,54 34,00 0,59 45,73 0,73 b 59,10 0,89 c
5. Óxido cuproso 29,70 0,55 45,74 0,74 70,95 1,01 a 82,61 1,15 ab
EE (±) 0.07 0,05 0,06 0,06
Medias con letras diferentes, en el sentido de las columnas difieren por la prueba de Duncan
P ≤0,05
Resultados similares han sido obtenidos por diversos autores respecto a la eficacia de
T. harzianum y T. viride sobre M. roreri, entre ellos Bastos (1998), quien determinó la
efectividad de T. viride sobre el hongo fitopatógeno en Colombia en un 98%. Resultados
similares presentó Osorio (2010), al comprobar la eficiencia de T. harzianum en campo,
como biocontrolador sobre M. roreri, al reducir la enfermedad hasta en 17%.
Estos resultados concuerdan con los de Seng et al. (2014), quienes informaron que
Trichoderma es capaz de reducir la incidencia de infección de M. roreri en 11% a los 35
días, al aplicar spray de Trichoderma spp. con tierra, ceniza y otras fuentes de carbón en
plantaciones de cacao.

Resultados y Discusión
96
De esta manera se confirmó que la selección y especificidad de los aislados de
Trichoderma constituyen un elemento importante a tomar en cuenta para ser utilizados
como agentes de control biológico (Acevedo y Arcia, 2006).
Ayala (2008), al determinar la efectividad técnica de fungicidas químicos en plantaciones
de cacao obtuvo valores que no sobrepasaron el 55,70% luego de seis meses de
investigación.
Algunos autores señalan la potencialidad de T. harzianum en el control de hongos
fitopatógenos como R. solani, Fusarium oxysporum f. sp. cubense (E. F. Smith) Snyd &
Hans., lo que resalta su alta capacidad antagónica; con efectividad técnica que superó el
80% (Bernal et al., 2004; García et al., 2006; Reyes et al., 2008; Pérez et al., 2009).
La producción promedio anual de los pequeños productores de cacao en sistemas
agroforestales, alcanza los 200 kg ha-1 al año. El hecho de que la aplicación de aislados
nativos de T. harzianum y T. viride reduzca la incidencia de la enfermedad hasta en un
40% con una eficacia técnica de 80,39%, demuestra la importancia de reproducir
artesanalmente los hongos antagonistas a nivel local, que además de ser económicos, se
encuentran disponibles por los productores en el momento requerido para realizar
aplicaciones en campo, e incrementan la producción de cacao orgánico.
Si se considera que el costo de producción de T. harzianum y T. viride (dosis de 1013
esporas ha-1) en condiciones de laboratorio es de USD 15,02 más el costo de aplicación de
USD 19,2 esto nos da un costo total de aplicación de USD 34,22 ha-1, lo que representa un
rubro adicional en el presupuesto del sistema productivo, pero significativamente
económico en relación con la producción que pasaría de 200 a 320 kg ha-1 de cacao seco
por año, lo que representa un incremento de la producción de USD 240 al año por hectárea.
De esta manera se comprobó la eficiencia de T. harzianum y T. viride como biocontrolador
de M. roreri al aplicarlo en campo, con eficiencias de 80,39% y 60,94%, respectivamente.

Resultados y Discusión
97
En este caso los resultados obtenidos permiten contar con tres aislados nativos de
Trichoderma, provenientes de plantaciones orgánicas de cacao en la Amazonía
ecuatoriana, debidamente caracterizados y evaluados in vitro y en campo, para el manejo
biológico de M. roreri en cacao, y de esta forma disminuir las pérdidas ocasionadas por
este hongo fitopatógeno en campo, lo cual resulta una alternativa de producción para los
pequeños productores, quienes no contaban con estrategias biológicas para mitigar la
Moniliasis. Así, este constituye el primer registro en el país del control de esta enfermedad
con el empleo de los antagonistas T. harzianum y T. viride.

Conclusiones

Conclusiones
98
5. CONCLUSIONES
1. Las características culturales, morfológicas, fisiológicas, moleculares y patogénicas
de los 20 aislados obtenidos con síntomas de Moniliasis correspondieron a la
especie Moniliophthora roreri.
2. Se identificaron cuatro géneros de hongos anamorfos, asociados a mazorcas de
cacao con síntomas de Moniliasis, correspondientes a Cylindrocladium,
Dichobotrys, Colletotrichum y Phytophthora.
3. El cultivar Nacional resultó más susceptible a los aislados de M. roreri, en mazorcas
de cacao, en relación a la respuesta diferencial de los genotipos CCN-51, EET-95,
EET-96, EET-103.
4. Se obtuvieron tres aislados de hongos antagonistas: UEA-Th1 y UEA-Th2 que
fueron caracterizados e identificados a nivel de especie como Trichoderma
harzianum, y UEA-Tv3 como Trichoderma viride.
5. UEA-Tv3 Trichoderma viride y UEA-Th1 Trichoderma harzianum manifestaron un
efecto marcado de los mecanismos de acción de competencia, micoparasitismo y
antibiosis sobre M. roreri.
6. Trichoderma viride, y la combinación de Trichoderma viride más Trichoderma
harzianum mostraron una eficacia técnica superior al 69,94% sin diferencias con el
control químico, en condiciones de campo.

Recomendaciones

Recomendaciones
99
6. RECOMENDACIONES
1. Demostrar la patogenicidad de los géneros de hongos anamorfos asociados a la
Moniliasis de cacao, obtenidos en el presente trabajo.
2. Emplear los aislados UEA-Mr6.10, UEA-Mr1.6, UEA-Mr1.16, UEA-Mr7.13,
UEA-Mr7.17 y UEA-Mr2.7 en programas de selección de mejoramiento genético
en la Moniliasis de cacao en condiciones amazónicas.
3. Utilizar los genotipos CCN-51, EET-95, EET-96, EET-103, los que presentaron
resistencia a Moniliasis en ensayos de campo.
4. Evaluar la posible utilización de las cepas UEA-Tv3 y UEA-Th2 para la
formulación de biopreparados en el control de M. roreri.

Referencias Bibliográficas

Referencias Bibliográficas
7. REFERENCIAS BIBLIOGRÁFICAS
1. Abbott, W. (1925). A method of computing the effectiveness of an insecticide.
Econ. Entomol. (18), 265-267.
2. ACEPROCACAO. (2012). Proyecto de Fortalecimiento de la Asociación
Ecuatoriana de Cacao Nacional Fino de Aroma. Ecuador. pp. 8-9.
3. Acevedo, R. y Arcia, A. (2006). Control Biológico de Sclerotium cepivorum por
Trichoderma sp. in vitro. Fitopal. Venez. 1(1): 34.
4. Acosta-Suárez, M., Pichardo, T., Roque, B., Cruz-Martín, M., Mena, E., Leiva-
Mora, M., Castro, R. y Alvarado-Capó, Y. (2013). Antagonismo in vitro de
Trichoderma harzianum, Rifai contra Mycosphaerella fijiensis Morelet.
Biotecnología Vegetal 13(4): 231-235.
5. Agrios, G. (2005). Plant Pathology. Fifth edition. Academic Press. San Diego-
California, United States of America. 948 pp.
6. Aime, M. y Phillips-Mora, W. (2005). The causal agents of witches’ broom and
frosty pod rot of cacao (chocolate, Theobroma cacao) from a new lineage of
Marasmiaceae. Mycology 97(5): 1012-1022.
7. Akrofi, A. (2015). Phytophthora megakarya a review on its status as a pathogen on
cacao in west Africa. African Crop Science Journal 23(1): 67-87.
8. Alarcón, J., Arévalo, E., Díaz, A. y Galindo, J. (2012). Manejo fitosanitario del
cultivo de cacao. ICA (Instituto Colombiano Agropecuario). Produmedios. Bogotá,
Colombia. 15 pp.
9. Altschul, S., Gish, W., Miller, E., Meyers, E. y Lipman, D. (1990). Basic local
alignment search tool. J. Mol. Biol. 215, 403-10.
10. Alvindia, D. (2012). Inhibitory influence of biocontrol agents, plant oils and an
inorganic salt on Mycosphaerella fijensis and Cordana musae, the causal pathogen
of black sigatoka and leaf spot of banana African. Journal of Microbiology
Research 6(19): 4179-4184.
11. Amores, F., Agama, J., Mite, F., Jiménez, J., Loor, G. y Quiroz, J. (2009). EET-544
y EET-558 nuevos clones de cacao nacional para la producción bajo riego en la

Referencias Bibliográficas
península de Santa Elena. Boletín divulgativo No. 134. Estación Experimental
Tropical Pichilingue. Quevedo, Ecuador. 47 pp.
12. Ampuero, E. (1967). Monilia pod rot of cocoa. Cocoa Grower’s Bulletin. 9,1518.
13. ANECACAO. (2015a). Evaluación de las exportaciones de cacao. Disponible en
http://www.anecacao.com/es/historia-del-cacao/. (Consultado: 29-04-2013).
14. ANECACAO. (2015b). Exportación ecuatoriana de cacao 2015. Boletín
Divulgativo. Elaborado por Ricky Moncayo. Guayaquil, Ecuador. 6 pp.
15. ANECACAO. (2015c). Estadísticas, boletín estadístico mensual. Disponible en
http://www.anecacao.com/es/estadisticas/. (Consultado: 04-08-2015).
16. Arbeláez, L. (2010). Análisis de la diversidad intraespecie de Moniliophthora
roreri (Cif) Evans et al., por medio de marcadores morfológicos y genéticos. Tesis
de Magister en Biotecnología. Universidad Nacional de Colombia. 80 pp.
17. Arcos, M. (2011). Obtención y evaluación de cepas nativas de Trichoderma spp. en
el biocontrol de Botrytis cinerea en el cultivo de rosas. Tesis de Ingeniero
Agropecuario. Escuela Superior Politécnica del Ejército, Ecuador. 154 pp.
18. Argüello, O. (2000). Manejo Integrado de la Moniliasis del cacao en Santander.
Tecnología para el mejoramiento del sistema de producción de cacao. Impresores
Colombianos. Bucaramanga, Colombia. 84 pp.
19. Avendaño, C., Mendoza, A., Hernández, E., López G., Martínez, M., Caballero, J.,
Guillén, S. y Espinosa, S. (2010). Mejoramiento genético participativo en cacao
(Theobroma cacao L.). 79 pp.
20. Ayala, M. (2008). Manejo integrado de Moniliasis (Moniliophthora roreri) en el
cultivo de cacao (Theobroma cacao L.) mediante el uso de fungicidas, combinado
con labores culturales. Tesis de Ingeniero Agropecuario. Escuela Superior
Politécnica del Litoral, Ecuador. 115 pp.
21. Bailey, B., Bae, H., Strem, M., Crozier, J., Thomas, S., Samuels, G., Vinyard, B. y
Holmes, K. (2008). Antibiosis, mycoparasitism, and colonization success for
endophytic Trichoderma isolates with biological control potential in Theobroma
cacao. Biological Control. 46(1): 24-35.

Referencias Bibliográficas
22. Barrera-Necha, L., Bautista-Baños, S., Flores-Moctezuma, H. y Rojas, A. (2008).
Efficacy of essential oils on the conidial germination, growth of Colletotrichum
gloesporiodes (Penz.) Penz. and Sacc and control of postharvest disease in papaya
(Carica papaya L.). Plant Pathology 7(2): 174-178.
23. Barros, O. (1977). Investigaciones sobre el hongo Monilla roreri Cif & Par.
causante de la pudrición acuosa de la mazorca del cacao, sus daños y control. El
Cacaotero Colombiano 3: 42-52.
24. Bartley, B. (2005). The genetic diversity of cacao and its utilization. Cabi
Publishing. Massachusetts, United States of America. 357 pp.
25. Bastos, G. (1998). Resultados preliminares sobre la eficacia de Trichoderma viride
en el control de Monilia (Moniliophthora roreri). Bogota, Colombia. 107 pp.
26. Bastos, C., y Bezerra, J. (2010). Podridao de frutos do cautiverio causada por
Cylindrocladium scoparium MORG. Agrotrópica 22(2): 107-110.
27. Batista, L. (2009). Guía Técnica el Cultivo de Cacao en la República Dominicana.
CEDAF. Santo Domingo, República Dominicana. 250 pp.
28. Bell, D., Well, D., y Markham, C. (1982). In vitro antagonism of Trichoderm
species against six fungal plant pathogens. Phytopathology 72: 379-382.
29. Benito, J., Arévalo, E., y Garcés, L. (2007). Manejo Integrado de cacao para el
control de enfermedades con énfasis en Monilia. En Memorias Foro Binacional
ICT-INIA. Situación Actual y perspectivas de investigación en cacao (Theobroma
cacao L.) en el Perú. APP Cacao Proyecto INIA/ICT. Perú, Ecuador.
30. Bernal, A., Andreu, C., y Moya, M. Utilización de Trichoderma spp. como
alternativa ecológica para el control de Fusarium oxysporum sp. cubense (EF
Smith) Snyd & Hans. CubA, 2004. Disponible en
http://www.virtualcentr.org/es/enllBTJ%20Tallr/bernalalezander.htm. (Consultado:
22-04-2013).
31. Bettiol, W., Rivera, M., Mondino, P., Montealegre, J., y Colmenárez, Y. (2014).
Control biológico de plantas de América Latina y el Caribe. pp. 221-225.
32. BIOSCIENCE. (2004). Databases. Classification based on 9th edition of Dictionary
of the Fungi. Disponible en

Referencias Bibliográficas
http://www.indexfungorum.org./Names/NamesRecord.asp?RecordID=219848.
(Consultado: 08-11-2012).
33. Brenes, O. (1983). Evaluación de la resistencia a Monilia roreri y su relación con
algunas características morfológicas del fruto de cultivares de cacao (Theobroma
cacao L.). Tesis de Maestro en Ciencias. CATIE Centro Agronómico Tropical de
Investigación y Enseñanza, Costa Rica. 60 pp.
34. CNSV (Centro Nacional de Sanidad Vegetal). (2011). Manual de funciones para las
Estaciones territoriales de Protección de Plantas. MINAG. 45 pp.
35. Campbell, C. y Madden, L. (1990). Monitoring epidemics: disease, in Introduction
to Plant Disease Epidemiology. John Wiley. New York. pp. 107-128.
36. Campuzano, H. (1976). Fluctuación de poblaciones de esporas de Monilia roreri
Cif & Par., y viabilidad durante un ciclo completo de afección. Noticias
Fitopatológicas 5, 107.
37. Campuzano, H. (1980). La moniliasis de cacao. Cacaotero Colombiano 13: 21-24.
38. CATIE. 2012. Catálogo de clones de cacao seleccionados por el CATIE para
siembras comerciales. Manual Técnico N° 105. Turrialba-Costa Rica. 70 pp.
39. Ciferri, R. y Parodi, E. (1933). Descrizione del fungo che causa la Moniliasis del
cacao. Phytopathologische Zeitschrift 5: 539-542.
40. Concepción-Brindis, A., Bautista-Muñoz, C., Boldo-León, X., Ortiz-García, C.,
Hernández-Vélez, R. y Martínez-Carrera, D. (2015). Determination of extracellular
enzyme activity and identification of beta-glucosidase genes from Moniliophthora
roreri. Revista Mexicana de Fitopatología 32: 61.
41. Cook, R. y Baker, K. (1983). The nature and practice of biological control of plants
pathogens. American Phytopathological Society Press. United States of America.
539 pp.
42. Correa, J.; Castro, S., y Coy, J. (2014). Estado de la Moniliasis del cacao causada
por Moniliophthora roreri en Colombia. Acta Agronómica DOI:
10.15446/acag.v63n4.42747.

Referencias Bibliográficas
43. Cruz, S. (1993). Determinación de fuentes de resistencia de cacao de origen
nacional al ataque de Monilia roreri (Cif. & Par). Tesis de Ingeniero Agropecuario.
Universidad de Guayaquil, Ecuador. 101 pp.
44. Cuervo-Parra, J., Sánchez-López, V., Ramírez-Suero, M. y Ramírez-Lepe, M.
(2011). Morphological and molecular characterization Moniliophthora roreri
causal agent of frosty pod rot of cocoa tree in Tabasco Mexico. Plant Pathology
Journal 10(3): 122-127.
45. Cuhna, R. (2006). Estrutura genética de populações de Crinipellis perniciosa e
Moniliophthora roreri utilizando marcadores RAPD e SSR. Tesis de Doctor en
Agronomía (Genética y Mejoramiento de plantas). Universidad Estadual Paulista,
Brasil. 128 pp.
46. Chet, I., Harman, G., Baker, R. (1981). Trichoderma hamatum: Its hypal interaction
with Rhizoctonia solani and Pytium spp. Microbial Ecology 7: 29-38.
47. Dennis, C. y Wester, J. (1971). Antagonistic properties of species groups of
Trichoderma I. Production of non-volatile antibiotics. Transactions of the British
Mycological Society 57, 25-39.
48. Desrosiers, R.; Von Buchwald, A. y Bolaños, C. 1955. Effect of rainfall on the
incidence of monilia pod rot in Ecuador. FAO. Plant Protection Bulletin 3(2): 161-
164.
49. DNA Baser Sequence Assembler v4. (2013). Heracle BioSoft. Disponible en
www.DnaBaser.com. (Consultado: 04-03-2013).
50. ECORAE. (2001). Compendio de recomendaciones tecnológicas para los
principales cultivos de la Amazonía ecuatoriana. Nina Comunicaciones. Quito,
Ecuador. 13 pp.
51. Edgar, R. (2004). MUSCLE: multiple sequence alignment with high accuracy and
high throughput. Nucleic Acids Research 32: 1792-1797.
52. El productor. (2012). El periódico del campo. Artículos técnicos. Agrícolas.
Rehabilitación de Huertas de Cacao. Disponible en
http://elproductor.com/2012/11/22/rehabilitacion-de-huertas-de-cacao/
(Consultado: 12-08-2015).

Referencias Bibliográficas
53. Enríquez, G. (2010). Cacao Orgánico. Guía para productores ecuatorianos. Quito,
Ecuador. 360 pp.
54. Errington, J. (2003). Regulation of endospores formation in Bacillus subtilis.
Microbiol. 1: 117-126.
55. Esquivel, F., Flores, M., Domínguez, M. y Márquez, F. (2012). Ficha técnica
Escoba de bruja del cacao Moniliophthora perniciosa. Sistema Nacional de
Vigilancia Epidemiológica Fitosanitaria, SINAVEF. México. 21 pp.
56. Estrella, E. y Cedeño, J. (2012). Medidas de control de bajo impacto ambiental para
mitigar la moniliasis (Moniliophthora roreri (Cif. y Par) Evans et al.) en cacao
Híbrido Nacional x Trinitario en Santo Domingo de los Tsáchilas. Informe Técnico
Proyecto de Investigación, Santo Domingo, Ecuador. 139 pp.
57. Evans, H., Edwards, D. y Rodríguez, M. (1977). Research on cocoa diseases in
Ecuador: past and present. PANS (Pest Artic. News Summ.) 23: 68-80.
58. Evans, H., Stalpers, J., Samson, A. y Benny, G. (1978). On the taxonomy of
Monilla roreri, an important pathogen of Theobroma cacao in South America.
Canadian Journal of Botany 56: 2528- 2532.
59. Evans, C. (1981). Pod Rot of cacao caused by Moniliophthora (Monilla) roreri.
Phytopathological 44 pp.
60. Evans, H. (1986). A reassessment of Moniliophthora roreri (Monilia) pod rot of
cocoa. Cocoa Grower´s Bulletin 37: 34-43.
61. Evans, C., Holmes, K., Phillips, W. y J. Wilkinson. (2002). What is a name:
Crinipellis, the final resting place for the frosty pod rot pathogen of cacao?
Mycologist 16(4): 148- 152.
62. Evans, H., Holmes, K. y Reid, A. (2003a). Phylogeny of the frosty pod rot pathogen
of cocoa. Plant Pathology 52: 476-485.
63. Evans, H., Holmes, K. y Thomas, S. (2003b). Endophytes and mycoparasites
associated with and indigenous forest tree, Theobroma gileri, in Ecuador and a
preliminary assessment of their potential as biocontrol agents of cocoa diseases.
Mycological Progress 2: 149-160.

Referencias Bibliográficas
64. FAO. (2010). Proyecto calidad de los alimentos vinculados con el origen y las
tradiciones en América Latina. La cadena de valor del cacao en el Ecuador,
diagnóstico actual. En: Taller Nacional La Denominación de Origen para el Cacao
Arriba del Ecuador. Guayaquil, Ecuador.
65. FEDECACAO (Federación Nacional de Cacaoteros). (2004). Reconocimiento y
control de la Monilia del cacao. Colombia. 4 pp.
66. Feitosa, M., Pimente, C., Olivieira, V. y Faria, B. (1986). Cylindrocladium
floridanum, fungo patogenio en cacaoteiros (Theobroma cacao) no estado de São
Paulo. Fitopatología Brasileira 11(3): 455-463.
67. Fernández, R. y Suárez, C. (2009). Antagonismo in vitro de Trichoderma
harzianum Rifai against Fusarium oxysporum Schlecht f. sp. passiflorae en
maracuya (Passiflora edulis Sims var. Flavicarpa) del municipio Zona Bananera
Colombiana. Rev. Fac. Nal. Agr. Medellín 62(1): 4743-4748.
68. Flood, J. y R. Murphy. (2004). Cocoa futures: A source book of some important
issues facing the cocoa industry. The Commodities Press. 163 pp.
69. Fulton, R. (1989). The cacao disease trilogy black pod, monilia pod rot, and witches
broom. Plant Disease 73: 601-603.
70. Fundación Hondureña de Investigación Agrícola (FHIA). (2003). Identificación y
control de la Moniliasis del cacao. Cortés, Honduras. 24 pp.
71. García, R., Ricia, R., Zambrano, C. y Gutiérrez, L. (2006) Desarrollo de un
fungicida biológico con base a una cepa del hongo Trichoderma harzianum
proveniente de la Región Andina Venezolana. En Memorias del Taller
Latinoamericano Biocontrol con Trichoderma y otros antagonistas. Instituto de
Investigaciones de Sanidad Vegetal. Ciudad de La Habana.
72. García, C. (2007). Caracterización del sistema proteolítico de Moniliophthora
roreri (Cif. & Par) Evans et al., causante de la Moniliasis del cacao. Tesis para
aspirar al título de Maestra en Ciencias. Especialista en Producción
Agroalimentaria en el Trópico. Instituto de Enseñanza e Investigación en Ciencias
Agrícolas. Tabasco. México. 116 pp.

Referencias Bibliográficas
73. García, L. (2010). Catálogo de cultivares de cacao del Perú. Ministerio de
Agricultura y Riego. Lima, Perú. 111 pp.
74. Gardes, A. y Brund, T. (1993). ITS primers with enhanced specificity for
Basidiomycetes-Application to the identification of mycorrhizae and rusts. Mol.
Ecol. 2: 113-118.
75. Garrett, M., Robinson, P. (1969). A stable inhibition of spore germination produced
by fungy. Archives of Microbiology 67:370-377.
76. Gonzales, M. (2001). Utilización de Trichoderma spp. para el control de hongos
patógenos de la semilla y del suelo en el cultivo de frijol. Tesis de Máster en
Protección Vegetal. Universidad Agraria de La Habana. La Habana, Cuba. 105 pp.
77. González, I., Infante, D., Peteira, B., Martínez, B., Arias, Y., González, N. y
Miranda, I. (2011). Caracterización bioquímica de aislados de Trichoderma spp.
promisorios como agentes de control biológico. II. Expresión de actividad
glucanasa. Rev. Protección Vegetal 26(1): 13-17.
78. González, A. y D. Roble. (2014). Aislamiento y caracterización del hongo
Moniliophthora roreri (monilia) en frutos de Theobroma cacao L. (cacao) del
cultivar San José del Real de la Carrera, Usulutan. Tesis de Licenciado en Química
y Farmacia. Universidad de El Salvador. San Salvador, El Salvador. 193 pp.
79. Guédez, C., Cañizarez, L., Castillo, C., Olivar, R. (2009). Efecto antagónico de
Trichoderma harzianum sobre algunos hongos patógenos postcosecha de Fragaria
spp. Revista de la Sociedad Venezolana de Microbiología 29(1): 44-49.
80. Guerrero, R. y Arias D. (2006). Determinación de dos especies de hongos del
género Trichoderma (T. koningiopsis-T. stromaticum) para el control de
enfermedades de la mazorca de cacao (Theobroma cacao). Tesis de Ingeniero
Agrónomo. Universidad Estatal de Quevedo. Quevedo - Ecuador. 53 pp.
81. Griffith, G., Nicholson, J., Nenninger, A., Birch, J. y Hedger, N. (2003). Witches
brooms and frosty pods: Two major pathogens of cacao, New Zealand. Journal of
Botany 41(3): 423-435.
82. Hardy, F. (1961). Manual de cacao. Instituto Interamericano de Ciencias Agrícolas.
Turrialba, Costa Rica. 429 pp.

Referencias Bibliográficas
83. Harman, G. (2000). Myths and dogmas of control. Changes in perceptions derived
from research on Trichoderma harzianum T-22. Plant Disease 84(4): 377-393.
84. Harman, G. (2003). Trichoderma harzianum, T. viride, T. koningii, T. hamatum and
other spp. (Deuteromycetes, moniliales). Disponible en:
http://www.ibun.unal.edu.co/r2r7e.html. (Consultado: 26-03-2014).
85. Harman, G. (2006). Overview of Mechanisms and Uses of Trichoderma spp.
Phytopatology 96(2): 190-194.
86. Harman, G y Shoresh, M. (2007). The mechanisms and applications of symbiotic
opportunistic plant symbionts. In: M. Vurro and J. Gressel (Eds). Novel
Biotechnologies for Biocontrol Agent Enhancement and Management. Springer
Dordrecht. pp. 131-155.
87. Hernández, T., Ríos, M. y Arévalo, E. (1991). Enfermedades del cacao en sistema
de producción en la Amazonía Peruana. Programa de Producción Agroindustrial
AD/PERU/459 UNFDAC-PNUD/OSP, Tingo María - Perú. pp. 37-44.
88. Hernández, A., Sierra, A. y Carr, A. (2006). Evaluación in vitro del antagonista de
especies de Trichoderma sobre hongos fitopatógenos que afectan las vitroplantas de
piña (Ananas comosus L. Merr): En Memorias del Taller Latinoamericano.
Biocontrol con Trichoderma y otros antagonistas. Instituto de Investigaciones de
Sanidad Vegetal. Ciudad de La Habana. 150 pp.
89. Hernández, M., Sánchez, P., García, O., Mayek, P., González, P., Quiroz, V.
(2011). Caracterización molecular y agronómica de aislados de Trichoderma spp.
nativos del noreste de México. Rev. Colomb. Biotecnol. 13(2): 176-185.
90. Holliday, P. (1970). Monilia roreri. C.M.I. Descriptions of pathogenic fungi and
bacteria 226. Commowealth Agricultural Bure, London and Reading.
91. Holmes, K., Schroers, H., Thomas, S., Evans, H. y Samuels, G. (2004). Taxonomy
and biocontrol potential of a new species of Trichoderma from the Amazon basin
of South America. Mycological Progress 3(2):199-210.
92. Howell, C. (2003). Mechanisms employed by Trichoderma species in the biological
control of plant diseases: The history and evolution of current concepts. Plant
Disease 87(1): 4-10.

Referencias Bibliográficas
93. IDIAF (Instituto Dominicano de Investigación Agropecuarias y Forestales). (2012).
Conozca y prevenga las enfermedades devastadoras del cacao. Programa de
mercados y competitividad. Disponible en:
http://www.idiaf.gov.do/publicaciones/Publications/cacao.enfermedades/HTML/fil
es/assets/downloads/Untitled-1.indd.pdf (Consultado: 23-03-2013).
94. IICA (Instituto Interamericano de Cooperación para la Agricultura). (2006).
Protocolo estandarizado de oferta tecnológica para el cultivo del cacao. Lima, Perú.
73 pp.
95. Infante, D., Martínez, B., González, N. y Reyes, Y. (2009). Mecanismos de acción
de Trichoderma frente a hongos fitopatógenos. Protección Vegetal 24(1): 14-21.
96. INAMHI (INSTITUTO NACIONAL DE METEOROLOGÍA E HIDROLOGÍA).
(2014). Anuario Meteorológico. Quito, Ecuador. 149 pp.
97. INIAP (INSTITUTO NACIONAL DE INVESTIGACIONES
AGROPECUARIAS). (2000). Programa nacional de cacao y café. Boletín
Informativo. Características agronómicas de clones recomendadas por INIAP.
Pichilingue, Ecuador. 45 pp.
98. INIAP. (2011). Caracterización de los Sistemas de Producción de la Amazonía
ecuatoriana. Informe Final. EECA – INIAP. Joya de los Sachas, Ecuador. 150 pp.
99. INIAP. (2012). Guía del manejo integrado de enfermedades del cultivo de cacao
(Theobroma cacao L.) en la Amazonía ecuatoriana. Joya de los Sachas, Ecuador.
20 pp.
100. Isla, E. y Andrade, B. (2009). Manual para la producción orgánica en las
comunidades nativas de la cordillera del Cóndor. Proyecto Paz y Conservación
Binacional en la Cordillera del Cóndor; Ecuador-Perú Fase II (Componente
Peruano). Lima, Perú. 88 pp.
101. Jaimes, Y. y F. Aranzazu. (2010). Manejo de las enfermedades del cacao
(Theobroma cacao L.) en Colombia, con énfasis en monilia (Moniliophthora
roreri). CORPOICA, Produmedios, Bogotá. 90 pp.
102. Jiménez, J. (1986). Combate biológico de la Moniliasis del cacao (Theobroma
cacao) causada por Moniliophthora roreri mediante bacterias epifitas en la zona

Referencias Bibliográficas
Atlántica de Costa Rica. Tesis de Magister Scientiae. Universidad de Costa Rica y
el Centro Agronómico Tropical de Investigación y Enseñanza. Turrialba, Costa
Rica. 118 pp.
103. Jiménez, J., Galindo, J., Ramírez, C. y Enríquez, G. (1988). Evaluación del
combate biológico y químico de la Moniliasis (Moniliophthora roreri) del cacao en
Costa Rica. En Décima Conferencia Internacional de Investigación en cacao.
pp. 453-456.
104. Johnson, E. (1946). An Improved Slide Culture Technique for the Study and
Identification of Pathogenic Fungi. J Bacteriol 51(6): 689-694.
105. Johnson, J., Bonilla, J. y Agüero, L. (2008). Manual de manejo y producción del
cacaotero. León. Nicaragua. 40 pp.
106. Krauss, U., Hoopen, M., Hidalgo, E., Martínez, A., Arroyo, C., García, J.,
Portuguez, A. y Sánchez, V. (2003). Manejo integrado de la moniliasis
(Moniliophthora roreri) del cacao (Theobroma cacao). Talamanca, Costa Rica. 10:
37-38.
107. Lecuona, R. (1996). Microorganismos patógenos empleados en el control
microbiano de insectos plaga. Argentina. 338 pp.
108. Leickfeldt, E., Samuels, G., Nirenberg, H. y Petrini, O. (1999). A Morphological
and molecular perspective of Trichoderma viride: Is It One or Two Species? Appl.
Envirom. Microbiol. 65(6): 2418-2428.
109. Lee, S. y Taylor, J. (1992). Phylogeny of five fungus-like protoctistan
Phytophthora species, inferred from the internal transcribed spacer of ribosomal
DNA. J. Mol. Biol. Evol. (9): 636-653.
110. León, R., Pino, S., Martínez, B., Liriano, R., Núñez, D. (2012). Aislamiento y
selección de cepas de Trichoderma y su efecto antagonista frente a Sclerotium
rolfsii, Rhizoctonia sp. y Fusarium sp. Centro Agrícola 39(2): 43-48.
111. Lerch, G. (1977). La experimentación agrícola en las Ciencias biológicas y
agrícolas. Científico-Técnico. La Habana, Cuba. 205 pp.
112. Li, S., Yang, L., Zhou, B., Li, Z. y Chen, Y. (2002). Trichoderma viride. Institute of
Microbiology, Yunnan University, China. 55 pp.

Referencias Bibliográficas
113. Lilly, V. y Barnet, H. (1951). Physiology of the fungi. McGraw-Hill. New York,
United States of America. 244 pp.
114. Macagnan, D., Romeiro, R. y Pomella, A. (2005). Eficiencia de baterias do tipo
actinomicetos e endoporogenicas da germinaçao de basidoporos de Crinipellis
perniciosa em frutos de cacau. In Annais VIII Reuniao de control biologóico de
Fitopatógenos. Ministerio de Agricultura pecuária e abastecimiento CEPLAC
(BRA). pp. 97-98.
115. Maridueña, M. (2011). Estudio de la micobiota patogénica de cacao criollo
(Theobroma cacao) en cinco provincias de la costa ecuatoriana y evaluación de la
efectividad in vitro de los bioles locales para su control. Tesis de Máster en
Biotecnología Agrícola con mención en Agricultura Orgánica. Escuela Superior
Politécnica del Litoral, Guayaquil, Ecuador. 154 pp.
116. Martínez, B., Reyes, Y., Infante, D., González, E., Baños, H. y Cruz, A. (2008).
Selección de aislamientos de Trichoderma spp. candidatos a biofungicidas para el
control de Rhizoctonia sp. en arroz. Protección Vegetal 23(2): 118-125.
117. Martínez, B., Infante, D. y Reyes, Y. (2013). Trichoderma spp. y su función en el
control de plagas en los cultivos. Protección Vegetal 28 (1): 1-11.
118. Matamoros, A. 2007. Gestión de la Biodiversidad Amazónica en Ecuador.
PROGRAMA OTCA-BID ATN/DC-9251-R6. 118 pp.
119. Medeiros, F., Pomella, A. De Souza, J., Niella, G., Valle, R., Bateman, R., Fravel,
D., Vinyard, B., y Hebbar, P. (2010). A novel, integrated method for management
of witches broom diseases in cacao, in Bahia, Brazil. Crop Protection 29: 704-713.
120. Meinhardt, L., Rincones, J., Bailey, B., Aime, C., Griffith, G., Zhang, D. y Pereira,
G. (2008). Moniliophthora perniciosa, the causal agent of witches´ broom disease
of cacao: what´s new from this old foe?. Molecular Plant Pathology 9(5): 577-588.
121. Meinhardt, L, Lacerda, G., Thomazella, D., Teixeira, P., Falsarella, M., Schuster,
S., Carlson, J., Guiltinan, M., Mieczkowsi, P., Farmer, A., Ramaraj, T., Crozier, J.,
Davis, R., Shoa, J., Melnick, G. y Bailey, B. (2014). Genoma and secretoma
analysis of the hemibiotrophic fungal pathogen, Moniliophthora roreri, which

Referencias Bibliográficas
causes frosty pod rot disease of cacao: mechanisms of the biotrophic and
necrotrophic phases. BMC Genomics 15: 164.
122. Meléndez, L. (1993). Microambiente, cantidad de esporas en el aire e incidencia del
hongo Moniliophthora roreri (Cif. & Par). Evans et al. bajo tres sistemas de manejo
de sombra leguminosas en cacao (Theobroma cacao L.). Tesis de Magister
Scientiae en Ciencias Agrícolas y Recursos Naturales. Universidad de Costa Rica.
CATIE. Turrialba, Costa Rica. 120 pp.
123. Meléndez, L. y Somarriba, E. (1999). Microambiente y cantidad de esporas de
Moniliophthora roreri bajo tres sistemas de manejo de sombra en leguminosas en
cacao. Agroforestería en las Américas 6(23): 39-41.
124. Mfegue, V., Herail, C., Adreit, H., Mbenoun, M., Techou, Z., Ten, M., Tharreau,
D. y Ducamp, M. (2012). Microsatellite markers for population studies of
Phytophthora megakarya (Pythiaceae), a cacao pathogen in Africa. Am. J. Bot.
September DOI: 10.3732/ajb.1200053.
125. Moreno, A. y Flores, J. (2011). Fomento de la cadena de valor de cacao en
organizaciones de pequeños productores de Esmeraldas y Napo. GIZ. Quito.
Ecuador. 104 pp.
126. Najar, T. y Thomas, S. (2001). El efecto de los microorganismos eficaces en la
supresión del hongo Moniliophthora roreri bajo condiciones de campo y
laboratorio con inoculación artificial. Tesis de Ingeniero Agrónomo. Universidad
Earth. Guácimo, Costa Rica.
127. Navarro, M. y Mendoza, I. (2009). Cultivo del cacao en sistemas agroforestales.
Guía técnica para promotores. Río San Juan, Nicaragua. 70 pp.
128. Negrone, C., León, M. y Villalba, N. (2008). Evaluación del comportamiento de
plantines de Pinus taeda L. con y sin aplicación de fertilización starter
creciendo en sustrato con Trichoderma harzianum. Tesis de Ingeniero Agrónomo.
Universidad de la República. Montevideo, Uruguay. 60 pp.
129. Oliveira, M. y Luz, E. (2005). Doenças do cacauteiro (Theobroma cacao). Manual
de Fitopatología. Ilhéus, Brasil. CEPLAC/ CEPEC/SEFIT. 132 pp.

Referencias Bibliográficas
130. Olivera Costa, V. y Rodríguez, D. (2014). Evaluación del crecimiento, sanidad y
resistencia a heladas de tres híbridos de Eucalyptus grandis con aplicación
de bioestimulantes, Trichoderma harzianum (Trichosoil®) y quitosano
(Biorend®), en plantación. Tesis de Ingeniero Agrónomo. Universidad de la
República. Montevideo, Uruguay.
131. Ongena, M., Henry, G. y Thonart, P. (2009). The roles of cyclic lipopeptides in the
biocontrol activity of plant diseases. Plant Pathology in the 21 st. Century Springer
Verlag. Berlin. pp. 59-69.
132. Ordoñez, L., Gavilánez, C. y Salazar, A. (2011). Secuestro de Carbono en Biomasa
Aérea en Sistemas Agroforestales de Cacao y Café ubicados en la Reserva de
Biosfera Sumaco. Programa GTZ – GESOREN. Quito, Ecuador. 44 pp.
133. Osorio, R. (2010). Estudio del efecto de Trichoderma harzianum en el control de
Moniliophthora roreri en plantas de Theobroma cacao en la provincia de
Esmeraldas. Tesis de Ingeniero Agroindustrial. Escuela Politécnica Nacional.
Quito, Ecuador. 125 pp.
134. Paredes, N. (2009). Manual de cultivo de cacao para la Amazonía ecuatoriana.
Instituto Nacional Autónomo de Investigaciones Agropecuarias, Estación
Experimental Central de la Amazonía DENAREF-Unidad de recursos
fitogenéticos. Manual No. 76. INIAP, Quito, Ecuador. 43 pp.
135. Párraga, J. y Zambrano, J. (2012). Capacidad antagónica de Trichoderma spp., a
nivel in vitro frente a los hongos fitopatógenos Crinipellis perniciosa y
Moniliophthora roreri. Tesis de Ingeniero Agrícola. Escuela Superior Politécnica
Agropecuaria de Manabí Manuel Félix López. Calceta, Ecuador. 67 pp.
136. Peralvo, D. y Saavedra, L. (2006). Validación de biopreparados en base a bacterias
epifitas para el control de la moniliasis en Cacao fino de aroma en el cantón
Valencia de la provincia de Los Ríos. Tesis de Ingeniero Agropecuario. Escuela
Superior Politécnica del Ejército. Santo Domingo, Ecuador. 139 pp.
137. Pérez, J. (2009). Evaluación y caracterización de secciones clonales de cacao
(Theobroma cacao L.) del programa de mejoramiento del CATIE. Tesis de

Referencias Bibliográficas
Magister Scientiae en Agricultura Ecológica. CATIE (Centro Agronómico Tropical
de Investigación y Enseñanza). Turrialba, Costa Rica. 164 pp.
138. Pérez, L., Batlle, A., Chacón, J., Montenegro, V. (2009) Eficacia de Trichoderma
harzianum A34 en el biocontrol de Fusarium oxysporum F. sp. Cubense, agente
causal de la marchitez por Fusarium o mal de Panamá de los bananos en Cuba.
Fitosanidad 13(4):259-263.
139. Pérez, S. (2013). Kallari, historia de un grupo de artesanos y agricultores
emprendedores y patriotitas de la provincia de Napo, Ecuador. Estudio de caso.
Quito, Ecuador. 17 pp.
140. Pérez, E., Milanés, P., Bernal, A., Leiva, M., Garcés, G., Lobato, L., Cañar, L.,
Sierra, Y., Mena, O. (2013). Antagonismo "in vitro" de Trichoderma harzianum
sobre aislados camagüeyanos de Bipolaris oryzae y Sarocladium oryzal. Centro
Agrícola 40(3): 29-36.
141. Phillips, W. (1986). Evaluación de la resistencia de cultivares de cacao (Theobroma
cacao L.) a Moniliophthora roreri (Cif. y Par.). Tesis de Máster en Ciencias.
Universidad de Costa Rica, San José. Centro Agronómico Tropical de Investigación
y Enseñanza, Turrialba (Costa Rica). 119 pp.
142. Phillips-Mora, W.; Galindo, J. (1988). Evaluación de la resistencia de cultivares de
cacao (Theobroma cacao) Moniliophthora roreri (Cif. y Par.). Proceedings of the
Tenth International Cocoa Research Conference. Santo Domingo, Dominican
Republican: Cocoa Producers Alliance. pp. 685-689.
143. Phillips-Mora, W. (2003). Origin, Biogeography, Genetic Diversity and Taxonomic
Affinities of the cacao Fungus Moniliophthora roreri as determined using
Molecular, Phytopathological and Morpho-Physiological Evidence. Tesis de Doctor
en Ciencias.UK: University of Reading. 373 pp.
144. Phillips-Mora, W. (2004). La moniliasis de cacao: una seria amenaza para el cacao
en México. En Memorias de Simposio Nacional de Manejo Fitosanitario de cultivos
tropicales. UACH. Tabasco, México. 170 pp.
145. Phillips-Mora, W., Castillo, J., Krauss, V., Rodriguez, E. y Wilkinson J. (2005).
Evaluation of cacao (Theobroma cacao) clones against seven Colombian isolates of

Referencias Bibliográficas
Moniliophthora roreri from four pathogen genetic groups. Plant Pathology 54, 483-
490.
146. Phillips, M., Coutiño, F., Ortiz, P., López, J., Hernández, A. y Aime, C. (2006).
First report of Moniliophthora roreri causing frosty pod rot (moniliasis disease) of
cocoa in México. Plant pathology 55-58.
147. Phillips-Mora, W. (2006a). Cincuenta años de la presencia de Moniliasis de cacao
en Centro América: Cronología de su impacto y su dispersión desde Panamá hasta
México. In International Cocoa Research Conference. San José, Costa Rica. Actas
Proceeding Nigeria, COPAL (1): 1039-1047.
148. Phillips-Mora, W. (2006b). La moniliasis del cacao: un enemigo que podemos y
debemos vencer. Taller regional andino de aplicación tecnológica en el cultivo de
cacao. Quevedo, Ecuador. pp. 21-25.
149. Phillips-Mora, W. y Wilkinson, M. (2007). Frosty pod of cacao: A disease with a
limited geographic range but unlimited potential for damage. Symposium Cacao
Diseases: Important threats to chocolate Production Worldwide. Phytopathology
97(12): 1644-1647.
150. Phillips-Mora, W., Aime, M. y Wilkinson, M. (2007). Biodiversity and
biogeography of the cacao (Theobroma cacao) pathogen Moniliophthora roreri in
tropical America. Plant Pathology 56: 911-922.
151. Phillips-Mora, W., Baqueros, F., Melnick, RL., y Bailey, BA. 2015. First report of
frosty pod rot caused by Moniliophthora roreri on cacao in Bolivia. New Disease
Reports 31:29. DOI: 10.5197/j. 2044-0588.
152. Porras, V. (1983). Epifitiología de la moniliasis (Monilia roreri Cif y Par.) del
cacao y su relación con la producción del árbol en la zona de Matina. El cacaotero
colombiano 25: 28-29.
153. Porras, V. (1985). Determinación de la estabilidad de la Resistencia a Monilia
roreri en cultivares de cacao en dos zonas de Costa Rica. Tesis de Magister
Scientiae. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE).
Turrialba, Costa Rica. 124 pp.

Referencias Bibliográficas
154. Porras, V. y J. Sánchez. (1991). Enfermedades del cacao. Fundación Hondureña de
Investigación Agrícola. Instituto Interamericano de Cooperación para la
Agricultura. Serie Comunicación Tecnología y Desarrollo. Fascículo No. 5. La
Lima. Cortez. Honduras. 14 pp.
155. Pozzo, T., Cabero, K., Nordberg, N., Karlsson, E. y Liden, G. (2015). Cellulolytic
and xylanolytic activities of Bionectria and Hypocrea strains isolated from Quinoa
and Brave straw species native to the Bolivian Altiplano. Plant Sciences, California,
United States of America.
156. PROAMAZONIA (Programa para el Desarrollo de la Amazonía). (2003).
Caracterización de las zonas productoras de cacao en el Perú y su competitividad.
Lima, Perú. 208 pp.
157. Punja, Z. y Grogan, R. (1983). Hyphall interactions and antagonism among isolates
and single basidiospores strains of Athelia (Sclerotium rolfsii). Phytopathology
72: 379-382.
158. Ramírez, S. (2008). La Moniliasis un desafío para lograr la sostenibilidad del
sistema cacao México. Tecnología en Marcha 21(1): 97-110.
159. Reyes, R., Martínez, B. y Infante, D. (2008). Evaluación de la actividad antagónica
de trece aislamientos de Trichoderma spp. sobre Rhizoctonia sp. Protección Vegetal
23(2): 112-117.
160. Riddel, R. (1950). Permanent strained mycologycal preparation obtained by slide
cultures. Micologia (82): 265-270.
161. Rifai, M. (1969). A revision of the genus Trichoderma. Res. Mycol. 116: 1-56.
162. Robles, B. (2008). Validación de biopesticidas en base a bacterias epifitas para el
control de la moniliasis (Moniliophthora roreri Cif y Par. Evans et al.) en el
cultivo de cacao híbrido CCN-51 en Santo Domingo, provincia Santo Domingo de
los Tsáchilas. Tesis de Ingeniero Agropecuario. Escuela Politécnica del Ejército.
Santo Domingo de los Tsáchilas, Ecuador. 31 pp.
163. Rodríguez, L. (2004). Caracterización de clones de cacao por respuesta a monilia
(Moniliophthora roreri (Cif & Par.) Evans et al.) en Santander. CORPOICA.
Santander, Colombia. 6 pp.

Referencias Bibliográficas
164. Rojas, E., Rehner, S., Samuels, G., VanBael, S., Here, E., Cannon, P., Chen, R.,
Pang, J., Wang, R., Zhang, Y., Peng, Y. y Sha, T. (2010). Colletotrichum
gloesporioides s.l. associated with Theobroma cacao and other plants in Panamá:
multilocus phylogenies distinguish host-associated pathogens from asymptomatic
endophytes. Mycologia 102(6): 1318-1338.
165. Romero-Arenas, O., Huerta, M., Damián, M., Domínguez, F. y Arellano, D. (2009).
Características de Trichoderma harzianum, como agente limitante en el cultivo de
hongos comestibles. Biotecnología 11(2): 143-151.
166. Romero, G., Olivera Costa, V. y Rodríguez, D. (2015). Trichoderma harzianum
como agente de control biológico. Universidad de la República Montevideo,
Uruguay. 65 pp.
167. Rondón, J. y Cumana, L. (2005). Revisión Taxonómica del género Theobroma
(Sterculiaceae) en Venezuela. Acta. Bot. Venez. 28(1): 113-133.
168. Rorer, J. (1918). Enfermedades y plagas del cacao en el Ecuador y métodos
apropiados al cultivo. Trad. al español por Abelardo Pachano. Guayaquil, Ecuador.
Asociación de Agricultores. pp. 17-40.
169. Rossi, C. (2010). Evaluación de Trichoderma harzianum como agente biopromotor
y de biocontrol en plantines de Eucalyptus dunii Maiden. Tesis de Ingeniero
Agrónomo. Universidad de la República. Montevideo, Uruguay. 129 pp.
170. Rumbos, R., Hernández, J., Moya, A., Quevedo, H. y Gómez, C. (2005).
Comportamiento in vitro del hongo (Moniliophthora roreri Cif & Par) causante de
la moniliasis de cacao en la zona sur del lago de Maracaibo. Congreso
Latinoamericano de Fitopatología. En Memorias de Congreso Venezolano de
Fitopatología. Barquisimeto, Venezuela. 37 pp.
171. Samaniego, J., Ulioa, A. y Herrera, T. (1989). Hongos del suelo antagónicos de
Phymatorichum omnivorum, Fitopatología Mexicana 7(1): 86-95.
172. Samuels, G., Dodd, S., Bing-Sheng, L., Petrini, O., Schroers, H. y Druzhinina, I.
(2006a). The Trichoderma koningii aggregate species. Studies in Mycology 56: 67-
133. DOI: 3114/sim.2006.56.03.

Referencias Bibliográficas
173. Samuels, G., Suárez, C., Solís, K., Holmes, K., Thomas, S., Ismael, A., Evans, H.
(2006b). Trichoderma theobromicola and T. paucisporum: two new species isolated
from cacao in South America. Mycological Research 110: 381-392.
174. Samuels, G., Ismaiel, A., Souza, J., Chaverri, P. (2012a). Trichoderma stromaticum
and its overseas relatives. Micol. Progress 11: 215-255. DOI: 10.1007/s11557-011-
0743-4.
175. Samuels, G., Ismael, A., Mulaw, T., Szakacs, G., Druzhinina, I., Kubicek, C. y
Jaklitsch, W. (2012b). The Longibrachiatum Clade of Trichoderma: a revision with
new species. Fungal Diversity 55: 77-108.
176. Sambrook, J. y Rusell, D. (2001). Molecular Cloning. A Laboratory Manual. 3ra
ed. Cold Spring Harbor Laboratory. New York. 2100 pp.
177. Sánchez, J., Brenes, O., Phillips, W. y Enríquez, G. (1987). Methodology for
inoculating pods whit the fungus Moniliophthora (Monilla) roreri. Proceedings of
the Tenth International Cocoa Research Conference. Santo Domingo, Dominican
Republican: Cocoa Producers Alliance. pp. 467-471.
178. Sánchez, J. y González, L. (1989). Metodología para evaluar la susceptibilidad a
moniliasis en cultivares de cacao (Theobroma cacao). Turrialba, Costa Rica.
39: 461-468.
179. Sánchez, L., Gamboa, E. y Rincón, J. (2003). Control químico y cultural de la
moniliasis (Moniliophthora roreri Cif & Par) del cacao (Theobroma cacao L.) en el
estado Barinas. México. Revista Facultad de Agronomía 20(2): 188-194.
180. Sánchez, H. (2011). Determinación de la actividad enzimática extracelular de
Endo-1,3(4)-ß-glucanasa y amplificación de genes Endo-1,3(4)-ß-glucanasa a partir
de Moniliophthora roreri. Tesis de Maestro en ciencias en Producción
Agroalimentaria en el Trópico. Escuela de Posgraduados. Tabasco, México.
181. Sánchez, H., Bautista, C., Boldo, X., Ortiz, F., Ramos, J. y Carrera, D. (2011).
Endo-1,4-beta-glucanases enzymatic activity from Moniliophthora roreri grown in
solid state fermentation. Ciencia de Alimentos e Ingeniería, Colegio de
Postgraduados, Periférico Carlos A. Molina, km 3.5 Carr. Cardenas-Huimanguillo,
H. Cárdenas, Tabasco 86500, México. 115 pp.

Referencias Bibliográficas
182. Sánchez, F., Garcés, F. (2012). Moniliophthora roreri (Cif y Par) Evans et al. en el
cultivo de cacao. Scientia Agropecuaria 3: 249-258.
183. Sánchez-Mora, F., Garcés, R., Vera, F., Ramos, A., Troya, F. y Díaz, G. (2011).
Cuantificación de enfermedades en mazorcas de cacao (Theobroma cacao L.) en la
zona central del Litoral Ecuatoriano. En Memorias del VIII Simposio Internacional
de Recursos Genéticos para América Latina y El Caribe, Quito. Ecuador. 523 pp.
184. Sandoval, G., Ambrecht, H. y Granada, G. (1987). Posibilidades de control
biológico de moniliasis de cacao. El Cacaotero Colombiano 10(34): 39.
185. Saquicela, D. (2010). Evaluación económica de los componentes del manejo
integrado para el control de enfermedades de cacao tipo Nacional. Tesis de
Ingeniero Agropecuario. (ESPE) Escuela Politécnica del Ejército. Santo Domingo,
Ecuador. 13 pp.
186. Schmitz, W. (1985). Studies in the Atlantic coast of Costa Rica on the
epidemiology of the fungus Moniliophthora roreri Evans et al. in cacao
(Theobroma cacao L.) Tesis de Doctor en Ciencias. Göttingen, Alemania. Georg
August University. 190 pp.
187. Seng, J., Herrera, G., Vaughan, C., McCoy, M. (2014). Uso de Trichoderma fungi
in spray solutions to reduce M. roreri infection of Theobroma cacao fruits in
Northeastern Costa Rica. Biología Tropical 62(3): 899-907.
188. Shaner, E. y Finney, F. (1977). The effect of nitrogen fertilization on the expression
of slow-mildewing resistance in Knox wheat. Phytopathology 67: 1051-1056.
189. Solís, K. (1999). Determinación de organismos antagónicos a Monilia roreri a
partir de mazorcas de cacao dejadas en el suelo. Tesis de Ingeniero Agrónomo.
Universidad de Guayaquil, Guayaquil, Ecuador. 55 pp.
190. Solís, K. y Suárez-Capello, C. (2006). Uso de Trichoderma spp. para control del
complejo Moniliasis-Escoba de bruja del cacao en Ecuador. Instituto Ecuatoriano
de Investigaciones Agropecuarias. Quevedo, Ecuador. 8 pp.
191. Stackebrandt, E. y Goebel, B. (1994). Taxonomic note: a place for DNA-DNA
reassociation and 16s rRNA sequence analysis in the present species definition in
bacteriology. International Journal of Systematics Bacteriology 44: 846-849.

Referencias Bibliográficas
192. Stamps, J. (1985). Phytophthora palmivora. CMI Descriptions of Pathogenic Fungi
and Bacteria. Nº 831. Set Nº 84.
193. Sterling, A., Hermida, M., Rodríguez, C., Salas, Y., Nieto, M., Caicedo, D. (2015).
Reacción a Moniliophthora roreri en Theobroma spp. en Caquetá, Colombia.
Summa Phytopathol. 41(3): 183-190.
194. Suárez, C. (1971). Estudio del mecanismo de penetración y del proceso de
infección de Monilia roreri Cif & Par. en frutos de cacao (Theobroma cacao L.).
Tesis de Ingeniero Agrónomo. Escuela Superior Politécnica del Litoral. Guayaquil,
Ecuador. 59 pp.
195. Suárez, L. (2006). Aislamiento e identificación de M. roreri causante de la monilla
en municipios de nororiente colombiano y ensayos preliminares para su control
biológico. Universidad Francisco de Paula Santander. Respuestas 11(1): 3-8.
196. Suárez, C. (2008). Alternativas tecnológicas para el manejo integrado de plagas y
enfermedades del cultivo de cacao. En Congreso Regional sobre alternativas
tecnológicas en la Amazonia Ecuatoriana. INIAP (Instituto Nacional de
Investigaciones Agropecuarias). Joya de los Sachas, Ecuador. 30 pp.
197. Suárez, L., y Rangel, A. (2014). Aislamiento de microorganismos para control
biológico de Moniliophthora roreri. Acta Agronómica 62(4): 370-378.
198. Suárez, L. (2016). Identificación molecular de aislamientos de Moniliophthora
roreri en huertos de cacao de Norte de Santander, Colombia. Acta Agronómica
65 (1): 51-57.
199. Suárez-Castellá, M., Martín, L., Triana, R., Pérez, Z., Rodríguez, M., Agromonte,
D., Sarría, Z. (2009). Procedimiento metodológico para la conformación,
implementación y perfeccionamiento de sistemas de calidad para biofábricas.
Biotecnología Vegetal 9(3): 141-152.
200. Sun, Z., Li, S. y Sun, M. (2015). Selection of reliable reference genes for gene
expression studies in Clonostachys rosea 67-1 under sclerotial induction.
Microbiol. Methods 114: 62-65.

Referencias Bibliográficas
201. Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6:
Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. and Evol.
30: 2725-2729.
202. Tiburcio, L., Costa, F., Carazolle, C., Mondego, C., Schuster, E., Carlson, J.,
Guiltinan, J., Bailey, A., Mieczkowski, P., Meinhardt, L. y Pereira, W. (2010).
Genesa Acquired by Horizontal Transfer Are Potentially in Moniliophthora
perniciosa and Moniliophthora roreri, two of the Mayor Pathogens of cacao.
Journal of Molecular Evolutions 70(1): 85-97.
203. Torres de la Cruz, M. (2010). Progreso temporal y manejo integrado de la
moniliasis Moniliophthora roreri (Cif & Par.). Evans et al. del cacao (Theobroma
cacao) en Tabasco México. Tesis de Doctor en Ciencias, Fitosanidad. Colegio de
Postgraduados. Montecillo, México. 99 pp.
204. Torres de la Cruz, M., Ortiz, C., Téliz, D., Mora, A. y Nava, C. (2011). Temporal
progress and integrated management of frosty pod rot (Moniliophthora roreri) of
cocoa in Tabasco Mexico. Plant Pathology 93(1): 31-36.
205. Townsend, G. y Heuberger, J. (1943). Methods for estimating losses caused by
diseases in fungicide experiments. Plant Dis. Rep. 27: 340-343.
206. Urdaneta, G. y Delgado, A. (2007). Identificación de la micobiota del filoplano del
cacaotero (Theobroma cacao L.), en el municipio Carraciolo Parra Olmedo, Estado
Mérida. Venezuela. Revista Facultad de Agronomía. 24: 47-68.
207. Uquillas, L. (2004). Inducción de la germinación para mejorar la eficiencia de dos
agentes antagónicos para el control de la Monilia (Crinipellis roreri) del cacao
(Theobroma cacao). Tesis de Magister Scientiae. Centro Agronómico Tropical de
Investigación y Enseñanza (CATIE). Turrialba, Costa Rica. 85 pp.
208. Van der Plank, J. (1963). Plant Diseases: Epidemics and Control. Academic Press.
London. 349 pp.
209. Vásquez, D.; Montes, R.; Pérez, A. y Flores, H. (2014). Aceites esenciales y
extractos acuosos para el manejo in vitro de Fusarium oxysporum f. sp. lycopersici
y F. solani. Revista Mexicana de Fitopatología 31(2): 170-179.

Referencias Bibliográficas
210. Villamil, J., Blanco, J. y Rosero, S. (2012). Evaluación in vitro de microorganismos
nativos por su antagonismo contra Moniliophthora roreri Cif & Par en Cacao
(Theobroma cacao L.). Universidad Nacional de Colombia. Medellín, Colombia.
Revista Facultad Nacional de Agronomía 65(1): 6305-6315.
211. Villamil, J., Viteri, S. y Villegas, W. (2015). Aplicación de antagonistas
microbianos para el control biológico de Moniliophthora roreri (Cif & Par) en
Theobroma cacao L. bajo condiciones de campo. Medellín. Revista Facultad
.Nacional de Agronomía. 68(1): 6441-7450.
212. Villavicencio, M. (2010). Caracterización morfológica, fisiológica y patogénica de
Moniliophthora roreri aislados de cinco provincias de la Costa ecuatoriana. Tesis
de Ingeniero Agrícola y Biológico. Escuela Superior Politécnica del Litoral.
Guayaquil, Ecuador. 142 pp.
213. Villegas, C. (1979). Moniliasis; Bibliografía parcialmente anotada. Comp. por
C. Villegas y G. Enríquez. Turrialba, Costa Rica. CATIE. 45 pp.
214. Vinale, F., Sirasithamparamb, K., Ghisalbertic, E., Marraa, R., Woo, L., Lorito, M.
(2008). Trichoderma-plant-pathogen interactions. Soil Biology & Biochemistry. 40:
1-10.
215. White, J., Bruns, T., Lee, S. y Taylor, J. (1990). Amplification and Direct
sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A
guide to methods and applications. Eds. Innis, M.A., D.H. Gelfand, J.J. Sninsky and
T.J. White. Academic Press, New York. 315-322 pp.

Anexos

ANEXOS
Anexo 1. Fincas en las que se realizó la recolección de las mazorcas de cacao
FINCA PROPIETARIO PROVINCIA CANTÓN SECTOR EDAD DE LA
PLANTACIÓN
(años)
1 Sr. Carlos Pozo Napo Tena Colonia Bolívar 7
2 Sr. Cesar Dahua Napo Tena Ahuano 8
5 Sra. Mariana Rodríguez Napo Arosemena
Tola Arosemena Tola 7
6 Sr. Ángel Pozo Napo Tena Puerto Misahualli 7
7 Sr. Wilson Avilés Pastaza Arajuno Las peñas del pozo 8
8 Sr. Patricio Carreño Morona
Santiago Palora La Comanche 5
Anexo 2. Ubicación de los sitios donde se recolectaron las mazorcas con síntomas de
Moniliasis, A Ecuador, B cantones Napo, Pastaza y Morona Santiago, C fincas objeto de
estudio

Anexo 3. Datos climatológicos de los lugares de procedencia de los aislados de M. roreri
PROVINCIAS ALTITUD
(msnm)
TEMPERATURA
(º C)
PRECIPITACIÓN
(mm)
H.R
(%)
PASTAZA 1 100 23 ± 2 4 158.8 88
NAPO 600 24 ± 2 4 195.8 84
MORONA SANTIAGO 1 250 22 ± 2 3 119.9 92

Anexo 4. Escala de clasificación de síntomas según Sánchez et al. (2003)
Valor
Interna
(% de almendras
afectadas)
Externa
(Clasificación de síntomas)
0 0 Fruto sano
1 0 - 20 Presencia de puntos aceitosos (hidrosis)
2 21 - 40 Presencia de tumefacción y/o madurez
prematura
3 41 - 60 Presencia de mancha chocolate
4 61 - 80 Presencia de micelio que cubre hasta la cuarta
parte de la mancha parda
5 > 81 Presencia de micelio que cubre hasta la cuarta
parte de la mancha chocolate

Anexo 5. Composición del medio V8 modificado
Medio de cultivo Fórmula
V8 modificado 200 mL jugo V8, 15 g agar, 15 g de
extracto de malta (Difco), 3 g Carbonato
de calcio, esto se afora a 1 000 mL de
agua destilada.
Composición del Jugo V8: 45 mg de
sodio, 700 mg de potasio, 7 g
carbohidratos, 5 g azúcar y 1 g proteína.

Anexo 6. Escala de valoración de severidad externa propuesta por Brenes (1983)
Escala Características Correspondencia visual
Grado 0 Fruto sano
Grado 1 Puntos aceitosos
Grado 2 Tumefacción o clorosis
Grado 3 Mancha (necrosis)
Grado 4 Micelio hasta un 25% de
la mancha
Grado 5 Micelio en más del 25%
de la mancha

Anexo 7. Escala de valoración de severidad interna propuesta por Sánchez et al. (1987)
Escala Características Correspondencia visual
Grado 0 Cero área necrosada
Grado 1 1-20% del área necrosada
Grado 2 21-40% área necrosada
Grado 3 41-60% área necrosada
Grado 4 61-80% área necrosada
Grado 5 100% área necrosada
Anexo 8. Identificación de hongos anamorfos asociados a frutos de cacao con síntomas de
Moniliasis en la provincia del Napo
Finca Parroquia Sector Géneros de hongos
anamorfos
Finca 1 Ahuano Colonia Bolívar Cylindrocladium sp.
Finca 2 Ahuano Ahuano Colletotrichum sp.
Finca 3 Archidona San Pablo Dichobotrys sp.
Finca 4 Archidona Centro Shiguango Cylindrocladium sp.
Finca 5 Arosemena Tola Arosemena Tola Phytophthora sp.
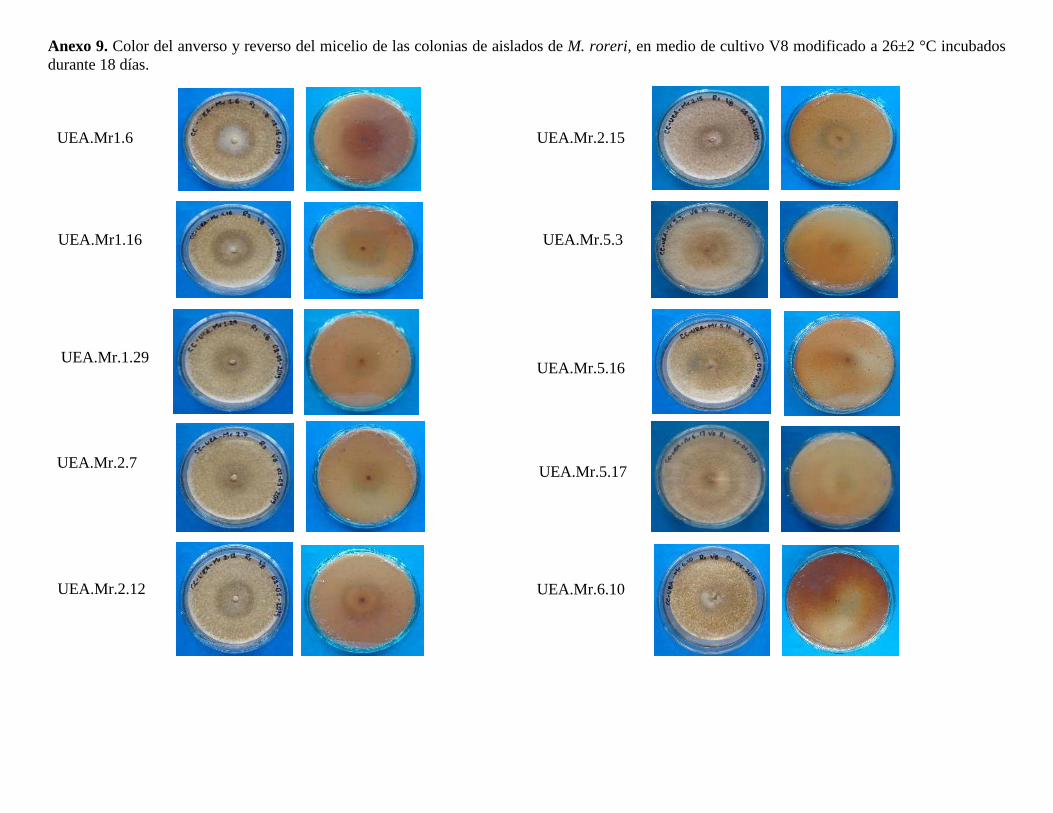
UEA.Mr1.6 UEA.Mr.2.15
UEA.Mr1.16 UEA.Mr.5.3
UEA.Mr.1.29
UEA.Mr.5.16
UEA.Mr.2.7
UEA.Mr.2.12
UEA.Mr.5.17
UEA.Mr.6.10
Anexo 9. Color del anverso y reverso del micelio de las colonias de aislados de M. roreri, en medio de cultivo V8 modificado a 26±2 °C incubados
durante 18 días.

UEA.Mr.7.13
UEA.Mr.7.16
UEA.Mr.8.9
UEA.Mr.8.10
Mr
UEA.Mr.8.11 UEA.Mr.7.17
UEA.Mr7.20 UEA Mr 8.19
UEA.Mr.7.28 UEA.Mr 8.20
Color del anverso y reverso del micelio de las colonias de aislados de M. roreri, en medio de cultivo V8 modificado a 26±2 °C incubados durante 18
días.

Anexo 10. Cuantificación del ADN de los hongos fitopatógenos
#
Sample
ID User name Date and Time
Nucleic
Acid Conc. Unit A260 A280 260/280 260/230
Sample
Type Factor
1 1 Usuario 25/12/2015 12:14:19 234.3 ng/B5l 8.686 3.921 2.22 2.25 DNA 50
2 2 Usuario 25/12/2015 12:14:57 50.6 ng/B5l 1.011 0.487 2.08 1.25 DNA 50
3 3 Usuario 25/12/2015 12:15:22 34.3 ng/B5l 0.686 0.341 2.01 2.26 DNA 50
4 4 Usuario 25/12/2015 12:15:48 44.2 ng/B5l 0.884 0.441 2 2.01 DNA 50
5 5 Usuario 25/12/2015 12:16:10 243.4 ng/B5l 6.871 3.091 2.22 2.43 DNA 50
6 6 Usuario 25/12/2015 12:16:38 27.1 ng/B5l 0.341 0.181 1.88 1.98 DNA 50
7 7 Usuario 25/12/2015 12:17:10 365 ng/B5l 9.299 4.117 2.26 2.43 DNA 50
8 8 Usuario 25/12/2015 12:17:34 20.1 ng/B5l 0.203 0.105 1.93 1.29 DNA 50
9 9 Usuario 25/12/2015 12:18:00 26.1 ng/B5l 0.322 0.161 2.01 1.92 DNA 50
10 10 Usuario 25/12/2015 12:18:25 26.2 ng/B5l 0.324 0.168 1.92 0.92 DNA 50
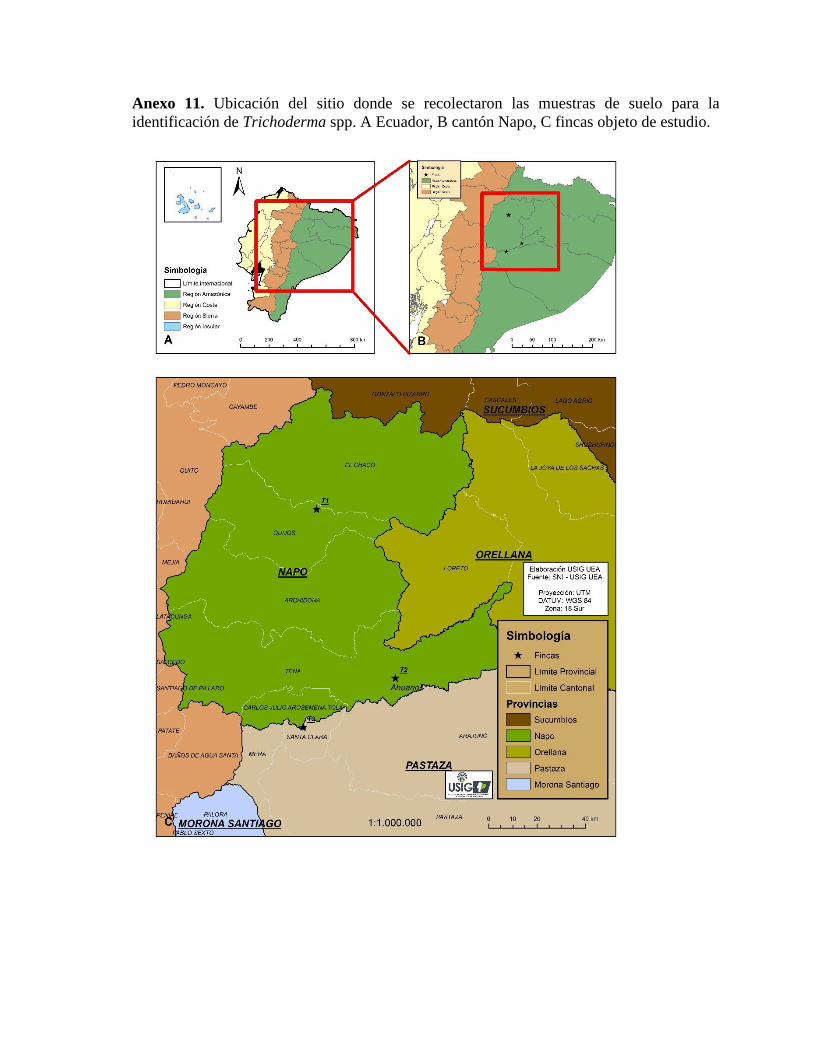
Anexo 11. Ubicación del sitio donde se recolectaron las muestras de suelo para la
identificación de Trichoderma spp. A Ecuador, B cantón Napo, C fincas objeto de estudio.
Related Documents











