Chapter 23 23.1 Introduction We carried out research on “Global Change Impacts on Terrestrial Ecosystems in Monsoon Asia” (TEMA) as a GCTE core research project from 1995 to 2003. The pur- pose of TEMA was to predict the effects of elevated CO 2 and climate change on the distribution and structure of forest ecosystems in eastern Asia, and to determine the associated feedback effects to the global carbon cycle (Hirose et al. 1998). The target region is characterized by continuous forested biomes from tropic to sub-arctic zones under prevailing humid climates, and by the on- going pressures of land use and pollution. The compari- son between a latitudinal gradient where seasonality is amplified with latitude, and an altitudinal gradient in tropical mountains without seasonal thermal change enables us to analyze and describe the climate-vegeta- tion linkage. We made detailed investigations mainly in such core sites as Mount Kinabalu (Malaysian Borneo), the Lake Biwa basin (central Japan), and the Tomakomai Forest (northern Japan), and in many satellite sites scatteed across the target region. The interim achieve- ments of TEMA have been published in separate papers as well as special issues (Hirose and Walker 1996; Nakashizuka et al. 1999). The overall synthesis will be published as a journal special issue (Kohyama et al. 2005). We do not summarize these overall results here. Our purpose in this chapter is to emphasize certain unique aspects of TEMA, which were not emphasized in other GCTE activities. We particularly focused on (1) the linkage of physiological processes of foliage canopies to landscape-scale processes of population demography and community dynamics, (2) integration of forest ecosystem processes into watershed-scale budgets, and (3) attention to lake ecosystems characterizing watershed-scale land systems. Other GCTE research has focused heavily on responses of physiological and ecosystem processes to global change. However, the prediction of the long-term response of for- est systems to global change requires the coupling of eco- system physiology and tree population demography. To interface the gap between them, TEMA focused on the branch-module-based reconstruction of tree foliage devel- opment at smaller scale, and the shift of vegetation zones determined by the dispersion of propagules and the sup- pression by resident vegetation at larger spatial scales. Another aspect of TEMA was watershed-scale inte- gration. Recently growing evidence shows that the ma- jority of lotic and lentic waters are supersaturated in terms of CO 2 (Cole et al. 1994), suggesting that the in- land aquatic systems function to return C that was fixed in the terrestrial ecosystems to the atmosphere through the biological activities. The inland aquatic systems also function to bury terrigenous organic carbon. According to Dean and Gorham (1998), lakes, reservoirs and wet- lands bury organic carbon more than the oceans do, al- though these cover less than 2% of Earth’s surface. Thus, the aquatic systems are likely “hot spots” for carbon pro- cessing in terrestrial ecosystems (Hanson et al. 2004). We therefore focused on carbon flux along the atmo- sphere-forest-stream-lake continuum at a watershed scale. We found that the stream chemistry reflects biogeochemi- cal metabolism at upstream forest ecosystems. In addition, we found that the carbon metabolism and biogeochemical cycling of inland aquatic systems are tightly linked with allochthonous organic matter input from the watershed and function to determine the fate of terrigenous carbon from the forests. These achievements from TEMA suggest that the effects of global and regional human impacts on terrestrial ecosystems should be examined at the scale of a watershed, as a unit of spatial integration. 23.2 Responses of Plant Communities to the Global Change: Scaling from Leaf to Landscape Through Individual Plant In many natural plant communities, the local environ- ment one individual experiences is different from that for others. Plants in a stand compete for resources with each other, and competition alters morphological and physiological traits of plants. These alterations in turn change the microclimate and resource acquisition of in- dividuals in the stand, and consequently influence the growth of individuals (Hikosaka et al. 1999). However, studies on plant individuals growing in a stand are very few, which may make it difficult to predict responses of plants to CO 2 at a stand level. Terrestrial Ecosystems in Monsoon Asia: Scaling up from Shoot Module to Watershed Takashi Kohyama · Jotaro Urabe · Kouki Hikosaka · Hideaki Shibata · Takahito Yoshioka · Eiichi Konohira Jun Murase · Eitaro Wada

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chapter 23
23.1 Introduction
We carried out research on “Global Change Impacts onTerrestrial Ecosystems in Monsoon Asia” (TEMA) as aGCTE core research project from 1995 to 2003. The pur-pose of TEMA was to predict the effects of elevated CO2and climate change on the distribution and structure offorest ecosystems in eastern Asia, and to determine theassociated feedback effects to the global carbon cycle(Hirose et al. 1998). The target region is characterized bycontinuous forested biomes from tropic to sub-arcticzones under prevailing humid climates, and by the on-going pressures of land use and pollution. The compari-son between a latitudinal gradient where seasonality isamplified with latitude, and an altitudinal gradient intropical mountains without seasonal thermal changeenables us to analyze and describe the climate-vegeta-tion linkage. We made detailed investigations mainly insuch core sites as Mount Kinabalu (Malaysian Borneo),the Lake Biwa basin (central Japan), and the TomakomaiForest (northern Japan), and in many satellite sitesscatteed across the target region. The interim achieve-ments of TEMA have been published in separate papersas well as special issues (Hirose and Walker 1996;Nakashizuka et al. 1999). The overall synthesis will bepublished as a journal special issue (Kohyama et al. 2005).We do not summarize these overall results here.
Our purpose in this chapter is to emphasize certainunique aspects of TEMA, which were not emphasized inother GCTE activities. We particularly focused on (1) thelinkage of physiological processes of foliage canopies tolandscape-scale processes of population demography andcommunity dynamics, (2) integration of forest ecosystemprocesses into watershed-scale budgets, and (3) attention tolake ecosystems characterizing watershed-scale land systems.Other GCTE research has focused heavily on responses ofphysiological and ecosystem processes to global change.However, the prediction of the long-term response of for-est systems to global change requires the coupling of eco-system physiology and tree population demography. Tointerface the gap between them, TEMA focused on thebranch-module-based reconstruction of tree foliage devel-opment at smaller scale, and the shift of vegetation zones
determined by the dispersion of propagules and the sup-pression by resident vegetation at larger spatial scales.
Another aspect of TEMA was watershed-scale inte-gration. Recently growing evidence shows that the ma-jority of lotic and lentic waters are supersaturated interms of CO2 (Cole et al. 1994), suggesting that the in-land aquatic systems function to return C that was fixedin the terrestrial ecosystems to the atmosphere throughthe biological activities. The inland aquatic systems alsofunction to bury terrigenous organic carbon. Accordingto Dean and Gorham (1998), lakes, reservoirs and wet-lands bury organic carbon more than the oceans do, al-though these cover less than 2% of Earth’s surface. Thus,the aquatic systems are likely “hot spots” for carbon pro-cessing in terrestrial ecosystems (Hanson et al. 2004).
We therefore focused on carbon flux along the atmo-sphere-forest-stream-lake continuum at a watershed scale.We found that the stream chemistry reflects biogeochemi-cal metabolism at upstream forest ecosystems. In addition,we found that the carbon metabolism and biogeochemicalcycling of inland aquatic systems are tightly linked withallochthonous organic matter input from the watershedand function to determine the fate of terrigenous carbonfrom the forests. These achievements from TEMA suggestthat the effects of global and regional human impacts onterrestrial ecosystems should be examined at the scale of awatershed, as a unit of spatial integration.
23.2 Responses of Plant Communities to the GlobalChange: Scaling from Leaf to LandscapeThrough Individual Plant
In many natural plant communities, the local environ-ment one individual experiences is different from thatfor others. Plants in a stand compete for resources witheach other, and competition alters morphological andphysiological traits of plants. These alterations in turnchange the microclimate and resource acquisition of in-dividuals in the stand, and consequently influence thegrowth of individuals (Hikosaka et al. 1999). However,studies on plant individuals growing in a stand are veryfew, which may make it difficult to predict responses ofplants to CO2 at a stand level.
Terrestrial Ecosystems in Monsoon Asia:Scaling up from Shoot Module to Watershed
Takashi Kohyama · Jotaro Urabe · Kouki Hikosaka · Hideaki Shibata · Takahito Yoshioka · Eiichi KonohiraJun Murase · Eitaro Wada

286 CHAPTER 23 · Terrestrial Ecosystems in Monsoon Asia: Scaling up from Shoot Module to Watershed
One of the important foci of TEMA was a process-based analysis of environmental responses in populationdemography in plant communities. We adopted severalapproaches. We carried out an experimental study ofmonospecific stands as the simplest plant community.We also constructed a shoot-module-based simulationmodel, and an individual-plant-based model, to predictthe response of plant communities to global change atdifferent spatial scales.
23.2.1 Competition among Individual Plantsin Even-Aged Monospecific Standsat Elevated CO2
Competition among individuals in plant populations iscategorized into two modes, i.e., symmetric and asym-metric competition (Weiner 1990). Symmetric competi-tion indicates that individuals in a stand acquire re-sources in proportion to their sizes while in asymmetriccompetition large individuals acquire resources more thanproportionately. For instance competition for light is asym-metric in dense stands (Ford and Diggle 1981; Weiner 1986;Jurik 1991; Nagashima 1999; Hikosaka et al. 1999), whilethat for nutrients is more symmetric (Wilson 1988;Weiner et al.1997; Hikosaka and Hirose 2001). The modeof competition is critical in development of size inequal-ity in the stand. Size inequality is generally assessed withthe coefficient of variation (Weiner 1990). Symmetriccompetition, where plant growth is proportional to thesize, does not alter size inequality, while asymmetric com-petition increases size inequality in the stand.
Since diffusion of CO2 within plant stands is very fast,competition for CO2 is unlikely to occur among individu-als (Jones 1992). Even though elevated CO2 may benefitall individuals in the stand, the enhancement of growthby elevated CO2 may indirectly alter the mode of compe-tition (Wayne and Bazzaz 1997). There are two alterna-tive hypotheses in this respect. One is that elevated CO2makes the competition more asymmetric and increasessize inequality in the stand, because enhanced growth oflarger individuals suppresses light acquisition of smallerindividuals. The other is that elevated CO2 reduces thedegree of asymmetry in competition and consequentlysize inequality. This is because the end-product inhibi-tion of photosynthesis due to elevated CO2 (Stitt andKrapp 1999) may be stronger in larger plants exposed tohigh light, and because the reduction of the light com-pensation point of photosynthesis at elevated CO2 maybenefit smaller individuals growing in the shade (Os-borne et al. 1997).
To test these hypotheses, we established even-agedmonospecific stands of an annual, Chenopodium album,at ambient and doubled CO2 with high and low nutrientavailabilities in open top chambers (Nagashima et al.2003). The growth of individual plants was monitored
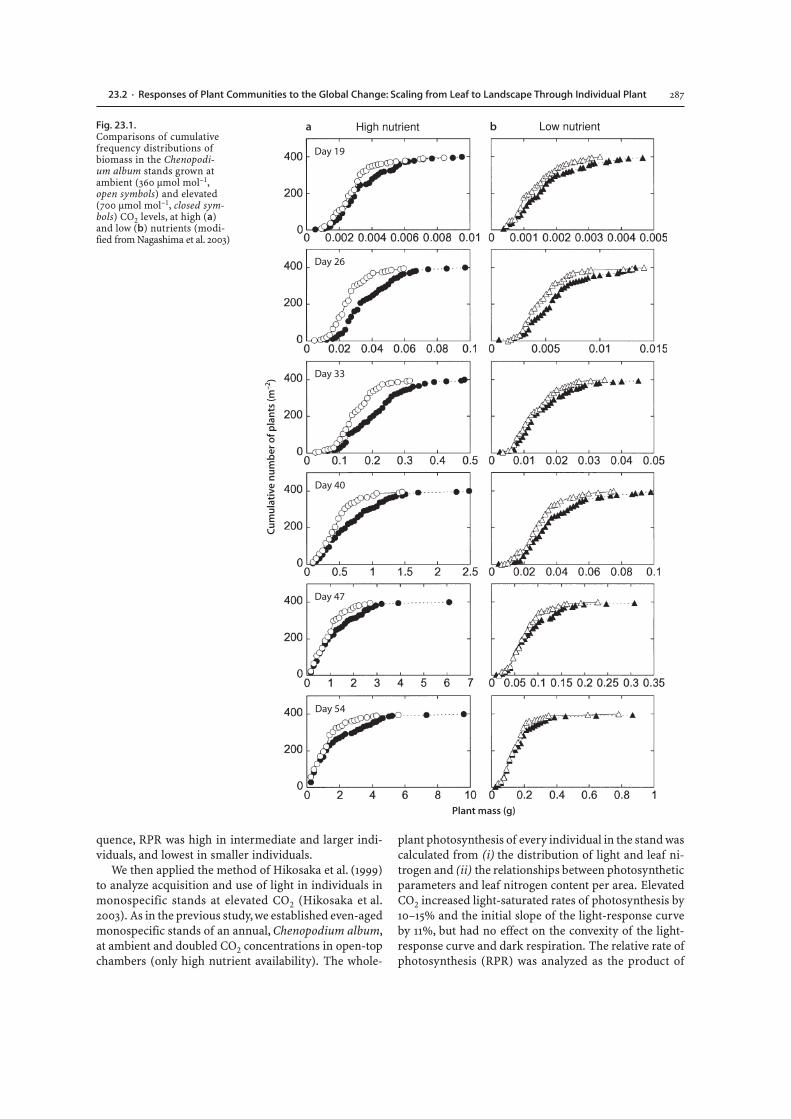
non-destructively every week until flowering. ElevatedCO2 significantly enhanced plant growth at high nutri-ents, but did not at low nutrients. The size inequality ex-pressed as the coefficient of variation tended to increaseat elevated CO2. Size structure of the stands was ana-lyzed by the cumulative frequency distribution of plantsize (Fig. 23.1). At early stages of plant growth, CO2 el-evation benefited all individuals and shifted the wholesize distribution of the stand to larger size classes. Atlater stages, dominant individuals were still larger at el-evated than at ambient CO2, but the difference in smallsubordinate individuals between two CO2 levels becamesmaller. Although these tendencies were found at bothnutrient availabilities, the difference in size distributionbetween CO2 levels was larger at high nutrients. The CO2elevation did not significantly enhance the growth rateas a function of plant size except for the high nutrientstand at the earliest stage. This indicates that the higherbiomass at elevated CO2 at later stages in the high nutri-ent stand was caused by the larger size of individuals atthe earliest stage. Therefore, the effects of elevated CO2on stand structure and size inequality are strongly de-pended on the growth stage and nutrient availability.
The emerging question is the ecophysiological basisof competition at elevated CO2. As mentioned above, thedifference in size structure results from different size-dependent growth rates of individuals within the stand.In a dense stand, large dominant individuals are betterat capturing light. Small subordinate individuals, on theother hand, need less investment for biomass in supporttissues because they maintain leaves at lower positions(Givnish 1982), and they can increase biomass allocationfor leaf expansion, and partly ameliorate the limited lightavailability (Anten and Hirose 1998). To indicate the ef-ficiency of biomass use to capture light, Hirose andWerger (1995) introduced the index Φmass, that is photonflux captured per unit aboveground mass. They suggestedthat Φmass might not differ between dominant and sub-ordinate species in multispecies systems. However, plantgrowth is determined not only by the amount of acquiredresources, but also by the efficiency of resource use(growth per unit amount of resource acquired). Hikosakaet al. (1999) defined light-use efficiency of photosynthe-sis (LUE) as photosynthesis per unit photon intercep-tion, and described the photosynthesis of individuals asthe product of Φmass and LUE: RPR = Φmass × LUE, whereRPR is the relative photosynthetic rate (photosyntheticrate per unit aboveground mass). Provided that plantgrowth is proportional to leaf photosynthesis, RPR isclosely related to the relative growth rate. With a modifi-cation of the canopy photosynthesis model of Hirose andWerger (1987), Hikosaka et al. (1999) estimated the pho-tosynthetic rate of individuals in a natural monospecificstand of an annual, Xanthium canadense. They foundthat Φmass was higher in larger individuals, while LUEwas the highest in intermediate individuals. As a conse-
287
quence, RPR was high in intermediate and larger indi-viduals, and lowest in smaller individuals.
We then applied the method of Hikosaka et al. (1999)to analyze acquisition and use of light in individuals inmonospecific stands at elevated CO2 (Hikosaka et al.2003). As in the previous study, we established even-agedmonospecific stands of an annual, Chenopodium album,at ambient and doubled CO2 concentrations in open-topchambers (only high nutrient availability). The whole-
plant photosynthesis of every individual in the stand wascalculated from (i) the distribution of light and leaf ni-trogen and (ii) the relationships between photosyntheticparameters and leaf nitrogen content per area. ElevatedCO2 increased light-saturated rates of photosynthesis by10–15% and the initial slope of the light-response curveby 11%, but had no effect on the convexity of the light-response curve and dark respiration. The relative rate ofphotosynthesis (RPR) was analyzed as the product of
Fig. 23.1.Comparisons of cumulativefrequency distributions ofbiomass in the Chenopodi-um album stands grown atambient (360 µmol mol–1,open symbols) and elevated(700 µmol mol–1, closed sym-bols) CO2 levels, at high (a)and low (b) nutrients (modi-fied from Nagashima et al. 2003)
23.2 · Responses of Plant Communities to the Global Change: Scaling from Leaf to Landscape Through Individual Plant
288 CHAPTER 23 · Terrestrial Ecosystems in Monsoon Asia: Scaling up from Shoot Module to Watershed
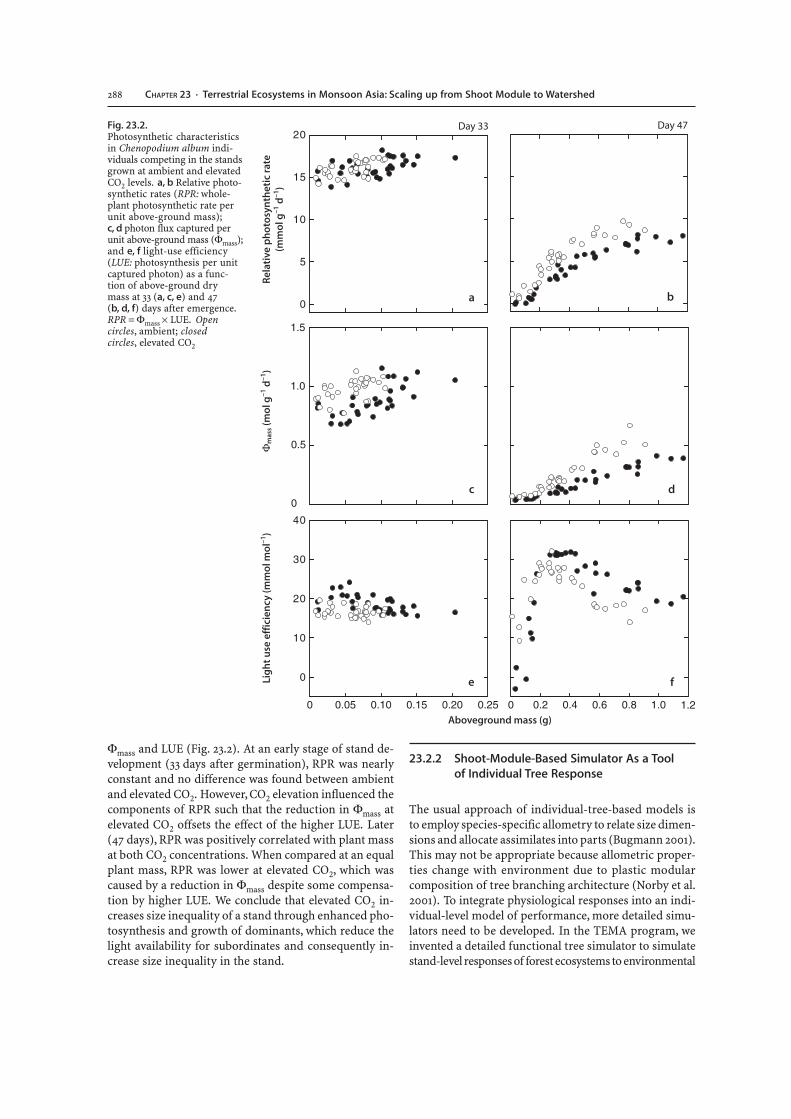
Φmass and LUE (Fig. 23.2). At an early stage of stand de-velopment (33 days after germination), RPR was nearlyconstant and no difference was found between ambientand elevated CO2. However, CO2 elevation influenced thecomponents of RPR such that the reduction in Φmass atelevated CO2 offsets the effect of the higher LUE. Later(47 days), RPR was positively correlated with plant massat both CO2 concentrations. When compared at an equalplant mass, RPR was lower at elevated CO2, which wascaused by a reduction in Φmass despite some compensa-tion by higher LUE. We conclude that elevated CO2 in-creases size inequality of a stand through enhanced pho-tosynthesis and growth of dominants, which reduce thelight availability for subordinates and consequently in-crease size inequality in the stand.
23.2.2 Shoot-Module-Based Simulator As a Toolof Individual Tree Response
The usual approach of individual-tree-based models isto employ species-specific allometry to relate size dimen-sions and allocate assimilates into parts (Bugmann 2001).This may not be appropriate because allometric proper-ties change with environment due to plastic modularcomposition of tree branching architecture (Norby et al.2001). To integrate physiological responses into an indi-vidual-level model of performance, more detailed simu-lators need to be developed. In the TEMA program, weinvented a detailed functional tree simulator to simulatestand-level responses of forest ecosystems to environmental
Fig. 23.2.Photosynthetic characteristicsin Chenopodium album indi-viduals competing in the standsgrown at ambient and elevatedCO2 levels. a, b Relative photo-synthetic rates (RPR: whole-plant photosynthetic rate perunit above-ground mass);c, d photon flux captured perunit above-ground mass (Φmass);and e, f light-use efficiency(LUE: photosynthesis per unitcaptured photon) as a func-tion of above-ground drymass at 33 (a, c, e) and 47(b, d, f) days after emergence.RPR = Φmass × LUE. Opencircles, ambient; closedcircles, elevated CO2

289
change. This simulator, named PipeTree, describes current-year shoot growth in three-dimensional space (Kubo andKohyama 2005). Light conditions that regulate photosyn-thesis are calculated at each shoot module by monitor-ing obstruction with in a hemispherical space. Alloca-tion of assimilates into three-dimensional plant archi-tecture is constrained by current-year shoot morphol-ogy including leaf/stem ratio, branching rules, and a so-called pipe-model relationship that sap wood area at anyposition of a stem is proportional to the cumulative leafarea above that position (Shinozaki et al. 1964).
Using observed data of a monospecific quasi-even-aged natural stand of subalpine fir (Abies veitchii) popu-lation in a subalpine forest of central Japan, Kubo andKohyama (2005) revised the model structure and tunedphysiological and morphological parameters. Figure 23.3demonstrates a three-dimensional view of a fir standsimulated by PipeTree. The model quantitatively repro-duced phenomena of amplified size hierarchy, self-thin-ning, and allometry shift with stand age. The model alsoshowed that allometry shifts towards slenderer tree formsunder elevated atmospheric CO2.
Physiological-process-based detailed models likePipeTree can provide reasonable parameterization, and/or functional responses of parameters with environmen-tal change, for coarse-scaled individual-based and othermodels of forest ecosystem dynamics.
23.2.3 Modeling the Shift of Forest Zonation
Available simulation tools of vegetation dynamics atlarger scales up to the Earth do not explicitly examinethe limit of seed dispersal, which is an essential featureof plants as sessile living organisms. The decline of spe-cies diversity is related to the dispersal limit coupled withhabitat fragmentation; meanwhile the spread of alienspecies worldwide is caused by artificial dispersal.
Therefore, reasonable models need to incorporate dis-persal process.
In the TEMA program, we developed two models withdispersal processes to simulate vegetation shifts with glo-bal warming. Kohyama and Shigesada (1995) and Kohyama(2005) extended a tree-size-structure-based model intoa geographic landscape. In the model, a tree speciespopulation is distributed in geographic space (distribu-tion range) and at each location (size distribution andpatch age structure). Dispersal capacity is expressed bya coefficient of dispersal at the smallest size. The dis-persal coefficient (with units of squared distance) is simi-lar to a diffusion coefficient but is defined not for a unittime but for a unit seeding event. When run for a sce-nario of a century-long warming followed by fixed warmclimate, the model projected that extant forests would re-spond to warming without remarkable time lags by chang-ing size structure and biomass. However for the bound-aries between forest zones there were extreme time lags. Ittook thousands of years to attain new steady-state bound-aries, and a tenfold increase of dispersal coefficients in-creased the movement of boundaries less than two-fold.This delay of boundary shifts was brought about by the‘mass effect’ of existent forests (adapted to cooler condi-tions) that prevented the invasion of more-adapted forestsin warmer conditions. Because species have broader po-tential niches than realized niches, and because establishedtrees are long lived, such competitive depression of bound-ary shifts by resident forest zones resulted. If there wereno neighbor forest types, a forest species could readily shiftits distribution without an obvious time lag.
Takenaka (2005) proposed another type of vegetationdispersal simulator, based on a lattice model where onecell of the lattice corresponds to an individual tree. Dis-persal probability is expressed there by an exponentialfunction from the mother-tree cell. Establishment occursonly when seeds are dispersed to vacant cells, or canopygaps. The model similarly simulated the millennial delayof forest boundary shifts. It also suggested that speciesdiversity declines at the range of forest type shift due torandom drift, and one species that happened to estab-lish in a new site eventually dominated the landscape.
The framework of these models needs to be incorpo-rated into dynamic global vegetation models (DGVMs)to predict changes in biome zones, or the failure of adap-tive distribution of biomes, in response to global change.
23.3 Carbon Budget at the Forest Watershed Scale
Carbon biogeochemistry in forest watersheds consistsof photosynthesis, autotrophic and heterotrophic res-piration, changes in biomass above and below theground, litterfall, decomposition of organic matter,changes in the storage of soil organic matter, and leach-ing of dissolved and particulate carbon to groundwater
Fig. 23.3. A 44 year old stand of Japanese subalpine fir forest, simu-lated by PipeTree. Tree individuals are explicitly described with apopulation of shoot modules with three-dimensional locations
23.3 · Carbon Budget at the Forest Watershed Scale

290 CHAPTER 23 · Terrestrial Ecosystems in Monsoon Asia: Scaling up from Shoot Module to Watershed
and streams (Fig. 23.4). Here, we focus on current re-sults and a synthesis of the biogeochemical processesof carbon in forest watersheds based mainly on workconducted during TEMA.
23.3.1 Carbon Exchange betweenAtmosphere-Forest-Stream Boundaries
Eddy-correlation techniques for assessing CO2 flux in theforest canopy provide quantitative information on netphotosynthesis and respiration (for both vegetation andmicroorganisms), also referred to as net ecosystem ex-change (NEE). NEE is a critical index for the quantita-tive assessment of the extent to which atmospheric car-bon dioxide has been sequestrated by the ecosystem. Inthe Horonai River basin located in northern Japan, theNEE flux observed using the eddy correlation method(1999–2001) was 258 g C m–2 yr–1 (SD = 36), with atmo-spheric carbon dioxide sequestrated mainly during thegrowing season from June to September (Tanaka andTanaka 2002; Shibata et al. 2005). Yamamoto et al. (1999)reported that NEE flux ranged from 65 to 136 g C m–2 yr–1
(mean = 106) in a temperate forest located in central Ja-pan, which suggests that the inter-annual variation inregional climate is reflected in the inter-annual variationin NEE flux. Carbon exchange between forests and theatmosphere is driven by biotic factors such as photosyn-thesis, respiration, and nutrient conditions, and abioticfactors like solar radiation, temperature, air stability andothers. While the expected increase in anthropogenicnitrogen pollution in the Asian region over the next fewdecades (Galloway and Cowling 2002) is likely to affectthe future dynamics of NEE in this region, the effect ofnitrogen deposition on carbon sequestration is still un-der discussion (Lloyd 1999; Nadelhoffer et al. 1999).
In the NEE budget analyses, increases in above-groundbiomass and leaching in streams are useful measures thatcan be employed to better understand the role of vegeta-tion and soil in the carbon sequestration from a quanti-tative perspective. Since the budget for NEE (input) andleaching in streams (output) is indicative of the net stor-age or release of carbon (so called net ecosystem pro-duction (NEP)) in the watershed, respectively, the differ-ence between NEP and the carbon accumulated in thevegetation corresponds to the net carbon storage (or re-lease) for the soil system. In the Horonai River basin, theannual rate of increase of carbon in vegetation, the ex-tent of leaching to streams, and the net carbon storagein the soil were 108, 4 and 146 g C m–2 yr–1, respectively(Table 23.1, Hiura 2005; Shibata et al. 2001; Shibata et al.2005). Furthermore, it was found that 41% and 57% ofthe NEE were retained in vegetation and soil, respectively.Since the mean residence time of carbon is markedly dif-ferent for vegetation and soil, the relative partitioning ofsequestrated carbon in the ecosystem is a very importantconsideration when predicting the effect of carbon dynam-ics on environmental change. However, the cumulative ef-fect of possible sampling errors means that budget analy-ses are inherently difficult. These include: (i) difficulties inmeasuring eddies during periods of high atmospheric sta-bility and the irregularity of the canopy surface, (ii) thelarge spatial heterogeneity in the vegetation of the water-shed, (iii) the large temporal and spatial variation in resi-dence times associated with the leaching of carbon intostreams. Consequently, indirect or direct estimates of thecarbon contributions from above and below-ground bio-mass, combined with the quantification of fine root pro-duction and mortality, will be one of the most effectivemeans of ameliorating the effect of the aforementioneduncertainties associated with assaying carbon storage inthe ground (Shutou and Nakane 2004; Satomura et al. 2003).
23.3.2 Transport of Dissolved Organic CarbonAssociated with Dissolved Nitrogenfrom Terrestrial to Aquatic Ecosystems
Although the annual flux of dissolved carbon into streamswas generally very small when compared with that asso-ciated with gas exchange between the terrestrial ecosys-
Fig. 23.4. Compartment model of carbon flow in a forested wa-tershed
291
tem and the atmosphere, the supply of dissolved organiccarbon to lotic and lentic ecosystems is an importantsource of energy for aquatic biota and as a source of sec-ondary emissions of carbon dioxide from water surfaces.The quantitative and qualitative fate of dissolved organiccarbon was different for each watershed. This is due tothe heterogeneity expressed in terms of microbiologicalactivities, reaction to the solid phase, hydrological flow-pass in the ground, and the interrelationship betweencarbon and nitrogen cycling in forest ecosystems (Shibataet al. 2001; Neff and Asner 2001; Katsuyama and Ohte2002; Konohira and Yoshioka 2005). Consequently, theflux and concentration of dissolved organic carbon instream water can be a useful biogeochemical signatureof carbon cycling in catchment ecosystems. Nitrate (NO3
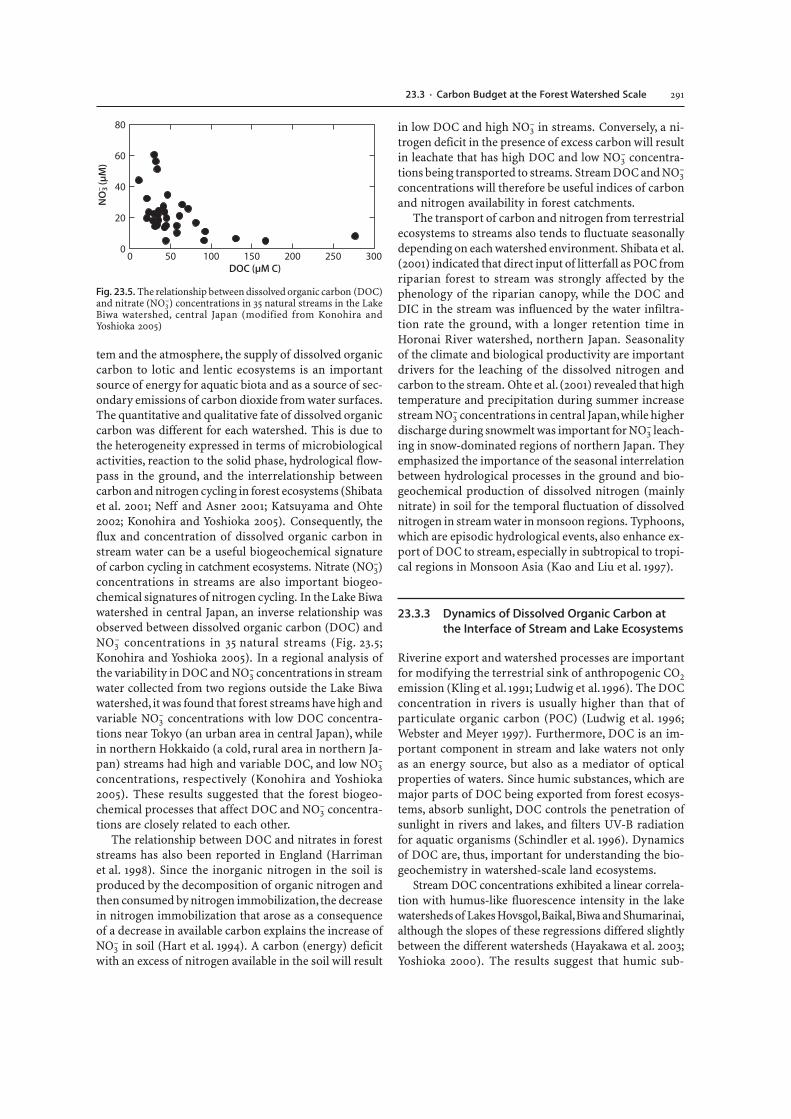
–)concentrations in streams are also important biogeo-chemical signatures of nitrogen cycling. In the Lake Biwawatershed in central Japan, an inverse relationship wasobserved between dissolved organic carbon (DOC) andNO3
– concentrations in 35 natural streams (Fig. 23.5;Konohira and Yoshioka 2005). In a regional analysis ofthe variability in DOC and NO3
– concentrations in streamwater collected from two regions outside the Lake Biwawatershed, it was found that forest streams have high andvariable NO3
– concentrations with low DOC concentra-tions near Tokyo (an urban area in central Japan), whilein northern Hokkaido (a cold, rural area in northern Ja-pan) streams had high and variable DOC, and low NO3
–
concentrations, respectively (Konohira and Yoshioka2005). These results suggested that the forest biogeo-chemical processes that affect DOC and NO3
– concentra-tions are closely related to each other.
The relationship between DOC and nitrates in foreststreams has also been reported in England (Harrimanet al. 1998). Since the inorganic nitrogen in the soil isproduced by the decomposition of organic nitrogen andthen consumed by nitrogen immobilization, the decreasein nitrogen immobilization that arose as a consequenceof a decrease in available carbon explains the increase ofNO3
– in soil (Hart et al. 1994). A carbon (energy) deficitwith an excess of nitrogen available in the soil will result
in low DOC and high NO3– in streams. Conversely, a ni-
trogen deficit in the presence of excess carbon will resultin leachate that has high DOC and low NO3
– concentra-tions being transported to streams. Stream DOC and NO3
–
concentrations will therefore be useful indices of carbonand nitrogen availability in forest catchments.
The transport of carbon and nitrogen from terrestrialecosystems to streams also tends to fluctuate seasonallydepending on each watershed environment. Shibata et al.(2001) indicated that direct input of litterfall as POC fromriparian forest to stream was strongly affected by thephenology of the riparian canopy, while the DOC andDIC in the stream was influenced by the water infiltra-tion rate the ground, with a longer retention time inHoronai River watershed, northern Japan. Seasonalityof the climate and biological productivity are importantdrivers for the leaching of the dissolved nitrogen andcarbon to the stream. Ohte et al. (2001) revealed that hightemperature and precipitation during summer increasestream NO3
– concentrations in central Japan, while higherdischarge during snowmelt was important for NO3
– leach-ing in snow-dominated regions of northern Japan. Theyemphasized the importance of the seasonal interrelationbetween hydrological processes in the ground and bio-geochemical production of dissolved nitrogen (mainlynitrate) in soil for the temporal fluctuation of dissolvednitrogen in stream water in monsoon regions. Typhoons,which are episodic hydrological events, also enhance ex-port of DOC to stream, especially in subtropical to tropi-cal regions in Monsoon Asia (Kao and Liu et al. 1997).
23.3.3 Dynamics of Dissolved Organic Carbon atthe Interface of Stream and Lake Ecosystems
Riverine export and watershed processes are importantfor modifying the terrestrial sink of anthropogenic CO2emission (Kling et al. 1991; Ludwig et al. 1996). The DOCconcentration in rivers is usually higher than that ofparticulate organic carbon (POC) (Ludwig et al. 1996;Webster and Meyer 1997). Furthermore, DOC is an im-portant component in stream and lake waters not onlyas an energy source, but also as a mediator of opticalproperties of waters. Since humic substances, which aremajor parts of DOC being exported from forest ecosys-tems, absorb sunlight, DOC controls the penetration ofsunlight in rivers and lakes, and filters UV-B radiationfor aquatic organisms (Schindler et al. 1996). Dynamicsof DOC are, thus, important for understanding the bio-geochemistry in watershed-scale land ecosystems.
Stream DOC concentrations exhibited a linear correla-tion with humus-like fluorescence intensity in the lakewatersheds of Lakes Hovsgol, Baikal, Biwa and Shumarinai,although the slopes of these regressions differed slightlybetween the different watersheds (Hayakawa et al. 2003;Yoshioka 2000). The results suggest that humic sub-
Fig. 23.5. The relationship between dissolved organic carbon (DOC)and nitrate (NO3
–) concentrations in 35 natural streams in the LakeBiwa watershed, central Japan (modified from Konohira andYoshioka 2005)
23.3 · Carbon Budget at the Forest Watershed Scale

292 CHAPTER 23 · Terrestrial Ecosystems in Monsoon Asia: Scaling up from Shoot Module to Watershed
stances were predominant in stream DOC, and the chemi-cal composition of the stream dissolved organic matterwas relatively homogeneous among watersheds. How-ever, such a correlation was not found in the pelagic ar-eas of these lakes. The surface DOC concentrations ofLake Biwa mainly fluctuated during the period at whichstratification occurred (from March to November), witha concomitant gradual decrease being observed in thehumus-like fluorescence intensity. These findings indi-cate the production of autochthonous DOC and that thehumic substances were derived from the watershed (al-lochthonous) and subsequently decomposed in the lake.The increase in the protein-like fluorescence observedin the surface layers of Lake Biwa also suggested the auto-chthonous production of dissolved organic matter(Mostofa et al. in review). Since humic substances arehighly resistant to biological degradation, the change inthe humus-like fluorescence intensity observed in the lakemay instead be attributed to photodegradation. Thechange in the position of the peak humus-like fluores-cence, determined using a three-dimensional excitationemission matrix, suggested that the alteration of the hu-mic substances could be attributed to solar irradiation(Hayakawa et al. 2003; Mostofa et al. in review).
23.4 Carbon Budget and Functions of the Lake BiwaEcosystem
To understand key processes determining the functionof aquatic habitats in terrestrial carbon balance, we mea-sured and analyzed the budget and the fate of C in a lakeecosystem in the TEMA program. We chose Lake Biwafor this model study. Lake Biwa is the largest lake in Ja-pan and one of the ancient lakes, with a surface area of674 km2, a mean depth of 41 m, a watershed of 3 848 km2
and a water retention time of ca. 5.5 years. The lake wasoriginally oligotrophic. However, due to development inthe watershed for the past 40 years, the lake has become
moderately eutrophic. At present, the lake is mesotrophicand supplies water resources for 14 million peoples liv-ing in the Kinki District of Japan. The following esti-mates were made in the north basin, which accounts for99% of the water volume of this lake.
23.4.1 Carbon Budget in the Lake
The concentration of particulate organic C (POC) in LakeBiwa ranged from 0.2 to 1.1 g C m–3 in the surface mix-ing layer (0–15 m deep), but was less than 0.3 g C m–3 inthe hypolimnion (>15–20 m deep). The pattern coincidedwith seasonal and vertical changes in phytoplanktonabundance (Yoshida et al. 2001). Similarly, the concen-tration of dissolved organic C (DOC) was high in sum-mer in the surface layer (1.4 g C m–3) but became verti-cally homogeneous in winter (~1.0 g C m–3). Based onmonitoring data, the annual input of allochthonous or-ganic C (POC + DOC) per unit area of Lake Biwa was9.6 g C m–2 yr–1. The net primary production rate inthe surface layer in the Lake Biwa ranged from 0.3 to2.7 g C m–2 d–1 and algae fixed 323 g C m–2 annually(Urabe et al.1999; Yoshimizu et al. 2001). Thus, auto-chthonous input of organic C is 30 times higher thanthe allochthonous input (Fig. 23.6). Nonetheless, theshare of allochthonous C was <40% of DOC in thesurface mixing layer and 50–60% in the hypolimnion(Yoshioka 2000), implying that allochthonous or-ganic C is less respired compared with autochthonousorganic C. Interestingly, organic C corresponding to 90%of the allochthonous input flowed out annually from thenorth basin of Lake Biwa (Yoshimizu et al. 2002). Sincemost organic C from the inflow rivers was humicsubstance and since its concentration was seasonallystable in the lake water (Yoshioka 2000), a substantialfraction of allochthonous organic matter seemed toflow out from the lake without being consumed effi-ciently within the lake.
Fig. 23.6.Carbon budget at an offshoresite of Lake Biwa

293
Of the organic C in the surface water, 78 g C m–2 yr–1
was transported annually to the profundal layer(Yoshimizu et al. 2001), implying that 75% of theorganic C was respired within the surface layer. From theage of lake sediments determined by 210Pb and 137Cs andthe abundance of organic C at the lake bottom (Muraseand Sakamoto 2000; Urabe et al. 2005), the annual rateof organic C burial at the lake bottom was estimated tobe 7–28 g C m–2 yr–1. This implies that 64–90% of theorganic C exported from the surface layer is respired ordecomposed at the profundal layer and/or the lake sedi-ments. This number is smaller than burial rates in lakeswith surface areas < 500 km2 but larger than those inlarger lakes such as Lake Micigana and Lake Bikal (e.g.,Dean and Gorham 1998). In addition, it is in accordancewith the oxygen consumption rate (~180 g O2 m–2) dur-ing the stagnant period in the hypolimnion of the LakeBiwa (Kumagai and Fushimi 1995). According to itsδ15N signature, 40% of organic matter in the lake sedi-ments was estimated to be terrestrial in origin (Muraseand Sakamoto 2000). This suggests that allochthonousorganic matter is selectively stored in the lake sedimentsdue to less efficient utilization by living organisms.
These estimates show that the P/R ratio of Lake Biwais 1.02~1.09, implying that C fixation slightly exceeds CO2production. This is because the allochthonous organicmatter is much less discharged compared with primaryproduction and less efficiently utilized by organisms.Note that the present C budgets did not consider hori-zontal transports of organic matters within the lake. Inthis lake, pCO2 at the surface water was less than satura-tion level from spring to fall, but its annual mean was397 ppmv because CO2 in the water was supersaturatedin winter (Urabe et al. 2005). Thus, as a whole, the LakeBiwa ecosystem functions as a C source. Indeed, thereare estimates that the community respiration rate of het-erotrophs exceeds the net primary production at an off-shore site. The slight discrepancy between the C budgetsof the biological community and the pCO2 observationsmay be due to decomposition of organic matter producedat the littoral area.
23.4.2 Metabolism in the Lake Sediments
The lake sediments are characterized by the concentratedaccumulation of organic carbon. Standing stocks of or-ganic carbon in the upper most layer (2 cm) of the sedi-ment were estimated to be 9.2 × 1010 g C for the wholelake (Murase and Sakamoto 2000). This value is 2.5 timeshigher than the estimated amount of carbon in the wholelake water (3.6 × 1010 g C). The littoral sediments and thedeeper layer of the profundal sediments showed highercontribution of terrestrial organic matter than the sur-face layer of the profundal sediments (Murase andSakamoto 2000).
Except for the top few millimeters, the bottom sedi-ments of the lake are anoxic and methanogenesis is themajor terminal carbon metabolism. Potential methaneproduction rates of the profundal sediment (0–25 cm indepth) ranged from 2.4–24 mg C m–2 d–1 (Murase andSugimoto 2002). Carbon isotopic data for methane andCO2 in the sediment suggested that hydrogenotrophicmethanogenesis was the major pathway (Murase andSugimoto 2001). A part of the produced methane wasstored in adsorbed form in the lake sediment (Sugimotoet al. 2003), but some was diffused up to the surface, where80–90% of diffused methane was oxidized (Murase et al.2005). The methane oxidation rates at the sediment sur-face are comparable to the rate of oxic carbon mineral-ization (17–20 mg C m–2 d–1) and these rates accountedfor 50–160% of the estimated carbon sedimentation rate(Murase, unpublished). Our estimates suggested that theincrease in oxygen consumption rate by 50% potentiallymakes the surface sediment anoxic causing the cata-strophic change in gas metabolism of the sediment. Inaddition to eutrophication, an increase in temperatureof the bottom water due to the global warming (Hayamiand Fujiwara 1999), which caused enhancement of mi-crobial activities, prolonged stagnant periods, and lowersolubility of oxygen, would accelerate the oxygen deple-tion in the sediment.
The potential methane production rate in the littoralsediments was approximately 5 to 10 times higher thanthat in the profundal sediments (Murase and Sugimoto2002). At the littoral area, substantial methane diffusedfrom the sediment across the oxic-anoxic boundary intothe lake water during the stagnant period (Murase et al.2003). Disturbance of the surface sediment by internalwaves of the lake seemed to accentuate transportation ofmethane from the sediment to the lake (Sakai et al. 2002),which can be major sources of dissolved methane in thewhole lake water (Murase et al. 2003).
23.4.3 Terrestrial Environment and Functionof Lake Ecosystems
Considering the watershed and lake surface areas, thedischarge rate of allochthonous organic carbon to, andcarbon burial rate in, lake Biwa are two orders of magni-tude lower than the NEE in the temperate forests in Ja-pan, but almost comparative to the outflow rate of or-ganic carbon to streams from the forests (Sect. 23.3.1 andTable 23.1). Thus, standing water is a potentially impor-tant arena determining the fate of organic carbon leach-ing to streams from the forests. This suggests that evenslight changes in the carbon budget in the forests mayhave serious impacts on lake ecosystems.
According to del Giorgio et al. (1994), the P/R ratio oflake ecosystems becomes > 1 with increasing the trophiccondition. This implies that the function of lake ecosys-
23.4 · Carbon Budget and Functions of the Lake Biwa Ecosystem

294 CHAPTER 23 · Terrestrial Ecosystems in Monsoon Asia: Scaling up from Shoot Module to Watershed
tems is changed from C sink to C source with increasingnutrient loading from the watershed. In Lake Biwa, theprimary production rate is nutrient limited (Urabe et al.1999). Therefore, increase in nutrient loadings wouldincrease C sequestration from the atmosphere throughincreased C fixation rate and push this lake toward aC sink. Progress of further eutrophication due to anthro-pogenic activities in the watershed, however, may turnthe lake ecosystem to vent gasses that are more efficientfor warming (Fig. 23.7). An increase in the sinking fluxof organic C due to increased primary production pro-motes oxygen consumption in the profundal layer and lakesediments. Global warming would accelerate the oxygenconsumption at the lake bottom by increasing water tem-perature at the profundal zone and reducing verticalmixing in winter. This may, in turn, promote the emis-sion of methane, a more efficient gas for global warmingthan CO2, from the lake sediments to the atmosphere.
As such, it is most likely that the function of lake eco-systems in terms of being a C sink or a C source dependson the land cover and land use, and thus the anthropo-genic activities, in the watershed. Unfortunately, only afew studies have included aquatic habitats when carbonbalance of terrestrial ecosystems is examined in Asianarea (Kao and Liu 1997; Ludwig et al. 1996; Sarin et al.2002; Shibata et al. 2001). Considering future anthropo-genic development in Monsoon Asia and the potentialimportance of inland aquatic systems as a hot spot whereterrigenous carbon is processed, studies at a watershedscale are essential to assess impacts of changes in car-bon budgets in terrestrial ecosystems under elevated CO2and climate change.
23.5 Summary and Conclusions
A GCTE core project “Global change impacts on terres-trial ecosystems in Monsoon Asia” (TEMA) has been car-ried out from 1995 to 2003. This chapter overviews theTEMA-employed unique approach of integrating acrossdifferent scales, i.e., from a plant leaf to watershed bud-gets, targeting on the eastern Asian region. We particu-larly focused on the linkage between physiological pro-cesses of foliage canopy and landscape-scale processes
of plant demography and plant community dynamics,where individual plant processes were integrated fromphysiology, and we projected the change in geographicpattern from individual plant processes. We evaluatedthe watershed unit where freshwater chemistry providesa signature of biogeochemical characteristics of terres-trial ecosystems. Stream chemistry controls the trophiccondition of lake ecosystems, which can contribute toglobal change particularly through methane emission.Integration at the scale of watersheds will contribute,within the scope of the new GLP in relation to LOICZ, tothe validation of the impact of environmental change onhuman society, and the impact of human activities onwatershed-scale environments.
References
Anten NPR, Hirose T (1998) Biomass allocation and light partition-ing among dominant and subordinate individuals in Xanthiumcanadense stands. Ann Bot 82:665–673
Bugmann HKM (2001) A review of forest gap models. ClimaticChange 51:259–305
Carignan RD, Planas, Vis C (2000) Planktonic production and respi-ration in oligotrophic shield lakes. Limnol Oceanogr 45:189–199
Cole JJ, Caraco NF, Kling GW, Kratz TK (1994) Carbon dioxide super-saturation in the surface waters of lakes. Science 265:1568–1570
Dean WE, Gorham E (1998) Magnitude and significance of carbonburial in lakes, reservoir, and peatlands. Geology 26:535–538
del Giorgio, Paul A, Peters RH (1994) Patterns in planktonic P:Rratio in lakes: influence of lake trophy and dissolved organiccarbon. Limnol Oceanogr 39:772–787
Ford ED, Diggle PJ (1981) Competition for light in a plant monoc-ulture modelled as a spatial stochastic process. Ann Bot 48:481–500
Galloway JN, Cowling EB (2002) Reactive nitrogen and the world:200 years of Change. Ambio 31:64–71
Givnish TJ (1982) On the adaptive significance of leaf height in for-est herbs. Am Nat 120:353–381
Gurung TB, Urabe J (1999) Temporal and Vertical difference in fac-tors limiting growth rate of heterotrophic bacteria in Lake Biwa.Microb Ecol 38:136–145
Hanson PC, Pollard A, Bade DL, Predick K, Carpenter SR, Foley J(2004) A model of carbon evasion and sedimentation in tem-perate lakes. Global Change Biology 10:1285–1298
Harriman R, Curtis C, Edwards AC (1998) An empirical approachfor assessing the relationship between nitrogen deposition andnitrate leaching from upland catchments in The United King-dom using runoff chemistry. Water Air Soil Pollut 105:193–203
Hart SC, Nason GE, Myrold DD, Perry DA (1994) Dynamics of grossnitrogen transformation in an old-growth forest: The carbonconnection. Ecology 75:880–891
Fig. 23.7.Changes in lake ecosystemfunctions in terms of C sinks/sources as a function of in-creases in nutrient loadings.Auto-C: autochthonous or-ganic carbon, and Allo-C: allo-chthonous organic carbon

295
Hayakawa K, Sekino T, Yoshioka T, Maruo M, Kumagai M (2003) Dis-solved organic carbon and fluorescence in Lake Hovsgol: factorsreducing humic content of the lake water. Limnology 4:25–33
Hayami Y, Fujiwara T (1999) Warming of hypolimnetic water in LakeBiwa. Oceanography in Japan 8:197–202 (in Japanese)
Hikosaka K, Hirose T (2001) Nitrogen uptake and use of competingindividuals in a Xanthium canadense stand. Oecologia 126:174–181
Hikosaka K, Sudoh S, Hirose T (1999) Light acquisition and use byindividuals competing in a dense stand of an annual herb,Xanthium canadense. Oecologia 118:388–396
Hikosaka K, Yamano T, Nagashima H, Hirose T (2003) Light-acqui-sition and use of individuals as influenced by elevated CO2 ineven-aged monospecific stands of Chenopodium album. FunctEcol 17:786–795
Hirose T, Walker B (ed) (1996) Global change and terrestrial eco-systems in monsoon Asia. Vegetatio 121:1–191
Hirose T, Werger MJA (1987) Maximizing daily canopy photosyn-thesis with respect to the leaf nitrogen allocation pattern in thecanopy. Oecologia 72:520–526
Hirose T, Werger MJA (1995) Canopy structure and photon flux par-titioning among species in a herbaceous plant community. Ecol-ogy 76:466–474
Hirose T, Kohyama T, Oshima Y (1998) GCTE activities in Japan.Global Environmental Research 1:19–24
Hiura T (2005) Estimation of aboveground biomass and net biom-ass increment in a cool temperate forest on a landscape scale.Ecol Res 20:271–277
Jones HG (1992) Plants and microclimate, 2nd edn. Cambridge Uni-versity Press, Cambridge
Jurik TW (1991) Population distribution of plant size and light en-vironment of giant ragweed (Ambrosia trifida L.). Oecologia87:539–550
Kao SJ, Liu KK (1997) Fluxes of dissolved and nonfossil particulateorganic carbon from an Oceania small river (Lanyang Hsi) inTaiwan. Biogeochemistry 39:255–269
Katsuyama M, Ohte N (2002) Determining the sources of stormflowfrom the fluorescence properties of dissolved organic carbon ina forested headwater catchment. J Hydrol 268:192–202
Kling GW, Kipphut GW, Miller MC (1991) Arctic lakes and streamsas gas conduits to the atmosphere: Implications for tundra car-bon budgets. Science 251:298–301
Kohyama T (2005) Scaling up from shifting gap mosaic to geo-graphic distribution in the modeling of forest dynamics. EcolRes 20:305–312
Kohyama T, Shigesada N (1995) A size-distribution-based model offorest dynamics along a latitudinal gradient. Vegetatio 121:117–126
Kohyama T, Canadell J, Ojima DS, Pitelka L (ed) (2005) Forest eco-systems and environments: scaling up from shoot module towatershed. Ecol Res 20:241–385
Konohira E, Yoshioka T (2005) Dissolved organic carbon and ni-trate concentrations in stream. A useful index indicating car-bon and nitrogen availability in catchments – Ecological Re-search 20:359–365
Kubo T, Kohyama T (2005) Abies population dynamics simulatedby a functional-structural tree model. Ecol Res 20:255–269
Kumagai M, Fushimi H (1995) Inflows due to snowmelt: Physicalprocesses in a lakrge lake, Lake Biwa, Japan. Coastal and Estua-rine Studies 48:129–139
Lloyd J (1999) The CO2 dependence of photosynthesis, plant growthresponses to elevated CO2 concentrations and their interactionwith soil nutrient status, II. Temperate and boreal forest pro-ductivity and the combined effects of increasing CO2 concen-trations and increased nitrogen deposition at a global scale.Funct Ecol 13:439–759
Ludwig W, Probst JL, Kempe S (1996) Predicting the oceanic inputof organic carbon by continental erosion. Global BiogeochemCycles 1:23–41
Mostofa KMG, Yoshioka T, Konohira E, Tanoue E, Hayakawa K, Ta-kahashi M (2005) Three-dimensional fluorescence as a tool forinvestigating the dynamics of dissolved organic matter in theLake Biwa watershed. Limnology 6:101–115
Murase J, Sakamoto M (2000) Horizontal distribution of carbonand nitrogen and their isotopic compositions in the surface sedi-ment of Lake Biwa. Limnology 1:177–184
Murase J, Sugimoto A (2001) Spatial distribution of methane in theLake Biwa sediments and its carbon isotopic compositions.Geochemical J 35:257–263
Murase J, Sugimoto A (2002) Seasonal and spatial variations ofmethane production in mesotrophic lake sediments (Lake Biwa,Japan) Verh Int Ver Limnol 28:971–974
Murase J, Sakai Y, Sugimoto A, Okubo K, Sakamoto M (2003) Sourcesof dissolved methane in Lake Biwa. Limnology 4:91–99
Murase J, Sakai Y, Kametani A, Sugimoto A (2005) Dynamics ofmethane in a mesotrophic Lake Biwa, Japan. Ecol Res 20:377–385
Nadelhoffer KJ, Emmett BA, Gundersen P, Kjonaas OJ, KoopmansCJ, Schleppi P, Tietema A, Wright RF (1999) Nitrogen depositionmakes a minor contribution to carbon sequestration in temper-ate forests. Nature 398:145–148
Nagashima H (1999) The processes of height-rank determinationamong individuals and neighborhood effects in Chenopodiumalbum L. stands. Ann Bot 83:501–507
Nagashima H, Yamano T, Hikosaka K, Hirose T (2003) Effects ofelevated CO2 on the size structure of even-aged monospecificstands of annual, Chenopodium album. Global Change Biol 9:619–629
Nakashizuka T, Kohyama T, Whitmore TC, Ashton PS (ed) (1999)Tree diversity and dynamics of western Pacific and eastern Asianforests. J Veg Sci 10:763–860
Neff JC, Asner GP (2001) Dissolved organic carbon in terrestrialecosystems: synthesis and a model. Ecosystems 4:29–48
Norby RJ, Ogle K, Curtis PS, Badeck F-W, Huth A, Hurtt GC, KohyamaT, Penuelas J (2001) Aboveground growth and competition inforest gap models: an analysis for studies of climatic change.Climatic Change 51:415–447
Ohte N, Tokuchi N, Shibata H, Tsujimura M, Tanaka T, Mitchell MJ(2001) Hydrobiogeochemistry of Forest ecosystems in Japan.Hydrol Process 15:1771–1789
Osborne CP, Drake BG, LaRoche J, Long SP (1997) Does long-termelevation of CO2 concentration increase photosynthesis in for-est floor vegetation? Indian strawberry in a Maryland forest.Plant Physiol 114:337–344
Sakai Y, Murase J, Sugimoto A, Okubo K, Nakayama E (2002) Resus-pension of bottom sediment by an internal wave in Lake Biwa.Lakes and Reservoirs 7:339–344
Sarin MM, Sudheer AK, Balakrishna K (2002) Significance of river-ine carbon transport: A case study of a large tropical river,Godavari (India). Science in China (Series C) 45 supplement:97–108
Satomura T, Nakatsubo T, Horikoshi T (2003) Estimation of thebiomass of fine roots and mycorrhizal fungi: a case study ina Japanese red pine (Pinus densiflora) stand. J Forest Res 8:221–225
Schindler DW, Curtis PJ, Parker BR, Stainton MP (1996) Conse-quences of climate warming and lake acidification for UV-Bpenetration in North American boreal lakes. Nature 379:705–708
Shibata H, Mitsuhashi H, Miyake Y, Nakano S (2001) Dissolved andparticulate carbon dynamics in a cool-temperate forested basinin northern Japan. Hydrol Process 15:1817–1828
Shibata H, Hiura T, Tanaka Y, Takagi K, Koike T (2005) Carbon cy-cling and budget in a forested basin of southwestern Hokkaido,northern Japan. Ecol Res 20:325–331
Shinozaki K, Yoda K, Hozumi K, Kira T (1964) A quantitative analy-sis of plant form – the pipe model theory I. basic analyses. JpnJ Ecol 14:97–105
Shutou K, Nakane K (2004) Change in soil carbon cycling for standdevelopment of Japanese Cedar (Cryptomeria japonica) planta-tions following clear-cutting. Ecol Res 19:233–244
Stitt M, Krapp A (1999) The interaction between elevated carbondioxide and nitrogen nutrition: the physiological and molecu-lar background. Plant, Cell and Environment 22:583–621
Sugimoto A, Dan J, Kumai T, Murase T (2003) Adsorption as a meth-ane storage process in natural lake sediment. Geophys Res LettDoi: 10.1029/2003GL018162
Takenaka A (2005) Local coexistence of tree species and the dy-namics of global distribution pattern along environmental gra-dient: a simulation study. Ecol Res 20:297–304
References

296 CHAPTER 23 · Terrestrial Ecosystems in Monsoon Asia: Scaling up from Shoot Module to Watershed
Tanaka Y, Tanaka N (2002) Carbon dioxide exchange and ecosys-tem respiration in a cool-temperate deciduous forest, In: WadaE (eds.): Response of terrestrial ecosystem to global climatechange, Annual synthesis report of IGBP-MEXT, pp 111–118 (inJapanese)
Urabe J, Sekino T, Nozaki K, Tsuji A, Yoshimizu C, Kagami M,Koitabashi T, Miyazaki T, Nakanishi M (1999). Light, nutrientsand primary productivity in Lake Biwa: an Evaluation of thecurrent ecosystem situation. Ecol Res 14:233–242
Urabe J, Yoshida T, Gurung TB, Sekino T, Tsugeki N, Nozaki K, MaruoM, Nakayama E, Nakanishi M (2005) The production to respira-tion ratio and itsi implication in Lake Biwa. Ecol Res 20:367–375
Wayne PM, Bazzaz FA (1997) Light acquisition and growth by com-peting individuals in CO2-enriched atmospheres: consequencesfor size structure in regenerating birch stands. J Ecol 85:29–42
Webster JR, Meyer JL eds (1997) Stream organic matter budgets. JNorth Am Benth Soci 16:3–161
Weiner J (1986) How competition for light and nutrients affects sizevariability in Ipomoea tricolor populations. Ecology 67:1425–1427
Weiner J (1990) Asymmetric competition in plant populations.Trend Ecol Evol 5:360–364
Weiner J, Wright DB, Castro S (1997) Symmetry of below groundcompetition between Kochia scoparia individuals. Oikos 79:85–91
Wilson JB (1988) The effect of initial advantage on the course ofplant competition. Oikos 51:457–472
Yamamoto S, Murayama S, Saigusa N, Kondo H (1999) Seasonal andinter-annual variation of CO2 flux between a temperate forestand atmosphere in Japan. Tellus 51B:402–413
Yoshida T, Kagami M, Gurung TB, Urabe J (2001) Seasonal succes-sion of zooplankton in Lake Biwa. Aquat Ecol 35:19–29
Yoshimizu C, Yoshida T, Nakanishi M, Urabe J (2001) Effects of zoop-lankton on the sinking flux of organic carbon in Lake Biwa. Lim-nology 2:37–43
Yoshimizu C, Urabe J, Sugiyama S, Maruo M, Nakayama E, NakanishiM (2002) Carbon and phosphorus budgets in the pelagic areaof Lake Biwa, the largest lake in Japan. Verh Int Verein Limnol24:1409–1414
Yoshioka T (2000) Characterization of dissolved organic matterusing chemical, isotopic and optical properties. In: Report ofthe Grant-in-Aid for Scientific Research (C), No. 10640474,fromthe Ministry of Education, Culture, Sports, Science, and Tech-nology, Japan, pp 53 (in Japanese)
Yoshioka T, Tanoue E, Ohta K, Masuzawa T, Konohira E, Wu F,Mostofa KG, Ohte N, Kawasaki M, Okazaki R, Katsuyama M, Ta-kahashi M, Hayakawa K, Ueda T, Boontanon N, Yoh M, Ueda S(2001) Response of the terrestrial ecosystem from the viewpointof material cyclings in freshwater system. In Wada E (ed) Re-port of Response of Terrestrial Watershed Ecosystem to Globalenvironmental Change, Scientific Research of Priority Area B(No. 11213101), the Ministry of Education, Culture, Sports, Sci-ence and Technology, Japan.175–194 (in Japanese)
Related Documents











