Systematic Entomology (2009), 34, 649–687 Taxonomic revision of the Mecyclothorax beetles (Coleoptera: Carabidae) of Oahu: epithets as epitaphs for an endangered fauna? JAMES K. LIEBHERR Department of Entomology, Cornell University, Ithaca, New York, U.S.A. Abstract. The Mecyclothorax Sharp beetle fauna of Oahu, Hawaii is revised, including the description of 13 new species—M. acherontius, M. uncinus, M. lobatus, M. dentatus, M. calceus, M. euryoides, M. invictus, M. ovatulus, M. lemur, M. satyrus, M. ramsdalei, M. pelops and M. niobe spp.n.—to complement the six previously described species. Several cryptic species complexes characterized by similar exter- nal anatomy and highly divergent male genitalia are present in the fauna, including the M. brevis (Blackburn) species complex—four species sympatrically distributed on Tantalus, a mountain on the northern margin of Honolulu—and the sympatric sister species M. carteri (Perkins) and M. invictus from Mt. Kaala in the western Waianae Range. This repeated pattern of closely related species characterized by divergent male genitalia and relatively static external anatomy is also observed in the Mecyclothorax fauna of Molokai, an island of moderate geologic age within the Hawaiian archipelago, but not in the Mecyclothorax fauna of the most recently emer- gent Hawaii Island. Species distributions are generally also much more restricted on the older islands. These results are consistent with the occurrence of rampant, unfixed infraspecific genitalic variation early in an island radiation, with subsequent fixation of divergent genitalic characters occurring as speciation proceeds. Three distinct areas of endemism are defined by distributions of Oahu Mecyclothorax : the Waianae Range, the north Koolau Range and the south Koolau Range. Species distributions in two other carabid beetle genera in Hawaii containing native species—Blackburnia Sharp and Bembidion Latreille—congruently define these areas. The historical range of sampling dates is compared for Mecyclothorax species across the three areas, with the greatest diminution of recently collected species-level diversity characterizing the south Koolau Range, where several Mecyclothorax species are characterized both by restricted geo- graphic distributions within the developmental footprint of Honolulu, and by a lack of recent field collections, suggesting that they may qualify for endangered species status. Introduction The currently described diversity of native Hawaiian insects, totalling more than 5000 species (Eldredge and Evenhuis, 2003), is estimated to have been derived from approximately 250 original colonists (Zimmerman, 1948; Liebherr, 2001a). The most diverse lineages, including Hyposmocoma Wals- ingham (Lepidoptera: Cosmopterygidae) (Zimmerman, 1978b; Correspondence: James K. Liebherr, Department of Entomology, John H. & Anna B. Comstock Hall, Cornell University, Ithaca, NY 14853-2601, U.S.A. E-mail: [email protected] Rubinoff & Haines, 2005) and the Hawaiian Drosophilidae (Diptera) (Hardy, 1965; Magnacca & O’Grady, 2008) have radiated spectacularly. Carabid beetles (Coleoptera: Carabidae) constitute the most commonly encountered native insect preda- tors in Hawaii, their numerical abundance complemented by sizeable species-level diversity: the two most diverse lineages include 135 species of Blackburnia Sharp (Tribe Platynini; Liebherr & Zimmerman, 2000; Liebherr, 2001b, 2003, 2006a; Liebherr & Short, 2006), and over 200 species of Mecyclotho- rax Sharp (Tribe Psydrini; Britton, 1948; Liebherr, 2005a, 2006b, 2008a). The Blackburnia radiation is distributed on all © 2009 The Author Journal compilation © 2009 The Royal Entomological Society 649

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Systematic Entomology (2009), 34, 649–687
Taxonomic revision of the Mecyclothorax beetles(Coleoptera: Carabidae) of Oahu: epithetsas epitaphs for an endangered fauna?
J A M E S K. L I E B H E R R
Department of Entomology, Cornell University, Ithaca, New York, U.S.A.
Abstract. The Mecyclothorax Sharp beetle fauna of Oahu, Hawaii is revised,including the description of 13 new species—M. acherontius, M. uncinus, M. lobatus,M. dentatus, M. calceus, M. euryoides, M. invictus, M. ovatulus, M. lemur, M. satyrus,M. ramsdalei, M. pelops and M. niobe spp.n.—to complement the six previouslydescribed species. Several cryptic species complexes characterized by similar exter-nal anatomy and highly divergent male genitalia are present in the fauna, includingthe M. brevis (Blackburn) species complex—four species sympatrically distributedon Tantalus, a mountain on the northern margin of Honolulu—and the sympatricsister species M. carteri (Perkins) and M. invictus from Mt. Kaala in the westernWaianae Range. This repeated pattern of closely related species characterized bydivergent male genitalia and relatively static external anatomy is also observed inthe Mecyclothorax fauna of Molokai, an island of moderate geologic age within theHawaiian archipelago, but not in the Mecyclothorax fauna of the most recently emer-gent Hawaii Island. Species distributions are generally also much more restricted onthe older islands. These results are consistent with the occurrence of rampant, unfixedinfraspecific genitalic variation early in an island radiation, with subsequent fixation ofdivergent genitalic characters occurring as speciation proceeds. Three distinct areas ofendemism are defined by distributions of Oahu Mecyclothorax : the Waianae Range, thenorth Koolau Range and the south Koolau Range. Species distributions in two othercarabid beetle genera in Hawaii containing native species—Blackburnia Sharp andBembidion Latreille—congruently define these areas. The historical range of samplingdates is compared for Mecyclothorax species across the three areas, with the greatestdiminution of recently collected species-level diversity characterizing the south KoolauRange, where several Mecyclothorax species are characterized both by restricted geo-graphic distributions within the developmental footprint of Honolulu, and by a lack ofrecent field collections, suggesting that they may qualify for endangered species status.
Introduction
The currently described diversity of native Hawaiian insects,totalling more than 5000 species (Eldredge and Evenhuis,2003), is estimated to have been derived from approximately250 original colonists (Zimmerman, 1948; Liebherr, 2001a).The most diverse lineages, including Hyposmocoma Wals-ingham (Lepidoptera: Cosmopterygidae) (Zimmerman, 1978b;
Correspondence: James K. Liebherr, Department of Entomology,John H. & Anna B. Comstock Hall, Cornell University, Ithaca, NY14853-2601, U.S.A. E-mail: [email protected]
Rubinoff & Haines, 2005) and the Hawaiian Drosophilidae(Diptera) (Hardy, 1965; Magnacca & O’Grady, 2008) haveradiated spectacularly. Carabid beetles (Coleoptera: Carabidae)constitute the most commonly encountered native insect preda-tors in Hawaii, their numerical abundance complemented bysizeable species-level diversity: the two most diverse lineagesinclude 135 species of Blackburnia Sharp (Tribe Platynini;Liebherr & Zimmerman, 2000; Liebherr, 2001b, 2003, 2006a;Liebherr & Short, 2006), and over 200 species of Mecyclotho-rax Sharp (Tribe Psydrini; Britton, 1948; Liebherr, 2005a,2006b, 2008a). The Blackburnia radiation is distributed on all
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society 649

650 J. K. Liebherr
of the high Hawaiian islands, from Kauai to Hawaii Island. Itsphylogenetic position within Pacific Platynini dates the radia-tion’s Hawaiian origin to the Early Miocene (Liebherr, 2005b),considerably before the origin of Kauai, the current oldest highisland dated to 5.1 Ma (Carson & Clague, 1995). By contrast,the greater species-level diversity of Mecyclothorax beetlesmay have evolved subsequent to the lineage’s colonization ofMaui Nui (Britton, 1948; Liebherr, 2008a): the time of originof the oldest fragment, West Molokai, is dated at 1.9 Ma.
Mecyclothorax beetles have radiated in other portions ofthe generic range, with the evolutionary products often repre-sented by geographically restricted, precinctive species. Suchradiations have been documented for the Queensland Alps ofAustralia (Baehr, 2003), the various massifs of Tahiti (Perrault,1992) and the mountains of New Guinea (Baehr, 2008; Lieb-herr, 2008b). Brachyptery is prevalent among species in theseradiations; indeed, all species of the Tahitian, New Guineanand Hawaiian Island radiations lack metathoracic flight wings.Closely related species are most often allopatrically distributed,suggesting that their phylogenetic relationships may be the keyto understanding the biogeographic history of the areas occu-pied by the species, as proposed by Rosen (1978) and Nelson& Platnick (1981).
Previous taxonomic revisions of Hawaiian Mecyclothoraxhave identified areas of endemism defined by precinctivespecies in Maui (Liebherr, 2005a), Molokai (Liebherr, 2006b)and Hawaii Island (Liebherr, 2008a). This revision extends tax-onomic understanding to Oahu, with the 19 recognized speciesplaced within the context of species occupying the neighbour-ing island of Molokai, as well as the other principal islands ofMaui and Hawaii (Britton, 1948). This revision, like those ofthe Mecyclothorax faunas of Molokai and Hawaii Island, per-mits recognition of putative adelphotaxic species along withthe definition of areas of endemism. The areas of endemismdefined by the Oahu Mecyclothorax species are congruent withthose defined by species in two other Hawaiian carabid beetleradiations: species of the genera Blackburnia Sharp (Lieb-herr & Zimmerman, 2000) and Bembidion Latreille (Liebherr,2008c). Unlike the faunas of the other islands studied to date,the Oahu Mecyclothorax fauna has been greatly influenced bythe development of a major metropolitan centre, namely Hon-olulu, the capital and principal urban center for Hawaii. Thehistorical development of Hawaii has brought with it abundantalien arthropod species that have had a profoundly negativeinfluence on the native fauna (Howarth, 1990). With Honoluluas the principal port of entry for Hawaii, Oahu is the epi-centre for this alien onslaught (Nishida, 2002). This revisionestablishes the basis for diagnosis and identification for the resi-dent Oahu Mecyclothorax species. Several of the species newlydescribed from Tantalus, a mountain in the southern Koolausthat historically housed a diverse insect fauna (Perkins, 1906),have not been observed in nature for the past century. Othernewly described species have been only recently discovered onthe summit of Mt. Kaala in the western Waianae Range. Inte-grating this taxonomic revision with those of Blackburnia andBembidion, given the congruent areas of endemism defined bythe three groups, the Oahu Mecyclothorax, Blackburnia and
Bembidion species can serve as indicators for the continuedassessment of ecosystem health in Oahu (Howarth, 1990).
Taxonomic history
The Reverend Thomas Blackburn collected the first OahuMecyclothorax specimens in 1877, describing three species in1878. Twenty-two specimens labelled by Blackburn have beenexamined for this revision, and these include representativesof three other species he did not recognize. Sharp (1903) sub-sequently revised the Oahu Mecyclothorax with the additionalbenefit of 56 specimens collected by R. C. L. Perkins andAlbert Koebele during the Fauna Hawaiiensis survey (Man-ning, 1986). Sharp recognized only Blackburn’s three species,although his study material included representatives of anothersix, including the three species missed by Blackburn.
After the Fauna Hawaiiensis field collecting was completedin 1901, Perkins and W. M. Giffard continued to collectOahu Mecyclothorax, chiefly from Tantalus (Perkins, 1906),a mountain top just north of Honolulu made easily accessibleby the building of the ‘government road’ up the ridge to theeast of Pauoa Valley (Beasley & Taylor, 1899). From 1902 until1909, Perkins and Giffard collected 152 specimens representingthe three Blackburn names plus five other undescribed species.Perkins (1917) described one of those new species collected in1902—M. carteri (Perkins)—that he had found at about 600 melevation in the Waianae Range (Fig. 1).
Subsequently, Britton (1948) revised the Oahu Mecyclotho-rax based on the 31 specimens remaining in the Natural HistoryMuseum, London after partitioning of the Fauna Hawaiien-sis material with Bishop Museum, Honolulu (Manning, 1986).In his revision, Britton recognized and described two newspecies, resulting in a new total of six described species,although the material at his disposal included representativesof another three. In parallel, Elwood Zimmerman studied the168 specimens held in the Bishop Museum, including theBishop Museum’s cut of Fauna Hawaiiensis specimens (Man-ning, 1986), the Perkins/Giffard material from 1902 to 1909,and subsequent specimens collected by Swezey (1931) andZimmerman. Although Zimmerman did not publish on thesespecies, specimen labels indicate that he recognized threenew species to complement specimens he assigned to five ofthe six previously described species. Male genitalic dissec-tions of specimens labelled as new species by Zimmerman,and Bishop Museum specimens labelled ‘compared with type,E.C.Z.’ strongly support Zimmerman making the dissectionsand type comparisons in association with his several extendedvisits to the Natural History Museum, London during 1949 and1954–1956 (Zimmerman, 1958, 1978a, 1978b).
Materials and methods
Taxonomic material
This revision of the Oahu Mecyclothorax is based on242 specimens deposited in the following institutions: ANIC,
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 651
Australian National Insect Collection, Canberra, Australia;BMNH, the Natural History Museum, London, U.K.; BPBM,Bernice P. Bishop Museum, Honolulu, U.S.A.; CAS, Califor-nia Academy of Sciences, San Francisco, CA, U.S.A.; CUIC,Cornell University Insect Collection, Ithaca, NY, U.S.A.;HDAC, Hawaii Department of Agriculture, Honolulu, HI,U.S.A.; MNHP, Museum National d’Histoire Naturelle, Paris,France; NMNH, National Museum of Natural History, Wash-ington, D.C., U.S.A.; SAMA, South Australian Museum, Ade-laide, Australia.
Recent specimen sampling involved sifting litter and moss,with subsequent field examination of the siftate allowingdiscovery of the active adult beetles. Moss growing on low-stature ohia (Metrosideros polymorpha) trees or logs wassprayed with synthetic pyrethrin insecticide, with specimensexiting the moss collected from a sheet placed below thesprayed limb or log.
In the laboratory, specimens were examined under halogenlight using both ring light and paired fibre-optic wand lightsources. Genitalic dissections were conducted after soakingspecimens in cold distilled water, if necessary, to removethem from archival mounting cards, and then relaxing them innear-boiling distilled water containing a drop of Kodak Photo-Flo® detergent (Kodak Corp., Rochester, NY). Male genitaliawere dissected along with the associated ninth tergite. Theaedeagus was cleared overnight in cold KOH, neutralized indilute 10% acetic acid, and placed in glycerine overnight beforeexamination. When the aedeagal internal sac was everted,eversion was attempted under KOH or dilute acetic acid using afinely ground, hooked minuten nadeln inserted into the medianlobe ostium to pull on the dorsal surface of the internal sacdorsal plate. Female reproductive tracts were removed with theentire abdomen, cleared overnight in cold KOH, and dissected
from the associated tergites and sternites. The ninth-segmentlaterotergites, associated gonocoxites, and internal structures—reproductive tract, gut and pygidial defensive glands—wereneutralized in dilute acetic acid, stained in Kodak ChlorazolBlack® suspended in methyl cellosolve, and mounted ontemporary glycerine microslides for viewing.
Illustrations
Dorsal habitus photographs and photographs of the maleaedeagus and female reproductive tract were made using aMicroptics Digital Imaging System using a Nikon D1X cam-era and Nikon CAPTURE 2.0 software. Specimens were pho-tographed on a transmissible-light stage, augmented whenappropriate with two additional fibre-optic light sources. Lightwas provided by a Microptics ML1000 fibre-optic illumina-tion system running off a DynaLite 2000DR capacitor source.Multiple stacked images were integrated using the COMBINE
Z5.3 public domain software (Hadley, 2006). Owing to thedensity and complexity of the female reproductive tract dissec-tion, those photographs were used as the basis for line drawingsmade with the aid of an Olympus BH-2 phase-contrast com-pound microscope.
Descriptive taxonomy and terminology
For all species, diagnoses are accompanied by comprehen-sive descriptions that include setational and microsculpturalcharacters. Genitalic characters are described for all speciesrepresented by multiple specimens. Unique female specimensaccorded holotype status were not dissected. Species areassigned to Britton’s (1948) groups based on diagnostic char-acters used in his key. Britton’s classification may or may not
Fig. 1. Hawaiian Island of Oahu, with threeareas of endemism supported by distributionsof Mecyclothorax species.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

652 J. K. Liebherr
define monophyletic groups. Nonetheless, putative synapomor-phies and a preliminary assessement of monophyly are pre-sented for each of Britton’s species groups represented in theOahu fauna. Synapomorphies are assessed relative to characterstates observed in Mecyclothorax punctipennis (MacLeay) andM. ambiguus (Erichson) of Australia, those extralimital speciesmost similar to the Hawaiian species M. montivagus (Black-burn) of Maui (Britton, 1948; Liebherr, 2008a). A robust testof monophyly for the various lineages must await compre-hensive description of the estimated 116 species—at least 69new—from Lanai and Maui. Britton’s practical classificationis advantageous as it is based on easily viewed external char-acters, thereby allowing assignment of species to groups ofsimilar habitus. Thus, for the present, users of this taxonomicrevision can identify species-level taxa grouped by relativelylimited patristic distance.
The dichotomous key to adults was developed using LucidProfessional 2.1 software (CPITT, 2001). Qualitative charac-ters were entered into a Lucid matrix, with various pathwaysinvestigated for cleaving the various species-level taxa.
Date–locality information for all specimens is maintainedelectronically, with data fields described in Liebherr & Zim-merman (2000), and the database updated from Liebherr(2000). Literature citations are based on the Hawaiian Ter-restrial Arthropod Database—Bibliography (G. Nishida & N.Evenhuis, personal communication). Label data are transcribedverbatim for primary types, with the individual labels on thespecimen pin delimited by the symbol ‘/’. Paratype data com-prise a sequence of the following fields: locality, date, eleva-tion, habitat or collecting method, collector, and repository withnumber of specimens. To conserve print space, if paratypeswere collected under different circumstances from the samelocality, the locality field is retained only for the first record inthe sequence, with subsequent topotypic records delimited bycommas.
Although the terms anterior and posterior are properlymodified to anteriad and posteriad when relative positions arediscussed, the more commonly used constructions anterad andposterad are used herein (Krombein & Pulawski, 1994).
A limited number of ratios proved useful for quantifyingbody shape. Eye development was quantified using an ocularratio: the ratio of the greatest distance across the compoundeyes divided by the minimum interocular distance across thefrons. This ratio is generally larger when the eyes are moredeveloped; however, if the dorsal eye margins converge onthe frons, the ratio may be large although the eyes are smalland protrude little. An additional ratio—ocular lobe ratio—isused to assist describing this condition. This ratio is definedas the distance from the front to hind margin of the eyemeasured from above, over the distance likewise measuredfrom the front margin of the eye to the postocular groove.The postocular groove occurs at the juncture of the ocularlobe and the gena posterad the eye. The smaller this ratio,the less the eye covers the ocular lobe. Pronotal breadth wasquantified using the ratio of maximum pronotal width overmedian pronotal length; MPW/PL. The basal constriction ofthe pronotum was quantified using the ratio of the maximum
pronotal width over the basal pronotal width. Basal pronotalwidth is easily measured when the laterobasal pronotal marginsare sinuate and hind angles are present. For the Mecyclothoraxbrevis (Blackburn) species complex, where the pronotal hindangles are much reduced and the laterobasal margin convexlycurved, the point along the margin of minimum radial curvaturewas used to represent the hind angle, with the basal widthmeasured between those points.
Standardized body length was determined as the sum of threemeasurements: (i) the median distance from the labral apicalmargin to the cervical ridge of the head capsule (estimatedwhen necessary by the lateral curvature of the cervical ridge);(ii) median pronotal length; plus (iii) the distance from thebasal ridge of the scutellum to the apex of the longer elytron,measured parallel to the elytral suture. Such a measurementunderestimates the apparent size, as it neglects mandibularextension and any distension of the abdomen, but it can bemade on preserved specimens in various postures.
Lateral elytral setae are described by the formula a + b,where a is the number of setae continuously grouped inthe eighth elytral interval just posterad the humerus, and b
is the number of setae arrayed in the apical portion of theeighth interval just anterad the subapical sinuation. If thenumber of setae is shown to vary infraspecifically withineither setal group, the less common number of setae (c) isgiven parenthetically; for example a + b(c). If a seta is presentbetween the anterior and posterior series, ‘1’ is added betweenbounding ‘+’ signs, within parentheses if that condition isuncommonly observed.
Male aedeagal structures follow the terminology of Liebherr(2006b, 2008a), which is based in part on homologies presentedin Maddison (1993). Female reproductive tract charactersfollow the terminology of Liebherr & Will (1998). The majordeviation from that treatment involves the presence of amembranous fold of cuticle immediately mesad the base ofgonocoxite 1 (e.g. Fig. 9). Although unsclerotized, this foldof membrane is always present in Mecyclothorax females,raising the prospect that it is homologous at some level withthe gonocoxal ramus observed in Psydrus piceus LeConte(Psydrini) and Broscus cephalotes (L.) (Broscini) (Liebherr &Will, 1998).
In the instance of allopatrically distributed cryptic species—for example M. simiolus (Blackburn), M. lemur sp.n. andM. satyrus sp.n.—one male representative of each collect-ing series was dissected, thereby permitting assignment ofseries to species. When cryptic species were distributed sym-patrically or parapatrically—for example the M. brevis speciescomplex—all available males were either dissected or theirgenitalic configuration was directly examined if the medianlobe apex extended from the abdomen. After series wereassigned to species through male dissection, representative con-serial females were dissected in order to elucidate charactersof the female reproductive tract. Finally, the external anatomi-cal characters of these genitalically associated specimens wereexamined to finalize diagnoses, partitioning infraspecific mor-phological variation from the subtle interspecific diagnosticcharacters. The number of individuals dissected is provided
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 653
parenthetically at the beginning of the ‘Male genitalia’ and‘Female reproductive tract’ sections of the descriptions.
Results
Key to the adults of Mecyclothorax from Oahu
1. Pronotum with lateral seta just anterad midpoint of length. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2
– Pronotum glabrous, without lateral seta . . . . . . . . . . . . . . . 72(1). Pronotal laterobasal margin straight or convex, the hind
angle evident but very obtuse, angle defined as change incurvature of basal and laterobasal margin. . . . . . . . . . . . . .3
– Pronotal laterobasal margin markedly sinuate, hind anglenearly right (Fig. 2A); discal elytral striae 1–3 markedlypunctate, striae 4–5 comprising series of shallowly con-nected punctures near midlength of elytra; distributed innorth Koolau Range (Fig. 5) . . . . . . . M. acherontius sp.n.
3(2). Frons medially between eyes covered with dis-tinct isodiametric sculpticells, the sculpticells convex withdeep margins resulting in granulate surface(Fig. 3A) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
– Frons medially between eyes covered with more indistinct—in some specimens difficult to trace—microsculpture con-sisting of transverse sculpticells or isodiametric sculpticellsirregularly arranged in transverse rows, the surface glossy(e.g. Fig. 3B) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
4(3). Elytral intervals 1–2 on disc covered with indistinct iso-diametric sculpticells in transverse rows, elytral intervals3–5 on disc with greater fraction of transverse sculpticells,the elytral disc glossy owing to shallow sculpticells mar-gins; male aedeagal median lobe apex hooked (Fig. 6A);distributed in Waianae Range (Fig. 7) . M. uncinus sp. n.
– Elytral intervals 1–5 uniformly covered with distincttransverse mesh microsculpture, lateral breadth of sculp-ticells 2–4× length; male aedeagal median lobe withrounded apex, and with large dorsoapical claw-like pro-jection (Fig. 6B–D); distributed in south Koolau Range(Fig. 10A) . . . . . . . . . . . . . . . . . . . . . . M. brevis (Blackburn)
5(3). Elytral discal intervals covered with elongate transversemesh microsculpture, the sculpticells longenough so that microsculpture comprises transverse par-allel lines on lateral intervals 4–5; striae 6 and 7 bothshallower than striae 1–5; aedeagal median lobe ventralsurface evenly curved to very slightly recurved, dorsalsurface evenly convex or with small tooth-like projection(Fig. 6H–I, K) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
– Elytral discal intervals covered with transverse meshmicrosculpture that includes some sculpticells with lateralbreadth 2× length, lateral intervals covered with elon-gate transverse mesh microsculpture; stria 6 as deeplyincised as stria 5 near midlength of elytron, stria 7 maybe shallower or not near midlength; aedeagal medianlobe with ventral surface recurved and broadly distended,dorsal surface expanded into a very large lobe-like pro-jection (Fig. 6F–G); distributed in south Koolau Range(Fig. 10B) . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. lobatus sp.n.
6(5). Pronotum narrower (Fig. 2E), ratio of maximum pronotalwidth (MPW) to median pronotal length (PL) 1.25–1.33;elytral discal striae distinctly incised, smooth to slightlyirregular along length, but without distinct punctures;aedeagal median lobe with distinct tooth on dorsal sur-face, ventroapical surface expanded just apical ostialopening (Fig. 6H–I); distributed in south Koolau Range(Fig. 10B) . . . . . . . . . . . . . . . . . . . . . . . . . . M. dentatus sp. n.
– Pronotum broader (Fig. 2F), ratio of MPW/PL1.35–1.41; elytral discal striae shallower, lined withdistinct elongate punctures; aedeagal median lobe dor-sal surface evenly convex, apex elongate with paralleldorsal and ventral margins, the tip narrowly rounded(Fig. 6K); distributed in south Koolau Range(Fig. 10A) . . . . . . . . . . . . . . . . . . . . . . . . . . . M. calceus sp. n.
7(1). Second elytral stria as strongly impressed as sutural striaat elytral midlength, striae 3–4 evident, punctate nearmidlength . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
– Second elytral stria much shallower than sutural striaat elytral midlength, striae 3–4 either very shallow andimpunctate, or comprising series of isolatedpunctures . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
8(7). Elytral disc and margins concolorous, or lateral marginsnarrowly paler; elytral striae 1–5 impressed, sutural striadeepest, striae 6–7 obsolete, traceable as series of isolatedpunctures . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
– Elytra with broadly flavous lateral, basaland sutural margins, the disc of each elytrondarkened by a piceous cloud (Fig. 2G); elytralstriae 1–7 impressed, evident, although suturalstria deepest; distributed in south Koolau Range(Fig. 10C) . . . . . . . . . . . . . . . . M. flavomarginatus (Britton)
9(8). Pronotum cordate, base narrow, ratio of maximum pronotalwidth (MPW) to basal pronotal width (BPW) 1.40–1.48(Fig. 4A–D); pronotal base with either round punctures orlongitudinal wrinkles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
– Pronotum broad basally, ratio MPW/BPW = 1.21–1.24(Fig. 2H); pronotal base smooth, without evident puncta-tion or wrinkles; distributed in Waianae Range(Fig. 13A) . . . . . . . . . . . . . . . . . . . . . . . . . M. euryoides sp.n.
10(9). Elytra broad basally, humeri extended laterally so thatelytra are more quadrate (Fig. 4A,B) . . . . . . . . . . . . . . . . 11
– Elytra narrow basally, the humeri rounded so that elytraare ovoid (Fig. 4C–D) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
11(10). Pronotal disc glossy, microsculpture reduced in places,where present sculpticells are very elongate, marginsdefining parallel lines irregularly joined into a mesh;discal elytral intervals 2–4 with elongate transversemesh microsculpture, the transverse sculpticells inter-mixed with parallel transverse lines, the surface withat most an indistinct silvery iridescence; male aedeagalmedian lobe slender, lobe apex extended to narrowlyrounded tip (Fig. 11D–F); distributed in Waianae Range(Fig. 7) . . . . . . . . . . . . . . . . . . . . . . . . . . . M. carteri (Perkins)
– Pronotal disc with distinct transverse meshmicroscuplture, the lateral breadth of sculpticells2–3 × length; discal elytral intervals 2–4 with
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

654 J. K. Liebherr
Fig. 2. Oahu Mecyclothorax species, dorsal view. A, M. acherontius ; B, M. uncinus ; C, M. brevis ; D, M. lobatus ; E, M. dentatus ; F. M. calceus;G. M. flavomarginatus ; H, M. euryoides .
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 655
Fig. 3. Head capsule, dorsal view, illustrat-ing different microsculptures observed in theM. brevis species complex. A, M. brevis ; B,M. dentatus .
evident, complete transverse-line microsculpture, thesurface with cupreous iridescence; male aedeagalmedian lobe robust, lobe apex short, broadly rounded(Fig. 11G–I); distributed in Waianae Range(Fig. 13B) . . . . . . . . . . . . . . . . . . . . . . . . . . . M. invictus sp.n.
12(10). Elytral discal intervals glossy, microsculpture obsolete;pronotal hind angle distinctly angulate, apex of anglenarrowly rounded; distributed in south Koolau Range(Table 1) . . . . . . . . . . . . . . . . . . . . M. oahuensis (Blackburn)
– Elytral discal intervals with distinct microsculptureconsisting of transverse parallel lines; pronotal hindangle less angulate, the basal margin curved anteradto meet sinuate laterobasal margin, angle apexrounded; distributed in south Koolau Range (Table 1). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. ovatulus sp.n.
13(7). Pronotal laterobasal margin distinctly sinuate, hind angleright or nearly so (Fig. 4E–G) . . . . . . . . . . . . . . . . . . . . . . 14
– Pronotal laterobasal margin straight to convex, hindangle broadly obtuse to obsolete, the margin rounded(Fig. 4H–K) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
14(13). Pronotal laterobasal sinuation short, lateral margins diver-gent nearly from hind angles (Fig. 4F, G); surface of elytraldiscal intervals glossy without evidence of microsculpture(125×) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
– Pronotal laterobasal sinuation elongate, marginsparallel for short distance before hind angles(Fig. 4E); surface of elytral discal intervals glossy,but with transverse mesh microsculpture clearlyvisible (125×); distributed in Waianae Range(Fig. 13C) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. lemur sp.n.
15(14). Eyes larger, outer surface convex, ocular ratio1.40–1.43 (Fig. 4F); pronotal base with distinct anddeep punctures; elytral basal groove somewhat curved,elytral humeri broader; male aedeagal median lobebroad to apex, tip acuminate, apical face straight(Fig. 15A, B); distributed in south Koolau Range(Fig. 16A) . . . . . . . . . . . . . . . . . . . . M. simiolus (Blackburn)
– Eyes smaller, little protruded, ocular ratio1.37–1.38 (Fig. 4G); pronotal base with smaller,
shallower punctures; elytral basal groove shorter, humerinarrower; male aedeagal median lobe with large api-cal hook, ventroapical tip broadly rounded(Fig. 15D, E); distributed in north Koolau Range(Fig. 5) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. satyrus sp.n.
16(13). Elytral discal striae 1–3 distinctly punctate, with striaeeither continuously depressed, or represented by series ofisolated punctures (Fig. 4I–K); pronotal lateral marginaldepression narrow throughout length, pronotum moreorbicular, MPW/PL = 1.20–1.25 . . . . . . . . . . . . . . . . . . . 17
– Elytral striae nearly impunctate, sutural stria slightlyirregular but without any discernible punctation, stria 2with minute punctulae, striae 3–4 shallow, smooth(Fig. 4H); pronotal lateral marginal depression mod-erately broad in basal half, pronotum broader,MPW/PL = 1.36; distributed in Waianae Range(Fig. 13C) . . . . . . . . . . . . . . . . . . . . . . . . . M. ramsdalei sp.n.
17(16). Pronotal hind angles obtusely angulate, defined by junctureof straight laterobasal margin and anteriorly curved basalmargin (Fig. 4J, K); eyes smaller, less convex, ocular ratio1.31–1.35; standardized body length 3.7–4.2 mm . . . . 18
– Pronotal hind angles obsolete, laterobasal and basal mar-gins not defining a discrete obtuse angle (Fig. 4I); eyeslarger, more convex, ocular ratio 1.37–1.41; standard-ized body length 4.3–4.7 mm; distributed in south KoolauRange (Fig. 16B) . . . . . . . . . . . . . . . . . M. tantalus (Britton)
18(17). Elytra slightly iridescent owing to very fine transverse-line microsculpture observable on intervals 2–4 near ely-tral midlength (80×); elytra more elongate, apex narrowlyrounded, humeri narrowed basally (Fig. 4J); standardizedbody length 3.8–4.2 mm; distributed in north KoolauRange (Fig. 5) . . . . . . . . . . . . . . . . . . . . . . . . .M. pelops sp.n.
– Elytral surface glossy, without evidence of anymicrosculpture; elytra broadly ovoid, apex broadlyrounded, humeri more broadly extended basally(Fig. 4K); standardized body length 3.7–3.8 mm; distributed in south Koolau Range(Fig. 10C) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. niobe sp.n.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

656 J. K. Liebherr
Fig. 4. Oahu Mecyclothorax species, dorsal view. A, M. carteri ; B, M. invictus ; C, M. oahuensis ; D, M. ovatulus ; E, M. lemur ; F, M. simiolus ;G, M. satyrus ; H. M. ramsdalei ; I, M. tantalus ; J, M. pelops ; K, M. niobe.
Mecyclothorax Sharp, 1903
For most recent and complete synonymy see Liebherr & Marris(2009).
Mecyclothorax palustris group
Diagnosis. Species diagnosable by: (i) elytral microsculp-ture distinct; (ii) elytral disc flatter medially, not greatly
elevated relative to lateral elytral margins; (iii) lateral ely-
tral striae—at least the seventh—reduced, and mesal striae—
at least striae 1–3—developed on disc; (iv) elytral stria
2 less impressed than sutural stria at apex; (v) pronotal
laterobasal margins sinuate, the hind angles distinct, acute to
slightly obtuse; and (vi) standardized body length moderate,
4.0–6.0 mm (Liebherr, 2006b, 2008a). The cordate prono-
tum and reduced apex of elytral stria 2 are synapomorphous,
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 657
although these synapomorphies may define this group as para-phyletic relative to other groups exhibiting greater reductionsin elytral striae, for example the M. scaritoides group.
The M. palustris group also includes three Molokai species,two species from Lanai, one described species from Maui,and four from Hawaii Island (Britton, 1948; Liebherr,2006b, 2008a).
Mecyclothorax acherontius sp.n.(Figs 2A; 5)
Type material. Holotype female: Pupukea Oahu, HawaiiXII-17-1933/N.L.H. Krauss/HOLOTYPE Mecyclothorax ache-rontius J.K. Liebherr 2008 (red, black-bordered label) (BPBM).
Diagnosis. Most similar to M. palustris (Sharp) of Molokai(Liebherr, 2006b), but differing by: (i) the small, indistinctpunctures surrounded by smooth cuticle on the median base ofthe pronotum, versus distinct punctures set in an irregular sur-face, those punctures near the disc strigiform in M. palustris;(ii) eyes small, little projected from ocular lobes that meet headcapsule behind eye at obtuse angle, ocular ratio 1.50, versuseyes more convex, ocular ratio 1.54, with ocular lobes meet-ing gena at deep groove behind eye; (iii) pronotal laterobasalmargins slightly divergent from hind angle, the hind anglesdistinctly obtuse (Fig. 2A), versus laterobasal margins slightlyconvergent or parallel anterad hind angles, the hind anglesnearly right to slightly obtuse in M. palustris. Standardizedbody length 4.9 mm.
Description. Head capsule with frons broadly convex,frontal grooves shallow, minute transverse wrinkles mesadfrontal grooves and paired supraorbital setae; ocular loberatio 0.83; mentum toothed medially, the projection trian-gular, the apex a right angle. Pronotum transverse, maxi-mum pronotal width/median pronotal length 1.35; pronotalbase unmargined, evenly convex between the obtuse hindangles; pronotal base nearly smooth medially, punctures therevery shallow, lateral punctures deeper, but surrounded bysmooth cuticle; median longitudinal impression very finelyincised, shallow; anterior transverse impression very broad,shallow, smooth, traceable only near broadly rounded frontangles; lateral marginal depression broadest near front angle,lined there with isodiametric sculpticells, narrower near lat-eral seta, slightly upturned; laterobasal depression continu-ous with lateral marginal depression, broadly and smoothlyelevated to hind angles. Elytra subquadrate, broad basally;elytral striae 1–5 continuous, impressed on disc, stria 6 dis-continous, a series of punctures variously joined by the strialdepression, stria 7 absent; parascutellar striole and stria 1continous to base, stria 1 smooth and continuous to apex,stria 2 traceable to apex but markedly shallower, striae 3–6reduced to obsolescence anterad point of juncture of striae 4and 5; two dorsal elytral setae positioned just mesad third striaand depressing half of third interval; elytral humeri broadlyrounded, basal groove rounded; lateral elytral setae in series
Fig. 5. Distribution of Mecyclothorax species in the north KoolauRange area of endemism: •, M. acherontius ; ©, M. satyrus; �,sympatric M. satyrus and M. pelops .
of 6 + 6. Coloration of dorsal body surface rufopiceous, thebasal three antennomeres and palps brunneous; ventral bodysurface piceous, contrasted with rufoflavous elytral epipleura;legs rufobrunneous, with piceous infuscation medially alonglength of femora. Microsculpture of frons reduced isodiamet-ric, the sculpticells nearly indistinguishable; pronotal disc withobsolete irregular transverse mesh microsculpture, the meshesreduced to transverse lines over portions of the disc; discal ely-tral intervals covered with distinct sculpticells, ranging fromnearly isodiametric to transverse (i.e. twice as broad as long)with sculpticells of varying shapes associated with strial andsetal depressions.
Female reproductive tract. The lone female holotype spec-imen was not dissected.
Distribution. This species is recorded from the northKoolau Range locality of Pupukea (Fig. 5), currently a campof the Boy Scouts of America.
Etymology. The species epithet is a latinization of theGreek Acherontieos, that is, ‘pertaining to the River Acheron’,a river in the underworld (Jaeger, 1955). The name assumesthat this species exhibits a habitat preference much like that ofthe anatomically similar riparian species M. palustris (Sharp)of Molokai.
Mecyclothorax brevis group
Diagnosis. Species of this group are characterized by twosynapomorphies: (i) continuous presence of elytral striae 1–7
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

658 J. K. Liebherr
from the disc to the elytral apex, although striae 6 and 7 may beshallower (their relative depressed condition must be assessedwith the lateral surface of the elytron perpendicular to the fieldof view); (ii) obsolescent, glabrous pronotal hind angles, theangles marked at most by a change in curvature of the convexlaterobasal margin.
This group is centred on Oahu, with the only other memberbeing M. latissimus Liebherr (2006b) of Molokai. Given theunique derivation of the rounded pronotal base and thewell-developed striae, this geographically restricted group isprovisionally considered monophyletic.
Within the treatment of this group, all Oahu species arediagnosed, with the associated species descriptions based oncomparison with the comprehensive description of the mostprevalently collected species, M. brevis.
Mecyclothorax uncinus sp.n.(Figs 2B; 6A; 7)
Type material. Holotype male (card mounted with left pro-and mesolegs separately glued to card, and male genitalia inplastic genitalia vial): T. brevis var. D.S. Oahu Perkins 670(on obverse of card)/Hawaiian Is. R.C.L. Perkins (yellow-linedlabel)/Sharp Coll 1905-313/HOLOTYPE Mecyclothorax unci-nus J.K. Liebherr 2008 (red, black-bordered label) (BMNH).
Diagnosis. Best distinguished from other species of theM. brevis complex by the male aedeagal median lobe with itshooked apex, and well-developed dorsal ostial microtrichialpatch (Fig. 6A). Externally it can be distinguished from theother species of the complex by: (i) elytra with reducedmicrosculpture of isodiametric and transverse sculpticells onthe discal intervals, transverse sculpticells predominating onthe lateral intervals, the surface glossy; (ii) elytral humeri pro-jected anterad, the elytral basal groove distinctly curved to thenarrowly rounded humerus (Fig. 2B); and (iii) elytral stria 7 asdeeply impressed as stria 6. Standardized body length 4.4 mm.
Description. Deviating from the description of M. brevisby the following. Head capsule with indistinctly convex frons,delimited laterally by frontal grooves that do not depress thefrontal surface; anterior of two supraorbital setae separatedfrom frontal grooves by a narrow carina; outer surface of eyesprojected slightly beyond curvature defined by posterior por-tion of ocular lobe, ocular ratio 1.45, ocular lobe ratio 0.74;labrum five-setose (median seta unpaired in unique holotype).Pronotum transverse, MPW/PL 1.38; median base coveredwith shallow, longitudinal wrinkles lined with distinct isodi-ametric sculpticells; median longitudinal impression deep andcontinuous from basal margin to disc, broader and shalloweron disc; anterior transverse impression well defined, crossedby longitudinal wrinkles giving a punctate appearance; cal-losity anterad transverse impression depressed relative to discposterad transverse impression; pronotal front angles projected,rounded; pronotal lateral marginal depression broad enoughto be lined with distinct isodiametric sculpticells throughout
pronotal length, outer half of marginal depression translucentthroughout. Elytra with discal striae smooth, evenly impressed,only the slightest irregularities in the deepest portions of thestriae; parascutellar striole present, shallow, impunctate; para-scutellar seta present just mesad stria 1; two dorsal elytral setaepresent just mesad third stria, at about 0.3× and 0.6× ely-tral length; lateral elytral setae arranged as 7 + 6. Tarsomeresbroadened apically, fourth metatarsomere apical breadth about0.67× median length; basal two metatarsomeres gracile, dorsalsurface convex between dorsolateral sulci. Coloration of dor-sal body surface glossy brunneous, elytral disc broadly darker,glossy piceous; ventral coloration as M. brevis. Microsculp-ture on frons a uniformly upraised isodiametric mesh, with-out the formation of wrinkles; pronotum covered with distinctmicrosculpture, a mixture of isodiametric and transverse sculp-ticells organized in transverse rows.
Male genitalia (n = 1). In addition to the characters in thediagnosis, the male aedeagal median lobe is tubular, withoutan accessory subapical projection (Fig. 6A; Liebherr, 2006b);median lobe apex with straight apical face, the lobe tip definingan obtusely rounded right angle; internal sac spiculate andmelanized on the ventral surface of the apical portion; dorsalplate large and robust, the dorsal surface defined by a distinctlysclerotized peritreme.
Distribution. The Perkins’ lot 670 ‘Anonymous, (N.D.), towhich the holotype specimen was assigned, is recorded fromthe lee, or west side, of Mt. Kaala between ‘2000+ −3000 ft;’>610–914 m elevation (Fig. 7).
Etymology. The species epithet is the Latin adjectiveuncinus, meaning hooked or barbed (Brown, 1956), anddenoting the condition of the male aedeagal median lobe apex.
Mecyclothorax brevis (Blackburn, 1878)(Figs 2C; 3A; 6B–E; 8A; 9A, B; 10A)
Cyclothorax brevis Blackburn 1878: 123.Thriscothorax brevis, Sharp 1903: 267; Perkins 1906: 48(biology).Mecyclothorax brevis, Britton 1948: 156.
Type material. Lectotype male (card mounted with malegenitalia in plastic genitalia vial) hereby designated: cardwith two-lined Blackburn Oahu code on obverse (Zimmerman,1957), ‘brev.’ on reverse/Type (round, red-bordered label)/Hawaiian Is. Rev.T. Blackburn. 1888–1930. (yellow-linedlabel)/LECTOTYPE Cyclothorax brevis BlackburnJ.K. Liebherr 1998 (red, black-bordered label) (BMNH). Thislectotype designation is required because the original typeseries included specimens of this species and also one specimenof M. lobatus (SAMA).
Diagnosis. This species is distinguished by the distinctisodiametric microsculpture on the frons, the sculpticell mar-gins upraised to the degree that they produce a wrinkled,granulate, and irregularly iridescent surface (Fig. 3A). Mecy-clothorax brevis can be distinguished from its microsympatric
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 659
Fig. 6. Male aedeagal median lobe and parameres, right lateral view except where noted. Collecting locality given in brackets for specimens ofM. brevis species complex. A, M. uncinus, internal sac everted (Mt. Kaala); B, M. brevis (Tantalus, 550 m el.); C, M. brevis (Oahu; Blackburnlectotype); D, M. brevis (Tantalus, 550 m el.); E, M. brevis, ventral view (Tantalus, 550 m el.); F, M. lobatus (Tantalus, 610 m el.); G, M. lobatus(Tantalus, 610 m el.); H, M. dentatus (Pauoa, 460 m el.); I, M. dentatus, internal sac everted (Oahu; Perkins lot 787); J, M. dentatus, ventral view(Pauoa, 460 m el.); K, M. calceus (Tantalus 400 m el.); L, M. flavomarginatus ; M, M. flavomarginatus, ventral view. Abbreviations: af, apical face;asp, accessory subapical projection; dop, dorsal ostial microtrichial patch; dp, dorsal plate; dpp, dorsal plate peritreme; ve; ventral expansion.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687
660 J. K. Liebherr
Fig. 7. Distribution of Mecyclothorax species in the Waianae Rangearea of endemism: •, M. uncinus ; ©, M. carteri .
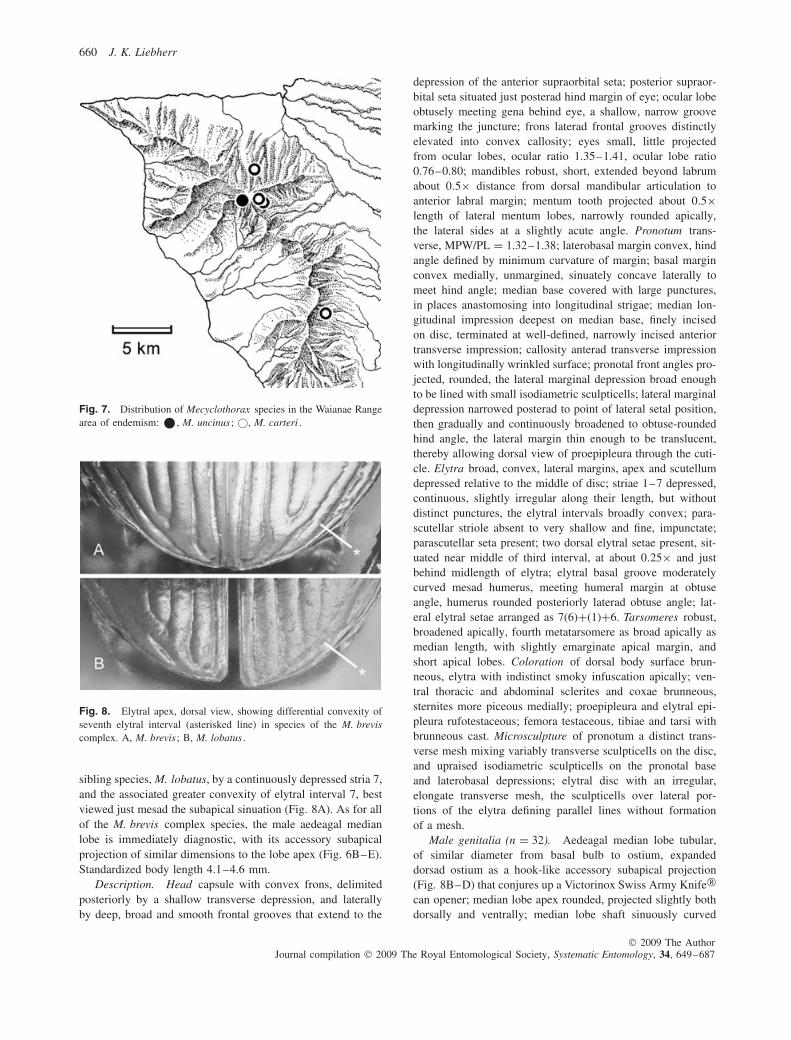
Fig. 8. Elytral apex, dorsal view, showing differential convexity ofseventh elytral interval (asterisked line) in species of the M. breviscomplex. A, M. brevis ; B, M. lobatus .
sibling species, M. lobatus, by a continuously depressed stria 7,and the associated greater convexity of elytral interval 7, bestviewed just mesad the subapical sinuation (Fig. 8A). As for allof the M. brevis complex species, the male aedeagal medianlobe is immediately diagnostic, with its accessory subapicalprojection of similar dimensions to the lobe apex (Fig. 6B–E).Standardized body length 4.1–4.6 mm.
Description. Head capsule with convex frons, delimitedposteriorly by a shallow transverse depression, and laterallyby deep, broad and smooth frontal grooves that extend to the
depression of the anterior supraorbital seta; posterior supraor-bital seta situated just posterad hind margin of eye; ocular lobeobtusely meeting gena behind eye, a shallow, narrow groovemarking the juncture; frons laterad frontal grooves distinctlyelevated into convex callosity; eyes small, little projectedfrom ocular lobes, ocular ratio 1.35–1.41, ocular lobe ratio0.76–0.80; mandibles robust, short, extended beyond labrumabout 0.5× distance from dorsal mandibular articulation toanterior labral margin; mentum tooth projected about 0.5×length of lateral mentum lobes, narrowly rounded apically,the lateral sides at a slightly acute angle. Pronotum trans-verse, MPW/PL = 1.32–1.38; laterobasal margin convex, hindangle defined by minimum curvature of margin; basal marginconvex medially, unmargined, sinuately concave laterally tomeet hind angle; median base covered with large punctures,in places anastomosing into longitudinal strigae; median lon-gitudinal impression deepest on median base, finely incisedon disc, terminated at well-defined, narrowly incised anteriortransverse impression; callosity anterad transverse impressionwith longitudinally wrinkled surface; pronotal front angles pro-jected, rounded, the lateral marginal depression broad enoughto be lined with small isodiametric sculpticells; lateral marginaldepression narrowed posterad to point of lateral setal position,then gradually and continuously broadened to obtuse-roundedhind angle, the lateral margin thin enough to be translucent,thereby allowing dorsal view of proepipleura through the cuti-cle. Elytra broad, convex, lateral margins, apex and scutellumdepressed relative to the middle of disc; striae 1–7 depressed,continuous, slightly irregular along their length, but withoutdistinct punctures, the elytral intervals broadly convex; para-scutellar striole absent to very shallow and fine, impunctate;parascutellar seta present; two dorsal elytral setae present, sit-uated near middle of third interval, at about 0.25× and justbehind midlength of elytra; elytral basal groove moderatelycurved mesad humerus, meeting humeral margin at obtuseangle, humerus rounded posteriorly laterad obtuse angle; lat-eral elytral setae arranged as 7(6)+(1)+6. Tarsomeres robust,broadened apically, fourth metatarsomere as broad apically asmedian length, with slightly emarginate apical margin, andshort apical lobes. Coloration of dorsal body surface brun-neous, elytra with indistinct smoky infuscation apically; ven-tral thoracic and abdominal sclerites and coxae brunneous,sternites more piceous medially; proepipleura and elytral epi-pleura rufotestaceous; femora testaceous, tibiae and tarsi withbrunneous cast. Microsculpture of pronotum a distinct trans-verse mesh mixing variably transverse sculpticells on the disc,and upraised isodiametric sculpticells on the pronotal baseand laterobasal depressions; elytral disc with an irregular,elongate transverse mesh, the sculpticells over lateral por-tions of the elytra defining parallel lines without formationof a mesh.
Male genitalia (n = 32). Aedeagal median lobe tubular,of similar diameter from basal bulb to ostium, expandeddorsad ostium as a hook-like accessory subapical projection(Fig. 8B–D) that conjures up a Victorinox Swiss Army Knife®can opener; median lobe apex rounded, projected slightly bothdorsally and ventrally; median lobe shaft sinuously curved
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 661
Fig. 9. Female reproductive tract, ventral view (A, C, E, G) and left gonocoxa, ventral view (B, D, F, H). A, B, M. brevis ; C, D, M. lobatus ;E, F, M. dentatus ; G, H, M. flavomarginatus. Abbreviations: alf, apicolateral fringe setae; am, apicomedial seta; an, apical nematiform setae; bc,bursa copulatrix; co, common oviduct; de, dorsal ensiform seta; gc1, basal gonocoxite 1; gc2, apical gonocoxite 2; le, lateral ensiform setae; sg,spermathecal gland; sp, spermatheca.
laterally (Fig. 6E), with accessory subapical projection offsetleftwards relative to the lobe apex; aedeagal internal sacnarrow, membranous, without microtrichial fields (Fig. 6D);dorsal plate small, lightly sclerotized. The range of variationin median lobe configuration is limited to: (i) differentialdevelopment of the accessory subapical projection apex, fromblunt (Fig. 6B) to acuminate (Fig. 6D); and (ii) differentialexpansion of the apex, resulting in a rounded tip in lateralview (Fig. 6C) to a slightly hooked tip (Fig. 6B).
Female reproductive tract (n = 7). Bursa copulatrixcolumnar, broadly rounded apically, apex may be expandedslightly relative to bursal base near common oviduct juncture(Fig. 9A), bursa copulatrix length 2.5–3.2× gonocoxal length;spermathecal gland duct short, length subequal to slightlyshorter than spermathecal gland reservoir, the gland duct andreservoir assembly about as long as spermathecal duct; basalgonocoxite 1 (Fig. 9B) with apical fringe of two to four setae
(count may vary by two in an individual), and zero to twomedioapical setae (count may vary by two in an individual,median count per side is one seta); apical gonocoxite 2 broadbasally, gradually narrowed to subacuminate tip, bearing twolateral ensiform setae, one dorsal ensiform seta and two apicalnematiform setae.
Distribution. Distributed on Tantalus, with recorded local-ities between 545 and 614 m elevation (Fig. 10A). Perkins’specimens labelled ‘beyond Tantalus toward Pauoa’, and ‘Tan-talus, Manoa side’ suggest localities some small distance awayfrom the actual summit area.
Habitat. Mecyclothorax brevis has been collected amonggrass roots, and under an Elaeocarpus, or Kalia tree. Writingabout the insects of Tantalus, Perkins (1906: 48) stated:‘Thriscothorax [= Mecyclothorax ] brevis is not rare and isalways found on the ground . . .’
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

662 J. K. Liebherr
Fig. 10. Distribution of Mecyclothorax species in the south KoolauRange area of endemism: A, •, M. brevis, ©, M. calceus; B,•, M. lobatus, ©, M. dentatus ; C, •, M. flavomarginatus,©, M. niobe.
Mecyclothorax lobatus sp.n.(Figs 2D; 6F, G; 8B; 9C, D; 10B)
Cyclothorax brevis (Blackburn, 1878): 123 (in part).
Type material. Holotype male (card-mounted, dissectedmale genitalia in plastic genitalia vial): 2000 ft T. 4 II 06(in pencil on reverse of card)/Tantalus 2000 ft/W. M. Giffard
4.II.06/ W.M.Giffard Collection/Mecyclothorax brevis com-plex male 7 J.K. Liebherr 2008/HOLOTYPE Mecyclothoraxlobatus J.K. Liebherr 2008 (red, black-bordered label) (BPBM).Paratypes , HAWAII, Oahu: ‘Blackburn 2-line Oahu code’(Zimmerman, 1957) (Blackburn) (SAMA, 1); Tantalus, underrocks, 18.xii.1929 (Williams) (HDAC, 1), 545 m, 20.v.1906(Giffard) (BPBM, 1), 545 m, 11.vi.1906 (Giffard) (CUIC, 2),605 m, 1.ii.1906 (Giffard) (BPBM, 4), 605 m, 3.ii.1906 (Gif-fard) (BPBM, 3), 605 m, 9.ix.1906 (Giffard) (BPBM, 1).
Diagnosis. Frons and vertex covered with a mix of isodi-ametric and transverse microsculpture, the sculpticells nearlyflat so that the surface is glossy, not granulate (Fig. 3B); apexof interval 7 nearly flat mesad subapical sinuation (Fig. 8B)owing to shallowness of elytral stria 7, stria 6 nearly as deeplyimpressed at midlength as striae 1–5. The male aedeagus isdistinguished by the extremely large accessory subapical pro-jection, and the expanded ventral surface (Fig. 6F, G). Stan-dardized body length 4.2–4.6 mm
Description. Head capsule with convex frons and deep,broad and smooth frontal grooves; ocular ratio 1.33–1.42, ocu-lar lobe ratio 0.76–0.80. Pronotum transverse, MPW/PL =1.32–1.40; in different individuals, laterobasal margin slightlysinuate, straight, or slightly convex anterad hind angle, in theformer two instances hind angles distinctly obtuse; medianbase covered with large punctures that are longitudinallyextended near the disc, resulting in longitudinal strigae; ante-rior transverse impression smooth, callosity anterad transverseimpression slightly upraised but smooth; pronotal front anglesonly slightly projected, narrowly rounded, the lateral marginaldepression only slightly widened near front, very narrowposterad to position of lateral seta, which is situated alongmesal margin of lateral depression; lateral marginal depres-sion relatively broad and of equal width from midpoint to hindangle. Elytra narrowed basally, the humeri narrowly roundedposterad angular juncture with slightly curved basal groove;striae 1–6 depressed, continuous, striae 1–2 minutely punctateon disc, elytral intervals slightly convex; parascutellar stri-ole present, shallow, two- to four-punctate; parascutellar setapresent; two dorsal elytral setae present, situated near mid-dle of third interval, at about 0.33× and 0.67× elytral length;lateral elytral setae arranged as 7(6)+(1)+6. Tarsomeres andColoration as in M. brevis. Microsculpture on pronotal disc anirregular transverse mesh, with elongate transverse sculpticellsarranged in parallel over lateral portions of the discal sur-face defining parallel lines, pronotal base covered with trans-verse sculpticells; elytral disc with distinct transverse meshmicrosculpture.
Male genitalia (n = 4). Aedeagal median lobe appearingto be an exaggeration of features observed in M. dentatus,especially with regard to the elongate accessory subapicalprojection, marked ventral expansion, and broadened apex(Fig. 6F–H); dorsal plate of internal sac moderate (judgedin uneverted dissections), larger than observed in M. brevis,although smaller than in M. uncinus (Fig. 6G); spiculation ofinternal sac moderate, without any major fields of microtrichial
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 663
spines. Variation in the male aedeagaus is limited to differentialbreadth of the accessory subapical projection (Fig. 6F, G).
Female reproductive tract (n = 1). Bursa copulatrix colum-nar, apex broadly rounded and slightly broader than medianportion of bursa (Fig. 9C), bursa copulatrix length 2.4× gono-coxal length; spermathecal gland duct length short, subequalto spermathecal gland reservoir, gland duct and reservoirassembly shorter than spermathecal duct; basal gonocoxite 1(Fig. 9D) with apical fringe of two to three setae and twomedioapical setae; apical gonocoxite 2 broad basally, narrowedlaterally to acuminate tip, bearing two lateral ensiform setae,one dorsal ensiform seta and two apical nematiform setae.
Distribution. Specimens labelled with a specific localityare known from Tantalus between 550 and 614 m elevation(Fig. 10B).
Habitat. The only specimen labelled with any ecologicalinformation was collected ‘under rocks’.
Etymology. The species epithet is the Latin adjectivelobatus, meaning lobed, denoting the condition of the maleaedeagal median lobe.
Mecyclothorax dentatus sp.n.(Figs 2E; 3B; 6H–J; 9E, F; 10B)
Type material. Holotype male (card-mounted, dissectedmale genitalia in glass genitalia vial): ‘785’ on reverse ofcard/Hawaiian Is. R.C.L. Perkins (yellow-lined label)/SharpColl. 1905-313/brevis/HOLOTYPE Mecyclothorax dentatusJ.K. Liebherr 2008 (red, black-bordered label) (BMNH).Paratypes , HAWAII, Oahu: (Koebele) (BPBM, 5); Honolulu,R.C.L.P. lot no. 710, 1897 (Koebele) (BPBM, 1); High Mts. nrHonolulu, R.C.L.P. Lot no. 884, xii.1900 (Perkins) (ANIC, 1;BMNH, 4; BPBM, 3); Honolulu Mts., 455 m, 1902 (Perkins)(BPBM, 3), i.1903 (Perkins) (BPBM, 1); Honolulu Mts.,R.C.L.P. lot no. 785, viii.1900 (Perkins) (BMNH, 2), R.C.L.P.lot no. 787, xii.1900 (Perkins) (BPBM, 2); Honolulu, Pauoa,455 m (Perkins) (BPBM, 2); Tantalus, 425 m, v.1906 (Giffard)(BPBM, 2).
Diagnosis. This species and M. calceus are distinguishedfrom all other species of this group by the reduced elytralstriae 6 and 7. In M. dentatus stria 6 is continuous althoughshallower than stria 5, and stria 7 is reduced to obsolescence,its presence defined by a series of broad, shallow punctationsindistinctly joined by barely impressed longitudinal impres-sions. The pronotum of M. dentatus is also more quadratethan in all other species (Fig. 2E), with MPW/PL in the range1.25–1.33 (versus a range of 1.32–1.40 in the other species).Mecyclothorax dentatus can be diagnosed from M. calceus bythe smoother elytral striae that irregularly waver along theirlength, but lack distinct punctures. The male aedeagal medianlobe bears a tooth-like accessory subapical projection (Fig. 6J,K), and a broadly expanded apex. Standardized body length4.0–4.6 mm.
Description. Head capsule with indistinctly convex frons,frontal grooves deeper near frontal suture, shallower and lin-ear mesad anterior supraorbital seta; outer surface of eyes asconvex, to more convex than curvature defined by posteriorportion of ocular lobe, ocular ratio 1.38–1.46, ocular lobe ratio0.73–0.84. Pronotum with laterobasal margin straight anteradobtuse hind angle; basal margin convex medially, unmargined,indistinctly concave laterally to meet obtuse hind angle; medianbase covered with large punctures associated with longitudi-nal wrinkles producing strigose appearance; median longitudi-nal impression deepest near anterior portion of median base,finely incised on disc, and terminated at medially obsolete,narrowly incised and shallow anterior transverse impression;pronotal front angles projected and tightly rounded, the lat-eral marginal depression broad enough to be lined with smallisodiametric sculpticells at the front angle, but very narrowfrom just posterad front angle to posterad lateral seta, thenwidened slightly to laterobasal depression; lateral margin nar-row enough so that it is difficult to view proepipleura throughit. Elytra subovoid, the basal groove moderately curved tomeet rounded humeral margin, humerus broadly convex later-ally and posteriorly; striae 1–7 depressed, continuous, slightlyirregular along their length, but without distinct punctures (asin M. brevis); parascutellar striole present, shallow, smooth;parascutellar seta present; two dorsal elytral setae present,situated near middle of third interval, at about one-fourthlength and just behind midlength of elytra; lateral elytral setaearranged as 7(5 − 6) + (1) + 6. Tarsomeres and Coloration asin M. brevis. Microsculpture on frons and vertex consistingof isodiametric and slightly transverse sculpticells arranged intransverse rows, some slightly wrinkled in the centre of frons,but not to the degree observed in M. brevis (Fig. 3A); pronotaldisc covered with regular, elongate transverse mesh, prono-tal base with mix of isodiametric and transverse sculpticells;elytral disc with elongate transverse mesh microsculpture, themesh becoming less regular in the lateral intervals.
Male genitalia (n = 4). Aedeagal median lobe evenlycurved (Fig. 6I) to slightly recurved (Fig. 6H) ventrally, dor-sally expanded as a tooth-like accessory subapical projectionthat is offset to the left relative to apex in ventral view (Fig. 6J);median lobe apex broadened dorsally and ventrally, the tiptightly rounded and expanded ventrally, the dorsal margin api-cad ostium rounded in some individuals (Fig. 6H), acutelyangulate in others (Fig. 6I); aedeagal internal sac with finespicules on ventral surface, dorsal plate of moderate length,lightly sclerotized (Fig. 6H). Aedeagal variation is encom-passed by a larger dorsal tooth associated with a slight ventralexpansion of the median lobe (Fig. 6H), versus a smaller dorsaltooth with no expansion (Fig. 6I).
Female reproductive tract (n = 2). Bursa copulatrixbroadly columnar, of equal breadth throughout length (Fig. 9E),bursa copulatrix length 2.8–3.0× gonocoxal length; sperma-thecal gland duct short, length subequal to spermathecal glandreservoir, gland and reservoir assembly shorter than sperma-thecal duct; basal gonocoxite 1 (Fig. 9F) with apical fringe oftwo to three setae, and two to four medioapical setae (count
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

664 J. K. Liebherr
may vary by two in an individual); apical gonocoxite 2 mod-erately broad basally, narrowed to acuminate tip, bearing twolateral ensiform setae, one dorsal ensiform seta and two apicalnematiform setae.
Distribution. This species is recorded specifically fromTantalus at elevations of 425–455 m, and Pauoa near Tantalusat 455 m elevation (Fig. 10B).
Etymology. The species epithet is the Latin adjectivedentatus, meaning toothed, denoting the condition of the maleaedeagal median lobe.
Mecyclothorax calceus sp.n.(Figs 2F; 6K; 10A)
Type material. Holotype male (card-mounted, dissectedmale genitalia in plastic genitalia vial): T. 1300 1 III 08 (in pen-cil on obverse of card)/W.M.Giffard Collection/Mecyclothoraxbrevis complex male 22 J.K. Liebherr 2008/HOLOTYPEMecyclothorax calceus J.K. Liebherr 2008 (red. black-borderedlabel) (BPBM). Paratypes , card-mounted specimens withoutlabelling on separate cards but on same pin, male dissectionsin plastic genitalia vials labelled 1 and 2 to correspond to num-bered cards (BPBM, 2).
Diagnosis. As in the preceding species, M. calceus haselytral striae 6 and 7 reduced relative to stria 5, although stria7 is better developed than in M. dentatus, being continuouslyimpressed. The discal elytral striae are lined with rudimentarypunctures, and the striae are straight, not wavering between thepunctures. The male aedeagal median lobe is slightly expandeddorsally, although without a distinct projection (Fig. 6K),and the median lobe apex is narrowly elongate and curvedventrad, with a tightly rounded tip. A narrowly elongatemedian lobe apex is shared with M. latissimus of Molokai(Liebherr, 2006b: figs 93; 94), although the apex is longer andmore curved ventrally in M. calceus. Standardized body length4.2–4.4 mm.
Description. Head capsule with slightly convex fronsdelimited laterally by distinct, narrowly incised frontal groovesthat extend posteriorly mesad the anterior supraorbital seta, thedepression of the anterior seta separated from frontal groove bya narrow carina; eyes little projected beyond curvature definedby posterior portion of ocular lobe, ocular ratio 1.40–1.44,ocular lobe ratio 0.81–0.84. Pronotum transverse, MPW/PL= 1.35–1.41; laterobasal margin convex to slightly sinuateanterad obtuse hind angle; median base covered with inter-mittent, shallow punctures interspersed between fine, shallowlongitudinal wrinkles; median longitudinal impression deepestnear anterior of median base, very fine and shallowly incisedon disc; anterior transverse impression obsolete medially, morewell defined near slightly protruded, rounded front angle; areaanterad transverse impression smooth; lateral marginal depres-sion broad enough near front angle to be lined with smallisodiametric sculpticells, narrowed posteriorly to point of lat-eral setal position, then gradually and continuously broadened
to obtuse-rounded hind angle; behind the point of insertionof the lateral seta, lateral marginal depression broad and thinenough to be translucent, thereby allowing dorsal view ofproepipleura. Elytra broad, convex, humeral margin narrowlyrounded laterad juncture with distinctly curved basal groove;parascutellar striole present, shallow, smooth; parascutellar setapresent; two dorsal elytral setae present, situated close to thirdstria, at about 0.3× and 0.6× elytral length; lateral elytral setaearranged as 7 + 6. Tarsomeres and Coloration as in M. brevis.Microsculpture of frons and vertex as in M. dentatus, a mixtureof upraised isodiametric and transverse sculpticells arranged intransverse rows with some wrinkling medially; pronotal discwith elongate transverse mesh microsculpture, more irregularlaterally forming transverse lines, pronotal base with irregularisodiametic mesh associated with punctures and wrinkles; ely-tral disc glossy, with reduced, elongate transverse sculpticellsloosely joined into a mesh.
Male genitalia (n = 3). Although the aedeagal medianlobe does not bear a distinct dorsal projection, the dorsalsurface is expanded relative to the overall diameter of thelobe at approximately the position of the accessory toothobserved in M. dentatus. Mecyclothorax latissimus of Molokailacks any such dorsal expansion, with its median lobe broadestat about midlength. The dorsal expansion in the apical halfof the median lobe of M. calceus can thus be consideredsynapomorphic with the greater development of subapicalprojections in M. brevis, M. lobatus and M. dentatus. Based onuneverted dissections, the internal sac does not bear spicules,and the dorsal plate is of moderate length.
Distribution. Although the card upon which the holotypeis mounted bears a cursory pencil inscription with a locality of‘T.’, and ‘1300’, there can be little doubt that the type localityis Tantalus 1300 ft. This conclusion is definitively supported bylabelling of other specimens, for which Giffard wrote ‘T.’, anelevation and a date on the underside of the mounting card, andthen added two printed labels to the pin bearing: (i) ‘Tantalus’and an elevation in feet; and (ii) his name and the date. Anexample of this convention is shown above for the holotype ofM. lobatus. The two paratypes of M. calceus bear no labellingother than that of ‘W.M. Giffard Collection’, but the genitaliaagree, and they are mounted on similar, although not identical,cards. Thus it is assumed they also came from the type localityof Tantalus, 1300 ft (Fig. 10A).
Giffard’s incomplete labelling may be attributed to thefact that these specimens were among the last Mecyclotho-rax he collected on Tantalus. In 1906 he collected 2 M. brevis,1 M. dentatus and 13 M. lobatus individuals from the moun-tain. During 1907 he collected 27 M. brevis, and in 1908 hecollected 23 M. brevis and the 3 M. calceus. Although he con-tinued to collect on Oahu until 1919, he collected no moreMecyclothorax specimens.
Etymology. The species epithet calceus is the Latin noundenoting shoe or slipper, which refers to the boot-like apex ofthe male aedeagal median lobe.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 665
Mecyclothorax flavomarginatus group
Diagnosis. Species of this group exhibit reduction ofthe elytral striae, with the first or sutural stria completeand deep but the second and more lateral striae obsoleteapically, a synapomorphy shared with the M. scaritoides group.Both groups are also characterized by synapomorphous eyereduction; the ocular ratio of all species in both groupsranging from 1.31 to 1.43, and the ocular lobe ratio small,0.64–0.72. In species of the M. flavomarginatus group, theelytra have striae 1–4, or 1–5, or even 1–6 present discally,although striae 5 and 6 are shallower than those closer tothe suture, symplesiomorphic relative to the more reducedelytral striation in the M. scaritoides group. The glabrouspronotum is subcordate or cordate, with the laterobasal marginsinuate anterad the pronotal hind angle. The pronotal lateralmarginal depression is autapomorphously broad enough so thatthe lateral margin is upturned, and the outer portion of theexplanate lateral margin is translucent. The body size is smallto moderate in beetles of this group; standardized body length3.4–4.7 mm.
This species group is best represented on Oahu, withM. impunctatus Liebherr (2006b) from Molokai, and M. sharpiBritton (1948) from Maui being the only other describedspecies. The group is not known from Hawaii Island (Lieb-herr, 2008a).
Mecyclothorax flavomarginatus Britton(Figs 2G; 6L, M; 9G, H; 10C)
Mecyclothorax flavomarginatus Britton 1948: 128.Cyclothorax oahuensis Blackburn 1878: 123 (in part).Metrothorax oahuensis, Perkins 1917: 250 (misidentification).
Type material. Holotype female (card-mounted): ‘female’symbol on obverse, ‘769’ on reverse of card/Type (round,red-bordered label)/oahuensis/Honolulu Oahu 2000 ft PerkinsVII 1901/Holotype Mecyclothorax flavomarginatus E.B. Brit-ton 1939 (BMNH).
Diagnosis. Instantly recognizable by the broadly flavouselytral margins contrasted with the piceous cloud on eachelytral disc (Fig. 2G), head and pronotum rufous with a piceouscast; pronotum subcordate, the laterobasal margin slightlysinuate anterad the obtuse, rounded hind angle; pronotalbase smooth, although an undulated, discontinuous transversedepression partially defines a basal marginal bead; elytral striae1–7 visible, punctures indistinctly indicated in striae 2–4.Standardized body length 3.8–3.9 mm.
Description. Head narrow, the ocular lobes little pro-jected from gena; eyes not projected from ocular lobes; ocularratio 1.39–1.44, ocular lobe ratio 0.65–0.70; frons broadlyconvex, the frontal grooves broad, rounded in deepest por-tion near frontal suture, terminated just mesad depression ofanterior supraorbital seta; posterior supraorbital seta immedi-ately dorsad dorsal terminus of postocular groove. Pronotum
moderately transverse, MPW/PL = 1.29; median base delim-ited from disc by line of minute punctures; median longitu-dinal impression absent on base, broad and shallow on discwhere it is lined with sculpticells; anterior transverse impres-sion broad, shallow, smooth, delimiting smoothly convex ante-rior callosity; front angles little projected, broadly rounded,the lateral marginal depression broad, translucent from frontto hind angle, only slightly narrowed near midlength; hindangles obtuse, rounded at apex, the laterobasal margin indis-tinctly sinuate anterad angle. Elytra broad, subquadrate, humeribroadly rounded laterad little-curved basal groove; parascutel-lar striole broad, shallow, parascutellar seta present in orlaterad striole; two dorsal elytral setae in subfoveate depres-sions, the anterior depression adjoining stria 3, the posteriorin middle of interval 3; lateral elytral setae arranged as 7 + 6.Tarsomeres short, robust, metatarsomere 4 apical breadth 1.5×median length, apical margin emarginate, but apical lobes short.Coloration of ventral sclerites dark brunneous, femora, tibiaeand tarsi flavous. Microsculpture on frons a well-developedtransverse mesh, sculpticells isodiametric on vertex; pronotaldisc covered with indistinct transverse-line microsculpture, thesculpticells irregularly joined by crosslinks to form an inter-mittent mesh; discal elytral intervals 1–3 glossy at convexcentres between striae, depressed areas associated with striaelined with transverse sculpticells; lateral elytral intervals cov-ered with transverse-line microsculpture loosely joined intoa mesh; elytral apex covered with indistinct transverse-linemicrosculpture.
Male genitalia (n = 1). Aedeagal median lobe graduallynarrowed from basal bulb to tightly rounded apex, ventralsurface very slightly curved before parallel-sided, slightlydownturned apex (Fig. 6L); median lobe curved to right inventral view (Fig. 6M), apical face nearly straight.
Female reproductive tract (n = 1). Bursa copulatrixbroadly rounded apically, broader in basal half (Fig. 9G), bur-sal length 2.9× gonocoxal length; spermathecal gland ductlength subequal to globose reservoir, the gland assembly quiteshort, its length about half length of spermathecal duct andreservoir; basal gonocoxite 1 with apical fringe of two tothree setae, one to two setae present near medioapical margin(Fig. 9H); apical gonocoxite 2 narrow basally, subtriangular,bearing two lateral ensiform setae, one dorsal ensiform seta,and two apical nematiform setae.
Distribution. This species is recorded from Tantalus(BPBM), with the specification ‘makai Government road’,interpreted here to mean a locality below the current Tanta-lus Drive, somewhat to the west of Tantalus, and on the valleyslopes above Kanealole Stream.
Mecyclothorax euryoides sp.n.(Figs 2H; 11A–C; 12A, B; 13A)
Type material. Holotype female: Puu Kalena 19 April1931 Oahu/O. H. Swezey Collector/Cibotium chamissoi/HOLOTYPE Mecyclothorax euryoides J.K. Liebherr 2008(red, black-bordered label) (BPBM). Paratypes , HAWAII,
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

666 J. K. Liebherr
Oahu: Mt. Kaala, 1.ix.1929 (Swezey) (BPBM, 1), 1195 m,pyrethrin fog mossy Metrosideros, 29.vii.2006 (Polhemus)(BPBM, 1), 1210 m, moss, vii.1954 (Ford) (BPBM, 1), beatingMetrosideros/fern/moss, 22.iv.1991 (Liebherr) (CUIC, 2).
Diagnosis. A larger and broader bodied species than theothers in this group, the pronotum broad basally, with latero-basal margin broadly elevated laterad the laterobasal depres-sion, and the elytra broad basally (Fig. 2H), standardized bodylength 4.6–4.7 mm; pronotal base nearly smooth with at mostindistinct, irregularly distributed longitudinal wrinkles; ely-tral striae 1–5 very shallow on disc, striae 2–5 irregularlyexpanded suggesting indistinct punctures; elytral humeri pro-jected anterad, the lateral elytral marginal depression broadestat and immediately posterad subangulate humerus. The nearlyflat elytral intervals are covered with a well-developed elon-gate transverse mesh microsculpture resulting in an indistinctblue iridescence.
Description. Head capsule narrow and elongate, eyessmall and little protruded from ocular lobes, ocular ratio1.36–1.40, ocular lobe ratio 0.67–0.72; frons convex betweenshallow, ill-defined frontal grooves, the grooves widest justposterad the frontal suture where they encompass a shallowpit; anterior supraorbital seta joined posteriorly to a shallowgroove that encompasses the posteriodorsal eye margin; poste-rior supraorbital seta located anteriodorsad the dorsal terminusof postocular groove. Pronotum robust, abacoid, with broad,medially unmargined base that is depressed relative to disc,the base delimited from disc by indistinct longitudinal wrin-kles; median longitudinal impression indistinctly indicated onbase, nearly obsolete on disc, very shallow; anterior transverseimpression very shallow, crossed by faint wrinkles, distinctonly close to nonprotruded front angle; lateral marginal depres-sion narrowest in apical third of length, widened posterad toposition of distinct laterobasal sinuation that defines nearlyright hind angle; laterobasal depression bordered laterally bycallous-like, broadly upraised lateral margin, and basally bythin, lateral marginal bead. Elytra broad, convex, the subangu-late humerus extended forward laterad curved basal groove;parascutellar striole smoothly impressed; parascutellar setapresent laterad striole or in base of stria 1; striae 1–4 evi-dent on disc, stria 1 deepest and smooth, striae 2–3 shallowerand evidently punctate, stria 4 very shallow and nearly smooth;two dorsal elytral setae close together in middle of third inter-val, 0.3× and 0.5× elytral length; lateral elytral setae arrangedas 7 + 6. Tarsomeres elongate, gracile, metatarsomere 4 apicalwidth 1.3× median length, apically emarginate, with apicallobes short, 0.25× median length. Coloration of head andpronotum brunneous with piceous cast; elytra rufobrunneous,sutural interval and lateral elytral margin paler, apex broadlypaler, all rufoflavous; ventral thoracic and abdominal stern-ites rufobrunneous, the abdominals paler laterally; proepipleurapaler, rufoflavous, elytral epipleura and femora bright flavous,tibiae and tarsi flavous with a brunneus cast. Microsculptureon frons consisting of flat isodiametric sculpticells in trans-verse rows, the surface glossy, vertex with more transversely
stretched sculpticells; pronotal disc glossy, covered with shal-low transverse-line microsculpture that is difficult to traceowing to the glossy surface.
Male genitalia (n = 2). Aedeagal median lobe robust,very broad throughout midlength (Fig. 11A), the apex beyondostium parallel-sided and downcurved to a tighty roundedtip; median lobe slightly curved rightwards in ventral view(Fig. 11C); aedeagal internal sac elongate, the basal portionbasad dorsal plate as long as lobe itself (Fig. 11B); inter-nal sac with lightly sclerotized ventral ostial microtrichialpatch, and dorsal ostial microtrichial patch comprising stoutspicules.
Female reproductive tract (n = 1). Bursa copulatrix elon-gate and very broad throughout (Fig. 12A), apex broadlyrounded, bursal length 4.6× gonocoxal length and 2.0× bur-sal breadth (dissection compressed under microslide coverslip); spermathecal gland assembly elongate, the length of ductplus reservoir nearly twice length of spermathecal duct plusreservoir; basal gonocoxite 1 (Fig. 12B) with apical fringe oftwo to four setae, two setae present near medioapical margin;apical gonocoxite 2 broad basally, subtriangular, apex sub-acuminate, and bearing two lateral ensiform setae, one dorsalensiform seta and two apical nematiform setae.
Distribution. Recorded from the Waianae Range, on Mt.Kaala and Puu Kalena (Fig. 13A).
Habitat. All records of this species are associated witharboreal microhabitats. Beetles have been collected in moss,Cibotium chamissoi tree fern, by beating ferns and mossyMetrosideros polymorpha (ohia), and by applying pyrethrin foginsecticide to mossy ohia.
Etymology. The species epithet euryoides is a compound-ing of the Greek stem eury, meaning wide or broad, and theModern Latin suffix—oides, denoting likeness of form (Jaeger,1955).
Mecyclothorax carteri (Perkins)(Figs 4A; 7; 11D–F; 12C, D)
Metrothorax carteri Perkins 1917: 249; Swezey 1931: 423(distribution).Mecyclothorax carteri, Britton 1948: 130.Metrothorax oahuensis (Swezey, 1954: 8) (misidentifica-tion, biology)
Type material. Lectotype female (card-mounted) herebydesignated: ‘Metrothorax carteri. Type. R.C.L.P. Waianae Mts.2000+ ft’ in ink on obverse of card, ‘female’ symbol in pencilin lower left of card, ‘Waianae Mts.’ on reverse of card/1318(yellow label)/LECTOTYPE Metrothorax carteri Perkins J.K.Liebherr 1998 (red, black-bordered label) (BPBM). This lec-totype designation is required because Perkins (1917) did notspecify that the specimen was unique.
Diagnosis. Externally, beetles of this dorsally concolorousspecies are nearly indistinguishable from individuals belonging
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687
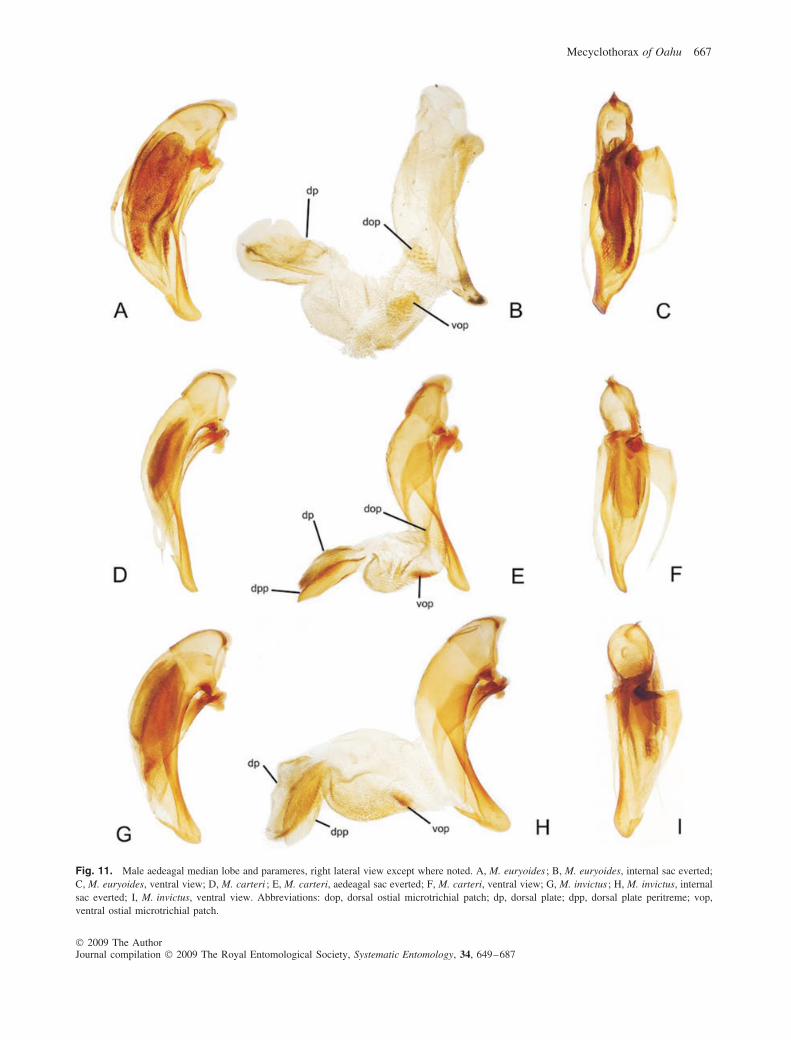
Mecyclothorax of Oahu 667
Fig. 11. Male aedeagal median lobe and parameres, right lateral view except where noted. A, M. euryoides ; B, M. euryoides, internal sac everted;C, M. euryoides, ventral view; D, M. carteri ; E, M. carteri, aedeagal sac everted; F, M. carteri, ventral view; G, M. invictus ; H, M. invictus, internalsac everted; I, M. invictus, ventral view. Abbreviations: dop, dorsal ostial microtrichial patch; dp, dorsal plate; dpp, dorsal plate peritreme; vop,ventral ostial microtrichial patch.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687
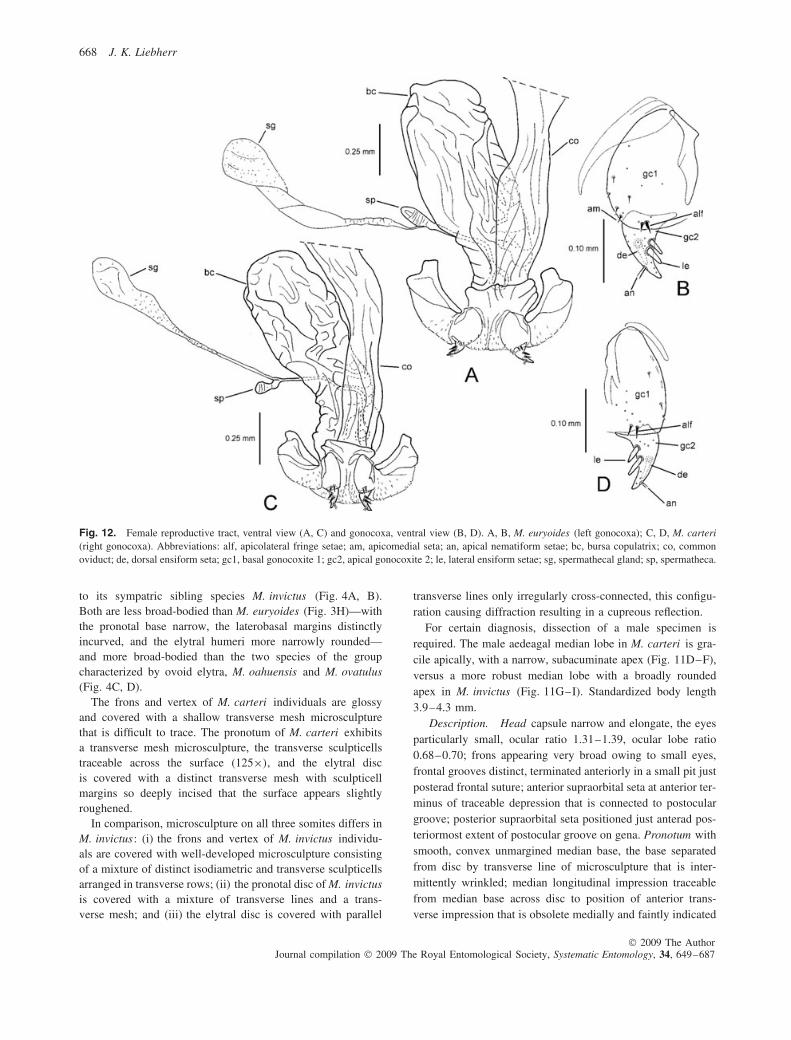
668 J. K. Liebherr
Fig. 12. Female reproductive tract, ventral view (A, C) and gonocoxa, ventral view (B, D). A, B, M. euryoides (left gonocoxa); C, D, M. carteri(right gonocoxa). Abbreviations: alf, apicolateral fringe setae; am, apicomedial seta; an, apical nematiform setae; bc, bursa copulatrix; co, commonoviduct; de, dorsal ensiform seta; gc1, basal gonocoxite 1; gc2, apical gonocoxite 2; le, lateral ensiform setae; sg, spermathecal gland; sp, spermatheca.
to its sympatric sibling species M. invictus (Fig. 4A, B).Both are less broad-bodied than M. euryoides (Fig. 3H)—withthe pronotal base narrow, the laterobasal margins distinctlyincurved, and the elytral humeri more narrowly rounded—and more broad-bodied than the two species of the groupcharacterized by ovoid elytra, M. oahuensis and M. ovatulus(Fig. 4C, D).
The frons and vertex of M. carteri individuals are glossyand covered with a shallow transverse mesh microsculpturethat is difficult to trace. The pronotum of M. carteri exhibitsa transverse mesh microsculpture, the transverse sculpticellstraceable across the surface (125×), and the elytral discis covered with a distinct transverse mesh with sculpticellmargins so deeply incised that the surface appears slightlyroughened.
In comparison, microsculpture on all three somites differs inM. invictus: (i) the frons and vertex of M. invictus individu-als are covered with well-developed microsculpture consistingof a mixture of distinct isodiametric and transverse sculpticellsarranged in transverse rows; (ii) the pronotal disc of M. invictusis covered with a mixture of transverse lines and a trans-verse mesh; and (iii) the elytral disc is covered with parallel
transverse lines only irregularly cross-connected, this configu-ration causing diffraction resulting in a cupreous reflection.
For certain diagnosis, dissection of a male specimen isrequired. The male aedeagal median lobe in M. carteri is gra-cile apically, with a narrow, subacuminate apex (Fig. 11D–F),versus a more robust median lobe with a broadly roundedapex in M. invictus (Fig. 11G–I). Standardized body length3.9–4.3 mm.
Description. Head capsule narrow and elongate, the eyesparticularly small, ocular ratio 1.31–1.39, ocular lobe ratio0.68–0.70; frons appearing very broad owing to small eyes,frontal grooves distinct, terminated anteriorly in a small pit justposterad frontal suture; anterior supraorbital seta at anterior ter-minus of traceable depression that is connected to postoculargroove; posterior supraorbital seta positioned just anterad pos-teriormost extent of postocular groove on gena. Pronotum withsmooth, convex unmargined median base, the base separatedfrom disc by transverse line of microsculpture that is inter-mittently wrinkled; median longitudinal impression traceablefrom median base across disc to position of anterior trans-verse impression that is obsolete medially and faintly indicated
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687
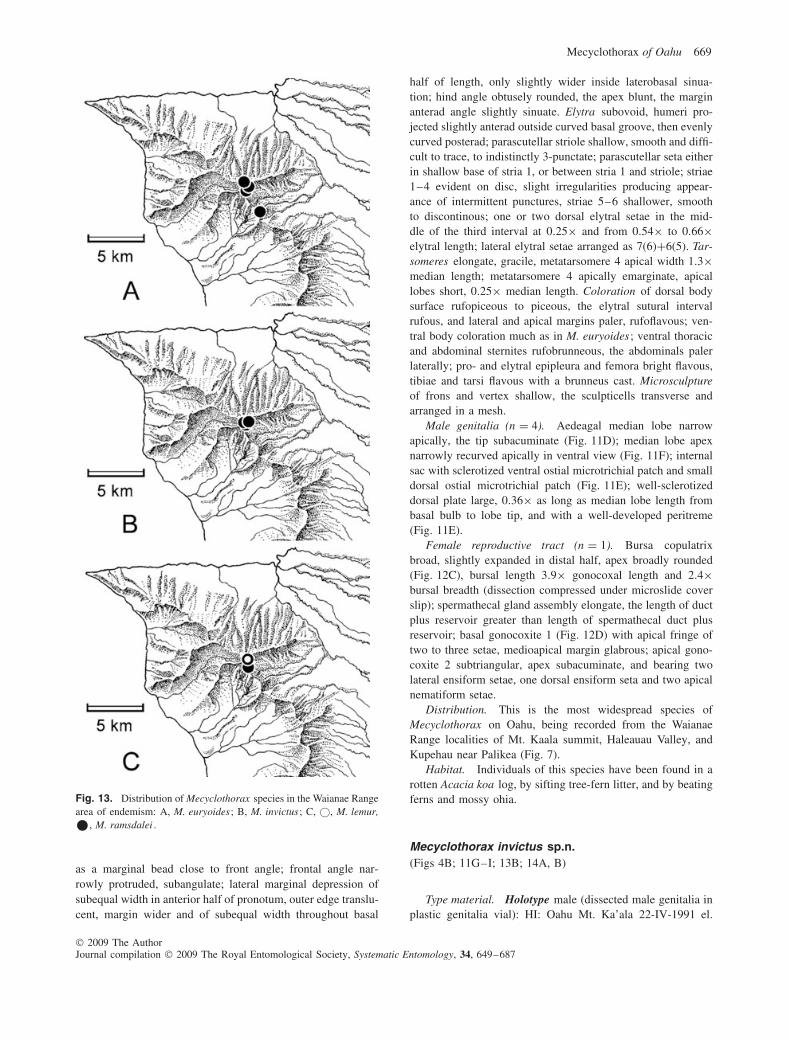
Mecyclothorax of Oahu 669
Fig. 13. Distribution of Mecyclothorax species in the Waianae Rangearea of endemism: A, M. euryoides ; B, M. invictus ; C, ©, M. lemur,•, M. ramsdalei .
as a marginal bead close to front angle; frontal angle nar-rowly protruded, subangulate; lateral marginal depression ofsubequal width in anterior half of pronotum, outer edge translu-cent, margin wider and of subequal width throughout basal
half of length, only slightly wider inside laterobasal sinua-tion; hind angle obtusely rounded, the apex blunt, the marginanterad angle slightly sinuate. Elytra subovoid, humeri pro-jected slightly anterad outside curved basal groove, then evenlycurved posterad; parascutellar striole shallow, smooth and diffi-cult to trace, to indistinctly 3-punctate; parascutellar seta eitherin shallow base of stria 1, or between stria 1 and striole; striae1–4 evident on disc, slight irregularities producing appear-ance of intermittent punctures, striae 5–6 shallower, smoothto discontinous; one or two dorsal elytral setae in the mid-dle of the third interval at 0.25× and from 0.54× to 0.66×elytral length; lateral elytral setae arranged as 7(6)+6(5). Tar-someres elongate, gracile, metatarsomere 4 apical width 1.3×median length; metatarsomere 4 apically emarginate, apicallobes short, 0.25× median length. Coloration of dorsal bodysurface rufopiceous to piceous, the elytral sutural intervalrufous, and lateral and apical margins paler, rufoflavous; ven-tral body coloration much as in M. euryoides; ventral thoracicand abdominal sternites rufobrunneous, the abdominals palerlaterally; pro- and elytral epipleura and femora bright flavous,tibiae and tarsi flavous with a brunneus cast. Microsculptureof frons and vertex shallow, the sculpticells transverse andarranged in a mesh.
Male genitalia (n = 4). Aedeagal median lobe narrowapically, the tip subacuminate (Fig. 11D); median lobe apexnarrowly recurved apically in ventral view (Fig. 11F); internalsac with sclerotized ventral ostial microtrichial patch and smalldorsal ostial microtrichial patch (Fig. 11E); well-sclerotizeddorsal plate large, 0.36× as long as median lobe length frombasal bulb to lobe tip, and with a well-developed peritreme(Fig. 11E).
Female reproductive tract (n = 1). Bursa copulatrixbroad, slightly expanded in distal half, apex broadly rounded(Fig. 12C), bursal length 3.9× gonocoxal length and 2.4×bursal breadth (dissection compressed under microslide coverslip); spermathecal gland assembly elongate, the length of ductplus reservoir greater than length of spermathecal duct plusreservoir; basal gonocoxite 1 (Fig. 12D) with apical fringe oftwo to three setae, medioapical margin glabrous; apical gono-coxite 2 subtriangular, apex subacuminate, and bearing twolateral ensiform setae, one dorsal ensiform seta and two apicalnematiform setae.
Distribution. This is the most widespread species ofMecyclothorax on Oahu, being recorded from the WaianaeRange localities of Mt. Kaala summit, Haleauau Valley, andKupehau near Palikea (Fig. 7).
Habitat. Individuals of this species have been found in arotten Acacia koa log, by sifting tree-fern litter, and by beatingferns and mossy ohia.
Mecyclothorax invictus sp.n.(Figs 4B; 11G–I; 13B; 14A, B)
Type material. Holotype male (dissected male genitalia inplastic genitalia vial): HI: Oahu Mt. Ka’ala 22-IV-1991 el.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

670 J. K. Liebherr
1210 m J.K. Liebherr under bog boardwalk/Mecyclothorax nspcarteri broad ‘male’ #2 J.K. Liebherr 2005/HOLOTYPE Mecy-clothorax invictus J.K. Liebherr 2008 (red, black-borderedlabel) (CUIC). Paratypes , HAWAII, Oahu: Mt. Kaala sum-mit, 1195 m, pyrethrin fog mossy Metrosideros, 29.vii.2006(Polhemus) (BPBM, 1), sifting Cibotium/Metrosideros litter,29.vii.2006 (Liebherr) (CUIC, 4), 1210 m, sifting tree-fern lit-ter, 18.V.1991 (Liebherr) (BPBM, 2; CUIC, 5).
Diagnosis. To diagnose this species from its siblingspecies M. carteri and the other Oahu species of theM. flavomarginatus group, see the M. carteri diagnosis. Stan-dardized body length 3.7–4.4 mm.
Description. The following description presents attributesthat deviate from information in the above description ofM. carteri, and so the M. carteri description can be used forM. invictus, substituting only the following limited informa-tion. Head capsule with eyes variably projected from ocularlobes, ocular ratio 1.33–1.40, ocular lobe ratio 0.67–0.69. Ely-tra consistently with two dorsal elytral setae in the middleof the third interval, the anterior at 0.25× elytral length, andthe posterior just behind midlength of the elytra; elytral setaearranged as 7(6)+(1)+6(5).
Male genitalia (n = 3). Aedeagal median lobe broadbasally to beyond ostium, the apex broadly rounded (Fig. 11G);median lobe broad throughout length in ventral view, notrecurved, but gradually narrowed to a broad apex (Fig. 11I);internal sac elongate, with a small ventral ostial microtrichialpatch, and without a dorsal ostial microtrichial patch (Fig. 11H);dorsal plate very large, 0.4× as long as median lobe frombasal bulb to lobe tip (Fig. 11G, H), and with an evident per-itreme.
Female reproductive tract (n = 1). Bursa copulatrix nar-row, elongate (Fig. 14A), bursal length 4.0× gonocoxallength and 3.6× bursal breadth (dissection compressed undermicroslide cover slip); spermathecal gland assembly elongate,the length of duct plus reservoir 1.5× length of spermathecalduct plus reservoir; basal gonocoxite 1 (Fig. 14B) with api-cal fringe of three setae, medioapical margin glabrous; apicalgonocoxite 2 narrow, apex narrowed, subacuminate, and bear-ing two lateral ensiform setae, one dorsal ensiform seta, andtwo apical nematiform setae.
Distribution. Known from several proximate localities onthe summit plateau of Mt. Kaala (Fig. 13B).
Habitat. Most individuals of this species have been col-lected from ground-level microhabitats, including under a bogboardwalk, and by sifting litter from Cibotium tree ferns andMetrosideros (ohia). One individual was collected by pyrethrinfogging a mossy ohia.
Etymology. The Latin adjective invictus means strong,unconquered, or invincible, an appropriate name for an OahuMecyclothorax species that has persevered until the presentday. Moreover, this species is proposed as the adelphotaxonof M. carteri, named by Perkins (1917) to honour George R.Carter, the second territorial governor, whose surname is basedon the Latin cartero-, meaning strong.
Mecyclothorax oahuensis (Blackburn)(Fig. 4C)
Cyclothorax oahuensis Blackburn 1878: 123.Metrothorax oahuensis, Sharp 1903: 274; Perkins 1917: 250.Mecyclothorax oahuensis, Britton 1948: 130.
Type material. Lectotype female (card-mounted) herebydesignated: card with two-lined Blackburn Oahu code (Zim-merman, 1957), and ‘female’ symbol in pencil on obverse,‘oahuensis’ on reverse/Type (round, red-bordered label)/Hawaiian Is. Rev.T. Blackburn. 1888–1930./LECTOTYPECyclothorax oahuensis Blackburn J.K. Liebherr 1998 (red,black-bordered label) (BMNH). This lectotype is requiredbecause specimens labelled as M. oahuensis in Blackburn’shand also include a specimen of M. flavomarginatus (SAMA),and the holotype of M. ovatulus (BMNH). The designation alsoends confusion regarding the primary type specimen, as (Brit-ton 1948: 130) apparently mistook the specimen for a male,and mistakenly designated it a holotype.
Diagnosis. Among species of the M. flavomarginatusgroup, M. oahuensis is distinguished by the absence of dis-cernible microsculpture on the elytra; the discal intervalsglossy. The pronotum has a narrow base, and the elytra areovoid with narrow, rounded humeri (Fig. 4C). The discal por-tions of striae 1–4 are distinctly punctate. As in M. ovatulus,this species is characterized by small individuals; standardizedbody length 3.4 mm.
Description. Head capsule narrow, appearing subtrian-gular owing to the reduced eyes, ocular ratio 1.34, ocularlobe ratio 0.62; frontal grooves broadly depressing lateral por-tions of frons, terminated posteriorly in deep depression ofanterior supraorbital seta; posterior supraorbital seta in broadshallow depression dorsad postocular groove. Pronotum sub-cordate, moderately transverse, MPW/PL = 1.24; basal mar-gin nearly evenly convex between bluntly obtuse hind angles;median base medially convex, sparsely covered with distinct,isolated punctures that are closest where base meets disc;median longitudinal impression deepest near anterior marginof base, finely incised on disc; anterior transverse impressionsmooth, shallow, broad, delimiting smoothly convex anteriorcallosity; front angles slightly protruded, tightly rounded; lat-eral marginal depression of subequal width throughout anterior0.66× of pronotal length, margin upturned, evenly broadenedand increasingly translucent to laterobasal sinuation; latero-basal margins parallel for short distance anterad hind angles,then distinctly divergent anterad. Elytra narrowed basally, thehumeri subangulate laterad distinctly curved basal groove;parascutellar striole markedly incised basally where it is con-tinuous with basal groove; parascutellar seta present in base ofstria 1; striae 1–4 deep and distinctly punctate on disc, striae5–6 shallower but still with evident punctulae; stria 2 trace-able to apex, but very shallow, smooth; two dorsal elytral setaein subfoveate depressions set in the middle of third interval, at0.28× and 0.50× elytral length; lateral elytral setae arranged as
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687
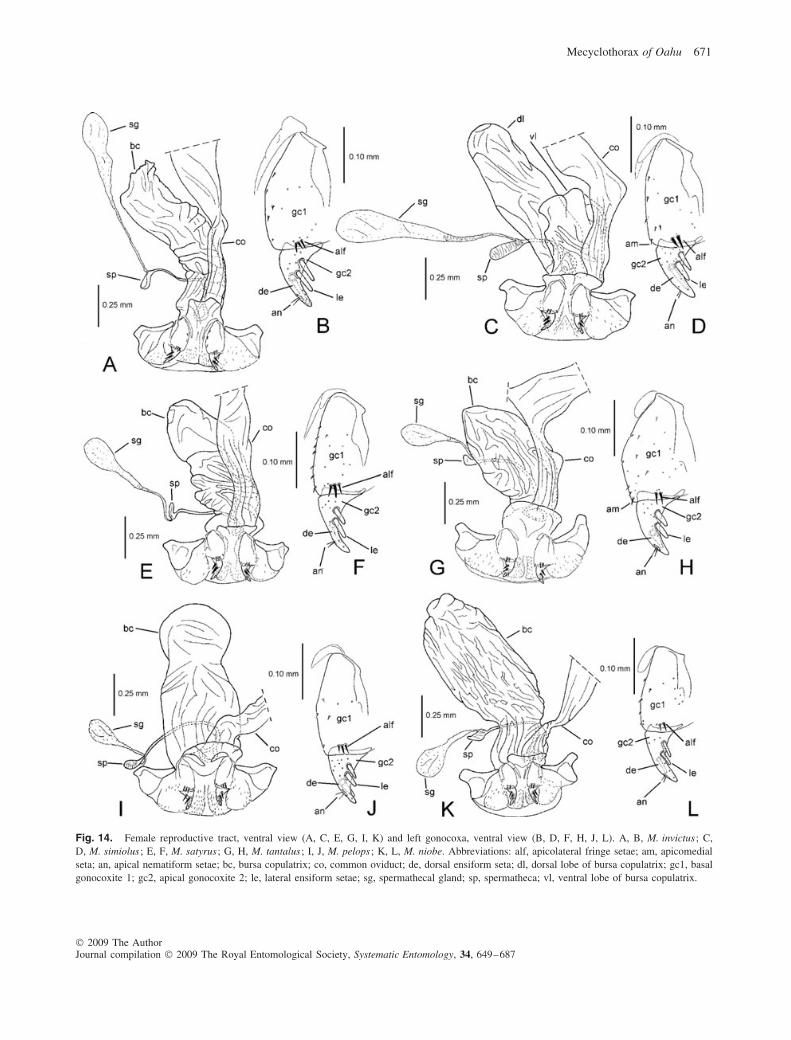
Mecyclothorax of Oahu 671
Fig. 14. Female reproductive tract, ventral view (A, C, E, G, I, K) and left gonocoxa, ventral view (B, D, F, H, J, L). A, B, M. invictus ; C,D, M. simiolus ; E, F, M. satyrus ; G, H, M. tantalus ; I, J, M. pelops ; K, L, M. niobe. Abbreviations: alf, apicolateral fringe setae; am, apicomedialseta; an, apical nematiform setae; bc, bursa copulatrix; co, common oviduct; de, dorsal ensiform seta; dl, dorsal lobe of bursa copulatrix; gc1, basalgonocoxite 1; gc2, apical gonocoxite 2; le, lateral ensiform setae; sg, spermathecal gland; sp, spermatheca; vl, ventral lobe of bursa copulatrix.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

672 J. K. Liebherr
6 + 6. Tarsomeres short, robust, metatarsomere 4 apical width1.6× median length; metatarsomere 4 appearing straight in dor-sal view, the minimal emargination obscured from view bymetatarsomere 5. Coloration of head, pronotal disc, and baseof elytral disc concolorous, brunneous; apex of elytral discwith diffusely darker piceous cloud; elytral marginal depres-sion and interval 8 flavous along elytral sides, elytral apexbroadly flavous, suture (stria 1) paler, rufoflavous. Microsculp-ture on frons of indistinct, transversely stretched sculpticells,the microsculpture traceable on the glossy surface, vertex withmore evident mix of transverse and isodiametric sculpticells;pronotal disc glossy, with transverse-line microsculpture inter-mittently traceable on the glossy surface.
Female reproductive tract. The lone female lectotypespecimen was not dissected.
Distribution. Based on Blackburn’s two-line symbol onthe mounting card, this species was collected on Oahu,although not in the Waianae Mountains, as he labelled speci-mens from that range using two parallel lines crossed by twooblique slash marks (Zimmerman, 1957). Thus by eliminationand Blackburn’s description of his usual collecting activitieson Oahu (Blackburn, 1885), the type locality of this species isdesignated as the south Koolau Range (Fig. 1).
Mecyclothorax ovatulus sp.n.(Fig. 4D)
Cyclothorax oahuensis Blackburn 1878: 123 (in part).
Type material. Holotype female (card-mounted): card withBlackburn’s two-line Oahu code (Zimmerman, 1957), and‘female’ symbol in pencil on obverse, ‘oahuensis’ on reverse/Paratype (round yellow-bordered label)/Hawaiian Is. Rev.T.Blackburn. 1888–1930. (yellow-lined label)/Mecyclothoraxcarteri Perkins E.B. Britton. det. 1945/HOLOTYPE Mecy-clothorax ovatulus J.K. Liebherr 2008 (red, black-borderedlabel) (BMNH).
Diagnosis. In overall appearance much like M. oahuensis,with small-sized body—standardized body length 3.4 mm—and narrow pronotal base and narrow, rounded humeri(Fig. 4D), but distinguishable by the presence of dense, trans-verse elytral microsculpture consisting of elongate sculpticellsarranged in parallel lines, resulting in a silvery iridescence.Elytral striae 1–4 are very broad and shallow, with the onlyindication of puncturation being slight irregularities in directionalong the strial length. The pronotal hind angles are defined byrounded expansions of the lateral margin, and are not as welldeveloped as the distinct, obtuse hind angles of M. oahuensis.
Description. Head capsule appearing broad owing to thereduced, flattened eyes, ocular ratio deceptively large at 1.39because internal eye margins converge on frons, ocular loberatio 0.71; frontal margins broad anteriorly, lined with iso-diametric sculpticells, divergent laterally mesad supraorbitalsetae and posteriorly approaching postocular groove; posterior
supraorbital seta in broad depression lined with isodiamet-ric microsculpture. Pronotum subcordate, moderately trans-verse, ratio of maximum pronotal width to median prono-tal length 1.24; basal margin evenly curved between apicallyrounded hind angles; base medially smooth, with nine to tendistinct punctures laterally bordering laterobasal depression;median longitudinal impression shallowly and finely incisedon disc; anterior transverse impression obsolete medially, deepand smooth laterally, defining smoothly convex anterior cal-losity; front angles not protruded to slightly protruded (uni-laterally varying); marginal lateral depression broad enoughto be translucent posterad front angle, slightly narrower tomidlength, and broader, translucent along the margin to shal-lowly excavated laterobasal sinuation. Elytral humeri angu-late laterad distinctly curved basal groove; parascutellar stri-ole distinctly impressed, continuous with basal groove, indis-tinctly 4-punctate; striae 1–6 shallowly and broadly depressed,lined with transverse miscrosculpture; two dorsal elytral setaein depressions spanning half the width of third interval, at0.25× and 0.56× elytral length; lateral elytral setae arrangedas 5 + 6. Tarsomeres short, robust, apical width of metatar-somere 4 about 1.7× median length; metatarsomere 4 shal-lowly emarginate apically, ventral excavation obscured in dor-sal view by metatarsomere 5. Coloration of head and pronotaldisc brunneous; elytral disc darker, rufopiceous; elytral lateralmargins narrowly flavous, elytral apex slightly more broadlyflavous, suture rufoflavous. Microsculpture indistinct on centreof frons, an intermittently traceable transverse mesh contrastedwith distinct isodiametric sculpticells in frontal grooves; prono-tal disc glossy, transverse-line microsculpture difficult to tracein the direct reflection from fibre-optic wand light sources.
Female reproductive tract. The lone female holotype spec-imen was not dissected.
Distribution. For the same reasons as specified forM. oahuensis, the holotype of this species is assumed to havebeen collected in the south Koolau Range.
Etymology. The species epithet ovatulus compounds theLatin ovat-, or egg-shaped, ovoid, with the adjectival suffix-ulus, denoting a tendency. The name signifies the ovatepronotum with its nearly rounded laterobasal margin.
Mecyclothorax scaritoides group
Diagnosis. Species of this group exhibit extreme reductionof elytral striation, with the first or sutural stria being the onlystria present on the elytral apex, and also the deepest stria in thebasal half of the elytra (Britton, 1948). However, this derivedstate may not define a monophyletic group, as other body-shapecharacters are incongruent with extreme strial reduction. Forexample, some M. scaritoides group species exhibit markedlyconvex upper body surfaces, with the very narrow pronotalmargins greatly depressed relative to the pronotal disc, and theelytral disc inflated enough in several species—for instanceM. simiolus (Fig. 4F)—that the apicolateral elytral marginsare obscured from dorsal view. Other species are less dor-sally inflated; for example M. lemur (Fig. 4E) or M. niobe(Fig. 4K). Among Molokai species, dorsally inflated species
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 673
occur in both the M. scaritoides and the M. ducalis groups(Liebherr, 2006b). For the Oahu species herein assigned to thisgroup based on elytral striation, M. ramsdalei is much moreflattened dorsoventrally (Fig. 4H), although this species sharessynapomorphously rounded pronotal hind angles with the moreconvex M. tantalus, M. pelops and M. niobe (Fig. 4H–K).
Described species characterized by the extreme reductionof elytral striation used to define this group otherwise occurin the windward forests of Molokai (three species), Maui(four species) and Hawaii Island (six species) (Britton, 1948;Liebherr, 2006b, 2008a).
Mecyclothorax lemur sp.n.(Figs 4E; 13C)
Type material. Holotype female: HI: Oahu Mt. Ka’ala22-IV-1991 1210 m el. J.K. Liebherr under bog boardwalk/HOLOTYPE Mecyclothorax lemur J.K. Liebherr 2008 (red,black-bordered label) (CUIC).
Diagnosis. Beetles of large body size, standardized bodylength 4.8 mm, with the pronotum uniquely characterized bya narrow base and extremely elongate laterobasal sinuation(Fig. 4E). Mecyclothorax simiolus is the most similar species inthe group, although the pronotum of M. simiolus is much lesscordate (Fig. 4F), and the beetles are smaller, with standardizedbody length 4.0–4.6 mm. The elytra of M. lemur are coveredwith a regular, transverse-mesh microsculpture that may belost in the reflection of incident light on the glossy surface, buteasily viewed outside the area of direct reflection.
Description. Head with broadly convex frons and ver-tex, frontal grooves sinuously recurved from frontal sutureto posterior terminus mesad anterior supraorbital seta, thegrooves mesally bordering lateral bulbous convexity; posteriorsupraorbital seta in shallow depression lined with indistinctmicrosculpture; eye not projected from curvature of ocularlobe, dorsal surface of eye slightly elevated relative to itsdorsal margin, ocular ratio 1.41, ocular lobe ratio 0.70. Prono-tum median base distinctly punctate, 20–25 punctures eachside of midline, basal margin nearly straight between sharp,slightly obtuse hind angles, the margin slightly expanded medi-ally; median longitudinal impression basally connected to alongitudinal puncture, very finely incised on disc; anteriortransverse impression obsolete, evident as a marginal beadonly immediately mesad slightly projected, tightly roundedfront angle; lateral marginal depression narrow throughoutlength, raised into a broad bead at laterobasal sinuation, themarginal bead continued for a short distance mesad hind angle.Elytra ovoid, basal groove distinctly curved to meet angu-late humerus; lateral margin sloping away abruptly lateradhumerus; parascutellar striole discontinuous, basally evidentnear basal groove, 2-punctate apically; parascutellar seta sit-uated mesad base of sutural stria 1; stria 1 represented byisolated punctures near base, continuous and punctate on disc,smooth and continous in apical third of length; striae 2–4extremely shallow, represented by shallow isolated punctures
on disc; two dorsal elytral setae in middle of flat interval3, at 0.25× and 0.45× of elytral length; lateral elytral setaearranged as 5 + 5. Tarsomeres gracile, metatarsomere 4 apicalwidth subequal to median length; metatarsomere 4 moderatelyemarginate apically, length of the longer outer apical lobe 0.5×tarsomere median length. Coloration of dorsal body surfacepiceous, elytral lateral marginal depression and sutural intervalslightly paler, rufopiceous; thoracic and abdominal ventritesrufopiceous to piceous, pro- and elytral epipleura, and femoraflavous, tibiae and tarsi with brunneous cast. Microsculpture onfrons a traceable transverse mesh interspersed with transversewrinkles, vertex with more isodiametric mesh; pronotal disccovered with evident transverse mesh microsculpture, sculpti-cell breadth 2–3× length.
Female reproductive tract. The lone female holotype spec-imen was not dissected.
Distribution. Known only from the summit bog of Mt.Kaala (Fig. 13C).
Habitat. The holotype was collected under a bog board-walk pressed into the bog surface, sympatric and synchronicin time and space with one specimen of M. carteri.
Etymology. The Latin noun lemur denotes a ghost of thedeparted, now applied to a group of monkey-like animals(Brown, 1956), and evokes Blackburn’s adjectival ‘simiolus’,meaning a small male ape.
Mecyclothorax simiolus (Blackburn)(Figs 4F; 14C, D; 15A–C; 16A)
Cyclothorax simiolus Blackburn 1878: 123.Metrothorax simiolus, Sharp 1903: 274; Perkins 1906: 48(biology).Mecyclothorax simiolus, Britton 1948: 141.
Type material. Lectotype male (card-mounted): Black-burn two-lined Oahu code (Zimmerman, 1957) on obverse,‘simiolus’ on reverse/Type (round, red-bordered label)/LECTOTYPE Cyclothorax simiolus Blackburn J.K. Liebherr1998 (red, black-bordered label) (BMNH). Designating theBMNH lectotype and comparing it with a limited numberof specimens from Bishop Museum allowed transport of thespecies concept without the associated loan of the primary typespecimen.
Diagnosis. These beetles are characterized by a pronotumwith brief, abrupt laterobasal sinuations (Fig. 4F), the lateralmargins anterad the nearly right hind angles parallel for onlya short distance before the hind angles. The pronotal baseis moderately constricted, the ratio of maximum pronotalwidth to basal pronotal width 1.52–1.54. The pronotal lateralmarginal depression is very narrow in the apical half, andthe pronotal disc is broadly convex. The elytral disc isbroadly convex and inflated, so that in dorsal view thelateroapical and apical margins are obscured by the inflateddisc. Contra the elytral microsculpture of the most similar
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

674 J. K. Liebherr
species M. lemur, the discal elytral intervals of M. simiolusare completely glossy and without evident microsculpture, theonly indication of elytral microsculpture limited to indistincttransverse sculpticells lining the strial punctures. Standardizedbody length 4.0–4.6 mm.
Description. Head much like M. lemur, sinuously recurvedfrontal grooves terminated posteriorly mesad anterior supraor-bital setal depression, grooves mesally defining a lateral bul-bous convexity; posterior supraorbital seta in dorsal terminus ofpostocular groove; eyes and ocular lobe somewhat projected,ocular ratio 1.40–1.43, ocular lobe ratio 0.71–0.77. Prono-tum subcordate, broadly convex, base sparsely punctate, aboutten isolated punctures each side of midline; median longitu-dinal impression shallow and finely incised on disc; anteriortransverse impression narrowly obsolete medially, evidencedlaterally as finely incised margin to anterior callosity; frontangles not protruded, rounded, with lateral marginal depressionpresent immediately posterad angle, but completely obsoletebehind that to laterobasal sinuation, the lateral margin a raisedbead for much of its length; lateral margin at hind angle a broadbead that continues onto lateral reaches of pronotal base. Elytrabroadly rounded basally, basal groove curved to very obtuselymeet humeral margin; parascutellar striole shallow, discontin-uous, with 4–5 punctures along length; parascutellar seta inbase of sutural stria 1; striae 1–4 punctate on disc, stria 1continuously depressed with larger punctures, striae 2–4 withpunctures increasingly more isolated; only stria 2 continuousto apex, the other striae absent there; two dorsal elytral setaein depressions spanning half of elytral interval 3, at 0.25× andabout 0.60× elytral length; lateral elytral setae arranged as 6 +6. Tarsomeres short and robust, metatarsomere 4 apical width1.4× median length; metatarsomere 4 slightly emarginate api-cally, the ventral emargination obscured in dorsal view bytarsomere 5. Coloration brunneous over dorsal body surface,only the elytral sutural interval and lateral marginal depres-sion slightly paler, rufous and rufoflavous, respectively; ventralbody coloration as in M. lemur. Microsculpture obscure toabsent on frons and vertex, the only evidence being indistinctsculpticells in the frontal grooves; pronotal disc with broadtransverse irregularities and intermittently traceable transverse-line microsculpture the only interruptions to the glossy surface.
Male genitalia (n = 7). Aedeagal median lobe broadbasally, narrowed at ostium to an acuminate tip, the apicalface straight dorsad tip (Fig. 15A); median lobe evenly curvedrightwards to apex in ventral view (Fig. 15C); median lobeinternal sac with dorsal ostial microtrichial patch composed ofstout spike-like microtrichia (Fig. 15B); well-sclerotized dorsalplate moderately sized, 0.32× as long as median lobe measuredfrom basal bulb to tip, and with well-sclerotized peritreme.
Female reproductive tract (n = 2). Bursa copulatrixbilobed, a shorter ventral lobe that is 2.0–2.1× gonocoxallength and a longer dorsal lobe that is 3.6–3.8× gonocoxallength (Fig. 14C), ventral lobe slightly broader and stainedmore darkly with chlorazol black than dorsal lobe; sperma-thecal gland assembly elongate, length of duct plus reservoir1.6× length of spermathecal duct plus reservoir; basal gono-coxite 1 (Fig. 14D) with apical fringe of two or three setae,
a single small seta at medioapical margin; apical gonocoxite2 broad basally, evenly narrowed to subacuminate apex, andbearing two lateral ensiform setae, one dorsal ensiform seta,and two apical nematiform setae.
Distribution. This is the most repeatedly collected speciesof Oahu Mecyclothorax, and it is known from the south KoolauRange localities of Tantalus, Pauoa Valley, Konahuanui, Paloloand above Niu Valley (Fig. 16A).
Habitat. One specimen was collected on Tantalus bysifting leaves. Perkins collected one specimen 2–5.xi.1892 aslot 38 (Anonymous, N.D.), which corresponds to the followingentry in his collecting journal: ‘Went up Pauoa Valley on toKonahuanui. . . . Nothing of note except a Cyclothorax in stemof tree fern . . . (Perkins, 1892: 7)’. Perkins subsequently wroteof collecting on Tantalus: ‘Thriscothorax brevis is not rare. . . and Metrothorax [=Mecyclothorax ] simiolus is sometimesfound with it, but more often in cavities of tree ferns or rottingwood (Perkins, 1906: 48)’.
Mecyclothorax satyrus sp.n.(Figs 4G; 5; 14E, F; 15D–F)
Type material. Holotype male (card-mounted): ‘778.’ onreverse of card/Wahiawa IV 1901/Hawaiian Is. R.C.L. Perkins.(yellow-lined label)/Sharp Coll. 1905-313/HOLOTYPE Mecy-clothorax satyrus J.K. Liebherr 2008 (red, black-borderedlabel) (BMNH). Paratypes , HAWAII, Oahu: Castle Tr., 665 m,moss on trees, 03.iv.1938 (Zimmerman) (BPBM, 1), Kalu-anui, 760 m, 03.iv.1938 (Valentine) (BPBM, 1); Kaipapau,head of valley, under moss, 12.ix.1939 (Anderson) (BPBM,1); Kaluanui Str. head, 850 m, ohia w/moss, pyrethrin fog,21.x.2002 (Polhemus) (NMNH, 1); Poamoho Tr., ix.1958(Ford) (BPBM, 1), 750 m, sifting ground litter, 2.vi.1977(Howarth) (BPBM, 1), 700 m, on trail at night, 5.v.1995 (Lieb-herr) (CUIC, 1), 760 m, beating vegetation at night, 5.v.1995(Asquith) (CUIC, 1); Puu Kaaumakua, 760 m, 28.iv.1938(Valentine) (BPBM, 3), beating vegetation, 6–7.v.1996 (Pol-hemus/Asquith/Ewing) (CUIC, 1); Puu Ohulehule, 8.vi.1924(Swezey) (HDAC, 1); Schofield Waikane Tr., 760 m, moss,28.iv.1938 (Zimmerman) (BPBM, 1).
Diagnosis. A species of glossy upper body surface asis M. simiolus, but generally of smaller stature, standardizedbody length 3.7–4.6 mm and with a more constricted pronotalbase (Fig. 4G); the ratio of maximum pronotal width tominimum basal pronotal width measured just anterad hindangles 1.60–1.65. The elytral humeri are also narrower than inM. simiolus, with the short elytral basal groove abruptly joinedto the broadened elytral lateral depression at the subangulatehumerus. The eyes of M. satyrus individuals are also smallerand flatter, with the ocular ratio 1.37–1.38, and ocular loberatio 0.66.
Description. Head capsule broad across convexly pro-jected ocular lobes, eyes small, outer surface defining samecurvature as posterior portion of ocular lobe; frontal grooves
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687
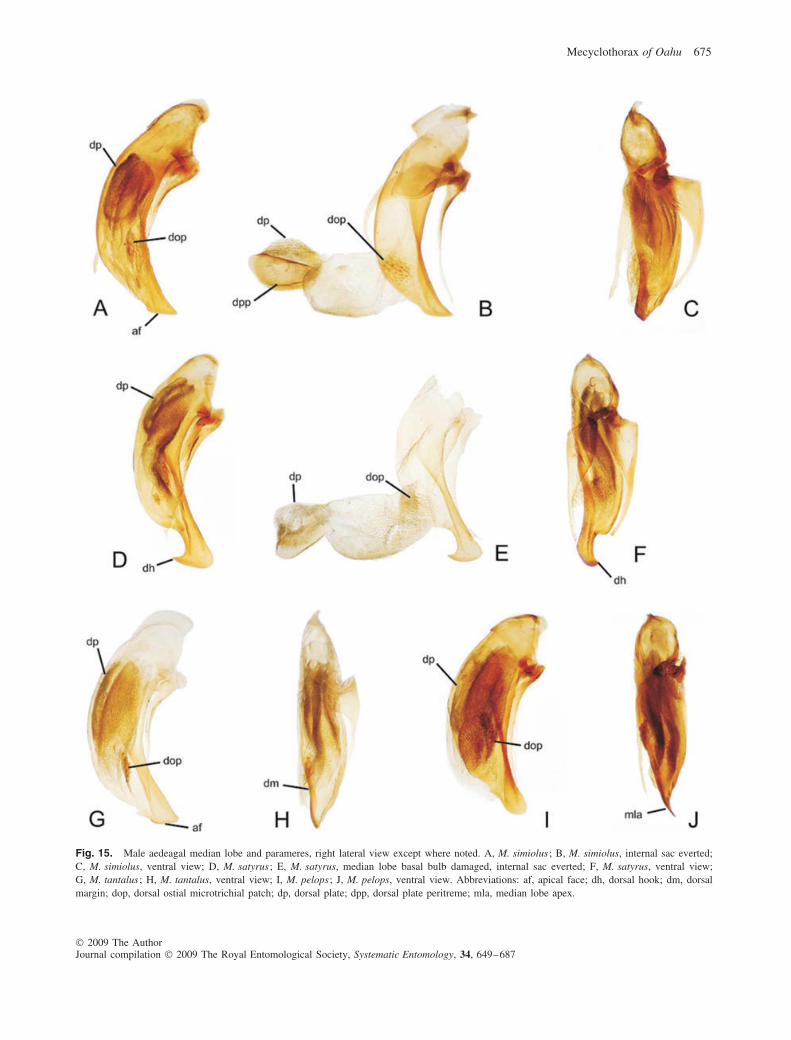
Mecyclothorax of Oahu 675
Fig. 15. Male aedeagal median lobe and parameres, right lateral view except where noted. A, M. simiolus ; B, M. simiolus, internal sac everted;C, M. simiolus, ventral view; D, M. satyrus ; E, M. satyrus, median lobe basal bulb damaged, internal sac everted; F, M. satyrus, ventral view;G, M. tantalus ; H, M. tantalus, ventral view; I, M. pelops ; J, M. pelops, ventral view. Abbreviations: af, apical face; dh, dorsal hook; dm, dorsalmargin; dop, dorsal ostial microtrichial patch; dp, dorsal plate; dpp, dorsal plate peritreme; mla, median lobe apex.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

676 J. K. Liebherr
Fig. 16. Distribution of Mecyclothorax species in the south KoolauRange area of endemism: A, M. simiolus ; B, M. tantalus .
deep, finely incised, sinuously terminated immediately mesadanterior supraorbital seta, separated from setal depression byfine carina; posterior supraorbital seta in dorsal terminus ofpostocular groove. Pronotum cordate; base smooth medially,sparsely punctate laterally, approximately 15 isolated puncturesmesad deep, pit-like laterobasal depression; median longitudi-nal impression posteriorly terminated in small lenticular punc-ture on base, finely incised, shallow on disc; anterior transverseimpression shallow and broad medially, more finely incisedlaterally, defining a marginal bead just mesad finely rounded,non-protruded front angle; lateral marginal depression slightlybroader just posterad front angle, extremely narrow posteradto midlength, only slightly broader to laterobasal sinuation;marginal bead little upraised outside laterobasal depression.Elytra narrow basally, subangulate humerus indicated by junc-ture of nearly straight and finely incised basal groove andexpanded elytral lateral depression; parascutellar striole shal-low and broad, 2–3-punctate; parascutellar seta present justmesad base of sutural stria 1; striae 1–4 punctate on disc, stria1 deepest, striae 2–3 somewhat shallower, and punctures ofstria 4 nearly isolated by cuticle coplanar with intervals; elytralstriae 5–6 traceable by shallow, isolated punctures; all striaesave stria 1 absent from elytral apex; two dorsal elytral setaein minute depressions near the middle of interval 3, at 0.25×and 0.55–0.60× elytral length; lateral elytral setae arranged
as 6 + 6. Tarsomeres short and robust, metatarsomere 4 api-cal width 1.5× median length; metatarsomere 4 apical marginemarginate. Coloration of dorsal body surface brunneous, ely-tral lateral margins narrowly rufoflavous, apex more broadlyflavous, suture hardly paler, rufous; ventral body surface as inM. lemur. Microsculpture obscure on frons and vertex, trace-able over portions of frontal grooves; pronotal disc glossy, veryfine parallel lines visible over portions of surface; elytral discglossy, microsculpture absent.
Male genitalia (n = 3). Aedeagal median lobe broad toostium, as in M. simiolus, but terminated apically in a verylarge dorsal hook (Fig. 15D); in ventral view, the left mar-gin of median lobe abruptly constricted apicad ostium, thehooked tip recurved to the left distad the apical constriction(Fig. 15F); internal sac with dorsal ostial microtrichial patch;dorsal plate of moderate size, well sclerotized (Fig. 15D, E)(sclerotization difficult to judge in single everted and over-cleared dissection).
Female reproductive tract (n = 1). Bursa copulatrix elon-gate, moderately broad throughout (Fig. 14E), bursal length3.6× gonocoxal length and 2.7× bursal breadth; sperma-thecal gland assembly moderate, the length of reservoir plusduct subequal to length of spermathecal duct and reservoir;basal gonocoxite 1 (Fig. 14F) with apical fringe of three setae,medioapical margin with seta near apex, but angled marginglabrous; apical gonocoxite 2 (Fig. 14F) moderately broadbasally, narrowed in apical third of length to acuminate apex,and bearing two lateral ensiform setae, one dorsal ensiformseta, and two apical nematiform setae.
Distribution. Known from the north Koolau Range locali-ties of the Castle Trail in Kaluanui Valley, Kaipapau Valley, thePoamoho Trail summit, Puu Ohulehule, the Schofield–Waikanetrail summit, and Puu Kaaumakua (Fig. 5). The type locality—Perkins’ ‘Wahiawa’—is interpreted as the crest of the KoolauRange uphill from Wahiawa, probably along the present-daySchofield–Waikane Trail.
Habitat. Individuals of this species have been foundon the ground and in semiarboreal microhabitats associatedwith moss. Ground-level records involve sifting ground litter,looking under moss on the ground, and collecting an individualwalking on a trail at night. Other beetles have been collected bybeating vegetation during daytime and nighttime, by searchingin moss on trees, and by applying pyrethrin fog insecticide toohia trees.
Etymology. As with the species epithets of the previoustwo species, the Latin noun satyrus denotes a kind of ape or awoodland deity (Brown, 1956).
Mecyclothorax ramsdalei sp.n.(Figs 4H; 13C)
Type material. Holotype female: Hawaiian Is.: Oahu I.Waianae Mts, Mt. Ka’ala N.A.R. 21◦30.333′N 158◦0.8.773′WA.S. Ramsdale leg. 13 March 2005 11 am ex litter & moss(sift & berlese)/HOLOTYPE Mecyclothorax ramsdalei J.K.Liebherr 2008 (red, black-bordered label) (BPBM).
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 677
Diagnosis. A species characterized by the flatter bodyshape of the M. flavomarginatus group, but sharing the reducedelytral striation and more rounded pronotal shape with the fol-lowing three species. The pronotum is transverse, MPW/PL =1.36, with the lateral marginal depression narrower and opaquein the anterior half and broader and marginally translucentnearer the obtuse-rounded hind angles. The sutural stria 1 isdeep, smooth and continuous from the parascutellar seta to theelytral apex. Striae 2–3 are nearly as deep near the elytralmidlength, much shallower in the basal half, and only stria2 is shallowly traceable near the elytral apex. This species ismost similar overall to M. pelops (Fig. 4J), but that species ischaracterized by a narrower body, and punctate discal elytralstriae 1–3. Standardized body length 3.8 mm.
Description. Head broad, frons broadly convex, eyessmall; anterior supraorbital seta at apex of equilateral triangulardepression extended to eye; ocular ratio 1.36, ocular lobe ratio0.68; frontal grooves nearly straight, narrowly incised, joinedto frontal suture at pit, extended posterad anterior supraorbitalseta; posterior supraorbital seta dorsad posterior terminus ofpostocular groove. Pronotum with broadly obtuse hind angles,the margin rounded at apex of the angle and laterobasal marginonly indistinctly sinuate anterad angle; median base indistinctlylongitudinally wrinkled medially, with four to five indistinctpunctures each side set in irregularly wrinkled surface; medianlongitudinal impression distinct on base, finely incised on disc;anterior transverse impression distinct throughout, althoughshallow medially, defining marginal bead laterally that termi-nates at slightly protruded, rounded front angle. Elytra broadbasally, basal groove straight laterad scutellum to base of stria2, then curved evenly anterad to broadly rounded humerus;parascutellar striole shallow, continous, with single puncturenear apex; parascutellar seta present immediately mesad baseof sutural stria 1; striae 1–6 evident to traceable on disc, stria 1crenulate, irregularly broadened along length, striae 2–6 pro-gressively shallower, of similar depth along length but withslight deviations in direction giving appearance of indistinctpunctation; stria 1 deeply incised to apex, striae 2–4 traceableapically but progressively shallower; two dorsal elytral setae inshallow depressions spanning half of interval 3, at 0.23× and0.58× elytral length; lateral elytral setae arranged as 6 + 6.Tarsomeres short and robust, apical width of metatarsomere4 1.5× median length; metatarsomere 4 moderately emarginateapically, the lateral lobes approximately 0.5× median length.Coloration of head and pronotum brunneous, elytral disc some-what darker, rufopiceous, elytral lateral margin narrowly paler,elytral apex more broadly paler, rufoflavous to flavous atthe margin; ventral body surface coloration as in M. lemur.Microsculpture an evident transverse mesh on frons, more iso-diametric on vertex; pronotal disc evenly covered with elongatetransverse mesh microsculpture, the shallowly margined sculp-ticells best viewed outside the reflection of direct light source;elytral disc covered with dense transverse lines only occasion-ally joined longitudinally, the sculpticells narrow enough thatlight is diffracted by the surface, resulting in an indistinct bluishiridescence.
Female reproductive tract. The lone female holotype spec-imen was not dissected.
Distribution. Like five other species, M. ramsdalei isknown from the summit plateau of Mt. Kaala, albeit from alocality towards the southern margin of the summit based onGPS data associated with the specimen (Fig. 13C).
Habitat. The lone record for this species is from a ground-level microhabitat, through sifting ground litter and moss.
Etymology. This eponymic species epithet memorializesthe collector of the unique holotype, Alastair S. Ramsdale, andcommemorates his contributions to the Hawaiian entomologi-cal community.
Mecyclothorax tantalus Britton(Figs 4I; 14G, H; 15G, H; 16B)
Mecyclothorax tantalus Britton 1948: 122.Metrothorax rotundicollis, Perkins 1917: 250 (misidentifica-tion).
Type material. Holotype female (card-mounted): Oahusummit of Mt. Tantalus/Identified by me in Honolulu asrotundicollis Molokai (Perkins’ handwriting)/Holotype Mecy-clothorax tantalus E.B. Britton 1940 (BMNH).
Diagnosis. Convex beetles sharing the glossy piceousupper body surface of M. simiolus (Fig. 4F, I), but with apronotum almost completely lacking hind angles; the onlyevidence of hind angles being a blunt change in curvatureof the laterobasal margin. As in the convex M. simiolus, thepronotal lateral marginal depression is extremely narrow, andthe apical margin of the elytra is obscured by the inflateddisc. However, M. tantalus is characterized by broader elytra,with the humerus rounded, and the base extended laterallysomewhat, resulting in a subquadrate elytral form. Standardizedbody length 4.3–4.7 mm.
Description. Head with broad, shallow, undulated frontalgrooves, the posterior reaches of which are defined by anarrow carina rimming depression of anterior supraorbital seta;posterior supraorbital seta dorsad and posterad dorsal terminusof postocular groove; eyes slightly distended from ocular lobes,ocular ratio 1.37–1.41, ocular lobe ratio 0.71–0.78. Pronotuma transverse ovoid, hind angles defined by slight change incurvature of laterobasal margin; median base evenly convexbetween slightly recurved sinuations mesad obtusely roundedhind angles; median base with 15–20 punctures each side, thebasalmost punctures equidistant from basal margin, producingan indistinct marginal bead; median longitudinal impressionrepresented by shallow depression on base, finely incised andvery shallow on disc; anterior transverse impression finelyincised throughout, shallowest medially, and laterally definingmarginal bead that extends onto little-protruded, rounded frontangle; lateral marginal depression broad inside front angle,extremely narrow posterad to hind angle; basolateral marginalbead posterad the laterobasal depression broadened medially
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

678 J. K. Liebherr
to meet indistinct basal marginal bead. Elytra inflated, withbroadly rounded humeri, the basal elytral groove and humeralmargin equally recurved; parascutellar striole very shallowwith three isolated punctures; parascutellar seta present, suturalstria 1 obsolete posterad position of seta; elytral striae 1–4evident on disc, stria 1 deepest and distinctly punctate, striae2–4 progressively shallower with smaller punctures; stria 1deep, smooth to apex, stria 2 traceable to apex althoughshallow, striae 3–4 obsolete before position where they wouldfuse in striate elytra; two dorsal elytral setae in middle ofinterval 3, at 0.25× and 0.60× elytral length; lateral elytralsetae arranged as 6 + 6. Tarsomeres short and robust, apicalwidth of metatarsomere 4 1.6× median length; metatarsomere2 apically emarginate, outer apical lobe 0.5× median length.Coloration of head, pronotum and elytra concolorous piceous,elytral lateral marginal depression and apex only narrowlypaler, rufoflavous; ventral thoracic and abdominal scleritesand coxae piceous, proepipleura also piceous, and elytralepipleura marginally pale, flavous, but the mesal marginadjoining metepisternum piceous; femora and tibiae flavous.Microsculpture absent from frons and vertex, the glossysurface interrupted by transverse wrinkles; pronotal disc glossy,but with indistinct transverse mesh microsculpture visiblethrough the reflection; elytral disc glossy, indistinct transversesculpticells visible in strial punctuations.
Male genitalia (n = 1). Aedegal median lobe broad basally,apex beyond ostium blunt, apical face nearly straight, tiptightly rounded (Fig. 15G), appearing much like a less angu-late version of the M. simiolus aedeagus (Fig. 15A); medianlobe apex nearly straight in ventral view, the dorsal marginslightly concave basad rounded tip; internal sac with dorsalostial microtrichial patch, and elongate dorsal plate that is0.37× as long as median lobe measured from basal bulb to tip.
Female reproductive tract (n = 1). Bursa copulatrix broad,moderately elongate, tapered to a fusiform apex (Fig. 14G),bursal length 3.4× gonocoxal length and 2.3× bursal breadth;spermathecal gland assembly short, length of gland duct andreservoir subequal to length of spermathecal duct and reser-voir; basal gonocoxite 1 (Fig. 14H) with apical fringe of twosetae, one seta present on medioapical margin; apical gono-coxite 2 broad basally, narrowed to subacuminate apex, andbearing two lateral ensiform setae, one dorsal ensiform seta,and two apical nematiform setae.
Distribution. Known from Tantalus at elevations of545–614 m, and from Puu Lanipo, both in the south KoolauRange (Fig. 16B).
Mecyclothorax pelops sp.n.(Figs 4J; 5; 14I, J; 15I, J)
Type material. Holotype female (card-mounted with dis-sected female genitalia and associated structures in plasticgenitalia vial, sternites glued separately on card): HAWAI-IAN ISLANDS Oahu I. Koolau Mts. Poamoho Trail, 750 m,2.VI.1977/sifting ground litter/F.G. Howarth Collector
BISHOP/HOLOTYPE Mecyclothorax pelops J.K. Liebherr2008 (red, black-bordered label) (BPBM). Paratypes , identicaldate–locality data (BPBM, 2).
Diagnosis. Like those of M. ramsdalei and the nextspecies, M. niobe, beetles of M. pelops are small-bodied withreduced eyes; standardized body length 3.7–4.3 mm, ocularratio 1.32–1.35, and ocular lobe ratio 0.69–0.70. The prono-tum is subquadrate—versus transverse in M. ramsdalei or morerounded in M. niobe (Fig. 4H, J, K)—with distinct obtuse-rounded hind angles and with only the slightest laterobasalsinuation anterad the angles. The elytra are moderately nar-row basally, with the humeri broadly rounded. The elytraare covered with evident transverse-line microsculpture as inM. ramsdalei, but unlike the glossy discal elytral intervals char-acterizing M. niobe. Unlike the smooth elytral striae character-izing M. ramsdalei, the discal striae of M. pelops are punctate.
Description. Head with deep, undulated frontal groovesthat terminate posteriorly at hind margin of anterior supraor-bital setal depression; posterior supraorbital seta dorsad andposterad dorsal terminus of poctocular groove. Pronotum rel-atively narrow, MPW/PL = 1.25–1.29; median base nearlysmooth, minute punctures each side of midline interspersedwith more longitudinal wrinkles near juncture with disc,basal marginal bead present; median longitudinal impressionwidest at front of base, finely incised, shallow on disc; ante-rior transverse impression obsolete medially, deepened lat-erally to become marginal bead mesad slightly protruded,tightly rounded front angles; lateral marginal depression nar-row throughout anterior half of length, even at front angles,and slightly broader, flatly explanate in basal half anteradlaterobasal sinuation; basal margin upraised posterad shal-low laterobasal depression. Elytra with basal groove brieflycurved mesad the more broadly curved humerus; lateralmarginal depression gradually diminished to smoothly meetbasal groove; parascutellar striole very shallow, 2–3-punctate;parascutellar seta mesad very shallow base of sutural stria 1;striae 1–4 evident as continuous depressions on disc, stria 1deepest, with minute punctures along length isolated by deeplyimpressed strial segments, cuticle between shallower puncturesof striae 2–3 nearly coplanar with intervals, and stria 4 shal-lowest and nearly smooth; stria 5 evidenced by very shallow,isolated punctures; stria 1 continuous and deep to apex, stria 2traceable nearly to apex, but ultimately absent; two dorsal ely-tral setae present, the anterior in middle of third interval, theposterior adjacent to stria 2, at 0.25× and 0.67× elytral length;lateral elytral setae arranged as 6 + 6. Tarsomeres moder-ately elongate and gracile, metatarsomere 4 apical width 1.3×median length; metatarsomere 4 apically emarginate, outer api-cal lobe 0.5× as long as median length. Coloration of dorsalbody surface a uniform brunneous, a piceous cloud on apex ofeach elytral half, bordered by rufoflavous suture, and broadlyflavous apex. Microsculpture on frons an elongate transversemesh mixed with transverse lines, vertex covered with mix ofisodiametric and transverse sculpticells; pronotal disc coveredwith transverse-line microsculpture intermixed with a minority
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 679
of transverse sculpticells; elytra covered with transverse-linemicrosculpture with little tendency to form a mesh.
Male genitalia (n = 1). Aedeagal median lobe broadbasally, apically narrowed to a broadly rounded tip (Fig. 15I);in ventral view the median lobe shaft straight, only the verynarrow blade-like apex curved to the right (Fig. 15J); inter-nal sac with well-developed dorsal ostial microtrichial patch,dorsal plate elongate, 0.38× as long as median lobe measuredfrom basal bulb to tip.
Female reproductive tract (n = 1). Bursa copulatrixextremely broad basally, with slight constriction defining anapical bulb, surface membranous as indicated by faint wrin-kling of cuticle when dissection is compressed under microslidecover slip (Fig. 14I), bursal length 3.3× gonocoxal length,2.3× bursal breadth; spermathecal gland assembly short, lengthof gland duct and reservoir 0.50× length of spermathecal ductand reservoir; basal gonocoxite 1 (Fig. 14J) with apical fringeof two or three setae, apicomedial margin either with a singleseta or glabrous; apical gonocoxite 2 broadly subtriangular,lateral margin moderately excavated, apex acuminate, bearingtwo lateral ensiform setae, one dorsal ensiform seta, and twoapical nematiform setae.
Distribution. Known only from the type locality at thesummit of the Poamoho Trail in the north Koolaus (Fig. 5),where it is sympatric with M. satyrus.
Habitat. The only record for this species involves collect-ing the beetles from sifted ground litter.
Etymology. Pelops was dismembered by his father Tanta-lus, cooked and and offered up as food for the Gods, but thetreachery was discovered and Zeus resurrected him (Graves,1988). The species epithet recognizes the similarity betweenM. tantalus and M. pelops, as well as the continued residenceof M. pelops in nature.
Mecyclothorax niobe sp.n.(Figs 4K; 10C; 14K, L)
Type material. Holotype female (card-mounted with dis-sected female genitalia and associated structures in plastic gen-italia vial, sternites glued separately on card): ‘Palolo, OahuColl.O.H.S.’ on obverse, ‘13 December 2008’ on reverse/Mecyclothorax n. sp. nr. tantalus E.C.Z. / Mecyclothorax n. sp.tantalus palolo ‘female’ #1 J.K. Liebherr 2008/HOLOTYPEMecyclothorax niobe J.K. Liebherr 2008 (red, black-borderedlabel) (BPBM). Paratype, Hawaii, Oahu: Olympus, 8-ix-1912(Swezey) (BPBM).
Diagnosis. Small, broad, convex beetles—standardizedbody length 3.6 mm—characterized by a subglobose pronotum,the pronotal hind angles very obtuse and blunt (Fig. 4K). Asin M. pelops, the eyes are reduced, little convex and small indiameter, with the ocular ratio 1.31–1.32 and the ocular loberatio 0.58. Besides the smaller size and broader body, M. niobeis diagnosed from M. pelops by the total lack of microsculptureon the pronotal and elytral discs, both somites with a glossy
surface devoid of any consistently traceable microsculpture.Moreover, the discal elytral striae 1–3 are more distinctlypunctured in M. niobe.
Description. Head appearing very broad owing to thereduced eyes and broad frons; frontal grooves deep andstraight, extended from pit at juncture with frontal suture tomesal margin of depression surrounding anterior supraorbitalseta; posterior supraorbital seta dorsad the dorsal terminus ofpostocular groove, twice as far from groove as groove is fromeye. Pronotum subglobose and convex, the hind angles markedonly by a change in curvature of the basal and lateral margins;median base completely smooth, inflated; basal marginal beadobsolete medially, present and narrow laterad midline, broad-ened laterally to form posterior wall of very small laterobasaldepression; median longitudinal impression very shallow andtraceable only on disc; anterior transverse impression obso-lete medially, deeper laterally to become marginal bead mesadslightly protruded, tightly rounded front angle; lateral marginaldepression narrow throughout length from front to hind angle,but broad enough so that outer margin is slightly upturned. Ely-tra globose, basal groove indistinctly curved laterad scutellum,then broadly curved anterad to define humerus at base of lateralelytral depression; parascutellar striole deep, 3-punctate; paras-cutellar seta present mesad the base of sutural stria 1; striae1–4 evident on disc, stria 1 distinctly punctate, segments ofstria between punctures nearly coplanar with adjoining inter-vals, striae 2–3 with punctures as large as in stria 1 but moreisolated; stria 4 defined by series of isolated punctures; stria1 deep and continuous to apex, stria 2 absent in apical 0.1×elytral length; two dorsal elytral setae in middle of third elytralinterval, at 0.25× and 0.60× elytral length; lateral elytral setaearranged as 6 + 6. Tarsomeres robust and short, apical widthof metatarsomere 4 1.9× median length; metatarsomere 4 api-cal margin nearly straight, apical lobes not evident. Colorationof dorsal body surface rufous, apical portions of elytral halvesdarkened by piceous cloud that is margined medially by flavoussutural interval, and more broadly apically by flavous apex;ventral body surface coloration as in M . lemur. Microsculp-ture absent from centre of frons, an indistinct transverse meshpresent laterally, vertex covered with indistinct isodiametricsculpticells; pronotal disc glossy, with only the most indis-tinct transverse lines intermittently traceable on surface; ely-tral surface predominantly glossy, indistinct transverse meshmicrosculpture traceable over portions of disc.
Female reproductive tract (n = 1). Bursa copulatrix veryelongate, broadened apically and constricted in basal quarterof length (Fig. 14K), bursal length 5.0× gonocoxal length,and 2.6× apical bursal width; spermathecal gland assemblyshort, length of gland duct and reservoir 0.67× length ofspermathecal duct and reservoir; basal gonocoxite 1 (Fig. 14L)with apical fringe of two setae, medioapical margin glabrous;apical gonocoxite 2 narrow, apical third of length narrowed tosubacuminate apex, and bearing two lateral ensiform setae, onedorsal ensiform seta, and two apical nematiform setae.
Distribution. Known from the proximate localities ofPalolo and Olympus on the crest of the south Koolau Range(Fig. 10C).
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

680 J. K. Liebherr
Etymology. Niobe, daughter of Tantalus and sister toPelops, boasted to the goddess Leto about her comparativebounty of children, whereupon Leto commanded her twochildren Artemis and Apollo to kill Niobe’s seven sons andseven daughters (Graves, 1988). As M. niobe is known fromonly two specimens collected nearly 100 years ago in areasnow encroached upon by Honolulu that suffer concomitantincreased presence of invasive plants (Cuddihy & Stone, 1990),this species’ fate likely reflects the demise of Niobe’s line.
Discussion
The Oahu Mecyclothorax fauna comprises several sympatric ornarrowly parapatric cryptic species complexes. These includethe species of the M. brevis group differentially distributedon the slopes of Tantalus in the south Koolau Range, andthe sister species pair M. carteri and M. invictus sympatri-cally distributed on the summit of Mt. Kaala in the WaianaeRange. In both instances, male genitalic configurations are thebest means to reliably diagnose species. The geographic distri-butions of Oahu Mecyclothorax species define three discrete,highly isolated areas of endemism on Oahu. Those areas havebeen differentially impacted by human development, allowinga fine-scale view of species endangerment for Honolulu County(= Oahu), a county exhibiting one of the highest endangermentrates in the United States of America (Dobson et al., 1997;Rutledge et al., 2001).
Cryptic species and genitalic evolution
It has been long and widely known that male genitalia maydiagnose closely related species of arthropods that may dif-fer only subtly in other characters. Yet the apparently obviousdifferences portrayed in male genitalia may reside in other non-biological phenomena (Huber, 2003). These may include ana priori assumption of the utility of genitalia for diagnosis,coupled with the examination of an inadequate number of spec-imens to assess variablity. This mistake may be compoundedfor populations at different localities when we extend incorrectqualitative assessments of sympatric individuals to allopatricpopulations. In this instance we underestimate geographic vari-ation and overestimate the diagnostic utility of genitalia. Wemay also emphasize changes in the shape of genitalia as diag-nostic, when the underlying basis for such shapes may reside indevelopmental history, or in the influence of the environment.And finally, we may simply assume that genitalic differencesare diagnostic, when they might represent polymorphisms.
The cryptic species complexes of Oahu Mecyclothorax, aspresented above, seem immune to such criticisms. For theM. brevis complex, all specimens are sympatric or narrowlyparapatric, being found on the slopes of Tantalus. Adequatetaxonomic material collected by Perkins and Giffard allowthe multiple dissections that permit evaluation of infraspecificgenitalic variability versus interspecific morphological dis-continuities (Fig. 6A–K). Moreover, in this species complex,although male genitalia are the most definitive means by
which to identify specimens to species, other characters suchas microsculpture on the vertex (Fig. 3) and elytra, pronotalshape (Fig. 4E, F), and development of the seventh elytralinterval (Fig. 8) all serve to support the species diagnoses.Corresponding differences exist in the dimensions and con-figurations of the female bursa copulatrix (Fig. 9A, C, E),although specimen sampling is more limited for this set ofcharacters. The single geographically outlying species in thiscomplex from Mt. Kaala—M. uncinus—differs distinctly inmale genitalia (Fig. 6A), but it can also be diagnosed byreduced microsculpture, and anteriorly projected elytral humeri(Fig. 2B). The second sympatric species pair, M. carteri andM. invictus, is also characterized by distinctively diagnosablemale genitalia (Fig. 11D–I), as well as by consistent differ-ences in the microsculpture of the head, pronotum and elytra.Thus the Oahu Mecyclothorax fauna includes several com-plexes of species whose morphological transformations havecentred on changes to the male genitalia, microsculpture, andthe shapes of various somites.
Similar diagnostic differences are observed in the MolokaiMecyclothorax fauna, although on this island the majority ofcryptic species pairs are allopatric. Six adelphotaxic speciespairs are distributed allopatrically in eastern Molokai, centredon the Kainalu Valley and summit crest above Wailau Valley,with their adelphotaxa distributed to the west in the highlandsbordering Pelekunu Valley (Liebherr, 2006b). In all instancesmale genitalia and microsculptural difference diagnose thesesister pairs, although in two instances the configuration of thefemale bursa copulatrix can also diagnose the species.
An additional partially sympatric Molokai species pair—M. lissopterus Liebherr and M. arcuatus Liebherr—deviatesfrom all others in that the male aedeagus and female bursacopulatrix covary in size, with both the aedeagus and the bursaof M. lissopterus being distinctly and diagnosably smaller. Inthis instance, the aedeagus of a male M. arcuatus could not beaccommodated within the bursa of a female M. lissopterus.Liebherr (2006b) hypothesized that simultaneous evolutionof both the male genitalia and female reproductive tract,mediated by homologous gender-specific development of maleand female imaginal discs during metamorphosis to the imago,effected this morphological transformation.
These situations are very different from that observed amongspecies of Hawaii Island Mecyclothorax (Liebherr, 2008a).The fauna of this youngest of the Hawaiian Islands is repletewith extremely variable species, with variation in both externaland internal characters represented both infrapopulationally andgeographically. Among the most geographically widespreadspecies in this fauna found across all four major volcanoes thatmake up the island—Kohala, Mauna Kea, Hualalai and MaunaLoa including Kilauea—M. deverilli (Blackburn) exhibits sub-stantial variation in male genitalia. Males from the variouspopulations differ dramatically in median lobe configuration(Liebherr, 2008a: Figs 67–78), although not in the configura-tion of the internal sac, the membranous structure everted fromthe median lobe during copulation at whose apex lies the gono-pore. In this species, only infrapopulational variation detected
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 681
through numerous dissections precluded a mistaken dismem-berment of the species into numerous artificial taxa. Anotherwidespread species, M. konanus Sharp, compounds male gen-italic diversity with variation in external characters such aspronotal shape and microsculpture.
Based on evidence from the Mecyclothorax faunas of thesethree islands, it would appear that speciation on the olderislands—Oahu can be dated to 3.3 Ma, and Molokai to 1.7 Ma(Carson & Clague, 1995)—has proceeded to fix genitalic aswell as external morphological variation to a much greaterdegree than has occurred on the youngest island, Hawaii, datedto 0.5 Ma. The discrete genitalic configurations observed inthe males of Molokai and Oahu, compared with the more sub-tle external changes in shape and microsculpture, may reflectobservational bias. Nonetheless, the relatively greater changesin male genitalia versus female genitalia, although exceptionssuch as M. lissopterus and M. arcuatus can be cited, suggestthat sexual selection may be operating during the evolutionof these Mecyclothorax faunas. The male genitalic variationwithin widespread species on Hawaii Island indicates thatfemales from there are provided variable information on malesduring copulation. Given the limited evidence obtainable froma taxonomic study, the presence in widespread species of sub-stantial variation in male genitalia coupled with more subtlevariation in female bursal configuration favours the hypothesisof female-choice-based sexual selection (Eberhard, 1985). OnOahu, female choice among the reproductive advances of con-specific and non-conspecific males could certainly take placein the two cryptic species complexes made up of sympatricor narrowly parapatric species. However, the adoption of thishypothesis as a general one must take into account similarobservations of male variation and relative female similaritybetween allopatric sister species on Molokai. Presumably thesespecies were isolated geographically, with differences leadingto speciation evolving since separation. Females might stillchoose among males within their population, but, at present,populations of each species are isolated from each other, pre-cluding reinforcement. Thus acceptance of female choice asthe basis for genitalic variation across Hawaiian Mecyclotho-rax must posit a continuing dialogue between the sexes withregard to genitalic variation and its assessment during mating.This mutual assessment would take place within the contextof isolated populations affected by different selective regimesand processes brought into play by limited population sizes,suggesting that sexual selection might not be the sole cause ofrapid diversification.
The Hawaiian Drosophilidae has been proposed as the sig-nal example of sexual selection leading to rampant diver-sification within the Hawaiian Islands (Kaneshiro, 1987;Kaneshiro & Boake, 1987). Within the Hawaiian fauna,species of Drosophila Fallen exhibit substantial secondarymale structures—head shape, tibial brushes—that are used incomplicated pre-mating displays (Kaneshiro, 1983). Infraspe-cific and interspecific variation in such structures, such asthe foretibial ‘cilia’ of the planitibia group of picture-wingedDrosophila (Carson et al., 1982), provide an example of mor-phological characters that can be immediately connected to
a specific mating behavior—pre-copulatory courtship. Muchas in Mecyclothorax, variation in setal numbers is highestwithin populations of the most recently evolved Hawaii Islandspecies, D. silvestris (Perkins), with much less infraspecificvariation exhibited within the species from Oahu and Molokai,namely D. hemipeza (Hardy) and D. differens (Hardy &Kaneshiro). Male genitalic characters are highly congruent withthese secondary sexual characters (Kaneshiro et al., 1995), andhighly predictive phylogenetically at the species-group level(Bonacum et al., 2005). However, given the relative homo-geneity of male genitalia within species groups (Kaneshiro,1976) and the asymmetrical mating success during hybridiza-tion of various species (Kaneshiro, 1983, 1990), it is apparentthat the configuration of male genitalia may be of lesser signif-icance for individual mating success in Hawaiian Drosophila.
The second major division of Hawaiian drosophilids basedon mating behaviour, Scaptomyza Hardy (O’Grady et al.,2003), may be more relevant to a discussion of male geni-talic evolution in Mecyclothorax. Scaptomyza exercise ‘simpleassault-type courtship’ (Kaneshiro, 1983), at the same timeexhibiting much less dramatic external morphological differen-tiation. Like Mecyclothorax, sympatric species as well as somespecies allopatrically distributed on neighbouring islands, forexample Maui and Hawaii, can only be diagnosed by charac-ters of the male claspers, ninth tergite, and anal plates (Hardy,1965). As of now, however, it is not possible to determine theamount of male genitalic variation among Scaptomyza speciesin order to assess whether a pattern of diminished variationon older islands is shared with Mecyclothorax. For example,in the Scaptomyza subgenus Elmomyza Hackman—treated assubgenus Trogloscaptomyza Hardy by Hardy (1965) and Hardy& Wheeler (1966)—only 17 of 77 species for which numbersof male specimens were specified included type series of morethan ten specimens. Fully 20 others were described from uniquemales or a female only, 15 from two male types, and 8 morefrom only three male types. A more complete array of dis-sected individuals of the various taxa from various localitiesis required before we can assess similarities to the pattern ofdiminished variation associated with island age observed inMecyclothorax.
Although the most dramatic genitalic evolution among theOahu Mecyclothorax species involves the male aedeagus, onedramatic instance of a discrete change in the female reproduc-tive tract also has occurred. The bursa of M. simiolus is par-tially divided, so that a smaller ventral lobe is situated betweenthe larger, plesiomorphic dorsal bursal lobe and the commonoviduct (Fig. 14C). Both lobes connect to the vaginal areabetween the termini of the ventrally located common oviductand the dorsally located spermathecal duct. Analogous bipartitebursae have been observed in two other unrelated Mecyclotho-rax species, M. oculatus Sharp of Molokai (Liebherr, 2006b)and M. oopteroides Liebherr & Marris (2009) of the ChathamIslands, New Zealand. Beyond arguing for parallel evolutionof this condition based on phylogeny, the relative dimensionsof the dorsal and ventral lobes in the various species arguefor homoplastic origins of these structures. In M. oculatus, theventral lobe is larger than the dorsal lobe (Liebherr, 2006b:
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

682 J. K. Liebherr
fig. 125), and of the same dimensions as the single lobe in thesister species, M. dunbarorum Liebherr. By contrast, in bothM. simiolus and M. oopteroides, the dorsal lobe is larger (e.g.Fig. 14C) and of similar dimensions to the single bursal lobeof closely related species; for example M. satyrus (Fig. 14E)for the former and M. otagoensis Liebherr and Marris for thelatter. Liebherr & Marris (2009) argued that the function of abipartite bursa copulatrix may have more to do with ovipositionor fertilization behaviour than with mating. A shorter ventrallobe could be used to position and facilitate ejection of theegg during oviposition. Conversely, a longer ventral lobe couldhold one egg while a second was being fertilized, its anteriorend positioned near the spermathecal duct opening (Chapman,1998). Thus the condition observed in M. dunbarorum couldconceivably support oviposition of more than one fertilized eggat a time.
Areas of endemism and area relationships
Previous revisionary studies of Hawaiian Carabidae demon-strated the existence of three areas of endemism on Oahu:the Waianae Range, and a southern and northern divisionof the Koolau Range (Fig. 1). For the genus Blackburnia(Liebherr & Zimmerman, 2000), the Waianae Range houses18 species, of which eight are precinctive to the mountainrange. The much longer Koolau Range houses 24 Blackburniaspecies, with 15 species precinctive to the range. Most Koolau-specific species of Blackburnia are distributed throughout thenorth–south length of the Koolaus, although B. hihia Liebherrand B. fordi Liebherr are restricted to the northern portions,and B. blaptoides (Blackburn) and B. paloloensis Liebherr arerestricted to the southern parts. Among the nine native Oahuspecies of Bembidion (Liebherr, 2008c), four are widespreadacross the island; however, one species triplet has a repre-sentative species in the three areas: B. paratomarium Lieb-herr in the Waianaes, B. gagneorum Liebherr in the northernKoolaus, and B. atomarium (Sharp) in the southern Koolaus.Bembidion koebelei (Sharp) is also restricted to the south-ern Koolaus, although it appears to be a peripheral isolate ofB. ignicola, a broadly distributed species known from all theislands.
Species distributions in the present revisionary study arecongruent with this pattern, with the extremely similar, andputative cladistic triplet of species—M. lemur of the Waianaes,M. satyrus of the northern Koolaus, and M. simiolus ofthe southern Koolaus—representing the areas with closelyrelated species, as in the Bembidion triplet. In these instanceswhere closely related species occupy all recognized areas ofendemism, the vicariant events that isolated these areas wouldserve as the most parsimonious explanations underlying thespeciation events leading to the present-day species.
Other insect groups so far revised for Oahu generally donot exhibit species distributions as restricted as those exhibitedby Mecyclothorax and Bembidion. Although species of thecricket genera Prognathogryllus Brunner v. W., LeptogryllusPerkins, Laupala Otte, and Trigonidium Rambur (Orthoptera:Gryllidae) are restricted to either the Waianae or Koolau
Ranges, those species known from the Koolaus have beensampled at various points along the length of the range (Otte,1994). Like the carabid beetle species, all crickets are flightless,suggesting that the coherence of species distributions alongthe length of the Koolau Range has been maintained bymeans of terrestrial movement of individuals. The fact thatthe Mecyclothorax and Bembidion beetles are much smaller inbody size (∼ 3–5 mm versus 8–30 mm) may underlie theirgreater isolation and propensity to speciate.
The Oahu species of Nesiomiris Kirkaldy (Heteroptera:Miridae) include N. contortus Gagne from the northernKoolaus and N. planatus Gagne from the southern Koolaus,although both species are also found respectively in thenorthern and southern Waianae Range (Gagne, 1997). Inthis instance, shared history with the Waianae Range isimplied, with the Koolaus differentially occupied by the twospecies. Nesiomiris uncinatus Gagne is endemic to the southernKoolaus—Tantalus and Kaau Crater—with competition with itsclose relative M. beardsleyi Gagne suggested as the reason forthis limited relictual distribution (Gagne, 1997).
Based on the finest-grain response of insect taxa to thepresent-day geography of Oahu, it is clear that three distinctareas of endemism can be recognized, and that these areasshould be the minimal basis for decisions made regardingconserving a comprehensive representation of the Oahu insectfauna.
The underlying cause for the biogeographic subdivision ofthe Koolau Range is no doubt in part due to the extreme lengthand very limited breadth of the habitable montane communitiesdistributed along the range. The crest of the Koolaus lies wellbelow 1000 m elevation throughout its length, with the majorpeaks Lanihuli and Konahuanui reaching 845 and 960 m alongthe north and south sides of Nuuanu Valley in the southernKoolaus. This long windward stretch currently supports mixedfern shrubland, a plant community including Dicranopterisand Dicranopterygium fern species, as well as shrub speciesof Dubautia and Styphelia, and low-stature ohia (Gagne &Cuddihy, 1990).
This community would not have been continuously dis-tributed along the current Koolau crest prior to the collapse ofthe Koolau Volcano’s seaward side (Dana, 1849, 1889; Mooreet al., 1989, 1994), for at that time the present-day KoolauRange would have constituted the dry leeward slopes of thevolcano. Moreover, at the time of the collapse, the Island ofOahu stood at least 1000 m higher in elevation (Moore &Clague, 1992), being subsequently depressed by volcanic load-ing on the Pacific Plate owing to the development of Maui Nui(Price & Elliott-Fisk, 2004). Thus, during the development ofthe Koolau Volcano before 2.6 Ma (Carson & Clague, 1995)until its collapse less than a million years later, the broad wind-ward face would have been an optimal area for habitation byMecyclothorax beetles. The Koolau collapse has been vari-ably dated as 3.3–2.5 Ma based on foraminiferal and micro-fossil assemblages in sediments deposited on debris blocksmoved during the Nuuanu landslide (Morgan et al., 2006), or as2.1–1.78 Ma based on palaeomagnetic data (Herrero-Berveraet al., 2002).
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 683
That the Oahu Mecyclothorax have close relationships withthe Molokai fauna is demonstrated by the M. brevis speciesgroup, which contains the five Oahu species plus a singleMolokai species, M. latissimus. The rounded pronotum, convexdorsal body surface, and well-impressed lateral elytral striae ofthese species are all synapomorphies that support monophyly ofthis group. Mecyclothorax acherontius of the northern Koolausis extremely similar to M. palustris of Molokai, althoughphylogenetic relationships of these two species versus severalothers in the species group from Lanai and Maui have yet tobe proposed.
Because there are no Mecyclothorax species known fromKauai, and because the most plesiotypic Hawaiian species arerestricted to Maui (Britton, 1948; Liebherr, 2006b, 2008a),occupation of the geologically oldest part of the Hawaiianrange of Mecyclothorax—Oahu—is of interest. If Oahu werecolonized about 2.0 Ma as part of a primitively cosmopolitandistribution that included Oahu Nui (i.e. Oahu, the PenguinBank, plus western Molokai; Price & Elliott-Fisk, 2004), thatcolonization could have been accomplished over land. Thisprimordial distribution would have been vicariated first by theNuuanu landslide that plunged the northeastern half of KoolauVolcano into the sea, breaking the connection between the northand south Koolau areas, and subsequently by inundation of thePenguin Bank between Oahu and Molokai, vicariating Oahufrom Molokai. A strictly vicariant hypothesis entails additionalspecifications concerning taxic distributions. First, none ofthe most generalized species of Hawaiian Mecyclothorax—the M. montivagus group (Britton, 1948; Liebherr, 2006b,2008a)—presently occur on Molokai or Oahu, instead beingrestricted to East Maui (i.e. Haleakala Volcano) and HawaiiIsland (Liebherr, 2008a). As Haleakala did not begin itsdevelopment until 750 000 years ago (Carson & Clague, 1995),the acceptance of vicariant origins for the Oahu Mecyclothoraxwould entail several additional hypotheses. These include:(i) representatives of the plesiotypic M. montivagus groupwere primordially present on Molokai and Oahu, but arecurrently undiscovered or extinct there; (ii) representatives ofthe other various lineages present on Oahu now classified in theM. palustris, M. brevis, M. flavomarginatus and M. scaritoidesspecies groups were also primordially in place on both Oahuand Molokai. These would have been vicariated first by theNuuanu landslide, resulting in origin of precinctive northernKoolau taxa, and later by the sundering of the PenguinBank, isolating southern Oahu from Molokai. The absence ofM. montivagus group taxa from both Molokai and Oahu isecologically and geologically credible, as both islands currentlylack the elevational stature needed for the high-elevationscrubland habitats occupied by members of the group on Mauiand Hawaii I. (Britton, 1948; Liebherr, 2008a). However, bothMolokai and Oahu were of significantly greater stature in theirshield-building phases, during which scrubland habitats wouldhave been widespread. Extinction events and area relationshipsof extant taxa posited by these hypotheses could be testedusing a reconciled tree biogeographic analysis (Liebherr &Zimmerman, 2000; Liebherr, 2001b) based on a cladistichypothesis for all Hawaiian Mecyclothorax.
Species endangerment
All of the 19 species of Mecyclothorax from Oahu arerestricted to one of the three recognized areas of endemism(Table 1). The earliest collections for each of the species areclearly dependent on the historical patterns of entomologicalactivities in Oahu, with Blackburn’s (1878) species collectedfirst during his various day-long excursions into the KoolauMountains (Blackburn, 1885). The Waianaes were exploredlater, although Blackburn spent 3 days there on one trip(Perkins in Swezey, 1931). In addition, the northern Koolauswere accessed by Perkins at various points during the early1900s, although he had comparatively little success collectingMecyclothorax there.
Blackburn was the first to collect M. brevis and M. lobatus(Table 1) on Tantalus; however, the other two narrowly
Table 1. Temporal limits of collections and associated collectorsfor Oahu Mecyclothorax species from the three recognized areas ofendemism.
Species Collector Year
Waianae RangeM. uncinus Perkins 1896M. carteri Perkins 1902
Kavanaugh 1991M. invictus Liebherr 1991
Polhemus 2006M. euryoides Swezey 1929
Polhemus 2006M. lemur Liebherr 1991M. ramsdalei Ramsdale 2005North Koolau RangeM. acherontius Krauss 1933M. satyrus Perkins 1901
Polhemus 2002M. pelops Howarth 1977South Koolau RangeM. brevis Blackburn ∼1877a
Perkins 1908M. calceus Giffard 1908M. dentatus Koebele 1897
Giffard 1906M. lobatus Blackburn ∼1877a
Williams 1929M. flavomarginatus Blackburn ∼1877a
Perkins 1906M. oahuensis Blackburn ∼1877a
M. ovatulus Blackburn ∼1877a
M. simiolus Blackburn ∼1877a
Zimmerman 1937M. tantalus Perkins 1903
Smith 1965M. niobe Swezey 1908
Swezey 1912
aSpecimens described in Blackburn (1878) were collected after his1876 arrival in Honolulu (Lea, 1912). Blackburn’s specimen ofM. ovatulus is assumed contemporaneous with his other specimens.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

684 J. K. Liebherr
parapatric members of the species complex—M. dentatus andM. calceus—were not collected until the later period of col-lecting by the territorial entomologists R. C. L. Perkins (Scott,1956) and Albert Koebele (Perkins, 1943), and by the ama-teur entomologist Walter Giffard (Swezey & Fullaway, 1931).This discrepancy is probably a result of the different meansof access to Tantalus enjoyed by the various entomologists.Blackburn’s access would have been restricted to a ridge trailstarting in the Punahou area, and traversing Ualakaa, Kakea,and reaching Tantalus and the summit, then called Puu Ohia,from the south (Lyons et al., 1881). By 1899, a governmentroad had been built from the Punchbowl, winding up PauoaRidge and reaching Tantalus from the west (Beasley & Tay-lor, 1899). Perkins stated that the mountain is ‘accessible up to1500 ft elevation, either by riding, driving or walking, and theactual summit can be reach in an hour and a half on foot fromHonolulu itself’ (1906: 39). Elevations given by Giffard, suchas 1300, 1500 and 1800 ft, are herein interpreted as elevationsalong the track of this government road to the summit. Perkinsalso referenced the road in his labeling; for example, speci-mens of M. flavomarginatus collected ‘makai Government road(BPBM)’. Perkins (1906) commented that he did not conduct‘systematic collecting’ on Tantalus until 1900, although he hadvisited it for 1 day in 1892.
Viewing last dates of collections for the various OahuMecyclothorax (Table 1), only four of ten species knownfrom the south Koolau Range area of endemism have beencollected during the past 100 years. Such long-term absenceof collections is not observed for the other two areas; forexample, five of six Waianae species have been collectedrecently—indeed, three of them for the first time since 1991.The lack of specimens from the south Koolau Range isnot based on a comparative lack of collecting effort. Thesum since 1908 of collecting days on which at least onecarabid beetle was collected amounts to 223 days for theWaianaes, 134 days for the north Koolau Range area asdefined here (Fig. 1), and 263 days for the south KoolauRange (Liebherr, 2000). Therefore the lack of collections ofMecyclothorax from the south Koolaus appears to reflect agenuine decrease in species abundance, or at least decreasedspecies apparency to a long, diverse line of entomologists.This decrease was noticed by Perkins (1906: 39): ‘in thepast 5 years again I notice many changes all tending to itsimpoverishment [of Tantalus] as a collecting ground’ (Perkins,1906: 39).
Endangerment status may be defined by the size of thespecies’ distributional ranges (IUCN, 2001), with criticallyendangered status assigned for species known from rangesencompassing less than 50 km2, and endangered species sta-tus for species with ranges of less than 100 km2. The IUCNcategories of critically endangered and endangered generallycorrespond to the United States Endangered Species Act sta-tus ranks GH (possibly extinct) and G1 (critically imperiled)(Wilcove & Master, 2005). All Oahu Mecyclothorax species(Figs 5; 7; 10; 13; 16) are known from distributions con-siderably smaller than 50 km2. The most expansive distribu-tions estimated using area of occupancy criterion B (IUCN,
2001: 12) include M. satyrus at 23 km2 (Fig. 5), M. carteri at∼20 km2 (Fig. 7), and M. simiolus at 16 km2 (Fig. 16A). Allother distributional ranges are defined by the type localitiesalone, or relatively proximate localities defining areas of occu-pancy considerably smaller than 10 km2 (e.g. Figs 5; 7; 10;13; 16B).
Given that several species pairs allopatrically occupy differ-ent areas of endemism, and that the areal extent comprisingmontane habitats within those areas is limited, Oahu Mecy-clothorax species may have inhabited primordial distributionalranges smaller than 100 km2, or conceivably even less than50 km2. Thus using the criteria of the IUCN (2001), theycould have qualified for endangered or critically endangeredstatus before humans colonized Hawaii. In these instances thegeographically based endangerment criteria of the IUCN areinsensitive to the varied biogeographic histories of Oahu Mecy-clothorax. It would seem more appropriate to ordinate thevarious species of a taxonomic group occupying any partic-ular geographic area on a scale of endangerment. Species suchas those of the M. brevis species complex that occupy proxi-mate geographic ranges defined principally by elevation wouldthus be given a higher endangerment index than allopatric,closely related species occupying broader ranges, for exampleM. satyrus and M. simiolus. Orthogonal to such spatial ordi-nation, one should assess the temporal distribution of spec-imen collections—as provided in IUCN (2001) populationaltrend criteria—to give a greater endangerment status to speciescollected repeatedly only during earlier times than to thoseobserved also more recently. Finally, those species known fromonly a single collection event should most sensibly be classifiedas data-deficient (IUCN, 2001).
Given these preconditions, M. brevis, M. dentatus,M. flavomarginatus and M. niobe—all occupying extremelyrestricted ranges within the south Koolau Range area ofendemism, and all repeatedly collected then subsequently andcontinuously absent from field collections (Table 1)—merit thehighest endangerment status. Other geographically restricted,southern Koolau species such as M. lobatus, M. simiolus andM. tantalus merit somewhat lower status, as their earlier fieldcollections are complemented by more recent retrievals fromnature. The numerous species known only from single collect-ing events or unique holotypes—M. acherontius, M. calceus,M. lemur, M. oahuensis, M. ovatulus, M. pelops, M. ramsdalei,M. uncinus—enter the taxonomic pantheon based on theirspecific epithets. Whether these names serve as epitaphs orcelebrate viable evolutionary entities can be determined onlyby continued field survey.
Acknowledgements
I thank the following curators and their institutions for loansthat permitted the reassociation of historically collected tax-onomic material essential to this revision: Thomas A. Weir(ANIC); Conrad P.D.T. Gillett (BMNH); G. Allan Samuelsonand Shepherd Myers (BPBM); David H. Kavanaugh (CAS);Bernarr Kumashiro (HDAC); Thierry Deuve (MNHP); Terry
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 685
L. Erwin (NMNH); Jan Forrest (SAMA). I thank Nigel Stork,Stuart Hine, Chris Lyal, Roger Booth and Max Barclay forassisting me during repeated visits to the Natural HistoryMuseum, London. Gordon Nishida and Neal L. Evenhuisdeveloped and maintain the Hawaiian Terrestrial ArthropodDatabase at the Bishop Museum, Honolulu. The State ofHawaii Department of Land and Natural Resources, Castle &Cooke Land Company, and the U.S. Department of the Armypermitted access to the Poamoho Trail. Betsy Harrison Gagneand Dan Polhemus are thanked for their continued support,and for access to Mt. Kaala Natural Area Reserve. I thankAnita Manning for helpful advice on historical maps of Oahu.The Oahu stipple map was graciously provided by Dan Otte.This research was supported by National Science FoundationREVSYS award DEB-0315504.
References
Anonymous N.D. Key to numbers & localities of Mr. Perkins’ Hawai-ian insects. Dept. of Entomology Library, The Natural HistoryMuseum, London.
Baehr, M. (2003) Psydrine ground beetles (Coleoptera: Carabidae: Psy-drinae), excluding Amblytelini, of eastern Queensland rainforests.Memoirs of the Queensland Museum, 49, 65–109.
Baehr M. (2008) Two new species of the genus Mecyclothorax Sharpfrom New Guinea (Coleoptera: Carabidae: Psydrinae). Tijdschriftvoor Entomologie, 151, 133–140.
Beasley, T.D. & Taylor, J.T. (1899) Oahu, Hawaiian Islands. J.T.Taylor, San Francisco. URL www.davidrumsey.com [accessed 23December 2008].
Blackburn, T. (1878) Characters of new genera and descriptions of newspecies of Geodephaga from the Hawaiian Islands II. EntomologistsMonthly Magazine, 15, 119–123.
Blackburn, T. (1885) Mr. Blackburn’s resume of his journeys andcollecting in the archipelago. Memoirs on the Coleoptera of theHawaiian Islands (ed. by T. Blackburn and D. Sharp) Vol. 3,pp. 197–208. Transactions of the Royal Dublin Society, New Series.
Bonacum, J., O’Grady, P.M., Kambysellis, M. & DeSalle, R. (2005)Phylogeny and age of diversification of the planitibia speciesgroup of the Hawaiian Drosophila. Molecular Phylogenetics andEvolution, 37, 73–82.
Britton, E.B. (1948) A revision of the Hawaiian species of Mecy-clothorax (Coleoptera: Carabidae). Occasional Papers of BerniceP. Bishop Museum, 19, 107–166.
Brown, R.W. (1956) Composition of Scientific Words. SmithsonianInstitution Press, Washington, DC.
CPITT (Centre for Pest Information Technology & Transfer, Universityof Queensland) (2001) Lucid Professional. CSIRO Publishing,Collingwood, Victoria.
Carson, H.L. & Clague, D.A. (1995) Geology and biogeography ofthe Hawaiian Islands. Hawaiian Biogeography, Evolution on a HotSpot Archipelago (ed. by W.L. Wagner and V.A. Funk), pp. 14–29.Smithsonian Institution Press, Washington, DC.
Carson, H.L., Val, F.C., Simon, C.M. & Archie, J.W. (1982) Mor-phometric evidence for incipient speciation in Drosophila silvestrisfrom the Island of Hawaii. Evolution, 36, 132–140.
Chapman, R.F. (1998) The Insects: Structure and Function. CambridgeUniversity Press, Cambridge.
Cuddihy, L. & Stone, C.P. (1990) Alteration of Native HawaiianVegetation: Effects of Humans, Their Activities and Introductions.University of Hawaii Press, Honolulu.
Dana, J.D. (1849) Geology. United States Exploring Expedition 10. C.Sherman, Philadelphia.
Dana, J.D. (1889) Points in the geological history of the Islands Mauiand Oahu. American Journal of Science, 37, 81–103.
Dobson, A.P., Rodriguez, J.P., Roberts, W.M. & Wilcove, D.S. (1997)Geographic distribution of endangered species in the United States.Science, 275, 550–553.
Eberhard, W.G. (1985) Sexual Selection and Animal Genitalia. HarvardUniversity Press, Cambridge, MA.
Eldredge, L.G. & Evenhuis, N.L. (2003) Hawaii’s biodiversity: adetailed assessment of the numbers of species in the HawaiianIslands. Bishop Museum Occasional Papers, 76, 1–28.
Gagne, W.C. (1997) Insular evolution, speciation, and revision of theHawaiian genus Nesiomiris (Hemiptera: Miridae). Bishop MuseumBulletins in Entomology, 7, 1–226.
Gagne, W.C. & Cuddihy, L.W. (1990) Vegetation. Manual of theFlowering Plants of Hawaii (ed. by W.L. Wagner, D.R. Herbstand S.H. Sohmer), pp. 45–114. University of Hawaii Press/BishopMuseum Press, Honolulu.
Graves, R. (1988) The Greek Myths, Complete and Unabridged Editionin One Volume. Moyer Bell Ltd., Mt. Kisco, NY.
Hadley A. (2006) CombineZ5 (computer program for stackingdigital images). Free Software Foundation Inc., Boston, MA.URL www.hadleyweb.pwp.blueyonder.co.uk/CZ5/combineZ5.htm[accessed 30 December 2008].
Hardy, D.E. (1965) Diptera: Cyclorrapha II. Series Schizophora,Section Acalypterae I, Family Drosophilidae. Insects of Hawaii, 12,vii, 1–814.
Hardy, D.E. & Wheeler, M.R. (1966) Descriptions and notes onHawaiian Drosophilidae (Diptera). Studies in Genetics, 3, 195–244(University of Texas Publication No. 6615).
Herrero-Bervera, E., Canon-Tapia, E., Walker, G.P.L. & Guerrero-Garcıa, J.C. (2002) The Nuuanu and Wailau giant landslides:insights from paleomagnetic and anisotropy of magnetic suscepti-bility (AMS) studies. Physics of the Earth and Planetary Interiors,129, 83–98.
Howarth, F.G. (1990) Hawaiian terrestrial arthropods: an overview.Bishop Museum Occasional Papers, 30, 4–26.
Huber, B.A. (2003) Rapid evolution and species-specificity of arthro-pod genitalia: fact or artifact? Organisms, Diversity and Evolution,3, 63–71.
IUCN (Species Survival Commission) (2001) IUCN Red List Cate-gories and Criteria (Version 3.1). IUCN—The World ConservationUnion, Gland, Switzerland, Cambridge, U.K. URL www.iucn.org[accessed 30 December 2008].
Jaeger, E.C. (1955) A Source-Book of Biological Names and Terms.Charles C. Thomas, Springfield, IL.
Kaneshiro, K.Y. (1976) A revision of generic concepts in the biosys-tematics of Hawaiian Drosophilidae. Proceedings of the HawaiianEntomological Society, 22, 255–279.
Kaneshiro, K.Y. (1983) Sexual selection and direction of evolutionin the biosystematics of Hawaiian Drosophilidae. Annual Review ofEntomology, 28, 161–178.
Kaneshiro, K.Y. (1987) Speciation via sexual selection versus adaptiveevolution in Hawaiian Drosophila. Insect Science and Applications,8, 447–452.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

686 J. K. Liebherr
Kaneshiro, K.Y. (1990) Natural hybridization in Drosophila, withspecial reference to species from Hawaii. Canadian Journal ofZoology, 68, 1800–1805.
Kaneshiro, K.Y. & Boake, R.B. (1987) Sexual selection and speciation:issues raised by Hawaiian Drosophila. Trends in Ecology andEvolution, 2, 207–212.
Kaneshiro, K.Y., Gillespie, R.G. & Carson, H.L. (1995) Chromosomesand male genitalia of Hawaiian Drosophila: tools for interpretingphylogeny and geography. Hawaiian Biogeography, Evolution on aHot Spot Archipelago (ed. by W.L. Wagner and V.A. Funk), pp.57–71. Smithsonian Institution Press, Washington, DC.
Krombein, K.V. & Pulawski, W.J. (1994) Biosystematic studies ofCeylonese wasps, XX: a revision of Tachysphex Kohl, 1883, withnotes on other oriental species (Hymenoptera: Sphecidae: Larrinae).Smithsonian Contributions to Zoology, 552, 1–106.
Lea, A.M. (1912) The late Rev. Canon Thomas Blackburn, B.A., andhis entomological work. Transactions and Proceedings of the RoyalSociety of South Australia, 36, iii–xl.
Liebherr, J.K. (2000) Appendix, Insects of Hawaii 16. An OddysseyProduction, Ithaca, NY (electronic database available from theauthor).
Liebherr, J.K. (2001a) Preface to the reissue of Insects of Hawaii.Insects of Hawaii, 1,(reissue) v–xv.
Liebherr, J.K. (2001b) Blackburnia gastrellariformis sp.n. (Coleoptera:Carabidae), from Molokai: successful prediction of a new taxon byreconciled tree analysis. Insect Systematics and Evolution, 32, 133–141.
Liebherr, J.K. (2003) Blackburnia lata sp.n. (Coleoptera: Carabidae)from Kauai: morphological transformation in the arboreal microhab-itat. Insect Systematics and Evolution, 34, 41–52.
Liebherr, J.K. (2005a) New species of Mecyclothorax (Coleoptera:Carabidae, Psydrini) from Polipoli, Maui define an area of endemismon Haleakala Volcano, Hawaii. Journal of the New York Entomolog-ical Society, 113, 97–128.
Liebherr, J.K. (2005b) Vanuatu Platynini (Coleoptera: Carabidae):Miocene diversification on the Melanesian Arc. Invertebrate Sys-tematics, 19, 263–295.
Liebherr, J.K. (2006a) Recognition and description of Blackburniakavanaughi, new species (Coleoptera: Carabidae) from Kauai,Hawaii. Journal of the New York Entomological Society, 114, 17–27.
Liebherr, J.K. (2006b) Taxonomic revision of the Mecyclothoraxbeetles (Coleoptera: Carabidae, Psydrini) of Molokai, Hawaii andrecognition of areas of endemism on Kamakou Volcano. Journal ofthe New York Entomological Society, 114, 179–281.
Liebherr, J.K. (2008a) Taxonomic revision of the Mecyclothorax Sharpof Hawaii Island: abundant genitalic variation in a nascent islandradiation. Deutsche Entomologische Zeitschrift, 55, 19–78.
Liebherr, J.K. (2008b) Mecyclothorax kavanaughi sp.n. (Coleoptera:Carabidae) from the Finisterre Range, Papua New Guinea. Tijdschriftvoor Entomologie, 151, 147–154.
Liebherr, J.K. (2008c) Taxonomic revision of Hawaiian BembidionLatreille (Coleoptera: Carabidae, Bembidiini) with a discussion oftheir reductive and derivative evolutionary specializations. Annals ofCarnegie Museum, 77, 19–78.
Liebherr, J.K. & Marris, J.W.M. (2009) Revision of the New Zealandspecies of Mecyclothorax Sharp (Coleoptera: Carabidae: Psydrinae,Mecyclothoracini) and the consequent removal of several species toMeonochilus gen.n. (Psydrinae, Meonini). New Zealand Entomolo-gist, 32, 5–22.
Liebherr, J.K. & Short, A.E.Z. (2006) Blackburnia riparia, new species(Coleoptera: Carabidae, Platynini): a novel element in the Hawaiian
riparian insect fauna. Journal of the New York Entomological Society,114, 1–16.
Liebherr, J.K. & Will, K.W. (1998) Inferring phylogenetic relationshipswithin Carabidae (Insecta, Coleoptera) from characters of the femalereproductive tract. Phylogeny and Classification of Caraboidea (ed.by G.E. Ball, A. Casale and A. Vigna Taglianti), pp. 107–170. AttiMuseo Regionale di Scienze Naturali, Torino.
Liebherr, J.K. & Zimmerman, E.C. (2000) Hawaiian Carabidae(Coleoptera), Part 1: introduction and Tribe Platynini. Insects ofHawaii, 16, 1–494.
Lyons, C.J., Alexander, W.D., Brown, J.F., Monsarrat, M.D., Webster,W. and Covington, R. (1881) Oahu, Hawaiian Islands. Hawai-ian Government Survey, Boston. URL www.davidrumsey.com[accessed 23 December 2008].
Maddison, D.R. (1993) Systematics of the holarctic beetle subgenusBracteon and related Bembidion (Coleoptera: Carabidae). Bulletinof the Museum of Comparative Zoology, 153, 143–299.
Magnacca, K.N. & O’Grady, P.M. (2008) Revision of the‘nudidrosophila’ and ‘atelodrosophila’ species groups of HawaiianDrosophila (Diptera: Drosophilidae), with descriptions of twenty-two new species. Systematic Entomology, 33, 395–428.
Manning, A. (1986) The Sandwich Islands Committee, BishopMuseum, and R.C.L. Perkins: cooperative zoological exploration andpublication. Bishop Museum Occasional Papers, 26, 1–46.
Moore, J.G. & Clague, D.A. (1992) Volcano growth and evolution ofthe Island of Hawaii. Geological Society of America Bulletin, 104,1471–1484.
Moore, J.G., Clague, D.A., Holcomb, R.T., Lipman, P.W., Normark,W.R. & Torresan, M.E. (1989) Prodigious submarine landslides onthe Hawaiian Ridge. Journal of Geophysical Research, 94(B12),17465–17484.
Moore, J.G., Normark, W.R. & Holcomb, R.T. (1994) Giant Hawaiianlandslides. Annual Review of Earth and Planetary Sciences, 22, 119–144.
Morgan, J.K., Resig, J.M. & Siesser, W.G. (2006) Microfossil assem-blages on Tuscaloosa Seamount and their constraints on the ages ofthe Nuuanu landslide, north of Oahu, HI. Journal of Volcanologyand Geothermal Research, 151, 269–278.
Nelson, G. & Platnick, N. (1981) Systematics and Biogeography,Cladistics and Vicariance. Columbia University Press, New York.
Nishida, G.M. (ed.) (2002) Hawaiian Terrestrial Arthropod Checklist,4th edn. Hawaii Biological Survey/Bishop Museum, Honolulu, HI.
O’Grady, P., Bonacum, J., DeSalle, R. & Val, F. (2003) Theplacement of Engiscaptomyza, Grimshawomyia, and Titanochaeta,three clades of endemic Hawaiian Drosphilidae (Diptera). Zootaxa,159, 1–16.
Otte, D. (1994) The Crickets of Hawaii. The Orthopterists’ Society atThe Academy of Natural Sciences, Philadelphia.
Perkins, R.C.L. (1892) Notes on Collecting, Oahu, 1892 . BishopMuseum archives, Ms. SC Perkins, Box 1, p. 13.
Perkins, R.C.L. (1906) The insects of Tantalus. Proceedings of theHawaiian Entomological Society, 1, 38–51.
Perkins, R.C.L. (1917) New Hawaiian Caraboidea. EntomologistsMonthly Magazine, 53, 246–250.
Perkins, R.C.L. (1943) The early work of Albert Koebele in Hawaii.The Hawaiian Planters Record, 47, 2–5.
Perrault, G.G. (1992) Endemism and biogeography among TahitianMecyclothorax species (Coleoptera: Carabidae: Psydrini). The Bio-geography of Ground Beetles of Mountains and Islands (ed. by G. R.Noonan, G. E. Ball and N. E. Stork), pp. 201–215. Intercept, Ltd.,Andover, U.K.
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687

Mecyclothorax of Oahu 687
Price, J.P. & Elliott-Fisk, D. (2004) Topographic history of the MauiNui Complex, Hawaii, and its implication for biogeography. PacificScience, 58, 27–45.
Rosen, D. (1978) Vicariant patterns and historical explanation inbiogeography. Systematic Zoology, 27, 159–188.
Rubinoff, D. & Haines, W.P. (2005) Web-spinning caterpillar stalkssnails. Science, 309, 575 (supplemental on-line materials, 3 pp., 5figs).
Rutledge, D.T., Lepczyk, C.A., Jialong, X & Liu, J. (2001) Spatiotem-poral dynamics of endangered species hotspots in the United States.Conservation Biology, 15, 475–487.
Scott, H. (1956) Robert Cyril Layton Perkins. Biographical Memoirsof Fellows of the Royal Society, 2, 215–236.
Sharp, D. (1903) Coleoptera, Caraboidea. Fauna Hawaiiensis (ed.by D. Sharp), Vol. 3, 175–292. Cambridge University Press,Cambridge.
Swezey, O.H. (1931) Haleauau Valley, the locality where theRev. Thomas Blackburn collected in the Waianae Moun-tains. Proceedings of the Hawaiian Entomological Society, 7,422–424.
Swezey, O.H. (1954) Forest Entomology in Hawaii, Vol. 44. BerniceP. Bishop Museum Special Publication, Honolulu, HI.
Swezey, O.H. & Fullaway, D.T. (1931) Walter M. Giffard, a biograph-ical sketch. Proceedings of the Hawaiian Entomological Society, 7,347–351.
Wilcove, D.S. & Master, L.L. (2005) How many endangered speciesare there in the United States? Frontiers in Ecology and theEnvironment, 3, 414–420.
Zimmerman, E.C. (1948) Introduction. Insects of Hawaii, 1, 1–206.Zimmerman, E.C. (1957) Ephemeroptera—Neuroptera—Trichoptera
and supplement to volumes 1 to 5. Insects of Hawaii, 6, 1–209.Zimmerman, E.C. (1958) Lepidoptera: Pyraloidea. Insects of Hawaii,
8, 1–456.Zimmerman, E.C. (1978a) Microlepidoptera: Part I: Monotrysia,
Tineoidea, Tortricoidea, Gracillarioidea, Yponomeutoidea, and Alu-citoidea. Insects of Hawaii, 9, xvi, 1–882.
Zimmerman, E.C. (1978b) Microlepidoptera: Part II: Gelechoidea.Insects of Hawaii, 9, 883–1903.
Accepted 3 March 2009
© 2009 The AuthorJournal compilation © 2009 The Royal Entomological Society, Systematic Entomology, 34, 649–687
Related Documents











