Taxon-speci¢c and seasonal variations in £agellates grazing on heterotrophic bacteria in the oligotrophic Lake Annecy ^ importance of mixotrophy Isabelle Domaizon , Sylvie Viboud, Dominique Fontvieille Universite ¤ de Savoie, CISM, UMR CARRTEL, 73376 Le Bourget du Lac, France Received 12 November 2002; received in revised form 22 September 2003; accepted 26 September 2003 First published online 23 October 2003 Abstract We investigated the taxonomic composition of flagellate assemblages and taxon-specific bacterial grazing rates of heterotrophic and mixotrophic flagellates in the oligotrophic Lake Annecy (France). The comparison of bacterial grazing rates to bacterial production demonstrated a high transfer efficiency from the bacterial compartment up to flagellates. Per capita grazing rates ranged from 1.2U10 3 to 5.1U10 6 bacteria l 31 h 31 for heterotrophic flagellates, and from 4.8U10 6 to 6.8U10 7 bacteria l 31 h 31 for mixotrophic flagellates. The main bacterial grazers were Katablepharis within heterotrophic flagellates and Dinobryon within mixotrophic flagellates. Our results show that bacterial ingestion by a given flagellate taxon changed seasonally and could vary up to 30-fold. We also provide evidence that mixotrophic flagellates represent an important link in the flux of materials through planktonic food webs in Lake Annecy, suggesting that the introduction of mixotrophs within functional groups could improve our understanding of carbon flux pathways. ȣ 2003 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords : Heterotrophic nano£agellate ; Mixotrophic £agellate ; Grazing rate ; Lake Annecy ; Microbial food web 1. Introduction The recognition of the qualitative and quantitative im- portance of the microbial food web in freshwater ecosys- tems has greatly modi¢ed our approach to plankton ecol- ogy [1^3]. The ecological roles of heterotrophic £agellates and ciliates in freshwater pelagic environments were re- cently reviewed by Nakano [4], focusing especially on their roles as consumers of microbial plankton, food resources for zooplankton, and regenerators of inorganic nutrients. It is now well known that bacterivorous protists have the potentiality to regulate bacterial abundance and produc- tion and are therefore a keystone group in the transfer of picoplanktonic carbon to higher trophic levels [5^7]. How- ever, although heterotrophic nano£agellates (HNF) are generally recognised as the primary consumers of bacte- rioplankton and picoplankton in both marine and fresh- water ecosystems [8^12], the number of records that show a lack of coupling between bacteria and HNF has in- creased in recent years [5,13,14]. Based on their investiga- tion on bacterial grazing rates of heterotrophic £agellates, Cleven and Weisse [9] concluded that it seems obvious from the high variability of the data produced in the liter- ature that we need to learn more about ingestion rates within a given £agellate population. Moreover, recent studies show the importance of mixotrophs as bacterial consumers [15^21], while their signi¢cance in regulating bacterial communities is still under discussion. Only a few studies have investigated seasonal succession and tax- on-speci¢c grazing rates of £agellates [2,9,10,22^24]. None of these studies were under strict oligotrophic conditions and mixotrophic species were rarely taken into account. During this study, we investigated the taxonomic com- position of £agellates and their bacterial consumption in the oligotrophic Lake Annecy. The goals were (i) to inves- tigate the relative importance of di¡erent £agellate taxa (heterotrophic and mixotrophic £agellates) in bacterial regulation through grazing and (ii) to assess seasonal changes in taxon-speci¢c bacterial uptake rates. We as- 0168-6496 / 03 / $22.00 ȣ 2003 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi :10.1016/S0168-6496(03)00248-4 * Corresponding author. E-mail address : [email protected] (I. Domaizon). FEMS Microbiology Ecology 46 (2003) 317^329 www.fems-microbiology.org

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Taxon-speci¢c and seasonal variations in £agellates grazing onheterotrophic bacteria in the oligotrophic Lake Annecy ^
importance of mixotrophy
Isabelle Domaizon �, Sylvie Viboud, Dominique FontvieilleUniversite¤ de Savoie, CISM, UMR CARRTEL, 73376 Le Bourget du Lac, France
Received 12 November 2002; received in revised form 22 September 2003; accepted 26 September 2003
First published online 23 October 2003
Abstract
We investigated the taxonomic composition of flagellate assemblages and taxon-specific bacterial grazing rates of heterotrophic andmixotrophic flagellates in the oligotrophic Lake Annecy (France). The comparison of bacterial grazing rates to bacterial productiondemonstrated a high transfer efficiency from the bacterial compartment up to flagellates. Per capita grazing rates ranged from 1.2U103 to5.1U106 bacteria l31 h31 for heterotrophic flagellates, and from 4.8U106 to 6.8U107 bacteria l31 h31 for mixotrophic flagellates. Themain bacterial grazers were Katablepharis within heterotrophic flagellates and Dinobryon within mixotrophic flagellates. Our results showthat bacterial ingestion by a given flagellate taxon changed seasonally and could vary up to 30-fold. We also provide evidence thatmixotrophic flagellates represent an important link in the flux of materials through planktonic food webs in Lake Annecy, suggesting thatthe introduction of mixotrophs within functional groups could improve our understanding of carbon flux pathways.9 2003 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Heterotrophic nano£agellate ; Mixotrophic £agellate ; Grazing rate; Lake Annecy; Microbial food web
1. Introduction
The recognition of the qualitative and quantitative im-portance of the microbial food web in freshwater ecosys-tems has greatly modi¢ed our approach to plankton ecol-ogy [1^3]. The ecological roles of heterotrophic £agellatesand ciliates in freshwater pelagic environments were re-cently reviewed by Nakano [4], focusing especially on theirroles as consumers of microbial plankton, food resourcesfor zooplankton, and regenerators of inorganic nutrients.It is now well known that bacterivorous protists have thepotentiality to regulate bacterial abundance and produc-tion and are therefore a keystone group in the transfer ofpicoplanktonic carbon to higher trophic levels [5^7]. How-ever, although heterotrophic nano£agellates (HNF) aregenerally recognised as the primary consumers of bacte-rioplankton and picoplankton in both marine and fresh-
water ecosystems [8^12], the number of records that showa lack of coupling between bacteria and HNF has in-creased in recent years [5,13,14]. Based on their investiga-tion on bacterial grazing rates of heterotrophic £agellates,Cleven and Weisse [9] concluded that it seems obviousfrom the high variability of the data produced in the liter-ature that we need to learn more about ingestion rateswithin a given £agellate population. Moreover, recentstudies show the importance of mixotrophs as bacterialconsumers [15^21], while their signi¢cance in regulatingbacterial communities is still under discussion. Only afew studies have investigated seasonal succession and tax-on-speci¢c grazing rates of £agellates [2,9,10,22^24]. Noneof these studies were under strict oligotrophic conditionsand mixotrophic species were rarely taken into account.
During this study, we investigated the taxonomic com-position of £agellates and their bacterial consumption inthe oligotrophic Lake Annecy. The goals were (i) to inves-tigate the relative importance of di¡erent £agellate taxa(heterotrophic and mixotrophic £agellates) in bacterialregulation through grazing and (ii) to assess seasonalchanges in taxon-speci¢c bacterial uptake rates. We as-
0168-6496 / 03 / $22.00 9 2003 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.doi :10.1016/S0168-6496(03)00248-4
* Corresponding author.E-mail address: [email protected] (I. Domaizon).
FEMSEC 1591 26-11-03
FEMS Microbiology Ecology 46 (2003) 317^329
www.fems-microbiology.org

sume that functional guilds could be more easily de¢nedfrom measured bacterivory impact of the di¡erent taxaand then the identi¢cation of relevant functional groupswithin protists would lead to a better understanding of thefunctioning and regulation of microbial food webs.
The study is based on measurements of abundance andbiomass of heterotrophic bacteria and £agellates, measure-ments of production (primary production and bacterialproduction), and evaluation of bacterial grazing rates of£agellates by the £uorescent microspheres method. Thiswork is part of a larger study in which dynamic and sea-sonal bacterial grazing of ciliates and metazooplankton isinvestigated in addition to £agellate grazing in order toquantify carbon £ux from the microbial food web up tohigher trophic levels and the relative importance of top-down and bottom-up forces in the regulation of bacterialpopulations.
2. Materials and methods
2.1. Study site and sampling
Lake Annecy (24.5 km2, altitude 446 m, latitude:45.5‡N, longitude: 6.1‡E) is one of the largest alpine lakesin France [25]. This lake has a maximal depth of 65 mand is divided into two basins. Although nutrient concen-trations are particularly low, phosphorus especially ap-pearing as a limiting factor for algal production (highN/P), the oligotrophic Lake Annecy is characterised by ahigh ¢sh production according to the scienti¢c monitoringthat has been conducted on the lake since the mid-1960s[26].
Our study was focussed on the epilimnion (0^10 m) atthe routine sampling station located in the northern basinof the lake where water depth is at its maximum (65 m).This sampling station was de¢ned according to previousinvestigations conducted as part of the water quality mon-itoring [26]. The study period lasted from April to Novem-ber 2001. Samples were taken in biweekly to monthly in-tervals (sampling dates : 3 and 18 April, 10 and 21 May,5 and 19 June, 17 July, 14 August, 18 September, 15 Oc-tober, 27 November).
2.2. Physicochemical variables
Data on ambient parameters (temperature, dissolvedoxygen, conductivity, chlorophyll a, nutrients) were col-lected by the CARRTEL-INRA team (Thonon-Les-Bains)as part of the scienti¢c monitoring of the lake and havebeen summarised in an annual report [26].
2.3. Bacterial production and primary production
Bacterial production was measured by incorporation of[3H]thymidine according to Fuhrman and Azam [27] from
a sub-sample of the water used for microbe counting. Fif-teen millilitres of the sample were incubated after adding[3H]methylthymidine (9 nM ¢nal concentration, specif-ic activity : 82 Ci mmol31) and non-labelled thymidine(9 nM ¢nal concentration) at in situ temperature in thedark. The samples were treated in duplicate and a forma-lin-¢xed control (2% ¢nal concentration) was systemati-cally included. After 2 h incubation, 1 ml of formalinwas added before macromolecules were extracted in 10%ice-cold trichloroacetic acid for 30 min. The samples were¢ltered onto a polycarbonate membrane (0.22 Wm poresize) and rinsed with cold trichloroacetic acid before beingradioassayed by liquid scintillation counting (Packard Tri-Carb Liquid Scintillation Analyser 1500). Cell concentra-tion was computed from the production using a conver-sion factor (3.86U1018 cells mol31 [H3]Thymidine (TdR))proposed by Strofek [28].
Photosynthetic assimilation was measured by the 14Cmethod according to Steeman Nielsen [29,30]. After 4 hincubation the samples (two light bottles and two darkbottles) were immediately ¢ltered onto 0.45 Wm pore sizecellulose acetate membranes and were radioassayed by liq-uid scintillation counting. The assimilation rate (Wg C l31
h31) was computed according to the ¢ltered volume, theincubation time, the concentration of available mineralcarbon and the amount of radiolabelled carbon.
2.4. Staining and enumeration of heterotrophic bacteria and£agellates
Abundance of heterotrophic bacteria was determinedfrom formaldehyde-¢xed samples (¢nal concentration2%). Counting was carried out under an epi£uorescencemicroscope (Nikon Eclipse TE200) after staining withDAPI (1 Wg l31) and ¢ltration through black polycarbon-ate membranes (0.2 Wm pore size) according to the proto-col described by Porter and Feig [31]. Bacterial biovolumewas measured by a semi-automatic image analysis system(Lucia 4.6, Laboratory Imaging) and was converted to car-bon units using the conversion factor (220 fg C Wm33)proposed by Simon and Azam [32].
Flagellates were ¢xed with glutaraldehyde (1% ¢nal con-centration), stained with primulin [33] and collected onto ablack polycarbonate membrane (0.8 Wm pore size). For£agellates, slides were prepared within 24 h after samplingand were stored at 325‡C to minimise losses of auto£uo-rescence [34]. Slides were observed at a 1250U magni¢ca-tion using an epi£uorescence microscope (Nikon EclipseTE200) under UV light for heterotrophic bacteria andHNF, and under blue light for pigmented £agellates.The C content of £agellates was then calculated using aconversion factor of 220 fg C Wm33 [35].
2.5. Flagellate grazing
Grazing rates of £agellates on heterotrophic bacterial
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329318
communities were estimated from the ingestion of £uores-cent beads [2,36]. The protocol was similar to that used byCarrias et al. [10] and Thouvenot et al. [22]. Experimentswere conducted in duplicate in 250-ml glass containerswhere tracer particles were added. A stock solution oftracer particles (0.5 Wm) was prepared from a concentratedsolution of Fluoresbrite Plain Microspheres (Polysciences)and was treated with bovine serum albumin (0.5 mg ml31)to avoid clumping of particles [10,36]. The microspheresize was chosen according to the size of the bacteria inLake Annecy (mean length of heterotrophic bacteria dur-ing the study was 0.7 M 0.28 Wm). The ¢nal concentrationof microbeads in the experimental bottles was around2U105 beads ml31 while bacterial abundance variedfrom 1.1 to 3.6 cells ml31 during the study period. There-fore, the concentration of microspheres in each experimen-tal bottle was from 5 to 20% of the bacterial concentrationin the lake. Bead concentration was estimated by epi£uor-escence microscopy after ¢ltration onto polycarbonate ¢l-ters (0.2 Wm pore size). The plankton from 0 to 10 m wereacclimatised for at least 5 min in 250-ml glass bottles be-fore beads were injected. Based on preliminary measure-ments of predation kinetics in Lake Annecy, we chose anincubation time of 15 min. At all sampling dates, we con-ducted the grazing experiment in two experimental bottlesin which we took and analysed samples at 0 min and 15min. The incubation was stopped by adding ice-cold glu-taraldehyde (2% ¢nal concentration). The microbeads in-gested were enumerated after ¢ltering sub-samples fromeach bottle (30 ml) onto a 0.8-Wm polycarbonate mem-brane and staining with primulin as described for nano-£agellate enumeration. Flagellates and microbeads wereobserved at a 1250U magni¢cation under UV light andblue light.
Clearance rate (nl ind31 h31) was calculated for eachtaxon by dividing the number of ingested beads per hourby the bead concentration in the bottle. Ingestion rate ofeach taxon (bacteria £agellate31 h31) was calculated bymultiplying the corresponding clearance rate by the heter-otrophic bacterial concentration (abundance actually mea-sured on the day of experiment). The grazing impact (bac-teria h31 l31) of a taxon was estimated by multiplying itsingestion rate by its actual concentration. Bacterial grazingimpact of di¡erent £agellate groups (heterotrophic, mixo-trophic) was computed by summing grazing impacts of alltaxa belonging to the same group. In order to compareloss of bacterial production due to £agellate grazing, bac-terial ingestion rates were converted into carbon units us-ing bacterial biovolumes and a conversion factor of 220 fgC Wm33.
2.6. Analysis
Statistical treatments mainly consisted of correlationanalysis (Pearson correlation factor) to search for empiri-cal relationships between variables.
3. Results
3.1. Ambient parameters
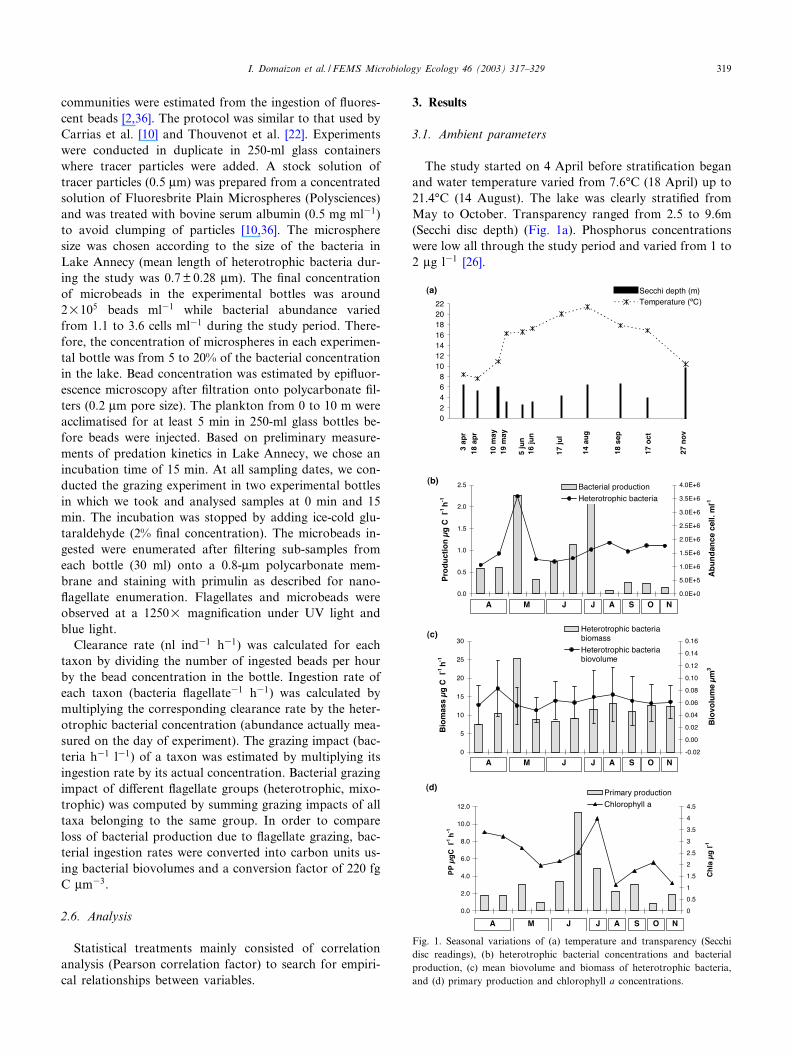
The study started on 4 April before strati¢cation beganand water temperature varied from 7.6‡C (18 April) up to21.4‡C (14 August). The lake was clearly strati¢ed fromMay to October. Transparency ranged from 2.5 to 9.6m(Secchi disc depth) (Fig. 1a). Phosphorus concentrationswere low all through the study period and varied from 1 to2 Wg l31 [26].
Fig. 1. Seasonal variations of (a) temperature and transparency (Secchidisc readings), (b) heterotrophic bacterial concentrations and bacterialproduction, (c) mean biovolume and biomass of heterotrophic bacteria,and (d) primary production and chlorophyll a concentrations.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329 319
3.2. Concentration, biomass and production ofheterotrophic bacteria
Abundance and biomass of heterotrophic bacteria re-spectively ranged from a minimum of 1.1U106 bacteriaml31 and 23.2 Wg C l31 (3 April) to a maximum of3.6U106 bacteria ml31 and 78.9 Wg C l31 on 19 May(Fig. 1b). Filamentous forms were not observed duringthe study period. Heterotrophic bacteria were mainly rep-resented by small-sized free cells (mean biovolume: 0.06Wm3) (Fig. 1c). No correlation appeared between hetero-trophic bacteria (abundance or biomass) and temperature,chlorophyll a concentration or primary production.
Bacterial production reached its maximum (2.25 Wg Cl31 h31) on 19 May and the annual mean was 0.77 Wg Cl31 h31 (Fig. 1c). A second peak was observed in Junewhen the maximal chlorophyll a concentration was ob-served. A signi¢cant positive correlation appeared betweenbacterial production and chlorophyll a concentration(r= 0.679; P= 5%).
3.3. Phytoplankton primary production and chlorophyll aconcentration
Phytoplankton primary production varied between 0.9(October) and 11.3 Wg C l31 h31 (June) in the 0^10-msurface layer (Fig. 1d). This low production is typical ofthis type of oligotrophic system. Similar values were mea-sured during a previous study [37].
Chlorophyll a concentration ranged from 1.13 to 3.99Wg l31, the maximal concentration being recorded duringJune and July (Fig. 1d). For the last 4 years [38] similarlylow values have usually been observed in the euphoticzone of Lake Annecy.
3.4. Taxonomic composition, abundance, biomass of£agellates
Annual means for total £agellate abundance and bio-mass were respectively 2.8U103 cells ml31 and 16 Wg Cl31. HNF represented from 12 to 83% of the total £agel-late abundance. This percentage reached a maximum inMay. For all other sampling dates, pigmented £agellatesoutnumbered HNF. Due to a higher mean biovolume thanheterotrophs, pigmented £agellates dominated £agellatebiomass in the epilimnion throughout the whole studyperiod.
3.4.1. Non-pigmented £agellatesIn most cases, £agellates were identi¢ed to the family or
genus level. Within heterotrophic £agellates, major taxasuch as Kinetoplastids, Choano£agellates and Chrysomo-nads (Heterokonts) could be identi¢ed under the epi£uo-rescence microscope. Some other £agellates, particularlycolourless Cryptomonads (6 Wm long) and small uni£agel-lated cells (3 Wm long) which could not unequivocally be
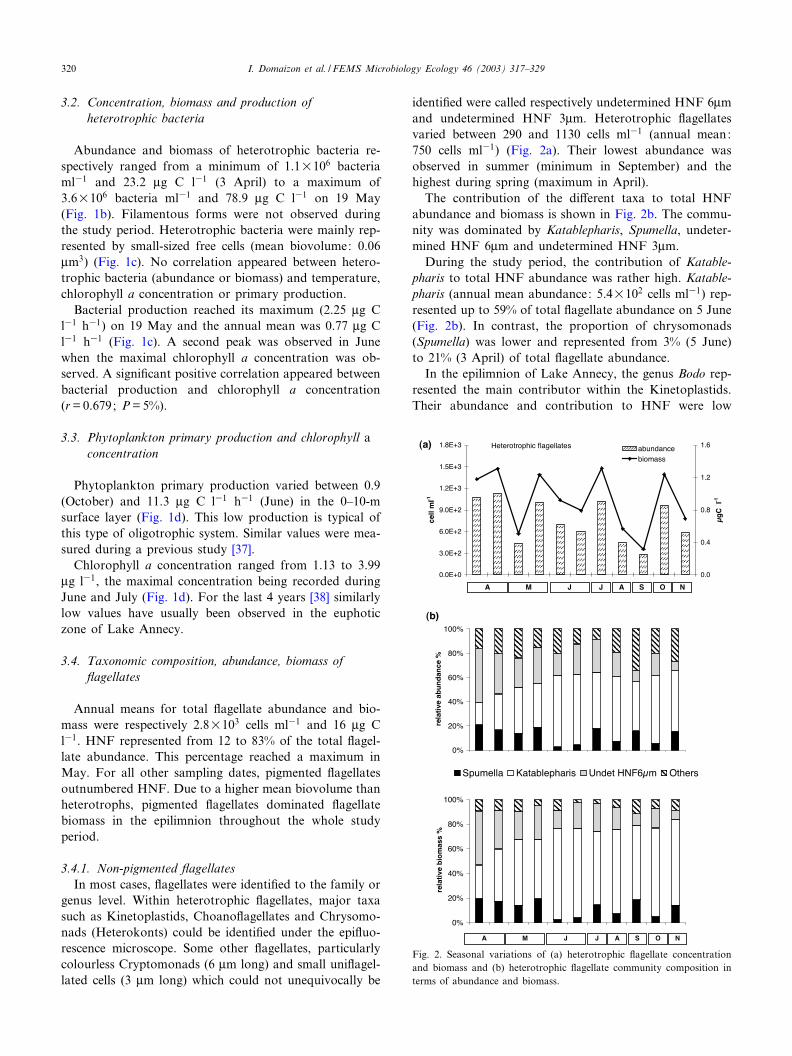
identi¢ed were called respectively undetermined HNF 6Wmand undetermined HNF 3Wm. Heterotrophic £agellatesvaried between 290 and 1130 cells ml31 (annual mean:750 cells ml31) (Fig. 2a). Their lowest abundance wasobserved in summer (minimum in September) and thehighest during spring (maximum in April).
The contribution of the di¡erent taxa to total HNFabundance and biomass is shown in Fig. 2b. The commu-nity was dominated by Katablepharis, Spumella, undeter-mined HNF 6Wm and undetermined HNF 3Wm.
During the study period, the contribution of Katable-pharis to total HNF abundance was rather high. Katable-pharis (annual mean abundance: 5.4U102 cells ml31) rep-resented up to 59% of total £agellate abundance on 5 June(Fig. 2b). In contrast, the proportion of chrysomonads(Spumella) was lower and represented from 3% (5 June)to 21% (3 April) of total £agellate abundance.
In the epilimnion of Lake Annecy, the genus Bodo rep-resented the main contributor within the Kinetoplastids.Their abundance and contribution to HNF were low
Fig. 2. Seasonal variations of (a) heterotrophic £agellate concentrationand biomass and (b) heterotrophic £agellate community composition interms of abundance and biomass.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329320
throughout the study period (from 9 to 93 cells ml31, i.e.from 3 to 10% of the total HNF abundance; Fig. 2b).Such a low concentration in oligo- to mesotrophic lakesis not surprising for this genus [9,10], Auer and Arndt [39]reported that the Bodonid Bodo saltans clearly increases inboth frequency and abundance with increasing trophicstatus.
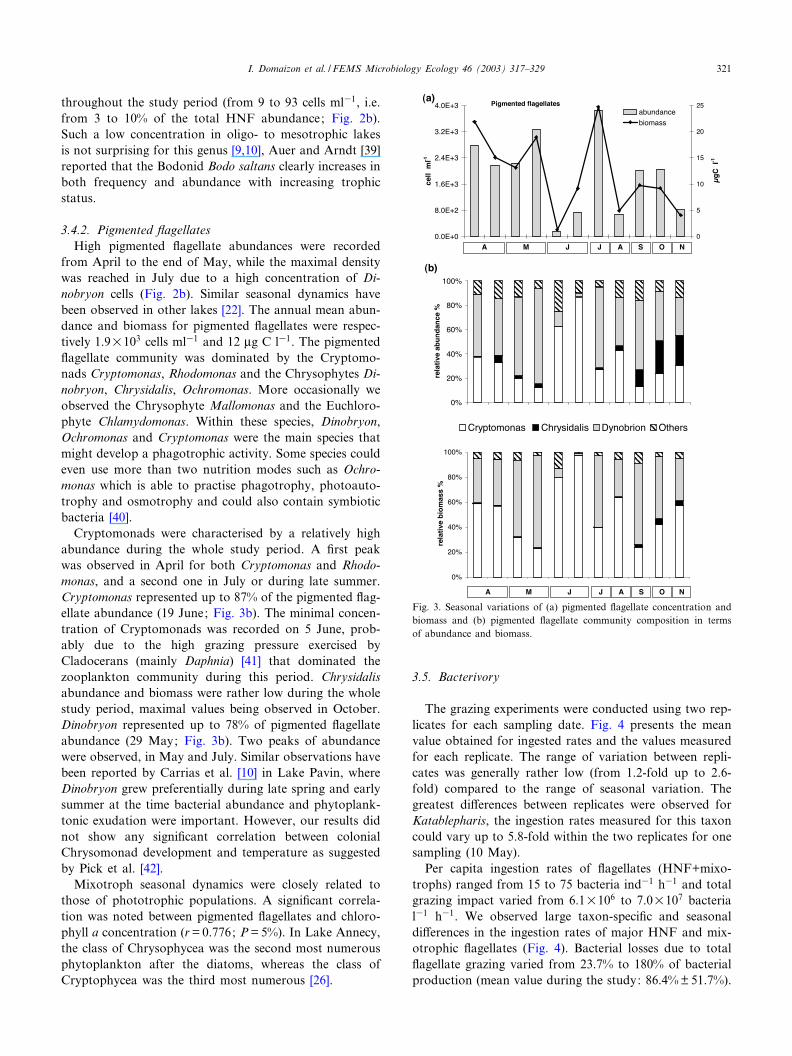
3.4.2. Pigmented £agellatesHigh pigmented £agellate abundances were recorded
from April to the end of May, while the maximal densitywas reached in July due to a high concentration of Di-nobryon cells (Fig. 2b). Similar seasonal dynamics havebeen observed in other lakes [22]. The annual mean abun-dance and biomass for pigmented £agellates were respec-tively 1.9U103 cells ml31 and 12 Wg C l31. The pigmented£agellate community was dominated by the Cryptomo-nads Cryptomonas, Rhodomonas and the Chrysophytes Di-nobryon, Chrysidalis, Ochromonas. More occasionally weobserved the Chrysophyte Mallomonas and the Euchloro-phyte Chlamydomonas. Within these species, Dinobryon,Ochromonas and Cryptomonas were the main species thatmight develop a phagotrophic activity. Some species couldeven use more than two nutrition modes such as Ochro-monas which is able to practise phagotrophy, photoauto-trophy and osmotrophy and could also contain symbioticbacteria [40].
Cryptomonads were characterised by a relatively highabundance during the whole study period. A ¢rst peakwas observed in April for both Cryptomonas and Rhodo-monas, and a second one in July or during late summer.Cryptomonas represented up to 87% of the pigmented £ag-ellate abundance (19 June; Fig. 3b). The minimal concen-tration of Cryptomonads was recorded on 5 June, prob-ably due to the high grazing pressure exercised byCladocerans (mainly Daphnia) [41] that dominated thezooplankton community during this period. Chrysidalisabundance and biomass were rather low during the wholestudy period, maximal values being observed in October.Dinobryon represented up to 78% of pigmented £agellateabundance (29 May; Fig. 3b). Two peaks of abundancewere observed, in May and July. Similar observations havebeen reported by Carrias et al. [10] in Lake Pavin, whereDinobryon grew preferentially during late spring and earlysummer at the time bacterial abundance and phytoplank-tonic exudation were important. However, our results didnot show any signi¢cant correlation between colonialChrysomonad development and temperature as suggestedby Pick et al. [42].
Mixotroph seasonal dynamics were closely related tothose of phototrophic populations. A signi¢cant correla-tion was noted between pigmented £agellates and chloro-phyll a concentration (r= 0.776; P= 5%). In Lake Annecy,the class of Chrysophycea was the second most numerousphytoplankton after the diatoms, whereas the class ofCryptophycea was the third most numerous [26].
3.5. Bacterivory
The grazing experiments were conducted using two rep-licates for each sampling date. Fig. 4 presents the meanvalue obtained for ingested rates and the values measuredfor each replicate. The range of variation between repli-cates was generally rather low (from 1.2-fold up to 2.6-fold) compared to the range of seasonal variation. Thegreatest di¡erences between replicates were observed forKatablepharis, the ingestion rates measured for this taxoncould vary up to 5.8-fold within the two replicates for onesampling (10 May).
Per capita ingestion rates of £agellates (HNF+mixo-trophs) ranged from 15 to 75 bacteria ind31 h31 and totalgrazing impact varied from 6.1U106 to 7.0U107 bacterial31 h31. We observed large taxon-speci¢c and seasonaldi¡erences in the ingestion rates of major HNF and mix-otrophic £agellates (Fig. 4). Bacterial losses due to total£agellate grazing varied from 23.7% to 180% of bacterialproduction (mean value during the study: 86.4% M 51.7%).
Fig. 3. Seasonal variations of (a) pigmented £agellate concentration andbiomass and (b) pigmented £agellate community composition in termsof abundance and biomass.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329 321
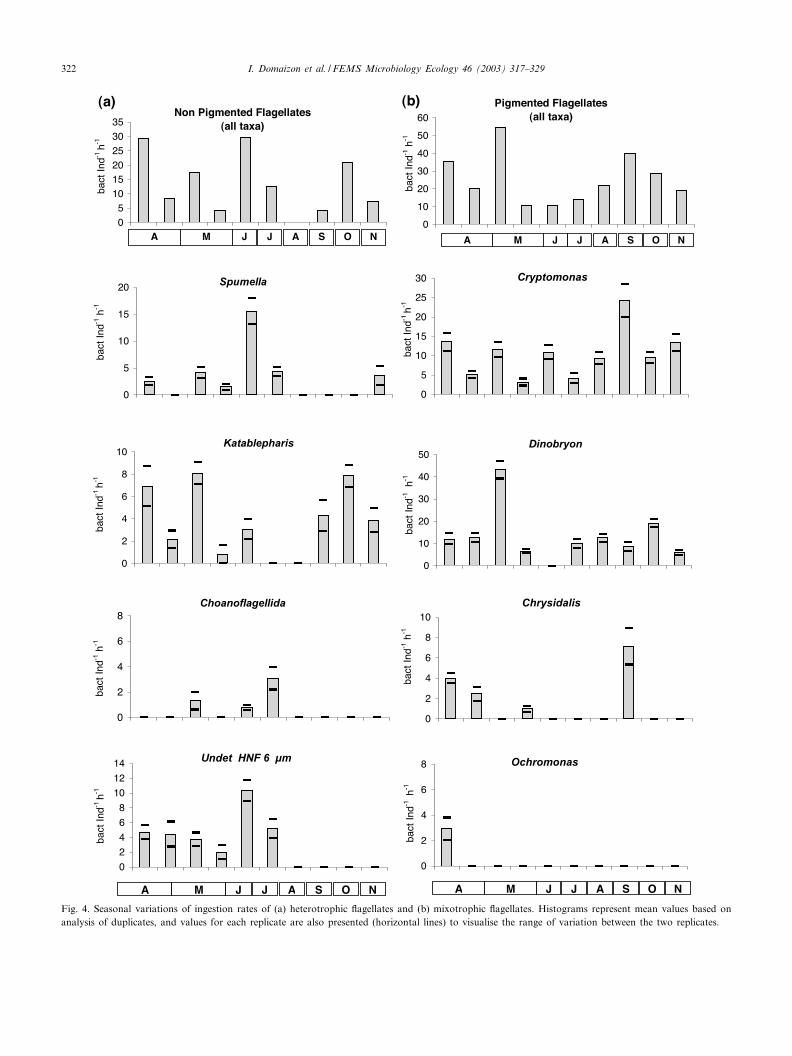
Fig. 4. Seasonal variations of ingestion rates of (a) heterotrophic £agellates and (b) mixotrophic £agellates. Histograms represent mean values based onanalysis of duplicates, and values for each replicate are also presented (horizontal lines) to visualise the range of variation between the two replicates.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329322
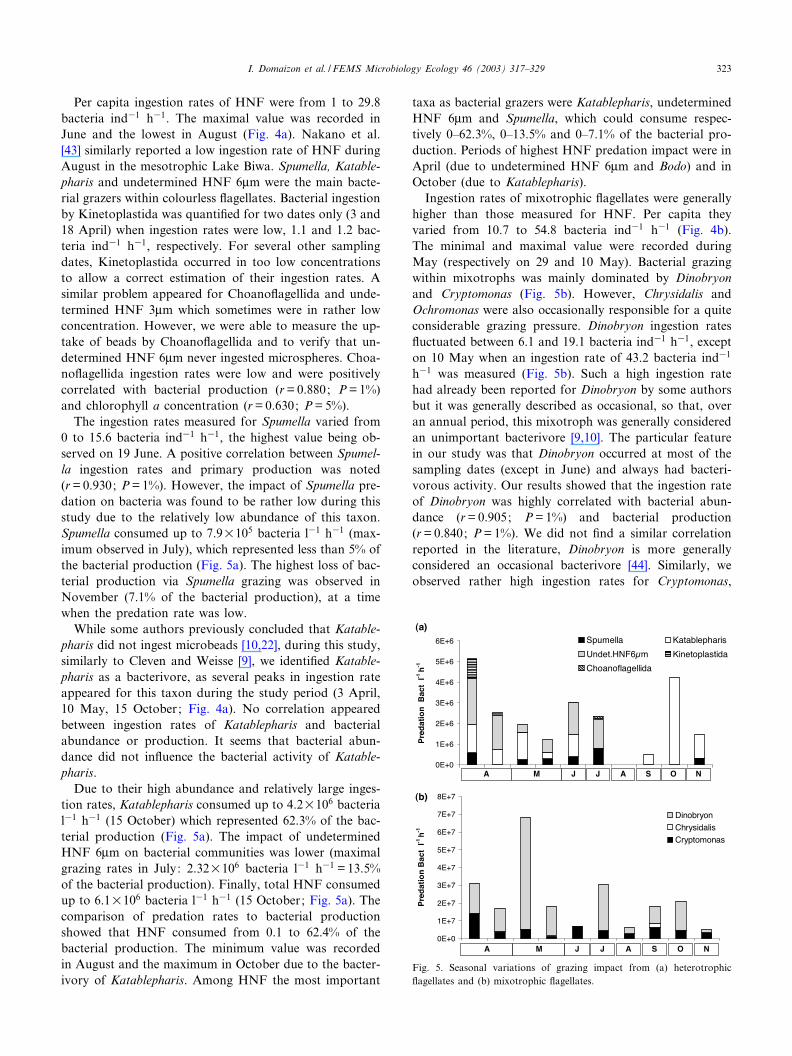
Per capita ingestion rates of HNF were from 1 to 29.8bacteria ind31 h31. The maximal value was recorded inJune and the lowest in August (Fig. 4a). Nakano et al.[43] similarly reported a low ingestion rate of HNF duringAugust in the mesotrophic Lake Biwa. Spumella, Katable-pharis and undetermined HNF 6Wm were the main bacte-rial grazers within colourless £agellates. Bacterial ingestionby Kinetoplastida was quanti¢ed for two dates only (3 and18 April) when ingestion rates were low, 1.1 and 1.2 bac-teria ind31 h31, respectively. For several other samplingdates, Kinetoplastida occurred in too low concentrationsto allow a correct estimation of their ingestion rates. Asimilar problem appeared for Choano£agellida and unde-termined HNF 3Wm which sometimes were in rather lowconcentration. However, we were able to measure the up-take of beads by Choano£agellida and to verify that un-determined HNF 6Wm never ingested microspheres. Choa-no£agellida ingestion rates were low and were positivelycorrelated with bacterial production (r= 0.880; P= 1%)and chlorophyll a concentration (r= 0.630; P= 5%).
The ingestion rates measured for Spumella varied from0 to 15.6 bacteria ind31 h31, the highest value being ob-served on 19 June. A positive correlation between Spumel-la ingestion rates and primary production was noted(r= 0.930; P= 1%). However, the impact of Spumella pre-dation on bacteria was found to be rather low during thisstudy due to the relatively low abundance of this taxon.Spumella consumed up to 7.9U105 bacteria l31 h31 (max-imum observed in July), which represented less than 5% ofthe bacterial production (Fig. 5a). The highest loss of bac-terial production via Spumella grazing was observed inNovember (7.1% of the bacterial production), at a timewhen the predation rate was low.
While some authors previously concluded that Katable-pharis did not ingest microbeads [10,22], during this study,similarly to Cleven and Weisse [9], we identi¢ed Katable-pharis as a bacterivore, as several peaks in ingestion rateappeared for this taxon during the study period (3 April,10 May, 15 October; Fig. 4a). No correlation appearedbetween ingestion rates of Katablepharis and bacterialabundance or production. It seems that bacterial abun-dance did not in£uence the bacterial activity of Katable-pharis.
Due to their high abundance and relatively large inges-tion rates, Katablepharis consumed up to 4.2U106 bacterial31 h31 (15 October) which represented 62.3% of the bac-terial production (Fig. 5a). The impact of undeterminedHNF 6Wm on bacterial communities was lower (maximalgrazing rates in July: 2.32U106 bacteria l31 h31 = 13.5%of the bacterial production). Finally, total HNF consumedup to 6.1U106 bacteria l31 h31 (15 October; Fig. 5a). Thecomparison of predation rates to bacterial productionshowed that HNF consumed from 0.1 to 62.4% of thebacterial production. The minimum value was recordedin August and the maximum in October due to the bacter-ivory of Katablepharis. Among HNF the most important
taxa as bacterial grazers were Katablepharis, undeterminedHNF 6Wm and Spumella, which could consume respec-tively 0^62.3%, 0^13.5% and 0^7.1% of the bacterial pro-duction. Periods of highest HNF predation impact were inApril (due to undetermined HNF 6Wm and Bodo) and inOctober (due to Katablepharis).
Ingestion rates of mixotrophic £agellates were generallyhigher than those measured for HNF. Per capita theyvaried from 10.7 to 54.8 bacteria ind31 h31 (Fig. 4b).The minimal and maximal value were recorded duringMay (respectively on 29 and 10 May). Bacterial grazingwithin mixotrophs was mainly dominated by Dinobryonand Cryptomonas (Fig. 5b). However, Chrysidalis andOchromonas were also occasionally responsible for a quiteconsiderable grazing pressure. Dinobryon ingestion rates£uctuated between 6.1 and 19.1 bacteria ind31 h31, excepton 10 May when an ingestion rate of 43.2 bacteria ind31
h31 was measured (Fig. 5b). Such a high ingestion ratehad already been reported for Dinobryon by some authorsbut it was generally described as occasional, so that, overan annual period, this mixotroph was generally consideredan unimportant bacterivore [9,10]. The particular featurein our study was that Dinobryon occurred at most of thesampling dates (except in June) and always had bacteri-vorous activity. Our results showed that the ingestion rateof Dinobryon was highly correlated with bacterial abun-dance (r= 0.905; P= 1%) and bacterial production(r= 0.840; P= 1%). We did not ¢nd a similar correlationreported in the literature, Dinobryon is more generallyconsidered an occasional bacterivore [44]. Similarly, weobserved rather high ingestion rates for Cryptomonas,
Fig. 5. Seasonal variations of grazing impact from (a) heterotrophic£agellates and (b) mixotrophic £agellates.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329 323

varying from 3 to 24 bacteria ind31 h31, whereas thisgenus is usually considered a negligible bacterivore [24]
During our study, due to their high abundance and in-gestion rates, mixotrophic £agellates, and particularly Di-nobryon, appeared as major bacterial grazers (Fig. 5a,b).Mixotrophs consumed up to 6.8U107 bacteria l31 h31
(maximum observed in April). A quite similar result wasobserved in July when mixotrophic £agellates were respon-sible for the loss of 49.1% of the bacterial production. Themean loss of bacterial production due to grazing by mix-otrophic £agellates was 152% ( M 98.2). On particular datesduring spring, late summer and autumn (May, August,September, October) grazing measurements showed thatmixotrophic £agellate grazing was higher than bacterialproduction, mainly due to the bacterivory of Dinobryon ;mixotrophic £agellates removed 193% of the bacterial pro-duction on 29 May. Of course, the rather large uncertaintyassociated with such comparisons of bacterial productionand grazing loss rates derived from microsphere or £uo-rescently labelled bacteria (FLB) experiments must be con-sidered. Any extrapolation of the data to discuss the reg-ulation impact of £agellates on bacteria should beconsidered in view of this uncertainty. However, our re-sults clearly demonstrated that mixotrophs were importantbacterial grazers in Lake Annecy.
4. Discussion
Our study is the ¢rst to provide information on thecomposition, dynamics, and grazing impact of £agellatesin the oligotrophic Lake Annecy. A few studies have in-vestigated bacterial grazing by HNF-speci¢c taxa, and ex-tremely rarely studies have dealt with taxon-speci¢c bac-terial grazing for both HNF and mixotrophic £agellates,particularly under oligotrophic conditions. Concentrationsof HNF measured in Lake Annecy were close to thosemeasured in other lakes [9,10,21]. The main di¡erenceconcerning this group was the large dominance of Cryp-tomonad forms (Katablepharis, undetermined HNF 6Wm)compared to Chrysomonad cells (Spumella) which wereusually reported as the dominant species within HNF[2,9,10,22,45]. As regards pigmented £agellates the speci¢cfeature in Lake Annecy was the numerical importanceof Dinobryon which was observed on nine out of the11 sampling dates, while in several lakes a high occur-rence of mixotrophic species and particularly Dinobryonwas restricted to a relatively short period during summer[9,46].
4.1. Potential shortcomings of the £uorescent microbeadsmethod
Bene¢ts and advantages of microspheres and FLB tech-niques have been extensively discussed in the literature[9,22,47^50]. In this study we used the tracer approach
as a ‘near standard’ method but of course, we have toconsider possible problems inherent in the technique.
First, some species may have had some selective grazingbehaviour against microspheres as arti¢cial preys, and theextrapolation of the data should be considered accord-ingly. However, Boegnik et al. [51] recently reported thatthe types of particles (inert or living) do not induce selec-tivity for interception-feeding bacterivorous nano£agel-lates. The authors showed that the introduction of beadsinto natural food (mixture of bacteria) did not modify theingestion rates of three di¡erent £agellates (Spumella,Ochromonas, Cafeteria) and that no signi¢cant di¡erenceappeared between beads and bacteria in terms of capturee⁄ciency. This study [51] concluded that selective fooduptake rather depended on food concentration for Spu-mella and Ochromonas. A signi¢cant selection of beads(beads rejected or non-ingested) appeared only for rela-tively high food concentrations (from 2.5U107 cells ml31
to 3.3U107 cells ml31). During our experiment we did notobserve such high concentrations of bacteria and we as-sumed that there was no strong selection between beadsand bacteria.
Secondly, a problem inherent in the use of microspheresis the possible quick egestion of ingested beads and FLBby some Chrysomonads as demonstrated by Boegnik et al.[52]. We relied on the conclusions of several authors whopreviously demonstrated that the number of microspheresingested by £agellates increases with time up to a maximalvalue [47,50,53]. They considered that during the linearincrease of particle numbers in vacuoles, there is no eges-tion of particles. After the plateau is reached, the particlenumber in £agellates is relatively constant through timeand represents a balance between ingestion and egestion.Based on this predation kinetics, the ingestion rates areassessed by using data in the linear part of the curve[47,50,53]. We used this kind of predation kinetics tochoose the time of incubation used during our experiment.Predation kinetics were performed on three dates (April,June, September) for the microbial communities in LakeAnnecy. The incubation times tested were 0, 5, 10, 15, 20,25 and 30 min. We observed a linear increase of the in-gested particle numbers during a period which rangedfrom 15 to 30 min according to taxa. During this linearincrease we considered that there was no egestion. Eges-tion appeared after 20 min for Dinobryon, while for Choa-no£agellates we observed a linear increase in ingested par-ticles for up to 30 min. For Spumella and Katablepharis,egestion appeared after 15 min and we noticed that noparticles were ingested by Spumella for a 5-min incuba-tion. As a result of these observations, for the seasonalstudy of grazing we chose to minimise handling e¡ectand counting, and we carried out the enumeration of in-gested microspheres for two incubation times only: 0 minand 15 min.
Thirdly, the size selectivity of bacterivorous £agellates isan often neglected problem in FLB or microspheres tech-
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329324

niques. We cannot exclude the possibility that larger bac-terivores such as Katablepharis prefer larger feed sizes anddo not e⁄ciently graze on small particles such as the mi-crobeads.
However, the use of microspheres still appeared to us tobe the best compromise for the simultaneous estimation ofbacterial consumption by £agellates (results presented inthis paper), ciliates and metazooplankton (Domaizon etal., in preparation), because beads resist digestion andare easy to see in guts or vacuoles. Associated with ataxonomic identi¢cation of grazers, this method has theprimary advantage of taking into account the variabilityof ingestion rate within each taxon and is especially usefulfor considering the still poorly studied bacterivorous ac-tivity of mixotrophic species.
4.2. Interest of taxon-speci¢c ingestion rates
We observed large di¡erences between taxon-speci¢c in-gestion rates of major £agellates (both heterotrophs andmixotrophs) and high seasonal variations. Per capita in-
gestion rates of HNF ranged from 0 to 30 bacteria HNF31
h31 ; Cleven and Weisse [9], who considered HNF grazingin Lake Constance by the FLB method, reported verysimilar values of ingestion rates (from 0 to 31 bacteriaind31 h31). As regards mixotrophs, we measured per cap-ita ingestion rates varying from 10 to 55 bacteria ind31
h31 which demonstrated that uptake rates by mixotrophsmay also greatly vary seasonally. In order to comparetaxon ingestion rates of £agellates we included this studyin literature data (Table 1), assuming that the micro-spheres technique and the alternative methods such asFLB were internally consistent, and ignoring problemsoriginating from di¡erent experimental protocols. As con-cerns HNF and more particularly Spumella, Choano£agel-lida, Bodonids, our data fall generally within the range buttoward the lower end of previously published data (Table1) [2,9,10,22,24,45,54^56]. In contrast, Katablepharis,which is generally considered a facultative bacterivore[10,22], appeared to have quite considerable bacterivorousactivity ; however, its bacterial uptake rates varied season-ally by a factor of up to 8. The larger range for seasonal
Table 1Ingestion rates of heterotrophic and mixotrophic £agellates obtained by various methods in lakes di¡ering in their trophic status
Ingestion rates (bacteria ind31 h31) Methoda Site Trophy degree Reference
Heterotrophic £agellatesSpumella-Monas-like cells 0^15.6 Microspheres Lake Annecy Oligotrophic this study
1.6^27 Microspheres Lake Pavin Oligo-mesotrophic [10]0^14.1 Microspheres Reservoir Sep Oligo-mesotrophic [22]10^14 FLB Lake Erie Mesotrophic [45]10^15 FLB Lake Constance Meso-eutrophic [24]
(Spumella 2^6 Wm) 0^5 FLB Lake Constance Meso-eutrophic [9](Spumella s 6 Wm) 6^31 FLB Lake Constance Meso-eutrophic [9]
14^38 Live observation [54]3^23 Microspheres Lake Oglethorpe Eutrophic [2]21 FLB Reservoir Rimov Eutrophic [55]
Katablepharis 0^8.1 Microspheres Lake Annecy Oligotrophic this study0^5 FLB Lake Constance Meso-eutrophic [9]
Choano£agellida 0^3.1 Microspheres Lake Annecy Oligotrophic this study1.7^33.6 Microspheres Lake Pavin Oligo-mesotrophic [10]0^11 FLB Lake Constance Meso-eutrophic [9]13^73 Microspheres [56]13^37 FLB Lake Erie Mesotrophic [45]8^42 Microspheres Lake Oglethorpe Eutrophic [2]53 FLB Reservoir Rimov Eutrophic [55]
Bodonids 0^10 Microspheres Lake Annecy Oligotrophic this study36 FLB Reservoir Rimov Eutrophic [55]
Mixotrophic £agellatesCryptomonas 3^24 Microspheres Lake Annecy Oligotrophic this study
0^14 Microspheres Reservoir Sep Oligo-mesotrophic [22]Dinobryon 6^43.2 Microspheres Lake Annecy Oligotrophic this study
2.4^35.3 Microspheres Lake Pavin Oligo-mesotrophic [10]63.2^137.6 Microspheres Reservoir Sep Oligo-mesotrophic [22]
D. bavaricum 8^38 Microspheres Lake Oglethorpe Eutrophic [2]D. cylindricum 6^12 Microspheres Lake Oglethorpe Eutrophic [2]
0^5.4 FLB Arti¢cial pond Eutrophic [16]Ochromonas 0^3 Microspheres Lake Annecy Oligotrophic this study
6.4^87 Microspheres Lake Pavin Oligo-mesotrophic [10]2^53 Microspheres Lake Oglethorpe Eutrophic [2]0^38.3 FLB Arti¢cial pond Eutrophic [16]
aFLB: £uorescent labelled bacteria
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329 325

variations was observed for Spumella within heterotrophic£agellates (from 0 to 15 bacteria ind31 h31) and for Di-nobryon within mixotrophs (from 6 to 43 bacteria ind31
h31).The large di¡erences between taxon-speci¢c ingestion
rates of major £agellates and high seasonal variationsshow that all HNF did not act similarly as primary bac-terivores. These results suggest that considering the taxo-nomic composition of £agellates may provide a betterunderstanding of trophic structures and thus energy £owwithin the microbial loop. When assessments of overallHNF grazing are made, they are generally based on thebacterial abundance/HNF abundance ratio. We have de-rived this type of overall evaluation from our data withthe aim of making a comparison with our taxon-speci¢cassessment of bacterial grazing. It is generally consideredthat a minimal proportion of 1000 bacteria per heterotro-phic £agellate characterises a microbial food web where£agellates preferentially consume bacteria [57]. Duringour study this ratio was generally higher than 1000 andvaried from 981 to 8275 (mean value: 3239) showing thatbacteria in the euphotic zone of Lake Annecy were abun-dant enough to support the development of the heterotro-phic £agellates that were present. Jugnia et al. [57] pro-posed using the formula established by Starink et al. [58]to evaluate the fraction of the bacterial production con-sumed by heterotrophic £agellates. When applied to ourdata, this formula suggested that the fraction of bacterialproduction consumed by HNF would vary from 20% to109% (annual mean 55%). The results we obtained fromthe microspheres method showed that HNF consumedfrom 0.1 to 62.4% of the bacterial production (annualmean: 17.9%). For most of the sampling dates, the impactof HNF predation measured by the microspheres methodwas then lower than that estimated from the ratio bacte-ria/HNF according to the Starink formula. The estimatedHNF predation based on the microsphere method washigher for one sampling date only, in October, whenboth the ingestion rates and the abundance of Katablepha-ris were particularly high.
The signi¢cantly di¡erent results given by these twoapproaches highlight the di⁄culty in comparing studieswhere impact of predation is assessed by di¡erent meth-ods.
4.3. Mixotrophy in the microbial loop of Lake Annecy
A few studies concluded that mixotrophic £agellates areunimportant bacterial grazers in freshwater pelagic foodwebs [9,10,46]. On the contrary, we observed that mixo-trophic £agellates could play an important role in the reg-ulation of bacterial communities in Lake Annecy.Although per capita ingestion rates of HNF could behigher than ingestion rates of mixotrophs, the total graz-ing impact of mixotrophs, particularly due to Dinobryon,and to a lesser extent to Cryptomonas, was most of the
time higher than the impact of heterotrophs. On average,only 17.9% ( M 19.2) of the bacterial production was re-moved by HNF during the study period, while the meanloss of bacterial production due to mixotrophs grazingwas 152% ( M 98.2). Cleven and Weisse [9] reported similarresults for the HNF grazing impact in Lake Constance.The authors observed that only 9.7% ( M 12.4%) of thebacterial production was removed by HNF on an annualaverage. They suggested that grazers other than smallHNF signi¢cantly contributed to the total bacterial mor-tality.
We assume that the high ingestion rates of mixotrophic£agellates in the epilimnion of Lake Annecy could be aconsequence of the low soluble reactive phosphorus (SRP)concentrations. Olrik [44] demonstrated that it is possiblefor mixotrophic Chrysophytes to solve their demand forphosphorus by ingestion of bacteria. The results of Hitch-man and Jones [16] con¢rmed this observation showingthat, in Lily Pond (England), the Dinobryon populationingested FLB to supplement their own phosphate require-ments when SRP was low. Similarly, our results pointedout that when Dinobryon was present, its bacterial inges-tion rate was always important under the P-limited con-ditions that are quite permanent in the oligotrophic LakeAnnecy (SRP concentrations varied from 1 to 2 Wg l31
during the study [26]). Bird and Kal¡ [15] showed thatDinobryon were also able to assimilate organic carbonfrom the ingestion of prey, in addition to other nutrientssuch as phosphorus. Such results suggested that the im-portance of mixotrophic protists as bacterial grazers couldbe higher in oligotrophic systems than in systems withhigher trophic status. However, Sanders et al. [2] demon-strated that mixotrophic £agellates could play a signi¢cantrole as bacterivores under eutrophic conditions. In theeutrophic lake they studied, mixotrophs displayed a strongseasonal impact, the highest values being recorded in win-ter and spring. In Lake Annecy, the maximal impact ofmixotroph bacterivory was recorded in August.
Our results cannot be compared to other oligotrophicsystems, due to the absence of available data on taxon-speci¢c grazing in oligotrophic freshwater systems. How-ever, some comparisons can be made with marine systemswhere several recent studies and reviews deal with theimportance of mixotrophs [17,20,21,58^61]. As in fresh-water systems, in marine environments mixotrophy is con-sidered an adaptation to low food environments in orderto supplement nutrients [21,58,60,61]. For example, Hav-skum and Riemann [21] demonstrated that bacterivorousmixotrophic £agellates were responsible for 86% of theentire £agellate bacterivory in the upper layer of the Bayof Aarhus (Denmark) which is characterised by low salin-ity and low inorganic nutrients. However, Dolan and Pe-rez [59], discussing the costs, bene¢ts and possible uses ofmixotrophy in marine oligotrichs, suggested that it couldnot be closely linked to the exploitation of low food envi-ronments, but probably serves a variety of purposes. Sim-
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329326

ilarly, Havskum and Hansen [20] concluded that the addi-tional phototrophic metabolism of mixotrophs could notbe only related to the availability of dissolved inorganicnutrients but probably gives the mixotrophs a better abil-ity to compete with strictly heterotrophic protists in envi-ronments where prey concentration or production is low.This point was investigated by Rothhaupt [62] who dem-onstrated that both £agellates (obligate bacterivores andmixotrophs) e¡ectively competed for food bacteria, show-ing that Spumella built up lower biomass compared to thelarger Ochromonas due to the supplementary photosyn-thetic capabilities of the latter.
The apparent contradictions in the literature as regardsthe role of mixotrophs are probably due to the variety ofnutritional strategies that exist within mixotrophic protiststhemselves [17,63,64]. Di¡erent types of mixotrophs havebeen described, based on the extent to which phototrophyand phagotrophy are used [18,63,64]. Some mixotrophsare primarily autotrophic while others mainly use hetero-trophy to ful¢l their energy requirements. Some authorshave de¢ned di¡erent physiological types within mixo-trophs based on their nutritional behaviour [18,63,64].Mixotrophic £agellates identi¢ed in Lake Annecy essen-tially belong to type II of the Stickney model [18] andmore exactly to subgroups IIA and IIB which, accordingto Stoecker, primarily concern phototrophic species thatcan have a bacterivorous activity when dissolved inorganicnitrogen is limiting (e.g.Dinobryon) or when a trace organ-ic growth factor is limiting (e.g. Cryptomonas). Jones [63]categorised Cryptomonas as a primarily phototrophic spe-cies that ingests preys only at very low rates to meet re-quirements for cell maintenance. In contrast, during ourstudy, rather high ingestion rates were measured for Cryp-tomonas compared to those measured for HNF taxa.
Most of the studies dealing with the identi¢cation andthe role of mixotrophic groups were conducted in marinesystems [17,18,20,59,60,65], and references on lakes arestill lacking; however, it is clear that, in all systems, mix-otrophs increase the complexity of nutrients and energy£ows by functioning as both producers and consumers.Our study demonstrated the important role that mixo-trophs could play in the regulation of bacterial commun-ities stressing the importance in developing a functionalpartitioning of all microbial compartments where mixo-trophs especially would be included, and which could beincorporated into general models of food webs to betterdistinguish possible carbon pathways within the pelagicfood webs.
Acknowledgements
We thank Ian Jones (CEH Windermere, UK) for cor-recting the English text. We also wish to thank Dr J.F.Carrias for his advice concerning methodology on grazingexperiments, J.C. Hustache and J.P. Moille from the Hy-
drobiology Laboratory of INRA Thonon for their valua-ble help during ¢eld samplings. This work was supportedby the Re¤gion Rho“ne-Alpes as part of the Contrat PlanEtat Re¤gion (CPER) ‘Durable management of Lake An-necy ^ Productivity and Functioning’ coordinated by DrD. Gerdeaux, CARRTEL-INRA Thonon (74) France.
References
[1] Riemann, B. and Christo¡ersen, K. (1993) Microbial trophodynamicsin temperate lakes. Mar. Microb. Food Webs 7, 69^100.
[2] Sanders, R.W., Porter, K.G., Bennett, S.J. and Debiase, A.E. (1989)Seasonal patterns of bacterivory by £agellates, ciliates, rotifers andcladocerans in a freshwater planktonic community. Limnol. Ocean-ogr. 34, 673^687.
[3] Porter, K.G., Sherr, E.B., Sherr, B.F., Pace, M.L. and Sanders, R.W.(1985) Protozoa in planktonic food webs. J. Protozool. 32, 409^415.
[4] Nakano, S. (2000) The role of protists in microbial loop of lakeecosystems. Jap. J. Ecol. 50, 41^51.
[5] Wieltschnig, C., Kirschner, A.K.T., Steitz, A. and Velimirov, B.(2001) Weak coupling between heterotrophic nano£agellates and bac-teria in a eutrophic freshwater environment. Microb. Ecol. 42, 159^167.
[6] Ju«rgens, K., Arndt, H. and Zimmermann, H. (1997) Impact of meta-ozan and protozoan grazers on bacterial biomass distribution in mi-crocosm experiments. Aquat. Microb. Ecol. 12, 131^138.
[7] Hahn, M.W. and Ho£e, M.G. (2001) Grazing of protozoa and itse¡ect on populations of aquatic bacteria. FEMS Microbiol. Ecol. 35,113^121.
[8] Callieri, C., Karjalainen, S.M. and Passoni, S. (2002) Grazing byciliates and heterotrophic nano£agellates on picocyanobacteria inLago Maggiore. J. Plankton Res. 24, 785^796.
[9] Cleven, A.J. and Weisse, T. (2001) Seasonal succession and taxonspeci¢c bacterial grazing rates of heterotrophic nano£agellates inLake Constance. Aquat. Microb. Ecol. 23, 147^161.
[10] Carrias, J.F., Amblard, C. and Bourdier, G. (1996) Protistan bacter-ivory in an oligomesotrophic lake: Importance of attached ciliatesand £agellates. Microb. Ecol. 31, 249^268.
[11] Pernthaler, A., Simek, K., Sattler, B., Schwarzenbacher, B., Bobkova¤,J. and Psenner, R. (1996) Short term changes of protozoan controlon autotrophic picoplankton in an oligomesotrophic lake. J. Plank-ton Res. 18, 443^462.
[12] Sanders, R.W., Caron, D.A. and Berninger, U.G. (1992) Relation-ships between bacteria and heterotrophic nanoplankton in marineand fresh waters: an inter-ecosystem comparison. Mar. Ecol. Prog.Ser. 86, 1^14.
[13] Gasol, J.M. (1994) A framework for the assessment of top-down vsbottom-up control of heterotrophic nano£agellate abundance. Mar.Ecol. Prog. Ser. 113, 291^300.
[14] Sherr, E.B. and Sherr, B.F. (1994) Bacterivory and herbivory: Keyroles of phagotrophic protists in pelagic food webs. Microb. Ecol. 28,235.
[15] Bird, F.B. and Kal¡, J. (1986) Bacterial grazing by planktonic lakealgae. Science 231, 493^495.
[16] Hitchman, R.B. and Jones, H.L.J. (2000) The role of mixotrophicprotists in the population dynamics of the microbial food web in asmall arti¢cial ponds. Freshwater Biol. 43, 231^241.
[17] Sanders, R.W., Berninger, U.G., Lim, E.L., Kemp, P.F. and Caron,D.A. (2000) Heterotrophic and mixotrophic nanoplankton predationin the Sargasso Sea and on Georges Bank. Mar. Ecol. Prog. Ser. 192,103^118.
[18] Stickney, H.L., Hood, R.R. and Stoecker, D.K. (2000) The impact ofmixotrophy on planktonic marine ecosystems. Ecol. Modell. 125,203^230.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329 327

[19] Bouvier, T., Becquevort, S. and Lancelot, C. (1998) Biomass andfeeding activity of phagotrophic mixotrophs in the north-westernBlack Sea during the summer 1995. Hydrobiologia 363, 289^301.
[20] Havskum, H. and Hansen, A.S. (1997) Importance of pigmented andcolourless nanosized protists as grazers on nanoplakton in a phos-phate-depleted Norwegian fjord and in enclosures. Aquat. Microb.Ecol. 12, 139^151.
[21] Havskum, H. and Riemann, B. (1996) Ecological importance of bac-terivorous pigmented £agellates (mixotrophs) in the Bay of Aarhus,Denmark. Mar. Ecol. Prog. Ser. 137, 251^263.
[22] Thouvenot, A., Richardot, M., Debroas, D. and De¤vaux, J. (1999)Bacterivory of metazooplankton, ciliates and £agellates in a newly£ooded reservoir. J. Plankton Res. 21, 1659^1679.
[23] Simek, K., Vrba, J., Pernthaler, J., Posch, T., Hartman, P., Nedoma,J. and Psenner, R. (1997) Morphological and composition shifts in anexperimental bacterial community in£uenced by protists with con-trasting feeding modes. Appl. Environ. Microbiol. 63, 587^595.
[24] Ju«rgens, K. and Gu«de, H. (1991) Seasonal changes in the grazingimpact of phagotrophic £agellates on bacteria in Lake Constance.Mar. Microb. Food Webs 5, 27^37.
[25] Mouthon, J. and Dubois, J.P. (2001) Mollusc communities of thelittoral zone of Lake Annecy (Savoie, France). Ann. Limnol. 37,267^276.
[26] Balvay, G., Lazzarotto, J., Druart, J.C. and Guichard, V. (2002)Suivi de la qualite¤ des eaux du lac d’Annecy en 2001. Report SILAAnnecy, INRA-Thonon 220-2002.
[27] Fuhrman, J.A. and Azam, F. (1982) Thymidine incorporation as ameasure of heterotrophic bacterioplankton production in marine sur-face waters: Evaluation and ¢eld results. Mar. Biol. 66, 109^120.
[28] Stro¡ek, S. (1990) Les transferts verticaux de matie're et leur modi¢-cation par les bacte¤ries he¤te¤rotrophes ¢xe¤es sur particules en se¤dimen-tation dans les eaux de surface de deux grands lacs alpins (lac Le¤man,lac du Bourget, France). Thesis, Universite¤ Claude Bernard.
[29] Steemann Nielsen, E. (1952) The use of radioactive carbon (14C) formeasuring organic production in the sea. J. Cons. Int. Exploit. Mer18, 117^140.
[30] Steemann Nielsen, E. (1977) The carbon 14 technique for measuringorganic production by plankton algae. A report of the present knowl-edge. Folia Limnol. Scand., 17^45.
[31] Porter, K.G. and Feig, Y.S. (1980) The use of DAPI for identifyingand counting aquatic micro£ora. Limnol. Oceanogr. 25, 943^948.
[32] Simon, M. and Azam, F. (1989) Protein content and protein syn-thesis rates of planktonic marine bacteria. Mar. Ecol. Prog. Ser. 51,201.
[33] Caron, D.A. (1983) Technique for enumeration of heterotrophic andphototrophic nanoplankton, using epi£uorescence microscopy, andcomparison with other procedure. Appl. Environ. Microbiol. 46,491^498.
[34] Bloem, J., Ba«r-Gilissen, M.J.B. and Carpenter, T.E. (1986) Fixation,counting, and manipulation of heterotrophic nano£agellates. Appl.Environ. Microbiol. 52, 1266^1272.
[35] B[rsheim, K.Y. and Bratbak, G. (1987) Cell volume to cell carbonconversion factors for a bacterivorous monas sp. enriched from sea-water. Mar. Ecol. Prog. Ser. 36, 171^175.
[36] Pace, M.L. and Bailif, M.D. (1987) Evaluation of a £uorescent mi-crosphere technique for measuring grazing rates of phagotrophic mi-croorganisms. Mar. Ecol. Prog. Ser. 40, 185^193.
[37] Balvay, G., Blanc, P., Druart, J.C. and Guichard, V. (1999) Suivi dela qualite¤ du lac d’Annecy en 1998. Report SILA Annecy, INRA-Thonon 161-99.
[38] Balvay, G., Lazzarotto, J., Druart, J.C. and Guichard, V. (2001)Suivi de la qualite¤ des eaux du lac d’Annecy en 2000. Report SILAAnnecy, INRA-Thonon 201-2001.
[39] Auer, B. and Arndt, H. (2001) Taxonomic composition and biomassof heterotrophic £agellates in relation to lake trophy and season.Freshwater Biol. 46, 959^972.
[40] Doddema, H. and Van Der Veer, J. (1983) Ochromonas monicis sp.
nov. A particle feeder with bacterial endosymbionts. Cryptogam. Al-gol. 4, 89^97.
[41] Domaizon, I. and De¤vaux, J. (1999) Experimental study of the im-pacts of silver carp on plankton communities of eutrophic Villeretreservoir (France). Aquat. Ecol. 33, 193^204.
[42] Pick, F.R., Nalewajko, C. and Lean, D.R.S. (1984) The origin of ametalimnic chrysophyte peak. Limnol. Oceanogr. 29, 125^134.
[43] Nakano, S., Koitabashi, T. and Ueda, T. (1998) Seasonal changes inabundance of heterotrophic nano£agellates and their consumption ofbacteria in lake Biwa with special reference to trophic interactionswith Daphnia galeata. Arch. Hydrobiol. 142, 21^34.
[44] Olrik, K. (1998) Ecolgy of mixotrophic £agellates with special refer-ence to Chrysophyceae in Danish lakes. Hydrobiologia 369/370, 329^338.
[45] Hwang, S.G. and Health, R.T. (1997) Bacterial productivity andprotistan bacterivory in coastal and o¡shore communities of LakeErie. Can. J. Fish. Aquat. Sci. 54, 788^799.
[46] Weisse, T. and Mu«ller, H. (1998) Planktonic protozoa and the micro-bial food web in Lake Constance. Arch. Hydrobiol. Spec. Issue Ad-vanc. Limnol. 53, 223^254.
[47] Carrias, J.F. (1996) La boucle microbienne en milieu lacustre: Struc-ture et fonctionnement des communaute¤s picoplanctoniques et deprotistes £agelle¤s et cilie¤s. Thesis, Universite¤ Blaise Pascal.
[48] Landry, M.R. (1994) Methods and controls for measuring the graz-ing impact of planktonic protists. Mar. Microb. Food Webs 8, 37^57.
[49] Vaque¤, D., Gasol, J.M. and Marasse¤, C. (1994) Grazing rates onbacteria: the signi¢cance of methodology and ecological factors.Mar. Ecol. Prog. Ser. 109, 263^274.
[50] McManus, G.B. and Okubo, A. (1991) On the use of surrogate foodparticles to measure protistan ingestion. Limnol. Oceanogr. 36, 613^617.
[51] Boegnik, J., Matz, C., Ju«rgens, K. and Arndt, H. (2002) Food con-centration-dependent regulation of food selectivity of interception-feeding bacterivorous nano£agellates. Aquat. Microb. Ecol. 27,195^202.
[52] Boegnik, J., Matz, C., Ju«rgens, K. and Arndt, H. (2001) Confusingselective feeding with di¡erential digestion in bacterivorous nano£a-gellates. J. Eukaryot. Microbiol. 48, 425^432.
[53] Sherr, B.F., Sherr, E.B. and Fallon, R.D. (1987) Use of monodis-persed £uorescently labeled bacteria to estimate in situ protozoanbacterivory. Appl. Environ. Microbiol. 53, 958^965.
[54] Holen, D.A. and Boraas, M.E. (1991) The feeding behavior of Spu-mella sp. as a function of particle size implications for bacterial sizein pelagic systems. Hydrobiologia 220, 73^88.
[55] Simek, K., Hartman, P., Nedoma, J., Pernthaler, J., Springmann, D.,Vrba, J. and Psenner, R. (1997) Community structure, picoplanktongrazing and zooplankton control of heterotrophic nano£agellatres ina eutrophic reservoir during the summer phytoplankton maximum.Aquat. Microb. Ecol. 12, 49^63.
[56] Vaque¤, D. and Pace, L.M. (1992) Grazing on bacteria by £agellatesand cladocerans in lakes of contrasting food-web structure. J. Plank-ton Res. 14, 307^321.
[57] Jugnia, L.B., Tadonle¤ke¤, R.D., Sime-Ngando, T. and De¤vaux, J.(2000) The microbial food web in the recently £ooded Sep reservoir:Diel £uctuations in bacterial biomass and metabolitic activity in re-lation to phytoplankton and £agellate grazers. Microb. Ecol. 40, 317^329.
[58] Samuelsson, K., Berglund, J., Haeck, P. and Andersson, A. (2002)Structural changes in an aquatic microbial food web caused by in-organic nutrient addition. Aquat. Microb. Ecol. 29, 29^38.
[59] Dolan, J.R. and Perez, M.T. (2000) Costs, bene¢ts and characteristicsof mixotrophy in marine Oligotrichs. Freshwater Biol. 45, 227^238.
[60] Christaki, U., Van Wambeke, F. and Dolan, J.R. (1999) Nano£agel-lates (mixotrophs, heterotrophs, and autotrophs) in the oligotrophiceastern Mediterranean: standing stocks, bacterivory and relation-ships with bacterial production. Mar. Ecol. Prog. Ser. 181, 297^307.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329328

[61] Nygaard, K. and Tobiesen, A. (1993) Bacterivory in algae: a survivalstrategy during nutrient limitation. Limnol. Oceanogr. 38, 273^279.
[62] Rothhaupt, K.O. (1996) Laboratory experiments with a mixotrophicchrysophyte and obligately phagotrophic and phototrophic compet-itors. Ecology 77, 716^724.
[63] Jones, R.I. (2000) Mixotrophy in planktonic protist : an overview.Freshwater Biol. 45, 219^226.
[64] Stoecker, D.K. (1998) Conceptual models of mixotrophy in plank-tonic protists and some ecological and evolutionary implications.Eur. J. Protistol. 34, 281^290.
[65] Bockstahler, K.R. and Coats, D.W. (1993) Grazing of the mixotro-phic dino£agellate Gymnodinium sanguineum on ciliate populations ofChesapeake Bay. Mar. Biol. 116, 477^487.
FEMSEC 1591 26-11-03
I. Domaizon et al. / FEMS Microbiology Ecology 46 (2003) 317^329 329
Related Documents











