Research paper Targeting of Escherichia coli F4 fimbriae to Fcg receptors enhances the maturation of porcine dendritic cells Bert Devriendt a, *, Frank Verdonck a,b , Artur Summerfield d , Bruno M. Goddeeris a,c , Eric Cox a a Laboratory of Immunology, Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium b present affiliation: Ablynx nv, Technologiepark 4, 9052 Zwijnaarde, Belgium c Department Biosystems, Division Gene Technology, Faculty of Bioscience Engineering, KULeuven, Kasteelpark Arenberg 30, 3001 Heverlee, Belgium d Institute of Virology and Immunoprophylaxis, Sensemattstrasse 293, 3147 Mittelha ¨usern, Switzerland 1. Introduction Dendritic cells (DC) represent the most potent antigen presenting cells which are pivotal for the generation of adaptive immune responses and the defence against pathogens (Banchereau et al., 2000). Immature DC take up antigens (Ags) in the peripheral tissues, process them and then migrate to the lymph nodes to interact with and activate CD4 + and CD8 + naive T cells (Guermonprez et al., 2002). This process requires the maturation of DC, which involves not only the upregulation of several cell surface markers, such as MHCII, CD40, CD80 and CD86, but also a reduced Ag internalization capacity. In the T cell area of the local lymph nodes mature DC can activate T cells through the secretion of T cell polarizing cytokines and chemo- kines, which can influence the outcome of an immune response (Banchereau et al., 2000). DC take up extra- cellular Ag either by non-specific macropinocytosis or Veterinary Immunology and Immunopathology 135 (2010) 188–198 ARTICLE INFO Article history: Received 7 September 2009 Received in revised form 17 November 2009 Accepted 23 November 2009 Keywords: Porcine MoDC Fc receptors F4 fimbriae Immune complex ABSTRACT F4 + enterotoxigenic Escherichia coli (ETEC) infections are an important cause of postweaning diarrhoea in piglets and an oral immunization of piglets with purified F4 fimbriae protects them from a subsequent F4 + ETEC infection. However, oral immuniza- tion of suckling piglets is hampered due to the immature status of their immune system. Targeting of antigens to Fcg receptors (FcgR) on human and murine dendritic cells (DC) has been shown to enhance DC maturation and both humoral and cellular immune responses. To investigate the effect of F4 fimbriae incorporated in immune complexes (F4- IC) on porcine DC, we used porcine monocytic-derived DC (MoDC) as a model system. The results in this study demonstrate that FcgRI, II and III mRNA is expressed by porcine MoDC. Furthermore, we show that FcgRII and III are expressed on the cell surface and that F4-IC are internalized by MoDC via FcgR. This FcgR ligation induced a significantly enhanced expression of Major Histocompatibility complex (MHCII) class II and the costimulatory molecules CD80/86 and CD40 by MoDC compared with immature MoDC. Furthermore, the phagocytic capacity of F4-IC stimulated MoDC was reduced as evidenced by a reduced uptake of DQ-ovalbumin and FITC-dextran. In an allogenic and autologous mixed lymphocyte reaction, these F4-IC-activated MoDC showed an improved T cell stimulatory capacity in comparison with immature MoDC. The F4-IC induced DC maturation correlated with significant higher expression levels of several pro-inflammatory cytokines such as interleukine (IL) 1b, IL-6 and Tumor necrosis factor a, the chemokine IL-8 and IL- 12p40 in comparison with immature MoDC. Altogether, these results clearly demonstrate that FcgR engagement enhances the maturation of porcine MoDC, which may suggest that antigen targeting to FcgR on DC could improve vaccine design against infections. ß 2009 Elsevier B.V. All rights reserved. * Corresponding author. Tel.: +32 9 2647397; fax: +32 9 2647779. E-mail address: [email protected] (B. Devriendt). Contents lists available at ScienceDirect Veterinary Immunology and Immunopathology journal homepage: www.elsevier.com/locate/vetimm 0165-2427/$ – see front matter ß 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.vetimm.2009.11.013

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Veterinary Immunology and Immunopathology 135 (2010) 188–198
Research paper
Targeting of Escherichia coli F4 fimbriae to Fcg receptors enhances thematuration of porcine dendritic cells
Bert Devriendt a,*, Frank Verdonck a,b, Artur Summerfield d, Bruno M. Goddeeris a,c, Eric Cox a
a Laboratory of Immunology, Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgiumb present affiliation: Ablynx nv, Technologiepark 4, 9052 Zwijnaarde, Belgiumc Department Biosystems, Division Gene Technology, Faculty of Bioscience Engineering, KULeuven, Kasteelpark Arenberg 30, 3001 Heverlee, Belgiumd Institute of Virology and Immunoprophylaxis, Sensemattstrasse 293, 3147 Mittelhausern, Switzerland
A R T I C L E I N F O
Article history:
Received 7 September 2009
Received in revised form 17 November 2009
Accepted 23 November 2009
Keywords:
Porcine MoDC
Fc receptors
F4 fimbriae
Immune complex
A B S T R A C T
F4+ enterotoxigenic Escherichia coli (ETEC) infections are an important cause of
postweaning diarrhoea in piglets and an oral immunization of piglets with purified F4
fimbriae protects them from a subsequent F4+ ETEC infection. However, oral immuniza-
tion of suckling piglets is hampered due to the immature status of their immune system.
Targeting of antigens to Fcg receptors (FcgR) on human and murine dendritic cells (DC)
has been shown to enhance DC maturation and both humoral and cellular immune
responses. To investigate the effect of F4 fimbriae incorporated in immune complexes (F4-
IC) on porcine DC, we used porcine monocytic-derived DC (MoDC) as a model system. The
results in this study demonstrate that FcgRI, II and III mRNA is expressed by porcine MoDC.
Furthermore, we show that FcgRII and III are expressed on the cell surface and that F4-IC
are internalized by MoDC via FcgR. This FcgR ligation induced a significantly enhanced
expression of Major Histocompatibility complex (MHCII) class II and the costimulatory
molecules CD80/86 and CD40 by MoDC compared with immature MoDC. Furthermore, the
phagocytic capacity of F4-IC stimulated MoDC was reduced as evidenced by a reduced
uptake of DQ-ovalbumin and FITC-dextran. In an allogenic and autologous mixed
lymphocyte reaction, these F4-IC-activated MoDC showed an improved T cell stimulatory
capacity in comparison with immature MoDC. The F4-IC induced DC maturation
correlated with significant higher expression levels of several pro-inflammatory cytokines
such as interleukine (IL) 1b, IL-6 and Tumor necrosis factor a, the chemokine IL-8 and IL-
12p40 in comparison with immature MoDC. Altogether, these results clearly demonstrate
that FcgR engagement enhances the maturation of porcine MoDC, which may suggest that
antigen targeting to FcgR on DC could improve vaccine design against infections.
� 2009 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Veterinary Immunology and Immunopathology
journal homepage: www.e lsev ier .com/ locate /vet imm
1. Introduction
Dendritic cells (DC) represent the most potent antigenpresenting cells which are pivotal for the generation ofadaptive immune responses and the defence againstpathogens (Banchereau et al., 2000). Immature DC takeup antigens (Ags) in the peripheral tissues, process them
* Corresponding author. Tel.: +32 9 2647397; fax: +32 9 2647779.
E-mail address: [email protected] (B. Devriendt).
0165-2427/$ – see front matter � 2009 Elsevier B.V. All rights reserved.
doi:10.1016/j.vetimm.2009.11.013
and then migrate to the lymph nodes to interact with andactivate CD4+ and CD8+ naive T cells (Guermonprez et al.,2002). This process requires the maturation of DC, whichinvolves not only the upregulation of several cell surfacemarkers, such as MHCII, CD40, CD80 and CD86, but also areduced Ag internalization capacity. In the T cell area of thelocal lymph nodes mature DC can activate T cells throughthe secretion of T cell polarizing cytokines and chemo-kines, which can influence the outcome of an immuneresponse (Banchereau et al., 2000). DC take up extra-cellular Ag either by non-specific macropinocytosis or

B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198 189
more efficiently by receptor-mediated endocytosis (Sal-lusto and Lanzavecchia, 1994; Paillot et al., 2001). Differentreceptors are involved in antigen uptake by DC such as C-type lectins, scavenger receptors and FcR (FcgR, FceR, andFcaR, which bind, respectively, IgG, IgE and IgA). FcgR are afamily of membrane glycoproteins, belonging to the Igsuperfamily, which bind the Fc portion of IgG antibodies(Regnault et al., 1999). They can be divided into threesubclasses: FcgRI (CD64), FcgRII (CD32) and FcgRIII(CD16), which differ in binding affinity and subclass-specificity. FcgRI is a high affinity receptor, which bindsboth monomeric IgG and immune complexes, while FcgRIIand FcgRIII are low affinity receptors that only bindantigen complexed IgG. Both FcgRI and FcgRIII areactivating receptors, characterized by the presence of animmunoreceptor tyrosine-based activation motif (ITAM)motif, either intrinsic to the receptor or as part of anassociated subunit, the g chain (Ravetch and Bolland,2001). In contrast to FcgRIIA, which is an activatingreceptor, FcgRIIB is an inhibitory receptor, characterizedby an immunoreceptor tyrosine-based inhibition motif(ITIM), in its cytoplasmic domain (Ravetch and Bolland,2001). Activating and inhibitory FcgR mediate opposingfunctions, underlining the role of these receptors inregulating adaptive immune responses (Boruchov et al.,2005). In pig, all three FcgR subclasses are expressed(Halloran et al., 1994; Zhang et al., 2006; Qiao et al., 2006),however, hitherto only FcgRII and FcgRIII expression hasbeen demonstrated in porcine DC (Haverson et al., 2000;Carrasco et al., 2001; Balmelli et al., 2005; Bimczok et al.,2006). Previous studies with human and murine DC havedemonstrated that Ags incorporated in immune complexes(IC) are endocytosed more efficiently than soluble Agalone. Furthermore, this FcgR engagement by IC inducesthe maturation of DC, resulting in a strongly enhancedefficiency of Ag presentation to T cells (Liu et al., 2006;Regnault et al., 1999; Sedlik et al., 2003; Yada et al., 2003).To investigate if porcine DC react to FcgR targeting in asimilar manner as their human and murine counterparts,we used our F4 model antigen (Cox et al., 2002) to targetFcgR on porcine MoDC.
F4 fimbriae are proteinaceous filamentous adhesins,composed of repeating copies of the major fimbrialsubunit FaeG and some additional minor subunits (Bakkeret al., 1992a, 1992b), which are expressed on the surface ofF4+ enterotoxigenic Escherichia coli (ETEC). F4+ ETECinfections are an important cause of diarrhea in neonatal(Alexander, 1994) and recently weaned piglets (Hampson,1994). They adhere with their F4 fimbriae to F4-specificreceptors on the intestinal epithelium, subsequentlycolonize the small intestine and induce diarrhea. F4fimbriae are strong immunogenic antigens, since an oralimmunization of weaned piglets with purified F4 fimbriaeinduces a potent F4-specific intestinal immune responseprotecting the animals against a subsequent F4+ ETECchallenge infection (Van den Broeck et al., 1999a, 1999b;Verdonck et al., 2004). Recently, we demonstrated thatCD11R1+ intestinal DC are involved in the induction of amucosal immune response against F4+ ETEC infections(Devriendt et al., 2009). Although intestinal DC areprobably the prime target cells for orally administered
Ag/IgG immune complexes, they are not easily accessiblefor in vitro studies. In contrast, MoDC are a well-established model system in swine research (Carrascoet al., 2001; Paillot et al., 2001; Johansson et al., 2003;Bimczok et al., 2006, 2007; Pilon et al., 2009). Our resultsclearly show an enhanced functional maturation ofporcine MoDC after targeting F4 fimbriae to FcgR usingF4-specific antibodies, resulting in an enhanced produc-tion of the pro-inflammatory cytokines IL-1b, IL-6 andTNFa, the chemokine IL-8 and IL-12p40 in comparisonwith immature, F4- and IgG-stimulated MoDC. Further-more, we provide evidence that porcine MoDC mature inresponse to flagellin stimulation.
2. Materials and methods
2.1. Animals and blood samples
Heparinized blood samples were obtained from theexternal jugular vein of Belgian Landrace pigs kept as blooddonors under standard conditions at the Faculty ofVeterinary Medicine, Merelbeke, Belgium. All animalexperiments were in accordance with the local animalwelfare regulations of the Faculty of Veterinary Medicine.
2.2. Generation of F4/IgG immune complexes
F4 fimbriae were purified from the E. coli strain IMM01as previously described by Van den Broeck et al. (1999a).Briefly, bacteria were grown in Trypsone Soya Broth (TSB;Oxoid LTD., Basingstoke, Hampshire, UK) for 18 h at 37 8Cand 85 rpm. Subsequently, the F4 fimbriae were isolatedfrom the bacteria by mechanical shearing. After precipita-tion through addition of ammonium sulphate (40%, w/v),the fimbrial proteins were dialysed and filtrated (0.2 mm),the protein concentration was determined and the puritywas assessed by SDS-PAGE and Western blotting. Flagellinwas isolated from strain GIS26DFaeG using the sameprotocol as to purify F4 fimbriae. This strain is derived fromthe F4+ ETEC reference strain GIS26 by disruption of thegene encoding the major F4 fimbrial subunit FaeG(Verdonck et al., 2008). Porcine anti-F4 polyclonal Abswere affinity purified from heat-inactivated porcinehyperimmune serum, obtained from a pig intramuscularlyimmunized with purified F4 fimbriae. Different concen-trations of F4 fimbriae and IgG were incubated for 1 h at37 8C and the resulting F4-IC were analysed by multipleELISAs to determine the optimal concentrations at whichF4 fimbriae are completely covered with IgG, withoutprecipitation of F4-IC (data not shown).
2.3. Isolation and culture of cells
PBMC were isolated from whole blood by lymphoprepdensity gradient centrifugation. Monocytes were furtherenriched to a purity of >90% by positive immunomagneticbead selection (MACS; Miltenyi Biotec, Bergisch Gladbach,Germany) using the anti-CD172a mAb (clone: 74-12-15a;Pescovitz et al., 1984) and goat anti-mouse microbeadstogether with LS separation columns (MACS, MiltenyiBiotec). Cells were cultured in 24-well plates (Nunc,

B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198190
Thermo Fisher Scientific, Langenselbold, Germany) at adensity of 5� 105 cells/ml in phenol-red free Dulbecco’smodified Eagle’s Medium (DMEM; Gibco, Merelbeke,Belgium) supplemented with 10% (v/v) FCS (Greiner),100 U/ml penicillin (Gibco), 100 mg/ml streptomycin(Gibco), recombinant porcine (rp) GM-CSF (Inumaruet al., 1998) and rpIL-4 (prepared in our laboratory;Carrasco et al., 2001) and incubated at 37 8C in a humidifiedatmosphere at 5% CO2 to generate monocytic-deriveddendritic cells (MoDC) as previously described (Carrascoet al., 2001). On day 3 of the culture period, MoDC were fedby addition of fresh medium supplemented with rpGM-CSF and rpIL-4 at the same concentrations. On day 4 of theculture period, MoDC were treated for an additional 24 hwith different antigens: E. coli-derived LPS (1 mg/ml)(serotype O55:B5; Sigma, Bornem, Belgium), cholera toxin(CT) (1 mg/ml; Sigma), flagellin (50 mg/ml), F4 fimbriae(5 mg/ml), IgG (234 mg/ml) and F4-IC (5 mg/mlF4 + 234 mg/ml IgG), or were left untreated. After thisincubation period, MoDC were further processed forphenotyping, proliferation assays or cytokine expressionlevels as described below.
2.4. Analysis of FcgR mRNA expression by MoDC
Total RNA was isolated from 107 homogenized imma-ture MoDC using TRizol reagent (Invitrogen, Merelbeke,Belgium) and the RNeasy Mini kit (Qiagen, Venlo, TheNetherlands). RNA samples were treated with DNase I(Qiagen) and the absence of genomic DNA was verified byusing RNA samples as template in a PCR. The RNA wasquantified by measuring the optical density at 260 nm(OD260) and the OD260/OD280 and OD260/OD230 ratio’s werecalculated to assess RNA purity. RNA (3 mg) was convertedto cDNA and the FcgR mRNA expression was assessed withporcine FcgR-specific primers (Table 1). TLR5 expressionwas analysed with porcine TLR5-specific primers (Table 1).cDNA samples were denatured at 94 8C for 2 min, followedby 35 cycles of denaturing at 94 8C for 30 s, annealingtemperature (Table 1) for 30 s and elongation at 72 8C. ThemRNA expression of the glyceraldehyde-3-phosphatedehydrogenase (GAPDH) reference gene (Table 1) wasanalysed on the same samples as previously described(Melkebeek et al., 2007).
Table 1
Primer sequences, annealing temperature (Ta), amplicon size and accession nu
Gene symbol and (Acc number) Primer sequence
FcgRI F: GTGCTGAGAGCATCCTT
(DQ026063) R: CGCCTCACACCAGTATA
FcgRII F: GCTATTCCTGGCTCCTG
(DQ026064) R: AGTCTGGCACCTGTAGT
FcgRIII F: GCTCTGCTGCTTCTGGT
(AF372453) R: CTAGTCGCAATGGGTCA
TLR5 F: TTTCTGGCAATGGCTGG
(AB208697) R: TGGAGGTTGTCAAGTCC
GAPDH F: GGGCATGAACCATGAG
(AF017079) R: AAGCAGGGATGATGTT
Acc number, accession number; GAPDH, glyceraldehyde-3-phosphate dehydro
2.5. Analysis of FcgR-dependent binding and internalization
of F4-IC
IC are internalized through FcgR present on antigenpresenting cells. To determine if FcgR are involved in F4-ICuptake, purified F4 fimbriae were conjugated with 5(6)-carboxyfluorescein-N-hydroxysuccinimide ester (FluoS,480 Da) using the fluorescein labelling kit (Roche Diag-nostics, Basel, Switzerland). F4-FluoS fimbriae (1 mg) wereincubated with anti-F4 IgG (46 mg) for 1 h at 37 8C togenerate fluorescent F4-IC. Conjugation of F4 fimbriae withFluoS has no effect on IC formation (data not shown).Immature MoDC were harvested, washed and incubatedwith different concentrations of F4-FluoS for 5, 15, 30 or60 min at 4 8C or 37 8C to monitor the binding andinternalization of F4 fimbriae. To assay F4-IC bindingand internalization, immature MoDC were incubated withfluorescent F4-IC for 5, 15, or 30 min at 37 8C or 4 8C. For theinhibition experiments, immature MoDC were preincu-bated for 30 min at 4 8C with 50 mg/ml non-specificporcine monomeric IgG or heat-aggregated IgG (NordicImmunology, Tilburg, The Netherlands), after whichfluorescent F4-IC were added at the same time intervalsas mentioned above. Porcine IgG was heat-aggregated byincubation at 65 8C for 10 min. After the incubation periodwith either F4 or F4-IC, the cells were washed twice withice-cold PBS and propidium iodide (PI; 5 mg/ml) was addedto exclude dead cells from the analysis. Data were acquiredon a FACSCanto flow cytometer (FCM) with a minimumevent count of 30,000 and analysed with FACSDiva1
software (Becton Dickinson, Erebodegem, Belgium).
2.6. Phenotyping of MoDC
The cell surface expression of FcgR by immature MoDCwas assessed by FCM using FITC-conjugated mAbs againstporcine FcgRIII (clone G7; CD16; AbD Serotec, Oxford, UK)and human FcgRII (clone AT10; CD32; AbD Serotec), whichcrossreacts to porcine FcgRII (Balmelli et al., 2005).Immature MoDC were harvested, washed and incubatedwith pre-titrated saturating concentrations of the mAbs for30 min at 4 8C. Cells stained with FITC-conjugated isotype-matched irrelevant mAbs were used as negative controls.Next, the cells were washed and PI was added to the cells to
mbers.
Ta (8C) Amplicon size (bp)
GTC 53 210
ACC
TTC 53 244
GTC
TTC 53 283
CTC
ACA 55 319
ATG
AAGT 60 230
CTGG
genase.

B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198 191
exclude dead cells from the FCM analysis. MoDC cellsurface expression of MHCII, CD40 and CD80/86 afterstimulation was assessed by FCM using the mAb MSA3(Lunney et al., 1994), mAb G28-5 (anti-human, Bimczoket al., 2007) and a human CTLA4-muIg fusion protein(Ancell, Bayport, MN, USA) respectively, followed by FITC-conjugated F(ab’)2 sheep anti-mouse IgG Ab (Sigma).Briefly, MoDC were harvested and washed in stainingmedium (RPMI 1640 + 1% FCS), whereafter the cells wereincubated with pre-titrated saturating concentrations ofthe primary Abs for 20 min at 4 8C. Cells stained withisotype-matched irrelevant mAbs were used as negativecontrols. After washing, the cells were stained for 20 min at4 8C with FITC-conjugated secondary Abs in stainingmedium. Next, the cells were washed, PI was added anddata were acquired as described above.
2.7. Endocytic activity
The endocytic activity of Ag-stimulated MoDC wasevaluated by incubation of the cells with 10 mg/ml DQ-ovalbumin (DQ-OVA; Invitrogen Molecular Probes) inRPMI 1640 + 1% FCS for 1 h at 37 8C. DQ-OVA acquiresfluorescence after dequenching through proteolyticenzyme cleavage, permitting both the analyses of antigenuptake and processing by FCM. As a control, the uptake at4 8C was analysed. Alternatively, the receptor-mediatedendocytic activity was determined by uptake of FITC-conjugated dextran (Sigma). Briefly, MoDC stimulated inthe presence or absence of Ag were harvested, washed andincubated with 1 mg/ml FITC-conjugated dextran for 1 h at37 8C to measure uptake or at 4 8C to measure binding. PI�
MoDC were analysed for uptake of DQ-OVA or FITC-dextran by FCM with a minimum event count of 30 000.
2.8. Mixed lymphocyte reaction
The T cell stimulatory capacity of MoDC was analysed inan allogenic MLR. Lymphocytes were isolated from PBMCby depletion of CD172a+ cells using the MACS system.MoDC were treated with different antigens or leftuntreated for 24 h, whereafter the cells were harvested,washed and counted. MoDC were then cocultured intriplicate at titrated numbers with 2� 105 CD172a� cells inround-bottomed 96-well plates (Nunc). Cell cultures weremaintained in DMEM, 10% FCS, penicillin/streptomycinand 2-mercapto-ethanol (50 mM) at 37 8C in a humidifiedatmosphere at 5% CO2. After 4-5 days of culture, the cellswere pulse-labelled with 1 mCi/well [3H] methyl-thymi-dine (Amersham ICN, Bucks, UK) for another 18 h. Cellswere harvested onto glass fibre filters (Perkin-Elmer, LifeScience, Brussels, Belgium) and the [3H] methyl-thymidineincorporation was measured using a b-scintillationcounter (Perkin-Elmer).
The Ag-specific proliferation was assessed by usingCD172a� lymphocytes isolated from pigs immunized i.m.with 100 mg purified F4 fimbriae in incomplete Freund’sadjuvant (Van der Stede et al., 2003) and i.v. with 100 mg F4in PBS. MoDC were stimulated with either F4 fimbriae orF4-IC for 24 h or left untreated as mentioned above.Stimulated MoDC were subsequently cocultured at titrated
numbers with 2� 105 autologous lymphocytes for 4–5days and the [3H]methyl-thymidine incorporation wasmeasured as described above.
2.9. Determination of cytokine expression levels by ELISA
MoDC were generated and stimulated with thedifferent Ag, except for CT, as mentioned above. After24 h the cell-free culture supernatants were collected andthe porcine IL-1b, IL-6, IL-8, IL-10, IL-12p40 and TNFacytokine concentrations were determined with commer-cially available ELISA kits (Duoset, R&D systems) accordingto the manufacturer’s instructions. Briefly, microtiterplates were coated overnight at room temperature withcapture Ab specific for the analysed cytokines. The plateswere washed, blocked and samples and standards wereanalysed in duplicate with the detection Ab and thestreptavidin-HRP system. Optical densities were measuredin an ELISA plate reader at 450 nm. The cytokineconcentrations were calculated using DeltaSOFT JV 2.1.2software (BioMetallics, Princeton, NJ, USA) with a 5-parameter curve-fitting algorithm applied for standardcurve calculations.
2.10. Statistical analysis
Statistical analysis were performed using SPSS 16. One-way ANOVA with least significant difference (LSD) post hoctest was performed to compare the increase in theexpression of the cell surface molecules, the internaliza-tion of F4-IC and the proliferation of autologous lympho-cytes between the different treatments after square roottransformation of the data to homogenize variances. TheMann–Whitney U test was used to analyse the cytokineconcentrations between the different treatments. A p-value< 0.05 was considered statistically significant.
3. Results
3.1. Internalization of F4-IC is mediated through Fcgreceptors expressed by MoDC
We aimed to determine whether immature MoDC areable to internalise F4 fimbriae. Therefore, F4 fimbriae werefluorescently labelled with FluoS and immature MoDCwere incubated with different concentrations of F4-FluoS.Immature MoDC internalize F4 fimbriae both in a dose-dependent (Fig. 1A) and a time-dependent manner(Fig. 1B; uptake of 1 mg F4-FluoS). To observe a cleardifference in the uptake between F4 fimbriae and F4fimbriae incorporated in IC, we chose 1 mg F4-FluoS asworking concentration to generate fluorescent F4-IC.Although it has previously been demonstrated that porcineMoDC express functional FcgRIII (Carrasco et al., 2001) andare capable of IC uptake (Paillot et al., 2001), FcgRI and IIexpression has not been demonstrated. We detected FcgRI,FcgRII and FcgRIII mRNA expression by porcine immatureMoDC (Fig. 1C). Furthermore, a high cell surface expressionof FcgRIII by immature MoDC was detected, while FcgRII isexpressed at considerably lower levels (Fig. 1D). Asexpected, when the anti-F4 IgG concentration increases
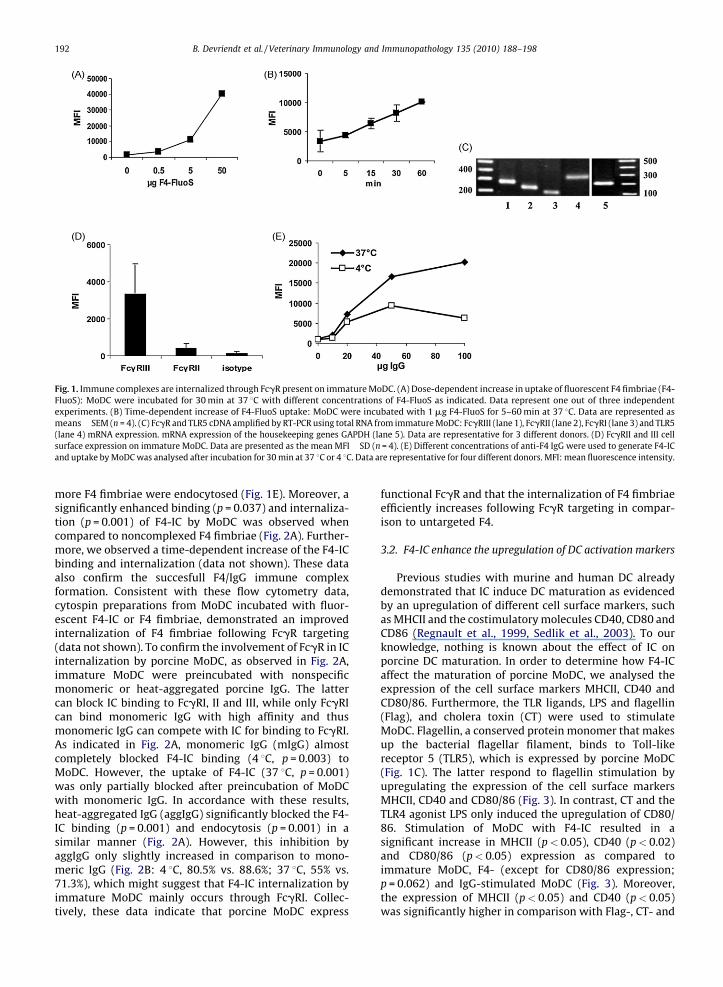
Fig. 1. Immune complexes are internalized through FcgR present on immature MoDC. (A) Dose-dependent increase in uptake of fluorescent F4 fimbriae (F4-
FluoS): MoDC were incubated for 30 min at 37 8C with different concentrations of F4-FluoS as indicated. Data represent one out of three independent
experiments. (B) Time-dependent increase of F4-FluoS uptake: MoDC were incubated with 1 mg F4-FluoS for 5–60 min at 37 8C. Data are represented as
means� SEM (n = 4). (C) FcgR and TLR5 cDNA amplified by RT-PCR using total RNA from immature MoDC: FcgRIII (lane 1), FcgRII (lane 2), FcgRI (lane 3) and TLR5
(lane 4) mRNA expression. mRNA expression of the housekeeping genes GAPDH (lane 5). Data are representative for 3 different donors. (D) FcgRII and III cell
surface expression on immature MoDC. Data are presented as the mean MFI� SD (n = 4). (E) Different concentrations of anti-F4 IgG were used to generate F4-IC
and uptake by MoDC was analysed after incubation for 30 min at 37 8C or 4 8C. Data are representative for four different donors. MFI: mean fluorescence intensity.
B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198192
more F4 fimbriae were endocytosed (Fig. 1E). Moreover, asignificantly enhanced binding (p = 0.037) and internaliza-tion (p = 0.001) of F4-IC by MoDC was observed whencompared to noncomplexed F4 fimbriae (Fig. 2A). Further-more, we observed a time-dependent increase of the F4-ICbinding and internalization (data not shown). These dataalso confirm the succesfull F4/IgG immune complexformation. Consistent with these flow cytometry data,cytospin preparations from MoDC incubated with fluor-escent F4-IC or F4 fimbriae, demonstrated an improvedinternalization of F4 fimbriae following FcgR targeting(data not shown). To confirm the involvement of FcgR in ICinternalization by porcine MoDC, as observed in Fig. 2A,immature MoDC were preincubated with nonspecificmonomeric or heat-aggregated porcine IgG. The lattercan block IC binding to FcgRI, II and III, while only FcgRIcan bind monomeric IgG with high affinity and thusmonomeric IgG can compete with IC for binding to FcgRI.As indicated in Fig. 2A, monomeric IgG (mIgG) almostcompletely blocked F4-IC binding (4 8C, p = 0.003) toMoDC. However, the uptake of F4-IC (37 8C, p = 0.001)was only partially blocked after preincubation of MoDCwith monomeric IgG. In accordance with these results,heat-aggregated IgG (aggIgG) significantly blocked the F4-IC binding (p = 0.001) and endocytosis (p = 0.001) in asimilar manner (Fig. 2A). However, this inhibition byaggIgG only slightly increased in comparison to mono-meric IgG (Fig. 2B: 4 8C, 80.5% vs. 88.6%; 37 8C, 55% vs.71.3%), which might suggest that F4-IC internalization byimmature MoDC mainly occurs through FcgRI. Collec-tively, these data indicate that porcine MoDC express
functional FcgR and that the internalization of F4 fimbriaeefficiently increases following FcgR targeting in compar-ison to untargeted F4.
3.2. F4-IC enhance the upregulation of DC activation markers
Previous studies with murine and human DC alreadydemonstrated that IC induce DC maturation as evidencedby an upregulation of different cell surface markers, suchas MHCII and the costimulatory molecules CD40, CD80 andCD86 (Regnault et al., 1999, Sedlik et al., 2003). To ourknowledge, nothing is known about the effect of IC onporcine DC maturation. In order to determine how F4-ICaffect the maturation of porcine MoDC, we analysed theexpression of the cell surface markers MHCII, CD40 andCD80/86. Furthermore, the TLR ligands, LPS and flagellin(Flag), and cholera toxin (CT) were used to stimulateMoDC. Flagellin, a conserved protein monomer that makesup the bacterial flagellar filament, binds to Toll-likereceptor 5 (TLR5), which is expressed by porcine MoDC(Fig. 1C). The latter respond to flagellin stimulation byupregulating the expression of the cell surface markersMHCII, CD40 and CD80/86 (Fig. 3). In contrast, CT and theTLR4 agonist LPS only induced the upregulation of CD80/86. Stimulation of MoDC with F4-IC resulted in asignificant increase in MHCII (p< 0.05), CD40 (p< 0.02)and CD80/86 (p< 0.05) expression as compared toimmature MoDC, F4- (except for CD80/86 expression;p = 0.062) and IgG-stimulated MoDC (Fig. 3). Moreover,the expression of MHCII (p< 0.05) and CD40 (p< 0.05)was significantly higher in comparison with Flag-, CT- and

Fig. 2. A) MoDC were incubated with F4 fimbriae (1 mg F4-FluoS) and
immune complexes (1 mg F4-FluoS + 46 mg anti-F4 IgG) for 30 min at
either 4 8C (binding) or 37 8C (internalization). For the FcgR inhibition
studies, MoDC were preincubated with either nonspecific monomeric
(mIgG) or heat-aggregated (aggIgG) porcine IgG for 30 min at 4 8C or
37 8C, after which MoDC were incubated for an additional 30 min with F4-
IC. Data are represented as means� SEM (n = 4–6). Letters indicate a
significant difference; (a) p< 0.05, (b)–(d) p< 0.01. (B) Inhibition of IC
binding (4 8C) and internalization (37 8C) after preincubation of MoDC with
either mIgG or aggIgG. Data are represented as means� SEM (n = 4–6).
Fig. 3. Phenotypic analysis of Ag-treated MoDC. Immature MoDC were
stimulated with different antigens as indicated in the x-axis and the
expression of the cell surface markers MHCII, CD80/86 and CD40 was
assayed by FCM. These data are represented as the mean relative increase
of marker expression (%� SEM; n = 16 individual monocyte donors) after
treatment as compared to immature MoDC. The relative marker expression
in % = 100� [(MFItreatment�MFIimm)/MFIimm]. F4-IC stimulated MoDC
significantly increase MHCII expression in comparison to immature DC
and all other Ags (*p< 0.05), whereas for CD80/86 expression F4-IC
significantly differs from immature DC and IgG (*p< 0.05). CD40
expression significantly increased after F4-IC stimulation in comparison
to all other stimuli (*p< 0.02).
B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198 193
LPS-stimulated MoDC (Fig. 3). These results indicate thatFcgR engagement significantly enhances the upregulationof MHCII, CD40 and CD80/86 expression on MoDC incomparison with immature, IgG- and F4-treated MoDC.
3.3. F4-IC stimulation reduces the phagocytic and endocytic
capacity
We next sought to investigate if the observed pheno-typical maturation of the treated MoDC correlated with afunctional maturation. Immature DC are proficient inphagocytosis, while mature DC downregulate their fluidphase uptake in preparation for efficient Ag presentation toT cells (Adams et al., 2004). Therefore, the phagocytic andantigen processing capacity of Ag-stimulated MoDC wasassessed through the uptake of DQ-OVA. As shown inFig. 4A, F4-IC stimulated MoDC display a reduced uptake ofDQ-OVA as compared to immature, F4- and IgG-treatedMoDC. Intriguingly, LPS and CT treatment did not result ina reduced DQ-OVA uptake. On the contrary, after CTtreatment we observed a higher uptake as compared toimmature MoDC, while LPS had almost no influence onDQ-OVA uptake. On the other hand, flagellin reduced theuptake of DQ-OVA. To confirm this reduction, the receptor-mediated endocytic activity of Ag-treated MoDC was
measured by administration of fluorescently labeleddextran (Fig. 4B), which binds to the mannose receptor.As expected, F4-IC treatment resulted in a reductionof FITC-dextran uptake in comparison with immature,F4- and IgG-treated cells. However, both CT and LPSinduced a reduction in FITC-dextran uptake, contradictingthe DQ-OVA data. Flagellin stimulation also reduced theinternalization of FITC-dextran by MoDC. Although notsignificant, these findings indicate that F4-IC stimulation ofMoDC downregulates the phagocytic and the receptor-mediated endocytic capacity in a more profound mannerthan either F4 or IgG stimulation.

Fig. 4. Downregulation of the endocytic and antigen processing capacity
of activated porcine MoDC. Both DQ-OVA (A) and FITC-dextran (B) uptake
is reduced in Ag-stimulated MoDC. MFI values were calculated by
subtracting MFI values obtained at 4 8C from those at 37 8C. These results
are representative for one out of 4 independent experiments.
Fig. 5. FcgR activation in F4-IC primed MoDC enhances the T cell
proliferation. (A) Allogenic MLR. Porcine MoDC were left untreated or
stimulated for 24 h with different agents as mentioned in the x-axis. The T
cell stimulatory capacity of stimulated MoDC was assessed by coculturing
2� 105 CD172a-depleted PBMC (T cell source) with graded numbers of
activated MoDC for 4 days. The proliferative responses were determined
by the incorporation of [3H]methyl-thymidine. Error bars indicate the
observed range within triplicates in one representative experiment out of
four using different donors. Data are represented as the mean cpm� SD.
(B) Antigen-specific autologous lymphocyte proliferation. After an
incubation period of 24 h, untreated or F4- and IC-stimulated MoDC were
cocultured with autologous lymphocytes isolated from F4 immunized pigs.
Proliferative responses were determined after 5 days by [3H]methyl-
thymidine incorporation. Data are shown as mean cpm� SEM (n = 3). The
proliferative responses of the CD172a� lymphocytes (no MoDC were added)
was below 3000 cpm. The letters indicate a significant difference between
the different stimulations: (a) IC to imm, p< 0.01; (b) F4 to imm, p< 0.02; (c)
IC to F4, p< 0.05.
B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198194
3.4. F4-IC activated MoDC induce a strong lymphocyte
proliferation
Another hallmark of DC maturation is their ability toactivate naive T cells due to the increased expression ofMHCII and costimulatory molecules. To confirm whetherthe observed maturation of MoDC after Ag treatmentcorrelates with the ability of mature DC to stimulateallogenic T cells, Ag-stimulated MoDC were coculturedwith allogenic T cells in a mixed lymphocyte reaction(MLR). F4-IC-treated MoDC were clearly more effective instimulating T cell proliferation in comparison withimmature, F4- and IgG-treated MoDC, especially at thelower DC to T cells ratio’s (Fig. 5A). Flagellin also induced ahigh T cell proliferation, whereas CT and LPS induced anintermediate T cell proliferation, as is the case for F4 andIgG. These MLR results suggest that IC and Flag stimulationinduced MoDC that were more potent activators ofallogenic T cells as compared to immature MoDC,indicating a functional MoDC maturation. Furthermore,IC stimulation enhances the functional DC maturation incomparison with F4 or IgG stimulation.
Ag-specific proliferation assays with F4-IC-pulsedMoDC and lymphocytes obtained from F4 immunized pigsrevealed a similar improvement of the stimulatory
capacity of MoDC after FcgR targeting (Fig. 5B). Whileboth F4- and F4-IC-stimulated MoDC induced a signifi-cantly enhanced Ag-specific T cell proliferation in compar-ison with immature DC (F4: p< 0.02; F4-IC: p< 0.01), F4-IC significantly heightened this proliferation as comparedto F4 stimulation (p< 0.05) (Fig. 5B).
3.5. F4-IC matured MoDC secrete predominantly pro-
inflammatory cytokines
Mature DC influence the outcome of immune responsesthrough the differentiation and polarization of T cells andthis is mediated by a combination of signals derived fromDC, including the secretion of T cell polarizing cytokines. Toinvestigate whether the observed phenotypical and func-tional maturation of MoDC induced after Ag treatmentcorrelates with a certain cytokine expression profile, wedecided to investigate the cytokine expression profile after

B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198 195
Ag induced MoDC maturation. The IL-1b, IL-6, IL-8, IL-10,IL12p40 and TNFa concentrations were determined in theculture supernatans of immature MoDC or MoDC that hadpreviously been stimulated with LPS, flag, F4, IgG or F4-IC.As shown in Fig. 6, both flagellin and F4-IC stimulationresulted in an significantly increased production of IL-1b(Flag: p = 0.001; IC: p = 0.001), IL-6 (Flag: p = 0.012; IC:p = 0.012), IL-8 (Flag: p = 0.001; IC: p = 0.001), IL-12p40(Flag: p = 0.027) and TNFa (Flag: p = 0.027; IC: p = 0.004) ascompared to immature MoDC. Furthermore, F4-IC stimu-lation significantly enhanced the production of thesecytokines in comparison with IgG (IL-1b: p = 0.001, IL-8:p = 0.002, IL-6: p= 0.027, TNFa: p = 0.015) and F4 treatment(IL-1b: p = 0.012, IL-8: p = 0.002, TNFa: p = 0.02). Althoughnot significant F4-IC stimulation seemed to increase IL-12p40 secretion in comparison with immature, F4- andIgG-stimulated MoDC and all treatments seemed to induceIL-10 production in comparison with immature MoDC.Interestingly, immature MoDC spontaneously secreted IL-10. LPS and F4 stimulation induced a significant increase inthe expression of IL-8 as compared to immature MoDC(p = 0.001). Furthermore, there was a tendency of increased
Fig. 6. Cytokine expression pattern of mature MoDC. Immature MoDC were st
collected and the concentration of IL-1b, IL-6, IL-8, IL-10, IL-12p40 and TNFa c
(n = 8 individual blood donors). Letters indicate a significant difference (p< 0.05) be
stimulation, while the triangle (D) indicates a significant difference (p< 0.05) bet
IL-6 and IL-10 expression by MoDC after F4 and LPSstimulation (Fig. 6). On the other hand, IgG stimulation didnot seem to affect cytokine synthesis. These results clearlydemonstrate that F4 targeting to FcgR on MoDC signifi-cantly upregulated the production of several cytokines incomparison with immature, F4- and IgG-stimulated MoDC.
4. Discussion
In the current study we demonstrated that porcine MoDCinternalize F4 fimbriae. Presumably this internalization ismediated through the recently discovered F4 receptor,porcine aminopeptidase N (Rasschaert et al., manuscript inpreparation), which is expressed on porcine MoDC (data notshown). However, targeting F4 fimbriae to FcgR signifi-cantly enhanced the endocytosis of F4 fimbriae by porcineMoDC, which implicates an FcgR-mediated process andcorroborates previous reports demonstrating the enhancedinternalization of antigens incorporated in immune com-plexes (Liu et al., 2006; Rawool et al., 2008). Although afunctional FcgRIII (CD16) expression and the uptake of IC byporcine MoDC was previously demonstrated (Carrasco et al.,
imulated with the indicated agents for 24 h. Cell-free supernatans were
ytokines were quantified by ELISA. Data are represented as means� SEM
tween F4-IC stimulation and immature MoDC (a), LPS (b), F4 (c) and IgG (d)
ween immature MoDC and Ag-stimulated MoDC.

B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198196
2001; Paillot et al., 2001), the effect of FcgR engagement byIC on the maturation of porcine MoDC had not beenevaluated. We show for the first time that immature porcineMoDC express FcgRI, II and III mRNA. Consistent with themRNA expression, porcine MoDC express FcgRII and FcgRIIIon the cell surface, although the former was detected atmuch lower expression levels than FcgRIII. The cell surfaceexpression of FcgRI still needs to be established, howevercurrently no porcine-specific or crossreactive FcgRI mAbsare available. These expression data seem to contradict FcgRexpression in human MoDC, which mainly express FcgRII atthe cell surface (Boruchov et al., 2005). Porcine MoDCendocytose F4-IC and both monomeric and multimeric IgGsignificantly inhibited this endocytosis, suggesting func-tional FcgR expression and an FcgR-mediated F4-ICinternalization. In comparison to monomeric IgG, multi-meric IgG did not significantly increase the inhibition andsince only FcgRI binds monomeric IgG, this suggests that F4-IC internalization is predominantly mediated via FcgRI,which is consistent with murine data (Rawool et al., 2008).Despite the fact that multimeric IgG binds to FcgRI, II and III,the F4-IC internalization was not completely inhibited,indicating that F4-IC are partially internalized via an FcgR-independent pathway.
Although previous studies with both human andmurine DC have shown that Ag targeting to FcgR enhancesDC maturation (Regnault et al., 1999; Sedlik et al., 2003;Liu et al., 2006), the effect of IC on porcine DC maturationwas not yet determined. Consistent with these earlierfindings, FcgR ligation by F4-IC resulted in a significantupregulation of the DC maturation markers MHCII, CD80/86 and CD40. Moreover, this phenotypic DC maturationwas accompanied by a reduced receptor-mediated endo-cytosis and macropinocytosis, corresponding to a shiftfrom antigen processing cells to antigen presenting cells.Furthermore, F4-IC matured MoDC showed an improvedallogenic T cell stimulatory capacity, providing additionalevidence that FcgR engagement by F4-IC induces afunctional DC maturation. In an autologous MLR, theinteraction of F4-IC with FcgR significantly augmented theefficiency of Ag-presentation to T lymphocytes in compar-ison with F4 primed DC. A recent study elegantly showedthat the enhanced Ag presentation by DC followingstimulation with multimeric IC is dependent on FcRn(Qiao et al., 2008). Given the polymeric nature of F4fimbriae, and consequently the generation of multimericIC, and the expression of FcRn by porcine MoDC (ourunpublished observations), we assume that a similarmechanism could be in play in the observed enhancementof Ag presentation by F4-IC matured MoDC. In humans,FcgR-mediated induction of DC maturation requires theblockade of the inhibitory FcgRIIB receptor (Dhodapkaret al., 2005). Despite the fact that porcine DC presumablyexpress the inhibitory FcgRIIB, DC maturation was stillpossible, indicating that coligation of activating andinhibitory FcgR not necessarily blocks porcine DC matura-tion.
DC, with their ability to stimulate naıve and antigen-specific T cells, can also potently influence innate andhumoral responses (Adams et al., 2004). Our datademonstrate that the F4-IC-induced DC maturation
correlated with a significantly increased secretion ofseveral pro-inflammatory cytokines, such as IL-1b, IL-6and TNFa, and the chemokine IL-8. IL-6 is known tostimulate B cells, while TNFa and IL-8 serve as chemoat-tractants for innate and adaptive immune effector cells,such as neutrophils and T cells (Gesser et al., 1996),underlining the function of DC in linking innate andadaptive immunity. Interestingly, IC stimulation failed toinduce a significant increase in IL-12p40 secretion.Anderson et al. (2004) demonstrated that FcgR ligationin human DC did not affect the expression of IL-12p40.Nevertheless, F4-IC matured MoDC were able to induce amore potent allogenic T cell proliferation compared toimmature and F4-stimulated DC. The cytokine expressiondata imply that FcgR ligation enables DC to recruit adiversified immune response against antibody-coatedpathogens. It’s also worth mentioning that in contrast toPilon et al. (2009) in our model immature MoDCspontaneously secrete IL-10, like immature human DC(Mnasria et al., 2008).
Cholera toxin has been implicated as a mucosaladjuvans for oral vaccination in piglets, demonstratingamplified immune responses to co-administered antigens(Verdonck et al., 2005). It was suggested that theinteraction of CT with DC is crucial for this activity(Gagliardi et al., 2002). However, Bimczok et al. (2007)reported that CT stimulation of porcine MoDC resulted inthe induction of a semi-mature phenotype. We alsoobserved only an increase in CD80/86 expression afterCT stimulation and no alteration in the endocytosis of OVA-dQ. In contrast, FITC-dextran uptake was reduced. Thisinconsistency could probably be due to the differentendocytosis mechanisms of OVA-dQ and FITC-dextran(Sallusto et al., 1995). Nevertheless, these findings and theMLR data further support the induction of a semi-matureporcine DC phenotype after CT treatment.
The TLR5 agonist flagellin seems to be a potent DCmaturation stimulus in humans, but in mice (Means et al.,2003; Dowling et al., 2008). We report for the first timethat porcine MoDC express TLR5 mRNA and mature inresponse to stimulation with ETEC-derived flagellin asevidenced by an increased expression of both MHCII andcostimulatory molecules, a reduced endocytotic capacityand an improved T cell stimulatory capacity. Flagellinstimulation also induced the expression of IL-1b, TNFa, IL-6, IL-8 and IL-12p40, implicating that flagelline stimulatedDC could direct T cells toward a TH1 response. In contrast toflagellin, porcine MoDC seemed rather unresponsive toLPS, since the latter failed to induce DC maturation and IL-12p40 cytokine secretion. These results are in line withearlier reports regarding LPS stimulation of porcine MoDC(Guzylack-Piriou et al., 2006; Pilon et al., 2009).
Oral F4 immunization of recently weaned pigletsprotects them against a subsequent F4+ ETEC challenge,which indicates the potent immunogenicity of F4 fimbriae(Van den Broeck et al., 1999a, 1999b; Verdonck et al.,2004). To prevent postweaning diarrhea, an active mucosalimmune response is required at the time of weaning.Therefore, piglets have to be immunized during thesuckling period, however attempts to immunize sucklingpiglets are hampered due to their immature immune

B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198 197
system (Snoeck et al., 2003). Several studies in mice andman have already demonstrated that Ag targeting to FcgRenhances both the humoral and cellular immunity(Getahun et al., 2004; Hjelm et al., 2006; Rawool et al.,2008) and abolishes the need for adjuvants (Adamovaet al., 2005; Getahun and Heyman, 2006). Moreover,previous studies have demonstrated that IC are transcy-tosed across the epithelial barrier via the neonatal Fcreceptor, FcRn (Yoshida et al., 2004), which is alsoexpressed by porcine enterocytes (Stirling et al., 2005).In addition, porcine small intestinal DC express FcgRIII(CD16) (Haverson et al., 2000; Bimczok et al., 2006),implying that FcgR-targeted Ag delivery to porcineintestinal DC could be used in oral vaccination. However,in vivo studies are required to address if F4-IC transcytosisoccurs in porcine enterocytes and if intestinal DC respondto FcgR targeting in a similar manner.
5. Conclusions
In conclusion, FcgR targeting to porcine MoDC sig-nificantly enhances the Ag internalization, DC maturationand Ag presentation to T cells, which corroborates previousexperiments with both human and murine DC. Moreover,the results further deepen our understanding of porcine DCbiology and suggest that F4-IC could offer a promisingcandidate for oral vaccine formulations for the mucosalimmunization of suckling piglets.
Conflict of interest
None.
Acknowledgements
We gratefully thank R. Cooman and G. De Smet for theirexcellent technical support and assistance with theanimals. We acknowledge Prof. Dr. S. Inumaru (Instituteof Animal Health, Ibaraki, Japan) for kindly providingrpGM-CSF, Prof. Dr. H.J. Rothkotter (Institute of Anatomy,Magdeburg, Germany) for the a-CD40 hybridoma SN andProf. Dr. A. Saalmuller (University of Veterinary Medicine,Vienna, Austria) for the MSA3 mAb. This research wasfunded by a PhD grant of the Institute for the Promotion ofInnovation through Science and Technology in Flanders(IWT-Vlaanderen). F. Verdonck had a postdoctoral grantfrom the FWO Vlaanderen.
References
Adams, S., O’Neill, D.W., Bhardwaj, N., 2004. Recent Advances in DendriticCell Biology. J. Clin. Immunol. 25 (3), 177–188.
Adamova, E., Walsh, M.C., Gosselin, D.R., Hale, K., Preissler, M.T., Graziano,R.F., Gosselin, E.J., 2005. Enhanced antigen-specific antibody andcytokine responses when targeting antigen to human FcGAMMAreceptor type I using an anti-human FcGAMMA receptor type I-streptavidin fusion protein in an adjuvant-free system. Immunol.Invest. 34 (4), 417–429.
Alexander, T.J.L., 1994. Neonatal diarrhoea in pigs. In: Gyles, C.L. (Ed.),Escherichia coli in Domestic Animals and Humans. CAB International,Oxon, UK, pp. 151–170.
Anderson, C.F., Lucas, M., Gutierrez-Kobeh, L., Field, A.E., Mosser, D.M.,2004. T cell biasing by activated dendritic cells. J. Immunol. 173 (2),955–961.
Bakker, D., Willemsen, P.T., Simons, L.H., van Zijderveld, F.G., de Graaf, F.K.,1992a. Characterization of the antigenic and adhesive properties ofFaeG, the major subunit of K88 fimbriae. Mol Microbiol. 6, 247–255.
Bakker, D., Willemsen, P.T., Willems, R.H., Huisman, T.T., Mooi, F.R.,Oudega, B., Stegehuis, F., de Graaf, F.K., 1992b. Identification of minorfimbrial subunits involved in biosynthesis of K88 fimbriae. J. Bacter-iol. 174, 6350–6358.
Balmelli, C., Vincent, I.E., Rau, H., Guzylack-Piriou, L., McCullough, K.,Summerfield, A., 2005. Fc gamma RII-dependent sensitisation ofnatural interferon-producing cells for viral infection and inter-feron-alpha responses. Eur. J. Immunol. 35 (8), 2406–2415.
Banchereau, J., Briere, F., Caux, C., Davoust, J., Lebecque, S., Liu, Y.J.,Pulendran, B., Palucka, K., 2000. Immunobiology of dendritic cells.Annu. Rev. Immunol. 18, 767–811.
Bimczok, D., Post, A., Tschernig, T., Rothkotter, H.J., 2006. Phenotype anddistribution of dendritic cells in the porcine small intestinal andtracheal mucosa and their spatial relationship to epithelial cells. CellTissue Res. 325 (3), 461–468.
Bimczok, D., Rau, H., Wundrack, N., Naumann, M., Rothkotter, H.J., McCul-lough, K., Summerfield, A., 2007. Cholera toxin promotes the genera-tion of semi-mature porcine monocyte-derived dendritic cells thatare unable to stimulate T cells. Vet. Res. 38 (4), 597–612.
Boruchov, A.M., Heller, G., Veri, M.C., Bonvini, E., Ravetch, J.V., Young, J.W.,2005. Activating and inhibitory IgG Fc receptors on human DCsmediate opposing functions. J. Clin. Invest. 115 (10), 2914–2923.
Carrasco, C.P., Rigden, R.C., Schaffner, R., Gerber, H., Neuhaus, V., Inumaru,S., Takamatsu, H., Bertoni, G., McCullough, K.C., Summerfield, A., 2001.Porcine dendritic cells generated in vitro: morphological, phenotypicand functional properties. Immunology 104 (2), 175–184.
Cox, E., Van der Stede, Y., Verdonck, F., Snoeck, V., Van den Broeck, W.,Goddeeris, B., 2002. Oral immunisation of pigs with fimbrial antigensof enterotoxigenic E. coli: an interesting model to study mucosalimmune mechanisms. Vet. Immunol. Immunopathol. 87, 287–290.
Devriendt, B., Gallois, M., Verdonck, F., Wache, Y., Bimczok, D., Oswald, I.P.,Goddeeris, B.M., Cox, E., 2009. The food contaminant fumonisin B1reduces the maturation of porcine CD11R1+ intestinal antigen pre-senting cells and antigen-specific immune responses, leading to aprolonged intestinal ETEC infection. Vet. Res. 40 (4), 40, doi:10.1051/vetres/2009023.
Dhodapkar, K.M., Kaufman, J.L., Ehlers, M., Banerjee, D.K., Bonvini, E.,Koenig, S., Steinman, R.M., Ravetch, J.V., Dhodapkar, M.V., 2005.Selective blockade of inhibitory Fcgamma receptor enables humandendritic cell maturation with IL-12p70 production and immunity toantibody-coated tumor cells. Proc. Natl. Acad. Sci. U.S.A. 102 (8),2910–2915.
Dowling, D., Hamilton, C.M., O’Neill, S.M., 2008. A comparative analysis ofcytokine responses, cell surface marker expression and MAPKs in DCsmatured with LPS compared with a panel of TLR ligands. Cytokine 41(3), 254–262.
Gagliardi, M.C., Sallusto, F., Marinaro, M., Vendetti, S., Riccomi, A., DeMagistris, M.T., 2002. Effects of the adjuvant cholera toxin on den-dritic cells: stimulatory and inhibitory signals that result in theamplification of immune responses. Int. J. Med. Microbiol. 291 (6–7), 571–575.
Gesser, B., Lund, M., Lohse, N., Vestergaard, C., Matsushima, K., SindetPe-dersen, S., Jensen, S.L., Thestrup-Pedersen, K., Larsen, C.G., 1996. IL-8induces T cell chemotaxis, suppresses IL-4, and up-regulates IL-8production by CD4(+) T cells. J. Leukoc. Biol. 59 (3), 407–411.
Getahun, A., Dahlstrom, J., Wernersson, S., Heyman, B., 2004. IgG2a-mediated enhancement of antibody and T cell responses and itsrelation to inhibitory and activating Fc gamma receptors. J. Immunol.172 (9), 5269–5276.
Getahun, A., Heyman, B., 2006. How antibodies act as natural adjuvants.Immunol. Lett. 104 (1–2), 38–45.
Guermonprez, P., Valladeau, J., Zitvogel, L., Thery, C., Amigorena, S., 2002.Antigen presentation and T cell stimulation by dendritic cells. Annu.Rev. Immunol. 20, 621–667.
Guzylack-Piriou, L., Piersma, S., McCullough, K., Summerfield, A., 2006.Role of natural interferon-producing cells and T lymphocytes inporcine monocyte-derived dendritic cell maturation. Immunology118 (1), 78–87.
Halloran, P.J., Sweeney, S.E., Strohmeier, C.M., Kim, Y.B., 1994. Molecularcloning and identification of the porcine cytolytic trigger molecule G7as a Fc gamma RIII alpha (CD16) homologue. J. Immunol. 153 (6),2631–2641.
Hampson, D.J., 1994. Postweaning Escherichia coli diarrhoea in pigs. In:Gyles, C.L. (Ed.), Escherichia coli in Domestic Animals and Humans.CAB International, Oxon, UK, pp. 171–191.

B. Devriendt et al. / Veterinary Immunology and Immunopathology 135 (2010) 188–198198
Haverson, K., Singha, S., Stokes, C.R., Bailey, M., 2000. Professional andnon-professional antigen-presenting cells in the porcine small intes-tine. Immunology 101, 492–500.
Hjelm, F., Carlsson, F., Getahun, A., Heyman, B., 2006. Antibody-mediatedregulation of the immune response. Scand. J. Immunol. 64 (3), 177–184.
Inumaru, S., Kokuho, T., Denham, S., Denyer, M.S., Momotani, E., Kitamura,S., Corteyn, A., Brookes, S., Parkhouse, R.M., Takamatsu, H., 1998.Expression of biologically active recombinant porcine GM-CSF bybaculovirus gene expression system. Immunol. Cell Biol. 76 (3),195–201.
Johansson, E., Domeika, K., Berg, M., Alm, G.V., Fossum, C., 2003. Char-acterisation of porcine monocyte-derived dendritic cells according totheir cytokine profile. Vet. Immunol. Immunopathol. 91 (3–4), 183–197.
Liu, Y., Gao, X., Masuda, E., Redecha, P.B., Blank, M.C., Pricop, L., 2006.Regulated expression of FcgammaR in human dendritic cells controlscross-presentation of antigen-antibody complexes. J. Immunol. 177(12), 8440–8447.
Lunney, J.K., Walker, K., Goldman, T., Aasted, B., Bianchi, A., Binns, R., et al.,1994. Overview of the first international workshop to define swineleukocyte cluster of differentiation (CD) antigens. Vet. Immunol.Immunopathol. 43, 193–206.
Means, T.K., Hayashi, F., Smith, K.D., Aderem, A., Luster, A.D., 2003. TheToll-like receptor 5 stimulus bacterial flagellin induces maturationand chemokine production in human dendritic cells. J. Immunol. 170(10), 5165–5175.
Melkebeek, V., Verdonck, F., Goddeeris, B.M., Cox, E., 2007. Comparison ofimmune responses in parenteral FaeG DNA primed pigs boostedorally with F4 protein or reimmunized with the DNA vaccine. Vet.Immunol. Immunopathol. 116 (3–4), 199–214.
Mnasria, K., Lagaraine, C., Velge-Roussel, F., Oueslati, R., Lebranchu, Y.,Baron, C., 2008. Anti-CD25 antibodies affect cytokine synthesis pat-tern of human dendritic cells and decrease their ability to primeallogeneic CD4+ T cells. J. Leukoc. Biol. 84 (2), 460–467.
Paillot, R., Laval, F., Audonnet, J.C., Andreoni, C., Juillard, V., 2001. Func-tional and phenotypic characterization of distinct porcine dendriticcells derived from peripheral blood monocytes. Immunology 102 (4),396–404.
Pescovitz, M.D., Lunney, J.K., Sachs, D.H., 1984. Preparation and charac-terization of monoclonal antibodies reactive with porcine PBL. J.Immunol. 133, 368–375.
Pilon, C., Levast, B., Meurens, F., Le Vern, Y., Kerboeuf, D., Salmon, H.,Velge-Roussel, F., Lebranchu, Y., Baron, C., 2009. CD40 engagementstrongly induces CD25 expression on porcine dendritic cells andpolarizes the T cell immune response toward Th1. Mol. Immunol.46 (3), 437–447.
Qiao, S., Zhang, G., Xia, C., Zhang, H., Zhang, Y., Xi, J., Song, H., Li, X., 2006.Cloning and characterization of porcine Fc gamma receptor II (Fcgam-maRII). Vet. Immunol. Immunopathol. 114 (1–2), 178–184.
Qiao, S.W., Kobayashi, K., Johansen, F.E., Sollid, L.M., Andersen, J.T., Mil-ford, E., Roopenian, D.C., Lencer, W.I., Blumberg, R.S., 2008. Depen-dence of antibody-mediated presentation of antigen on FcRn. Proc.Natl. Acad. Sci. U.S.A. 105 (27), 9337–9342.
Ravetch, J.V., Bolland, S., 2001. IgG Fc receptors. Annu. Rev. Immunol. 19,275–290.
Rawool, D.B., Bitsaktsis, C., Li, Y., Gosselin, D.R., Lin, Y., Kurkure, N.V.,Metzger, D.W., Gosselin, E.J., 2008. Utilization of Fc receptors as amucosal vaccine strategy against an intracellular bacterium, Franci-sella tularensis. J. Immunol. 180 (8), 5548–5557.
Regnault, A., Lankar, D., Lacabanne, V., Rodriguez, A., Thery, C., Rescigno, M.,Saito, T., Verbeek, S., Bonnerot, C., Ricciardi-Castagnoli, P., Amigorena, S.,1999. Fcgamma receptor-mediated induction of dendritic cell matura-tion and major histocompatibility complex class I-restricted antigen
presentation after immune complex internalization. J. Exp. Med. 189(2), 371–380.
Sallusto, F., Lanzavecchia, A., 1994. Efficient presentation of soluble antigenby cultured human dendritic cells is maintained by granulocyte/macro-phage colony-stimulating factor plus interleukin 4 and downregulatedby tumor necrosis factor alpha. J. Exp. Med. 179 (4), 1109–1118.
Sallusto, F., Cella, M., Danieli, C., Lanzavecchia, A., 1995. Dendritic cells usemacropinocytosis and the mannose receptor to concentrate macro-molecules in the major histocompatibility complex class II compart-ment: downregulation by cytokines and bacterial products. J. Exp.Med. 182 (2), 389–400.
Sedlik, C., Orbach, D., Veron, P., Schweighoffer, E., Colucci, F., Gamberale,R., Ioan-Facsinay, A., Verbeek, S., Ricciardi-Castagnoli, P., Bonnerot, C.,Tybulewicz, V.L., Di Santo, J., Amigorena, S., 2003. A critical role for Sykprotein tyrosine kinase in Fc receptor-mediated antigen presentationand induction of dendritic cell maturation. J. Immunol. 170 (2), 846–852.
Snoeck, V., Huyghebaert, N., Cox, E., Vermeire, A., Vancaeneghem, S.,Remon, J.P., Goddeeris, B.M., 2003. Enteric-coated pellets of F4 fim-briae for oral vaccination of suckling piglets against enterotoxigenicEscherichia coli infections. Vet. Immunol. Immunopathol. 96 (3–4),219–227.
Stirling, C.M., Charleston, B., Takamatsu, H., Claypool, S., Lencer, W.,Blumberg, R.S., Wileman, T.E., 2005. Characterization of the porcineneonatal Fc receptor-potential use for trans-epithelial protein deliv-ery. Immunology 114 (4), 542–553.
Van den Broeck, W., Cox, E., Goddeeris, B.M., 1999a. Receptor-dependentimmune responses in pigs after oral immunization with F4 fimbriae.Infect. Immun. 67 (2), 520–526.
Van den Broeck, W., Cox, E., Goddeeris, B.M., 1999b. Induction of immuneresponses in pigs following oral administration of purified F4 fim-briae. Vaccine 17 (15–16), 2020–2029.
Van der Stede, Y., Cox, E., Verdonck, F., Vancaeneghem, S., Goddeeris, B.M.,2003. Reduced faecal excretion of F4+—E. coli by the intramuscularimmunisation of suckling piglets by administration of 1a,25-dihy-droxyvitamin D3 or CpG-oligodeoxynucleotides. Vaccine 21, 1023–1032.
Verdonck, F., Cox, E., Van der Stede, Y., Goddeeris, B.M., 2004. Oralimmunization of piglets with recombinant F4 fimbrial adhesin FaeGmonomers induces a mucosal and systemic F4-specific immuneresponse. Vaccine 22 (31–32), 4291–4299.
Verdonck, F., Snoeck, V., Goddeeris, B.M., Cox, E., 2005. Cholera toxinimproves the F4(K88)-specific immune response following oralimmunization of pigs with recombinant FaeG. Vet. Immunol. Immu-nopathol. 103 (1–2), 21–29.
Verdonck, F., Joensuu, J.J., Stuyven, E., De Meyer, J., Muilu, M., Pirhonen, M.,Goddeeris, B.M., Mast, J., Niklander-Teeri, V., Cox, E., 2008. The poly-meric stability of the Escherichia coli F4 (K88) fimbriae enhances itsmucosal immunogenicity following oral immunization. Vaccine 26(45), 5728–5735.
Yada, A., Ebihara, S., Matsumura, K., Endo, S., Maeda, T., Nakamura, A.,Akiyama, K., Aiba, S., Takai, T., 2003. Accelerated antigen presentationand elicitation of humoral response in vivo by FcgammaRIIB- andFcgammaRI/III-mediated immune complex uptake. Cell. Immunol.225 (1), 21–32.
Yoshida, M., Claypool, S.M., Wagner, J.S., Mizoguchi, E., Mizoguchi, A.,Roopenian, D.C., Lencer, W.I., Blumberg, R.S., 2004. Human neonatalFc receptor mediates transport of IgG into luminal secretions fordelivery of antigens to mucosal dendritic cells. Immunity 20 (6),769–783.
Zhang, G., Qiao, S., Li, Q., Wang, X., Duan, Y., Wang, L., Xiao, Z., Xia, C., 2006.Molecular cloning and expression of the porcine high-affinity immu-noglobulin G Fc receptor (FcgammaRI). Immunogenetics 58 (10),845–849.
Related Documents


![Porcine Epidemic Diarrhea [Autosaved]](https://static.cupdf.com/doc/110x72/577c808c1a28abe054a92a69/porcine-epidemic-diarrhea-autosaved.jpg)








