Proc. Nat!. Acad. Sci. USA Vol. 77, No. 6, pp. 3620-43624, June 1980 Immunology Target selectivity of interferon-induced human killer lymphocytes related to their Fc receptor expression (natural killer cells) MARIA G. MASUCCI, GIUSEPPE MASUCCI, EVA KLEIN, AND WOLFGANG BERTHOLD* Department of Tumor Biology, Karolinska Institutet, S-104 01 Stockholm, Sweden Communicated by George Klein, February 25, 1980 ABSTRACT Human blood lymphocytes were fractionated on the basis of surface characteristics such as adherence to nylon wool and expression of erythrocyte (E) and Fc receptors. The various subsets were incubated with interferon for 3 hr. Two cell lines that differ in sensitivity to the natural killer effect, K562 and Daudi, were exposed to these lymphocytes (i.e., their sensitivity to interferon-activated killing was tested). Cell line Daudi, with a low sensitivity to the natural killer effect, was also affected by interferon-activated killing. The efficiency of the nonadherent subsets, separated accor in to the expression of E receptor, ranked similarly in natural killing (anti-K562) and interferon-activated killing (anti-K562 and anti-Daudi) in the following order: E receptor-negative cells, low-affinity E re- ceptor-positive cells, and high-affinity E receptor-positive cells. Further separation on the basis of Fc receptor expression re- vealed a difference between the two targets. The Fc receptor- positive and -negative cells that did not express high-affinity E receptors killed K562 with similar efficiency whereas Daudi cells were more sensitive to the effect of cells devoid of Fc re- ceptor. Results obtained with other targets suggested that T cell lines behave similarly to K562 and that the difference may be generally valid for T and B cell lines. Human blood lymphocytes have a lytic effect in vitro against certain tumor cell lines (1). The active population is heteroge- neous with regard to its cell surface characteristics (2-7). The "null" subset, obtained after removal of surface immuno- globulin-positive cells and readily resetting T lymphocytes, is highly enriched in activity (2-4). This subset contains a high proportion of Fc receptor (FcR)-positive cells. In addition, T lymphocytes with low-affinity sheep erythrocyte receptors (ERs) are also active (5-7). The killing mechanism is unknown. In short-term assays, T cell-derived lymphoid lines are more sensitive than B lines (8). Blood lymphocytes of the majority of donors do not kill Daudi cells, a Burkitt lymphoma-derived B cell line carrying Epstein-Barr virus (9). In long-term tests, this target is also lysed, however. Lysis begins after 3-4 hr, suggesting an activation of the effectors during this period. An apparently nondiscriminative cytotoxic potential is also generated in mixed cultures of lymphocytes and tumor cells. Fibroblasts derived from normal tissues did not generate such cytotoxicity, neither in allogeneic nor in autologous combina- tions (10, 11). In cultures in which immune interactions occur, interferon is produced. This is probably responsible for the triggering because the exogenous interferon treatment activates lymphocytes promptly for cytotoxicity (12-16). The Daudi line is sensitive to interferon-activated killing (IAK), and the kinetics of cytotoxicity in such a system is similar against Daudi and K562 cells (17). Cell line K562 is highly sensitive to natural killing (NK). Because the large difference in the sensitivity of Daudi and K562 to lymphocyte-mediated lysis disappears when inter- feron-triggered lymphocytes are used as effectors, it is likely that interferon activates the population to express its maximal cytotoxic potential. Two sets of experiments have provided evidence that the increased cytotoxicity of IAK as compared to NK in a given system is due to the recruitment of new killer cells and not to the increase of the number of targets killed by already active cells. The experiments involved comparative limiting dilution of NK and IAK effectors against Daudi target cells showing an increased number of active samples in IAK (17) and elimination of NK cells by preincubation of lymphocyte populations with fibroblast monolayers, which left a residue of cells that were not NK-active by themselves but could be induced by interferon treatment (11). Another suggestive result was obtained in an in vivo system. Rats were depleted of NK activity by radiation or drug treatment. Inoculation of interferon inducers led to the reappearance of cytotoxic cells after a short time (18). In order to characterize the cytotoxic precursors we have exposed peripheral lymphocyte subsets to interferon and tested their activity against K562 and Daudi cells. MATERIALS AND METHODS Fractionation of Effector Cells on the Basis of Surface Markers. Lymphocytes were separated from 200 ml of human blood collected from regular blood donors. Our aim was to obtain lymphocyte subpopulations that express ERs of different affinity with and without a concomitant FcR. Separation according to scheme A in Fig. 1 was carried out in four steps. The starting population was depleted of macro- phages as described (19). Thereafter it was divided on the basis of nylon wool adherence (step 1). The nonadherent population was rosetted with sheep erythrocytes (SRBC) in the presence of dextran. This procedure gave the Edx+ and Edx- subsets after centrifugation on Ficoll/Hypaque (step 2). After the in- dicator erythrocytes were removed from the Edx+ cells, both fractions were subdivided into EA-rosetting and EA-negative cells (step 3). The Edx+Fc+ and Edx+Fc- cells were divided into high- and low-affinity ER-carrying T cells by rerosetting them with SRBC in 10% (vol/vol) fetal calf serum (step 4). Cell yields in the fractions obtained in one experiment are included in Table 2. The Edx- cells lack receptors for SRBC. The Edx+ popula- tion contains all SRBC-rosetting cells, the majority with high- affinity (Ehigh) and low-affinity (E10,) receptors. These Ehigh and E10, were separated by rerosetting the Edx+ subset in 10% fetal calf serum, which left the Elo, cells without attachment of SRBC. They were thus recovered in the interphase. In a second series of experiments the separation was simpli- fied (scheme B, Fig. 1). Macrophage-deprived populations were Abbreviations: NK, natural killing; IAK, interferon-activated killing; FcR, Fc receptor; SRBC, sheep erythrocytes; ER, SRBC receptor; Edx, cells rosetting with SRBC in presence of dextran. * Present address: Dr. Karl Thomae GmbH, Abt. Biol. Forschung/ Mikrobiol. D-7950 Biberach/Riss, West Germany. 3620 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "ad- vertisement" in accordance with 18 U. S. C. §1734 solely to indicate this fact.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Proc. Nat!. Acad. Sci. USAVol. 77, No. 6, pp. 3620-43624, June 1980Immunology
Target selectivity of interferon-induced human killer lymphocytesrelated to their Fc receptor expression
(natural killer cells)
MARIA G. MASUCCI, GIUSEPPE MASUCCI, EVA KLEIN, AND WOLFGANG BERTHOLD*Department of Tumor Biology, Karolinska Institutet, S-104 01 Stockholm, Sweden
Communicated by George Klein, February 25, 1980
ABSTRACT Human blood lymphocytes were fractionatedon the basis of surface characteristics such as adherence to nylonwool and expression of erythrocyte (E) and Fc receptors. Thevarious subsets were incubated with interferon for 3 hr. Twocell lines that differ in sensitivity to the natural killer effect,K562 and Daudi, were exposed to these lymphocytes (i.e., theirsensitivity to interferon-activated killing was tested). Cell lineDaudi, with a low sensitivity to the natural killer effect, was alsoaffected by interferon-activated killing. The efficiency of thenonadherent subsets, separated accor in to the expression ofE receptor, ranked similarly in natural killing (anti-K562) andinterferon-activated killing (anti-K562 and anti-Daudi) in thefollowing order: E receptor-negative cells, low-affinity E re-ceptor-positive cells, and high-affinity E receptor-positive cells.Further separation on the basis of Fc receptor expression re-vealed a difference between the two targets. The Fc receptor-positive and -negative cells that did not express high-affinity Ereceptors killed K562 with similar efficiency whereas Daudicells were more sensitive to the effect of cells devoid of Fc re-ceptor. Results obtained with other targets suggested thatT celllines behave similarly to K562 and that the difference may begenerally valid for T and B cell lines.
Human blood lymphocytes have a lytic effect in vitro againstcertain tumor cell lines (1). The active population is heteroge-neous with regard to its cell surface characteristics (2-7). The"null" subset, obtained after removal of surface immuno-globulin-positive cells and readily resetting T lymphocytes, ishighly enriched in activity (2-4). This subset contains a highproportion of Fc receptor (FcR)-positive cells. In addition, Tlymphocytes with low-affinity sheep erythrocyte receptors(ERs) are also active (5-7). The killing mechanism is unknown.In short-term assays, T cell-derived lymphoid lines are moresensitive than B lines (8). Blood lymphocytes of the majority ofdonors do not kill Daudi cells, a Burkitt lymphoma-derived Bcell line carrying Epstein-Barr virus (9). In long-term tests, thistarget is also lysed, however. Lysis begins after 3-4 hr,suggesting an activation of the effectors during this period.An apparently nondiscriminative cytotoxic potential is also
generated in mixed cultures of lymphocytes and tumor cells.Fibroblasts derived from normal tissues did not generate suchcytotoxicity, neither in allogeneic nor in autologous combina-tions (10, 11). In cultures in which immune interactions occur,interferon is produced. This is probably responsible for thetriggering because the exogenous interferon treatment activateslymphocytes promptly for cytotoxicity (12-16). The Daudi lineis sensitive to interferon-activated killing (IAK), and the kineticsof cytotoxicity in such a system is similar against Daudi andK562 cells (17).
Cell line K562 is highly sensitive to natural killing (NK).Because the large difference in the sensitivity of Daudi andK562 to lymphocyte-mediated lysis disappears when inter-feron-triggered lymphocytes are used as effectors, it is likely
that interferon activates the population to express its maximalcytotoxic potential.Two sets of experiments have provided evidence that the
increased cytotoxicity of IAK as compared to NK in a givensystem is due to the recruitment of new killer cells and not tothe increase of the number of targets killed by already activecells. The experiments involved comparative limiting dilutionof NK and IAK effectors against Daudi target cells showing anincreased number of active samples in IAK (17) and eliminationof NK cells by preincubation of lymphocyte populations withfibroblast monolayers, which left a residue of cells that werenot NK-active by themselves but could be induced by interferontreatment (11). Another suggestive result was obtained in anin vivo system. Rats were depleted of NK activity by radiationor drug treatment. Inoculation of interferon inducers led to thereappearance of cytotoxic cells after a short time (18).
In order to characterize the cytotoxic precursors we haveexposed peripheral lymphocyte subsets to interferon and testedtheir activity against K562 and Daudi cells.
MATERIALS AND METHODSFractionation of Effector Cells on the Basis of Surface
Markers. Lymphocytes were separated from 200 ml of humanblood collected from regular blood donors. Our aim was toobtain lymphocyte subpopulations that express ERs of differentaffinity with and without a concomitant FcR.
Separation according to scheme A in Fig. 1 was carried outin four steps. The starting population was depleted of macro-phages as described (19). Thereafter it was divided on the basisof nylon wool adherence (step 1). The nonadherent populationwas rosetted with sheep erythrocytes (SRBC) in the presenceof dextran. This procedure gave the Edx+ and Edx- subsetsafter centrifugation on Ficoll/Hypaque (step 2). After the in-dicator erythrocytes were removed from the Edx+ cells, bothfractions were subdivided into EA-rosetting and EA-negativecells (step 3). The Edx+Fc+ and Edx+Fc- cells were dividedinto high- and low-affinity ER-carrying T cells by rerosettingthem with SRBC in 10% (vol/vol) fetal calf serum (step 4). Cellyields in the fractions obtained in one experiment are includedin Table 2.The Edx- cells lack receptors for SRBC. The Edx+ popula-
tion contains all SRBC-rosetting cells, the majority with high-affinity (Ehigh) and low-affinity (E10,) receptors. These Ehighand E10, were separated by rerosetting the Edx+ subset in 10%fetal calf serum, which left the Elo, cells without attachmentof SRBC. They were thus recovered in the interphase.
In a second series of experiments the separation was simpli-fied (scheme B, Fig. 1). Macrophage-deprived populations were
Abbreviations: NK, natural killing; IAK, interferon-activated killing;FcR, Fc receptor; SRBC, sheep erythrocytes; ER, SRBC receptor; Edx,cells rosetting with SRBC in presence of dextran.* Present address: Dr. Karl Thomae GmbH, Abt. Biol. Forschung/Mikrobiol. D-7950 Biberach/Riss, West Germany.
3620
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.

Proc. Natl. Acad. Sci. USA 77 (1980) 3621
Scheme AStep 1. Passage through wool
Step 2. E rosetting inpresence of dextran
Step 3. EA rosetting
Step 4. E resetting
Total population
Nylon passed
Edxg
Edx+EA+ Edx EA
FgI
I hEI IEhighEA* EjOwEA+ EhighEA- EjOwEA-
Nylon adherent
Edx-
Edx-EA' Edx-EA-
Scheme B
Total population
Step 1. E rosetting
Ehigh
Step 2. EA rosetting-1
E-EA* E-EA-FIG. 1. Separation of lymphocyte subsets. The total population was depleted of macrophages. Edx, cells resetting with sheep erythrocytes
in the presence of dextran.
depleted of high-affinity E-positive T lymphocytes by SRBCresetting in 10% fetal calf serum (step 1). The SRBC-negativepopulation collected from the interphase after centrifugationon Ficoll/Hypaque was subfractionated into EA-rosetting andEA-negative fractions (step 2).The E- population collected from the interphase was het-
erogeneous. Because the nylon wool adherent cells were notremoved, it contained, in addition to null cells, B cells and Tcells that bear low-affinity ER (i.e., the Elow fraction).
Separation of Cells on the Basis of Adherence to NylonWool. The procedure of Julius et al. (20) was used. A maximumof 80 X 106 cells were loaded on a 5-ml column. The attachedcells were recovered mechanically by pressing the nylon inRPMI medium containing 10% fetal calf serum at 370C.
Separation of Cells on the Basis of SRBC Rosetting Ca-pacity. SRBC were suspended in RPMI medium containing10% fetal calf serum with or without Dextran 70 (Pharmacia).Dextran enhances the E resetting (21). The mixture was incu-bated at 370C for 15 min and thereafter in ice for 1 hr. Afterincubation the cells were carefully resuspended and centrifugedon Ficoll/Hypaque at 500 X g for 25 min. The cells in the in-
terphase and pellet were collected. The indicator erythrocyteswere lysed by treatment with distilled water.
Separation of Cells on the Basis of EA Rosetting Capacity.Ox erythrocytes that had been washed three times were exposedto the maximal subagglutinating amount of rabbit anti-oxerythrocytes (kindly provided by B. Andersson) at 370C for 30min. After two washings, 2.5% of the EA suspension was addedat 4:1 volume ratio to a lymphocyte suspension (106/ml) inRPMI medium containing 10% fetal calf serum. The mixturewas centrifuged and incubated at 370C for 30 min. After in-cubation the cells were carefully resuspended and centrifugedon Ficoll/Hypaque at 500 X g for 25 min. The cells in the in-terphase and the pellet were collected. The indicator erythro-cytes were lysed by treatment with distilled water.
Interferon Treatment of Effector Lymphocytes. Lym-phocytes (5 X 106) from each subset were suspended in 1 ml ofRPMI medium containing 10% fetal calf serum. Parallel ali-quots were incubated for 3 hr. To one aliquot 1000 units of in-terferon (provided by G. Bodo) was added. Interferon was
prepared from Namalwa cells induced by Sendai virus. Thepreparation was purified to 2.2 X 105 International Units/mg
Table 1. Comparison of interferon (IF)-induced increase of cytotoxic activity against K562 and Daudi cellsK562 Daudi
% 51CrLymphocyte release
donor (a)
No. ofeffectors X 10-3*
-IF (b) +IF (c)
Increase incytotoxicpotentialt
(d)
% 5'Crrelease
(e)
No. ofeffectors x 10-3*
-IF (f) +IF (g)
Increase incytotoxicpotentialt
(h)
1 30 35.2 14.0 2.51 10 41.6 6.8 6.112 20 41.6 22.4 1.85 5 40.0 3.4 11.763 20 41.6 22.4 1.85 10 50.0 5.2 9.614 30 12.4 8.8 1.40 10 37.6 4.0 9.405 25 66.4 52.0 1.27 5 100.0 11.0 9.096 30 10.8 5.2 2.07 10 57.6 5.2 11.07
Mean SD 1.82 :+ 0.44 Mean ± SD 9.47 ± 1.93* The number of effectors that gave the indicated 51Cr release was derived from the dose-response curves. Columns b and
f, untreated cells; columns c and g, interferon-treated cells.t The increase was calculated as the ratio between the values in columns b and c for K562 and the values in columns f andg for Daudi.
Immunology: Masucci et al.
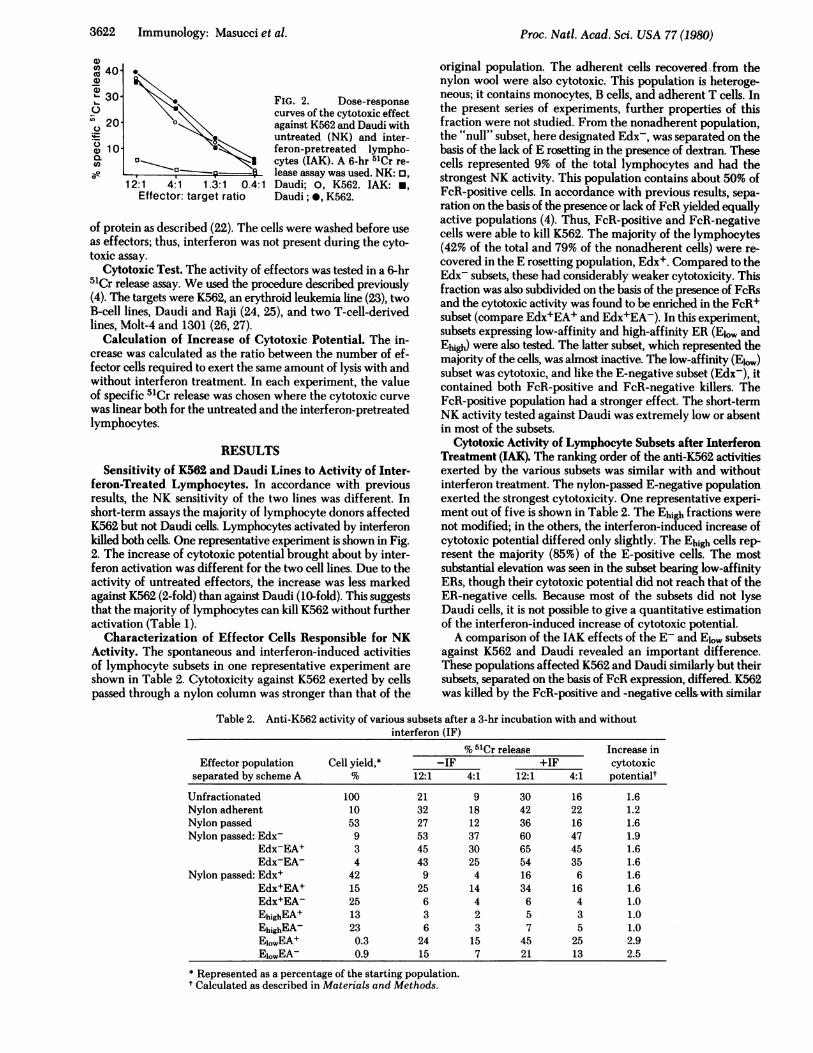
3622 Immunology: Masucci et al.
12:1 4:1 1.3:1 0.4:1Effector: target ratio
FIG. 2. Dose-responsecurves of the cytotoxic effectagainst K562 and Daudi withuntreated (NK) and inter-feron-pretreated lympho-cytes (LAK). A 6-hr 51Cr re-lease assay was used. NK:Daudi; 0, K562. IAK: *,Daudi; 0, K562.
of protein as described (22). The cells were washed before use
as effectors; thus, interferon was not present during the cyto-toxic assay.
Cytotoxic Test. The activity of effectors was tested in a 6-hr51Cr release assay. We used the procedure described previously(4). The targets were K562, an erythroid leukemia line (23), twoB-cell lines, Daudi and Raji (24, 25), and two T-cell-derivedlines, Molt-4 and 1301 (26, 27).
Calculation of Increase of Cytotoxic Potential. The in-crease was calculated as the ratio between the number of ef-fector cells required to exert the same amount of lysis with andwithout interferon treatment. In each experiment, the valueof specific 5'Cr release was chosen where the cytotoxic curvewas linear both for the untreated and the interferon-pretreatedlymphocytes.
RESULTSSensitivity of K562 and Daudi Lines to Activity of Inter-
feron-Treated Lymphocytes. In accordance with previousresults, the NK sensitivity of the two lines was different. Inshort-term assays the majority of lymphocyte donors affectedK562 but not Daudi cells. Lymphocytes activated by interferonkilled both cells. One representative experiment is shown in Fig.2. The increase of cytotoxic potential brought about by inter-feron activation was different for the two cell lines. Due to theactivity of untreated effectors, the increase was less markedagainst K562 (2-fold) than against Daudi (10-fold). This suggeststhat the majority of lymphocytes can kill K562 without furtheractivation (Table 1).
Characterization of Effector Cells Responsible for NKActivity. The spontaneous and interferon-induced activitiesof lymphocyte subsets in one representative experiment areshown in Table 2. Cytotoxicity against K562 exerted by cellspassed through a nylon column was stronger than that of the
original population. The adherent cells recovered-from thenylon wool were also cytotoxic. This population is heteroge-neous; it contains monocytes, B cells, and adherent T cells. Inthe present series of experiments, further properties of thisfraction were not studied. From the nonadherent population,the "null" subset, here designated Edx-, was separated on thebasis of the lack of E rosetting in the presence of dextran. Thesecells represented 9% of the total lymphocytes and had thestrongest NK activity. This population contains about 50% ofFcR-positive cells. In accordance with previous results, sepa-ration on the basis of the presence or lack of FcR yielded equallyactive populations (4). Thus, FcR-positive and FcR-negativecells were able to kill K562. The majority of the lymphocytes(42% of the total and 79% of the nonadherent cells) were re-covered in the E rosetting population, Edx+. Compared to theEdx- subsets, these had considerably weaker cytotoxicity. Thisfraction was also subdivided on the basis of the presence of FcRsand the cytotoxic activity was found to be enriched in the FcR+subset (compare Edx+EA+ and Edx+EA-). In this experiment,subsets expressing low-affinity and high-affinity ER (El., andEhigh) were also tested. The latter subset, which represented themajority of the cells, was almost inactive. The low-affinity (Eiow)subset was cytotoxic, and like the E-negative subset (Edx-), itcontained both FcR-positive and FcR-negative killers. TheFcR-positive population had a stronger effect. The short-termNK activity tested against Daudi was extremely low or absentin most of the subsets.
Cytotoxic Activity of Lymphocyte Subsets after InterferonTreatment (IAK). The ranking order of the anti-K562 activitiesexerted by the various subsets was similar with and withoutinterferon treatment. The nylon-passed E-negative populationexerted the strongest cytotoxicity. One representative experi-ment out of five is shown in Table 2. The Ehigh fractions werenot modified; in the others, the interferon-induced increase ofcytotoxic potential differed only slightly. The Ehigh cells rep-resent the majority (85%) of the E-positive cells. The mostsubstantial elevation was seen in the subset bearing low-affinityERs, though their cytotoxic potential did not reach that of theER-negative cells. Because most of the subsets did not lyseDaudi cells, it is not possible to give a quantitative estimationof the interferon-induced increase of cytotoxic potential.A comparison of the IAK effects of the E- and Elow subsets
against K562 and Daudi revealed an important difference.These populations affected K562 and Daudi similarly but theirsubsets, separated on the basis of FcR expression, differed. K562was killed by the FcR-positive and -negative cellswith similar
Table 2. Anti-K562 activity of various subsets after a 3-hr incubation with and withoutinterferon (IF)
% 5'Cr release Increase inEffector population Cell yield,* -IF +IF cytotoxic
separated by scheme A % 12:1 4:1 12:1 4:1 potentialt
Unfractionated 100 21 9 30 16 1.6Nylon adherent 10 32 18 42 22 1.2Nylon passed 53 27 12 36 16 1.6Nylon passed: Edx- 9 53 37 60 47 1.9
Edx-EA+ 3 45 30 65 45 1.6Edx-EA- 4 43 25 54 35 1.6
Nylon passed: Edx+ 42 9 4 16 6 1.6Edx+EA+ 15 25 14 34 16 1.6Edx+EA- 25 6 4 6 4 1.0EhighEA+ 13 3 2 5 3 1.0EhighEA- 23 6 3 7 5 1.0ElowEA+ 0.3 24 15 45 25 2.9EjowEA- 0.9 15 7 21 13 2.5
* Represented as a percentage of the starting population.t Calculated as described in Materials and Methods.
Proc. Nati. Acad. Sci. USA 77 (1980)
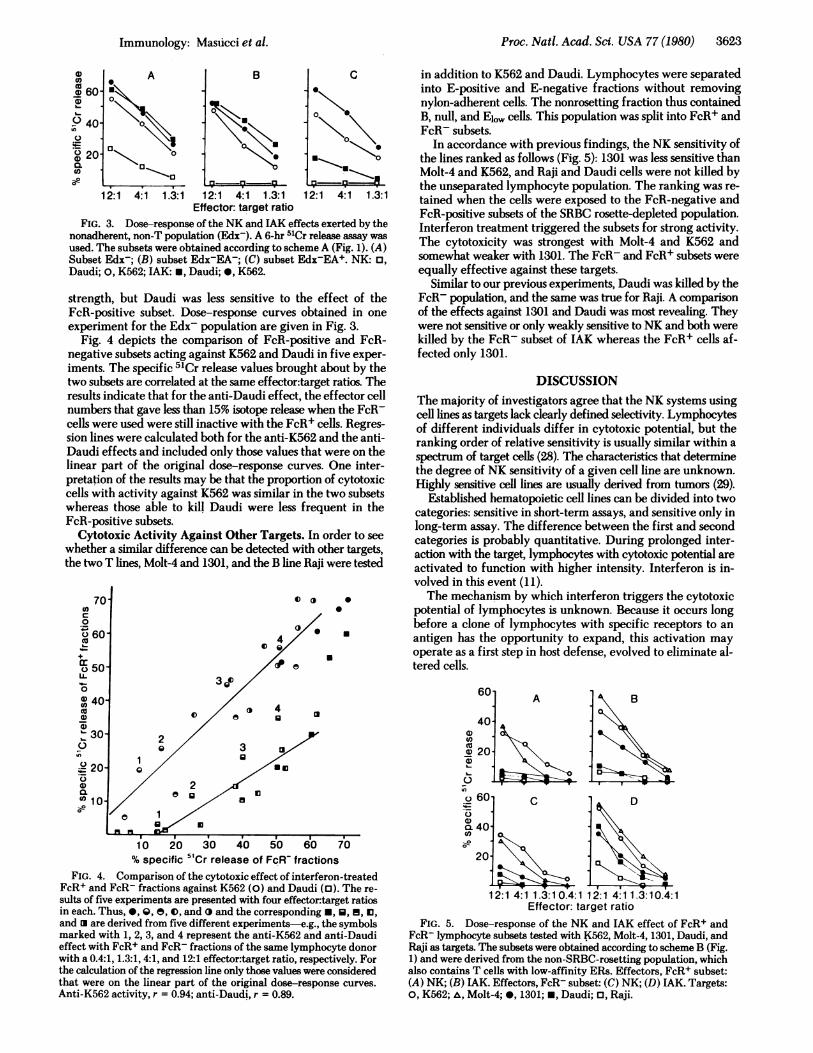
Proc. Natl. Acad. Sci. USA 77 (1980) 3623
C
I .N0
12:1 4:1 1.3:1 12:1 4:1 1.3:1 12:1 4:1 1.3:1Effector: target ratio
FIG. 3. Dose-response of the NK and IAK effects exerted by thenonadherent, non-T population (Edx-). A 6-hr 51Cr release assay wasused. The subsets were obtained according to scheme A (Fig. 1). (A)Subset Edx-; (B) subset Edx-EA-; (C) subset Edx-EA+. NK: o,Daudi; 0, K562; IAK: *, Daudi; *, K562.
strength, but Daudi was less sensitive to the effect of theFcR-positive subset. Dose-response curves obtained in oneexperiment for the Edx- population are given in Fig. 3.
Fig. 4 depicts the comparison of FcR-positive and FcR-negative subsets acting against K562 and Daudi in five exper-iments. The specific 5'Cr release values brought about by thetwo subsets are correlated at the same effector:target ratios. Theresults indicate that for the anti-Daudi effect, the effector cellnumbers that gave less than 15% isotope release when the FcR-cells were used were still inactive with the FcR+ cells. Regres-sion lines were calculated both for the anti-K562 and the anti-Daudi effects and included only those values that were on thelinear part of the original dose-response curves. One inter-pretation of the results may be that the proportion of cytotoxiccells with activity against K562 was similar in the two subsetswhereas those able to kill Daudi were less frequent in theFcR-positive subsets.
Cytotoxic Activity Against Other Targets. In order to seewhether a similar difference can be detected with other targets,the two T lines, Molt-4 and 1301, and the B line Raji were tested
o ( 0
*
.
3 j0
m
in addition to K562 and Daudi. Lymphocytes were separatedinto E-positive and E-negative fractions without removingnylon-adherent cells. The nonrosetting fraction thus containedB, null, and Elw cells. This population was split into FcR+ andFcR- subsets.
In accordance with previous findings, the NK sensitivity ofthe lines ranked as follows (Fig. 5): 1301 was less sensitive thanMolt-4 and K562, and Raji and Daudi cells were not killed bythe unseparated lymphocyte population. The ranking was re-tained when the cells were exposed to the FcR-negative andFcR-positive subsets of the SRBC rosette-depleted population.Interferon treatment triggered the subsets for strong activity.The cytotoxicity was strongest with Molt-4 and K562 andsomewhat weaker with 1301. The FcR- and FcR+ subsets wereequally effective against these targets.
Similar to our previous experiments, Daudi was killed by theFcR- population, and the same was true for Raji. A comparisonof the effects against 1301 and Daudi was most revealing. Theywere not sensitive or only weakly sensitive to NK and both werekilled by the FcR- subset of IAK whereas the FcR+ cells af-fected only 1301.
DISCUSSIONThe majority of investigators agree that the NK systems usingcell lines as targets lack clearly defined selectivity. Lymphocytesof different individuals differ in cytotoxic potential, but theranking order of relative sensitivity is usually similar within aspectrum of target cells (28). The characteristics that determinethe degree of NK sensitivity of a given cell line are unknown.Highly sensitive cell lines are usually derived from tumors (29).
Established hematopoietic cell lines can be divided into twocategories: sensitive in short-term assays, and sensitive only inlong-term assay. The difference between the first and secondcategories is probably quantitative. During prolonged inter-action with the target, lymphocytes with cytotoxic potential areactivated to function with higher intensity. Interferon is in-volved in this event (11).The mechanism by which interferon triggers the cytotoxic
potential of lymphocytes is unknown. Because it occurs longbefore a clone of lymphocytes with specific receptors to anantigen has the opportunity to expand, this activation mayoperate as a first step in host defense, evolved to eliminate al-tered cells.
60
a)
Coa)
a)
a)L-
QaeCo-10 20 30 40 50 60 70
% specific 51Cr release of FcR- fractionsFIG. 4. Comparison of the cytotoxic effect of interferon-treated
FcR+ and FcR- fractions against K562 (0) and Daudi (l). The re-sults of five experiments are presented with four effector:target ratiosin each. Thus, *, Q, e, 0, and and the corresponding *, U, f, 13,and are derived from five different experiments-e.g., the symbolsmarked with 1, 2, 3, and 4 represent the anti-K562 and anti-Daudieffect with FcR+ and FcR- fractions of the same lymphocyte donorwith a 0.4:1, 1.3:1, 4:1, and 12:1 effector:target ratio, respectively. Forthe calculation of the regression line only those values were consideredthat were on the linear part of the original dose-response curves.Anti-K562 activity, r = 0.94; anti-Daudi, r = 0.89.
12:1 4:1 1.3:10.4:1 12:1 4:11.3:10.4:1Effector: target ratio
FIG. 5. Dose-response of the NK and 1AK effect of FcR+ andFcR- lymphocyte subsets tested with K562, Molt-4, 1301, Daudi, andRaji as targets. The subsets were obtained according to scheme B (Fig.1) and were derived from the non-SRBC-rosetting population, whichalso contains T cells with low-affinity ERs. Effectors, FcR+ subset:(A) NK; (B) 1AK. Effectors, FcR- subset: (C) NK; (D) IAK. Targets:0, K562; A, Molt-4; 0, 1301; *, Daudi; 0, Raji.
am60
co
OD 60
° 40'u 20
C
Tat
A
I- a W"
B
4)Coco4)
z
._
0
4)
aP
Immunology: Masticei et al.
i:

3624 Immunology: Masucci et al.
The results with K562 and Daudi targets show that the killerlymphocyte population is heterogeneous with regard to theintensity of function. The majority kills the highly sensitiveK562 cells. Interferon stimulation brings them to high, perhapsfull, expression of their potential. As a consequence, inter-feron-treated cells can also kill targets that cannot be lysed bythe nontriggered NK effectors (like Daudi or Raji) with a lowerfunctional level. The threshold of the required intensity of lyticfunction probably differs for different targets, which is reflectedin the dose-response curves.The cytotoxicity of unmanipulated fresh lymphocytes is
mediated by a population that is heterogeneous with regard tosurface properties. B cells with surface Ig and strongly SRBCrosetting T cells are inactive. This latter fraction was nottriggered by interferon treatment. The "null" cells and the Tcells with relatively low-affinity ERs are responsible for the NKeffect, and interferon acts on these subsets.Our results confirm the findings of Trinchieri et al. (13), who
analyzed the lymphocyte populations with regard to three typesof cytotoxicity, NK, antibody-dependent cell-mediated cyto-toxicity (ADCC), and IAK. They separated the effectors on thebasis of nylon adherence and E resetting capacity and con-cluded that these three functions resided in the same subsets.Interferon did not recruit cytotoxic cells from populations withno or low NK effect. They characterized potentially cytotoxiccells as nonphagocytic, nonadherent, and devoid of surface Ig.
Without further fractionation, the null and "low-affinity"E populations give similar dose-response curves against Daudiand K562 in our experiments. Subfractionation on the basis ofFcR expression revealed certain selectivity, however, becauseDaudi was less sensitive to the effect of FcR-positive cells. Re-sults with additional targets suggested that this selectivity maybe general; T lines tended to resemble K562, whereas B lineslike Daudi were killed preferentially by FcR-negative cells.
Results obtained previously with blood lymphocytes of pa-tients with acute mononucleosis may be due to this differencebetween T and B lines (30). With a separation procedure similarto the one used in the present paper, elimination of FcR-positiveT cells (performed in order to deplete NK cells) abrogated ordecreased the cytotoxic activity against K562 and 1301 whereasDaudi was affected preferentially by FcR-negative T cells (31).The lymphocyte population contains T blasts in the acute phaseof IM and the proportion of FcR-positive T cells is lower thanin blood of healthy donors (32).The concomitant presence of cytotoxic cells with different
surface characteristics and selectivity was encountered previ-ously. In one of our studies (33), the specific allo-killer cells ofmixed lymphocyte cultures were enriched in the high-affinityER-carrying cell fractions. In another investigation (34) welooked for these killers in subsets separated according to FcRs;they were enriched in the FcR- subsets. The "anomalous" (i.e.,anti-K562) 'cytotoxicity that developed in parallel with thespecific cytotoxicity was stronger in the subset containing "low"E and null cells (30) and it was similar in the FcR-positive andnegative subsets (31).The practical implication of our finding is that results with
fractionated effector populations on the basis of FcR expressionmay differ according to the target cell type used. PreviouslyFcR-positive cells were considered to be mainly responsible forthe NK effect and specific cytotoxicities were searched for aftertheir removal. Because FcR-negative killers are more abundantin activated lymphocyte populations (33) and targets may varyin sensitivity to effectors according to expression of this receptor,removal of FcR-positive cells does not insure elimination of anondisease-related, indiscriminate cytotoxicity.
This investigation was supported by Grant 1 R01 CA25250-01 fromthe National Cancer Institute and by the Swedish Cancer Society.M.G.M. is the recipient of a fellowship from the Foundation "Blan-cefort-Boncompagni-Ludovisi, f6dd Bildt", Stockholm, Sweden. W.B.is the recipient of a European Molecular Biology Organization Fel-lowship.
1. Pross, H. F. & Baines, M. G. (1977) Cancer Immunol. Immu-nother. 3, 75-85.
2. Hersey, P., Edwards, A., Edwards, J., Adams, E., Milton, G. W.& Nelson, D. S. (1975) Int. J. Cancer 16, 173-183.
3. Bakacs, T., Gergely, P., Cornain, S. & Klein, E. (1977) Int. J.Cancer 19, 441-449.
4. Bakics, T., Gergely, P. & Klein, E. (1977) Cell. Immunol. 32,317-328.
5. West, H. W., Cannon, G. B., Kay, H. D., Ronnard, G. B. & Her-berman, R. B. (1977) J. Immunol. 118,355-361.
6. Bakics, T., Klein, E., Yefenof, E., Gergely, P. & Steinitz, M. (1978)Z. Immunitatsforsch.-Immunobiol. 154,121-134.
7. Kaplan, J. & Callewaert, D. (1978) J. Nati. Cancer Inst. 60,961-964.
8. Ono, A., Amos, D. B. & Koren, H. S. (1977) Nature (London) 266,546-547.
9. Masucci, M. G., Klein, E. & Argov, S. (1980) J. Immunol., inpress.
10. Chandon, M. R., Vanky, F., Carnaud, C. & Klein, E. (1975) Int.J. Cancer 15,342-350.
11. Saksela, E., Timonen, T. & Cantell, K. (1979) Scand. J. Immunol.10,257-266.
12. Virelizier, J. L., Allison, A. G. & de Maeyer, E. (1977) Infect.Immun. 17, 282-285.
13. Trinchieri, G., Santoli, D., Dee, R. R. & Knowles, B. (1978) J. Exp.Med. 147, 1299-1313.
14. Santoli, D., Trinchieri, G. & Koprowski, H. (1978) J. Immunol.121,532-538.
15. Svet-Moldavsky, G. J. & Chernyakhovskaya, I. J. (1967) Nature(London) 215, 1299-1300.
16. Einhorn, S., Blomgren, H. & Strander, H. (1978) Int. J. Cancer22,405-412.
17. Masucci, M. G., Masucci, G., Klein, E. & Berthold, W. (1980) inNew Trends in Human Immunology and Cancer Immunoth-erapy Symposium (Montpellier), in press.
18. Oehler, J. R., Lindsay, R. L., Nunn, M. E., Holden, H. T. &Herberman, R. B. (1978) Int. J. Cancer 21, 210-220.
19. Lundgren, G., Zukoski, Ch. F., and M6ller, G. (1968) Clin. Exp.Immunol. 3, 817-836.
20. Julius, M. H., Simpson, E. & Herzenberg, L. A. (1973) Eur. J.Immunol. 3, 645-649.
21. Brown, C. S., Halpern, H. & Wortis, H. H. (1975) Clin. Exp.Immunol. 20, 505-512.
22. Bodo, G. (1977) in Production and Purification of Lympho-bastoid Interferon, ed. Ikie, D. (Acad. Sci. Acts. Proc. Symp. onPreparation, Standardization and Clinical Use of Interferon,Zagreb, June 8-9), pp. 49-57.
23. Lozzio, C. B. & Lozzio, B. B. (1973) J. Nat. Cancer Inst. 50,535-538.
24. Klein, E., Klein, G., Nadkarni, J. S., Nadkarni, J. J., Wigzell, H.& Clifford, P. (1968) Cancer Res. 28, 1300-1310.
25. Pulvertaft, R. J. V. (1965) J. Clin. Pathol. 18,261-273.26. Minowada, J., Chnuma, T. & Moore, C. E. (1972) J. Natl. Cancer
Inst. 49, 891-895.27. McCarthy, R. E. (1965) Cancer 18,522-529.28. Callewaert, D. M., Kaplan, J., Johnson, D. F. & Peterson, W. D.
(1979) Cell. Immunol. 42, 103-112.29. Jondal, M., Spino, C. & Targan, S. (1978) Nature (London) 272,
62-64.30. Svedmyr, E. A. & Jondal, M. (1975) Proc. Nati. Acad. Sci. USA
72, 1622-1626.31. Bakacs, T., Svedmyr, E., Klein, E., Rombo, L. & Weiland, 0.
(1978) Cancer Lett. 4, 185-189.32. Haynes, B. F., Schooley, R. T., Grouse, J. E., Payling-Wright, C.
R., Dolin, R. & Fauci, A. S. (1979) J. Immunol. 2, 699-702.33. Poros, A. & Klein, E. (1979) Cell. Immunol. 46,57-68.34. Seeley, J. K., Masucci, G., Poros, A., Klein, E. & Golub, S. (1979)
J. Immunol. 123, 1303-1311.
Proc. Natl. Acad. Sci. USA 77 (1980)
Related Documents











