ELSEVIER Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358 Taphonomic comparison between Recent and fossil sand dollars James Nebelsick * Institute and Museum of Geology and Palaeontology, Sigwartstrasse 10, D-72076 Tu ¨bingen, Germany Received 28 January 1997; revised version received 9 July 1997; accepted 8 June 1998 Abstract The taphonomy of a Recent and a fossil sand dollar are compared. The recent Echinodiscus auritus originates from a shallow-water carbonate environment in the Red Sea. The fossil Parascutella ho ¨barthi is found in micaceous sands of the Lower Miocene Austrian Molasse Zone. Both species show strong similarities in constructional morphology including the flattened overall shape, details of the surface morphology as well as the presence of an internal support system. Three taphonomic features are remarkable different: (1) lethal and non-lethal predation (especially high in the recent E. auritus; (2) encrustation of the test (especially by coralline algae in the fossil Parascutella); and (3) radial cracking on implosion of the body cavity caused by sediment loading on the fossil tests. Differences in taphonomic signatures can be correlated with depositional environment (predation and encrustation) as well as factors related to burial of the fossil specimens (sediment loading and diagenesis). 1999 Elsevier Science B.V. All rights reserved. Keywords: taphonomy; echinoids; clypeasteroids; Miocene; predation; encrustation 1. Introduction The preservation of the echinoids as in all fossils is dependent on the nature and intensity of taphonomic processes affecting the organisms (Fig. 1), whereby a ‘poorly preserved’ skeleton can, in fact, convey more information concerning the palaeoecology of the fos- sil and its environment. The preservation of echinoids can be related to three main factors (Fig. 2). These are skeletal morphology, environmental factors, and temporal effects (Nebelsick, 1995). In this study a di- rect taphonomic comparison is made between a fossil sand dollar (Parascutella ho ¨barthi) from the Lower Miocene of the Austrian Molasse Zone and a Recent species from the Red Sea (Echinodiscus auritus), al- lowing the relative importance of these factors to be evaluated and compared. This direct comparison is L E-mail: [email protected] appropriate due to similarities not only in the con- structional morphology (flattened form with internal support systems) and test surface characters, but also in the environments in which they are found (shallow, subtidal sands). Numerous Recent observations con- cerning the biological and physical factors affecting the preservation of these sea-urchin tests can thus be incorporated in the comparison. Echinoderms obviously represent a special case among invertebrates as far as their skeletal construc- tion is concerned. Although unified by the possession of a stereom and a multiplated skeleton, echino- derms show a wide range of skeletal morphologies and preservation potentials. This has been shown in an increasing number of modern studies concerning specific echinoderm taxa or taphonomic processes affecting them (Meyer and Meyer, 1986; Kidwell and Baumiller, 1990; Donovan, 1991; Greenstein, 1991, 1992; Gordon and Donovan, 1992; Nebel- 0031-0182/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved. PII:S0031-0182(98)00211-9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358
Taphonomic comparison between Recent and fossil sand dollars
James Nebelsick *
Institute and Museum of Geology and Palaeontology, Sigwartstrasse 10, D-72076 Tubingen, Germany
Received 28 January 1997; revised version received 9 July 1997; accepted 8 June 1998
Abstract
The taphonomy of a Recent and a fossil sand dollar are compared. The recent Echinodiscus auritus originates froma shallow-water carbonate environment in the Red Sea. The fossil Parascutella hobarthi is found in micaceous sands ofthe Lower Miocene Austrian Molasse Zone. Both species show strong similarities in constructional morphology includingthe flattened overall shape, details of the surface morphology as well as the presence of an internal support system. Threetaphonomic features are remarkable different: (1) lethal and non-lethal predation (especially high in the recent E. auritus;(2) encrustation of the test (especially by coralline algae in the fossil Parascutella); and (3) radial cracking on implosion ofthe body cavity caused by sediment loading on the fossil tests. Differences in taphonomic signatures can be correlated withdepositional environment (predation and encrustation) as well as factors related to burial of the fossil specimens (sedimentloading and diagenesis). 1999 Elsevier Science B.V. All rights reserved.
Keywords: taphonomy; echinoids; clypeasteroids; Miocene; predation; encrustation
1. Introduction
The preservation of the echinoids as in all fossils isdependent on the nature and intensity of taphonomicprocesses affecting the organisms (Fig. 1), whereby a‘poorly preserved’ skeleton can, in fact, convey moreinformation concerning the palaeoecology of the fos-sil and its environment. The preservation of echinoidscan be related to three main factors (Fig. 2). Theseare skeletal morphology, environmental factors, andtemporal effects (Nebelsick, 1995). In this study a di-rect taphonomic comparison is made between a fossilsand dollar (Parascutella hobarthi) from the LowerMiocene of the Austrian Molasse Zone and a Recentspecies from the Red Sea (Echinodiscus auritus), al-lowing the relative importance of these factors to beevaluated and compared. This direct comparison is
Ł E-mail: [email protected]
appropriate due to similarities not only in the con-structional morphology (flattened form with internalsupport systems) and test surface characters, but alsoin the environments in which they are found (shallow,subtidal sands). Numerous Recent observations con-cerning the biological and physical factors affectingthe preservation of these sea-urchin tests can thus beincorporated in the comparison.
Echinoderms obviously represent a special caseamong invertebrates as far as their skeletal construc-tion is concerned. Although unified by the possessionof a stereom and a multiplated skeleton, echino-derms show a wide range of skeletal morphologiesand preservation potentials. This has been shown inan increasing number of modern studies concerningspecific echinoderm taxa or taphonomic processesaffecting them (Meyer and Meyer, 1986; Kidwelland Baumiller, 1990; Donovan, 1991; Greenstein,1991, 1992; Gordon and Donovan, 1992; Nebel-
0031-0182/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved.PII: S 0 0 3 1 - 0 1 8 2 ( 9 8 ) 0 0 2 1 1 - 9

350 J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358
Fig. 1. Schematic diagram of the taphonomic processes leading to different preservations of an echinoid. The presence of a taphonomicfeature is indicated by the thicker line (modified after Nebelsick, 1995).
sick, 1992, 1996; Nebelsick and Kampfer, 1994).The sand dollars represent extremely flattened formsof clypeasteroid sea-urchins (Durham, 1966; Mooi,1989) and are among the most successful groupsof Recent echinoderms (Smith, 1984; Mooi, 1990).These irregular echinoids show a wide geographicaldistribution occurring in both tropical and temperatesettings (Ghiold and Hoffman, 1984, 1986). They aregenerally restricted to shallow-water, higher-energyenvironments. They include both deposit feeding,endobenthic forms (such as Echinarachnius, Echino-discus, Encope and Melitta) (Bell and Frey, 1969;Stanley and James, 1971; Ebert and Dexter, 1975;Bentley and Cockcroft, 1995), as well as the par-tially exposed, suspension-feeding Dendraster ex-
centricus, an echinoid well known from the westcoast of North America (Birkeland and Chia, 1971;Timko, 1976; Smith, 1981; Beadle, 1989). Sanddollars have an extensive fossil record (Wagner,1974; Zinsmeister, 1980; Dodd et al., 1985; McKin-ney, 1985) including such extinct genera as Scutellaand Parascutella which are characteristic for manyMiocene rocks of the Mediterranean and Caribbeanareas as recorded not only in numerous histori-cal monographs, but also in the modern literature(Philippe, 1983; Boggild and Rose, 1984; Poddubiukand Rose, 1984; Courville et al., 1988). These sanddollars can occur as isolated specimens, but are oftenfound accumulated in great numbers in so called‘Scutella-sands’ (as in the study area).

J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358 351
Fig. 2. Taphonomic triangle showing three factors affectingtaphonomic processes (modified after Nebelsick, 1995).
2. Material and methods
2.1. Echinodiscus auritus from the Red Sea
Echinodiscus auritus Leske, 1778 is a large Indo-Pacific sand dollar reaching a maximum size of 15 cm.It shows a number of highly developed test character-istics. These include deeply incised posterior pairedlunules, a very complex polyfurcating food groovesystem leading to the centrally positioned peristome(mouth) (Fig. 3) and a cover of highly differentiatedminute spines. The periproct (anus) is located midwaybetween the peristome (mouth) and the posterior am-bitus (rim). It has an extensive, dense, internal supportsystem consisting of skeletal connections bridging thegap between the oral (lower) and apical (upper) sur-face. The internal supports are distributed betweenthe central cavity containing the main internal organsof the sea-urchin and the ambitus. Cage experimentsshowed that dead Echinodiscus loses its minute spineswithin a few days (Nebelsick and Kampfer, 1994).It is also susceptible to plate disarticulation, which,once initiated, quickly leads to the disintegration ofthe complete test into larger and smaller fragments(single plates).
Live and dead Echinodiscus were studied in theNorthern Bay of Safaga, Red Sea, Egypt, a shal-low-water carbonate environment (Nebelsick, 1992,1996; Nebelsick and Kampfer, 1994). This speciesis found in coastal sands in depths of less than 10
m. It is most common in a relatively restricted areawith poorly sorted, fine sands (Piller and Mansour,1990) where it reaches maximum densities of 2–3per m2. Both juveniles and adults are found in thesame environment. They live shallowly buried in thesubstrate and, if extracted, bury themselves within10 min. This distribution is similar to that observedfor another species of Echinodiscus (E. bisperfora-tus) from the South African south coast (Bentley andCockcroft, 1995).
2.2. Parascutella hobarthi from the Lower MioceneAustrian Molasse
P. hobarthi (Kuhn, 1936) is a Miocene sanddollar restricted to a single locality in the Mo-lasse Zone of Lower Austria (Kuhn, 1936; Schaffer,1962; Steininger, 1971). The accumulation of a largenumber of specimens allows detailed palaeoecolog-ical and taphonomic analysis. Numerous specimenswere examined from museum collections (Naturhis-torisches Museum, Vienna, Austria; Institute ofPalaeontology, Univ. Vienna, Hobarth Museum inHorn, Austria, and the Krahuletz Museum in Eggen-burg, Austria). Additional specimens were recoveredfrom field excavations which delivered importantsedimentological, palaeoecological and taphonomicinformation.
The sediments in which these echinoids arefound are known as the ‘Scutellensand’ and be-long to the Loibersdorf Formation. This Formationis part of a complex series of highly differentiated,shallow-water sediments near the southern marginof the Bohemian massif (Steininger and Roetzel,1991). They are Eggenburgian in age, a regionalstratigraphic stage of the Paratethys correspondingto the Burdigalian. Sedimentary data show verypoorly sorted, micaceous sands representing a shal-low-water nearshore environment (R. Roetzel, pers.commun., 1996). These sediments have never beenlithified, except for concretionary build-ups of whichthe echinoids often serve as a nucleus. The biota isdominated by the name-giving Parascutella. Otherfossils include coralline algae, mostly in associationwith the sea-urchin tests, bryozoans, calcitic-shelledbivalves including oysters and pectinids as well asthe spines of regular cidaroid echinoids and frag-ments of irregular spatangoid echinoids. The oysters

352 J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358
Fig. 3. Schematic morphological comparison of Echinodiscus and Parascutella. Note the differences in the ambitus (with and withoutlunules), the complexity of the food groove system and the position of the anus.
are commonly found in gregarious clumps. Mouldsof gastropods and aragonitic-shelled bivalves can befound in association with the lithification occurringaround the echinoid tests.
P. hobarthi is a relatively small sand dollar reach-ing a maximum size of 8 cm. It possesses only slightindentation along the ambitus and its food groovesystem is relatively simple (Fig. 3). The tubercu-lation (and hence spine differentiation, see Smith,1984) is not as complex as for Echinodiscus. Theperiproct is located along the posterior ambitus thusdistinguishing this genus from Scutella which pos-sess a periproct halfway between the peristome andposterior ambitus.
The taphonomic comparison of the echinoids wasaccomplished by visual recognition of different fea-
tures found on the tests (Fig. 4). In all, 23 mod-ern specimens and 73 fossil specimens were ana-lyzed. Additional information was gained by scan-ning electron microscope analysis of surface char-acters (Fig. 5). The results of the comparison areshown in Fig. 6.
3. Results
3.1. Taphonomy of Echinodiscus auritus
Dead tests of E. auritus show a very wide rangeof taphonomic features (Nebelsick and Kampfer,1994). Their preservation ranges from complete testswith very well preserved surface features (tubercles
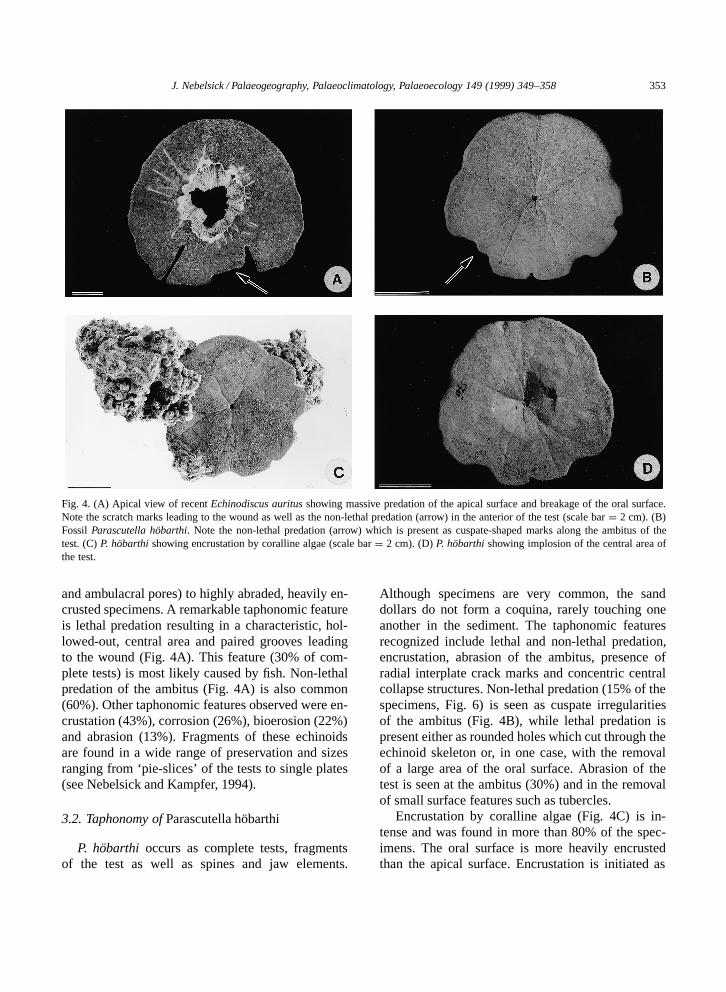
J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358 353
Fig. 4. (A) Apical view of recent Echinodiscus auritus showing massive predation of the apical surface and breakage of the oral surface.Note the scratch marks leading to the wound as well as the non-lethal predation (arrow) in the anterior of the test (scale bar D 2 cm). (B)Fossil Parascutella hobarthi. Note the non-lethal predation (arrow) which is present as cuspate-shaped marks along the ambitus of thetest. (C) P. hobarthi showing encrustation by coralline algae (scale bar D 2 cm). (D) P. hobarthi showing implosion of the central area ofthe test.
and ambulacral pores) to highly abraded, heavily en-crusted specimens. A remarkable taphonomic featureis lethal predation resulting in a characteristic, hol-lowed-out, central area and paired grooves leadingto the wound (Fig. 4A). This feature (30% of com-plete tests) is most likely caused by fish. Non-lethalpredation of the ambitus (Fig. 4A) is also common(60%). Other taphonomic features observed were en-crustation (43%), corrosion (26%), bioerosion (22%)and abrasion (13%). Fragments of these echinoidsare found in a wide range of preservation and sizesranging from ‘pie-slices’ of the tests to single plates(see Nebelsick and Kampfer, 1994).
3.2. Taphonomy of Parascutella hobarthi
P. hobarthi occurs as complete tests, fragmentsof the test as well as spines and jaw elements.
Although specimens are very common, the sanddollars do not form a coquina, rarely touching oneanother in the sediment. The taphonomic featuresrecognized include lethal and non-lethal predation,encrustation, abrasion of the ambitus, presence ofradial interplate crack marks and concentric centralcollapse structures. Non-lethal predation (15% of thespecimens, Fig. 6) is seen as cuspate irregularitiesof the ambitus (Fig. 4B), while lethal predation ispresent either as rounded holes which cut through theechinoid skeleton or, in one case, with the removalof a large area of the oral surface. Abrasion of thetest is seen at the ambitus (30%) and in the removalof small surface features such as tubercles.
Encrustation by coralline algae (Fig. 4C) is in-tense and was found in more than 80% of the spec-imens. The oral surface is more heavily encrustedthan the apical surface. Encrustation is initiated as
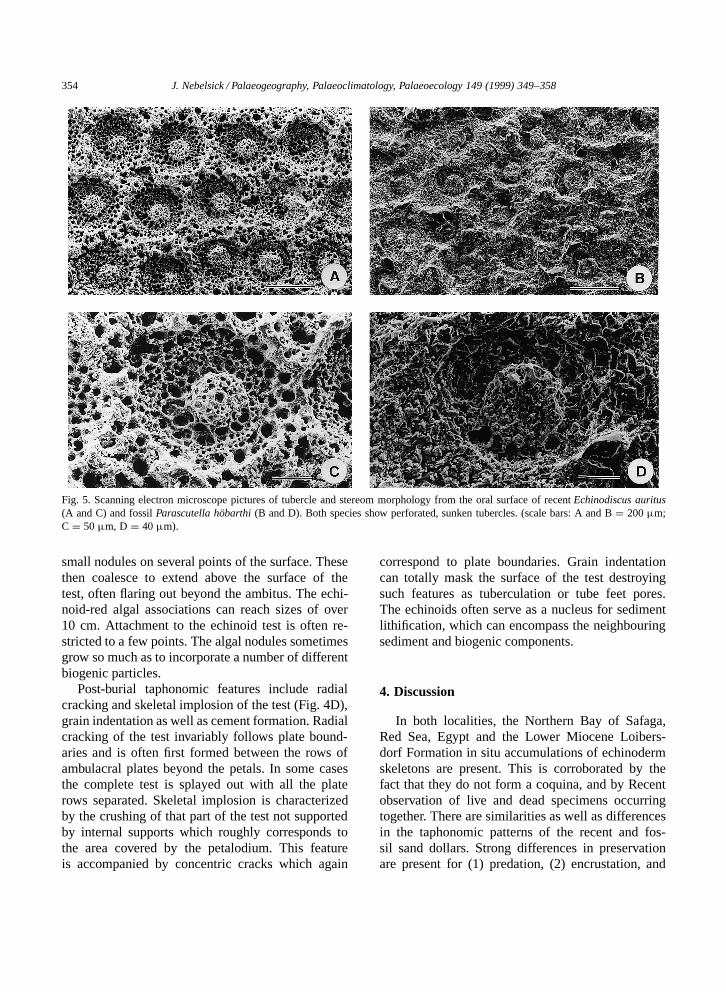
354 J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358
Fig. 5. Scanning electron microscope pictures of tubercle and stereom morphology from the oral surface of recent Echinodiscus auritus(A and C) and fossil Parascutella hobarthi (B and D). Both species show perforated, sunken tubercles. (scale bars: A and B D 200 µm;C D 50 µm, D D 40 µm).
small nodules on several points of the surface. Thesethen coalesce to extend above the surface of thetest, often flaring out beyond the ambitus. The echi-noid-red algal associations can reach sizes of over10 cm. Attachment to the echinoid test is often re-stricted to a few points. The algal nodules sometimesgrow so much as to incorporate a number of differentbiogenic particles.
Post-burial taphonomic features include radialcracking and skeletal implosion of the test (Fig. 4D),grain indentation as well as cement formation. Radialcracking of the test invariably follows plate bound-aries and is often first formed between the rows ofambulacral plates beyond the petals. In some casesthe complete test is splayed out with all the platerows separated. Skeletal implosion is characterizedby the crushing of that part of the test not supportedby internal supports which roughly corresponds tothe area covered by the petalodium. This featureis accompanied by concentric cracks which again
correspond to plate boundaries. Grain indentationcan totally mask the surface of the test destroyingsuch features as tuberculation or tube feet pores.The echinoids often serve as a nucleus for sedimentlithification, which can encompass the neighbouringsediment and biogenic components.
4. Discussion
In both localities, the Northern Bay of Safaga,Red Sea, Egypt and the Lower Miocene Loibers-dorf Formation in situ accumulations of echinodermskeletons are present. This is corroborated by thefact that they do not form a coquina, and by Recentobservation of live and dead specimens occurringtogether. There are similarities as well as differencesin the taphonomic patterns of the recent and fos-sil sand dollars. Strong differences in preservationare present for (1) predation, (2) encrustation, and
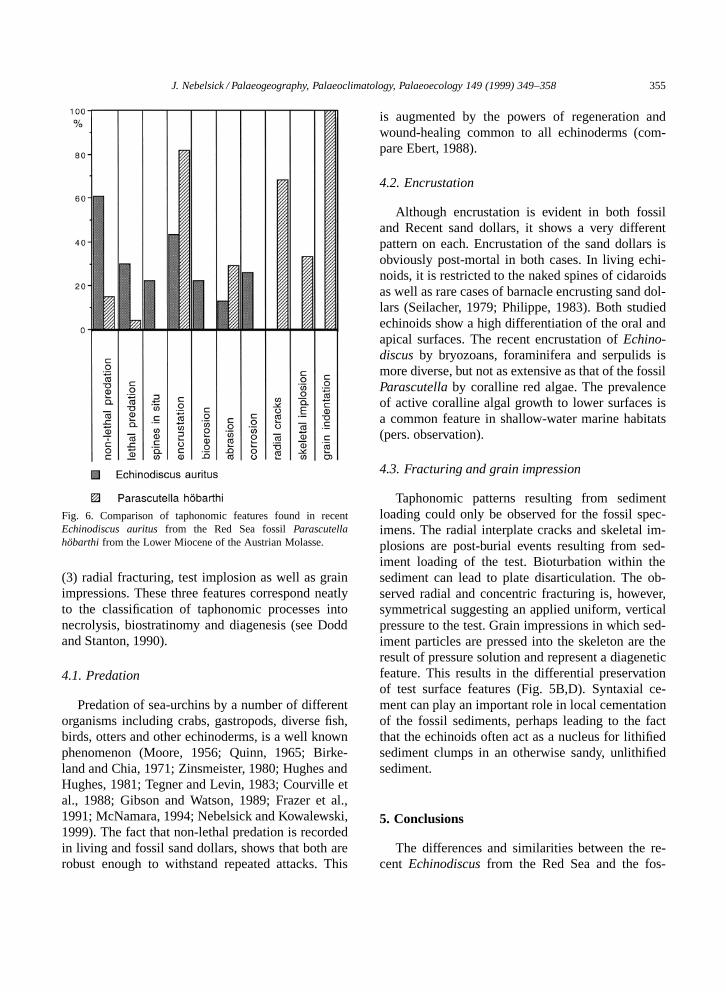
J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358 355
Fig. 6. Comparison of taphonomic features found in recentEchinodiscus auritus from the Red Sea fossil Parascutellahobarthi from the Lower Miocene of the Austrian Molasse.
(3) radial fracturing, test implosion as well as grainimpressions. These three features correspond neatlyto the classification of taphonomic processes intonecrolysis, biostratinomy and diagenesis (see Doddand Stanton, 1990).
4.1. Predation
Predation of sea-urchins by a number of differentorganisms including crabs, gastropods, diverse fish,birds, otters and other echinoderms, is a well knownphenomenon (Moore, 1956; Quinn, 1965; Birke-land and Chia, 1971; Zinsmeister, 1980; Hughes andHughes, 1981; Tegner and Levin, 1983; Courville etal., 1988; Gibson and Watson, 1989; Frazer et al.,1991; McNamara, 1994; Nebelsick and Kowalewski,1999). The fact that non-lethal predation is recordedin living and fossil sand dollars, shows that both arerobust enough to withstand repeated attacks. This
is augmented by the powers of regeneration andwound-healing common to all echinoderms (com-pare Ebert, 1988).
4.2. Encrustation
Although encrustation is evident in both fossiland Recent sand dollars, it shows a very differentpattern on each. Encrustation of the sand dollars isobviously post-mortal in both cases. In living echi-noids, it is restricted to the naked spines of cidaroidsas well as rare cases of barnacle encrusting sand dol-lars (Seilacher, 1979; Philippe, 1983). Both studiedechinoids show a high differentiation of the oral andapical surfaces. The recent encrustation of Echino-discus by bryozoans, foraminifera and serpulids ismore diverse, but not as extensive as that of the fossilParascutella by coralline red algae. The prevalenceof active coralline algal growth to lower surfaces isa common feature in shallow-water marine habitats(pers. observation).
4.3. Fracturing and grain impression
Taphonomic patterns resulting from sedimentloading could only be observed for the fossil spec-imens. The radial interplate cracks and skeletal im-plosions are post-burial events resulting from sed-iment loading of the test. Bioturbation within thesediment can lead to plate disarticulation. The ob-served radial and concentric fracturing is, however,symmetrical suggesting an applied uniform, verticalpressure to the test. Grain impressions in which sed-iment particles are pressed into the skeleton are theresult of pressure solution and represent a diageneticfeature. This results in the differential preservationof test surface features (Fig. 5B,D). Syntaxial ce-ment can play an important role in local cementationof the fossil sediments, perhaps leading to the factthat the echinoids often act as a nucleus for lithifiedsediment clumps in an otherwise sandy, unlithifiedsediment.
5. Conclusions
The differences and similarities between the re-cent Echinodiscus from the Red Sea and the fos-

356 J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358
sil Parascutella from the Miocene can be assessedby comparing constructional morphologies, environ-mental factors and temporal effects related to diage-nesis and sediment loading.
5.1. Constructional morphology
Both tests show similar constructional morpholo-gies. The excellent preservation of both Recent andfossil sand dollars as complete tests is primarily dueto the internal support system which not only con-nects the oral and apical surfaces, but also formsa meshwork connecting the separate plates to eachother. The presence of this system differentially af-fects the studied specimens. It allows the RecentEchinodiscus to be preserved as a ‘complete test’despite being hollowed out by predators; it becomesevident in the fossil Parascutella when sedimentpressure depresses the central area leaving the rimcomplete. Both Recent and fossil sand dollars havebetter preservation potentials than other echinodermswith which they occur.
5.2. Environmental factors
Differences in environmental factors are a moreimportant control of taphonomic patterns than vari-ations related to constructional morphology for thestudied material. The coarser fossil sediment mayrepresent a higher-energy environment which wouldexplain the higher degree of abrasion recorded forthe fossil specimens. The difference in predation pat-terns is another factor related to the environment.The fact that E. auritus originates from a tropicalenvironment with a very wide facies variation in-cluding reefs may enhance the presence of differentpotential predators. Both echinoids offer a similarsubstrate for encrusters as far as general size andsurface morphology is concerned. The differences inencrustation patterns thus also represent differencesin the ambient environment. The importance of deadParascutella tests as a substrate for coralline algae,which would otherwise be offered little other suitablesurfaces, has to be emphasized.
5.3. Time-related effects
Temporal effects affecting the sea-urchins rangefrom short-term time-averaging (see Nebelsick,
1992) to long-term diagenetic effects. A major diffi-culty in comparing these two species is that tapho-nomic processes would still have acted upon the Re-cent Echinodiscus had they not been removed fromthe study area. This may explain the under- or over-representation of certain taphonomic features. Thusthe spines found in situ in the recent echinoids willhave fallen off the test within a few days if left in thesediment. Continued surface residence time will alsoincrease the chance of abrasion and encrustation.This comparison of recent and fossil echinoids alsoshows how post-burial factors, including sedimentloading and grain indentations, can reduce the possi-bility of recognizing important taphonomic featuressuch as small-scale alterations of surface characters.
Acknowledgements
I would like to thank T. Haunold, S. Kampfer, L.Nebelsick, P. Pervesler, W. Piller, R. Roetzel and F.Steininger. This study was supported by the AustrianScience Foundation Projects: P5877 and P7507-Geoand P8869-GEO.
References
Beadle, S.C., 1989. Ontogenetic regulatory mechanisms, hete-rochrony, and eccentricity in dendrasterid sand dollars. Paleo-biology 15, 205–222.
Bell, B.M., Frey, R.W., 1969. Observations on ecology and thefeeding and burrowing mechanisms of Mellita quinquiesperfo-rata. J. Paleontol. 43, 553–560.
Bentley, A.C., Cockcroft, A.C., 1995. Sublittoral sand dollar(Echinodiscus bisperforatus) communities in two bays on theSouth African coast. S.Afr. J. Zool. 30, 5–18.
Birkeland, C., Chia, F.S., 1971. Recruitment risk, growth, ageand predation in two populations of sand dollars, Dendrasterexcentricus (Eschholtz). J. Exp. Mar. Biol. Ecol. 6, 265–278.
Boggild, G.R., Rose, E.P.F., 1984. Mid-Tertiary echinoid biofa-cies as palaeoenvironmental indices. Ann. Geol. Pays Hellen.32, 57–67.
Courville, P., Vincent, E., Thierry, J., David, B., 1988. La barre ascutelles du Burdigalien (Miocene) de Montbrison–Fontbonau(Bassin de Valreas, Vaucluse): du milieu de vie au milieu dedepot. Actes du 6 seminaire international sur les Echinoder-mes, pp. 3–16.
Dodd, J.R., Stanton, R.J., 1990. Paleoecology: Concepts andApplications. New York, 502 pp.
Dodd, J.R., Alexander, R.R., Stanton, R.J., 1985. Populationdynamics in Dendraster, Merriamaster, and Anadara from

J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358 357
the Neogene of the Kettleman Hills, California. Palaeogeogr.,Palaeoclimat., Palaeoecol. 52, 61–76.
Donovan, S.K., 1991. The taphonomy of echinoderms: calcare-ous multi-element skeletons in the marine environment. In:Donovan, S.K. (Ed.), The Processes of Fossilization. BelhavenPress, London, pp. 241–269.
Durham, J.W., 1966. Clypeasteroids. In: Moore, R.C. (Ed.),Treatise on Invertebrate Paleontology. Univ. Kansas Press,Lawrence, KA, pp. U450–U491.
Ebert, T.A., 1988. Growth, regeneration, and damage repair ofspines of the Slate-Pencil sea urchin Heterocentrotus mammil-atus (L.) (Echinodermata: Echinoidae). Pac. Sci. 42, 3–4.
Ebert, T.A., Dexter, D.M., 1975. A natural history study ofEncope grandis and Mellita grantii, two sand dollars in theNorthern Gulf of California, Mexico. Mar. Biol. 32, 397–407.
Frazer, T.K., Lindberg, W.J., Stanton, G.R., 1991. Predation onsand dollars by gray Triggerfish, Balistes capriscus, in thenortheastern gulf of Mexico. Bull. Mar. Sci. 48, 159–164.
Ghiold, J., Hoffman, A., 1984. Clypeasteroid echinoids and his-torical biogeography. Neues Jahrb. Geol. Palaontol. Monatsh.1984, 529–538.
Ghiold, J., Hoffman, A., 1986. Biogeography and biogeographichistory of clypeasteroid echinoids. J. Biogeogr. 13, 183–206.
Gibson, M.A., Watson, J.B., 1989. Predatory and non-preda-tory borings in echinoids from the upper Ocala Formation(Eocene), north-central Florida, U.S.A. Palaeogeogr., Palaeo-climatol., Palaeoecol. 71, 309–321.
Gordon, C.M., Donovan, S.K., 1992. Disarticulated echinoidossicles in paleoecology and taphonomy: the last interglacialFalmouth Formation of Jamaica. Palaios 7, 157–166.
Greenstein, B.J., 1991. An integrated study of echinoid taphon-omy: predictions for the fossil record of four echinoid families.Palaios 6, 519–540.
Greenstein, B.J., 1992. Taphonomic bias and the evolutionaryhistory of the Family Cidaridae (Echinodermata: Echinoidea).Paleobiology 18, 50–79.
Hughes, R.N., Hughes, P.I., 1981. Morphological and behav-ioral aspects of feeding in the Cassidae (Tonnacea, Mesogas-tropoda). Malacol. 20, 385–402.
Kidwell, S.M., Baumiller, T., 1990. Experimental disintegrationof regular echinoids: roles of temperature, oxygen and decaythresholds. Paleobiology 16, 247–271.
Kuhn, O., 1936. Eine neue Burdigalausbildung bei Horn. Sitzber.Osterr. Akad. Wiss. Math.-Naturwiss. Kl. 145, 35–45.
McKinney, M.L., 1985. The abundant occurrence of the Mid-dle Miocene sand dollar Abertella aberti in the HawthorneFormation of Florida. Southeast. Geol. 24, 155–158.
McNamara, K.J., 1994. The significance of gastropod predationto patterns of evolution and extinction in Australian Tertiaryechinoids. In: David, B., Guille, A., Feral, J.-P., Roud, M.(Eds.), Echinoderms through Time. Balkema, Rotterdam, pp.785–793.
Meyer, D.L., Meyer, K.B., 1986. Biostratinomy of recentcrinoids (Echinodermata) at Lizard Island, Great Barrier Reef,Australia. Palaios 1, 294–302.
Mooi, R., 1989. Living and fossil genera of the Clypeasteroida
(Echinoida: Echinodermata): an illustrated key and annotatedchecklist. Smithsonian Contrib. Zool. 448, 1–51.
Mooi, R., 1990. Paedomorphosis, Aristotoles lantern and theorigin of the sand dollars (Echinodermata: Clypeasteroida).Paleobiology 16, 25–48.
Moore, D.R., 1956. Observations on predation on echinodermsby three species of Cassidae. Nautilus 69, 73–76.
Nebelsick, J.H., 1992. The use of fragments in deducing echinoiddistribution by fragment identification in Northern Bay ofSafaga, Red Sea, Egypt. Palaios 7, 316–328.
Nebelsick, J.H., 1995. Actuopaleontological investigations onechinoids: the potential for taphonomic interpretation. In: Em-son, R.H., Smith, A.B., Campbell, A.C. (Eds.), EchinodermResearch. Balkema, Rotterdam, pp. 209–214.
Nebelsick, J.H., 1996. Biodiversity of shallow-water Red Seaechinoids: implications for the fossil record. J. Mar. Biol.Assoc. U.K. 76, 185–194.
Nebelsick, J.H., Kampfer, S., 1994. Taphonomy of Clypeasterhumilis and Echinodiscus auritus from the Red Sea. In: David,B., Guille, A., Feral, J.-P., Roux, M. (Eds.), EchinodermsThrough Time. Balkema, Rotterdam, pp. 803–808.
Nebelsick, J.H., Kowalewski, M., 1999. Drilling predation onRecent clypeasteroid echinoids from the Red Sea. Palaios 14,in press.
Philippe, M., 1983. Deformaton d’une scutelle (Echinoidea,Clypeasteroida) Miocene due a la fixation d’une balane. Hy-phothese Paleoecologique. Geobios 16, 371–374.
Piller, W., Mansour, A.M., 1990. The Northern Bay of Safaga(Red Sea, Egypt): an actuopalaeontological approach, II. Sedi-ment analysis and sedimentary facies. Beitr. Palaontol. Osterr.16, 1–102.
Poddubiuk, R.H., Rose, E.P.F., 1984. Relationships between Mid-Tertiary echinoid faunas from the central Mediterranean andeastern Caribbean and their palaeobiogeographic significance.Ann. Geol. Pays Hellen. 31, 115–127.
Quinn, B.G., 1965. Predation in sea urchins. Bull. Mar. Sci. 15,259–264.
Schaffer, H., 1962. Die Scutelliden des Miozans von Osterreichund Ungarn. Palaontol. Z. 36, 135–170.
Seilacher, A., 1979. Constructional morphology of sand dollars.Paleobiology 5, 191–221.
Smith, A.B., 1984. Echinoid Palaeobiology. George Allen andUnwin, London, 191 pp.
Smith, A.L., 1981. Comparison of macrofaunal invertebrates insand dollar (Dendraster excentricus) beds and in adjacentareas free of sand dollars. Mar. Biol. 65, 191–198.
Stanley, D.J., James, P.J., 1971. Distribution of Echinarachniusparma (Lamarck) and associated fauna on Sable Island Bank,Southeast Canada. Smithsonian Contrib. Earth Sci. 6, 1–24.
Steininger, F., 1971. Die Echinodermen der EggengurgerSchichtengruppe. In: Steininger, F.F., Senes, J. (Eds.), M1Eggenburgien, Ser. Chronostratigraphie und Neostratotypen.SAV, Bratislava, pp. 593–597.
Steininger, F., Roetzel, R., 1991. Geologische Grundlagen,Lithostratigraphie, Biostratigraphie und chronostratigraphischeKorrelation der Molasesedimente am Ostrand der BohmischenMasse. Arbeitstag. Geol. Bundesanst. 1, 102–108.

358 J. Nebelsick / Palaeogeography, Palaeoclimatology, Palaeoecology 149 (1999) 349–358
Tegner, M.J., Levin, L.A., 1983. Spiny lobsters and sea-urchins.Analysis of a predator–prey interaction. J. Exp. Mar. Biol.Ecol. 73, 125–150.
Timko, P.L., 1976. Sand dollars as suspension-feeders: a newdescription of feeding in Dendraster excentricus. Biol. Bull.151, 247–259.
Wagner, C.D., 1974. Fossil and Recent sand dollar echinoids ofAlaska. J. Palaeontol. 48, 105–123.
Zinsmeister, W.J., 1980. Observations on the predation of theclypeasteroid echinoid, Monophoraster darwini from the Up-per Miocene Enterrios Formation, Patagonia, Argentina. J.Paleontol. 54, 910–912.
Related Documents











