Physiologia Plantarum 138: 10–21. 2010 Copyright © Physiologia Plantarum 2009, ISSN 0031-9317 Systemin-dependent salinity tolerance in tomato: evidence of specific convergence of abiotic and biotic stress responses Francesco Orsini a,† , Pasquale Cascone b , Stefania De Pascale a , Giancarlo Barbieri a , Giandomenico Corrado b , Rosa Rao b and Albino Maggio a,∗ a Department of Agricultural Engineering and Agronomy, University of Naples Federico II, Via Universit ` a 100, 80055 Portici, Italy b Department of Soil, Plant and Environmental Science, University of Naples Federico II, Via Universit ` a 100, 80055 Portici, Italy Correspondence *Corresponding author, e-mail: [email protected] Received 7 July 2009 doi:10.1111/j.1399-3054.2009.01292.x Plants have evolved complex mechanisms to perceive environmental cues and develop appropriate and coordinated responses to abiotic and biotic stresses. Considerable progress has been made towards a better under- standing of the molecular mechanisms of plant response to a single stress. However, the existence of cross-tolerance to different stressors has proved to have great relevance in the control and regulation of organismal adaptation. Evidence for the involvement of the signal peptide systemin and jasmonic acid in wound-induced salt stress adaptation in tomato has been provided. To further unravel the functional link between plant responses to salt stress and mechanical damage, transgenic tomato (Lycopersicon esculentum Mill.) plants constitutively expressing the prosystemin cDNA have been exposed to a moderate salt stress. Prosystemin over-expression caused a reduction in stomatal conductance. However, in response to salt stress, prosystemin transgenic plants maintained a higher stomatal conductance compared with the wild-type control. Leaf concentrations of abscissic acid (ABA) and proline were lower in stressed transgenic plants compared with their wild-type con- trol, implying that either the former perceived a less stressful environment or they adapted more efficiently to it. Consistently, under salt stress, transgenic plants produced a higher biomass, indicating that a constitutive activation of wound responses is advantageous in saline environment. Comparative gene expression profiling of stress-induced genes suggested that the partial stomatal closure was not mediated by ABA and/or components of the ABA signal transduction pathway. Possible cross-talks between genes involved in wounding and osmotic stress adaptation pathways in tomato are discussed. Introduction The interaction among abiotic stress, herbivore and pathogen attacks in plants is subject to intense research Abbreviations – ABA, abscissic acid; ASA, ascorbate; BB, non-transgenic control cv. Better Boy; BBS, Better Boy plants over-expressing prosystemin cDNA; DW, dry weight; EC, electrical conductivity; ET, Ethylene; ISCI, Istituto Sperimentale per le Colture Industriali; JA, jasmonic acid; LSD, least significant difference; OA, osmotic adjustment; PCR, polymerase chain reaction; PR, pathogenesis-related; ROS, reactive oxygen species; RWC, relative water content. † Present address: Department of Agro-Environmental Sciences and Technologies, University of Bologna, Viale Fanin 44, 40127 Bologna, Italy because the hormone-mediated signalling network, which links biotic and abiotic stress responses, is essential for plant adaptation (Cheong et al. 2002). Early evidence for a cross-talk between abiotic and biotic stress 10 Physiol. Plant. 138, 2010

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Physiologia Plantarum 138: 10–21. 2010 Copyright © Physiologia Plantarum 2009, ISSN 0031-9317
Systemin-dependent salinity tolerance in tomato: evidenceof specific convergence of abiotic and biotic stress responsesFrancesco Orsinia,†, Pasquale Casconeb, Stefania De Pascalea, Giancarlo Barbieria, GiandomenicoCorradob, Rosa Raob and Albino Maggioa,∗
aDepartment of Agricultural Engineering and Agronomy, University of Naples Federico II, Via Universita 100, 80055 Portici, ItalybDepartment of Soil, Plant and Environmental Science, University of Naples Federico II, Via Universita 100, 80055 Portici, Italy
Correspondence*Corresponding author,e-mail: [email protected]
Received 7 July 2009
doi:10.1111/j.1399-3054.2009.01292.x
Plants have evolved complex mechanisms to perceive environmental cuesand develop appropriate and coordinated responses to abiotic and bioticstresses. Considerable progress has been made towards a better under-standing of the molecular mechanisms of plant response to a single stress.However, the existence of cross-tolerance to different stressors has proved tohave great relevance in the control and regulation of organismal adaptation.Evidence for the involvement of the signal peptide systemin and jasmonicacid in wound-induced salt stress adaptation in tomato has been provided.To further unravel the functional link between plant responses to salt stressand mechanical damage, transgenic tomato (Lycopersicon esculentum Mill.)plants constitutively expressing the prosystemin cDNA have been exposedto a moderate salt stress. Prosystemin over-expression caused a reductionin stomatal conductance. However, in response to salt stress, prosystemintransgenic plants maintained a higher stomatal conductance compared withthe wild-type control. Leaf concentrations of abscissic acid (ABA) and prolinewere lower in stressed transgenic plants compared with their wild-type con-trol, implying that either the former perceived a less stressful environment orthey adapted more efficiently to it. Consistently, under salt stress, transgenicplants produced a higher biomass, indicating that a constitutive activationof wound responses is advantageous in saline environment. Comparativegene expression profiling of stress-induced genes suggested that the partialstomatal closure was not mediated by ABA and/or components of the ABAsignal transduction pathway. Possible cross-talks between genes involved inwounding and osmotic stress adaptation pathways in tomato are discussed.
Introduction
The interaction among abiotic stress, herbivore andpathogen attacks in plants is subject to intense research
Abbreviations – ABA, abscissic acid; ASA, ascorbate; BB, non-transgenic control cv. Better Boy; BBS, Better Boy plantsover-expressing prosystemin cDNA; DW, dry weight; EC, electrical conductivity; ET, Ethylene; ISCI, Istituto Sperimentale perle Colture Industriali; JA, jasmonic acid; LSD, least significant difference; OA, osmotic adjustment; PCR, polymerase chainreaction; PR, pathogenesis-related; ROS, reactive oxygen species; RWC, relative water content.
†Present address: Department of Agro-Environmental Sciences and Technologies, University of Bologna, Viale Fanin 44,40127 Bologna, Italy
because the hormone-mediated signalling network,which links biotic and abiotic stress responses, isessential for plant adaptation (Cheong et al. 2002). Earlyevidence for a cross-talk between abiotic and biotic stress
10 Physiol. Plant. 138, 2010

responses was provided by Bressan et al. (1982), whodemonstrated that pathogenesis-related (PR) proteinsaccumulate in tomato cells exposed to salt stress and thateffector proteins involved in one of the tolerances haveprofitable characteristics for the other tolerances also.Subsequently, it was proved that ethylene and methyljasmonate synergistically induce members of the PRsuperfamily of defence genes (Xu et al. 1994). Currently,there is a general agreement on the presence of a linkbetween hormonal control and downstream activationof both osmotic and PR metabolic compounds.
Herbivore feeding is perhaps the most commonphysical damage that plants face during their life cycle.Insects, and more generally wounding, induce a systemicresponse that is rapidly propagated from the woundedsite to distal portions of the plant (Bergey et al. 1996). Intomato, the response to chewing insects and mechanicalwounding involves the release of a signal moleculecalled systemin (Pearce et al. 1991). This 18-aa peptideactivates defence responses by formation of hydrogenperoxides (Orozco-Cardenas and Ryan 1999), alterationsin ion fluxes across the plasma membrane (Lanfermeijeret al. 2008), phospholipase activation (Narvaez-Vasquezet al. 1999) and the synthesis of jasmonic acid (JA),a key molecule required for the long-distance signalresponse (Li et al. 2002). Most of our knowledge onsystemin-mediated physiological functions relates todirect and indirect plant defence mechanisms againstpests (Corrado et al. 2007, de la Noval et al. 2007,McGurl et al. 1994) and its role in the activationof the JA-pathway (Ryan and Pearce 2003). Becauseleaf piercing and wounding increase dehydration, it isexpected that plants have evolved coordinated responsesto these different stressors. Tomato plants grownin saline environment accumulate insect resistanceproteins (Dombrowski 2003) and similarly, woundedplants are more tolerant to salt stress (Capiati et al.2006). It has been proposed that the activation of broadand/or generic salt tolerance responses in plants attackedby herbivorous insects would present an energeticallyexpensive and probably unsustainable cost. For thisreason, it is not clear whether a wound-mediatedactivation of salt tolerance is a functional redundancy instress-signalling pathways or it is specifically as a result ofreactions that would protect the plant against defoliatinginsects (Dombrowski 2003, Capiati et al. 2006).
Several studies have indicated that stress responsesas well as other fundamental physiological processessuch as seed maturation and flowering are controlledby a concerted action of different signalling pathways.Nonetheless, this complex regulatory network is elicitedby a dynamic balance of a relatively reduced numberof primary endogenous signals (Bostok 2005). Among
these, it is known that abscissic acid (ABA), in additionto other fundamental functions (Wang et al. 2004),regulates adaptation to cold, salt and drought conditions(Maggio et al. 2002b, Verslues and Zhu 2005).
Components of ABA and JA/Ethylene-signalling path-ways are thought to be antagonistic in modulatingdefence and stress-responsive gene expression (Ander-son et al. 2004), but the interplay among differentmetabolic pathways is complex. The ABA-mediated sig-nalling pathway induced by drought stress upregulatesgenes that are involved in the JA-mediated woundingresponse, but negatively regulates JA/Ethylene-mediatedpathogen defence genes (Abe et al. 2003, Andersonet al. 2004, Boter et al. 2004). Overall, current evidenceimplies the existence of some specificity in the activationof coordinated responses in plants.
By using a combination of physiological andgene expression analyses, it is demonstrated thatover-expression of the systemin precursor in tomatogenerates pre-adaptation responses that lead to an ABA-independent salinity tolerance. This was sufficient toguarantee improved plant performances under salt stressconditions.
Materials and methods
Plant material, experimental set-up and stressconditions
Three experiments (A, B and C) were carried out at theUniversity of Napoli Federico II, Agriculture Faculty(40◦ 49′ N, 14◦ 15′ E, 30 m a.s.l.), in 2006 and2007. Seeds of transgenic tomato plants over-expressingthe prosystemin gene under the control of the 35 SRNA cauliflower mosaic virus (CaMV) promoter (BetterBoy plants over-expressing prosystemin cDNA, BBS)and relative non-transgenic control cv. Better Boy (BB)(Lycopersicon esculentum Mill.) were kindly donatedby Prof. C. Ryan (Washington University) (McGurlet al. 1994). The over-expression of the prosystemincDNA in transgenic BBS plants was confirmed byreverse transcriptase polymerase chain reaction (RT-PCR) analysis (see Appendix S1).
Seeds were sown on trays on 27 April 2006, 5 May2006 and 10 June 2007 for experiment A, B and C,respectively. At the stage of two fully expanded trueleaves, plants of uniform size were transferred to hydro-ponic systems (four plants per square meter). Rockwoolslabs (Grodan©) were used for experiment A, while plas-tic pots (diameter 20 cm) filled with perlite were usedfor experiments B and C. Experiments were conductedin a cold greenhouse (minimum inside air temperatureswere 18, 18 and 21◦C for experiment A, B and C, respec-tively; the maximum temperature was 38◦C during sunny
Physiol. Plant. 138, 2010 11

hours in late summer). A closed loop irrigation systemwith complete recycling of nutrient solution was used.Crop water uptake was weekly compensated with freshnutrient solution. The re-circulating nutrient solutionwas automatically pumped by a drip irrigation systemin daily irrigations ranging from four to six accord-ing to crop conditions. A modified Hoagland nutrientsolution (Hoagland and Arnon 1950) was preparedusing ammonium nitrate, calcium nitrate, potassiumsulphate, ammonium phosphate, ethylenediaminete-traacetic acid–iron complex, boric acid, magnesiumsulphate, manganese chloride, zinc sulphate, coppersulphate and molybdic acid; the final composition ofthe nutrient solution was: N = 18.25 mM (N–NO3 =17.00 mM, N–NH4 = 1.25 mM); P2O5 = 1.40 mM;K2O = 8.75 mM; S = 5.00 mM; Ca = 7.00 mM; Mg =3.50 mM; Cl = 10.00 mM; Na = 10.00 mM; Fe =15.00 μM; Cu = 0.75 μM; Zn = 7.00 μM; B = 50.00μM; Mn = 10.00 μM; Mo = 5.00 μM.
The following stress treatments were applied at thestage of four fully expanded leaves:
Experiment A. Tomato plants were irrigated withnutrient solution supplied with 0, 20 or 40 mMNaCl. Thirty-two (16 BB + 16 BBS) plants wereused for each salinity level. The experimental designwas a split-plot (replicated four times) with the threesalinity levels assigned to the main plots, and thetwo genotypes assigned to the sub-plot. The averageelectrical conductivities (EC) and pH of the three nutrientsolutions were 1.8 (pH 6.6), 4.0 (pH 6.7) and 5.9 (pH6.9) dS m−1 for 0, 20 and 40 mM NaCl, respectively.
Experiment B. Tomato plants were irrigated withnutrient solution containing either 0, 20 or 40 mMNaCl. Sixty-four plants (32 BB + 32 BBS) were usedfor each salinity level. The experimental design was asplit-plot (replicated four times) with the three salinitylevels assigned to the main plots, and the two genotypesassigned to the sub-plot. The average EC and pH of thethree nutrient solutions were 1.8, (pH 6.6), 4.0 (pH 6.8)and 6.1 (pH 6.9) dS m−1, for 0, 20 and 40 mM NaCl,respectively.
Experiment C (stress recovery). Sixty-four BB andBBS plants were salinized with a 40 mM NaCl nutrientsolution for 15 days (EC = 6.2 dS m−1). Subsequently,32 plants per genotype were irrigated with the standardnutrient solution (0 mM NaCl), while the remainingplants were maintained at a 40 mM NaCl irrigationregime. The experimental design was a randomizedblock, with four replications. The average EC and pH ofthe nutrient solutions were 1.9 (pH 6.6) and 6.3 (pH 6.9)dS m−1 for 0 and 40 mM NaCl, respectively.
Plant growth, physiological parameters and stressmetabolites accumulation
A week after the beginning of the salt treatments, thestomatal conductance was measured at noon on theabaxial surface of 16 of the youngest fully expandedleaves on four plants per treatment with a diffusionporometer (AP-4, Delta-T Devices, Cambridge, UK).On the same leaves, the total leaf water potentials(�t) were determined with a dew-point psychrome-ter (WP4, Decagon Devices, Washington, WA). Theosmotic potential (�π ) was estimated on frozen/thawedleaf samples and the pressure potential (�p) as the dif-ference between �t and �π , assuming a matrix potentialequal to 0. Leaf osmotic adjustment (OA) was deter-mined as the difference �πo Vo - �π V, where �πo
Vo is the product of (osmotic potential) × (osmotic vol-ume) of unstressed plants and �π V is the product of(osmotic potential) × (osmotic volume) of leaves fromsalinized plants. For each measurement, the osmoticvolume was approximated by the corresponding relativewater content (RWC) value calculated as: RWC = (leaffresh weight – leaf dry weight) (leaf saturated weight– leaf dry weight)−1. Leaf saturated weight was deter-mined after leaf immersion in distilled water for 24 h.ABA determinations were performed on crude extractsfrom 2 g of the youngest fully expanded leaves harvestedon three plants per genotype using an immunoassay kit(Hormondetek-Istituto Sperimentale per le Colture Indus-triali Research Institute for Industrial Crops, Bologna,Italy) (Quarrie et al. 1988). The leaf proline content wasdetermined according to Claussen (2005) on the sameplants used for ABA determinations.
Harvest was carried out from 4 to 24 July 2006, 12 to24 July 2006, and 5 to 25 August 2007 for experimentsA, B, and C, respectively. Fully ripe fruit yield wasquantified over five trusses per plant. Subsequently,plants were collected for biometric determinations. Atthe end of the experiments, plant leaf area was measuredusing a Li-Cor 3000 area meter (Li-Cor, Lincoln, NE) onthree plants per replicate. Fresh and dry (60◦C) weightswere measured separately on plants and fruits.
Results were analyzed by ANOVA and the Fisher’sleast significant differences (LSD) method was used forcomparing treatment group means at a significance levelof P ≤ 0.05. Fisher’s LSD (0.05) was used for compar-ing genotypes and salt stress treatments. Interaction LSD(0.05), when significant, was used to compare meansfrom any two genotypes within each salt stress treatment.
Molecular analysis
Total RNA isolation from leaves and first-strand cDNAsynthesis were performed according to previously
12 Physiol. Plant. 138, 2010
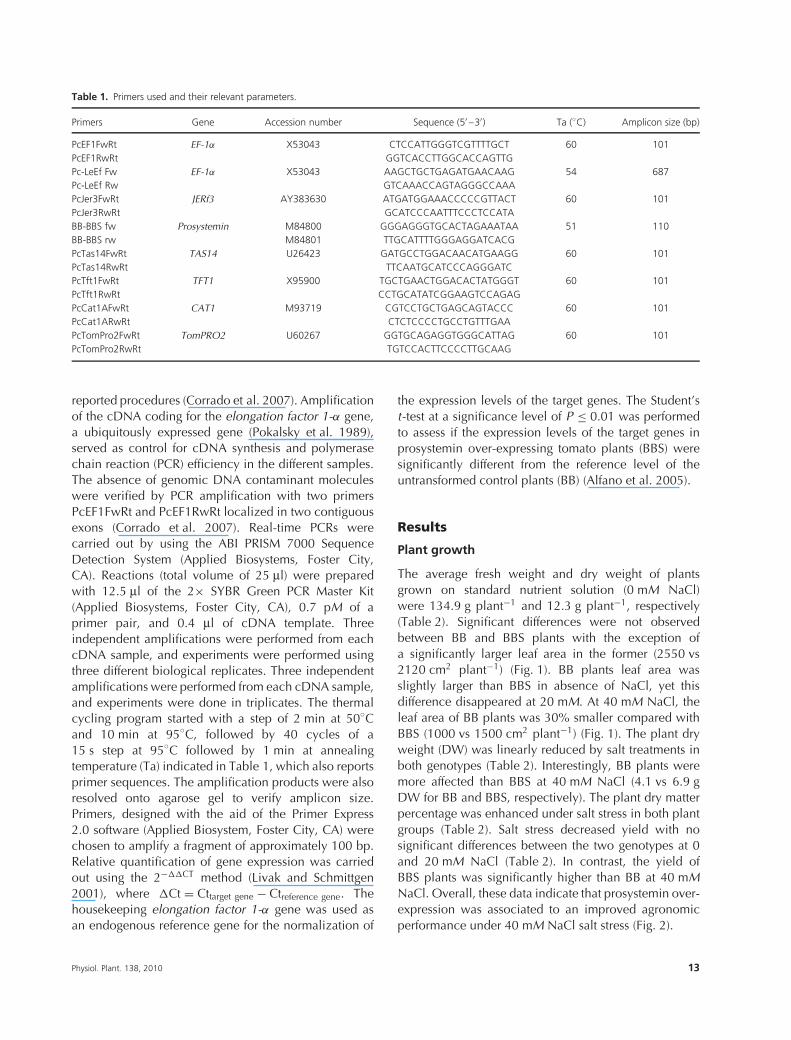
Table 1. Primers used and their relevant parameters.
Primers Gene Accession number Sequence (5′ –3′) Ta (◦C) Amplicon size (bp)
PcEF1FwRt EF-1α X53043 CTCCATTGGGTCGTTTTGCT 60 101PcEF1RwRt GGTCACCTTGGCACCAGTTGPc-LeEf Fw EF-1α X53043 AAGCTGCTGAGATGAACAAG 54 687Pc-LeEf Rw GTCAAACCAGTAGGGCCAAAPcJer3FwRt JERf3 AY383630 ATGATGGAAACCCCCGTTACT 60 101PcJer3RwRt GCATCCCAATTTCCCTCCATABB-BBS fw Prosystemin M84800 GGGAGGGTGCACTAGAAATAA 51 110BB-BBS rw M84801 TTGCATTTTGGGAGGATCACGPcTas14FwRt TAS14 U26423 GATGCCTGGACAACATGAAGG 60 101PcTas14RwRt TTCAATGCATCCCAGGGATCPcTft1FwRt TFT1 X95900 TGCTGAACTGGACACTATGGGT 60 101PcTft1RwRt CCTGCATATCGGAAGTCCAGAGPcCat1AFwRt CAT1 M93719 CGTCCTGCTGAGCAGTACCC 60 101PcCat1ARwRt CTCTCCCCTGCCTGTTTGAAPcTomPro2FwRt TomPRO2 U60267 GGTGCAGAGGTGGGCATTAG 60 101PcTomPro2RwRt TGTCCACTTCCCCTTGCAAG
reported procedures (Corrado et al. 2007). Amplificationof the cDNA coding for the elongation factor 1-α gene,a ubiquitously expressed gene (Pokalsky et al. 1989),served as control for cDNA synthesis and polymerasechain reaction (PCR) efficiency in the different samples.The absence of genomic DNA contaminant moleculeswere verified by PCR amplification with two primersPcEF1FwRt and PcEF1RwRt localized in two contiguousexons (Corrado et al. 2007). Real-time PCRs werecarried out by using the ABI PRISM 7000 SequenceDetection System (Applied Biosystems, Foster City,CA). Reactions (total volume of 25 μl) were preparedwith 12.5 μl of the 2× SYBR Green PCR Master Kit(Applied Biosystems, Foster City, CA), 0.7 pM of aprimer pair, and 0.4 μl of cDNA template. Threeindependent amplifications were performed from eachcDNA sample, and experiments were performed usingthree different biological replicates. Three independentamplifications were performed from each cDNA sample,and experiments were done in triplicates. The thermalcycling program started with a step of 2 min at 50◦Cand 10 min at 95◦C, followed by 40 cycles of a15 s step at 95◦C followed by 1 min at annealingtemperature (Ta) indicated in Table 1, which also reportsprimer sequences. The amplification products were alsoresolved onto agarose gel to verify amplicon size.Primers, designed with the aid of the Primer Express2.0 software (Applied Biosystem, Foster City, CA) werechosen to amplify a fragment of approximately 100 bp.Relative quantification of gene expression was carriedout using the 2−��CT method (Livak and Schmittgen2001), where �Ct = Cttarget gene − Ctreference gene. Thehousekeeping elongation factor 1-α gene was used asan endogenous reference gene for the normalization of
the expression levels of the target genes. The Student’st-test at a significance level of P ≤ 0.01 was performedto assess if the expression levels of the target genes inprosystemin over-expressing tomato plants (BBS) weresignificantly different from the reference level of theuntransformed control plants (BB) (Alfano et al. 2005).
Results
Plant growth
The average fresh weight and dry weight of plantsgrown on standard nutrient solution (0 mM NaCl)were 134.9 g plant−1 and 12.3 g plant−1, respectively(Table 2). Significant differences were not observedbetween BB and BBS plants with the exception ofa significantly larger leaf area in the former (2550 vs2120 cm2 plant−1) (Fig. 1). BB plants leaf area wasslightly larger than BBS in absence of NaCl, yet thisdifference disappeared at 20 mM. At 40 mM NaCl, theleaf area of BB plants was 30% smaller compared withBBS (1000 vs 1500 cm2 plant−1) (Fig. 1). The plant dryweight (DW) was linearly reduced by salt treatments inboth genotypes (Table 2). Interestingly, BB plants weremore affected than BBS at 40 mM NaCl (4.1 vs 6.9 gDW for BB and BBS, respectively). The plant dry matterpercentage was enhanced under salt stress in both plantgroups (Table 2). Salt stress decreased yield with nosignificant differences between the two genotypes at 0and 20 mM NaCl (Table 2). In contrast, the yield ofBBS plants was significantly higher than BB at 40 mMNaCl. Overall, these data indicate that prosystemin over-expression was associated to an improved agronomicperformance under 40 mM NaCl salt stress (Fig. 2).
Physiol. Plant. 138, 2010 13
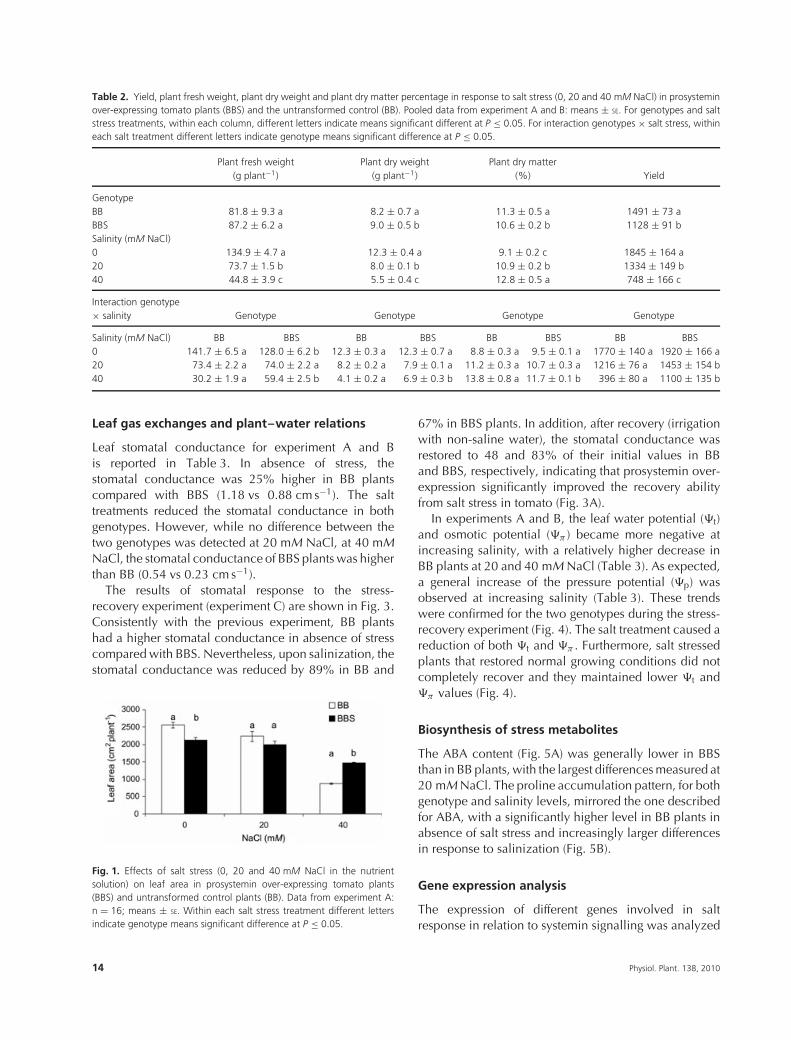
Table 2. Yield, plant fresh weight, plant dry weight and plant dry matter percentage in response to salt stress (0, 20 and 40 mM NaCl) in prosysteminover-expressing tomato plants (BBS) and the untransformed control (BB). Pooled data from experiment A and B: means ± SE. For genotypes and saltstress treatments, within each column, different letters indicate means significant different at P ≤ 0.05. For interaction genotypes × salt stress, withineach salt treatment different letters indicate genotype means significant difference at P ≤ 0.05.
Plant fresh weight Plant dry weight Plant dry matter(g plant−1) (g plant−1) (%) Yield
GenotypeBB 81.8 ± 9.3 a 8.2 ± 0.7 a 11.3 ± 0.5 a 1491 ± 73 aBBS 87.2 ± 6.2 a 9.0 ± 0.5 b 10.6 ± 0.2 b 1128 ± 91 bSalinity (mM NaCl)0 134.9 ± 4.7 a 12.3 ± 0.4 a 9.1 ± 0.2 c 1845 ± 164 a20 73.7 ± 1.5 b 8.0 ± 0.1 b 10.9 ± 0.2 b 1334 ± 149 b40 44.8 ± 3.9 c 5.5 ± 0.4 c 12.8 ± 0.5 a 748 ± 166 c
Interaction genotype× salinity Genotype Genotype Genotype Genotype
Salinity (mM NaCl) BB BBS BB BBS BB BBS BB BBS0 141.7 ± 6.5 a 128.0 ± 6.2 b 12.3 ± 0.3 a 12.3 ± 0.7 a 8.8 ± 0.3 a 9.5 ± 0.1 a 1770 ± 140 a 1920 ± 166 a20 73.4 ± 2.2 a 74.0 ± 2.2 a 8.2 ± 0.2 a 7.9 ± 0.1 a 11.2 ± 0.3 a 10.7 ± 0.3 a 1216 ± 76 a 1453 ± 154 b40 30.2 ± 1.9 a 59.4 ± 2.5 b 4.1 ± 0.2 a 6.9 ± 0.3 b 13.8 ± 0.8 a 11.7 ± 0.1 b 396 ± 80 a 1100 ± 135 b
Leaf gas exchanges and plant–water relations
Leaf stomatal conductance for experiment A and Bis reported in Table 3. In absence of stress, thestomatal conductance was 25% higher in BB plantscompared with BBS (1.18 vs 0.88 cm s−1). The salttreatments reduced the stomatal conductance in bothgenotypes. However, while no difference between thetwo genotypes was detected at 20 mM NaCl, at 40 mMNaCl, the stomatal conductance of BBS plants was higherthan BB (0.54 vs 0.23 cm s−1).
The results of stomatal response to the stress-recovery experiment (experiment C) are shown in Fig. 3.Consistently with the previous experiment, BB plantshad a higher stomatal conductance in absence of stresscompared with BBS. Nevertheless, upon salinization, thestomatal conductance was reduced by 89% in BB and
Fig. 1. Effects of salt stress (0, 20 and 40 mM NaCl in the nutrientsolution) on leaf area in prosystemin over-expressing tomato plants(BBS) and untransformed control plants (BB). Data from experiment A:n = 16; means ± SE. Within each salt stress treatment different lettersindicate genotype means significant difference at P ≤ 0.05.
67% in BBS plants. In addition, after recovery (irrigationwith non-saline water), the stomatal conductance wasrestored to 48 and 83% of their initial values in BBand BBS, respectively, indicating that prosystemin over-expression significantly improved the recovery abilityfrom salt stress in tomato (Fig. 3A).
In experiments A and B, the leaf water potential (�t)and osmotic potential (�π ) became more negative atincreasing salinity, with a relatively higher decrease inBB plants at 20 and 40 mM NaCl (Table 3). As expected,a general increase of the pressure potential (�p) wasobserved at increasing salinity (Table 3). These trendswere confirmed for the two genotypes during the stress-recovery experiment (Fig. 4). The salt treatment caused areduction of both �t and �π . Furthermore, salt stressedplants that restored normal growing conditions did notcompletely recover and they maintained lower �t and�π values (Fig. 4).
Biosynthesis of stress metabolites
The ABA content (Fig. 5A) was generally lower in BBSthan in BB plants, with the largest differences measured at20 mM NaCl. The proline accumulation pattern, for bothgenotype and salinity levels, mirrored the one describedfor ABA, with a significantly higher level in BB plants inabsence of salt stress and increasingly larger differencesin response to salinization (Fig. 5B).
Gene expression analysis
The expression of different genes involved in saltresponse in relation to systemin signalling was analyzed
14 Physiol. Plant. 138, 2010

BB BBS
Control
BB BBS
NaCl (40 mM)
Fig. 2. Prosystemin over-expressing tomato plants (BBS) and untransformed control plants (BB) irrigated with nutrient solution containing 0 (control)or 40 mM NaCl. Pictures refer to experiment B and were taken 20 days after salinization. White arrows indicated advanced wilting of older leaves inBB plants, which was absent in BBS plants.
Table 3. Total leaf water potential (�t), leaf osmotic potential (�π ), pressure potential (�p) and stomatal conductance (gs) in response to salt stress(0, 20 and 40 mM NaCl) in prosystemin over-expressing tomato plants (BBS) and the untransformed control (BB). Pooled data from experiment A andB: means ± SE. For genotypes and salt stress treatments, within each column, different letters indicate means significant difference at P ≤ 0.05. Forinteraction genotypes × salt stress, within each salt treatment different letters indicate genotype means significant difference at P ≤ 0.05.
gs �t �π �p
(cm s−1) (MPa) (MPa) (MPa)
GenotypeBB 0.77 ± 0.06 a −1.05 ± 0.08 a −2.01 ± 0.13 a 0.96 ± 0.06 aBBS 0.76 ± 0.03 a −0.76 ± 0.05 b −1.60 ± 0.08 b 0.84 ± 0.05 bSalinity
(mM NaCl)0 1.03 ± 0.03 a −047 ± 0.04 a −1.14 ± 0.06 a 0.68 ± 0.03 c20 0.87 ± 0.01 b −0.92 ± 0.05 b −1.82 ± 0.10 b 0.90 ± 0.07 b40 0.38 ± 0.03 c −1.33 ± 0.06 c −2.45 ± 0.10 c 1.12 ± 0.06 a
Interaction Genotype Genotype Genotype Genotypegenotype× salinity
Salinity BB BBS BB BBS BB BBS BB BBS(mM NaCl)
0 1.18 ± 0.02 a 0.88 ± 0.02 b −0.46 ± 0.09 a −0.47 ± 0.02 a −1.12 ± 0.12 a −1.17 ± 0.04 a 0.66 ± 0.04 a 0.70 ± 0.04 a20 0.89 ± 0.01 a 0.86 ± 0.02 a −1.10 ± 0.05 a −0.74 ± 0.03 b −2.15 ± 0.14 a −1.49 ± 0.06 b 1.06 ± 0.10 a 0.75 ± 0.06 b40 0.23 ± 0.02 a 0.54 ± 0.02 b −1.58 ± 0.04 a −1.08 ± 0.04 b −2.75 ± 0.10 a −2.15 ± 0.11 b 1.17 ± 0.09 a 1.07 ± 0.09 b
Physiol. Plant. 138, 2010 15
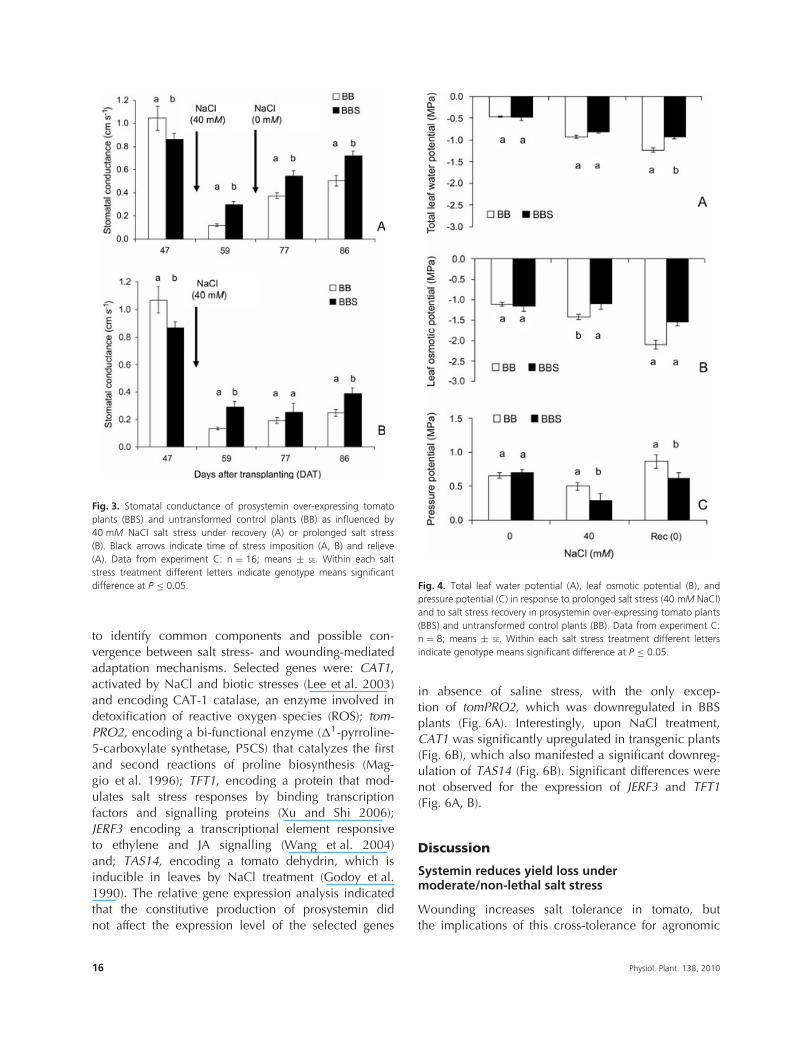
Fig. 3. Stomatal conductance of prosystemin over-expressing tomatoplants (BBS) and untransformed control plants (BB) as influenced by40 mM NaCl salt stress under recovery (A) or prolonged salt stress(B). Black arrows indicate time of stress imposition (A, B) and relieve(A). Data from experiment C: n = 16; means ± SE. Within each saltstress treatment different letters indicate genotype means significantdifference at P ≤ 0.05.
to identify common components and possible con-vergence between salt stress- and wounding-mediatedadaptation mechanisms. Selected genes were: CAT1,activated by NaCl and biotic stresses (Lee et al. 2003)and encoding CAT-1 catalase, an enzyme involved indetoxification of reactive oxygen species (ROS); tom-PRO2, encoding a bi-functional enzyme (�1-pyrroline-5-carboxylate synthetase, P5CS) that catalyzes the firstand second reactions of proline biosynthesis (Mag-gio et al. 1996); TFT1, encoding a protein that mod-ulates salt stress responses by binding transcriptionfactors and signalling proteins (Xu and Shi 2006);JERF3 encoding a transcriptional element responsiveto ethylene and JA signalling (Wang et al. 2004)and; TAS14, encoding a tomato dehydrin, which isinducible in leaves by NaCl treatment (Godoy et al.1990). The relative gene expression analysis indicatedthat the constitutive production of prosystemin didnot affect the expression level of the selected genes
Fig. 4. Total leaf water potential (A), leaf osmotic potential (B), andpressure potential (C) in response to prolonged salt stress (40 mM NaCl)and to salt stress recovery in prosystemin over-expressing tomato plants(BBS) and untransformed control plants (BB). Data from experiment C:n = 8; means ± SE. Within each salt stress treatment different lettersindicate genotype means significant difference at P ≤ 0.05.
in absence of saline stress, with the only excep-tion of tomPRO2, which was downregulated in BBSplants (Fig. 6A). Interestingly, upon NaCl treatment,CAT1 was significantly upregulated in transgenic plants(Fig. 6B), which also manifested a significant downreg-ulation of TAS14 (Fig. 6B). Significant differences werenot observed for the expression of JERF3 and TFT1(Fig. 6A, B).
Discussion
Systemin reduces yield loss undermoderate/non-lethal salt stress
Wounding increases salt tolerance in tomato, butthe implications of this cross-tolerance for agronomic
16 Physiol. Plant. 138, 2010
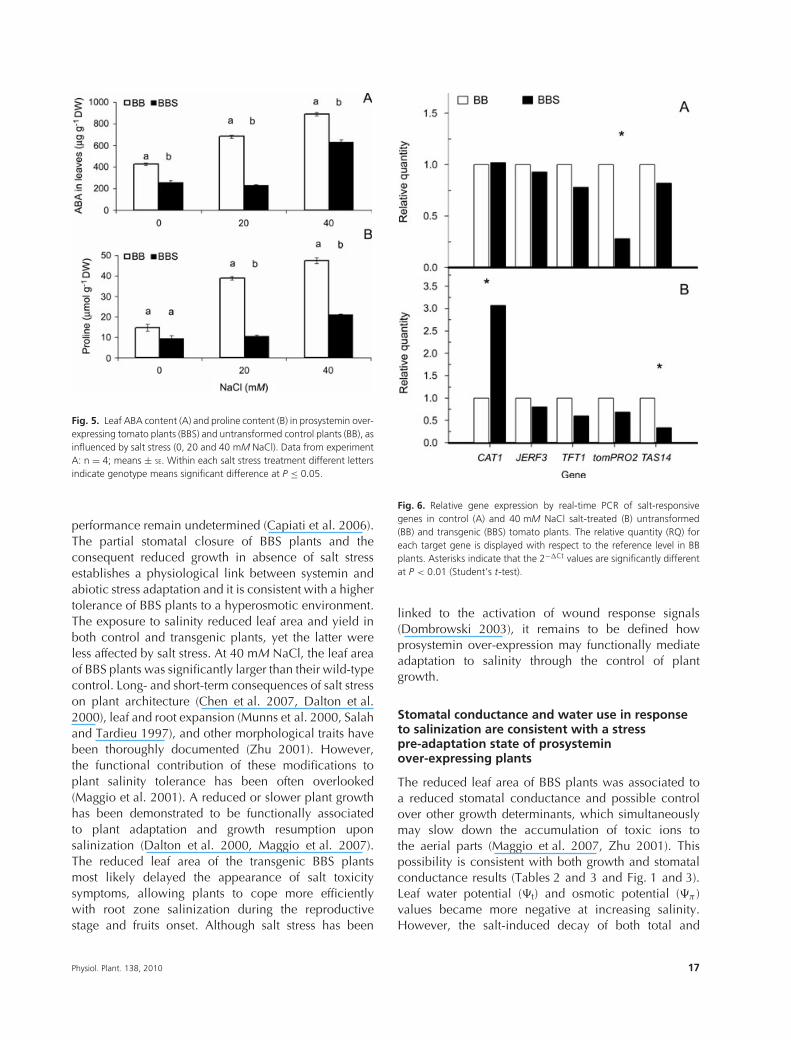
Fig. 5. Leaf ABA content (A) and proline content (B) in prosystemin over-expressing tomato plants (BBS) and untransformed control plants (BB), asinfluenced by salt stress (0, 20 and 40 mM NaCl). Data from experimentA: n = 4; means ± SE. Within each salt stress treatment different lettersindicate genotype means significant difference at P ≤ 0.05.
performance remain undetermined (Capiati et al. 2006).The partial stomatal closure of BBS plants and theconsequent reduced growth in absence of salt stressestablishes a physiological link between systemin andabiotic stress adaptation and it is consistent with a highertolerance of BBS plants to a hyperosmotic environment.The exposure to salinity reduced leaf area and yield inboth control and transgenic plants, yet the latter wereless affected by salt stress. At 40 mM NaCl, the leaf areaof BBS plants was significantly larger than their wild-typecontrol. Long- and short-term consequences of salt stresson plant architecture (Chen et al. 2007, Dalton et al.2000), leaf and root expansion (Munns et al. 2000, Salahand Tardieu 1997), and other morphological traits havebeen thoroughly documented (Zhu 2001). However,the functional contribution of these modifications toplant salinity tolerance has been often overlooked(Maggio et al. 2001). A reduced or slower plant growthhas been demonstrated to be functionally associatedto plant adaptation and growth resumption uponsalinization (Dalton et al. 2000, Maggio et al. 2007).The reduced leaf area of the transgenic BBS plantsmost likely delayed the appearance of salt toxicitysymptoms, allowing plants to cope more efficientlywith root zone salinization during the reproductivestage and fruits onset. Although salt stress has been
Fig. 6. Relative gene expression by real-time PCR of salt-responsivegenes in control (A) and 40 mM NaCl salt-treated (B) untransformed(BB) and transgenic (BBS) tomato plants. The relative quantity (RQ) foreach target gene is displayed with respect to the reference level in BBplants. Asterisks indicate that the 2−�Ct values are significantly differentat P < 0.01 (Student’s t-test).
linked to the activation of wound response signals(Dombrowski 2003), it remains to be defined howprosystemin over-expression may functionally mediateadaptation to salinity through the control of plantgrowth.
Stomatal conductance and water use in responseto salinization are consistent with a stresspre-adaptation state of prosysteminover-expressing plants
The reduced leaf area of BBS plants was associated toa reduced stomatal conductance and possible controlover other growth determinants, which simultaneouslymay slow down the accumulation of toxic ions tothe aerial parts (Maggio et al. 2007, Zhu 2001). Thispossibility is consistent with both growth and stomatalconductance results (Tables 2 and 3 and Fig. 1 and 3).Leaf water potential (�t) and osmotic potential (�π )values became more negative at increasing salinity.However, the salt-induced decay of both total and
Physiol. Plant. 138, 2010 17

osmotic potentials was observed in BBS plants onlyat advanced salinization (Table 3 and Fig. 4). The OAwas larger in BB plants (data not shown) confirmingthat these perceived more severe stressful conditions ascompared with BBS plants (Guerrier 1996). Althoughit cannot be fully excluded that BBS plants may haveactivated other mechanisms to counteract the effectsof salinization, the results obtained indicate that theseplants were less stress sensitive. This is in accordancewith the possibility that the perceived woundingstress, caused by the overproduction of systemin,contributed to a physiological pre-adaptation to otherenvironmental stresses (Hasegawa et al. 2000, Ryan andPearce 2003).
Systemin-dependent reduced stomatalconductance is not correlated to the leaf ABAcontent
The reduced stomatal conductance of BBS plants wasnot associated to an increased leaf ABA level, which wasalways lower in these plants compared with their relativecontrol. The ABA level was consistent with the prolineaccumulation pattern that was also lower in BBS plants.Interestingly, the concentration of these moleculesincreased in BBS plants only at the highest salt dose(Fig. 5). These results were confirmed by the constitutivedownregulation of tomPRO2, which is responsible forproline biosynthesis and is induced by ABA (Fig. 6A).Non-conventional mechanisms of stomatal closureunder salt stress have been documented, although thesehave been less explored than ABA-mediated stomatalresponses (Maggio et al. 2002b, Pei et al. 2000). Ithas been demonstrated that non-osmotically activeconcentrations of ascorbate (ASA) or its precursor L-galactono-1,4-lactone may activate stomatal closurewithout affecting either the ABA or the proline levels(Maggio et al. 2002a). Therefore, ASA-mediated stomatalclosure and ABA-mediated proline accumulation followdifferent signal transduction pathways, with the formerlikely being much more relevant to prosystemin-mediated responses. While the role of ABA in salttolerance is well documented, the association betweensystemin/JA-pathway and ABA is mainly correlative(Capiati et al. 2006). It has then been proposed thathigh cytosolic calcium concentrations, induced bysystemin and/or salt stress (Ryan 2000, Zhu 2002),activate proteins with calmodulin-like activities, suchas LeCDPK1, which are necessary for the onset ofsalt stress tolerance and the expression of insectresistance proteins (Capiati et al. 2006). On the otherhand, it is also possible that a local increase ofABA at the guard cells, mobilized from different
cellular compartments, was responsible for the observedstomatal closure (Tallman 2004). This hypothesis issupported by the observation that systemin activates theROS detoxification machinery (see activation of CAT1in BBS stressed plants; Fig. 6B) and protein kinasesactivity, which have been both associated to Ca2+
signalling in guard cells movements (Cheong et al. 2007,Constabel et al. 1995, Li et al. 1998, Schroeder et al.2001).
Effects of salt stress on gene expression
In absence of saline treatment, and with the exception oftomPRO2, there was no difference in the expressionlevels of CAT1, JERF3, TFT1 and TAS14, indicatingthat the observed salt tolerance of prosystemin over-expressing plants most probably does not occurvia conventional salt stress-induced pathways. Uponsalinization, while the expression of JERF3 and TFT1 didnot reveal differences between BB and BBS, an increasedexpression of CAT1 was observed in BBS plants.Systemin may have likely induced hypersensitivity ofCAT1 to salt stress and may have then contributedto facilitate adaptation, as catalases (and hydrogenperoxide) have extensively been linked to variousplant stress responses, including salinity (Willekenset al. 1997, Xing et al. 2007). Although it has beenreported that ABA, drought and salt stress inducethe expression of CAT1, the gene expression analysisobtained uncouples the ABA- from the salt stress-induced activation of CAT1, because the expressionlevel of this gene was not correlated to an increasedABA level (Xiong et al. 2002). It is possible thatintermediate components of the signal transductionpathway may function to divert proper signals to consentadaptation under different environmental conditions(Xing et al. 2007).
Overall, the results obtained support a model inwhich wounding is associated to a partial stomatalclosure that is not mediated by ABA and/or compo-nents of the ABA signal transduction pathway. Theconstitutive activation of the JA-pathway most likelydetermines a physiological state that directs resourcestowards the production of defensive proteins againstpests, pre-adapts plants to minimize water loss andnegatively regulates the production of hormones andmetabolites (ABA and proline), which will force plantsto invest additional resources to counteract a sec-ondary effect of herbivores. Further analyses are nec-essary to unravel how specificities and commonalitiesbetween different stress-related metabolic pathways areestablished. Nevertheless, it is ecologically and agro-nomically important that the wound-stimulated pre-
18 Physiol. Plant. 138, 2010

adaptation state turned out to be beneficial at advancedsalinization.
References
Abe H, Urao T, Ito T, Seki M, Kazuo Shinozaki K,Yamaguchi-Shinozaki K (2003) Arabidopsis AtMYC2(bHLH) and AtMYB2 (MYB) function as transcriptionalactivators in abscisic acid signaling. Plant Cell 15:63–78
Alfano G, Vitiello C, Caccioppoli C, Caramico T,Carola A, Szego MJ, McInnes RR, Auricchio A, Banfi S(2005) Natural antisense transcripts associated withgenes involved in eye development. Hum Mol Genet14: 913–923
Anderson JP, Badruzsaufari E, Schenk PM, Manners JM,Desmond OJ, Ehlert C, Maclean DJ, Ebert PR, Kazan K(2004) Antagonistic interaction between abscisic acidand jasmonate-ethylene signaling pathways modulatesdefense gene expression and disease resistance inArabidopsis. Plant Cell 16: 3460–3479
Bergey DR, Howe GA, Ryan CA (1996) Polypeptidesignaling for plant defensive genes exhibits analogies todefense signaling in animals. Proc Natl Acad Sci USA93: 12053–12058
Bostok RM (2005) Signal crosstalk and induced resistance:straddling the line between cost and benefit. Annu RevPhytopathol 43: 545–580
Boter, M, Ruız-Rivero, M, Abdeen, A, Prat, S (2004)Conserved MYC transcription factors play a key role injasmonate signaling both in tomato and Arabidopsis.Gene Dev 18: 1577–1591
Bressan RA, Handa AK, Handa S, Hasegawa, PM (1982)Growth and water relations of cultured tomato cells afteradjustment to low external water potentials. PlantPhysiol 70: 1303–1309
Capiati DA, Pais SM, Tellez-Inon MT (2006) Woundingincreases salt tolerance in tomato plants: evidence onthe participation of calmodulin-like activities incross-tolerance signalling. J Exp Bot 57: 2391–2400
Chen DS, Li YG, Zhou JC (2007) The symbiosis phenotypeand expression patterns of five nodule-specific genes ofAstragalus sinicus under ammonium and salt stressconditions. Plant Cell Rep 26: 1421–1430
Cheong YH, Chang HS, Gupta R, Wang X, Zhu T, Luan S(2002) Transcriptional profiling reveals novelinteractions between wounding, pathogen, abioticstress, and hormonal responses in Arabidopsis. PlantPhysiol 129: 661–677
Cheong YH, Pandey GK, Grant JJ, Batistic O, Li L, Kim BG,Lee SC, Kudla J, Luan S (2007) Two calcineurin B-likecalcium sensors, interacting with protein kinase CIPK23,regulate leaf transpiration and root potassium uptake inArabidopsis. Plant J 52: 223–239
Claussen W (2005) Proline as a measure of stress in tomatoplants. Plant Sci 168: 241–248
Constabel CP, Bergey DR, Ryan, CA (1995) Systeminactivates synthesis of wound-inducible tomato leafpolyphenol oxidase via the octadecanoid defensesignaling pathway. Proc Natl Acad Sci USA 92:407–411
Corrado G, Sasso R, Pasquariello M, Iodice L, Carretta A,Cascone P, Ariati L, Digilio M, Guerrieri E, Rao R (2007)Systemin regulates both systemic and volatile signalingin tomato plants. J Chem Ecol 33: 669–681
Dalton FN, Maggio A, Piccinni G (2000) Simulation ofshoot chloride accumulation: separation of physical andbiochemical processes governing plant salt tolerance.Plant Soil 219: 1–11
Dombrowski JE (2003) Salt stress activation ofwound-related genes in tomato plants. Plant Physiol132: 2098–3107
Godoy JA, Pardo JM, Pintor-Toro JA (1990) A tomatocDNA inducible by salt stress and abscisic acid:nucleotide sequence and expression pattern. Plant MolBiol 15: 695–705
Guerrier G (1996) Fluxes of Na+, K+ and Cl−, and osmoticadjustment in Lycopersicon pimpinellifolium andL. esculentum during short-and long-term exposures toNaCl. Physiol Plant 97: 583–591
Hasegawa PM, Bressan RA, Zhu J-K, Bohnert HJ (2000)Plant cellular and molecular responses to highsalinity. Annu Rev Plant Physiol Plant Mol Biol 51:463–499
Hoagland DR, Arnon DI (1950) The Water-CultureMethod for Growing Plants Without Soil. College ofAgriculture, University of California, Berkeley, CA, p 32
Lanfermeijer FC, Staal M, Malinowski R, Stratmann JW,Elzenga JT (2008) Micro-electrode flux estimationconfirms that the Solanum pimpinellifolium cu3 mutantstill responds to systemin. Plant Physiol 146: 129–139
Lee JT, Prasad V, Yang PT, Wu JF, David Ho TH,Charng YY, Chan MT (2003) Expression of ArabidopsisCBF1 regulated by an ABA/stress inducible promoter intransgenic tomato confers stress tolerance withoutaffecting yield. Plant Cell Environ 26: 1181–1190
Li J, Lee IRJ, Assmann SM (1998) Guard cells possess acalcium-dependent protein kinase that phosphorylatesthe KAT1 potassium channel. Plant Physiol 116:785–795
Li L, Li C, Lee GI, Howe GA (2002) Distinct roles forjasmonate synthesis and action in the systemic woundresponse of tomato. Proc Natl Acad Sci USA 99:6416–6421
Livak KJ, Schmittgen TD (2001) Analysis of relative geneexpression data using real-time quantitative PCR and the2(-Delta Delta C(T)) method. Methods 25: 402–408
Physiol. Plant. 138, 2010 19

Maggio A, Garcıa-Rıos M, Fujita T, Bressan R, JolyR, Hasegawa P, Csonka L (1996) The electronic plantgene register. Plant Physiol 112: 862
Maggio A, Hasegawa PM, Bressan RA, Consiglio MF,Joly RJ (2001) Unravelling the functional relationshipbetween root anatomy and stress tolerance. Aust J PlantPhysiol 28: 999–1004
Maggio A, Miyazaki S, Veronese P, Fujita T, Ibeas JI,Damsz B, Narasimhan ML, Hasegawa PM, Joly RJ,Bressan RA 2002a Does proline accumulation play anactive role in stress-induced growth reduction? Plant J31: 699–712
Maggio A, Mc Cully LG, Kerdnaimongkol K, Bressan RA,Hasegawa PM, Joly RJ 2002b The ascorbic acid cyclemediates signal transduction leading to stress-inducedstomatal closure. Funct Plant Biol 29: 845–852
Maggio A, Raimondi G, Martino A, De Pascale S (2007)Salt stress response in tomato beyond the salinitytolerance threshold. Environ Exp Bot 59: 276–282
McGurl B, Orozco-Cardenas M, Pearce G, Ryan CA(1994) Overexpression of the prosystemin gene intransgenic tomato plants generates a systemic signal thatconstitutively induces proteinase inhibitor synthesis.Proc Natl Acad Sci USA 91: 9799–9802
Munns R, Guo J, Passioura JB, Cramer GR (2000) Leafwater status controls day-time but not daily rates of leafexpansion in salt treated plants. Aust J Plant Physiol 27:949–957
Narvaez-Vasquez J, Florin-Christensen J, Ryan CA (1999)Positional specificity of a phospholipase A activityinduced by wounding, systemin, and oligosaccharideelicitors in tomato leaves. Plant Cell 11: 2249–2260
de la Noval B, Perez E, Martınez B, LeonO, Martınez-Gallardo N, Delano-Frier J (2007)Exogenous systemin has a contrasting effect on diseaseresistance in mycorrhizal tomato (Solanumlycopersicum) plants infected with necrotrophic orhemibiotrophic pathogens. Mycorrhiza 17: 449–460
Orozco-Cardenas M, Ryan CA (1999) Hydrogen peroxideis generated systemically in plant leaves by woundingand systemin via the octadecanoid pathway. Proc NatlAcad Sci USA 96: 65536–65557
Pearce G, Strydom D, Johnson S, Ryan CA (1991) Apolypeptide from tomato leaves induceswound-inducible inhibitor proteins. Science 253:895–898
Pei Z-M, Murata Y, Benning G, Thomine S, Klusener B,Allen GJ, Grill E, Schroeder JI (2000) Calcium channelsactivated by hydrogen peroxide mediate abscisic acidsignalling in guard cells. Nature 406: 731–734
Pokalsky AR, Hiatt WR, Ridge N, Rasmussen R,Houck CM, Shewmaker CK (1989) Structure andexpression of elongation factor 1 alpha in tomato.Nucleic Acids Res 17: 4661–4673
Quarrie SA, Whitford PN, Appleford NEJ, Wang TL,Cook SK, Henson IE, Loveyes BR (1988) A monoclonalantibody to (S)-abscissic acid: its characterization anduse in a radioimmunoassay for measuring abscissic acidin crude extracts of cereal and lupin leaves. Planta 173:330–339
Ryan CA (2000) The systemin signaling pathway:differential activation of plant defensive genes. BiochimBiophys Acta 1477: 112–121
Ryan CA, Pearce G (2003) Systemins: a functionallydefined family of peptide signals that regulate defensivegenes in Solanaceae species. Proc Natl Acad Sci USA100: 14577–14580
Salah HBH, Tardieu F (1997) Control of leaf expansionrate of droughted maize plants under fluctuatingevaporative demand: a superposition of hydraulic andchemical messages. Plant Physiol 114: 893–900
Schroeder JI, Kwak JM, Allen GJ (2001) Guard cell abscisicacid signalling and engineering drought hardiness inplants. Nature 410: 327–330
Tallman G (2004) Are diurnal patterns of stomatalmovement the result of alternating metabolism ofendogenous guard cell ABA and accumulation of ABAdelivered to the apoplast around guard cells bytranspiration? J Exp Bot 55: 1963–1976
Verslues PE, Zhu JK (2005) Before and beyond ABA:upstream sensing and internal signals that determineABA accumulation and response under abiotic stress.Biochem Soc Trans 33: 375–379
Wang H, Huang Z, Chen Q, Zhang Z, Zhang H, Wu Y,Huang D, Huang R (2004) Ectopic overexpression oftomato JERF3 in tobacco activates downstream geneexpression and enhances salt tolerance. Plant Mol Biol55: 183–192
Willekens H, Chamnongpol S, Davey M, Schraudner M,Langebartels C, Van Montagu M, Inze D, Van Camp W(1997) Catalase is a sink for H2O2 and is indispensablefor stress defence in C3 plants. EMBO J 16: 4806–4816
Xing Y, Jia W, Zhang J (2007) AtMEK1 mediatesstress-induced gene expression of CAT1 catalase bytriggering H2O2 production in Arabidopsis. J Exp Bot 58:2969–2981
Xiong L, Schumaker KS, Zhu JK (2002) Cell signallingduring cold, drought, and salt stress. Plant Cell 14:165–183
Xu WF, Shi WM (2006) Expression profiling of the 14-3-3gene family in response to salt stress and potassium andiron deficiencies in young tomato (Solanumlycopersicum) roots: analysis by real-time RT–PCR. AnnBot (Lond) 98: 965–974
Xu Y, Chang P, Liu D, Narasimhan ML, Raghothama KG,Hasegawa PM, Bressan RA (1994) Plant defense genesare synergistically induced by ethylene and methyljasmonate. Plant Cell 6: 1077–1085
20 Physiol. Plant. 138, 2010

Zhu, JK (2001) Plant salt tolerance. Trends Plant Sci 6:66–71
Zhu JK (2002) Salt and drought stress signal transduction inplants. Annu Rev Plant Biol 53: 247–273
Supporting Information
Additional Supporting Information may be found in theonline version of this article:
Appendix S1 Prosystemin gene expression in trans-formed plants (BBS) and untransformed control (BB).
Please note: Wiley-Blackwell are not responsible forthe content or functionality of any supporting materialssupplied by the authors. Any queries (other than missingmaterial) should be directed to the corresponding authorfor the article.
Edited by Z. Gong
Physiol. Plant. 138, 2010 21
Related Documents