470 J. Paleont., 82(3), 2008, pp. 470–491 Copyright 2008, The Paleontological Society 0022-3360/08/0082-470$03.00 SYSTEMATIC ANALYSIS OF UPPER TRIASSIC SILICIFIED SCLERACTINIAN CORALS FROM WRANGELLIA AND THE ALEXANDER TERRANE, ALASKA AND BRITISH COLUMBIA ANDREW H. CARUTHERS 1 AND GEORGE D. STANLEY JR. 2 1 Department of Earth and Ocean Sciences, The University of British Columbia, 6339 Stores Road, Vancouver, B.C. V6T 1Z4, Canada, [email protected]; and 2 The University of Montana Center for Paleontology, Missoula 59812, [email protected] ABSTRACT—Acid processing allowed systematic identification of 458 Upper Triassic silicified scleractinian corals (20 genera, 47 species) from the Alexander terrane (southeast Alaska) and Wrangellia (Wrangell Mountains, southern Alaska and Vancouver Island, Canada). Coral faunas, here presented, show taxonomic affinity with coeval collections from other Cordilleran terranes, specifically the Wallowa terrane (northeastern Oregon and Idaho) and Peru (South America) as well as the distant Tethys region. Genera from the Alexander terrane include: Kompsasteria Roniewicz, Gablonzeria Cuif, Cuifia Melnikova, Paracuifia Melnikova, Distichophyllia Cuif, Retiophyllia Cuif, Kuhnastraea Cuif, Margaros- milia Volz, Distichomeandra Cuif, Astraeomorpha Reuss, Pamiroseris Melnikova, Crassistella Roniewicz, Stylophyllum Frech, and Meandros- tylis Frech. Genera from Wrangellia include: Gablonzeria Cuif, Distichophyllia Cuif, Retiophyllia Cuif, Kuhnastraea Cuif, Margarosmilia Volz, Distichomeandra Cuif, Astraeomorpha Reuss, Parastraeomorpha Roniewicz, Chondrocoenia Roniewicz, Pamiroseris Melnikova, Crassistella Roniewicz, Ampakabastraea? Alloiteau, Recticostastraea Stanley and Whalen, Meandrostylis Frech, Anthostylis Roniewicz, and the new genus Campesteria n. gen. New species include: Gablonzeria grandiosa n. sp., Paracuifia smithi n. sp., Paracuifia jennieae n. sp., P. anomala n. sp., Retiophyllia dendriformis n. sp., R. obtusa n. sp., and Campesteria prolixia n. sp. INTRODUCTION S ILICIFIED CORALS were among the earliest Triassic fossils col- lected from the United States. Smith (1912, 1927) recognized Upper Triassic shallow-water marine invertebrate fossils from various localities along western North America. As reviewed by Stanley (1979, p. 4), Smith recognized what he believed to be a continuous coral reef zone fringing the coast of North America. These rocks are now regarded as belonging to a number of tec- tonostratigraphic terranes including well-known Wrangellia and the Alexander terrane (Fig. 1). Stanley (1979) placed Smith’s cor- al and spongiomorph reef occurrences within the emerging con- cepts of exotic or displaced terrane theory, thus explaining the high latitudinal anomalies. Smith’s (1927) systematic description of silicified coral faunas from Gravina Island (now assigned to the Alexander terrane) was updated and combined with coral fau- nas from other Cordilleran terranes (Montanaro-Gallitelli et al., 1979; and this contribution). Squires (1956) then described an- other silicified Upper Triassic site from a limestone quarry near Lewiston, Idaho, at Mission Creek. Being somewhat of an enig- ma, this particular locality has since been loosely tied with the Wallowa terrane (based on overlapping gastropod and coral tax- onomy) and recently dated as late Norian (Nuetzel and Erwin, 2004). Montanaro-Gallitelli et al. (1979) next described a small col- lection of processed silicified corals from the Wrangell Mountains, south-central Alaska, that were among United States Geological Survey (USGS) collections of N. J. Silberling (in Armstrong et al., 1969). Revisiting the USGS site at Green Butte (USGS M1708) in 2004, we collected additional material forming the basis of this contribution (for the purpose of curation, we gave this locality a Montana Invertebrate (MI) locality number, MI 9934). The site represents one the best Upper Triassic silicified shelly invertebrate localities in the Cordilleran region. Clapp and Shimer (1911) originally described and mapped the Upper Triassic rock sequences and rich coral deposits along the southern shore of Lake Cowichan, southern Vancouver Island, Canada; now recognized as the southernmost extension of greater Wrangellia. Clapp and Shimer collected and described a small number of unetched corals, bivalves and mollusks from this site, which they mistakenly identified as Jurassic. Stanley (1989) in- vestigated the paleoecology and interpreted the depositional en- vironments of the Upper Triassic fossils from Lake Cowichan, providing an informal species list, here revised. We further make known a new locality (informally identified by M. J. Orchard) from southern Wrangellia along the Tahsis Inlet (west coast, Van- couver Island), here identified as MI 9913. This study (and subsequent investigations) was aided by silic- ification, revealing intricate details otherwise obscured by calcite recrystallization from original aragonite. Previous work by Clapp and Shimer (1911) and Smith (1927) failed to exploit the oppor- tunities offered by etching the specimens in weak acids. While these studies were instrumental in making known the wide extent of the geographic distribution of Upper Triassic coral faunas in Wrangellia and the Alexander terrane, the total number of coral taxa was too small for statistical comparisons. Recent mapping and biostratigraphic work in Keku Strait (southeast Alaska) by Katvala and Stanley (in press) have uncov- ered a large number of new and well preserved Late Triassic silicified corals and other marine invertebrate fossils from the Al- exander terrane, well dated by conodonts as early Norian. These new findings have greatly expanded our understanding of the pre- served diversity and stratigraphic distribution for this part of the Alexander terrane. By incorporating the Keku Strait coral fauna with that from Gravina Island (Caruthers and Stanley, in press), a much stronger and more meaningful statistical analysis can be utilized to infer associated paleobiogeography (Caruthers, 2005). Caruthers and Stanley (in press) provide detailed biofacies and comparison of depositional environment for Upper Triassic (No- rian to Rhaetian) units within the Alexander terrane and Wran- gellia, also presenting an in-progress species list (Caruthers and Stanley, in press, appendix A) for identified silicified corals of this study. These two works provide a basis for Norian to Rhaetian paleobiogeography of the Alexander terrane and Wran- gellia through statistical analysis of coral species. Analysis con- cludes there is no taxonomic evidence to support an amalgamation in the Upper Triassic, and also suggests a close proximity (high similarity) between coral faunas from southern Wrangellia and Peru (also noted by Prinz, 1991; Prinz-Grimm, 1995 and Stanley, 1994). Paleogeographic results (and associated details for the analysis), paleoecology of the corals, techniques involved in ace- tic acid processing, and their significance in taxonomy can be found in Caruthers (2005) and Caruthers and Stanley (in press, figs. 3, 26). Detailed locality information, lithologic descriptions and geological maps for specimens described in this paper can be found in Caruthers and Stanley (in press, figs. 4, 6, 10, 21, 23). Table 1 presents geographic and geologic breakdown for localities according to terrane or craton. Cuif (1965–1976) initiated a new classification scheme for Up- per Triassic scleractinian corals that has been largely improved

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

470
J. Paleont., 82(3), 2008, pp. 470–491Copyright � 2008, The Paleontological Society0022-3360/08/0082-470$03.00
SYSTEMATIC ANALYSIS OF UPPER TRIASSIC SILICIFIED SCLERACTINIANCORALS FROM WRANGELLIA AND THE ALEXANDER TERRANE,
ALASKA AND BRITISH COLUMBIAANDREW H. CARUTHERS1 AND GEORGE D. STANLEY JR.2
1Department of Earth and Ocean Sciences, The University of British Columbia, 6339 Stores Road, Vancouver, B.C. V6T 1Z4, Canada,�[email protected]�; and 2The University of Montana Center for Paleontology, Missoula 59812, �[email protected]�
ABSTRACT—Acid processing allowed systematic identification of 458 Upper Triassic silicified scleractinian corals (20 genera, 47 species) fromthe Alexander terrane (southeast Alaska) and Wrangellia (Wrangell Mountains, southern Alaska and Vancouver Island, Canada). Coral faunas,here presented, show taxonomic affinity with coeval collections from other Cordilleran terranes, specifically the Wallowa terrane (northeasternOregon and Idaho) and Peru (South America) as well as the distant Tethys region. Genera from the Alexander terrane include: KompsasteriaRoniewicz, Gablonzeria Cuif, Cuifia Melnikova, Paracuifia Melnikova, Distichophyllia Cuif, Retiophyllia Cuif, Kuhnastraea Cuif, Margaros-milia Volz, Distichomeandra Cuif, Astraeomorpha Reuss, Pamiroseris Melnikova, Crassistella Roniewicz, Stylophyllum Frech, and Meandros-tylis Frech. Genera from Wrangellia include: Gablonzeria Cuif, Distichophyllia Cuif, Retiophyllia Cuif, Kuhnastraea Cuif, Margarosmilia Volz,Distichomeandra Cuif, Astraeomorpha Reuss, Parastraeomorpha Roniewicz, Chondrocoenia Roniewicz, Pamiroseris Melnikova, CrassistellaRoniewicz, Ampakabastraea? Alloiteau, Recticostastraea Stanley and Whalen, Meandrostylis Frech, Anthostylis Roniewicz, and the new genusCampesteria n. gen. New species include: Gablonzeria grandiosa n. sp., Paracuifia smithi n. sp., Paracuifia jennieae n. sp., P. anomala n. sp.,Retiophyllia dendriformis n. sp., R. obtusa n. sp., and Campesteria prolixia n. sp.
INTRODUCTION
SILICIFIED CORALS were among the earliest Triassic fossils col-lected from the United States. Smith (1912, 1927) recognized
Upper Triassic shallow-water marine invertebrate fossils fromvarious localities along western North America. As reviewed byStanley (1979, p. 4), Smith recognized what he believed to be acontinuous coral reef zone fringing the coast of North America.These rocks are now regarded as belonging to a number of tec-tonostratigraphic terranes including well-known Wrangellia andthe Alexander terrane (Fig. 1). Stanley (1979) placed Smith’s cor-al and spongiomorph reef occurrences within the emerging con-cepts of exotic or displaced terrane theory, thus explaining thehigh latitudinal anomalies. Smith’s (1927) systematic descriptionof silicified coral faunas from Gravina Island (now assigned tothe Alexander terrane) was updated and combined with coral fau-nas from other Cordilleran terranes (Montanaro-Gallitelli et al.,1979; and this contribution). Squires (1956) then described an-other silicified Upper Triassic site from a limestone quarry nearLewiston, Idaho, at Mission Creek. Being somewhat of an enig-ma, this particular locality has since been loosely tied with theWallowa terrane (based on overlapping gastropod and coral tax-onomy) and recently dated as late Norian (Nuetzel and Erwin,2004).
Montanaro-Gallitelli et al. (1979) next described a small col-lection of processed silicified corals from the Wrangell Mountains,south-central Alaska, that were among United States GeologicalSurvey (USGS) collections of N. J. Silberling (in Armstrong etal., 1969). Revisiting the USGS site at Green Butte (USGSM1708) in 2004, we collected additional material forming thebasis of this contribution (for the purpose of curation, we gavethis locality a Montana Invertebrate (MI) locality number, MI9934). The site represents one the best Upper Triassic silicifiedshelly invertebrate localities in the Cordilleran region.
Clapp and Shimer (1911) originally described and mapped theUpper Triassic rock sequences and rich coral deposits along thesouthern shore of Lake Cowichan, southern Vancouver Island,Canada; now recognized as the southernmost extension of greaterWrangellia. Clapp and Shimer collected and described a smallnumber of unetched corals, bivalves and mollusks from this site,which they mistakenly identified as Jurassic. Stanley (1989) in-vestigated the paleoecology and interpreted the depositional en-vironments of the Upper Triassic fossils from Lake Cowichan,providing an informal species list, here revised. We further makeknown a new locality (informally identified by M. J. Orchard)
from southern Wrangellia along the Tahsis Inlet (west coast, Van-couver Island), here identified as MI 9913.
This study (and subsequent investigations) was aided by silic-ification, revealing intricate details otherwise obscured by calciterecrystallization from original aragonite. Previous work by Clappand Shimer (1911) and Smith (1927) failed to exploit the oppor-tunities offered by etching the specimens in weak acids. Whilethese studies were instrumental in making known the wide extentof the geographic distribution of Upper Triassic coral faunas inWrangellia and the Alexander terrane, the total number of coraltaxa was too small for statistical comparisons.
Recent mapping and biostratigraphic work in Keku Strait(southeast Alaska) by Katvala and Stanley (in press) have uncov-ered a large number of new and well preserved Late Triassicsilicified corals and other marine invertebrate fossils from the Al-exander terrane, well dated by conodonts as early Norian. Thesenew findings have greatly expanded our understanding of the pre-served diversity and stratigraphic distribution for this part of theAlexander terrane. By incorporating the Keku Strait coral faunawith that from Gravina Island (Caruthers and Stanley, in press),a much stronger and more meaningful statistical analysis can beutilized to infer associated paleobiogeography (Caruthers, 2005).
Caruthers and Stanley (in press) provide detailed biofacies andcomparison of depositional environment for Upper Triassic (No-rian to Rhaetian) units within the Alexander terrane and Wran-gellia, also presenting an in-progress species list (Caruthers andStanley, in press, appendix A) for identified silicified corals ofthis study. These two works provide a basis for Norian toRhaetian paleobiogeography of the Alexander terrane and Wran-gellia through statistical analysis of coral species. Analysis con-cludes there is no taxonomic evidence to support an amalgamationin the Upper Triassic, and also suggests a close proximity (highsimilarity) between coral faunas from southern Wrangellia andPeru (also noted by Prinz, 1991; Prinz-Grimm, 1995 and Stanley,1994). Paleogeographic results (and associated details for theanalysis), paleoecology of the corals, techniques involved in ace-tic acid processing, and their significance in taxonomy can befound in Caruthers (2005) and Caruthers and Stanley (in press,figs. 3, 26). Detailed locality information, lithologic descriptionsand geological maps for specimens described in this paper can befound in Caruthers and Stanley (in press, figs. 4, 6, 10, 21, 23).Table 1 presents geographic and geologic breakdown for localitiesaccording to terrane or craton.
Cuif (1965–1976) initiated a new classification scheme for Up-per Triassic scleractinian corals that has been largely improved

471CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
FIGURE 1—Generalized map of western North America showing Wrangelliaand the Alexander terrane with approximate positions of study areas: WrangellMountains, Pender Point, Tahsis Inlet and Lake Cowichan (from Wrangellia),and Keku Strait and Gravina Island (from the Alexander terrane). Figure mod-ified from Jones et al. (1972), Jones et al. (1977), and Katvala and Stanley(in press).
and updated by Beauvais (1980) and Roniewicz (1989). Thisscheme, based largely on preserved microstructural differences inthe original aragonite skeletons of corals, differs from older clas-sification schemes of Reuss (1854, 1865), Frech (1890), and Volz(1896), which involve classification by morphological differences.A scheme based on microstructure is considerably more valid andprecise than those based on morphology, considering inconsisten-cies in coral preservation. However material described in this con-tribution from the Alexander terrane and Wrangellia have beenreplaced by silica and do not present any microstructure, andtherefore must be used as a proxy for the fine quality of aragoniticmaterial from the Alps.
Aside from destroying delicate coralline microstructure, fourmain aspects of the silicification process also are responsible foraltering the preserved morphology and should be taken into ac-count when reviewing the following identifications. These include(1) destroying detailed corallite walls, (2) lumping together thinlybladed septa and destroying their delicate arrangement, (3) pro-ducing an entirely diagenetic pseudo-columella which mimics es-sential characteristics of other unrelated species, and (4) obliter-ating minute ornamentation along septal surfaces (Caruthers andStanley, in press).
SYSTEMATIC PALEONTOLOGY
All described specimens are silicified and were isolated bytreatment with 10% acetic acid; residues were obtained using 200mesh sieves. Specimens are curated in the University of MontanaPaleontology Center, Missoula, MT and assigned University ofMontana Invertebrate Paleontology (UMIP) numbers. Fossil lo-calities were also given Montana Invertebrate (MI) numbers withUSGS locality numbers in parentheses (where applicable).
Abbreviations used.—UMIP—University of Montana Invertebrate Paleontology cata-
logue number; note multiple specimens may have one UMIPnumber.
D—Corresponds to diameter of colony; usually longest dimensionof colony (or fragment).
d—Corresponds to the corallite diameter (in mm) for individualcorallites within a single colony.
c-c—Distance between consecutive calical centers within mean-droid or pseudomeandroid colonies.
s—Relates to the number of counted septa within a given coral-lite.
s/3mm—Relates to the number of septa occurring in three milli-meters of a calice, usually used in cases with meandroid coralsor in cases where there are many fine septa warranting an av-erage instead of a direct count.
s/5mm—Corresponds to the number of septa present in five mil-limeters.
S1, S2, . . . —Corresponds to cycle (or order) of septal growth.
Class ANTHOZOA Ehrenberg, 1835Order SCLERACTINIA Bourne, 1900
Suborder PACHYTHECALIINA Eliasova, 1976Superfamily VOLZEIOIDEA Cuif, 1977
Family CYCLOPHYLLIIDAE Roniewicz, 1989Genus KOMPSASTERIA Roniewicz, 1989
Type species.⎯Isastraea oligocystis Frech, 1890.
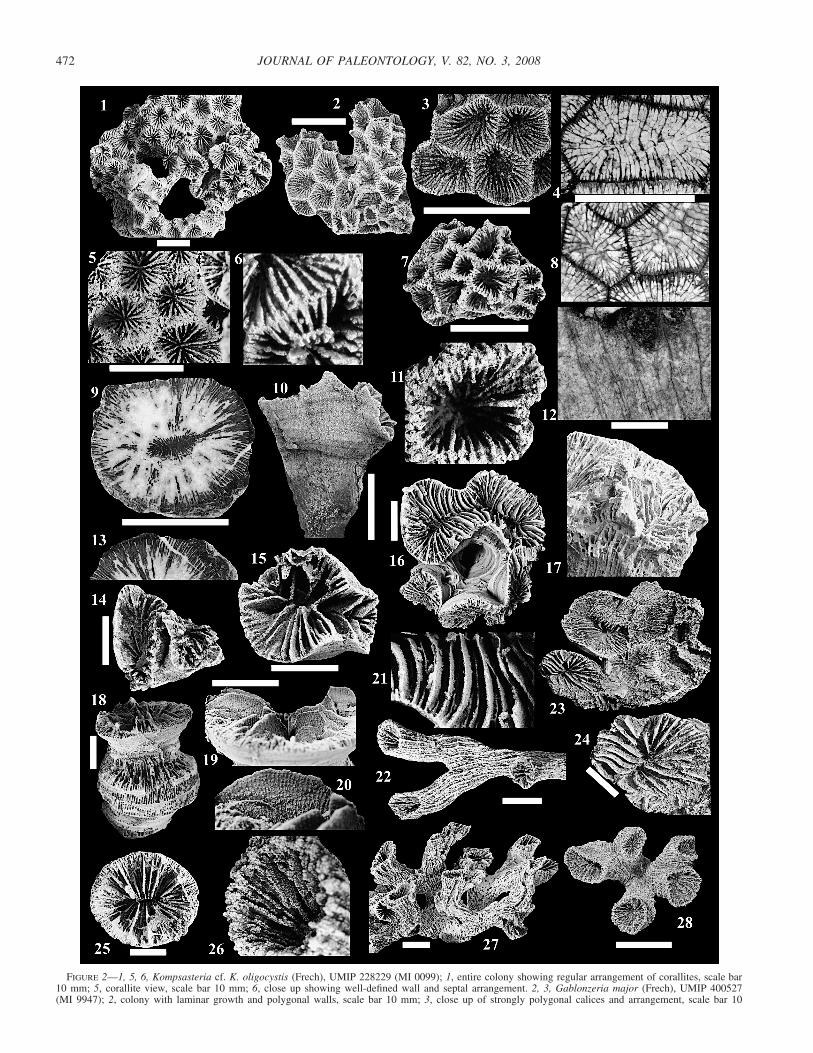
KOMPSASTERIA cf. K. OLIGOCYSTIS (Frech, 1890)Figure 2.1, 2.5, 2.6
Isastraea oligocystis FRECH, 1890, p. 24, pl. 5, fig. 8, 8a, 8b.Kompsasteria oligocystis (Frech) RONIEWICZ, 1989, p. 30, pl. 2, figs. 2, 3, pl.
3, figs. 1–5.
Description.⎯Colony lamellate, cerioid, D � 52 � 60 mm; corallites sub-circular to slightly polygonal; walls broad, lack detail; calices shallow, d �5.0–8.5 mm, s � 17–31 (22–28 in adults), differentiated to two or threecycles, extend to axis, no columella; granular silicification destroyed mostseptal detail along wall; S1 wedge-shaped, extend between three fourths andall the way to center; S2 thinner, extend one half length of S1; S3 rare, visibleas small projections.
Material examined.⎯A single silicified colony, UMIP 228229, from theCornwallis Limestone, Keku Strait, MI 0099 (M1911), southeast Alaska.
Occurrence.⎯Norian, Keku Strait, Alaska; Rhaetian, Northern CalcareousAlps.
Discussion.⎯Roniewicz (1989) identified this distinctive genusprimarily by septal microstructure and gross corallite and colonymorphology. Our specimen matches well with illustrations byRoniewicz (1989) regarding corallite diameter and colony mor-phology. Lack of significant detail in corallite walls may be at-tributed to weathering, abrasion, or poor preservation, and thuswarrants tentative identification. This is the first report of thisgenus and species outside the former Tethys region.
Family GABLONZERIIDAE Roniewicz, 1989Genus GABLONZERIA Cuif, 1976
Type species.⎯Isastraea profunda major Frech, 1890.
GABLONZERIA MAJOR (Frech, 1890)Figure 2.2, 2.3
Isastraea profunda (Reuss) var. major FRECH, 1890, p. 22, pl. 5, figs. 4, 5.Isastrea whiteavesi CLAPP AND SHIMER, 1911, pl. 40, fig. 9.
472 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
FIGURE 2—1, 5, 6, Kompsasteria cf. K. oligocystis (Frech), UMIP 228229 (MI 0099); 1, entire colony showing regular arrangement of corallites, scale bar10 mm; 5, corallite view, scale bar 10 mm; 6, close up showing well-defined wall and septal arrangement. 2, 3, Gablonzeria major (Frech), UMIP 400527(MI 9947); 2, colony with laminar growth and polygonal walls, scale bar 10 mm; 3, close up of strongly polygonal calices and arrangement, scale bar 10

473CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
TABLE 1—Locality information pertaining to geographic sub area, tectonostratigraphic terrane, formation, age, and Montana Invertebrate (MI) number. FromCaruthers and Stanley (in press).
Locality Symbol Sub area Terrane or craton Formation Age
Montanainvertebrate
number (MI)
— AL Southeast Alaska Alexander — Norian —Flounder Cove FC Keku Strait Alexander Cornwallis E. Norian MI 0099Gil Harbor GH Keku Strait Alexander Hound Isl. Vol. L. Norian MI 0087Southwest of Kousk SWK Keku Strait Alexander Cornwallis E. Norian MI 0074Big Spruce BS Keku Strait Alexander Cornwallis E. Norian MI 0056Cornwallis Peninsula East CPE Keku Strait Alexander Cornwallis E. Norian MI 0070Nehenta Bay NB Gravina Island Alexander Nehenta E. Norian MI 9930–32Phocena Bay PB Gravina Island Alexander Nehenta E. Norian MI 9933Nelson Cove NC Gravina Island Alexander Nehenta E. Norian MI 9935–38— NWR Southern Alaska N. Wrangellia — Norian —Green Butte GB Wrangell Mountains N. Wrangellia Chitistone E. Norian MI 9934— SWR Vancouver Island S. Wrangellia — Rhaetian —Lake Cowichan LC Vancouver Island S. Wrangellia Parsons Bay Rhaetian MI 8302Pender Point PP Vancouver Island S. Wrangellia Parsons Bay Rhaetian MI 9947Tahsis Inlet TS Vancouver Island S. Wrangellia Parsons Bay Rhaetian MI 9913— WA Eastern Oregon Wallowa — Norian —Hells Canyon HC Wallowa Mountains Wallowa Martin Bridge Norian MI 8417— PU Andean Highlands Peru — Norian ? —Shalipayco PU Andean Highlands Peru Pucara Gr. Norian ? MI 8811
←
mm. 7, 11, Gablonzeria profunda (Reuss), UMIP 303362 (MI 0099); 7, entire colony, scale bar 10 mm; 11, close up showing septal arrangement. 4, 8, 12,Gablonzeria grandiosa n. sp., holotype UMIP 400488 (MI 9930); 4, 8, thin section, surface view showing polygonal walls and arrangement, scale bar 10mm; 12, polished longitudinal view, note abundant dissepiments, scale bar 10 mm. 9, 13, Cuifia cf. C. marmorea Frech, UMIP 400482 (MI 9930); 9, polishedsection cut horizontally across calice showing heavy recrystallization, scale bar 50 mm; 13, close up with few well preserved septa. 14, Paracuifia smithi(Smith), UMIP 218066 (MI 8831); 14, poorly preserved fragmented colony, scale bar 10 mm. 10, 15, 16, 17, 21, Paracuifia jennieae n. sp.; 10, 15, paratypeUMIP 400296 (MI 9934); 16, 17, 21, holotype UMIP 400043 (MI 0099); 10, longitudinal view of 15, scale bar 10 mm; 15, surface view with initial start offlabellate growth, scale bar 10 mm; 16, surface view, growth form evident, scale bar 10 mm; 17, longitudinal view with abundant dissepiments and noepitheca; 21 close up of 16 showing septa curving toward axial fissure. 23, 24, Paracuifia anomala n. sp., holotype UMIP 400360 (MI 9938); 23, colonysurface view showing large, irregularly shaped calices; 24, close up with well preserved septa curving toward axis, no ornamentation, scale bar 10 mm. 18,19, 20, 25, Distichophyllia norica (Frech), UMIP 400182 (MI 0099); 18, longitudinal view of multi-generational stalk and epitheca, scale bar 10 mm; 19,well preserved septa with abundant ornamentation in vertical rows trending axially; 20, close up, scale bar 10 mm; 25, surface view, well-defined axial fissureand arrangement, scale bar 10 mm. 22, 26, 27, 28, Retiophyllia alfurica (Wilckens), three specimens UMIP 400526 (MI 9947); 22, lateral view showingbifurcation without epitheca, scale bar 10 mm; 26, close up of calice, ornamentation along lateral surfaces of septa; 27, lateral view of colony with uniquesnake-like stalks and trumpet-shaped corallites, scale bar 10 mm; 28, surface view of another colony, evident bifurcation and trifurcation of corallites onsingle stalk, scale bar 10 mm.
Elysastraea major (Frech) SQUIRES, 1956, p. 26, figs. 52, 53.Isastraea profunda major (Frech) CUIF, 1976, p. 114, text-fig. 13, pl. 11, fig.
1, pl. 12, figs. 4, 5.Gablonzeria major (Frech) MATZNER, 1986, pl. 9, fig. 8; RONIEWICZ, 1989,
pl. 4, figs. 2, 3, 4, pl. 5, figs. 1, 2.
Description.⎯Colonies laminar and mound shaped, D � 3.0 � 3.0 mm;corallites well preserved, strongly polygonal; walls distinct; calices deep, d� 5.5–7.5 mm; s � 33–54 in three cycles, no details or columella; S1, S2one half corallite radius with equal thicknesses; S3 thinner, wedge-shaped,one fourth length of radius tapering vertically toward each calical depression.
Material examined.⎯Five specimens from the Parson Bay Formation, Van-couver Island, Canada, at Pender Point (MI 9947), one from Clapp and Shi-mer, 1911.
Occurrence.⎯Rhaetian, Vancouver Island, Canada; Norian, Northern Cal-careous Alps; Lewiston, Idaho.
Discussion.⎯Our specimens match well with original illustra-tions by Frech (1890) and those of Roniewicz (1989), specificallyin corallite size, shape, and septal arrangement.
GABLONZERIA PROFUNDA (Reuss, 1854)Figure 2.7, 2.11
Isastraea profunda REUSS, 1854, p. 116, pl. 9, figs. 5, 6.(See RONIEWICZ, 1989; STANLEY, 1994.)Guembelastraea vancouverensis (Clapp and Shimer) PRINZ-GRIMM, 1995, p.
238, text-fig. 5, fig. 8g.
Description.⎯Colonies mound-shaped, cerioid; budding intracalicular, D� 22 � 18 mm; corallites oval, circular; walls shared; calices sloping, deep,d � 2.5–4.5 mm; s � 16–35 in three cycles, no columella; S1 thicker, wedge-shaped, straight or wavy, extend one half radius; S2 thinner, extend one fourthradius; S3 small, along wall.
Material examined.⎯Eight silicified specimens from the Cornwallis Lime-stone in Keku Strait, MI 0099 (M1911) and MI 0074 (M2136); fifteen fromthe Chitistone Formation, MI 9934 (M1708).
Occurrence.⎯Norian to Rhaetian, Lake Cowichan, Vancouver Island;Lime Peak, Yukon, Canada; Keku Strait, Wrangell Mountains, Lake Iliamna,Alaska; Hells Canyon, Wallowa Mountains, Oregon; Lewiston, Idaho; PilotMountains, Nevada; Lake Shasta, California; Zlambach beds, Austria; PamirMountains, Russia; Japan and northern Chile.
Discussion.⎯Gablonzeria profunda (Reuss) is a widely dis-persed, cosmopolitan species reported from many terranes of theNorth American Cordillera as well as Tethyan and non-Tethyanlocalities. Stratigraphically this species is found throughout theearly Norian to Rhaetian strata. Our material matches well withspecimens by Stanley and Whalen (1989, p. 812, figs. 6.10, 7.11).
GABLONZERIA GRANDIOSA new speciesFigure 2.4, 2.8, 2.12
Diagnosis.⎯Massive colony, cerioid with large polygonal cor-allites, deep calices and no columella. Intracalicular budding oc-curs by fusing septa. Three to four orders of septa present; S1reach central axis, straight, wavy or zigzag shaped.
Description.⎯Colony massive, cerioid, D � 195 mm; corallites sharp, po-lygonal; calices deep, d � 6.0 � 6.5 mm with s � 40 (newer), d � 6.0 �12.0 mm with s � 74–82 (older); septa preserved to three or four orders, nocolumella; S1 reach axis, wedge-shaped, thicker towards endotheca, straight,wavy or zigzag; S2 thinner, one half of S1; S3 one fourth length of S1; S4rare, visible along endotheca. Dissepiments between S1 and S2, rarely be-tween S3 and S4; intracalicular budding by fusing S1 across parent coralliteforming new wall; similar to cell division (Fig. 2.8).

474 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
Etymology.⎯Latin, grandiosum for the characteristically large corallites ofthis species in comparison to all other species of the genus.
Types.⎯Holotype: Figure 2.3, 2.4, 2.7 (UMIP 400488) investigated in thinsection from the Nehenta Limestone, MI 9930 on Gravina Island, southeastAlaska. Paratypes: none.
Discussion.⎯Corallite shape, mode of budding, and septal at-tributes (specifically differentiation, thickness, and appearance) inG. grandiosa n. sp. warrant its placement into genus GablonzeriaCuif. Gablonzeria grandiosa n. sp. has much larger corallite di-ameter, colony size, and calical depth in comparison to both G.profunda (Reuss) and G. major (Frech) in Roniewicz (1989, pl.4, figs. 1a, 2a–b).
Suborder CARYOPHYLLIINA Vaughan and Wells, 1943Superfamily REIMANIPHYLLIOIDEA Melnikova, 1974
Family REIMANIPHYLLIIDAE Melnikova, 1974Genus CUIFIA Melnikova, 1975
Type species.⎯Cuifia gigantella Melnikova, 1975.
CUIFIA cf. C. MARMOREA (Frech, 1890)Figure 2.9, 2.13
Montlivaltia marmorea FRECH, 1890, p. 41, pl. 11, fig. 6, 6A.‘‘Montlivaltia’’ marmorea Frech ZANKL, 1969, p. 31, pl. 2, fig. 12 (non text-
fig. 21).non Montlivaltia marmorea Frech SENOWBARI-DARYAN, 1980, p. 39, pl. 3,
fig. 4.Cuifia marmorea (Frech) RONIEWICZ, 1995, p. 15, pl. 1, figs. 1–3, pl. 2, figs.
1–3, pl. 4, fig. 3.
Description.⎯Poorly preserved solitary disk, d � 7.5 � 8.0 � 5.5 cm.Central portion heavily recrystallized and obliterated, s � 280 differentiatedto four or five orders, best preserved around wall. S1 thickest, extend radius;S2–S5 of equal thickness, obscured by recrystallization.
Material examined.⎯One nonsilicified specimen (sectioned) from the Neh-enta Limestone, Gravina Island, MI 9930, southeast Alaska.
Occurrence.⎯Early Norian, Gravina Island, Alaska; Norian, Europe (seeRoniewicz, 1995).
Discussion.⎯The specimen resembles material illustrated inFrech (1890) and Roniewicz (1995). We regard this identificationas tentative based on a single poorly preserved specimen.
Genus PARACUIFIA Melnikova, 2001
Type species.⎯Protoheterastraea magnifica Melnikova, 1984.
PARACUIFIA SMITHI new speciesFigure 2.14
Confusastraea grandissima SMITH (not Frech), 1927, p. 130, pl. 115, fig. 4.Margarastraea aff. deningeri Wilkens MONTANARO-GALLITELLI, RUSSO, AND
FERRARI, 1979, p. 148, pl. 4, figs. 7a, b.
Diagnosis.⎯Very large, irregularly shaped corallites tending tobe pseudomeandroid and slightly bifurcating colony growth. Sep-ta confluent and ornamented.
Description.⎯Colony highly fragmented, dendroid to pseudomeandroid;colony surrounded by thin epitheca, D � 2.3 cm; calices oval, deep, tapering,d � 10–20 mm; corallite wall thick. s � 3–4 (in 3 mm), 5–6 (in 5 mm),differentiated to two or three cycles, thick, wedge-shaped, no columella, or-namented; S1 longer, reach one half radius; S2 visible as small projectionsalong periphery.
Etymology.⎯Named in honor of J. P. Smith, who first described this spec-imen.
Types.⎯Holotype: silicified partial colony from Smith (1927, pl. 115, fig.4), U.S. National Museum of Natural History (USNM 174241). Paratype:silicified colony (MI 8831). Both from the Nehenta Formation, west coast ofGravina Island, southeast Alaska.
Occurrence.⎯Early Norian, Gravina Island, Alaska.Discussion.⎯Smith (1927) first described this species as Con-
fusastraea grandissima Frech. Roniewicz (1989, p. 68, pls. 16,17) later reassigned Frech’s type material from the Alps to Pa-laeastraea grandissima (Frech), which does not resemble Smith’sspecimen from Gravina Island. Montanaro-Gallitelli et al. (1979,p. 148, pl. 4, fig. 7) later reillustrated Smith’s type, placing it insynonymy with Margarastraea aff. M. deningeri Wilckens
(Wilckens, 1937, p. 179, pl. 7, fig. 1), an assignment that cannotbe maintained. As Smith pointed out, his Alaskan specimen isquite distinct with large corallite diameter. Pseudomeandroid in-tegration, intratentacular budding and intratrabecular linkages pro-duce oval calices, strong thick corallite wall, and thick, orna-mented, wedge-shaped septa. The intricate colony shape is veryevident in type material of Smith (1927, pl. 115, fig. 4) and thesecharacteristics justify reassignment to genus Paracuifia Melni-kova. Smith (1912, p. 112) also mentioned Palaeastraea gran-dissima from the Hosselkus Limestone in the Lake Shasta region(Eastern Klamath terrane), but since he did not illustrate it, wecan make no judgment on its taxonomy. With the erection ofParacuifia smithi n. sp., Palaeastraea grandissima is restricted tothe Tethys and no longer occurs in North America.
PARACUIFIA JENNIEAE new speciesFigure 2.10, 2.15, 2.16, 2.17, 2.21
Diagnosis.⎯Elevated circular growing flabellate colony de-fined by a single pseudomeandroid meander occasionally pinchedto form individual calices. Subhorizontally folded epitheca sur-rounds colony with horizontal dissepiments occurring regularly.Septa blade-like, extend to central fissure, bending to define cal-ice, no ornamentation.
Description.⎯Colony cone shaped, flabellate, stalk-like, D � 4.0 � 4.5� 5.0 cm, surrounded by a well preserved, thin, subhorizontally folded epi-theca, horizontal dissepiments exposed along worn surfaces. Corallites definedby single pseudomeandroid meander, occasionally pinched, forming deep, of-ten interconnected calices, d � 1.7 � 2.0 mm, c-c � 10–15 mm, s � 5–7(in 3 mm) and 9–11 (in 5 mm) in holotype and 54 in paratypes. Septa thin,slightly wedge-shaped with granular ornamentation; differentiated to three orfour cycles; S1 bend toward fissure defining calices, occasionally fuse to formindividual calices; S2 thinner, extend one half length of S1; S3 (and S4) small,project one eighth to one fourth size of S1.
Etymology.⎯After the given name of Jennie Mary Smoly/Caruthers, abeautiful, kind-hearted woman whose strength, determination and supportivenature will never be forgotten.
Types.⎯Holotype silicified: UMIP 400043 from the Cornwallis Limestone,MI 0099 (M1911), Keku Strait, southeast Alaska. Paratypes (three, one fig-ured) silicified: Figure 2.10, 2.15 (UMIP 400296) from the Chitistone For-mation, Wrangell Mountains, MI 9934 (M1708) south-central Alaska.
Discussion.⎯Particularly large corallite diameter, intricate cal-ical shape, wedge-shaped radial elements bending toward the cen-tral fissure and granular ornamentation on lateral septal surfaceshelp place this unique new species in genus Paracuifia Melnikova(Roniewicz, personal commun., 2007). Melnikova (2001) indi-cated dendroid integration, which is particularly evident in para-types of P. jennieae new species; the holotype reveals a mean-droid appearance. Paracuifia jennieae n. sp. is unique in that itforms a single sinuous elevated meander without adjacent coun-terparts.
PARACUIFIA ANOMALA new speciesFigure 2.23, 2.24
Diagnosis.⎯Colony thin, laminar; unique large irregularly-shaped cerioid corallites and folded epitheca. Calices shallow,broad, contain well-defined blade-like septa; S1 and S2 equallength, thickness, reach center.
Description.⎯Colony thin, laminar, D � 10 cm; corallites sub-phaceloidto meandroid, slightly elevated, large, irregular, circular and elongate; d �20–35 mm (50 mm longest), new corallites fuse with existing upon growth,surrounded by thin, subhorizontally folded epitheca. Calices shallow, broad,well-defined walls; s � 40 (circular calice), 6–8 (in 5 mm of elongate calices),poorly preserved, granular, differentiated to three or four cycles, bending to-ward center; S1 and S2 thickest, most pronounced, extend entire radius; S3and S4 thinner, one fourth to one half length of S1.
Etymology.⎯Greek, anom- for its irregularly shaped, large corallites.Types.⎯Holotype silicified: UMIP 400360 from the Nehenta Formation,
MI 9938, Gravina Island, southeast Alaska. Paratypes: none.Discussion.⎯Paracuifia anomala new species has large irreg-
ularly shaped cerioid corallites, slightly elevated, resemblingthose from genus Paracuifia Melnikova (Roniewicz, personal

475CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
commun., 2007). Paraquifia anomala n. sp. differs from Mar-garastraea klipsteini Frech in Volz (1896, pl. 5, fig. 7a–d) withmuch larger corallites, far fewer septa (40 in comparison to 100in M. klipsteini Frech), thick S1 and S2 bending to accentuate thecalice (Fig. 2.24), thin, subhorizontally folded epitheca surround-ing the colony, and new corallites growing into and fusing withthe existing colony upon growth.
Family DISTICHOPHYLLIIDAE Cuif, 1977Genus DISTICHOPHYLLIA Cuif, 1975a
Type species.⎯Montlivaltia norica (Frech, 1890).
DISTICHOPHYLLIA NORICA (Frech, 1890)Figure 2.18, 2.19, 2.20, 2.25
Montlivaltia norica FRECH, 1890, p. 39, pl. 3, figs. 8, 9, pl. 10, figs. 1–5, pl.18, fig. 17.
(See Roniewicz, 1989; Stanley, 1994; Melnikova, 2001.)
Description.⎯Solitary, variable in height, flat, conical or cylindrical withthin to moderately thick epitheca; d � 1.3 � 2.3 cm to 4.5 � 4.0 cm; reju-venation through intracalicular budding; forming large multi-generationalstalks; s � 58–140, differentiated to four cycles around fissure; S1, S2 thick-est, uniform, more pronounced, extend to center with ornamentation (orspines) along lateral surfaces parallel to axis (Fig. 2.19, 2.20); S3 one half tothree quarters length of S1, no ornamentation; S4 (if present) thinnest, onequarter to one half radius.
Material examined.⎯Fifteen silicified corals from the Cornwallis Lime-stone, MI 0099 (M1911), MI 0087 (M1912), MI 0074 (M2136), MI 0070,Keku Strait, southeast Alaska. Twenty-seven silicified corals from the Chiti-stone Formation, MI 9934 (M1708), in the Wrangell Mountains, southcentralAlaska.
Occurrence.⎯Extremely widespread species. Rhaetian, Vancouver Island(Lake Cowichan); Norian, Wrangell Mountains, Keku Strait, Long Creek,Alaska; Hells Canyon, Wallowa Mountains, Oregon; Lewiston, Idaho; PilotMountains, Nevada; Austria; Mt. Everest, Tibet; Timor; southeastern PamirMountains and Koryak Highlands, northeastern Russia; Japan.
Discussion.⎯Specimens match well with illustrations ofSquires (1956), Montanaro-Gallitelli et al. (1979), Stanley andWhalen (1989), and descriptions of septal detail in Roniewicz(1989). We observe three to four orders of septa compared withfive described by Roniewicz (1989). Zankl (1969) and Squires(1956) reported lateral budding and Frech (1890; pl. 10, figs.1–5) illustrated specimens with twisting or sunken axial fissures,to which none of our specimens match.
Genus RETIOPHYLLIA Cuif, 1967Type species.⎯Thecosmilia fenestrata Reuss, 1854 in Frech,
1890.Discussion.⎯Found in Upper Triassic material from both Eu-
rope and North America, this very speciose branching genus issubject to constant revision. Species were identified, in part, bycomparisons to Roniewicz’ (1989) detailed species differentiation(exclusive of microstructure) including: 1) number of septa, 2)costae, 3) presence and nature of connecting processes, 4) natureof endotheca, and 5) angle of bifurcation.
RETIOPHYLLIA ALFURICA (Wilckens, 1937)(Figure 2.22, 2.26, 2.27, 2.28)
Thecosmilia alfurica WILCKENS, 1937, pl. 9, figs. 1, 2.
Description.⎯Low growing, dendroid, epitheca absent or thin, no con-necting processes; corallites bifurcate (and trifurcate); calices circular or oval,tapering (trumpet shaped), d � 14 � 8 mm (adults) and 6.5 � 3.5 mm(juveniles); s � 31–34 (juveniles) and 50–63 (adults), differentiated into threeor four cycles, variable thicknesses around calice. S1 most prominent, extendone half of radius, ornamentations along lateral surfaces (Fig. 2.26); S2 thin-ner, one half length of S1; S3 hard to distinguish from endotheca, only assmall projections; S4 rare, resembles S3.
Material examined.⎯Four silicified colonies and many fragments fromnorthern Vancouver Island, Canada (MI 9947).
Occurrence.⎯Rhaetian, northern Vancouver Island, Canada; Norian, Ser-am, Molluccas.
Discussion.⎯Wilckens (1937) originally described and illus-trated this species from the island of Seram. It is apparent that
the holotype was not etched and therefore the specimen does notshow colony growth form because it is covered in matrix.
Retiophyllia alfurica (Wilckens) differs from other species inthe genus by characteristic trumpet-shaped corallites displayingregular bifurcation (and occasional trifurcation; Fig. 2.28) uponbudding, supported by irregular (bending) stalks. Retiophyllia al-furica (Wilckens) could not be compared with the North Ameri-can species R. quesnelliana Stanley and Senowbari-Daryan be-cause of poor preservation and heavy recrystallization; however,R. wrangelliana (in Montanaro-Gallitelli et al., 1979) does notdisplay trumpet-shaped corallites and is highly fragmented.
RETIOPHYLLIA CAESPITOSA (Reuss, 1865)(Figure 3.1, 3.2, 3.5, 3.6)
Thecosmilia caespitosa REUSS, 1865, p. 159, pl. 3, fig. 3; FRECH, 1890, p. 7,pl. 1, figs. 1–12, (non fig. 13); CUIF, 1966, p. 534, text-fig. 2B.
Thecosmilia caespitosa (Reuss) n.v. pauciseptata HAAS, 1909, p. 144, pl. 5,fig. 1.
Distichophyllum caespitosum (Reuss), CUIF, 1967, p. 129, text-fig. 3, pl. 4,fig. 3; CUIF, 1975b, p. 375, text-figs. 35b, 36; MELNIKOVA AND BYCHKOV,1986, p. 47, pl. 13, fig. 2.
Retiophyllia caespitosa (Reuss), RONIEWICZ, 1989, pl. 8, figs. 2, 4, 9, pl. 9,fig. 3; MELNIKOVA, 2001, pl. 15, fig. 1.
Description.⎯Colony phaceloid, frequently bifurcating (or trifurcating)surrounded by subhorizontal epithecal folds, thick calical rim and taperingdepression (8 mm deep), no columella. Corallites circular or oval, taper to-wards stalk (trumpet shaped); d � 6–9 mm; s � 35–39, wedge shaped, pre-served to three or four cycles. S1, S2 equal in thickness, S1 extends threefourths of radius, containing ornamentation (Fig. 3.1); S2 one half length ofS1, no clear ornamentation; S3 and S4 exist only along endotheca.
Material examined.⎯Thirty-five silicified colonies from the ChitistoneFormation, MI 9934 (M1708) in the Wrangell Mountains, south-central Alas-ka.
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; Norian, Cauca-sus, central and northeast Asia; Rhaetian, Northern Calcareous Alps.
Discussion.⎯Our specimens closely match Reuss’ (1865) orig-inal description and revisions by Roniewicz (1989). Retiophylliacaespitosa (Reuss) is distinctive with frequent bifurcation, pro-portionate thickness of all radial elements, club or wedge-shapedsepta, a thickened skeleton, and S1 septa that do not extend theentire length, resulting in free space. Retiophyllia caespitosaclosely resembles R. alfurica (Wilckens) in having trumpet shapedcorallites, commonly bifurcating (and trifurcating) upon budding,and ornamentation on S1 septa. However, these two species differwith respect to corallite size, frequency of budding, shape of stalk,and the presence of thick subhorizontal epithecal folds.
Stanley (1986, p. 27), in a list of verified taxa, showed R. caes-pitosa as occurring in Hells Canyon, Oregon (Wallowa terrane),but no illustration or systematics were given, thus precludingcomparison. Smith (1927, pl. 121, fig. 7) described Thecosmiliacf. T. caespitosa Reuss from Iliamna Lake, Alaska; however, il-lustrations do not show any distinguishing features to identify thatspecimen with those of Reuss (1865) or Roniewicz (1989). There-fore Smith’s identification is highly questionable, making R. caes-pitosa known only with certainty from the Wrangell Mountains(northern Wrangellia) in North America.
RETIOPHYLLIA CLATHRATA (Emmrich, 1853)(Figure 3.3, 3.4, 3.8, 3.9)
Thecosmilia clathrata Emmrich FRECH, 1890, (part) pl. 4, fig. 11.Thecosmilia fenestrata Reuss FRECH, 1890, (part) pl. 2, figs. 11, 12a, 12b.Calamophyllia suttonensis CLAPP AND SHIMER, 1911, p. 431, pl. 40, figs. 5,
7, pl. 42, fig. 15.Thecosmilia fenestrata Reuss SMITH (not Reuss), 1927, p. 128, pl. 105, figs.
1, 9.Thecosmilia cf. suttonensis (Clapp & Shimer) SQUIRES, 1956, p. 22, fig. 31.Thecosmilia wrangelliana RUSSO in Montanaro-Gallitelli, Russo, and Ferrari,
1979, pl. 4, fig. 3.Retiophyllia dawsoni (Clapp and Shimer) STANLEY AND WHALEN, 1989, fig.
5.8, 5.9, 5.13, 5.14.Retiophyllia clathrata (Emmrich) PRINZ, 1991, p. 156, pl. 1, fig. 4.
Description.⎯Colonies large, D � 25 � 20.5 � 9.0 cm, phaceloid, acutely

476 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
FIGURE 3—1, 2, 5, 6, Retiophyllia caespitosa (Reuss) four individual fragments, UMIP 400321 (MI 9934); 1, calical view showing ornamentation on septalsurfaces; 2, lateral view with thick folded epitheca clearly visible, scale bar 10 mm; 5, close up of corallite showing horizontal ornamentations along septalsurfaces; 6, surface view showing trifurcation of corallites, imbedded gastropod from silicification, scale bar 10 mm. 3, 4, 8, 9, Retiophyllia clathrata (Emmrich),

477CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
←
one large colony and one fragment UMIP 221120 (MI 8302); 3, longitudinal view of large colony showing sub-parallel branching and abundant connectingprocesses, scale bar 10 mm; 4, same colony rotated 90� axially; 8, close up of stalk showing hollow connecting process and dissepiments between adjacentsepta; 9, calice with dissepiments and granular ornamentation along septal surfaces, scale bar 5 mm. 10, 11, 12, Retiophyllia dawsoni (Clapp and Shimer),UMIP 400655 (MI 8302); 10, surface view showing multiple corallites and fused branches, scale bar is 10 mm; 11, lateral view of runner-type growth, scalebar 10 mm; 12, close up of oval calice with well preserved septa. 15, 16, Retiophyllia cf. R. frechi Roniewicz (MI 0099); 15, UMIP 228233, calical viewwith well preserved septa and dissepiments, scale bar 10 mm; 16, UMIP 228208, lateral view of septa with alternating thicknesses, scale bar 10 mm. 17, 18,Retiophyllia cf. R. norica (Frech), two fragments UMIP 303353 (MI 0099); 17, poorly preserved lateral surface, scale bar 10 mm; 18, surface view, S1 septabending toward calical center. 7, 14 Retiophyllia oppeli (Reuss), UMIP 400324 (MI 9934); 7, largest corallite on stalk, oval, scale bar 10 mm; 14, lateralview of thick folded epitheca and irregular branching, scale bar 10 mm. 22, 23, 25, Retiophyllia parvisepta (Squires) UMIP 400330 (MI 9934); 22, caliceshowing arrangement, wedge-shaped septa and pseudo-columella, scale bar 5 mm.; 23, acutely branching colony, thick folded epitheca and trifurcation ofcorallites, scale bar 10 mm; 25, calice showing the septa without ornamentation. 13, 20, 21, Retiophyllia cf. R. robusta Roniewicz, UMIP 228213 (MI 0099);13, calice with well preserved septa and axial fissure, scale bar 10 mm; 20, surface view showing trifurcation of corallites and thick epitheca, scale bar 10mm; 21, view of thick stalk and trifurcating corallites, scale bar 10 mm. 19, 27, 29, Retiophyllia tenuicosta Reuss, UMIP 400303 (MI 9934); 19, calice withseptal arrangement and granular ornamentation, scale bar 5 mm; 27, longitudinal view with thin epitheca exposed along the upper portion of the stalk, scalebar 10 mm; 29, trifurcating corallites and thin epitheca. 28, 30, 31, 32, 33, 34, Retiophyllia dendriformis n. sp., holotype UMIP 22066 (MI 8302); 28, 30,31, 32, surface views showing variable shape of calices and well preserved septa, scale bar on 30 is 10 mm; 33, longitudinal view with acute, bush-likebranching; 34, same colony rotated 90� axially, scale bar 100 mm. 24, 26, Retiophyllia obtusa n. sp., holotype UMIP 400337 (MI 9934); 24, calice withmoderately well preserved septa and granular ornamentation, scale bar 5 mm; 26, obtuse branching deep calices, and thick, folded epitheca, scale bar 10 mm.
branching; corallites circular or oval, bifurcate into long, parallel, equal-di-mensional stalks supported by hollow connecting processes along epitheca(Fig. 3.8); calices deep, d � 4–6 mm; s � 15–20 in four cycles, dissepimentsbetween all orders; S1 and S2 most prominent, equal length, extend to axis,S1 thickest, contain ornamentation (Fig. 3.9); S3 very thin, one half lengthof S1; S4 rarely observed, one fourth length of S1.
Material examined.⎯Four large silicified colonies from Lake Cowichan,southern Vancouver Island, Canada, MI 8302. Twenty fragmented and silic-ified colonies from the Chitistone Formation, MI 9934, Wrangell Mountains,south-central Alaska.
Occurrence.⎯Rhaetian, Vancouver Island, Canada; early Norian, WrangellMountains, Iliamna Lake, Cook Inlet, Alaska; Hells Canyon, Oregon; Norian,Lewiston, Idaho; northern Chile; Northern Calcareous Alps.
Discussion.⎯Reevaluation of the type material of Clapp andShimer (1911) yields evidence suggesting Calamophyllia sutto-nensis Clapp and Shimer (1911) should be placed in synonymywith R. clathrata (Emmrich), to which we assign our material.Similarities include mode of branching, equal bifurcation, fre-quency of connecting processes, and pinnular nature of septalgrowth. Specimens differ only with regard to number of septa(greater in Clapp and Shimer, 1911; and Frech, 1890). Prinz(1991) recognized this very characteristic reef-building Alpinespecies in silicified material from South America (northern Chile),suggesting comparisons with Clapp and Shimer’s species.
The highly fragmented R. dawsoni (Clapp and Shimer) in Stan-ley and Whalen (1989, fig. 5.8, 5.9, 5.13, 5.14) likely belongs toR. clathrata (Emmrich) based on corallite diameter, lack of epi-theca, and distinctive parallel branching pattern; however, septacount could not be made due to recrystallization within the calice.
RETIOPHYLLIA DAWSONI (Clapp and Shimer, 1911)(Figure 3.10–3.12)
Calamophyllia dawsoni CLAPP AND SHIMER, 1911, p. 431, pl. 40, fig. 1, pl.42, fig. 16.
‘‘Thecosmilia’’ dawsoni STANLEY, 1979, pl. 3, fig. 3.Retiophyllia dawsoni (Clapp and Shimer) MELNIKOVA AND BYCHKOV, 1986,
fig. 2, pl. 7, fig. 1.Retiophyllia pascoensis STANLEY, 1994, pl. 5, figs. 1–5, text-figs. 8, 9.
Description.⎯Colonies small, dendroid, densely packed, D � 3.0 � 2.6� 2.0 cm, surrounded by variably preserved epitheca; corallites circular,slightly oval, d � 3–5 mm, s � 12–17 differentiated into three size orders,no detail or ornamentation; S1 most prominent, extend three fourths of radius;S2 thinner, less abundant; S3 small, only along endotheca.
Material examined.⎯Original material from Clapp and Shimer (1911, pl.42, fig. 16). Two silicified specimens from the Chitistone Formation, MI 9934(M1708), Wrangell Mountains, south-central Alaska. Fifteen silicified frag-ments from Lake Cowichan, MI 8302, and another locality at Pender Point,MI 9947, Vancouver Island, Canada.
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; Rhaetian, LakeCowichan, Pender Point, Vancouver Island, Canada; Norian, Pucara Group,central Peru; Koryak Highlands, northeastern Russia.
Discussion.⎯Clapp and Shimer (1911) originally identify Cal-amophyllia dawsoni from Lake Cowichan in Vancouver Island,
Canada. Melnikova and Bychkov (1986) reaffirm the validity ofthis species, including it in the genus Retiophyllia Cuif. Stanley(1994) illustrated and described in much detail a new species,Retiophyllia pascoensis from the Pucara Group, Peru. It closelyresembles R. dawsoni Clapp and Shimer in corallite diameter,shape, septal attributes, thin epitheca, and distinct low growingcolonies along the sediment/water interface. Therefore, we in-clude R. pascoensis Stanley as R. dawsoni (Clapp and Shimer),attributing the fewer septa (30–50 in R. pascoensis Stanley, 12–17 in our material) to diagenesis.
Type material by Clapp and Shimer (1911, pl. 42, fig. 16) isheavily recrystallized, poorly illustrated upside down, and vague-ly described (precluding accurate comparisons). Furthermore, theartistic sketch (Clapp and Shimer, 1911, pl. 40, fig. 1) showingcritical septal details was not confirmed in our reevaluation of thetypes. The excellent topotype material of R. dawsoni we illustratehere supplements Clapp and Shimer’s original specimens. Thedescription in Stanley (1994) encompasses colony shape, growthform, and corallite morphology.
Our material resembles holotype material of both R. dawsoni(Clapp and Shimer) and R. pascoensis Stanley; in particular, thetwo dendroid growth forms or morphotypes (first mentioned byStanley, 1994) are recognized in material from this study. North-ern Wrangellia (Wrangell Mountains) contains only the verticallygrowing morphotype whereas southern Wrangellia (Lake Cowi-chan and Pender Point, Vancouver Island) contains vertical andhorizontally growing morphotypes. Retiophyllia dawsoni differsfrom R. oppeli (Reuss) in smaller corallites, straighter stalk, andmuch thinner (non-folded) epitheca. Retiophyllia dawsoni is en-demic to the terranes of the American Cordillera and unknown inthe Tethys, distributed from Peru to Alaska.
RETIOPHYLLIA cf. R. FRECHI Roniewicz, 1989(Figure 3.15, 3.16)
Thecosmilia fenestrata Reuss FRECH, 1890, p. 9, pl. 1, figs. 25–27, pl. 2, figs.1–11, 13–17 (non fig. 12).
Calamophyllia fenestrata Reuss CUIF, 1965, p. 533, text-fig. 2 A.Retiophyllia fenestrata (Reuss) CUIF, 1967, p. 130, text-fig. 4 A, B, pl. 4, fig.
4; CUIF, 1975a, p. 369, text-figs. 33–35.Retiophyllia frechi RONIEWICZ, 1989, p. 48, pl. 7, figs. 1–8, pl. 9, fig. 5, pl.
13, fig. 2.
Description.⎯Three fragmented colonies, no apparent branching or bud-ding; epitheca worn, exposing distal septal edges with alternating thickness,no dissepiments. Corallites variable, d � 12 � 13 mm and 17 � 18 mm, s� 48–56, differentiated into two or three cycles, no columella. S1 longer,wedge shaped; S2 thinner, extend from one half to same length as S1. Dis-sepiments observed along endotheca, between S1 and S2.
Material examined.⎯Three silicified fragments from the Cornwallis Lime-stone, Keku Strait, MI 0099 (M1911), southeast Alaska.
Occurrence.⎯Early Norian, Keku Strait, Alaska; Norian, Pucara Group,Peru; Rhaetian, Northern Calcareous Alps.

478 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
Discussion.⎯Longitudinal view shows septa with alternatingthickness and few dissepiments, similar to descriptions in Ron-iewicz (1989, p. 49), but differing with respect to corallite di-ameter and lack of connecting processes, and therefore warrantinga tentative identification with R. frechi Roniewicz.
RETIOPHYLLIA cf. R. NORICA (Frech, 1890)(Figure 3.17, 3.18)
Thecosmilia norica FRECH, 1890, p. 9, pl. 1, figs. 14–24.Thecosmilia norica var. densisepta HAAS, 1909, p. 145, pl. 5, fig. 3.Thecosmilia norica var. lobatisepta HAAS, 1909, p. 145, pl. 5, fig. 4.Paradistichophyllum noricum (Frech). MELNIKOVA, 1975, p. 90, pl. 15, figs.
3, 4.‘‘Thecosmilia’’ cf. ‘‘T.’’ fenestrata (Reuss) STANLEY, 1979, p. 56, pl. 3, fig. 1.Retiophyllia norica (Frech) RONIEWICZ, 1989, p. 52, pl. 8, figs. 5–8, pl. 9,
fig. 2, pl. 14, figs. 1, 2; STANLEY, 1994, p. 12, figs. 10.1, 10.2.
Description.⎯Poorly preserved phaceloid colonies, obtuse bifurcation;corallites circular to oval, d � 13–18 mm, horizontal dissepiments poorlypreserved along endotheca (rare); s � 28–36, differentiated into three to fourcycles, all thicker toward pellicular wall, no ornamentation or columella,curve toward center along budding margin (Fig. 3.18). S1 and S2 extend toradius, S1 thickest; S3 one half to three fourths length of S1; S4 (rare, thin)exist near endotheca.
Material examined.⎯Two silicified fragments from the Cornwallis Lime-stone, Keku Strait, MI 0099 (M1911), southeast Alaska; four from the Chi-tistone Formation, Wrangell Mountains, MI 9934 (M1708), south-centralAlaska.
Occurrence.⎯Norian, Keku Strait, Wrangell Mountains, Long Creek,Alaska; Pilot Mountains, Nevada; Lime Peak, Yukon, Canada; Sonora, Mex-ico; Northern Calcareous Alps, Austria.
Discussion.⎯Identification is tentative because of poor pres-ervation and small sample size. Specimens match the morpholog-ical description of Roniewicz (1989) with respect to corallite size,shape, septal arrangement, and angle of bifurcation. Our samplesalso resemble material by Frech (1890, pl. 1, figs. 14–24), Ron-iewicz (1989, pl. 8, figs. 5–8), and Yarnell (2000, pl. 2, figs.4, 5).
RETIOPHYLLIA OPPELI (Reuss, 1865)(Figure 3.7, 3.14)
Calamophyllia oppeli REUSS, 1865, p. 160, pl. 4, fig. 1.Thecosmilia oppeli (Reuss), FRECH, 1890, p. 10, pl. 2, figs. 18–20, 24, pl. 3,
figs. 4 A–E (non pl. 2, figs. 21–23); VINASSA DE REGNY, 1915, p. 89, pl.69, figs. 7, 8; MONTANARO-GALLITELLI, RUSSO, AND FERRARI, 1979, pl.4, fig. 2.
Retiophyllia oppeli (Reuss), RONIEWICZ, 1989, pl. 9, figs. 7, 8, pl. 10, figs.3, 4; PRINZ, 1991, pl. 1, fig. 5.
Description.⎯Phaceloid, uneven bifurcation, zigzag branches covered byuneven, thick epitheca. Corallites variable, sub-circular and oval; calices deep(2–3 mm), d � 4 and 12 mm, s � 20 and 47 respectively, two or three cyclespreserved; S1 thickest, wedge shaped, most prominent; S2 and S3 smaller,project one half the length of S1.
Material examined.⎯A single silicified colony from the Chitistone For-mation, Wrangell Mountains, MI 9934 (M1708), south-central Alaska.
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; Hells Canyon,Oregon; Lime Peak, Yukon, Canada; Sonora, Mexico; northern Chile;Rhaetian, Northern Calcareous Alps; Timor.
Discussion.⎯Retiophyllia oppeli (Reuss) is distinctive for itssmall corallite diameter and shape, frequent acute bifurcations,connecting processes, and S1 septa being twice or three times asthick as S2 or S3 septa (Roniewicz, 1989), to which our specimenmatches except for the presence of connecting processes. Mon-tanaro-Gallitelli et al. (1979, pl. 4, fig. 2) identified Thecosmiliaaff. T. oppeli (Reuss) from the Wrangell Mountains (M1708).
RETIOPHYLLIA PARVISEPTUM (Squires, 1956)(Figure 3.22, 3.23, 3.25)
Pinacophyllum parviseptum SQUIRES, 1956, p. 7, figs. 4–8.Pinacophyllum (?) parviseptum Squires MONTANARO-GALLITELLI, RUSSO,
AND FERRARI, 1979, pl. 1, fig. 1.Retiophyllia dawsoni (Clapp and Shimer) STANLEY AND WHALEN, 1989, fig.
7.6 (non fig. 5.8, 5.9, 5.13, 5.14).
Description.⎯Colonies phaceloid, acutely branching, epitheca folded; cor-allites circular, same diameter as stalk; calices deep, d � 3–6 mm, s � 18–33, wedge shaped, thin, differentiated to three cycles, no ornamentation (Fig.3.25); S1 and S2 most prominent, extending to axis, S3 shorter, only visiblearound periphery; columella absent (pseudocolumella diagenetic).
Material examined.⎯Six specimens from the Chitistone Formation, Wran-gell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; Hells Canyon,Oregon; late Norian, Lewiston, Idaho.
Discussion.⎯Our material clearly belongs to Retiophyllia Cuif,not Pinacophyllum Frech, as made by Squires (1956), eventhough microstructure is absent. We place this species in genusRetiophyllia Cuif based on acute dendroid branching, granularstraight, non-beaded septa reaching the axis without columella,smooth to slightly folded epitheca, and moderately deep calice(all common traits of the genus); however, the lack of distinguish-ing features does not enable specific identification.
Insufficient detail and corallite preservation preclude synonymywith Pinacophyllum cf. P. parviseptum Squires in Stanley andWhalen (1989, fig. 7.14). Retiophyllia dawsoni (Clapp and Shi-mer) in Stanley and Whalen (1989, fig. 7.6) resembles this speciesand is placed in synonomy, as evidenced by the thickly foldedepitheca, septal arrangement, calical depth, corallite shape, anddiameter. However, as pointed out by Squires (1956), a spiralbudding pattern within the bifurcation plane is not present.
RETIOPHYLLIA cf. R. ROBUSTA Roniewicz, 1989(Figure 3.13, 3.20, 3.21)
Thecosmilia norica FRECH, 1890, pl. 10, fig. 6.Retiophyllia robusta RONIEWICZ, 1989, pl. 8, figs. 1, 3, 10.
Description.⎯Colonies phaceloid, poorly preserved; pellicular walls sur-round entire colony, thicker near calice; corallites circular, d � 15–20 mm; s� 50–55, well preserved, differentiated into three cycles around calice; S1thickest, reach axis; S2 one half to three fourths the length of S1; S3 small,rare.
Material examined.⎯Two specimens from the Cornwallis Limestone,Keku Strait, MI 0099 (M1911), southeast Alaska.
Occurrence.⎯Early Norian, Keku Strait, Alaska; Rhaetian, Northern Cal-careous Alps.
Discussion.⎯Identification is tentative, warranted only by an-gle of bifurcation and colony shape, vaguely matching nonsilici-fied material of Roniewicz (1989); corallite diameter and numberof septa also match, but are on the higher end. Poor preservationand small sample size create doubt, so more material is neededfor further study.
Retiophyllia robusta Roniewicz previously was known onlyfrom Rhaetian deposits in the Alps (former Tethys), so if correct,this constitutes a first occurrence in the North American Cordil-lera (Alexander terrane).
RETIOPHYLLIA TENUICOSTA (Reuss, 1854)(Figure 3.19, 3.27, 3.29)
Rhabdophyllia tenuicosta REUSS, 1854, pl. 6, figs. 18–21.
Description.⎯Colonies phaceloid, fragmented, D � 22 � 8 mm, bifur-cation and trifurcation upon budding, partially covered by thin, slightly foldedepitheca, horizontal dissepiments occur along stalk; corallites circular; calicesdeep (1–2 mm), taper toward stalk, d � 3.0–5.5 mm; s � 12–26 in threecycles around axis; S1 most pronounced, wedge shaped, granular ornamen-tation on lateral surfaces (Fig. 3.19, 3.29), reach one half radius; S2 extendone fourth radius, thinner than S1; S3 wedge-shaped, visible along wall.
Material examined.⎯Three fragments from the Chitistone Formation,Wrangell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; Carnian, North-ern Calcareous Alps.
Discussion.⎯Material is similar to measurements and illustra-tions of type material in Reuss (1854).
Rhabdophyllia Edwards and Haime is not mentioned in recentUpper Triassic coral taxonomic revisions and therefore little isknown about the microstructure and associated septal details nec-essary to accurately establish this genus in modern taxonomic

479CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
schemes. Characteristics with Retiophyllia Cuif seem more tena-ble. Similarities include: septal arrangement, lack of columella,bifurcation or trifurcation of corallites, and subhorizontally foldedepitheca partially covering colony surfaces. Assignment to genusPinacophyllum Frech is not warranted because of the ornamen-tation (Fig. 3.19, 3.29) along the septa, which resembles that inRetiophyllia Cuif, rather than in Pinacophyllum Frech.
RETIOPHYLLIA DENDRIFORMIS new species(Figure 3.28, 3.30–3.34)
Thecosmilia cf. T. dawsoni (Clapp & Shimer) SQUIRES, 1956, p. 23, figs. 29–30.
Thecosmilia dawsoni (Clapp and Shimer) MONTANARO-GALLITELLI, RUSSO,AND FERRARI, 1979, pl. 4, fig. 1a, b.
Diagnosis.⎯Bush-like, frequent branching, bifurcating alongstalk with connecting processes. Corallites occasionally bulbousconstricted with a thin or non-existent epitheca; S1 septa regularlyprotrude above calical rim.
Description.⎯Colonies large, low-growing, dendroid, bush-like; denselypacked corallites from frequent, acute, extracalicular budding; epitheca thinor non-existent. Calices sub-circular or oval, occasionally quenched or con-stricted (Fig. 3.28, 3.30–3.32) with connecting processes between corallitesand neighboring stalks (Fig. 3.34); d � 5–9 mm, s � 23–32 in two or threecycles around the calice. S1, S2 extend radius, equally thick; S1 thicker, morepronounced, protrude above calical rim, questionable ornamentation; S3small, rare, wedge-shaped, project along pellicular wall.
Etymology.⎯Latin, dendros for the characteristically dense, acutelybranched corallites, similar to the branches of a tree.
Types.⎯Holotype silicified: figured UMIP 22066. Paratypes silicified: onelarge colony not figured (UMIP 400718) and many individual branches withgood septa and preservation (UMIP 221097) etched from Lake Cowichan,Vancouver Island, Canada, MI 8302.
Other material examined.⎯Twelve silicified fragments from the ChitistoneFormation, Wrangell Mountains, MI 9934 (M1708), south-central Alaska.
Occurrence.⎯Rhaetian, Lake Cowichan, Vancouver Island, Canada; lateNorian, Lewiston, Idaho; early Norian, Wrangell Mountains, Alaska.
Discussion.⎯This species is easily confused with R. clathrata(Frech), both occurring in great abundance within in situ phace-loid coral thickets at Lake Cowichan, Vancouver Island (Stanley,1989), with comparable corallite diameters and septal counts aswell. However, R. dendriformis n. sp. is distinguished throughmuch shorter, bush-like dendroid growth, constricted, pinched orbulbous calices, and high corallite density. Compared to our newspecies, R. clathrata (Frech) has extremely abundant connectingprocesses, implying a different kind of colony integration or adifferent mode of colony stabilization.
Specimens from Squires (1956) and Montanaro-Gallitelli et al.(1979) are fragmented and poorly preserved. Although they havedimensions matching both R. clathrata (Frech) and R. dendrifor-mis n. sp., growth forms of these fragments are dendroid withdensely packed corallites, not phaceloid with parallel branches,thus indicating inclusion in our new species, further showing sim-ilarity between Wrangellia and the Wallowa terrane.
RETIOPHYLLIA OBTUSA new species(Figure 3.24, 3.26)
Diagnosis.⎯Colony phaceloid, budding extracalicularly, dis-tinguished by unusual, obtuse branching form, surrounded bythick subhorizontally folded epitheca; corallites deep with sharpwalls, septa in three cycles extending to center, no columella.
Description.⎯Colony phaceloid, obtusely branching, D � 2.5 � 2.2 cm,fragmented, epitheca subhorizontally folded; corallites circular; walls, sharp,vertical; calices deep (6 mm), d � 6–7 mm, s � 20–38, uniform thickness,differentiated to two or three cycles, granular (Fig. 3.24); S1 thickest, mostpronounced, extend three fourths way to axis; S2 thinner, one half length ofS1; S3 exist along wall.
Etymology.⎯Latin, obtuse to describe the branching angle that distinguish-es it from other species in the genus.
Types.⎯Based on a single silicified holotype, UMIP 400337, from theChitistone Formation, MI 9934 (M1708), Wrangell Mountains, south-centralAlaska. Paratypes: None.
Discussion.⎯This species resembles R. parviseptum (Squires)but differs with an obtuse branching pattern. Retiophyllia parvi-septum further contains spiral budding, which is not observed inour new species.
Subfamily CORYPHYLLIINAE Beauvais, 1981Genus KUHNASTRAEA Cuif, 1976
Type species.⎯Phyllocoenia decussate Reuss, 1854.Remarks.⎯Cuif (1976) and Roniewicz (1989) gave detailed
descriptions based largely on microstructural details and mor-phology. Pseudocerioid integration, large intercorallite space, andring-like structures between epitheca and endotheca are traits,which stand out and were used to identify this genus (Cuif, 1976;Roniewicz, 1989, pl. 18, fig. 1).
KUHNASTRAEA DECUSSATA (Reuss, 1854)(Figure 4.1, 4.2, 4.5)
Phyllocoenia decussata REUSS, 1854, p. 99, pl. 13, figs. 2, 3.(See Roniewicz, 1989; Stanley and Whalen, 1989.)
Description.⎯Colonies plate-like, cerioid; calices circular, regularlyspaced, shallow, walls unshared, d � 5.5–7.0 mm; s � 23–35, differentiatedto three cycles around calice; S1 most pronounced, do not extend to centerleaving open space; S2 extend one half to full length of S1; S3 least pro-nounced; pseudo-ornamentation from granular silicification.
Material examined.⎯Four specimens from the Cornwallis Limestone,Keku Strait, southeast Alaska, MI 0099 (M1911) and MI 0056 (M2135);twelve from the Chitistone Formation, Wrangell Mountains, south-centralAlaska, MI 9934 (M1708).
Occurrence.⎯Rhaetian, Vancouver Island, Canada; Northern CalcareousAlps; Norian, Gravina Island, Keku Strait, Wrangell Mountains, Iliamna Lake,Peninsula, Alaska; Hells Canyon, Oregon; Shasta County, California; PilotMountains, Nevada.
Discussion.⎯Our specimens from Alaska and Canada containvery traditional skeletal morphology corresponding with K. de-cussata (Reuss) as revised in Roniewicz (1989). The pronouncedring-like structure, similar septal attributes, very densely packedcorallites, and little or no intercorallite space (Fig. 4.2, 4.5) matchvery well with material from Stanley and Whalen (1989, figs. 5.5,5.15, 7.1).
KUHNASTRAEA INCRASSATA (Frech, 1890)(Figure 4.3, 4.4)
Phyllocoenia incrassata FRECH, 1890, pl. 8, figs. 1–14.Isastrea whiteavesi CLAPP AND SHIMER, 1911, pl. 40, fig. 10.(See Roniewicz, 1989; Stanley and Whalen, 1989.)
Description.⎯Colonies subphaceloid to cerioid, calices oval, deep, d �5.5–7.0 mm, regularly spaced around stalk leaving large intercorallite space;s � 23–35, differentiated to three cycles around calice; S1 most pronounced,do not extend to center leaving open space, ornamented lateral surfaces (Fig.4.4); S2 extend one half to full length of S1; S3 least pronounced.
Material examined.⎯Three specimens from Lake Cowichan, VancouverIsland, Canada, MI 8302.
Occurrence.⎯Rhaetian, Vancouver Island, Canada; Norian-Rhaetian,Northern Calcareous Alps; early Norian, Hells Canyon, Oregon.
Discussion.⎯Despite having slightly larger corallite diameter,our material matches very well with K. incrassata (Frech) as re-vised in Roniewicz (1989). Evaluation of silicified material fromClapp and Shimer (1911) and non-type specimens from Stanleyand Whalen (1989) reveals a definite presence of this species inthe North American Cordillera. Clapp and Shimer (1911, pl. 40,fig. 10) mistakenly identified the Alpine variety as a new speciesIsastrea whiteavesi.
KUHNASTRAEA COWICHANENSIS (Clapp and Shimer, 1911)(Figure 4.6, 4.9, 4.10)
Isastrea cowichanensis CLAPP AND SHIMER, 1911, p. 429, pl. 41, fig. 11.(See Stanley and Whalen, 1989.)
Description.⎯Colonies mound shaped, platy; D � 3 � 4 cm; corallitescircular, oval; walls unshared, variable intercorallite space forms ring-likestructure (Fig. 4.10), prolonged costae tend to meet, do not fuse; calices deep,sloping, 1–2 mm, d � 3–4 mm, no columella; s � 20–30 differentiated to

480 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
FIGURE 4—1, 2, 5, Kuhnastraea decussata (Reuss), UMIP 400328 (MI 9934); 1, 2, colony with dense regular arrangement of corallites and ring-likestructures, scale bar on 1 is 10 mm; 5, close up of calice. 3, 4, Kuhnastraea incrassata (Frech) UMIP 21065 (MI 8302); 3, surface view of colony withabundant intercorallite space and thin epitheca, scale bar 10 mm; 4, close up of 3 showing well preserved calice and septa. 6, 9, 10, Kuhnastraea cowichanensis

481CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
←
(Clapp and Shimer), UMIP 400188 (MI 0099); 6, regular arrangement of corallites in surface view, scale bar 10 mm; 9, a single calice with abundantornamentation along septal surfaces; 10, surface view showing excellent preservation, arrangement and evident ring-like structure. 7, 8, 12, Margarosmiliacf. M. charlyana (Frech), two specimens UMIP 400335 (MI 9934); 7, close up of septa and well preserved pinnular ornamentation along lateral surfaces; 8,surface view showing distinct sub-phaceloid integration and well defined corallites, scale bar 10 mm; 12, lateral view of another specimen with thin, foldedepitheca and fused lower-portion of colony, scale bar 10 mm. 11, 15, Margarosmilia cf. M. richthofeni Volz, UMIP 400192 (MI 0099); 11, surface view ofirregular shaped interconnected corallites, scale bar 10 mm; 15, view of thin, well preserved septa and sporadic dissepiments. 13, 14, Margarosmilia confluens(Munster), UMIP 400334 (MI 9934); 13, calical view of well preserved lateral ornamentation on septal surfaces, scale bar 10 mm; 14, sub-phacelloid colonywith thin epitheca and well-preserved calices. 16, 20, Distichomeandra austriaca (Frech), latex mold UMIP 226955 (MI 9913); 16, surface view with sharp,distinct, walls and well defined pseudomeandroid corallites, scale bar 10 mm; 20, close up of distinct corallite shape and well preserved septa. 18, 19,Distichomeandra minor (Frech), UMIP 228270 (MI 0099); 18, adjacent corallites with sharp walls and septa extending to axial fissure; 19, colony withpseudomeandroid integration, scale bar 10 mm. 17, 22, Astraeomorpha crassisepta Reuss; 17, UMIP 228358 (MI 0087) lateral view of well preserveddissepiments between adjacent septa, scale bar 10 mm; 22, UMIP 400546 (MI 0099) well preserved, disorderly shared septa, occasionally joined formingtriple junction (arrow), scale bar 5 mm. 21, 25, Astraeomorpha confusa (Winkler), UMIP 221066 (MI 9947); 21, surface view with characteristic corallitearrangement; 25, close up of thin, curved, septa, forming columella, scale bar 10 mm. 26, 27, Parastraeomorpha cf. P. similis Roniewicz, UMIP 21092 (MI8302); 26, colony with poorly preserved corallites, scale bar 5 mm; 27, close up of distinct septa and shallow calices. 23, 24, Chondrocoenia schafhaeutli(Winkler), UMIP 400643 (MI 8302); 23, well preserved corallites regularly arranged with evident columella; 24, close up showing distinct septa confluent,thin corallite wall and button-like columella, scale bar 10 mm. 28, 29, Chondrocoenia cf. C. paradoxa (Melnikova), UMIP 400525 (MI 9934); 28, 29 surfaceviews showing confluent thin S1 septa extending to central axis, scale bar on 28 is 10 mm.
three or four cycles; S1 club-shaped, protrude one half to three fourths radius;S2 shorter, thinner, one half length of S1; S3 thin, against wall; S1 and S2have spiny ornamentation parallel to central axis (Fig. 4.9).
Material examined.⎯Six specimens from the Cornwallis Limestone, KekuStrait, southeast Alaska, MI 0099 (M1911), MI 0056 (M2135), MI 0070; fourspecimens from the Chitistone Formation, south-central Alaska, MI 9934(M1708); and two from the Parson Bay Formation, Lake Cowichan, Vancou-ver Island, Canada, MI 8302.
Occurrence.⎯Rhaetian, Vancouver Island, Canada; early Norian, GravinaIsland, Keku Strait, Wrangell Mountains, Alaska; Hells Canyon, Oregon; Ko-ryak terrane, Siberia.
Discussion.⎯The type specimen, Isastrea cowichanensis Clappand Shimer (1911), is vaguely described and adversely affectedby over-silicification, providing little or no detail in regards toseptal attributes and corallite morphology, which precludes tax-onomic understanding. However, several authors illustrated anddescribed specimens identified as, or in synonymy with, K. cow-ichanensis Clapp and Shimer from several Cordilleran terrane lo-calities (Smith, 1927; Montanaro-Gallitelli et al., 1979; Melnikovaand Bychkov, 1986; Stanley and Whalen, 1989, fig. 5.1), whichwe used to identify our material.
Family MARGAROPHYLLIIDAE Volz, 1896Genus MARGAROSMILIA Volz, 1896
Type species.⎯Montlivaltia zieteni KLIPSTEIN, 1843.
MARGAROSMILIA CHARLYANA (Frech, 1890)(Figure 4.7, 4.8, 4.12)
Thecosmilia charlyana FRECH, 1890, p. 11, pl. 3, figs. 5–5B, pl. 5, figs.6–7A.
Margarosmilia charlyana (Frech) RONIEWICZ, 1989, p. 77, pl. 19, figs. 1–4,pl. 22, figs. 3, 4; STANLEY, 1994, pl. 5, figs. 6–8, text-fig. 10; MELNIKOVA,2001, p. 53, pl. 16, fig. 2.
Description.⎯Colonies subphaceloid, D � 3.5 � 2.7 � 1.2 cm, growingfrom central point of attachment, subhorizontally folded epitheca, individualbranches fused except for calice; budding extracalicularly, flat, slightly ele-vated, giving subphaceloid appearance (Fig. 4.8). Corallites circular, d � 10–13 mm, s � 43–49 (rarely 30 or 60), equally thick, differentiated into fourcycles around calice, no columella. S1 and S2 most prominent, extendingradius, containing numerous, vertically oriented, jagged, pinnular structuresalong lateral surfaces (Fig. 4.7); S3 and S4 shorter, reaching one fourth toone half length of S1 septa.
Material examined.⎯Four colonies from the Chitistone Formation, Wran-gell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; Norian, PucaraGroup, central Peru; Norian-Rhaetian, Northern Calcareous Alps.
Discussion.⎯Roniewicz (1989) and Stanley (1994) describedthis species with great morphological detail, paying particular at-tention to the fused, closely spaced, multi-branching bifurcatingor trifurcating nature of the colony with sunken calices, whichour material highly resembles, but with larger corallites.
Roniewicz (1989) noted M. zieteni (Klipstein) (adult individu-al) figured in Cuif (1975b) to have diameters 13�16 mm, which
match perfectly with our specimens, suggesting similar morphol-ogies between M. zieteni (Klipstein) and M. charlyana (Frech).Our material also superficially resembles a specimen illustratedby Laube (1865, pl. 5, fig. 4) as Thecosmilia rugosa Laube butwith a much larger colony size.
MARGAROSMILIA cf. M. RICHTHOFENI Volz, 1896(Figure 4.11, 4.15)
Margarosmilia richthofeni VOLZ, 1896, pl. 1, figs. 15, 16.
Description.⎯Colony mound-shaped, polycentric, D � 4.5 � 2.7 cm, thinepitheca. Corallites irregular, slightly flabellate, interconnected, d � 20–25mm, s � 112 and 144, septa differentiated into five cycles. S1 thickest; S1,S2 and S3 extend to central fissure, occasionally bending, dissepiments occursporadically.
Material examined.⎯A single silicified colony from Keku Strait, southeastAlaska, MI 0099 (M1911).
Occurrence.⎯Early Norian, Keku Strait, Alaska; Carnian, Dolomite Alps.Discussion.⎯This assignment is tentative, based only on one
specimen presenting two moderately well preserved corallites thatreveal similarities with Volz’s (1896, pl. 1, figs. 15, 16) species.We here emend the diagnosis of the genus by Roniewicz (1989);specifically its polycentric integration, numerous septa, and link-ages (dissepiments) along the endotheca. However, our specimendoes not possess prominent sharp spines, which is a distinguishingfactor of the genus.
MARGAROSMILIA CONFLUENS (Munster, 1841)(Figure 4.13, 4.14)
Cyathophyllum confluens MUNSTER, 1841, pl. 2, fig. 16.Montlivaltia zieteni KLIPSTEIN, 1843, p. 289, pl. 20, fig. 1a–b.Montlivaltia zieteni (Klipstein) VOLZ, 1896, pl. 1, figs. 1–7, text-fig. 18; CUIF,
1974, fig. 29.Synastrea zieteni D’ORBIGNY, 1849, p. 208.Thamnastrea goldfussi EDWARDS AND HAIME, 1851, p. 113.?Paradistichophyllum minimum MELNIKOVA, 1975, p. 39, pl. 17, fig. 2.?Thecosmilia minima MELNIKOVA, 1986, p. 28, pl. 2, fig. 4.?Retiophyllia minima ILJINA AND MELNIKOVA, 1986, p. 50, pl. 14, fig. 1, pl.
15, figs. 4, 5.Margarosmilia minima (Melnikova) MELNIKOVA, 2001, p. 54–55, pl. 15,
fig. 4.
Description.⎯Colonies dendroid, D � 2.6 cm; branching acutely, fused,longitudinal section exposed by thin or missing epitheca, dissepiments rare;corallites circular, tapering, d � 5–6 mm, s � 22–36 differentiated to threeor four cycles; S1, S2 extend to central axis, no columella, display ornamen-tation along lateral septal surfaces (Fig. 4.13); S3 (S4 rare) thin, less pro-nounced, extend one fourth to one half distance of S1.
Material examined.⎯Five silicified fragments from the Chitistone For-mation, Wrangell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Early Norian, Wrangell Mountains Alaska; Rhaetian, Pa-mirs, central Iran, northeastern Caucasus.
Discussion.⎯General morphology matches the emended diag-nosis of the genus by Roniewicz (1989) with respect to phaceloid

482 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
integration and ornament of sharp spines along septa; however,corallite diameters are very small when compared to material cit-ed in Roniewicz (1989; referring to material figured in Cuif,1975b). Our material further resembles illustrations in Volz (1896,pl. 1, figs. 1–7) in colony size, fused branches, corallite diameter,thin septa (S1, S2 extending to center), and no columella. Ron-iewicz (1989) placed material from Volz in synonymy with M.confluens (Munster) based on microstructural similarities.
Genus DISTICHOMEANDRA Cuif, 1976
Type species.⎯Distichomeandra spinosa CUIF, 1976.
DISTICHOMEANDRA AUSTRIACA (Frech, 1890)(Figure 4.16, 4.20)
Isastraea austriaca FRECH, 1890, p. 23, pl. 6, figs. 1–3, 5, 6, (non pl. 6,fig. 4).
Isastraea eucystis FRECH, 1890, p. 26, pl. 6, figs. 10, 10A (non pl. 7, figs.11, 11a, 12, 12A).
‘‘Isastraea eucystis’’ (Frech) MATZNER, 1986, pl. 9, fig. 7.Margarastraea eucystis (Frech) STANLEY, 1979, p. 56, pl. 1, fig. 2.Distichomeandra austriaca (Frech) RONIEWICZ, 1989, p. 79, pl. 20, figs. 1,
6, pl. 21, figs. 5, 6, pl. 22, fig. 1; STANLEY, 1994, p. 12, fig. 9.1, 9.2, 9.10,9.11.
Distichomeandra cf. austriaca (Frech) STANLEY AND SENOWBARI-DARYAN,1999, p. 794, fig. 5.7–5.10.
Description.⎯Colonies nonsilicified and silicified, flat, laminar, D �10�15 cm (largest colony), pseudomeandroid; corallites interconnected, me-ander, long and sinuous with scattered, well defined calices separated by sharpdistinct walls, c-c � 3–6 mm, s � 7–9 (in 3 mm), 11–14 (in 5 mm), differ-entiated into two cycles; S1 and S2 equally thick, moderately well preserved,and extend to fissure bending to define each calice.
Material examined.⎯A single poorly preserved silicified specimen (notillustrated) from the Cornwallis Limestone, Keku Strait, southeast Alaska, MI0056 (M2135); two nonsilicified specimens (one illustrated) from the ParsonsBay Formation in Tahsis Inlet, Vancouver Island, Canada, MI 9913.
Occurrence.⎯Early Norian, Keku Strait, Alaska; Pilot Mountains, Nevada;Sonora, Mexico; late Norian to Rhaetian, Tahsis Inlet, Vancouver Island; No-rian, Quesnellia reef, British Columbia, Canada; Lime Peak, Yukon Canada;Northern Calcareous Alps.
Discussion.⎯Samples match well with pseudomeandroid spec-imens illustrated in Frech (1890), Roniewicz (1989, pl. 20, fig. 1)and Yarnell (2000, pl. 3, fig. 1). Stanley et al. (1994) pointed outthat this species has both pseudomeandroid and cerioid integra-tion.
DISTICHOMEANDRA MINOR (Frech, 1890)(Figure 4.18, 4.19)
Isastraea norica var. minor FRECH, 1890, p. 26, pl. 6, fig. 9.Distichomeandra minor (Frech) RONIEWICZ, 1989, p. 81, pl. 20, figs. 2–5, pl.
21, figs. 1, 2.
Description.⎯Colonies laminar, mound-shaped, D � 3.3 cm, cerioid andpseudomeandroid; corallites circular, oval and slightly polygonal, well definedwalls, d � 5–8 mm, s � 38–48, confluent, extend to center; papillate colu-mella, differentiated to four cycles, subequal thickness. S1 and S2 equallength, extend to central axis or fissure; S3 one half length of S1; S4 small,only visible along endotheca.
Material examined.⎯Two silicified samples from the Cornwallis Lime-stone Keku Strait, southeast Alaska, MI 0099 (M1911).
Occurrence.⎯Early Norian, Keku Strait, Alaska; Rhaetian, Northern Cal-careous Alps.
Discussion.⎯Roniewicz (1989) noted that D. minor differsfrom D. austriaca by smaller, more lobate calices containing moreabundant septa of subequal thicknesses and larger dissepimentsconnecting them. Distichomeandra minor also resembles Mean-drostylis irregularis Frech in corallite size and shape; however,M. irregularis Frech has two elongate septa forming parallel toeach other along the central fissure of each meandering corallite(Roniewicz, 1989, pl. 39, figs. 10a, b). This new occurrence inthe Alexander terrane constitutes the first occurrence outside theformer Tethys region.
DISTICHOMEANDRA sp. A(Figure 5.15, 5.20)
Description.⎯Colonies mound shaped, D � 5 cm; corallites highly vari-able, cerioid to pseudomeandroid, budding frequently; calices sub-sphericaland oval shaped, deep (3 mm), d � 2.5–10 mm (most between 6–10 mm), s� 15–44, well preserved, differentiated to two or three cycles around center,no columella; S1 longest, most pronounced, wedge shaped thickening towardwall, granulations on lateral surfaces (Fig. 5.15; most prominent on S2 andS3) indicative of genus, curved toward axis; S2 thinner, wedge-shaped, onehalf to three quarters length of S1; S3 one quarter distance of S1.
Material examined.⎯Nine colonies from the Chitistone Formation, Wran-gell Mountains, south-central Alaska, MI 9934 (M1708); one colony from theCornwallis Limestone, Keku Strait, southeast Alaska, MI 0099 (M1911).
Occurrence.⎯Early Norian, Wrangell Mountains, Keku Strait, Alaska.Discussion.⎯Pronounced granulations on lateral septal sur-
faces, in conjunction with cerioid corallites, help place this coralwithin genus Distichomeandra Cuif (Roniewicz, personal com-mun., 2007). It cannot be confused with Elysastrea fischeri Laube,with which it shows similar gross morphology including coralliteshape, measurements, and septal arrangement; E. fischeri, on theother hand, possesses septal sides with short menianes (Ronie-wicz, personal commun., 2007). Though abundant throughout thisfield area, specimens do not show distinguishing species-levelcharacteristics enabling a new species.
DISTICHOMEANDRA sp. B(Figure 5.12, 5.16)
Description.⎯Branching, multiserial columns, D � 4.8 cm; highly polyg-onal, cerioid to pseudomeandroid corallites; walls sharp, calices moderatelydeep, d � 3–5 m. s � 27–36 pseudo confluent, occasionally aligned betweenadjacent corallites, but not completely; poorly preserved granulations on lat-eral surfaces; preserved to three cycles. S1 extend to axis form papillate col-umella, most pronounced; S2 equal thickness, extend one half to total lengthof S1, fuse when length permits; S3 wedge-shaped, project along wall, onefourth length of S1.
Material examined.⎯A single silicified specimen from the Chitistone For-mation, Wrangell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska.Discussion.⎯Pseudoconfluent septa with granulations on lat-
eral surfaces are indicative of genus Distichomeandra Cuif andwarrant its inclusion (Roniewicz, personal commun., 2007). ThisWrangellian coral has measurements ranging just below those ofD. minor (Frech), as well as being highly irregular in colony andcorallite growth form, indicating that it is a separate taxon. Itshows close resemblance to Isastraea haueri Laube var. minorVinassa de Regny (1915, pl. 71, figs. 5, 6). Because we have onlyone specimen, we refrain from making a new species until better-preserved specimens can be recovered. However, we wish to pointout the similarities with material from Timor and the Dolomites.
Suborder FUNGIINA Verrill, 1865, faute de mieuxSuperfamily PROCYCLOLITOIDEA Vaughan and Wells, 1943
Family ASTRAEOMORPHIDAE Frech, 1890Genus ASTRAEOMORPHA Reuss, 1854
Type species.⎯Astraeomorpha crassisepta Reuss, 1854.
ASTRAEOMORPHA CRASSISEPTA Reuss, 1854(Figure 4.17, 4.22)
Astraeomorpha crassisepta REUSS, 1854, p. 127, pl. 16, figs. 4–7.(See RONIEWICZ, 1989; STANLEY AND WHALEN, 1989; MELNIKOVA, 2001)
Description.⎯Colony thamnasteroid, flat, laminar to mound shaped, hemi-spherical, columnar; corallites densely packed, disorderly, c-c � 1.8–3.5 mm,s � 12–18, thick (0.5 mm), differentiated to two cycles, columella button-like; S1, S2 shared, equal thickness, S1 longer, S2 occasionally fusing withadjacent S2 forming triple-junction (Fig. 4.22, arrow). Longitudinal viewyields dissepiments along epitheca (Fig. 4.17).
Material examined.⎯Eight specimens from the Cornwallis Limestone,Keku Strait, southeast Alaska, MI 0087 (M1912), MI 0056 (M2135); fourfrom the Nehenta Formation, Gravina Island, southeast Alaska, MI 9935;fourteen from the Chitistone Formation, Wrangell Mountains, south-centralAlaska, MI 9934 (M1708); thirteen from the Parson Bay Formation, LakeCowichan Vancouver Island, Canada, MI 8302.

483CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
Occurrence.⎯Wide-ranging and abundant species. Norian, Keku Strait,Gravina Island, Wrangell Mountains, Alaska; Hells Canyon, Oregon; Lewis-ton, Idaho; Pucara Group, northern Peru; Northern Calcareous Alps andSouthern Alps, Austria and Italy; Tatra Mountains, Poland; middle Afghani-stan; central Iran; northern Caucasus Mountains, Pamir Mountains, and Ko-ryak Highland, Russia.
Discussion.⎯Roniewicz (1989) described in detail this wellknown Upper Triassic species, indicating its worldwide distribu-tion during Norian to Rhaetian time. Squires (1956) mistakenlyassigned a new species to this genus that cannot be distinguishedfrom the type material. Thamnastraea borealis Smith (1927) wasplaced in synonymy with A. crassisepta Reuss by Montanaro-Gallitelli et al. (1979) and Stanley and Whalen (1989); our spec-imens match well with silicified material of Squires (1956), Mon-tanaro-Gallitelli et al. (1979) and Stanley and Whalen (1989).
ASTRAEOMORPHA CONFUSA (Winkler, 1861)(Figure 4.21, 4.25)
Thamnastraea confusa WINKLER, 1861, p. 488, pl. 8, fig. 10. (See MELNI-KOVA, 2001; STANLEY, 1994.)
Description.⎯Colony thamnasteroid, laminar; corallites shallow, regularlyarranged in rows, c-c � 2–3 mm, s � 12–15, equally thick, unfused, sym-metrical, differentiated to three cycles, button-like columella (Fig. 4.25); S1and S2 extend between columella; S1 bend toward center; S2 straight; S3situated between calices, do not extend entire distance.
Material examined.⎯A single silicified colony from the Nehenta Forma-tion, Gravina Island, southeast Alaska, MI 9935; one from the Parson BayFormation, Lake Cowichan Vancouver Island, Canada, MI 8832.
Occurrence.⎯Norian, Gravina Island, Alaska; Alps; Caucasus; centralAsia; Timor; Rhaetian, Vancouver Island, Canada.
Discussion.⎯Astraeomorpha confusa (Winkler) is well docu-mented in European and North American localities (Montanaro-Gallitelli et al., 1979; Roniewicz, 1989; Stanley and Whalen,1989), closely resembling A. crassisepta Reuss in c-c, septal ar-rangement, colony integration, and growth form. Astraeomorphaconfusa (Winkler) differs from A. crassisepta by having corallitesarranged in rows with thinner, symmetrical, and denser septa thanA. crassisepta Reuss. Roniewicz (1989) noted difficulties distin-guishing A. crassisepta from A. confusa, confirming a close re-lationship between the two species.
Genus PARASTRAEOMORPHA Roniewicz, 1989Type species.⎯Parastraeomorpha minuscula RONIEWICZ,
1989.
PARASTRAEOMORPHA cf. P. SIMILIS Roniewicz, 1989(Figure 4.26, 4.27)
Parastraeomorpha similis RONIEWICZ, 1989, p. 99, pl. 30, fig. 3; MELNIKOVA,2001, p. 62, pl. 24, fig. 1.
Parastraeomorpha cf. P. similis Roniewicz STANLEY, 1994a, p. 84, pl. 2, figs.7–8.
Astraeeomorpha confusa var. minor (Frech) STANLEY, 1979, pl. 1, fig. 8.
Description.⎯Colony ?columnar or laminar, subcerioid to thamnasteroid;corallite wall poorly developed; calices disorderly distributed, d � 1.5–2.2mm, s � 12–18, shared, slightly elevated, differentiated to three cycles; S1,S2, equal thickness, extend to center, S1 meet forming small, poorly devel-oped, easily mistakable columella; S3 rare, project along pseudo wall.
Material examined.⎯Two colonies from the Parson Bay Formation, LakeCowichan, Vancouver Island, Canada, MI 8302.
Occurrence.⎯Norian-Rhaetian, Northern Calcareous Alps; Norian, PamirMountains; Rhaetian, Vancouver Island, Canada; Norian, Pucara Group, cen-tral Peru.
Discussion.⎯Dimensions, septal arrangement and disorderlydistribution of corallites resemble type material from Roniewicz(1989, pl. 30, fig. 3) and non-type material from Stanley (1994b,pl. 2, figs. 7–8); however, poor preservation precludes definitiveidentification. Stanley (1994) noted a close relationship with P.minuscula Roniewicz, differentiated only through smaller calicaldiameters. Parastraeomorpha similis Roniewicz was reported byStanley (1994a) from South America (Peru), and Stanley (1979)reported and illustrated a small columnar growth form informally
identified as Astraeeomorpha confusa var. minor (Frech) from theLake Cowichan site on Vancouver Island, which is possibly as-signable to this species.
Suborder ARCHAEOCOENIINA Alloiteau, 1952Family ACTINASTRAEIDAE Alloiteau, 1952Genus CHONDROCOENIA Roniewicz, 1989
Type species.⎯Prionastraea schafhaeutli WINKLER, 1861.
CHONDROCOENIA SCHAFHAEUTLI (Winkler, 1861)(Figure 4.23, 4.24)
Prionastraea ? schafhautli WINKLER, 1861, p. 488, pl. 8, fig. 11.(See STANLEY, 1986; RONIEWICZ, 1989; MELNIKOVA, 2001.)
Description.⎯Colony cerioid, multiserial, columnar; corallites circular,walls weakly defined; columella button-like, calices shallow, c-c � 2.5–3.0mm, s � 27–34, nonconfluent, well preserved, differentiated to three cycles.S1 thickest, extend radius, fuse with columella; S2 three fourths radius, thin-ner; S3 one fourth to one half length of S1, occasionally same thickness asS2.
Material examined.⎯Three fragments from the Parson Bay Formation,Lake Cowichan, Vancouver Island, Canada, MI 8302.
Occurrence.⎯Very wide-ranging species. Rhaetian, Lake Cowichan, Van-couver Island; Northern Calcareous Alps, southern Alps (Lombardy), and theCarpathian Mountains; Norian, Lewiston, Idaho; Pamir Mountains, Russia;and the Pucara Group, Peru.
Discussion.⎯Corallite dimensions and general morphologymatch very well with specimens from Stanley (1986, 1994b),Roniewicz (1989), and Melnikova (2001). However, our speci-mens exhibit multiserial columnar growth, whereas most otherspecimens are either lamellate or hemispherical. Roniewicz(1989) mentioned that colonies occasionally branch, reaching upto 20 mm in diameter and 70 mm in length.
CHONDROCOENIA cf. C. PARADOXA (Melnikova, 1968)(Figure 4.28, 4.29)
Cyathocoenia paradoxa MELNIKOVA, 1968, p. 16, pl. 3, fig. 1, 2; 1975, p.61, pl. 2, figs. 1–3, 6, pl. 3, fig. 1; STANLEY AND WHALEN, 1989, p. 814,fig. 6.3.
Chondrocoenia paradoxa (Melnikova) MELNIKOVA, 2001, pl. 28, fig. 2.
Description.⎯Colony fragmented, laminar; corallites regularly spaced, cir-cular and polygonal; columella poorly defined or papillate; walls poorly de-fined and unshared; calices shallow, d � 3–4 mm, s � 14–22, differentiatedto two cycles; S1 most pronounced, extend radius, S2 thinner, visible alongendotheca.
Material examined.⎯A single silicified colony from the Chitistone For-mation, Wrangell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; Hells Canyon,Oregon; Norian-Rhaetian, Pamir Mountains.
Discussion.⎯Our specimen is identical to Cyathocoenia par-adoxa Melnikova in Stanley and Whalen (1989, fig. 6.3).
Suborder ASTRAEOINA Alloiteau, 1952Family PAMIROSERIIDAE Melnikova, 1984
Genus PAMIROSERIS Melnikova, 1971
Type species.⎯Thamnastraea meriani STOPPANI, 1858.
PAMIROSERIS MERIANI (Stoppani, 1858)(Figure 5.1, 5.2, 5.6, 5.7)
Thamnastraea meriani STOPPANI, 1858, p. 108, pl. 26, figs. 3–6.(See STANLEY AND WHALEN, 1989; MELNIKOVA, 2001.)
Description.⎯Colony thamnasteroid, flat, mound-shaped, occasionally us-ing spongiomorphs or other organisms as substrate; corallites slightly elevat-ed, circum-oral (fungiform), extracalicular budding, corallites grow separately,fuse with colony (three new; Fig. 5.2, 5.6). Calices shallow, c-c � 5–8 mm,s � 23–29 (max. 39) in four cycles; S1 (sometimes S2) shared, blade-like,extend to axis, thicker in middle (0.5 mm), confluent thinning between cor-allites, occasionally bending, fuse to form columella (poorly developed); S2extend length of S1, thinner, do not form columella; S3 one half to threefourths length of S1, same thickness as S2; S4 (rare) only along periphery.
Material examined.⎯Eight specimens from the Chitistone Formation,Wrangell Mountains, south-central Alaska, MI 9934 (M1708); three from the

484 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
FIGURE 5—1, 2, 6, 7, Pamiroseris meriani (Stoppani), UMIP 302898 (MI 0099); 1, close up of distal septa thinning between adjacent calices; 2, 6, 7surface views with distinct budding of new corallites on surface and thamnasteroid integration, scale bar on 6 is 10 mm. 4, 5, Pamiroseris borealis (Smith),UMIP 400498 (MI 9931); 4, close up of convex corallites with confluent, exert thin septa shared; 5, multi-serial colony with regularly spaced corallites, scale

485CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
←
bar 10 mm. 9, 13, Crassistella juvavica (Frech), UMIP 28362 (MI 0087); 9, well preserved surface view, polygonal corallites with sharp walls, scale bar 10mm; 13, corallites with well defined walls and septa forming button-like columella. 10, 11, Crassistella parvula (Melnikova), UMIP 303087 (MI 0099); 10,densely packed and well preserved corallites with moderately deep calices and confluent septa; 11, plate-like colony with multi-serial column, scale bar 10mm. 3, 8, ?Ampakabastraea cf. A. nodosa Cuif, UMIP 400524 (MI 9934); 3, 8, fragment with well preserved corallites, scale bar 10 mm. 15, 20, Disticho-meandra sp. A, UMIP 400308 (MI 9934); 15, view of unique S1 septa and septal arrangement aligned along shared wall of adjacent corallites; 20, mound-shaped colony with well preserved corallites, scale bar 10 mm. 12, 16, Distichomeandra sp. B, UMIP 400309 (MI 9934); 12, unique multi-serial, branching,columnar growth; 16, corallites well preserved septa extending to central axis forming papillate columella and wall structures, scale bar 10 mm. 19, 25,Recticostastraea wallowaensis Stanley and Whalen, UMIP 221125 (MI 8302); 19, small, lamellate, colony fragments with parallel and subparallel meanderingcorallites; 25, colony with moderately thick epitheca evident along ventral portion of colony, scale bar 10 mm. 14, 17, 18, 21, 22, Campesteria prolixia n.sp., UMIP 222068 (MI 8302); 14, septal view of ornamentation on lateral surfaces; 17, low-lying ridge between adjacent meanders, note S1 and S2 septaextending entire length; 18, radial septa forming individual calice, note columella, scale bar 10 mm; 21, 22, well-defined meander illustrated by convergingsepta along axial fissure, bending to accentuate. 30, 32, Stylophyllum cf. S. pygmaeum Frech, UMIP 228226 (MI 0099); 30, calice with incompletely joined,beaded septa, seemingly not well preserved, scale bar 10 mm; 32, lateral view of distinct branching pattern and thick folded epitheca, scale bar 10 mm. 28,29, Maeandrostylis grandiseptus Stanley and Whalen, UMIP 228272 (MI 0099); 28, multi-serial fragment with well preserved corallites, scale bar 10 mm;29, corallites with deep calices and shared septa. 23, 24, 26, Maeandrostylis vancouverensis (Clapp and Shimer), UMIP 400137 (MI 0087); 23, longitudinalsection with evident analogical spines; 24, surface view of regularly spaced corallites with deep calices, sharp well defined walls, scale bar 10 mm; 26, closeup of 24 with confluent septa only around the periphery of each calice. 27, 31, Anthostylis cf. A. acanthophora (Frech), UMIP 400716 (MI 8302); 27, multi-serial colony surface view with evident moderately well preserved polygonal walls, scale bar 10 mm; 31, view of confluent septa and deep calices.
Cornwallis Limestone, Keku Strait, southeast Alaska, MI 0099 (M1911) andMI 0056 (M2135).
Occurrence.⎯Early Norian, Keku Strait, Wrangell Mountains, Alaska;Hells Canyon, Oregon; Southern Dolomite Alps, northern Italy; southeasternPamirs; Rhaetian, Lombardy, Italy.
Discussion.⎯Stoppani (1858) originally described the speciesto which Melnikova (1971) placed in genus Pamiroseris Melni-kova. Later, Roniewicz (1989) gave morphological and micro-structural characteristics, unique to the genus, indicating pearl-like ornamentation (or granulation) along septal surfaces, partialcolumella, and abundant dissepiments along the endotheca.
Our specimens are identical to material from the Wallowa ter-rane, Hells Canyon, in Montanaro-Gallitelli et al. (1979, pl. 1, fig.7) and Stanley and Whalen (1989, figs. 6.1, 6.2, 6.5, 6.11). Mon-tanaro-Gallitelli et al. (1979) described Pamiroseris meriani(Stoppani) as easily recognizable, based on thamnasteroid inte-gration and circum-oral budding.
PAMIROSERIS BOREALIS (Smith, 1927)(Figure 5.4, 5.5)
Thamnastraea borealis SMITH, 1927, p. 131, pls. 8–10 (non figs. 6, 7).
Description.⎯Thamnasteroid, multiserial, producing knobby branches, D� 3.9 cm; corallites regularly spaced; shallow calices, c-c � 2–4 mm, s �17–29, confluent and exert, differentiated into three cycles. First cycle extendsto the center, second cycle nearly so while third cycle less developed. Lateralseptal ornamentation, trabecular; no columella but internal border encirclescentral cavity, producing paliform projection processes.
Material examined.⎯Two specimens from the Cornwallis Limestone,Keku Strait, MI 0056 (M2135), seven from the Nehenta Formation, GravinaIsland, MI 9931, southeast Alaska.
Occurrence.⎯Early Norian, Keku Strait, southwest coast of Gravina Is-land, Alaska.
Discussion.⎯Specimens resemble closely the type specimensof Thamnasteria borealis Smith (1927, pl. 115, figs. 8–10) fromGravina Island, southeast Alaska. Our material is better preserved,showing additional features of the septal ornament not visible inSmith’s material. Stanley and Whalen (1989) removed two para-types of Thamnasteria borealis Smith (1927, pl. 115, figs. 6, 7),recognizing that they belonged instead to Kuhnastraea incrassataFrech. Pamiroseris borealis (Smith), as presently revised, appearsendemic to the Alexander terrane and has not been reported else-where.
Genus CRASSISTELLA Roniewicz, 1989Type species.⎯Stephanocoenia juvavica FRECH, 1890.
CRASSISTELLA JUVAVICA (Frech, 1890)(Figure 5.9, 5.13)
Astrocoenia sp. FRECH, 1890, p. 36, text-fig. on p. 36.Isastraea profunda SMITH (not Reuss), part, 1927, p. 128, pl. 114, fig. 2.Isastraea parva SMITH, 1927, p. 128, pl. 114, fig. 9.
Cyathocoenia parva (Smith) MONTANARO-GALLITELLI, RUSSO, AND FERRARI,1979, pl. 1, fig. 6a, 6b.
Cyathocoenia aff. C. carinata (Cuif) MONTANARO-GALLITELLI, RUSSO, AND
FERRARI, 1979, p. 138, pl. 1, figs. 3a-4b.Rhaetiastraea cf. R. vesiculosa (MELNIKOVA in Melnikova and Bychkov)
STANLEY AND WHALEN, 1989, fig. 7.3–7.5.(See Roniewicz, 1989; Melnikova, 2001.)
Description.⎯Colonies laminar, encrusting, massive and hemispherical;corallites cerioid, polygonal, elongate along curves; walls sharp, distinct; col-umella, well defined, styliform; calices shallow, d � 2.0–3.5 mm, s � 15–32, nonconfluent, differentiated to two or three cycles; S1 and S2 fuse tocolumella; S1 thicker, wedge-shaped; S2 thinner; S3 (rare) project along walladhering to lateral surfaces of S1 and S2.
Material examined.⎯Twenty-seven specimens from the Cornwallis Lime-stone, Keku Volcanics, Keku Strait, and the Nehenta Formation, Gravina Is-land, southeast Alaska, MI 0099 (M1911), MI 0087 (M1912), MI 0074(M2136), MI 0056 (M2135), MI 9932; fifteen from the Chitistone Formation,Wrangell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Widely distributed species. Early Norian, Gravina Island,Keku Strait, Wrangell Mountains, Alaska; Hells Canyon, Oregon; Norian,Shasta County, California; Norian-Rhaetian, Northern Calcareous Alps; PamirMountains; and northeast Asia.
Discussion.⎯One of the most abundant species in this study,C. juvavica (Frech) is morphologically identical to illustrations ofCyathocoenia aff. C. carinata Cuif and Cyathocoenia parva(Smith) in Montanaro-Gallitelli et al. (1979, pl. 1, figs. 3, 4, pl.1, fig. 6) and is therefore placed in synonymy, differing withslightly smaller calices in Cyathocoenia parva (Smith). Growthform proved to be highly variable, exhibiting encrusting, lamel-late, and hemispherical growth in different specimens, likely re-flecting ecophenotypic variations to different depositional envi-ronments (Caruthers and Stanley, in press).
CRASSISTELLA PARVULA (Melnikova, 1982)(Figure 5.10, 5.11)
Toechastraea plana parvula DRONOV, GAZDZICKI, AND MELNIKOVA, 1982,p. 119, pl. 16, figs. 6–9; MELNIKOVA, 1986, p. 56, pl. 19, fig. 3.
Crassistella parvula (Melnikova) MELNIKOVA, 2001, pl. 28, fig. 3.
Description.⎯Plate-like, multiserial columnar growth, D � 4 cm; corallitescerioid, polygonal, distorted along curves; walls shared, not well pronounced;columella poorly developed; calices moderately deep, d � 1.5–3.0 mm, s �14–26 differentiated to three cycles, extend to axis, nonconfluent, trabecularwithout preserved ornamentation; S1 extend radius; S2 one half length; S3one fourth length.
Material examined.⎯A single silicified colony from the Cornwallis Lime-stone, Keku Strait, southeast Alaska, MI 0099 (M1911).
Occurrence.⎯Early Norian, Keku Strait, Alaska; Pamir Mountains; easternTethys.
Discussion.⎯Crassistella parvula (Melnikova) has the smallerd and s in comparison to C. vesiculosa (Melnikova in Melnikovaand Bychkov) and C. juvavica Roniewicz, which ranges betweenthe two. Corallite dimension, septal attributes, wall structure, and

486 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
calical depth match C. parvula (Melnikova) illustrated in Melni-kova (2001, pl. 28, fig. 3). This is the first occurrence of thistaxon outside the eastern Tethys region.
Genus AMPAKABASTREA Alloiteau, 1957Type species.⎯Ampakabastraea ampakabensis ALLOITEAU,
1957.
?AMPAKABASTRAEA cf. A. NODOSA Cuif, 1976(Figure 5.3, 5.8)
Ampakabastraea nodosa CUIF, 1976, p. 89, pl. 5, figs. 1–3.Ampakabastraea aff. nodosa Cuif MONTANARO-GALLITELLI, RUSSO, AND
FERRARI, 1979, p. 142, pl. 3, fig. 1.
Description.⎯Colonies small, fragmented, moderately well preserved, D� 1.4 cm; corallites cerioid; calices moderately deep, tapering with sharedwalls, d � 4–6 mm, s � 26–33, differentiated to three or four cycles; S1extend to axis, internal borders encircle the central cavity leaving an openspace; S2 extend to axis, do not fuse; S3 and S4 small, found along periphery.
Material examined.⎯Four fragments from the Chitistone Formation,Wrangell Mountains, south-central Alaska, MI 9934 (M1708).
Occurrence.⎯Early Norian, Wrangell Mountains, Alaska; early Norian,Turkey.
Discussion.⎯Our material matches, and is from, the same lo-cality as those illustrated in Montanaro-Gallitelli et al. (1979, pl.3, fig. 1); however, a tentative identification is given because oflimited material for study, high degree of fragmentation, and nodistinguishing microstructure in comparison to type material fromthe Alps. Our material also only has superficial similarity withillustrated type material in Cuif (1976, pl. 5, fig. 1), differing bythe presence of shared walls and a slight appearance of sharedsepta.
Genus RECTICOSTASTRAEA Stanley and Whalen, 1989Type species.⎯Recticostastraea wallowaensis STANLEY AND
WHALEN, 1989.
RECTICOSTASTRAEA WALLOWAENSIS Stanley and Whalen, 1989(Figure 5.19, 5.25)
Andrazella sp. MONTANARO-GALLITELLI, RUSSO, AND FERRARI, 1979, pl. 5,fig. 2a–c.
Recticostastraea wallowaensis STANLEY AND WHALEN, 1989, p. 811, fig. 6.6,6.7.
Description.⎯Colony laminar, plate-like, low-growing, D � 2 cm; coral-lites meandroid, small, forming series of parallel to subparallel, narrow, sin-uous meanders, c-c � 1.0–1.5 mm; septothecal ridges sharp, s � 11–15 (in3 mm), equally thick, differentiated to two cycles, extend between three quar-ters and the entire length to fissure.
Material examined.⎯Four specimens from the Parson Bay Formation,Lake Cowichan, Vancouver Island, Canada (MI 8302).
Occurrence.⎯Rhaetian, Vancouver Island, Canada; early Norian, HellsCanyon, Oregon.
Discussion.⎯Colony size, number of septa, meander layout,and distance between adjacent ridges is virtually identical to typematerial in Stanley and Whalen (1989, figs. 6.6, 6.7). This speciesis endemic to the North American Cordillera, previously onlybeing recognized from Hells Canyon, Oregon (Wallowa terrane)by both Stanley and Whalen (1989) and Montanaro-Gallitelli etal. (1979).
Genus CAMPESTERIA new genus
Type species.⎯Campesteria prolixia n. sp.Diagnosis.⎯Meandroid to pseudomeandroid colony with very
large corallites, plate-like, without defined walls or ridges; septamade of thick trabeculae, ornamented. Genus differs from Para-cuifia Melnikova and Margarastraea Volz (1896), by the presenceof trabecular tips along distal borders of septa, no corallite wallor ridge separating adjacent meanders, and septa completely in-terconnected.
Description.⎯Meandroid to pseudomeandroid, low-growing laminar col-ony without defined ridges or walls between adjacent meanders. Septa distinct,
composed of thick trabeculae, distal borders ornamented with trabecular tipsunique to the family.
Etymology.⎯Latin, campesteria for relatively flat, low-lying meanders in-dicative of colony shape.
CAMPESTERIA PROLIXIA new species(Figures 5.14, 5.17, 5.18, 5.21, 5.22, 6)
Margarastraea pulchra Montanaro-Gallitelli Russo, and Ferrari STANLEY,1994a, p. 85, pl. 3, figs. 1–3.
Diagnosis.⎯Corallites meandroid, no ridges, walls or epitheca;intercorallite spacing wide. Calices interconnected through abun-dant septal linkages. Septa ornamented with trabecular tips alongdistal portion.
Description.⎯Colony large, irregular meanders and pseudomeanders, lam-inar, D � 13 cm; without well-defined ridges, walls or epitheca (Fig. 6);corallites united along distal edges with intratrabecular linkages between ad-jacent corallites; arranged in linear meanders 10–15 mm apart; calices definedby radially converging septa (Fig. 5.18, 5.21), c-c � 13–15 mm, d � 10 mm.s � 6–9 (in 3 mm) 10–13 (in 5 mm), made of thick trabeculae, well orna-mented (Fig. 5.14) with trabecular tips along distal portion (Roniewicz, per-sonal commun., 2007); converge along fissure bending to accentuate calicesforming columella when radial (Fig. 5.18); differentiated to at least threecycles. S1, S2 prominent, extend entire length between adjacent meanders,terminating at fissure, thinner between meanders (very prominent in Fig. 6.1,6.2); S3 granular, extend three fourths distance to fissure; S4 (rare) highlyresemble S3.
Etymology.⎯Latin, prolixus for the unusually wide intercorallite space andextension of shared septa.
Types.⎯Holotype silicified, partial colony (UMIP 22089, Fig. 5.14, 5.17,5.18, 5.21, 5.22) from the Parson Bay Formation, Lake Cowichan (MI 8302).Paratypes UMIP 26953 (MI 9913) and UMIP 222068 (MI 8302), Figure 6;UMIP 22069 (MI 8302), UMIP 21548, 26942, 26943, 26954, 26943, and26932 (MI 9913) are not figured.
Occurrence.⎯Rhaetian, Lake Cowichan, Tahsis Inlet, Vancouver Island,Canada; Norian Pucara Group, Peru.
Discussion.⎯This coral is relatively abundant at Lake Cowi-chan and Tahsis Inlet, Vancouver Island, superficially resemblingMargarastraea klipsteini Frech in Volz (1896, p. 55, pl. 5, figs.7a–d) and M. deningeri Wilckens (1937, p. 179, pl. 7, fig. 1) withlow-lying meandroid corallites. This species, however, is distinct,having septa made of thick trabeculae showing teeth-like orna-mentation in the form of trabecular tips (unique to this genus).Within the type material of M. klipsteini and M. deningeri, distalseptal borders are smooth, having a midseptal zone of minitra-beculae, never forming thick distal ornamentation (Roniewicz,personal commun., 2007). This species was erroneously describedas Margarastraea pulchra Montanaro-Gallitelli from the PucaraGroup, central Peru, by Stanley (1994a). Reevaluation of this ma-terial shows the same septal ornamentation and definitive colu-mella present in our new species.
Suborder STYLOPHYLLINA Beauvais, 1981Family STYLOPHYLLIDAE Frech, 1890Genus STYLOPHYLLUM Reuss, 1854
Type species.⎯Stylophyllum polyacanthum REUSS, 1854.
STYLOPHYLLUM cf. S. PYGMAEUM Frech, 1890(Figure 5.30, 5.32)
Stylophyllum pygmaeum FRECH, 1890 (RONIEWICZ, 1989, pl. 36, fig. 9, pl.38, fig. 6).
Description.⎯Phaceloid, D � 4.5 � 3.5 cm, budding extracalicularly, fivedistinct buds; epitheca thick, folded; corallites subcircular to oval, d � 10–19 mm; s � 48–60, poorly preserved, irregular, beaded or incomplete, par-tially replaced by pyrite, differentiated to two cycles, extend toward axis, nocolumella.
Material examined.⎯One silicified colony from the Cornwallis Limestone,MI 0099 (M1911) in Keku Strait, southeast Alaska.
Occurrence.⎯Early Norian, Keku Strait, Alaska; Zlambach beds, Alps.Discussion.⎯Our specimen is very similar to illustrations in
Roniewicz (1989, pl. 36, fig. 9, pl. 38, fig. 6). Differing only incolony size, a new species is not warranted. Identification mustawait confirmation with additional specimens.
487CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
FIGURE 6—1, 2, 3, paratypes of Campesteria prolixia n. sp. from two different localities on Vancouver Island; 1, negative impression of colony surfaceUMIP 26953 (MI 9913) scale bar 10 mm; 2, silicified colony UMIP 222068 (MI 8302) showing distinct wide, low lying, ridges and pronounced thinning ofsepta distally between corallites, scale bar 10 mm; 3, artist sketch of the whole colony, showing irregularly encrusting colony. The material includes exampleswhere skeletons of the same species are preserved differently. In 1 the skeleton was dissolved but coral is preserved by moldic porosity. Silicification of theoriginal aragonite (2) results in finer details.
Genus MEANDROSTYLIS (Frech, 1890)Type species.⎯Stylophyllum irregulare FRECH, 1890.
MEANDROSTYLIS GRANDISEPTUS Stanley and Whalen, 1989(Figure 5.28, 5.29)
Coccophylum sp. MONTANARO-GALLITELLI, RUSSO, AND FERRARI, 1979, pl.3, figs. 7, 8.
Maeandrostylis grandiseptus STANLEY AND WHALEN, 1989, figs. 6.9, 7.2, 7.7,7.13.
Description.⎯Colony multiserial, columnar, cerioid, D � 2.5 � 1.2 cm;corallites broad, polygonal, regularly spaced, budding intracalicularly; calicesdeeply sunken, d � 6–8 mm, s � 26–39, moderately well preserved, massiveand exert, differentiated into four cycles extending to center, no columella;S1 and S2 best preserved, extend one half to three fourths radius; S3 onlyalong wall; S4 (rare) not well defined.
Material examined.⎯Two partial colonies from the Cornwallis Limestone,Keku Strait, southeast Alaska, MI 0099 (M1911).
Occurrence.⎯Early Norian, Keku Strait, Alaska; Hells Canyon, Oregon.Discussion.⎯Our specimens are identical to type material by
Stanley and Whalen (1989, figs. 6.9, 7.2, 7.7, 7.13), particularlywith respect to irregularly growing corallum, exceptionally largecalices and lack of a columella. Corallite diameters and septacount are on the higher end for this species; nevertheless they stillfall into the acceptable range established by Stanley and Whalen(1989).
MEANDROSTYLIS VANCOUVERENSIS (Clapp and Shimer, 1911)(Figure 5.23, 5.24, 5.26)
Isastraea vancouverensis CLAPP AND SHIMER, 1911, pl. 40, fig. 8, pl. 42, fig.17.
Isastraea vancouverensis (Clapp and Shimer) SMITH, 1927, pl. 112, figs. 1, 2.Isastraea profunda (Reuss) SMITH, 1927, figs. 5, 6.Elysastrea profunda (Reuss) SQUIRES, 1956, fig. 48.Gablonzeria profunda (Reuss) STANLEY, 1986, pl. 3.1, fig. 14.Chondrocoenia schafhaeutli (Winkler) STANLEY, 1994a, pl. 3, figs. 5, 6 [non
figs. 7, 8].
Description.⎯Colony laminar, encrusting, D � 11 cm, intracalicular bud-ding; walls steep, thin, semi-polygonal; endotheca tabuloid with analogicalspines projecting from internal septal borders (Fig. 5.23); calices deep, open,d � 1.5–4.0 mm, s � 15–28. Thick trabecular projections forming confluentsepta preserved to two cycles; S1 and S2 small, granular, equal thickness andlength, only along periphery.
Material examined.⎯Four specimens from the Cornwallis Limestone,Keku Strait, southeast Alaska, MI 0087 (M1912); one from the Nehenta For-mation, Gravina Island, southeast Alaska, MI 9935, MI 9933; twenty fromthe Parson Bay Formation, Lake Cowichan, Vancouver Island, Canada, MI8302.
Occurrence.⎯Rhaetian, Lake Cowichan, Vancouver Island, Canada; EarlyNorian, Keku Strait, Gravina Island, Alaska; Norian, Shasta County, Califor-nia; Norian, the Pucara Group, Peru; and Lewiston, Idaho.
Discussion.⎯Species are reassigned to genus MeandrostylisFrech, matching the diagnosis of the genus, with respect to thicktrabecular projections, tabuloid endotheca, analogical spines andconfluent septa (Roniewicz, personal commun., 2007). Mean-drostylis vancouverensis (Clapp and Shimer) has been previouslymisidentified by other workers and is quite common throughoutthe Upper Triassic of the North American Cordillera. Squires(1956, fig. 48) and Stanley (1986, pl. 3.1, fig. 14) illustrated anddescribed Elysastrea profunda (Reuss) from Idaho. Stanley(1994a) identified two specimens of Chondrocoenia schafhaeutli(Winkler) from American Museum of Natural History (AMNH)

488 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
TABLE 2—Geographic location (by terrane) and stratigraphic range of coral species identified in this study. (X) indicates presence, (FO) denotes a first timeoccurrence outside the Tethys, and stratigraphic ages: (C) Carnian, (EN) Early Norian, (N) Norian, (LN) Late Norian, and (R) Rhaetian.
Coral species AL
WR
SWR NWR WA
Tethys
West EastOutsideTethys
Strat.range
Kompsasteria cf. K. oligocystis (Frech) X(FO) X EN-RGablonzeria major (Frech) X X N-RG. profunda (Reuss) X X X X X X N-RG. grandiosa n. sp. X ENCuifia cf. C marmorea (Frech) X X EN/NParacuifia smithi n. sp. X ENP. jennieae n. sp. X ENP. anomala n. sp. X ENDistichophyllia norica (Frech) X X X X X X EN-RRetiophyllia alfurica (Wilckens) X(FO) X N-RR. caespitosa (Reuss) X(FO) X X EN-RR. clathrata (Emmrich) X X X X EN-RR. dawsoni (Clapp and Shimer) X X X EN-RR. cf. R. frechi Roniewicz X X EN-RR. cf. R. norica (Frech) X X X NR. oppeli (Reuss) X X X EN-RR. parvisepta (Squires) X X EN-LNR. cf. R. robusta Roniewicz X(FO) X EN-RR. tenuicosta (Reuss) X(FO) X C-ENR. dendriformis n. sp. X X EN-RR. obtusa n. sp. X ENKuhnastraea decussata (Reuss) X X X X X EN-RK. incrassata (Frech) X X N-RK. cowichanensis (Clapp and Shimer) X X X X EN-RMargarosmilia cf. M. charlyana (Frech) X(FO) X X EN-RM. cf. M. richthofeni Volz X(FO) X C-ENM. confluens (Munster) X(FO) X EN-RDistichomeandra austriaca (Frech) X X X EN-RD. minor (Frech) X(FO) X EN-RD. sp. A X X(FO) X C-END. sp. B X(FO) X C-ENAstraeomorpha crassisepta Reuss X X X X X X X N-RA. confusa (Winkler) X(FO) X(FO) X X X N-RParastraeomorpha cf. P. similis Roniewicz X X X N-RChondrocoenia schafhaeutli (Winkler) X X X X N-RC. cf. C. paradoxa (Melnikova) X X X ENPamiroseris meriani (Stoppani) X X X X X EN-RPamiroseris borealis (Smith) X ENCrassistella juvavica (Frech) X X X X X ENC. parvula (Melnikova) X(FO) X EN?Ampakabastraea cf. A. nodosa Cuif X ENRecticostrastraea wallowaensis Stanley and Whalen X X N-RCampesteria prolixia n. sp. X N-RStylophyllum cf. S. pygmaeum Frech X(FO) X EN-R?Meandrostylis grandisepta Stanley and Whalen X X ENM. vancouverensis (Clapp and Shimer) X X X EN-RAnthostylis cf. A. acanthophora (Frech) X X X X N-R
collections, Upper Triassic of Peru (Stanley, 1994a, pl. 3, figs. 5,6), which we here place in synonymy with Meandrostylis van-couverensis. Type material in Clapp and Shimer (1911, pl. 40,fig. 8, pl. 42, fig. 17) is poorly preserved, containing few distin-guishing features; however, examination of better preserved top-otype material (this study) reveals smaller calical diameters, ex-tremely short septa, and underdeveloped columella distinguishingthis species. Other specimens illustrated by Stanley (1994a) do,however, show affinities with genus Chondrocoenia Roniewicz.
Genus ANTHOSTYLIS Roniewicz, 1989Type species.⎯Coccophyllum acanthophorum FRECH, 1890.
ANTHOSTYLIS cf. A. ACANTHOPHORA (Frech, 1890)(Figure 5.27, 5.31)
Coccophyllum acanthophorum FRECH, 1890, p. 89, pl. 20, figs. 4–11.Coccophyllum acanthophorum Frech SQUIRES, 1956, p. 19, figs. 19–21; CUIF,
1973, p. 280, figs. 31, 32; CUIF, 1977, p. 17, pl. 1, figs. 12, 13, pl. 2, fig.5; STANLEY, 1979, pl. 2, figs. 3, 5–7; SENOWBARI-DARYAN, 1980, p. 40,pl. 4, fig. 6; STANLEY, 1986, pl. 3.1, figs. 8, 9.
Anthostylis acanthophora (Frech) RONIEWICZ, 1989, p. 133, pl. 41, figs. 5–7.(See Stanley, 1994.)
Description.⎯Colony laminar and columnar, multiserial; corallites cerioid,polygonal, walls well defined, sharp, no columella; calices deep, d � 1.5–2.0
mm, s � 20–25, nonconfluent, differentiated to three cycles; S1 and S2 equalthickness, slightly longer than S3.
Material examined.⎯Fourteen colony fragments from Lake Cowichan (MI8302) and Tahsis Inlet (MI 9913), Vancouver Island, Canada.
Occurrence.⎯Rhaetian, Vancouver Island, Canada; Norian, Lewiston, Ida-ho; Pucara Group, Peru; Norian to Rhaetian, Northern Calcareous Alps.
Discussion.⎯Anthostylis acanthophora (Frech) has a limitedgeographical range, only being found in the latest Norian toRhaetian deposits in the Northern Calcareous Alps and two Cor-dilleran terranes (Wrangellia and the Wallowa terrane of Idaho).
DISCUSSION AND CONCLUSIONS
The exquisitely preserved, silicified corals described hereinfrom Upper Triassic localities of the Alexander terrane and Wran-gellia (Table 1) greatly expand our understanding of the taxonom-ic, stratigraphic, and geographic relationships of scleractinian cor-als from not only these two allochthonous terranes but many otherterranes within the North American Cordillera, further establish-ing a rather large taxonomic dataset that can be used for statisticalcomparison. This work subsequently combines and updates sys-tematic coral identification lists from several smaller contribu-tions, filling gaps in the known taxonomy while shedding lighton worldwide scleractinian coral distribution.

489CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
Fourteen of forty-seven species (or 29%) identified and de-scribed in this study are here recorded for the first time outsidethe Tethys region, with seven new species: Gablonzeria grandio-sa, Retiophyllia dendriformis, Retiophyllia obtusa, Paracuifiasmithi, Paracuifia jennieae, Paracuifia anomala, and one new ge-nus, Campsteria, type species of which is Campesteria prolixian. sp. (Table 2). Twelve species (five cosmopolitan) within greaterWrangellia were found to be in common with the Wallowa terraneof northeast Oregon (Table 2), a connection that was previouslyunrecognized in statistical analysis of in-progress taxonomic coralidentifications (this data set) within Caruthers and Stanley (inpress). A Wrangellia/Wallowa terrane connection was first sug-gested based on lithology (Jones et al., 1977) and has since beenreintroduced through similarities in bivalves (Newton, 1983), gas-tropods (Blodgett, personal commun., 2004), and corals (thisstudy). When comparing the Wallowa and Alexander terranes,only eight species (five cosmopolitan) were found in common(Table 2), suggesting little proximity between the two terranes.
Our understanding of the stratigraphy and paleontology of theAlexander terrane has been greatly enhanced by recent field stud-ies (Katvala and Stanley, in press; Caruthers and Stanley, inpress), the focus of which revolved around age constraints andbiofacies comparison within coeval limestone units. Coral diver-sity from the Alexander terrane and Wrangellia is unusually highcompared with silicified faunas from other terranes (Clapp andShimer, 1911; Smith, 1927; Squires, 1956; Montanaro-Gallitelliet al., 1979; Stanley, 1979; Stanley and Whalen, 1989). Caruthersand Stanley (in press) provide discussion and elaboration, focus-ing on collection size, updating taxonomy, and describing varioustechniques involved with processing and extraction of silicifiedfaunas.
The phaceloid genus Retiophyllia Cuif presented the most di-verse species in our study. This appears also to be true in theNorthern Calcareous Alps (Roniwicz, 1989), where it is the prin-cipal reef-building genus (Flugel, 2002). Highly variable ingrowth form, corallite density and shape, as well as septal orna-mentation, Retiophyllia displays a variety of branching arrange-ments as well as angles of bifurcation within the colony. Twelvespecies were identified (Figs. 2.22, 2.26–2.28, 3; Table 2), ninefrom combined northern and southern Wrangellia, two from theAlexander terrane, and one species found overlapping betweenthe two terranes. Retiophyllia alfurica (Wilckens), and R. tenui-costa (Reuss) are new additions to the genus, previously recog-nized only from the former Tethys and eastern PanthalassanOcean. Retiophyllia parviseptum (Squires) is also a new addition,relatively well known within the North American Cordillera, pre-viously misidentified to genus Pinacophyllum Frech by Squires(1956). Retiophyllia clathrata (Emmrich) and R. dendriformis n.sp. are distinct in growth form, mode of budding, and septal at-tributes, and yet co-occur within the Rhaetian strata at Lake Cow-ichan (southern Wrangellia).
Having appropriate microstructure is important when consid-ering validity of known taxonomic classification schemes pro-posed by Cuif (1965–1976), Beauvais (1980) and Roniewicz(1989). For corals placed within genus ?Ampakabastraea Allo-iteau, it is important to note that morphology was the sole meansused for identification, owing to the fact that microstructure hasbeen obliterated by mineralogical replacement. For this reason anelement of caution is advised.
Few localities exist that span the Carnian-Norian boundary,with rare occurrences of early Norian forms. Most of our knowl-edge comes from younger (Norian, late Norian and Rhaetian) cor-al faunas occurring in the Alps (Roniewicz, 1989). Within thisstudy, we were fortunate to have eight early Norian localitiesencompassing both the Alexander terrane and northern Wrangel-lia, two of which (Flounder Cove, MI 0099, in Keku Strait, andGreen Butte, MI 9934, in the Wrangell Mountains) yield well-dated and immaculately preserved corals from strata very close
to the Carnian-Norian boundary (Katvala and Stanley, in press;Caruthers and Stanley, in press). An overall trend is exemplifiedin this study by coral species whose stratigraphic range co-occursthroughout the Carnian (C), early Norian (EN), and Rhaetian (R).Early Norian deposits from the Alexander terrane yield ten EN-R, nine N, eight EN, three N-R, and two C-EN species, whilesimilar aged localities from northern Wrangellia contain elevenEN-R, six N, four EN, three C-EN, and one N-R species (Table2). Corals from southern Wrangellia, on the other hand, were onlystudied from Rhaetian strata, therefore producing different results;seven EN-R and eleven N-R species (Table 2). The stratigraphicrange of five species originally described only from the CarnianDolomites in the Alps was extended to the early Norian andRhaetian. Within this study, both localities holding this mixedfauna contain limestone units straddling this boundary (Katvalaand Stanley, in press; Caruthers and Stanley, in press) and thushave a direct bearing on the debated impact of the Carnian/Norianextinction event. When viewing these results, it is important tonote that N-R corals contain an element of uncertainty (with re-spect to EN, N, or late Norian (LN) assignment), therefore ren-dering it necessary to separate EN-R from the more general N-Rgroup.
Altogether the exquisite silicification, extremely broad strati-graphic range, and abundant diversity in Upper Triassic coral fau-na from the Alexander terrane and Wrangellia suggest many com-mon species with the former Tethys Sea and eastern PanthalassanOcean, as well as other Cordilleran terranes, which is largely un-precedented and rare in the majority of Upper Triassic depositsfrom the North American Cordillera.
ACKNOWLEDGMENTS
We acknowledge a grant from the National Science Foundation (EAR-9624501) and Geological Society of America (7629-04). We wish to thank J.Baichtal Forest, Geologist, and the U.S. Forest Service for providing logisticaland field support at Keku Strait and Gravina Island, as well as E. Katvala fordetailed fieldwork, conodont identifications, and vast collections of KekuStrait fossils, and Dr. R. Blodgett for his field collaboration and for sharinghis knowledge of Alaskan geology. Dr. E. Roniewicz is thanked for her ex-pertise in the identification of problematic taxa and the many helpful sugges-tions throughout the review process, immensely improving the quality of thiscontribution. Further gratitude goes to D. Sharp and D. Rosenkrans (Wrangell-St. Elias National Park and Preserve) for collecting permits and logisticalsupport. Type material from previous collections made by Clapp and Shimer(1911) and Smith (1927) was kindly loaned to us from the National Museumof Natural History, Smithsonian Institution, and the Geological Survey ofCanada. The authors would like to dedicate this contribution to the memoryof Jennie Caruthers (mother); her soft, fun-loving demeanor and laughterbrightened each day. She will never be forgotten and truly was ‘‘Sweeter thanflowers.’’
REFERENCES
ALLOITEAU, J. 1952. Madreporaires post-paleozoiques, p. 539–684. In J. Piv-eteau (ed.), Triate de Paleontologie, Paris.
ALLOITEAU, J. 1957. Contribution a la systematique des Madreporaires fossils.Triate de Paleontologie, Paris, 462 p.
ARMSTRONG, A. K., E. M. MACKEVETT JR., AND N. J. SILBERLING. 1969.The Chitistone and Nizina Limestones of part of the southern WrangellMountains, Alaska—A preliminary report stressing carbonate petrographyand depositional environments. U.S. Geological Survey Professional Paper650-D:D49–D62.
BEAUVAIS, L. 1980. Sur la Taxonomie des Madreporaires mesozoiques. ActaPalaeontologica, 25:345–360.
BOURNE, G. C. 1900. The Anthozoa. In R. Lankester (ed.), Treatise on Zo-ology, II, Chapter VI, 84 p.
CARUTHERS, A. H. 2005. Upper Triassic carbonates and scleractinian coralsfrom Wrangellia and the Alexander terrane (Alaska and Vancouver Island,Canada): Depositional environments and paleobiogeography. UnpublishedM.S. thesis, University of Montana, Missoula, 211 p.
CARUTHERS, A. H. AND G. D. STANLEY JR. In press. Upper Triassic silicifiedshallow-water corals and other marine fossils from Wrangellia and the Al-exander terrane, Alaska and Vancouver Island, British Columbia. Geolog-ical Society of America Special Paper.
CLAPP, C. H. AND H. W. SHIMER. 1911. The Sutton Jurassic of the Vancouver

490 JOURNAL OF PALEONTOLOGY, V. 82, NO. 3, 2008
Group, Vancouver Island. Boston Society of Natural History, Proceedings,34(12):426–438.
CUIF, J. P. 1965. Sur les rapports des genres des madreporaires, MontlivaltiaLam. et Thecosmilia M. Edw. Et H. et leur presence au Trias. Bulletin dela Societe Geologique de France, serie 7, no. 7:530–536.
CUIF, J. P. 1966. Structure de quelques Polypiers phaceloides triasiques. Traitede Paleontologie, France, serie 7, no. 8:125–132.
CUIF, J. P. 1973. Recherches sur les Madreporaires du Trias. I. Famille desStylophyllidae. Traite de Paleontologie, serie 3, no. 97, Sciences de la Terre,17:211–291.
CUIF, J. P. 1974. Recherches sur les Madreporaires du Trias II. Astraeoidea.Revision des genres Montlivaltia et Thecosmilia. Etude de quelques typesstructuraux du Trias de Turquie. Bulletin du Museum National d’Histoirenaturelle, Paris, serie 3, no. 275, Sciences de la Terre, 40:293–400.
CUIF, J. P. 1975a. Recherches sur les Madreporaires du Trias II. Astraeoidea.Revision des genres Monlivaltia et Thecosmilia. Etude de quelques typestructuraux du Trias de Turquie. Bulletin du Museum National d’Histoirenaturelle, Paris, serie 3, no. 275, Sciences de la Terre, 40:293–400.
CUIF, J. P. 1975b. Recherches sur les Madreporaires du Trias III. Etude desstructures pennulaires chez les Madreporaires triasiques. Bulletin du Mu-seum National d’Histoire naturelle, Paris, serie 3, no. 310, Sciences de laTerre, 44:45–127.
CUIF, J. P. 1976. Recherches sur les Madreporaires du Trias IV. Formes cer-iomeandroides et thamnasterioides du Trias des Alpes et du Taurus sudan-atolien. Bulletin du Museum National d’Histoire naturelle, Paris, serie 3,no. 381, Sciences de la Terre, 53:65–194.
CUIF, J. P. 1977. Arguments pour une relation phyletique entre les Madre-poraires paleozoique et ceux de Trias. Implications systematiques etl’analyse microstructurale des Madreporaires triasiques. Memoires de laSociete Geologique de France, (n. ser.), Memoir 129, 54 p.
D’ORBIGNY, A. D. 1849. Note sur la classe des Amorphozoaires. Magazinede Zoologie pure et applique, series 2, no. 1:545–550.
DRONOV, V. I., A. GAZDZICKI, AND G. K. MELNIKOVA. 1982. Die triadischeRiffe in sudostlichen Pamir. Facies, 6:107–127.
EDWARDS, M. AND J. HAIME. 1851. Monographie des polypieres fossiles desterrains palaeozoiques, precedee d’un tableau general de la classificationdes polypes. In Archimes du Museum d’historie naturelle, Tome V, 89 p.
EHRENBERG, C. G. 1835. Dritter Beitrag zur Erkenntnis grosser Organisationin der Richtung des kleinsten Raumes. Abhandlungen der Koniglichen Aka-demie der Wissenschaften zu Berlin, p. 145–336.
ELIASOVA, H. 1976. Nouvelle famille du sous-ordre Amphiastracina Allo-iteau, 1952 (Hexacorallia), Tithonien de Tchecoslovaquie. Vestnik Ustred-niho ustavu geologickeho, 51:177–178.
EMMRICH, H. F. 1853. Geognostishe Beobachtungen aus den ostlichen bay-erischen und den angrenzenden osterreichischen Alpen. Jahrbuch d. K. K.Geology, Reichsanstalt, III–IV, 4(1):326–394.
FRECH, F. 1890. Die Korallenfauna der Trias. Die Korallen der juvavischenTriasprovinz. Palaeontographica, 37, 116 p.
FLUGEL, E. 2002. Triassic reef patterns, p. 391–464. In W. Kiessling, E. Flu-gel, and J. Golonka (eds.), Phanerozoic Reef Patterns. SEPM, no. 72, Tulsa.
HAAS, O. 1909. Bericht uber neue Aufsammlungen in den Zlambachmergelnder Fischerwiese bei Alt-Aussee. Beitrage zur Palaontologie in Geologie,Osterreich-Ungarns und des Orients. Mitteilungen des Geologie und Pa-laontologie Institut der Universitat Wien, 22:145–167.
ILLJINA, T. G. AND G. K. MELNIKOVA. 1986. Corals used as indicators forstratigraphy of carbonates, p. 30–67. In Parastratigraficheskie gruppy floryi fauny traisa [Parastratigraphic groups of the Triassic fauna and flora].Nedra, Leningrad.
KATVALA, E. C. AND G. D. STANLEY JR. In press. Conodont biostratigraphyand facies correlations in an Upper Triassic island arc, Keku Strait, south-east Alaska. Geological Society of America Special Papers.
KLIPSTEIN, A. 1843. Beitrage zur Geologischen Kenntniss der Ostlichen Al-pen. Gie�en, 311 p.
LAUBE, G. 1865. Die Fauna der Schichten von St. Cassian, I Abt. Akademieder Wissenschaften, Mathematisch-Naturwissenschaftlichen, Klasse Denk-schrift, 24:223–296.
MATZNER, C. 1986. The Zlambach beds (Rhaetian) of the Northern LimestoneAlps: Platform-slope-environments with allochthonous carbonate sedimen-tation. Facies, 14, 102 p.
MELNIKOVA, G. K. 1968. O rode Zchatzhozoenia (Zhekhazoralla). Paleonto-logicheskiy Zhurnal, 1:11–17.
MELNIKOVA, G. K. 1971. Novye dannye o morfologii, mikrostrukture i sys-tematike pozdnetriasovych Thamnasterioidea. Paleontologicheskiy Zhurnal,2:21–35.
MELNIKOVA, G. K. 1974. Osobennosti gistologicheskikh struktur i mikros-truktur septalnogo aparata u pozdnetriasovykh skleraktinii. Trudy (Institutgeologii i geofiziki. Sibirskoe Otdelenie. AN SSSR), 201:224–228.
MELNIKOVA, G. K. 1975. Pozdnetriasovie skleraktinii yugo-vostochnogo Pa-mira, Akademie Nauk. Tadschik. SSSR Institute of Geology, 234 p.
MELNIKOVA, G. K. 1984. Novye pozdnetriasovye korally otriada Archaeo-coeniida Alloiteau yugo-vostotshnogo Pamira, p. 42–55. In Novye vidyiskopayemoy flory I fauny Tadzhikistana. Donish, Dushanbe.
MELNIKOVA, G. K. 1986. Korally kak indikatory raschleneniya karbonatnykhtolshch. Trudy Vsesoyuznogo nauchno-issledovatelskogo geologicheskogoinstituta, 334:30–67.
MELNIKOVA, G. K. 2001. Chapter on corals, p. 30–80. In A. Yu. Rozanovand A. A. Shevyrev (eds.), Atlas of Triassic Invertebrates of the Pamirs.Nauka, Moscow, Russia, 190 p.
MELNIKOVA, G. K. AND Y. M. BYCHKOV. 1986. Pozdnetriasovye skleraktiniikhrebta Kenkeren (Koryarskoe nagor’e), p. 63–81. In Y. D. Zacharov andY. I. Onoprienko (eds.), Korreliatsia Permo-Triasovykh otlozhenii VoslokaSSSR. Institute of Biology and Pedology DVNTs AN SSSR, Vladivostok.
MONTANARO-GALLITELLI, E., A. RUSSO, AND P. FERRARI. 1979. Upper Tri-assic coelenterates of western North America. Bollettino della Societa Pa-leontologica Italiana, 18:133–156.
MUNSTER, G. G. 1841. Beitrage zur Geognosie und Petrefactenkunde dessudostlichen Tirols, vorzuglich der Schichten von St. Cassian. Bayreuth,162 p.
NEWTON, C. R. 1983. Paleozoogeographic affinities of Norian bivalves fromthe Wrangellian, Peninsular, and Alexander terranes, western North Amer-ica, p. 37–48. In C. H. Stevens (ed.), Pre-Jurassic Rocks in Western NorthAmerican Suspect Terranes. SEPM Pacific Section, Los Angeles.
PRINZ, P. 1991. Mesozoische Korallen aus Nordchile. Palaeontographica, A,216:47–209.
PRINZ-GRIMM, P. 1995. Triassiche Korallen der sudliche Zentral-Anden Geo-logica, Palaeontologica, 10:233–243.
REUSS, A. E. 1854. Beitrage zur Charakteristik der Kreideschichten in denOstalpen, besonders im Gosauthale und am Wolfgangsee. Akademie derWissenschaften, Mathematisch-Naturwissenschaftlichen, Klasse Denk-schrift, 7:67–133.
REUSS, A. E. 1865. Uber einige Anthozoen der Kossener Schichten und deralpinen Trias. Akademie der Wissenschaften, Mathematisch-Naturwissen-schaftlichen, Klasse Sitzungsber, 50(1):153–167.
RONIEWICZ, E. 1989. Triassic scleractinian corals of the Zlambach beds,Northern Calcareous Alps, Austria. Denkschrift Osterreiche Akademie Wis-senschaften, Vienna, 126, 152 p.
RONIEWICZ, E. 1995. Upper Triassic solitary corals from the Gosaukamm andother North Alpine regions. Osterreiche Akademie Wissenschaften, Vienna,202, 41 p.
SENOWBARI-DARYAN, B. 1980. Neue Kalkschwamme (Sphinctozoen) aus ob-ertriadischen Riffkalken von Sizilien. Mitteilungen der Gesellschaft Geo-logie und Bergbaustudenten Osterreich, 26:179–203.
SMITH, P. P. 1912. The occurrence of coral reefs in the Triassic of NorthAmerica. American Journal of Science, 33:92–96.
SMITH, J. P. 1927. Upper Triassic marine invertebrate faunas of North Amer-ica. U.S. Geological Survey Professional Paper 141, 262 p.
SQUIRES, D. F. 1956. A new Triassic coral fauna from Idaho. American Mu-seum Novitates, 1797, 27 p.
STANLEY, G. D., JR. 1979. Paleoecology, structure and distribution of Triassiccoral buildups in western North America. University of Kansas Paleonto-logical Contribution 65, 68 p.
STANLEY, G. D., JR. 1986. Late Triassic coelenterate faunas of western Idahoand northeastern Oregon. U.S. Geological Survey professional Paper 1435:23–39.
STANLEY, G. D., JR. 1989. An Upper Triassic reefal limestone, southern Van-couver Island, B.C. The Canadian Society of Petroleum Geologists Memoir13:766–775.
STANLEY, G. D., JR. 1994a. Upper Triassic corals from Peru. Palaeontogra-phica A, 233,1–6:75–98.
STANLEY, G. D., JR. 1994b. Upper Triassic spongiomorph and coral associ-ation dredged off the northwestern Australian shelf. AGSO Journal Austra-lian Geology and Geophysics, 15(1):127–133.
STANLEY, G. D., JR. AND M. T. WHALEN. 1989. Triassic corals and spon-giomorphs from Hells Canyon, Wallowa Terrane, Oregon. Journal of Pa-leontology, 63(6):800–819.
STANLEY, G. D. JR. AND B. SENOWBARI-DARYAN. 1999. Upper Triassic reeffauna from the Quesnel Terrane, central British Columbia, Canada. Journalof Paleontology, 73(5):787–802.
STANLEY, G. D. JR., C. GONZALEZ-LEON, M. R. SANDY, B. SENOWBARI-DARYAN, P. DOYLE, M. TAMURA, AND D. H. ERWIN. 1994. Upper Triassicinvertebrates from the Antimonio Formation, Sonora, Mexico. Paleontolog-ical Society Memoir 36, 33 p.
STOPPANI, A. 1858. Les petrification d’Esino ou description des fossils ap-purtenant au depot triassique superieur des environs d’Esino en Lombardie,Milano. J. Bernardoni, 113 p.
VAUGHN, T. W. AND J. W. WELLS. 1943. Revision of the suborders, familiesand genera of the Scleractinia. Geological Society of America Special Paper44, 363 p.

491CARUTHERS AND STANLEY—TRIASSIC SCLERACTINIAN CORALS FROM NORTH AMERICA
VINASSA DE REGNY, P. 1915. Triadische Algen, Spongien, Anthozoen undBryozoen aus Timor. Palaeontologie Timor, Joh. Wanner (ed.), 4(8):73–118.
VOLZ, W. 1896. Die Korallenfauna der Schichten von St-Cassian in Sud-Tyrol. Palaeontographica, 43, 124 p.
WILCKENS, O. 1937. Korallen und Kalkschamme aus dem obertriadischenPharetronenkalk von Seram (Molukken). Neues Jahrbuch fur Mineralogieund Palaontologie, Stuttgart, 77(B):171–211.
WINKLER, G. 1861. Der Oberkeuper, nach Studien in den bayerischen Alpen.Zeitschrift Deutsche Geologische Gesellschaft, 13:459–521.
YARNELL, J. M. 2000. Paleontology of two North American Triassic reeffaunas: Implications for terrane paleogeography. Unpublished M.S. thesis,University of Montana, Missoula, 141 p.
ZANKL, H. 1969. Der Hohe Goll. Aufbau und Lebensbild eines Dachsteinkalk-Riffes in der Obertrias der nordlichen Kalkalpen. Abhandlungen-sencken-bergiana Naturforschungs Gesellschaft, 519, 123 p.
ACCEPTED 8 NOVEMBER 2007
Related Documents











