APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Jan. 2003, p. 233–240 Vol. 69, No. 1 0099-2240/03/$08.000 DOI: 10.1128/AEM.69.1.233–240.2003 Copyright © 2003, American Society for Microbiology. All Rights Reserved. Synthesis of GDP-Mannose and Mannosylglycerate from Labeled Mannose by Genetically Engineered Escherichia coli without Loss of Specific Isotopic Enrichment Maria-Manuel Sampaio, 1 Helena Santos, 1 and Winfried Boos 2 * Instituto de Tecnologia Química e Biolo ´gica, Universidade Nova de Lisboa, 2780-156 Oeiras, Portugal, 1 and Department of Biology, University of Konstanz, 78457 Konstanz, Germany 2 Received 23 July 2002/Accepted 22 October 2002 We report the construction of an Escherichia coli mutant that harbors two compatible plasmids and that is able to synthesize labeled 2-O--D-mannosyl-D-glycerate from externally added labeled mannose without the loss of specific isotopic enrichment. The strain carries a deletion in the manA gene, encoding phosphomannose isomerase. This deletion prevents the formation of fructose-6-phosphate from mannose-6-phosphate after the uptake of mannose from the medium by mannose-specific enzyme II of the phosphotransferase system (PtsM). The strain also has a deletion of the cps gene cluster that prevents the synthesis of colanic acid, a mannose- containing polymer. Plasmid-encoded phosphomannomutase (cpsG) and mannose-1-phosphate guanylyltrans- ferase (cpsB) ensure the formation of GDP-mannose. A second plasmid harbors msg, a gene from Rhodothermus marinus that encodes mannosylglycerate synthase, which catalyzes the formation of 2-O--D-mannosyl-D- glycerate from GDP-mannose and endogenous glycerate. The rate-limiting step in 2-O--D-mannosyl-D-glyc- erate formation is the transfer of GDP-mannose to glycerate. 2-O--D-mannosyl-D-glycerate can be released from cells by treatment with cold-water shock. The final product is formed in a yield exceeding 50% the initial quantity of labeled mannose, including loss during preparation and paper chromatography. 2-O--D-Mannosyl-D-glycerate (MG) is one of the most widespread compatible solutes of thermophilic or hyperther- mophilic bacteria and archaea (21). This compound has oth- erwise been encountered only in the red algae of the order Ceramiales (4). The highly preferential distribution of MG among organisms requiring high temperatures for growth led to the hypothesis that it could play a role in the thermopro- tection of cell components in vivo. This speculation still lacks experimental proof, but at least in vitro, MG has been shown to be highly efficient in the protection of enzymes against thermal inactivation (3, 19). In order to study the metabolic fate of MG and the mech- anisms underlying its stabilizing effect on proteins, it is desir- able for this compound to be available in a labeled form with a high level of specific isotopic enrichment ( 14 C, 13 C); at present, such a compound is not commercially available. The biosynthesis of MG has been characterized in detail for the thermophilic bacterium Rhodothermus marinus (12) and for the hyperthermophilic archaeon Pyrococcus horikoshii (7). The known pathways involve the transfer of -mannosyl resi- dues from GDP-mannose to either D-glycerate, forming MG, or D-3-phosphoglycerate, followed by hydrolysis of mannosyl- 3-phosphoglycerate to MG (7, 12). Most organisms in which the synthesis of MG has been observed are not accessible for easy genetic manipulation (21). Thus, the conversion of a sim- ple radioactive sugar for the formation of MG is plagued by its fast metabolism after entering the producing organism. This situation is due to the common separation of catabolic and anabolic pathways seen for sugar-type compatible solutes. The strict separation of trehalose metabolism and its internal syn- thesis is a typical example of this phenomenon (10). Therefore, we thought of using a genetically altered Escherichia coli strain to synthesize MG from external mannose. E. coli does produce GDP-mannose, mainly for the synthesis of colanic acid via GDP-fucose (2, 9). The cpsB gene, encoding the enzyme for GDP-mannose synthesis (mannose-1-phosphate guanylyltrans- ferase), and the cpsG gene, encoding phosphomannomutase, form an operon that is part of the cps gene cluster necessary for the synthesis of colanic acid (25). By constructing an E. coli strain that is unable to metabolize mannose and by using plasmid-encoded enzymes, we were able to channel the uptake of mannose preferentially into conver- sion to GDP-mannose and MG. MATERIALS AND METHODS Bacterial strains and growth conditions. The strains and plasmids used in this study are described in Table 1. Strains were grown in minimal medium A (MMA) (15) or in NZA medium (22) supplemented with 0.2% glucose. For confirming strain markers, we used plates containing mannose and arabinose (0.2%) as required and glycerol (0.4%) as a carbon source. Unless stated otherwise, cells were centrifuged at 8,000 g for 7 min at 10°C. For transport assays, thin-layer chromatography (TLC) analysis, and preparative synthesis, cells were washed twice with MMA in the absence of a carbon source and resuspended in the same medium. The concentrations of antibiotics in solid and liquid media were as follows: ampicillin, 100 g/ml; chloramphenicol, 30 g/ml; tetracycline, 5 g/ml; and kanamycin, 100 g/ml. When needed, amino acids were added to the minimal medium at the follow- ing concentrations: histidine, 0.15 g/liter; isoleucine, 0.21 g/liter; leucine, 0.46 g/liter; and valine, 0.32 g/liter. The vitamins pyrodoxine and B 12 were used at 2.5 and 1 mg/liter, respectively. Genetic construction of the producer strain. P1vir transductions were per- formed as described by Miller (15); P1vir was from a laboratory stock. In order to use arabinose-induced promoters, it was necessary to introduce an araABC-leu * Corresponding author. Mailing address: Department of Biology, University of Konstanz, 78457 Konstanz, Germany. Phone: 49-7531- 882658. Fax: 49-7531-883356. E-mail: Winfried.Boos@Uni-Konstanz .de. 233 on July 4, 2016 by guest http://aem.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Jan. 2003, p. 233–240 Vol. 69, No. 10099-2240/03/$08.00�0 DOI: 10.1128/AEM.69.1.233–240.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Synthesis of GDP-Mannose and Mannosylglycerate from LabeledMannose by Genetically Engineered Escherichia coli
without Loss of Specific Isotopic EnrichmentMaria-Manuel Sampaio,1 Helena Santos,1 and Winfried Boos2*
Instituto de Tecnologia Química e Biologica, Universidade Nova de Lisboa, 2780-156 Oeiras, Portugal,1
and Department of Biology, University of Konstanz, 78457 Konstanz, Germany2
Received 23 July 2002/Accepted 22 October 2002
We report the construction of an Escherichia coli mutant that harbors two compatible plasmids and that isable to synthesize labeled 2-O-�-D-mannosyl-D-glycerate from externally added labeled mannose without theloss of specific isotopic enrichment. The strain carries a deletion in the manA gene, encoding phosphomannoseisomerase. This deletion prevents the formation of fructose-6-phosphate from mannose-6-phosphate after theuptake of mannose from the medium by mannose-specific enzyme II of the phosphotransferase system (PtsM).The strain also has a deletion of the cps gene cluster that prevents the synthesis of colanic acid, a mannose-containing polymer. Plasmid-encoded phosphomannomutase (cpsG) and mannose-1-phosphate guanylyltrans-ferase (cpsB) ensure the formation of GDP-mannose. A second plasmid harbors msg, a gene from Rhodothermusmarinus that encodes mannosylglycerate synthase, which catalyzes the formation of 2-O-�-D-mannosyl-D-glycerate from GDP-mannose and endogenous glycerate. The rate-limiting step in 2-O-�-D-mannosyl-D-glyc-erate formation is the transfer of GDP-mannose to glycerate. 2-O-�-D-mannosyl-D-glycerate can be releasedfrom cells by treatment with cold-water shock. The final product is formed in a yield exceeding 50% the initialquantity of labeled mannose, including loss during preparation and paper chromatography.
2-O-�-D-Mannosyl-D-glycerate (MG) is one of the mostwidespread compatible solutes of thermophilic or hyperther-mophilic bacteria and archaea (21). This compound has oth-erwise been encountered only in the red algae of the orderCeramiales (4). The highly preferential distribution of MGamong organisms requiring high temperatures for growth ledto the hypothesis that it could play a role in the thermopro-tection of cell components in vivo. This speculation still lacksexperimental proof, but at least in vitro, MG has been shownto be highly efficient in the protection of enzymes againstthermal inactivation (3, 19).
In order to study the metabolic fate of MG and the mech-anisms underlying its stabilizing effect on proteins, it is desir-able for this compound to be available in a labeled form witha high level of specific isotopic enrichment (14C, 13C); atpresent, such a compound is not commercially available.
The biosynthesis of MG has been characterized in detail forthe thermophilic bacterium Rhodothermus marinus (12) andfor the hyperthermophilic archaeon Pyrococcus horikoshii (7).The known pathways involve the transfer of �-mannosyl resi-dues from GDP-mannose to either D-glycerate, forming MG,or D-3-phosphoglycerate, followed by hydrolysis of mannosyl-3-phosphoglycerate to MG (7, 12). Most organisms in whichthe synthesis of MG has been observed are not accessible foreasy genetic manipulation (21). Thus, the conversion of a sim-ple radioactive sugar for the formation of MG is plagued by itsfast metabolism after entering the producing organism. Thissituation is due to the common separation of catabolic and
anabolic pathways seen for sugar-type compatible solutes. Thestrict separation of trehalose metabolism and its internal syn-thesis is a typical example of this phenomenon (10). Therefore,we thought of using a genetically altered Escherichia coli strainto synthesize MG from external mannose. E. coli does produceGDP-mannose, mainly for the synthesis of colanic acid viaGDP-fucose (2, 9). The cpsB gene, encoding the enzyme forGDP-mannose synthesis (mannose-1-phosphate guanylyltrans-ferase), and the cpsG gene, encoding phosphomannomutase,form an operon that is part of the cps gene cluster necessary forthe synthesis of colanic acid (25).
By constructing an E. coli strain that is unable to metabolizemannose and by using plasmid-encoded enzymes, we were ableto channel the uptake of mannose preferentially into conver-sion to GDP-mannose and MG.
MATERIALS AND METHODS
Bacterial strains and growth conditions. The strains and plasmids used in thisstudy are described in Table 1. Strains were grown in minimal medium A (MMA)(15) or in NZA medium (22) supplemented with 0.2% glucose. For confirmingstrain markers, we used plates containing mannose and arabinose (0.2%) asrequired and glycerol (0.4%) as a carbon source. Unless stated otherwise, cellswere centrifuged at 8,000 � g for 7 min at 10°C. For transport assays, thin-layerchromatography (TLC) analysis, and preparative synthesis, cells were washedtwice with MMA in the absence of a carbon source and resuspended in the samemedium.
The concentrations of antibiotics in solid and liquid media were as follows:ampicillin, 100 �g/ml; chloramphenicol, 30 �g/ml; tetracycline, 5 �g/ml; andkanamycin, 100 �g/ml.
When needed, amino acids were added to the minimal medium at the follow-ing concentrations: histidine, 0.15 g/liter; isoleucine, 0.21 g/liter; leucine, 0.46g/liter; and valine, 0.32 g/liter. The vitamins pyrodoxine and B12 were used at 2.5and 1 mg/liter, respectively.
Genetic construction of the producer strain. P1vir transductions were per-formed as described by Miller (15); P1vir was from a laboratory stock. In orderto use arabinose-induced promoters, it was necessary to introduce an araABC-leu
* Corresponding author. Mailing address: Department of Biology,University of Konstanz, 78457 Konstanz, Germany. Phone: 49-7531-882658. Fax: 49-7531-883356. E-mail: [email protected].
233
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

deletion. The nearby kefE::Tn10 pdx marker was introduced into strain MC1061,carrying �(araABC-leu). By selecting for Tetr and screening for loss of growth onarabinose and prototrophy for pyrodoxine, we obtained strain MS0501. A P1lysate of this strain was used to transfer �(araABC-leu) into strain SG20273,harboring a cps-his deletion. The resulting strain, MS1501, is still kefE::Tn10�(araABC-leu) �(cps-his). �cps was necessary to prevent the usage of GDP-mannose mainly for colanic acid synthesis. To remove kefE::Tn10, we used alysate of MC1061 to render MS1501 pdx� Tets, yielding strain MS1601. In orderto transfer the ptsG mutation of UE26 into MS1601, we introduced zce-726::Tn10, located at 24.25 min on the E. coli chromosome (24), into UE26 andobtained strain MS2601. A lysate of MS2601 allowed us to transfer the ptsGmutation into MS1601 and, by selecting for Tetr and screening for reducedgrowth on glucose, we obtained strain MS3601. The �(dgsA [mlc]-manA) dele-tion of strain PK2242 was introduced into MS3601 with the help of the nearbymarker nth::kan. By selecting for kanamycin resistance and screening for loss ofgrowth on mannose, we obtained strain MS4601. This producer strain is char-acterized by sensitivity to mannose and by slow growth on glucose. Additionally,MS4601 can be used to express arabinose-inducible genes. It has lost any sensi-tivity to arabinose. The final strain has the tendency to form mutations onLuria-Bertani plates, presumably because mannose is present in Luria-Bertanimedium, and mutations arise that render the strain mannose negative. In thepresence of 0.2% glucose or on glucose minimal medium plates, the strain isstable.
Preparation of plasmids and transformations were carried out according tostandard protocols (20).
Construction of plasmids. Two primers (5�-GTCTAGAATGGCGCAGTCGAAACTC-3� and 5�-CGCCTGCAGTTACTCGTTCAGCAACGTC-3�) con-taining the additional XbaI and PstI recognition sequences (underlined) wereused to amplify the genes cpsG (encoding phosphomannomutase) and cpsB(encoding mannose-1-phosphate guanylyltransferase) from genomic DNA of E.coli MC4100. The PCR product was digested with the corresponding restrictionenzymes and ligated into pTrc99B. The resulting plasmid was named pMS104and carries cpsG-cpsB under the control of an isopropyl-�-D-thiogalactopyrano-side (IPTG)-inducible promoter. To construct a vector carrying the mgs gene,encoding MG synthase of R. marinus, and compatible for use in combinationwith pMS104, we exchanged the antibiotic resistance of pMG37. The sequencecoding for chloramphenicol resistance, which was used for the exchange, wasobtained from digestion of pACYC184. In this way, we obtained plasmidpMS101. The producer strain was maintained free of plasmids. Only freshlytransformed strains were used for MG production.
[14C]MG and [14C]GDP-mannose production from [14C]mannose in wholecells. For evaluating the products formed in MS4601 cells, washed cultures ofcells with and without plasmids were exposed to external mannose (see Fig. 2).Transformed cells were grown in NZA medium with 0.2% glucose to an opticaldensity at 600 nm (OD600) of 0.5. After induction with 1 mM IPTG and further
growth for 6 h, cells were harvested and resuspended in MMA (without a carbonsource) to a final OD600 of 1.0. To 6 ml of this cell suspension, [14C]mannose wasadded to a final concentration of 0.078 �M (0.14 �Ci). After different timeintervals, 1-ml samples were withdrawn and centrifuged, and the pellet wasresuspended in 5 �l of 12% trichloroacetic acid (TCA). The mixture was kept onice for 10 min and centrifuged, and the supernatant was applied to a TLC plate(Silica 60; Merck). Chromatograms were run with a solvent system composed ofn-propanol and 25% ammonia (1:1, vol/vol). Authentic standards for mannose,mannose-1-phosphate, mannose-6-phosphate, GDP-mannose, and MG wereused for comparison. After being dried, the TLC plate was autoradiographed for3 days. Nonlabeled standards were visualized by spraying with �-naphthol–sulfuric acid solution followed by charring at 120°C (11).
To determine the rate of synthesis of MG, the above experiment was per-formed with a mixture of increasing concentrations of unlabeled mannose and0.078 �M (0.14 �Ci) [14C]mannose in the external medium (see Fig. 3). Toestimate the yield of MG production, 750 ml of a cell suspension (OD600, 2.0) inMMA was exposed to 5 �M mannose and incubated for 45 min at 42°C. Cellswere harvested, resuspended in 75 ml of ice-cold water, and incubated for 20 minin an ice bath. Cells were removed by centrifugation, and the supernatant solu-tion was lyophilized, dissolved in D2O, and analyzed by 1H nuclear magneticresonance (NMR) spectrometry. NMR spectra were acquired with a BrukerAMX300 spectrometer. The presence of MG was confirmed by spiking the NMRsample with the authentic compound. MG was quantified as previously described(23).
Preparative synthesis of [14C]MG. A culture of MS4601 transformed withpMS104 and pMS101 was grown in NZA medium with 0.2% glucose to an OD600
of 0.5. After induction with 1 mM IPTG, the cells were grown for an additional6 h, harvested, washed twice, and resuspended in 10 ml of MMA to a final OD600
of 2.0. [14C]Mannose was added to a final concentration of 1 �M, the cells wereincubated for 15 min at 42°C and centrifuged, and the pellet was quickly resus-pended in 2 ml of ice-cold water. After incubation on ice for 10 min, thesupernatant was recovered and lyophilized. The remainder of the sample wasdissolved in a minimal amount of water and applied to Whatman 3MM chro-matography paper. The chromatogram was developed with n-propanol and 25%ammonia (1:1, vol/vol) for 40 h, dried, and autoradiographed. The paper stripidentified by autoradiography as containing labeled MG was chromatographedwith water as the solvent; the eluate dripped into a glass vessel. The totalradioactivity was contained in 5 drops, which were kept frozen.
The final product was checked by TLC along with authentic unlabeled MG.Transport assays. MS4601, MS4601/pMS104, and MS4601/pMS104/pMS101
cells were grown in NZA medium with 0.2% glucose; 1 mM IPTG was added forinduction. Cells were harvested (8,000 � g, 7 min, 10°C), washed twice withMMA, and resuspended in MMA without a carbon source. To measure thetransport of mannose, we used a cell suspension with an OD600 of 1.5 to 2.0. To6 ml of culture, prewarmed for 2 min at 40°C, 0.3 �M [14C]mannose was added.
TABLE 1. Strains and plasmids used
Strain or plasmid Known genotype or description Reference or source
E. coli strainsMC4100 araD139 deoC1 flbB5301 ptsF25 rbsR relA1 rpsL150 � (argF-lac)U169 5SG20273 araD139 flbB5301 lon-100 ptsF25 rbsR relA1 rpsL150 � (argF-lac)U169 �(cps-his)273 �(lac-argF)U169 9MC1061 hsd merB araD139 �ara(ABC-leu)7679 �lacX74 galU galK rpsL thi 14PK2242 �(dgsA-manA) nth-1::kan 17Frag5 kefE::Tn10 pdx leu 13UE26 glk-7ptsG2 ptsM1 rpsL 1612078 zce-726::Tn10 24DH5� Cloning host Laboratory stockMS0501 MC1061 kefE::Tn10 pdx This studyMS1501 SG20273 �(araABC-leu)7679 kefE::Tn10 pdx This studyMS1601 MS1501 �(araABC-leu)7679 kefE� pdx� This studyMS2601 UE26 zce-726::Tn10 This studyMS3601 MS1601 ptsG2 zce-726::Tn10 This studyMS4601 MS3601 �(dgsA-manA) nth-1::kan This study
PlasmidspMG37 pKK223-3 harboring mgs from R. marinus under control of IPTG-inducible promoter; Ampr 12pACYC184 Ampr Cmr 6pTrc99B Ampr lacIq 1pMS101 pKK223-3 exchange of Cmr This studypMS104 PCR fragment containing cpsG and cpsB from E. coli in pTrc99B; Ampr This study
234 SAMPAIO ET AL. APPL. ENVIRON. MICROBIOL.
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from
For higher concentrations of mannose, cold substrate was premixed with radio-active substrate (we always used the same concentration of labeled material). Ateach time point, 1 ml of culture was filtered, and the rate of uptake of mannosewas determined by filtration. [14C]Mannose was purchased from AmershamPharmacia. It had a specific radioactivity of 296 mCi/mmol.
RESULTS
Construction of a strain that will produce MG with a highspecific radioactivity from external [14C]mannose. In thestrain which we constructed, mannose metabolism is directed
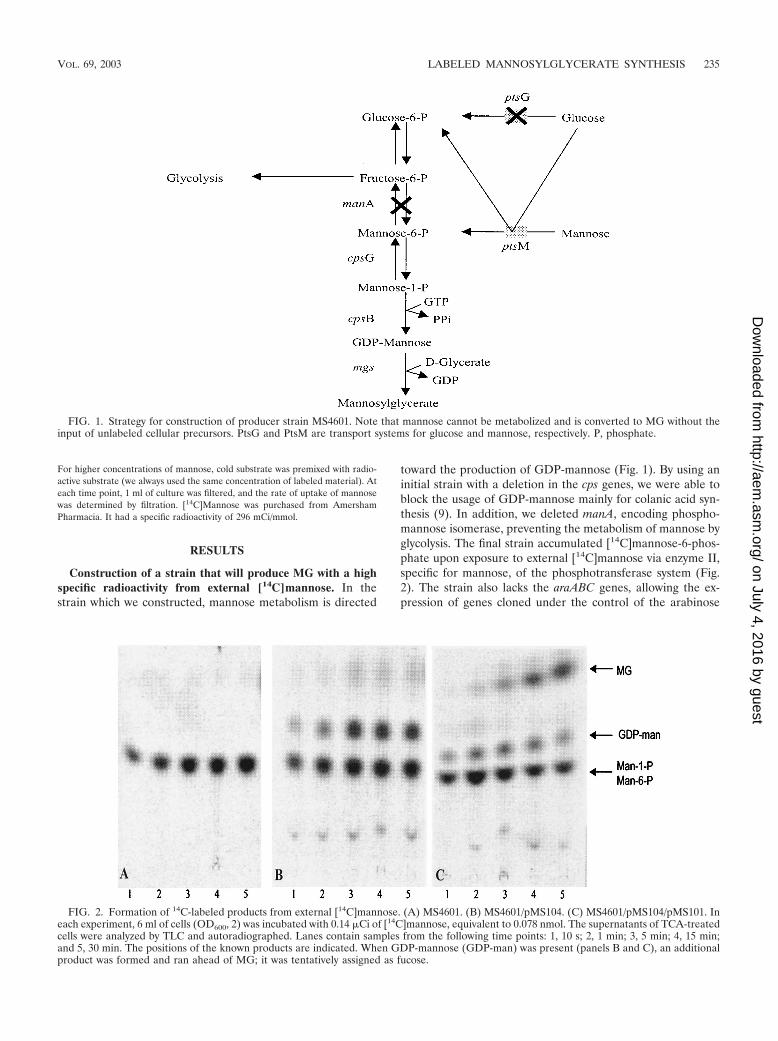
toward the production of GDP-mannose (Fig. 1). By using aninitial strain with a deletion in the cps genes, we were able toblock the usage of GDP-mannose mainly for colanic acid syn-thesis (9). In addition, we deleted manA, encoding phospho-mannose isomerase, preventing the metabolism of mannose byglycolysis. The final strain accumulated [14C]mannose-6-phos-phate upon exposure to external [14C]mannose via enzyme II,specific for mannose, of the phosphotransferase system (Fig.2). The strain also lacks the araABC genes, allowing the ex-pression of genes cloned under the control of the arabinose
FIG. 1. Strategy for construction of producer strain MS4601. Note that mannose cannot be metabolized and is converted to MG without theinput of unlabeled cellular precursors. PtsG and PtsM are transport systems for glucose and mannose, respectively. P, phosphate.
FIG. 2. Formation of 14C-labeled products from external [14C]mannose. (A) MS4601. (B) MS4601/pMS104. (C) MS4601/pMS104/pMS101. Ineach experiment, 6 ml of cells (OD600, 2) was incubated with 0.14 �Ci of [14C]mannose, equivalent to 0.078 nmol. The supernatants of TCA-treatedcells were analyzed by TLC and autoradiographed. Lanes contain samples from the following time points: 1, 10 s; 2, 1 min; 3, 5 min; 4, 15 min;and 5, 30 min. The positions of the known products are indicated. When GDP-mannose (GDP-man) was present (panels B and C), an additionalproduct was formed and ran ahead of MG; it was tentatively assigned as fucose.
VOL. 69, 2003 LABELED MANNOSYLGLYCERATE SYNTHESIS 235
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

promoter. A galU mutation (resulting in a defect in the syn-thesis of UDP-glucose) will prevent the synthesis of the inter-nal osmoprotectant trehalose (8).
Genes necessary for the expression of enzymes involved inthe synthesis of mannose-1-phosphate and GDP-mannose aswell as the enzyme MG synthase were introduced by using twoplasmids (pMS101 and pMS104).
Production of mannose-6-phosphate, GDP-mannose, andMG. Cells of strain MS4601 without plasmids, harboring plas-mid pMS104, or harboring plasmids pMS104 and pMS101were exposed to labeled mannose, and the internally formedproducts were analyzed by TLC (Fig. 2). In the absence of anyplasmid, one major spot, representing mannose-6-phosphate,was seen (Fig. 2A). In the presence of plasmid pMS104 (Fig.2B), the additional production of GDP-mannose was observed.Since mannose-6-phosphate and mannose-1-phosphate run atthe same positions in the solvent system used, the ratio of thesetwo compounds could not be determined. In the presence ofboth plasmids (Fig. 2C), an additional compound representingMG was detected. A weakly labeled spot running above MGwas also observed and tentatively assigned as fucose. The strainsynthesizing GDP-mannose but not MG synthesized smallamounts of a product running below mannose-1-phosphate ormannose-6-phosphate. This material was present in smalleramounts when MG synthase was present. Therefore, we be-lieve it to be a mannose product transferred to an unknownacceptor. Occasionally, we observed a spot running at the sameposition as free mannose. This spot was more prominent instrains lacking MG synthesis, indicating that GDP-mannose inthe absence of a suitable acceptor might be slowly degradedinternally to mannose.
Estimation of the rates of synthesis of MG and GDP-man-nose. Cells containing both plasmids were exposed to 0.078�M [14C]mannose in the presence of increasing amounts (upto 10 �M) of unlabeled mannose. The time-dependent forma-tion of labeled products was analyzed by TLC (Fig. 3). Under
the chosen conditions, external mannose was taken up by thecells in all experiments within 2 min. At a high specific radio-activity and a low external concentration of mannose (0.078�M), the synthesis of MG was immediate. Considering theinternal cell volume of the culture, the initial internal concen-tration of the precursors under these conditions was esti-mated to be about 80 �M. With decreasing specific activity andincreasing amounts of external mannose, the formation of la-beled MG seemingly declined, while free pools of mannose-6-phosphate or mannose-1-phosphate and GDP-mannose in-creased. We used an experiment in which cells were exposed to10 �M mannose (Fig. 3F) to estimate the in vivo rate ofproduction of MG. Under these conditions, the internal con-centration of GDP-mannose was on the order of 10 mM. Therate of MG formation was calculated by measuring the radio-activities of the different spots with phosphorimaging, takinginto account the dilution of the radioactivity. We found thatthe synthesis of MG (0.08 nmol/min/109 cells) was slow incomparison to the formation of GDP-mannose (0.25 nmol/min/109 cells). This has to be compared to the rate of mannoseuptake at 10 �M mannose, which is 2.9 nmol/min/109 cells (seebelow).
As an alternative method for extracting the labeled com-pounds, we treated the harvested cells with ice-cold water afterexposing them to different external concentrations of [14C]mannose for different times. The corresponding supernatantswere lyophilized and analyzed by TLC (Fig. 4). Again, it be-came apparent that the in vivo synthesis of MG was limited bythe conversion of GDP-mannose to MG. The cold-water shockreleased, without preference for any of the labeled compounds,at least 95% of the accumulated radioactivity, as demonstratedby TLC analysis of TCA-treated pellets (data not shown). Asshown in Fig. 4, when a higher initial concentration of MG(20 �M) was used, the percentage of radioactivity declinedover time. We believe this to be due to the release of labeledMG prior to the cold-water shock treatment. Indeed, small
FIG. 3. Time-dependent formation of [14C]MG from external [14C]mannose in MS4601/pMS104/pMS101. In each experiment, 6 ml of cells(OD600, 1) was incubated with 0.14 �Ci of [14C]mannose. Unlabeled mannose was added at various concentrations: A, 0.078 �M (no addition ofunlabeled mannose); B, 0.1 �M; C, 0.5 �M; D, 1 �M; E, 5 �M; and F, 10 �M. The supernatants of TCA-treated cells (5 �l) were analyzed byTLC and autoradiographed. Lanes contain samples from the following time points: 1, 10 s; 2, 5 min; 3, 20 min; 4, 40 min; and 5, 90 min.
236 SAMPAIO ET AL. APPL. ENVIRON. MICROBIOL.
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

amounts of labeled MG (only MG, no other labeled com-pound) could be detected in the medium supernatant prior tothe shock treatment (data not shown).
The synthesis of MG was confirmed by NMR analysis of thesupernatant solutions resulting from the cold-water shocktreatment of cells exposed to unlabeled mannose. The protonspectra showed that, in addition to MG, a variety of com-pounds were released into the external medium. The area ofthe anomeric proton resonance in MG was used for quantifi-cation, and an intracellular concentration of 5 mM was deter-mined (data not shown).
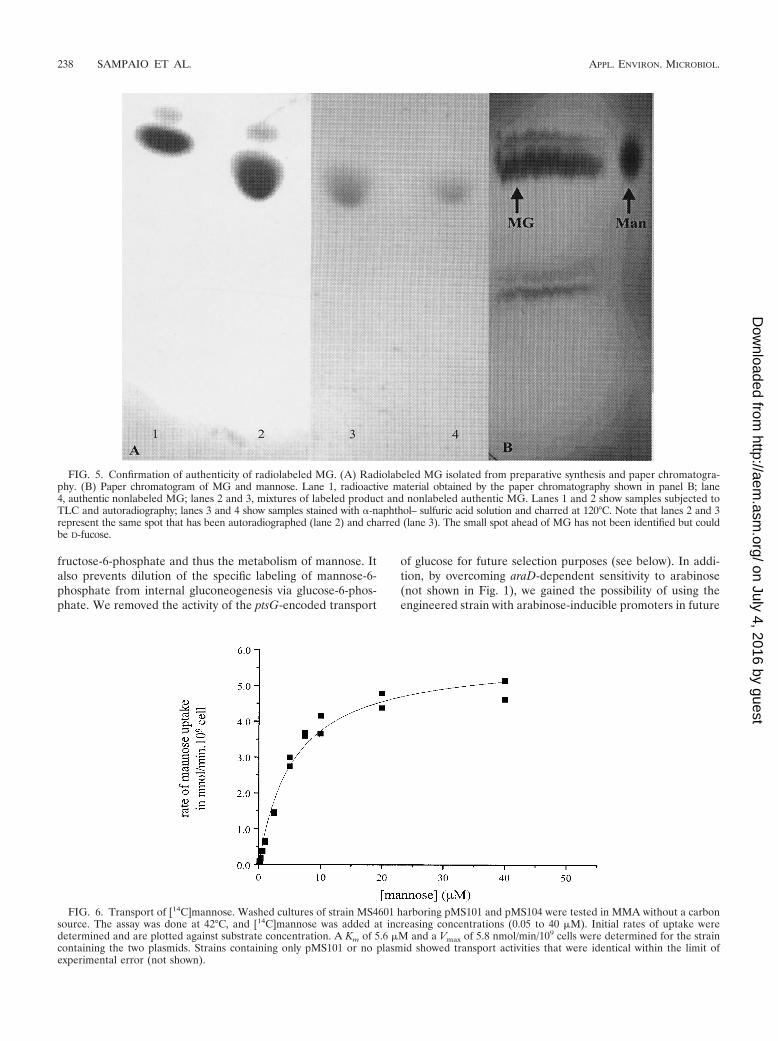
Preparative synthesis of [14C]MG. For the preparation ofradioactively labeled MG, undiluted [14C]mannose was used,and the final product was separated by paper chromatographyof material released from harvested cells by ice-cold watertreatment. The yield with respect to the input of mannose was56%. This value includes the loss of material during the dif-ferent procedures (cold shock, concentration, chromatogra-phy, and elution from paper). According to the metabolicscheme (Fig. 1), no dilution of the specific radioactivity shouldoccur. The isolated product was checked by TLC, and onemajor spot was detected. The radioactive material behaved likeauthentic nonlabeled MG (Fig. 5).
A sample of the medium supernatant (after cell harvest bycentrifugation) was analyzed by paper chromatography. Theonly radioactivity present (15% the initial radioactivity sup-plied as mannose) behaved like MG (data not shown). Thus,
internally produced MG does leak out, possibly carrier medi-ated, presumably after the concentration of internal MGreaches levels in the millimolar range.
Transport assays. The uptake of radioactive mannose wasanalyzed with MS4601, MS4601/pMS104, and MS4601/pMS104/pMS101 cells at different external mannose concentrations. In-itial rates were determined and plotted against mannose con-centrations. Figure 6 shows that uptake occurred with an ap-parent Km of 5.6 �M and a Vmax of 5.8 nmol/min/109 cells forthe strain with two plasmids. The Km and Vmax values obtainedfor MS4601 with pMS104 alone or with no plasmid were iden-tical within the limit of experimental error.
DISCUSSION
In this publication, we describe the production of labeledMG, an osmolyte that is preferentially found in thermophilicor hyperthermophilic prokaryotes (21). These organisms areuseful for isolating MG but do not allow the labeling of thiscompound, since labeled precursors added to the medium aredegraded in the cells and the precursors actually used for MGsynthesis are derived from internal metabolites and hence arenot labeled. E. coli does not produce MG and was thereforechosen for metabolic engineering to synthesize MG from ex-ternal mannose in the absence of mannose metabolism. Figure1 explains the simple strategy. The deletion of manA, encodingmannose-6-phosphate isomerase, prevents the formation of
FIG. 4. Release of labeled compounds after treatment with ice-cold water. MS4601/pMS101/pMS104 was grown in NZA medium with 0.2%glucose. Washed cultures were incubated with 1, 5, and 20 �M [14C]mannose containing the same total radioactivity (0.14 �Ci). At the indicatedtimes, 1-ml samples were withdrawn, harvested, and treated with ice-cold water. The supernatants were analyzed by TLC and autoradiographed.Time of harvest indicates the time of centrifugation; the radioactivity percentage indicates the amount of radioactivity with respect to the initialradioactivity present within the cells at the time indicated. Extraction with cold water released almost 100% of the radioactivity accumulated atthe time. The radioactivity remaining in the medium was not analyzed. P, phosphate.
VOL. 69, 2003 LABELED MANNOSYLGLYCERATE SYNTHESIS 237
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from
fructose-6-phosphate and thus the metabolism of mannose. Italso prevents dilution of the specific labeling of mannose-6-phosphate from internal gluconeogenesis via glucose-6-phos-phate. We removed the activity of the ptsG-encoded transport
of glucose for future selection purposes (see below). In addi-tion, by overcoming araD-dependent sensitivity to arabinose(not shown in Fig. 1), we gained the possibility of using theengineered strain with arabinose-inducible promoters in future
FIG. 5. Confirmation of authenticity of radiolabeled MG. (A) Radiolabeled MG isolated from preparative synthesis and paper chromatogra-phy. (B) Paper chromatogram of MG and mannose. Lane 1, radioactive material obtained by the paper chromatography shown in panel B; lane4, authentic nonlabeled MG; lanes 2 and 3, mixtures of labeled product and nonlabeled authentic MG. Lanes 1 and 2 show samples subjected toTLC and autoradiography; lanes 3 and 4 show samples stained with �-naphthol– sulfuric acid solution and charred at 120°C. Note that lanes 2 and 3represent the same spot that has been autoradiographed (lane 2) and charred (lane 3). The small spot ahead of MG has not been identified but couldbe D-fucose.
FIG. 6. Transport of [14C]mannose. Washed cultures of strain MS4601 harboring pMS101 and pMS104 were tested in MMA without a carbonsource. The assay was done at 42°C, and [14C]mannose was added at increasing concentrations (0.05 to 40 �M). Initial rates of uptake weredetermined and are plotted against substrate concentration. A Km of 5.6 �M and a Vmax of 5.8 nmol/min/109 cells were determined for the straincontaining the two plasmids. Strains containing only pMS101 or no plasmid showed transport activities that were identical within the limit ofexperimental error (not shown).
238 SAMPAIO ET AL. APPL. ENVIRON. MICROBIOL.
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

plasmid constructs. E. coli does contain the cpsG gene, encod-ing phosphomannose mutase (catalyzing the conversion ofmannose-6-phosphate to mannose-1-phosphate), and thecpsB gene, encoding GDP-mannose pyrophosphorylase (cata-lyzing the formation of GDP-mannose from GTP and man-nose-1-phosphate). However, these genes are contained in agene cluster responsible for the formation of colanic acid (2).Since the latter could interfere with the yield of MG, we choseto delete the cps cluster but introduced cpsB and cpsG on anIPTG-inducible plasmid. Finally, the mgs gene, encoding MGsynthase, was available only from a thermophilic organism (R.marinus). The activity of the encoded enzyme was optimal at85°C but very low at the ambient temperature of E. coli (12).Nevertheless, after induction, the enzyme was sufficiently ac-tive to detect the synthesis of MG (Fig. 2C and 3). The accu-mulation of mannose-1-phosphate or mannose-6-phosphatewas observed when GDP-mannose pyrophosphorylase was ab-sent (Fig. 2A and B), and the additional formation of GDP-mannose was observed in its presence (Fig. 2B). The fact thatthese compounds were metabolically stable proves the qualityof the engineered pathway.
The overall synthesis of MG appears to be limited mainly bythe transfer of �-mannosyl residues from GDP-mannose toD-glycerate. The external addition of glycerol or glycerate didnot increase the yield of MG (data not shown). Thus, theinternal pool of glycerate cannot be the limiting factor in MGsynthesis. Neither does mannose transport appear to be limit-ing in the producer strain, since the mannose uptake rate wasapproximately 70-fold higher than the rate of MG synthesis(5.8 compared to 0.08 nmol/min/109 cells). Surprisingly, therate of formation of GDP-mannose also was not very highdespite the fact that GDP-mannose pyrophosphorylase wasplasmid encoded and should have been present in largeamounts. This result suggests that the formation of GTP is thenext rate-limiting step after MG synthase activity is increased.
Engineered strain MS4601 is sensitive to mannose, mostlikely due to the accumulation of intermediates. By using thissensitivity as a selection marker, conceivably at increasing saltconcentrations, it might be possible to isolate mutations lead-ing to an overall higher rate of MG synthesis and thus increasethe concentration of internal MG as an effective protectiveosmolyte. Glucose used as a carbon source in the presence ofmannose would ensure the maintenance of PtsM as a port ofentrance for glucose and mannose, avoiding an undesiredmode of resistance (loss of mannose uptake). If successful, thestrain might also be used for the synthesis of large amounts ofunlabeled MG from mannose; in addition, with a small alter-ation in the strain (deletion of phosphofructokinase and intro-duction of wild-type mannose-6-phosphate isomerase), glucosemight be used as an inexpensive starting material for the syn-thesis of MG.
Although limited by several shortcomings (presence of twoplasmids and low rate of MG synthesis), the producer strainallows the high-yield formation of labeled MG from labeledmannose without the loss of specific radioactivity. Labeled MGcan now be used for studying the properties of this novelosmolyte. After exposure to limited amounts of mannose, up-take of mannose into cells, and the formation of internal MG,we did detect small quantities of labeled MG in the mediumwithout other labeled compounds. Thus, this material must
have leaked from the cells in a selective process. This obser-vation suggests the possibility of optimizing the production andselective release of MG after cloning of the responsible exitcarrier for MG.
ACKNOWLEDGMENTS
We gratefully acknowledge the receipt of bacterial strains from IanBooth, Jacqueline Plumbridge, and Susan Gottesman and plasmidsfrom Milton S. Da Costa and Miguel Valvano. We thank Tanja Epplerand Reinhold Horlacher for suggestions on plasmid and strain con-structions and Christoph Mayer for help with chromatography.
This work was supported by the European Community 5th Frame-work Program (project QLK3-CT-2000-00640) and by POCTI andFEDER (project 3571/BIO/2000). M.-M.S. was the recipient of PhDgrant BD71180/2000, FCT, Portugal. Support from the Fonds derChemischen Industrie is also acknowledged.
REFERENCES
1. Amann, E., B. Ochs, and K. J. Abel. 1988. Tightly regulated tac promotervectors useful for the expression of unfused and fused proteins in Esche-richia coli. Gene 69:301–315.
2. Andrianopoulos, K., L. Wang, and P. R. Reeves. 1998. Identification of thefucose synthetase gene in the colanic acid gene cluster of Escherichia coliK-12. J. Bacteriol. 180:998–1001.
3. Borges, N., A. Ramos, N. D. H. Raven, R. J. Sharp, and H. Santos. 2002.Comparative study of the thermostabilizing properties of mannosylglycerateand other compatible solutes on model enzymes. Extremophiles 6:209–216.
4. Bouveng, H., B. Lindberg, and B. Wickberg. 1955. Low-molecular carbohy-drates in algae. Acta Chem. Scand. 9:807–809.
5. Casadaban, M. J. 1976. Transposition and fusion of the lac genes to selectedpromoters in Escherichia coli using bacteriophage lambda and Mu. J. Mol.Biol. 104:541–555.
6. Chang, A. C., and S. N. Cohen. 1978. Construction and characterization ofamplifiable multicopy DNA cloning vehicles derived from the P15A crypticminiplasmid. J. Bacteriol. 134:1141–1156.
7. Empadinhas, N., J. D. Marugg, N. Borges, H. Santos, and M. S. da Costa.2001. Pathway for the synthesis of mannosylglycerate in the hyperthermo-philic archaeon Pyrococcus horikoshii. Biochemical and genetic character-ization of key enzymes. J. Biol. Chem. 276:43580–43588.
8. Giaever, H. M., O. B. Styrvold, I. Kaasen, and A. R. Strom. 1988. Biochem-ical and genetic characterization of osmoregulatory trehalose synthesis inEscherichia coli. J. Bacteriol. 170:2841–2849.
9. Gottesman, S., and V. Stout. 1991. Regulation of capsular polysaccharidesynthesis in Escherichia coli K12. Mol. Microbiol. 5:1599–1606.
10. Horlacher, R., and W. Boos. 1997. Characterization of TreR, the majorregulator of the Escherichia coli trehalose system. J. Biol. Chem. 272:13026–13032.
11. Jacin, H., and A. R. Mishkin. 1965. Separation of carbohydrates on borate-impregnated silica gel G plates. J. Chromatogr. 18:170–173.
12. Martins, L. O., N. Empadinhas, J. D. Marugg, C. Miguel, C. Ferreira, M. S.da Costa, and H. Santos. 1999. Biosynthesis of mannosylglycerate in thethermophilic bacterium Rhodothermus marinus. Biochemical and geneticcharacterization of a mannosylglycerate synthase. J. Biol. Chem. 274:35407–35414.
13. McLaggan, D., M. A. Jones, G. Gouesbet, N. Levina, S. Lindey, W. Epstein,and I. R. Booth. 2002. Analysis of the kefA2 mutation suggests that KefA isa cation-specific channel involved in osmotic adaptation in Escherichia coli.Mol. Microbiol. 43:521–536.
14. Meissner, P. S., W. P. Sisk, and M. L. Berman. 1987. Bacteriophage lambdacloning system for the construction of directional cDNA libraries. Proc. Natl.Acad. Sci. USA 84:4171–4175.
15. Miller, J. H. 1972. Experiments in molecular genetics. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
16. Nuoffer, C., B. Zanolari, and B. Erni. 1988. Glucose permease of Escherichiacoli. The effect of cysteine to serine mutations on the function, stability, andregulation of transport and phosphorylation. J. Biol. Chem. 263:6647–6655.
17. Plumbridge, J. 1998. Expression of ptsG, the gene for the major glucose PTStransporter in Escherichia coli, is repressed by Mlc and induced by growth onglucose. Mol. Microbiol. 29:1053–1063.
18. Postma, P. W., J. W. Lengeler, and G. R. Jacobson. 1993. Phosphoenolpyru-vate:carbohydrate phosphotransferase systems of bacteria. Microbiol. Rev.57:543–594.
19. Ramos, A., N. Raven, R. Sharp, S. Bartolucci, M. Rossi, R. Cannio, J.Lebbink, J. Van Der Oost, W. De Vos, and H. Santos. 1997. Stabilization ofenzymes against thermal stress and freeze-drying by mannosylglycerate.Appl. Environ. Microbiol. 63:4020–4025.
20. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
VOL. 69, 2003 LABELED MANNOSYLGLYCERATE SYNTHESIS 239
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from

21. Santos, H., and M. S. da Costa. 2001. Organic solutes from thermophiles andhyperthermophiles. Methods Enzymol. 334:302–315.
22. Schlegel, A., O. Danot, E. Richet, T. Ferenci, and W. Boos. 2002. The Nterminus of the Escherichia coli transcription activator MalT is the domain ofinteraction with MalY. J. Bacteriol. 184:3069–3077.
23. Silva, Z., N. Borges, L. O. Martins, R. Wait, M. S. da Costa, and H. Santos.1999. Combined effect of the growth temperature and salinity of the mediumon the accumulation of compatible solutes by Rhodothermus marinus andRhodothermus obamensis. Extremophiles 3:163–172.
24. Singer, M., T. A. Baker, G. Schnitzler, S. M. Deischel, M. Goel, W. Dove,K. J. Jaacks, A. D. Grossman, J. W. Erickson, and C. A. Gross. 1989. Acollection of strains containing genetically linked alternating antibiotic re-sistance elements for genetic mapping of Escherichia coli. Microbiol. Rev.53:1–24.
25. Stevenson, G., K. Andrianopoulos, M. Hobbs, and P. R. Reeves. 1996. Or-ganization of the Escherichia coli K-12 gene cluster responsible for produc-tion of the extracellular polysaccharide colanic acid. J. Bacteriol. 178:4885–4893.
240 SAMPAIO ET AL. APPL. ENVIRON. MICROBIOL.
on July 4, 2016 by guesthttp://aem
.asm.org/
Dow
nloaded from
Related Documents











