Please cite this article in press as: Fonseca-Maldonado R, et al. Synergistic action of co-expressed xylanase/laccase mixtures against milled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/10.1016/j.procbio.2014.03.027 ARTICLE IN PRESS G Model PRBI-10109; No. of Pages 10 Process Biochemistry xxx (2014) xxx–xxx Contents lists available at ScienceDirect Process Biochemistry jo ur nal home p age: www.elsevier.com/locate/procbio Synergistic action of co-expressed xylanase/laccase mixtures against milled sugar cane bagasse Raquel Fonseca-Maldonado a , Lucas F. Ribeiro a , Gilvan P. Furtado a , Letícia M. Arruda a , Luana P. Meleiro b , Juliana S. Alponti b , Carla Botelho-Machado b,e , Davi S. Vieira c , Eric Bonneil d , Rosa dos Prazeres Melo Furriel b , Pierre Thibault d , Richard J. Ward b,e,∗ a Departamento de Bioquímica e Imunologia, FMRP-USP, Universidade de São Paulo, São Paulo, Brazil b Departamento de Química, FFCLRP-USP, Universidade de São Paulo, São Paulo, Brazil c Instituto de Química, Universidade Federal do Rio Grande do Norte – Natal, Rio Grande do Norte, Brazil d Institute for Research in Immunology and Cancer, Université de Montréal, Canada e Laboratório Nacional de Ciência e Tecnologia do Bioetanol – CTBE/CNPEM, Campinas, São Paulo, Brazil a r t i c l e i n f o Article history: Received 10 December 2013 Received in revised form 31 January 2014 Accepted 28 March 2014 Available online xxx Keywords: Xylanase A CotA Synergic activity Mass spectrometry Pichia pastoris a b s t r a c t The primary plant cell wall is composed of cellulose, hemicellulose, lignin and protein in a stable matrix. The concomitant depolymerization of lignin by laccase and of hemicelluloses by xylanase can improve lignocellulose degradation in the production of second generation biofuels. A thermophilic variant of xylanase A (XynAG3) and the thermostable laccase (CotA), both from Bacillus subtilis, were produced in co-transformed Pichia pastoris strain GS115. Mobility changes in SDS-PAGE after Endo H digestion indicated that both enzymes were glycosylated. The maximum catalytic activity of the XynAG3 Pp and the CotA Pp was observed at 58 ◦ C and 75 ◦ C, respectively, and both enzymes presented high activity at pH 5.0. The half-life at 60 ◦ C of XynAG3 Pp and CotA Pp was 150 min and 540 min, respectively. The relative levels of CotA Pp and XynAG3 Pp in culture broths were altered by the concentration of methanol used for induction, and CotA Pp :XynAG3 Pp ratios of 1:1.5 and 1:2 were evaluated against milled sugar-cane bagasse. The highest activity was observed at a 1:2 ratio of CotA Pp :XynAG3 Pp , and was 44% higher as compared to the sum of the activities of the individual enzymes in the same assay conditions. These results demonstrate the synergistic action between an endoxylanase and a laccase against the natural lignocellulosic substrate. © 2014 Elsevier Ltd. All rights reserved. 1. Introduction Vegetal lignocellulosic material is a principal component of ter- restrial biomass and consists mainly of cellulose, hemicellulose and lignin, in variable amounts depending on the plant source. Cellulose is embedded in a complex matrix of hemicellulose and lignin [1], and therefore delignification is of importance for the exploitation of cellulose in industrial processes. Laccases (EC 1.10.3.2) decompose lignin by oxidation of a wide range of aromatic and polyphenolic compounds, which is coupled to the reduction of dioxygen to water [2], and xylanases (EC 3.2.1.8) depolymerize xylan, the second most abundant biopolymer after cellulose and the major hemicellulosic ∗ Corresponding author at: Departamento de Química – FFCLRP, Universidade de São Paulo, Avenida Bandeirantes 3900, Ribeirão Preto, SP, Brazil. Tel.: +55 16 3602 3875; fax: +55 16 3602 3848. E-mail address: [email protected] (R.J. Ward). polysaccharide found in plant cell walls [3]. Applications of mix- tures of laccases and xylanases have been reported [4], and more recently these enzymes have attracted considerable interest due to their potential for application in ethanol production using ligno- cellulosic residues, since removal of lignin and hemicelluloses can significantly enhance the hydrolysis of cellulose to release reducing sugars for subsequent fermentation to ethanol [5]. Laccases coordinate the binding of four copper atoms in three types of sites that differ in their environment and spectroscopic properties [6]. Redox potentials of laccases are usually lower in comparison with those of other lignin-modifying enzymes such as lignin peroxidase or manganese peroxidase [7], and they act in the degradation of polymers, the ring cleavage of aromatic compounds and the cross-linking of monomers. The best-studied bacterial lac- case is the CotA, the endospore coat component of Bacillus subtilis. The cotA gene encodes a 65-kDa protein that exhibits a high ther- mal stability with a half-life of 2 h at 80 ◦ C and an optimum catalytic temperature at 75 ◦ C [8]. http://dx.doi.org/10.1016/j.procbio.2014.03.027 1359-5113/© 2014 Elsevier Ltd. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

P
Sm
RLEa
b
c
d
e
a
ARRAA
KXCSMP
1
rliaclc[a
ST
h1
ARTICLE IN PRESSG ModelRBI-10109; No. of Pages 10
Process Biochemistry xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Process Biochemistry
jo ur nal home p age: www.elsev ier .com/ locate /procbio
ynergistic action of co-expressed xylanase/laccase mixtures againstilled sugar cane bagasse
aquel Fonseca-Maldonadoa, Lucas F. Ribeiroa, Gilvan P. Furtadoa, Letícia M. Arrudaa,uana P. Meleirob, Juliana S. Alpontib, Carla Botelho-Machadob,e, Davi S. Vieirac,ric Bonneild, Rosa dos Prazeres Melo Furrielb, Pierre Thibaultd, Richard J. Wardb,e,∗
Departamento de Bioquímica e Imunologia, FMRP-USP, Universidade de São Paulo, São Paulo, BrazilDepartamento de Química, FFCLRP-USP, Universidade de São Paulo, São Paulo, BrazilInstituto de Química, Universidade Federal do Rio Grande do Norte – Natal, Rio Grande do Norte, BrazilInstitute for Research in Immunology and Cancer, Université de Montréal, CanadaLaboratório Nacional de Ciência e Tecnologia do Bioetanol – CTBE/CNPEM, Campinas, São Paulo, Brazil
r t i c l e i n f o
rticle history:eceived 10 December 2013eceived in revised form 31 January 2014ccepted 28 March 2014vailable online xxx
eywords:ylanase AotAynergic activity
a b s t r a c t
The primary plant cell wall is composed of cellulose, hemicellulose, lignin and protein in a stable matrix.The concomitant depolymerization of lignin by laccase and of hemicelluloses by xylanase can improvelignocellulose degradation in the production of second generation biofuels. A thermophilic variant ofxylanase A (XynAG3) and the thermostable laccase (CotA), both from Bacillus subtilis, were producedin co-transformed Pichia pastoris strain GS115. Mobility changes in SDS-PAGE after Endo H digestionindicated that both enzymes were glycosylated. The maximum catalytic activity of the XynAG3Pp andthe CotAPp was observed at 58 ◦C and 75 ◦C, respectively, and both enzymes presented high activity atpH 5.0. The half-life at 60 ◦C of XynAG3Pp and CotAPp was 150 min and 540 min, respectively. The relativelevels of CotAPp and XynAG3Pp in culture broths were altered by the concentration of methanol used
ass spectrometryichia pastoris
for induction, and CotAPp:XynAG3Pp ratios of 1:1.5 and 1:2 were evaluated against milled sugar-canebagasse. The highest activity was observed at a 1:2 ratio of CotAPp:XynAG3Pp, and was 44% higher ascompared to the sum of the activities of the individual enzymes in the same assay conditions. Theseresults demonstrate the synergistic action between an endoxylanase and a laccase against the naturallignocellulosic substrate.
© 2014 Elsevier Ltd. All rights reserved.
. Introduction
Vegetal lignocellulosic material is a principal component of ter-estrial biomass and consists mainly of cellulose, hemicellulose andignin, in variable amounts depending on the plant source. Celluloses embedded in a complex matrix of hemicellulose and lignin [1],nd therefore delignification is of importance for the exploitation ofellulose in industrial processes. Laccases (EC 1.10.3.2) decomposeignin by oxidation of a wide range of aromatic and polyphenolic
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
ompounds, which is coupled to the reduction of dioxygen to water2], and xylanases (EC 3.2.1.8) depolymerize xylan, the second mostbundant biopolymer after cellulose and the major hemicellulosic
∗ Corresponding author at: Departamento de Química – FFCLRP, Universidade deão Paulo, Avenida Bandeirantes 3900, Ribeirão Preto, SP, Brazil.el.: +55 16 3602 3875; fax: +55 16 3602 3848.
E-mail address: [email protected] (R.J. Ward).
ttp://dx.doi.org/10.1016/j.procbio.2014.03.027359-5113/© 2014 Elsevier Ltd. All rights reserved.
polysaccharide found in plant cell walls [3]. Applications of mix-tures of laccases and xylanases have been reported [4], and morerecently these enzymes have attracted considerable interest due totheir potential for application in ethanol production using ligno-cellulosic residues, since removal of lignin and hemicelluloses cansignificantly enhance the hydrolysis of cellulose to release reducingsugars for subsequent fermentation to ethanol [5].
Laccases coordinate the binding of four copper atoms in threetypes of sites that differ in their environment and spectroscopicproperties [6]. Redox potentials of laccases are usually lower incomparison with those of other lignin-modifying enzymes such aslignin peroxidase or manganese peroxidase [7], and they act in thedegradation of polymers, the ring cleavage of aromatic compoundsand the cross-linking of monomers. The best-studied bacterial lac-
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
case is the CotA, the endospore coat component of Bacillus subtilis.The cotA gene encodes a 65-kDa protein that exhibits a high ther-mal stability with a half-life of 2 h at 80 ◦C and an optimum catalytictemperature at 75 ◦C [8].

ING ModelP
2 cess B
txoaGGs[
mctareiweipcmcateaekb
2
2
xa1dcG3wtcGarmatrpiatwwcoa5a
ARTICLERBI-10109; No. of Pages 10
R. Fonseca-Maldonado et al. / Pro
Endo-1,4-�-xylanases (EC 3.2.1.8) catalyze the hydrolysis ofhe �-1,4 glycosidic bonds in the xylan backbone to produceylooligosaccharides of varying length [9]. Xylanases from vari-us bacteria, fungi and yeasts have been characterized [10], andre found predominantly in glycosylhydrolase families GH5, GH8,H10 and GH11 and also in other families including GH7, GH16,H30, GH43 and GH62 [11]. The xylanase A from the mesophile B.
ubtilis (XynA) is a GH11 enzyme, and has been extensively studied12,13].
Several yeast expression systems have been developed for com-ercial production of recombinant proteins, and one of the most
ommonly used systems is the methylotrophic yeast Pichia pas-oris, where expression is driven by the strong promoter for thelcohol oxidase I (AOX1), which is induced by methanol and isepressed by other carbon sources such as glucose, glycerol, andthanol [14]. Another important feature of this system is the capac-ty for growth to very high cell densities in well-defined medium,
hich contributes to the efficient production and secretion of het-rologous proteins [15]. For many applications the cost of enzymess one of the main factors determining the economic viability of arocess. Cost reduction may be achieved by optimizing the reactiononditions so as to be compatible with temperature and pH require-ents of the enzymes, increasing enzyme production to lower unit
ost, and using enzyme combinations that promote synergy of cat-lytic function. In this study we have investigated the ability ofhe eukaryote P. pastoris to efficiently co-express two B. subtilisnzymes, a thermophilic XynA derived from directed evolutionnd the CotA. The pH and temperature optima of the heterologousnzymes were characterized, together with their thermal stability,inetics and synergistic effects against an insoluble lignocellulosiciomass.
. Methodology
.1. Construction of P. pastoris strains
The coding sequence of a thermophilic variant of theylanase A (xynAG3) created by directed evolution [16]nd the laccase gene, cotA, from Bacillus subtilis (BGSCA1 code 168), were cloned into pET28a as previouslyescribed [4]. A PCR reaction was performed with theonstruct pET28 cotA as template and the primers LacPN (5′-GCAGCCTCGAGAAAAGAGAGGCTGAAGCTAGCAGCCATCATCAT-′), LacPC (5′-GGGCCTAGGTTATTTATGGGGATCAGTTAT-3′)hich introduced restriction sites for XhoI and AvrII, respec-
ively (underlined sequences). Using the pET28 xynAG3onstruct as a PCR template, the primers XylPN (5′-GCAGCCTCGAGAAAAGAGAGGCTGAAGCTAGCACAGACTAC-3′)nd XylPC (5′-GATCAGCGGCCGCCGTTAGCTACCC-3′) introducedestriction sites for XhoI and NotI, respectively, in the ther-ophilic variant XynAG3. The PCR products encoding the CotA
nd the XynAG3 were digested with the appropriate restric-ion enzymes and cloned into plasmids pPIC9K and pPICZ�A,espectively. The pPIC9k cotA construct included an N-terminalolyhistidine sequence, and the pPICZ�A xynAG3 construct
ncluded a C-terminal polyhistidine sequence. The pPIC9k cotAnd pPICZ�A xynAG3 constructs were linearized with the restric-ion enzymes StuI and SacI, respectively, and the linearized DNAas used for transformation of P. pastoris GS115 by electroporationith a Gene Pulser (BioRad, Hercules, CA, USA). The linearized
onstruct pPIC9k cotA was inserted into the HIS4 gene by homol-
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
gous recombination and selected by histidine auxotrophismnd the linearized pPICZ�A xynAG3 was inserted within the′ AOX1 region and selected by zeocin resistance. To obtain
double transformant, the previously constructed P. pastoris
PRESSiochemistry xxx (2014) xxx–xxx
GS115/pPICZ�A xynAG3 was electroporated with the pPIC9k cotAconstruct. The strain transformed with double construct (P. pas-toris GS115/pPICZ�A xynAG3/pPIC9k cotA) was selected by zeocin(500 �g mL−1) resistance on MD agar plates (0.34% yeast nitrogenbase, 1% ammonium sulfate, 4 × 10−5% (w/v) biotin, 100 mMphosphate buffer, pH 6.0, 2% dextrose, 1.5% bacteriological agar).
2.2. Protein expression and purification
Both XynAG3 and CotA were expressed individually and ina coexpression mixture. For expression of the XynAG3, a sin-gle colony of P. pastoris GS115/pPICZ�A xynAG3 was inoculatedinto 100 mL of BMG medium (1.34% yeast nitrogen base withamino acids, 4 × 10−5% (w/v) biotin, 100 mM potassium phos-phate, pH 6.0, and 1.0% (v/v) glycerol) and grown at 30 ◦C in anorbital shaker until the culture reached an OD600 nm of 3. Cellswere harvested by centrifugation at 3000 × g for 5 min, and thecell pellet was resuspended to an OD600 nm of 1 in 100 mL BMMmedium (BMG medium in which glycerol was substituted witheither 0.5% or 1% methanol) and the culture was grown for 6–8days at 30 ◦C. Every 24 h methanol was added, and at the end ofthe culture, the cells were centrifuged (3000 × g, 15 min) and thesupernatant was stored at 4 ◦C. For expression of the CotA andXynAG3/CotA mixtures, P. pastoris GS115/pPIC9k cotA and the P.pastoris GS115/pPICZ�A xynAG3/pPIC9k cotA, the yeast nitrogenbase with amino acids was replaced with 0.34% yeast nitrogenbase without ammonium sulfate and without amino acids plus 1%ammonium sulfate and 0.25 mM CuSO4.
The supernatant expression medium of individual enzymesexpressed in P. pastoris, Xylanase AG3 (XynAG3Pp) and Laccase(CotAPp), was cleared of cell debris by centrifugation at 5000 g for7 min at 4 ◦C and filtration with a 0.2 �m filter. The filtrate wasconcentrated and the buffer exchanged with an Amicon StirredCell 10000 MWCO (Merck Millipore, Billerica, MA, USA). The con-centrate was applied to a HiTrapTM 3 mL Q-HP column (Promega,Madison, WI, USA) previously equilibrated with 500 mM NaCl,40 mM HEPES buffer pH 8.0. The column was washed with buffercontaining 40 mM HEPES pH 8.0, 300 mM NaCl, and 40 mM imi-dazole. Proteins were eluted with 40 mM HEPES buffer pH 8.0,500 mM NaCl and 300 mM imidazole at a flow rate of 1.5 mL min−1.Fractions containing the enzymes were concentrated and desaltedusing an Amicon Ultra-15 10000 MWCO (Merck Millipore, Bil-lerica, MA, USA). Protein purity was routinely checked usingSDS-polyacrylamide gel electrophoresis (SDS-PAGE).
2.3. Deglycosylation of recombinant proteins by endoglycosidaseH
The purified recombinant protein samples (10–20 �g) wereboiled for 5 min in 50 mM sodium citrate buffer (pH 5.5) contain-ing 1% SDS. After cooling, the samples were mixed with 250 U ofendoglycosidase H (Endo-H) (New England Biolab, Ipswich, MA,USA) and incubated at 37 ◦C for 3 h. The samples were subsequentlyanalyzed by SDS-PAGE and mass spectrometry.
2.4. Mass spectrometry analysis
After SDS-PAGE analysis, slices containing the protein bandsof the Endo H digested samples were cut from the gel, destainedwith 50% methanol, treated with 10 mM DTT (dithiothreitol) for1 h at 56 ◦C and alkylated with 55 mM chloroacetamide for 1 h at
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
25 ◦C. After washing in 50 mM ammonium bicarbonate, the gelslices were shrunk in 100% acetonitrile. Digestion with trypsin wasperformed in 50 mM ammonium bicarbonate for 8 h at 37 ◦C withagitation. The peptides were extracted with 90% acetonitrile/0.5 M

ING ModelP
cess B
ut
(eWC1n5rtTdoMetwHm
2
c1otab
2
3s0N
2t
omuseatTw5apsst
wCTuS
ARTICLERBI-10109; No. of Pages 10
R. Fonseca-Maldonado et al. / Pro
rea and dried under vacuum. Samples were solubilized in 5% ace-onitrile/0.2% formic acid prior to mass spectrometry analyses.
LC–MS/MS analyses were conducted on a nano-LC 2D pumpEksigent) coupled to a LTQ-Orbitrap Elite hybrid mass spectrom-ter via a nanoelectrospray ion source (Thermo Fisher Scientific,altham, MA). Peptides were loaded on a 360 �m ID × 4 mm,
18 trap column (prepared in house) prior to separation on a50 �m ID × 10 cm nano-LC column (Jupiter C18, 3 �m, 300 A, Phe-omex). Peptide elution was achieved using a linear gradient of–60% acetonitrile (0.2% formic acid) over 56 min with a flowate set at 600 nL min−1. Each full MS spectrum was followed bywelve MS/MS spectra on the most abundant multiply charged ions.andem MS experiments were performed using collision-inducedissociation in the high energy collision cell prior to mass analysisn the Orbitrap analyzer. The data were processed using the 2.3ascot (Matrix Science, Boston, MA, USA) search engine with tol-
rance parameters set to 15 ppm and 0.02 Da for the precursor andhe fragment ions, respectively. The selected variable modificationsere carbamidomethyl (C), deamidation (NQ), oxidation (M) andexNac (N). Tandem mass spectra were searched against a custom-ade database derived from the CotA amino-acid sequence.
.5. Circular dichroism spectroscopy
Purified protein samples of XynAG3Pp and CotAPp at con-entrations of 2 �M and 3.1 �M, respectively, were prepared in0 mM phosphate buffer (pH6.0). Circular dichroism (CD) spectraf XynAG3Pp and CotAPp were measured with a JASCO J-810 spec-ropolarimeter (JASCO Corporation, Tokyo, Japan) at 25 ◦C using
0.1 mm path-length quartz cuvette. The average of six scansetween 180 and 260 nm was recorded for each protein.
.6. UV–vis spectroscopy
UV–vis absorption spectra of CotAPp were measured between00 and 800 nm in 1.0-cm quartz cuvettes using a Cary 50 UV–vispectrophotometer. Measurements were performed at 25 ◦C using.5 mg mL−1 of protein in 10 mM HEPES buffer (pH7.5) and 100 mMaCl.
.7. Response surface methodology for estimation of pH andemperature optima
CotAPp activity was determined by the rate of oxidationf 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) byeasuring the increase in A420 (ε = 36000 M−1 cm−1). One activity
nit was defined as the amount of enzyme that oxidized 1 �mol ofubstrate per minute. Response Surface Methodology was used tostimate the optimum pH and temperature conditions for the CotActivity, in which a Central Composite Design (k = 2) with five cen-ral points and a total of 13 experiments (shown in supplementaryable S1A) was used. In all assays, 10 �L (1 �M) of purified enzymeas mixed with 0.5 mL of an aqueous ABTS solution (5 mM ABTS,
% PEG 5000) and 0.49 mL of 100 mM citrate/phosphate bufferdjusted to different pH and heated for 2 min at different tem-eratures in a Peltier heating system coupled to a Cary 50 UV–vispectrophotometer (Agilent Technologies, Santa Clara, CA). Theoftware Statistica 8 (StatSoft, Tulsa, OK, USA) was used to evaluatehe experimental data.
The optimum pH and temperature for the XynAG3Pp activityere also determined by Response Surface Methodology, and the
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
entral Composite Design experiment is shown in supplementaryable S1B. The xylanase activity of the XynAG3Pp was measuredsing 1% (w/v) oat spelt xylan as substrate (Sigma–Aldrich, Buchs,G, Switzerland) in 100 mM citrate/phosphate buffer adjusted to
PRESSiochemistry xxx (2014) xxx–xxx 3
different pH values. Data analysis was performed using the Statis-tica 8 software as described in the previous paragraph.
2.8. Enzyme kinetics measurements
The effect of the substrate concentration on the activity of puri-fied CotAPp was determined at 37 ◦C in 100 mM sodium acetatebuffer (pH 4.5) using increasing concentrations of ABTS (5–320 �M)in 1-cm cuvettes. Reactions were initiated by addition of 10 �L(3.1 �M) of the purified proteins, and the rates of oxidationobtained as described previously [4]. All enzymatic assays wereperformed in triplicate. The kinetic parameters maximum veloc-ity (Vmax), apparent dissociation constant for the substrate (K0.5),and catalytic constant (kcat) for the ABTS oxidation were calculatedby nonlinear regression fitting of the data to the semi-logarithmicform of the Hill equation using the software SigrafW [17]. The ther-mal stability was evaluated by incubating the purified enzyme at80 ◦C and 65 ◦C. After appropriate times samples were withdrawn,cooled, and residual activity was measured using the ABTS oxida-tion assay at 37 ◦C.
The effect of substrate concentration on xylanase activity wasstudied using oat spelt xylan as substrate at concentrations rangingfrom 1 to 12 mg mL−1 in 100 mM acetate buffer (pH 6.0). All reac-tions were initiated by addition of 20 �L of the purified enzymes(2 �M) to a final reaction volume of 1 mL, and after 10 min incuba-tion reducing sugars release was determined by the dinitrosalicylicacid (DNS) procedure [18]. All enzymatic assays were performedin triplicate, and the kinetic parameters were calculated using theSigrafW software [17]. Thermal stability was evaluated by incuba-tion of the purified enzymes at 60 ◦C and 70 ◦C, and residual activitywas measured at 37 ◦C using aliquots collected after different times.
2.9. Hydrolysis of milled sugarcane bagasse by mixtures ofXynAG3 and CotA
The enzymatic hydrolysis of a natural lignocellulosic sub-strate was performed using milled sugarcane bagasse (particle size0.2 mm), treated with 80% ethanol and washed thoroughly with100 mM acetate buffer (pH 5.0) to remove residual soluble sugars.A 1% (w/v) suspension of the treated and washed substrate wasprepared in the same buffer and mixed with 20× concentratedcrude co-expression culture supernatants after 120 h inductionwith either 0.5% or 1% methanol, or with the supernatants fromindividual cultures of the P. pastoris lines expressing XynAG3 andCotA. The reaction was incubated at 60 ◦C for 3 h in an orbitalshaker at 200 rpm to avoid substrate sedimentation, and the totalrelease of reducing sugar was measured by the DNS method [18].All experiments were carried out both in the presence and theabsence of 0.5 mM ABTS. After determination of the enzymaticactivities (U mL−1) of the two enzymes in the supernatants fromindividual cultures of the P. pastoris lines expressing CotAPp andXynAG3Pp, experiments measuring the total reducing sugar releaseusing milled sugarcane bagasse were also performed in which theCotAPp:XynAG3Pp molar ratio was varied from 5:1 to 1:5.
3. Results
3.1. Expression and purification
The genes encoding the laccase (CotA) and a thermophilic vari-ant of the xylanase A (XynAG3) from B. subtilis were successfullyengineered for heterologous expression in P. pastoris as active
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
secreted enzymes (CotAPp and XynAG3Pp, respectively) that accu-mulated in the culture broth. The catalytic activities of the twoenzymes in supernatants from 6 to 8 day cultures with inductionof protein expression by 0.5% or 1% (v/v) methanol were analyzed
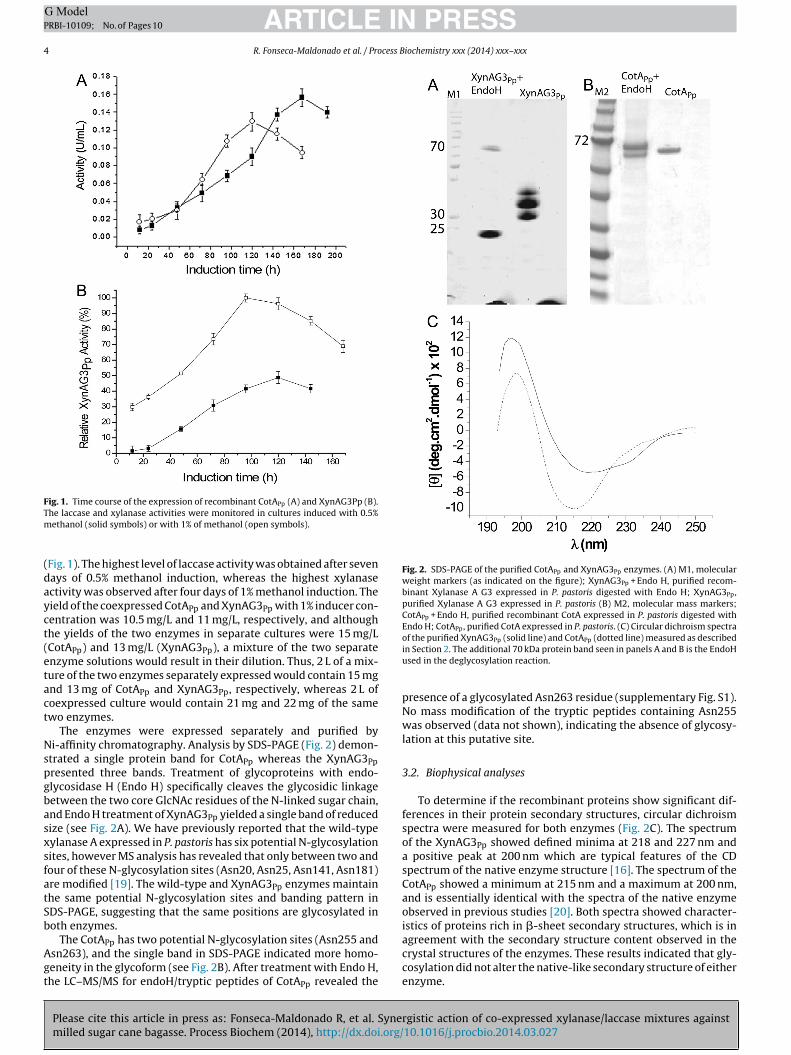
ARTICLE IN PRESSG ModelPRBI-10109; No. of Pages 10
4 R. Fonseca-Maldonado et al. / Process Biochemistry xxx (2014) xxx–xxx
Fig. 1. Time course of the expression of recombinant CotAPp (A) and XynAG3Pp (B).Tm
(dayct(etact
NspgbasxsfatSb
Agt
Fig. 2. SDS-PAGE of the purified CotAPp and XynAG3Pp enzymes. (A) M1, molecularweight markers (as indicated on the figure); XynAG3Pp + Endo H, purified recom-binant Xylanase A G3 expressed in P. pastoris digested with Endo H; XynAG3Pp,purified Xylanase A G3 expressed in P. pastoris (B) M2, molecular mass markers;CotAPp + Endo H, purified recombinant CotA expressed in P. pastoris digested withEndo H; CotAPp, purified CotA expressed in P. pastoris. (C) Circular dichroism spectraof the purified XynAG3 (solid line) and CotA (dotted line) measured as described
agreement with the secondary structure content observed in the
he laccase and xylanase activities were monitored in cultures induced with 0.5%ethanol (solid symbols) or with 1% of methanol (open symbols).
Fig. 1). The highest level of laccase activity was obtained after sevenays of 0.5% methanol induction, whereas the highest xylanasectivity was observed after four days of 1% methanol induction. Theield of the coexpressed CotAPp and XynAG3Pp with 1% inducer con-entration was 10.5 mg/L and 11 mg/L, respectively, and althoughhe yields of the two enzymes in separate cultures were 15 mg/LCotAPp) and 13 mg/L (XynAG3Pp), a mixture of the two separatenzyme solutions would result in their dilution. Thus, 2 L of a mix-ure of the two enzymes separately expressed would contain 15 mgnd 13 mg of CotAPp and XynAG3Pp, respectively, whereas 2 L ofoexpressed culture would contain 21 mg and 22 mg of the samewo enzymes.
The enzymes were expressed separately and purified byi-affinity chromatography. Analysis by SDS-PAGE (Fig. 2) demon-
trated a single protein band for CotAPp whereas the XynAG3Ppresented three bands. Treatment of glycoproteins with endo-lycosidase H (Endo H) specifically cleaves the glycosidic linkageetween the two core GlcNAc residues of the N-linked sugar chain,nd Endo H treatment of XynAG3Pp yielded a single band of reducedize (see Fig. 2A). We have previously reported that the wild-typeylanase A expressed in P. pastoris has six potential N-glycosylationites, however MS analysis has revealed that only between two andour of these N-glycosylation sites (Asn20, Asn25, Asn141, Asn181)re modified [19]. The wild-type and XynAG3Pp enzymes maintainhe same potential N-glycosylation sites and banding pattern inDS-PAGE, suggesting that the same positions are glycosylated inoth enzymes.
The CotA has two potential N-glycosylation sites (Asn255 and
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
Ppsn263), and the single band in SDS-PAGE indicated more homo-eneity in the glycoform (see Fig. 2B). After treatment with Endo H,he LC–MS/MS for endoH/tryptic peptides of CotAPp revealed the
Pp Pp
in Section 2. The additional 70 kDa protein band seen in panels A and B is the EndoHused in the deglycosylation reaction.
presence of a glycosylated Asn263 residue (supplementary Fig. S1).No mass modification of the tryptic peptides containing Asn255was observed (data not shown), indicating the absence of glycosy-lation at this putative site.
3.2. Biophysical analyses
To determine if the recombinant proteins show significant dif-ferences in their protein secondary structures, circular dichroismspectra were measured for both enzymes (Fig. 2C). The spectrumof the XynAG3Pp showed defined minima at 218 and 227 nm anda positive peak at 200 nm which are typical features of the CDspectrum of the native enzyme structure [16]. The spectrum of theCotAPp showed a minimum at 215 nm and a maximum at 200 nm,and is essentially identical with the spectra of the native enzymeobserved in previous studies [20]. Both spectra showed character-istics of proteins rich in �-sheet secondary structures, which is in
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
crystal structures of the enzymes. These results indicated that gly-cosylation did not alter the native-like secondary structure of eitherenzyme.

ARTICLE IN PRESSG ModelPRBI-10109; No. of Pages 10
R. Fonseca-Maldonado et al. / Process Biochemistry xxx (2014) xxx–xxx 5
Fig. 3. (A) UV–vis absorbance spectra of the purified CotAPp. See Section 2 for further experimental details. (B) Ribbon representation of a modeled three-dimensionals difiedm ack spf
wpcospTcaosAiTiTt
3a
pDcawfci**pfsawaatl
tructure of CotA from Bacillus subtilis with the N-glycosylation site at Asn263 mooiety is depicted as dark gray spheres, and the copper atoms are indicated by bl
rom the PDB file (Bento et al. [23]).
The environments of the T1 and T3 coppers of the CotAPp enzymeere observed by UV–vis absorbance spectroscopy (Fig. 3A). Theurified CotAPp exhibited the blue color that is characteristic ofopper laccases, and the absorption spectrum was similar to thatbserved of other glycosylated laccases [21,22]. The spectrumhowed an absorbance peak at 600 nm that is typical of a Type I cop-er binding, and which confers the intense blue color to the enzyme.he T3, or coupled binuclear copper site, consists of a pair of tightlyoupled copper ions and gives rise to the broad shoulder observedt 330 nm in the absorbance spectrum. A three-dimensional homol-gy model of the glycoprotein was constructed based on the B.ubtilis CotA crystal structure [23], and in this model structure thesn263 residue has been modified with an oligosaccharide consist-
ng of two GlcNAc and nine mannose saccharide residues (Fig. 3B).he model structure reveals that the glycosylation site in the CotAs located close to the surface cleft in which is located the external1 Cu2+, which is the site at where electron-donor molecules bindo the laccase.
.3. Effect of temperature and pH on xylanase and laccasectivity and thermal stability
The XynAG3Pp and CotAPp activities were measured at differentH values and temperatures, in which a Central Compositeesign with two variables was used to optimize reactiononditions. The models (Fig. 4A and B) were validated bynalysis of variance (ANOVA), which showed that the modelas significant at high confidence level (95%), with R2 = 0.97
or XynAG3Pp and R2 = 0.99 for CotAPp activities. The regressionoefficients (Table S2) of the polynomial expressions (CotAPp Activ-ty = −76.3888 + 16.2624 * pH − 1.8854 * pH2 + 1.3552 * T − 0.0099
T2 + 0.0232 * pH * T and XynA Activity = −3114.67 + 663.28 pH − 61.97 * pH2 + 54.03 * T − 0.49 * T2 + 0.82 * pH * T) showed low-values not only for all linear and quadratic functions, but alsoor the interaction between pH and temperature indicating theirignificant effect on enzymatic activities. The maximum xylanasectivity was at pH 5.8 and 58 ◦C and the maximum laccase activityas at pH 4.8 and 75 ◦C. To validate the models, the XynAG3Pp
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
ctivity was measured at pH 5 and a temperature of 60 ◦C, wheren activity of 362 ± 11 U mg−1 total protein was predicted, andhe experimentally determined activity was 355 ± 15 U mg−1. Foraccase activity under same conditions, the predicted activity was
with two N-acetylglucosamine and nine mannose residues. The oligosaccharideheres. The atomic coordinates of the native CotA from Bacillus subtilis were taken
10.5 ± 1.5 U mg−1, and the experimentally determined value was11.1 ± 1.1 U mg−1.
The thermostability of both enzymes was evaluated at two dif-ferent temperatures (Fig. 5). The XynAG3Pp showed half-lives of150 and 25 min at 60 ◦C and 70 ◦C, respectively (Fig. 5A). Ther-mal stability experiments with the CotAPp at both 65 ◦C and 80 ◦Cshowed an approximately 10-fold increase in activity during theinitial 100–150 min, and thereafter the CotAPp incubated at 80 ◦Cpresented 50% of its highest activity after 200 min, and at 65 ◦Cthe enzyme retained 50% of activity after 540 min (Fig. 5B). TheCotAPp showed a higher thermal activation than the same enzymeexpressed in Escherichia coli [4].
3.4. Kinetic parameters
The values calculated for the apparent dissociation constants(K0.5), turnover numbers (kcat), and catalytic efficiencies are pre-sented in Table 1. The K0.5 value for the hydrolysis of oat spelt xylanby XynAG3Pp was 1.3-fold higher than the previously reportedvalue of the unglycosylated enzyme expressed in E. coli [4], andconsequently the catalytic efficiency (kcat/K0.5) decreased by thesame factor. Glycosylation has no significant effect on kcat of theXynAG3Pp. In contrast, the maximum activity (Vmax) for the oxida-tion of ABTS by CotAPp showed an approximate 2-fold enhancementas compared to the unglycosylated enzyme, and consequently thekcat and the catalytic efficiency (kcat/K0.5) increased by the samefactor. The K0.5 value for the oxidation of ABTS by CotAPp was sim-ilar to the previously reported value of the unglycosylated enzyme[4].
3.5. Hydrolysis of milled sugarcane bagasse by mixtures ofXynAG3 and CotA
To evaluate the activity of the culture extracts against a nat-ural substrate, the co-expression system was used to producedifferent ratios of the XynAG3Pp and CotAPp enzymes. Enzymeproduction by the P. pastoris GS115/pPICZ�A xynAG3/pPI9k cotA(P. pastoris GS115 transformed with the coding sequences of both
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
enzymes), was induced with either 0.5% or 1% methanol, and afterfive days of induction the culture supernatant was concentrated20-fold and the enzyme levels of both extracts were measured.In the culture induced with 0.5% methanol, 3.1 nmol mL−1 of

ARTICLE IN PRESSG ModelPRBI-10109; No. of Pages 10
6 R. Fonseca-Maldonado et al. / Process Biochemistry xxx (2014) xxx–xxx
F e pana
Ctatttc
Fcf
ig. 4. Response surfaces (left hand side panels) and contour plots (right hand sidctivities. See Section 2 for further experimental details.
otAPp and 4.7 nmol mL−1 of XynAG3Pp were detected, and inhe culture induced with 1% methanol 4.6 nmol mL−1 of CotAPpnd 9.5 nmol mL−1 of XynAG3Pp were detected. The P. pastoris
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
ransformed with the coding sequences of either CotA or XynAG3ogether with P. pastoris transformed with the empty transforma-ion vector were subjected to methanol induction under the sameonditions. The amount of enzyme used in each experiment and
ig. 5. Thermostability of XynAG3Pp and CotAPp. (A) XynAG3Pp was incubated at 60 ◦C (ircles), and 80 ◦C (closed squares). Relative activities to the maximum values are shown,urther experimental details.
els) showing the effects of temperature and pH on CotAPp (A) and XynAG3Pp (B)
the reducing sugar released are shown in Table 2. The highestreducing sugar release was obtained with the extract induced with1% of methanol in the presence of ABTS (0.71 �mol of reducing
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
sugar) which contained a 2:1 ratio of XynAG3Pp/CotAPp. This is 22%higher as compared to the reducing sugar released with the extractinduced by 0.5% of methanol (0.58 �mol of reducing sugar), whichproduced a 1.5:1 ratio of XynAG3Pp/CotAPp. The extracts of the P.
open circles), and 70 ◦C (closed squares). (B). CotAPp was incubated at 65 ◦C (open and the values correspond to the averages of five measurements. See Section 2 for

ARTICLE IN PRESSG ModelPRBI-10109; No. of Pages 10
R. Fonseca-Maldonado et al. / Process Biochemistry xxx (2014) xxx–xxx 7
Table 1Kinetic parameters for CotAPp and XynAG3Pp.
Enzyme Substrate
ABTS Sylan
K0.5 (�M) kcat (s−1) kcat/K0.5 (mM−1 s−1) K0.5 (mg mL−1) kcat (s−1) kcat/K0.5 (mL mg−1 s−1)
CotAPp 27 8.6 319 nd nd ndXynAG3Pp nd nd nd 6.5 60 9.4
nd, not determined. Data represent the mean of two independent preparations.
Table 2Reducing sugar release from milled sugarcane bagasse treated with transformed P. pastoris culture supernatant concentrates. The table includes the amount of CotAPp andXynAG3Pp in nmol in each culture condition, together with the amount of reducing sugar released both in the presence and absence of ABTS. The results are presented as thetotal reducing sugar release in micromoles. Triplicate assays were performed in 100 mM sodium acetate buffer (pH 5.0) at 60 ◦C for 2 h (see Section 2 for further details).
0.5% of methanol induction 1% of methanol induction
Amount of enzyme(nmol)
Units of catalysis pergram of biomass(U g−1)
Reducing sugarreleased (�mol)
Amount of enzyme(nmol)
Units of catalysis pergram of biomass(U g−1)
Reducing sugarreleased (�mol)
Culture supernatantwith two enzymes
0.77 (CotAPp)1.18 (XynAG3Pp)
30 (CotAPp)500 (XynAG3Pp)
0.34 ± 0.016 0.77 (CotAPp)1.58 (XynAG3Pp)
30 (CotAPp)650 (XynAG3Pp)
0.38 ± 0.012
Culture supernatantwith twoenzymes + ABTS
0.77 (CotAPp)1.18 (XynAG3Pp)
30 (CotAPp)500 (XynAG3Pp)
0.58 ± 0.018 0.77 (CotAPp)1.58 (XynAG3Pp)
30 (CotAPp)650 (XynAG3Pp)
0.71 ± 0.015
Culture supernatantwith CotAPp
0.77 30 0.21 ± 0.011 0.77 30 0.21 ± ±0.014
Culture supernatantwith CotAPp + ABTS
0.77 30 0.3 ± 0.014 0.77 30 0.31 ± 0.014
Culture supernatantwith XynAG3Pp
1.18 500 0.1 ± 0.02 1.58 650 0.13 ± 0.017
Culture supernatantwith XynAG3Pp +ABTS
1.18 500 0.09 ± 0.013 1.58 650 0.13 ± 0.013
Culture Supernatantwithout enzymes
No enzymes No enzymes 0.0034 ± 0.0
Fw
pimcp1
otmtCo
ig. 6. Release of reducing sugar from milled sugarcane bagasse after treatmentith different ratios of CotA and XynAG3. See Section 2 for experimental details.
astoris double transformant had greater reducing sugar releas-ng activity than the sum of the action of the individual enzymes
easured separately, showing increases of 49% (i.e. 0.58 �mol asompared to 0.3 + 0.09 = 0.39 �mol) and 61% (i.e. 0.71 �mol as com-ared to 0.31 + 0.13 = 0.44 �mol) after induction with either 0.5% or% methanol, respectively, in the presence of ABTS.
With the aim of evaluating the degree of enzyme synergismbserved in the coexpression experiments, the total concentra-ion of reducing ends from soluble saccharides generated from
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
illed sugarcane bagasse was measured after treatment with mix-ures of CotAPp and XynAG3Pp in various ratios (see Fig. 6). EitherotAPp (CotAPp:XynAG3Pp of 1:0) or XynAG3Pp (CotAPp:XynAG3Ppf 0:1) alone showed reducing end releasing activity, and
16 No enzymes No enzymes 0.0031 ± 0.016
increasing the XynAG3Pp alone (CotAPp:XynAG3Pp of 0:2, 0:3 and0:5) had only a slight effect on reducing sugar release. The con-centration of soluble saccharide reducing ends was higher aftertreatment with an equimolar mixture of CotAPp and XynAG3Pp(CotAPp:XynAG3Pp of 1:1) than the sum of those obtained withCotAPp (CotAPp:XynAG3Pp of 1:0) or XynAG3Pp (CotAPp:XynAG3Ppof 0:1) alone, confirming the synergistic activity. However, increas-ing the proportion of the CotAPp (CotAPp:XynAG3Pp of 2:1, 3:1 and5:1) had no significant effect. In contrast, increasing the propor-tion of the XynAG3Pp in the mixtures (CotAPp:XynAG3Pp of 1:2, 1:3and 1:5) enhanced the synergistic effect of the enzymes. Increas-ing the CotAPp:XynAG3Pp ratio from 1:1 to 1:2 and from 1:2 to 1:3,increased the released reducing sugar by 18% and 39%, respectively.Above a CotAPp:XynAG3Pp ratio of 1:3, only a minor increase in totalreducing sugar release was observed, and at a CotAPp:XynAG3Ppratio of 1:5 the released reducing sugar was only 4% higher of thatat a ratio of 1:3.
4. Discussion
Synergism between exo-glucanases and endoxylanases is awell established effect and results in increased reducing sugarrelease [24], and crude extracts containing laccase–xylanase mix-tures derived from co-cultivation of two microbial sources hasbeen shown to improve biobleaching of wood pulps [25]. Thetwo enzymes were chosen on the basis of previously reportedresults which demonstrated synergy in the catalytic activitieswhen the two enzymes were fused to form a bifunctional chimera[4]. Furthermore, when expressed in E. coli the CotA presents an
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
optimum catalytic temperature of 75 ◦C that is similar to the tem-perature optimum of the XylanaseA G3, a thermostable variantof the xylanase A from B. subtilis created by directed evolution[16]. In the present study we have demonstrated the simultaneous

ING ModelP
8 cess B
hyctfaCc(r
xwoe7tvrCmghtftmEwtcsf[(epfePta
tcTsUtelbtcsrsa[nt
npti
ARTICLERBI-10109; No. of Pages 10
R. Fonseca-Maldonado et al. / Pro
eterologous expression of a laccase and an endoxylanase in theeast P. pastoris by the integrative transformation into the hostell genome of the two enzymes using different expression vec-ors and two rounds of different selections. Enzyme cost is a keyactor in economic viability of the application of enzyme mixtures,nd here we have shown that coexpression of the XynAG3Pp andotAPp show a significant gain in total protein yield (22.5 mg L−1) inomparison to mixtures of the same enzymes expressed separately15 mg L−1 and 13 mg L−1 for the separate CotAPp and XynAG3Pp,espectively).
Previous studies have demonstrated expression levels of variousylanase in Pichia pastoris varies over the range 3.7–324 U mL−1,ith the recombinant enzymes commonly presenting similar
r improved catalytic properties when compared to the nativenzymes [26]. In contrast, the XynAG3 expressed in E. coli retains0% activity after 30 min at 80 ◦C [16], which demonstrates thathe XynAG3Pp is less thermostable. This is consistent with pre-ious results that demonstrated a reduced thermostability of theecombinant XynA expressed in P. pastoris [19]. The XynAG3Pp andotAPp proteins produced in P. pastoris undergo post-translationalodification in the eukaryotic host cell, and both enzymes were
lycosylated although to different degrees. The XynAG3Pp showedeterogeneity in glycosylation site occupancy (Fig. 2A), whereashe CotAPp underwent modification at a single site. Glycoproteinsrom P. pastoris display N-linked glycosylation of the high mannoseype, and this pattern was confirmed by the shift in electrophoretic
igration and mass spectroscopic analysis of the CotAPp afterndoH digestion (Fig. 2B). The secondary structure of the enzymesas not affected by glycosylation (Fig. 2C), however a slightly per-
urbation in tertiary structure of CotAPp is suggested from thehanges in the copper centers as observed by UV–vis absorbancepectroscopy. The cloning and expression in P. pastoris GS115 of lac2rom Bacillus subtilis strain CICC20613 has recently been reported27]. The lac2 gene has 86% homology with a published cotA geneGeneID 936023), and the same predicted glycosylation sites, how-ver experimental analysis of the glycosylation of the recombinantrotein was not reported. The laccase gene from Bacillus licheni-
ormis, which shares 66% identity with CotA has been cloned andxpressed in P. pastoris [28]. Although both laccases expressed in. pastoris also show thermal activation, the optimum activity ofhe B. licheniformis laccase was at 70 ◦C, and the enzyme lost 65%ctivity after 2 h at 80 ◦C.
The UV–vis spectroscopy for CotAPp (Fig. 3A) showed an absorp-ion profile at 600 nm and at 330 nm and 720 nm that differs inomparison to the unglycosylated protein produced in E. coli [4].he type 3 Cu(I) may be present in either the reduced or oxidizedtate, however the prominent 330 nm absorption peak in the nearV, as well as the presence of the 720 nm peak are consistent with
he type 3 binuclear Cu(II) cluster in the oxidized state [29,30]. Thenvironment of the copper centers in the CotAPp can be altered byocal structural perturbations resulting from protein modificationy N-linked glycans (Fig. 3B), and it has been previously shownhat alteration of the environment of the type 3 copper in the lac-ase modulates the spectroscopic properties of the blue T1 copperite [31]. Previous studies of fungal laccases have suggested variousoles for carbohydrate moieties, such as stabilization of the proteintructure thereby favoring the correct incorporation of the coppertoms, directing protein secretion, and enhancing thermostability32]. Since the CotA is of bacterial origin and this enzyme is notaturally produced in the glycosylated form, it is not unexpectedhat glycosylation results in changes in the protein structure.
To create the double transformant, the homologous recombi-
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
ation insertion sites at either the AOX1 or the His4 loci in P.astoris GS115 in order to generate only Mut+ recombinants, andherefore different patterns of protein expression with alterationsn inducer concentration were unexpected. However we observed
PRESSiochemistry xxx (2014) xxx–xxx
that both the concentration of methanol and the time of induc-tion exerted distinct effects on the levels of CotAPp and XynAG3Ppactivity (Fig. 4). One possible explanation for the decreased CotAPpactivity with induction by 1% methanol may be susceptibility of theenzyme to proteases. Previous investigations have indicated thatproteolytic activity plays an important role in the determination ofthe levels of heterologous expression in P. pastoris [33,34], and athigh cell densities proteases can either be secreted or be releasedfrom dead cells [33]. In the case of XynAG3Pp the higher protein pro-duction on induction with 1% methanol may be due to the reducedsusceptibility of the enzyme to protease digestion, which in turnmay be the consequence the high glycan content.
The reduced catalytic efficiency parameter of the purifiedXynAG3Pp may be explained by the high degree of glycosylationthat limits access of the polysaccharide substrate to the enzymecatalytic site. The different effect of glycosylation on the CotAPpis perhaps due to the lower glycan content of the enzyme, andthat glycosylation in laccases is believed to play a role in copperretention [32,35]. Indeed, it is believed that the T2 copper is labilein many laccases [31]. The CotAPp is glycosylated at the Asn263residue that is located on a surface loop close to the T2 copper site,and the model of the glycoprotein generated in the present studysuggests that this modification may influence either affinity of theT2 copper, or binding of the ABTS modulator to the enzyme.
The Pareto chart (Fig. S2) derived from the biochemical analysesusing the surface response methodology shows that the interac-tions between pH and temperature have a significant effect onCotAPp and XynAG3Pp activities. Second order polynomial fittingmodels were used for both enzymes, in which the linear andquadratic terms of the two factors are important in the model,and in both cases the absence of a fit was not significant. Forboth the CotAPp and the XynAG3Pp the higher regression coeffi-cient for pH suggested that the catalytic activity is more sensitiveto changes in pH than temperature. The response surface method-ology is useful to rapidly identify the optimal conditions of pH andtemperature for the use of an enzyme mixture, which together withthe improved stability of the XynAG3Pp at 60 ◦C and compatibilityof both enzymes at pH 5.0 led us to choose these conditions for theexperiments using milled sugar cane bagasse as a natural substrate.
Milled sugarcane bagasse contains about 6% lignin and 55–60%hemicellulose, including xylans [36], and treatment with CotAPpled to the release of reducing sugar even though this enzyme lackshydrolytic activity. In the plant cell wall, the lignin and hemicellu-lose components are covalently linked, and as previously reportedthe treatment of a natural lignocellulosic substrate with CotAresults in depolymerization of lignin with a concomitant releaseof reducing sugars [20]. These findings are corroborated in the cur-rent study by the observation that the reducing sugar release dueto the CotAPp activity is higher in presence of ABTS, and we suggestthat this small molecule can penetrate the lignocellulose matrix toattack the lignin. The activity of the XynAG3Pp against the milledsugar cane bagasse is lower when compared to the soluble xylansubstrate, and this may be due to either differences in the com-position of the polysaccharides between the oat spelt and sugarcane xylans, or due to an altered conformation of the polysaccha-ride in the cell wall matrix or the presence of lignin which providesa protective barrier against enzyme attack. Thus the basis of thesynergistic effect of the enzyme mixtures may be explained bythe action of CotAPp against the lignin that exposes the xylan fromwithin the lignocellulose matrix, and the xylan is then hydrolyzedby the XynAG3Pp into reducing sugars. Alternatively, the mechan-ical pretreatment of the bagasse may lead to breakage of the xylan
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
polymers, and subsequent treatment with the laccase might releasexylo-oligomers from the lignocellulose matrix.
We observed that both induction by different concentrationsof methanol and the length of induction time during protein

ING ModelP
cess B
eaiphsccCiptmCtcbv
bbthbcebc
C
A
amtmpocpRmnma
A
GsF55iab
A
t
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLERBI-10109; No. of Pages 10
R. Fonseca-Maldonado et al. / Pro
xpression resulted in changes in the ratio of the secreted enzymes,nd at a CotAPp:XynAG3Pp ratio of 1:2 of (in the culture supernatantnduced with 1% methanol), more reducing sugar was releaseder total protein than at the CotAPp:XynAG3Pp ratio of 1:1.5. Theigher levels of reducing sugar at higher relative xylanase ratiosuggest that the xylan hydrolysis is the limiting factor in hemi-ellulose degradation in the milled sugar cane bagasse. This wasonfirmed by the observation that increasing the proportion ofotAPp had no influence on the synergistic effect, whereas increas-
ng the proportion of the XynAG3Pp in the mixture resulted inrogressively higher levels of reducing sugar release. Althoughhe CotAPp:XynAG3Pp ratio of 1:2 showed a significant improve-
ent in the enzyme synergy, the elevated synergy at higherotAPp:XynAG3Pp ratios showed that improvements in the sys-em are possible. The XynAG3Pp levels in the coexpression mixtureould be further increased by further variation in inducer levels, ory engineering the response of the AOX promoter in the expressionector.
In conclusion, our results have demonstrated synergismetween an endoxylanase and a laccase using milled sugar caneagasse as a lignocellulose substrate. Furthermore, we have shownhat coexpression of enzymes from a bacterial source in a eukaryoteost cell is a viable option for production of enzyme mixtures foriomass degradation, and that the composition of these mixturesan be influenced by the induction conditions. This study not onlyxpands the range of enzymes for use in lignocellulose degradation,ut also demonstrates the potential for the use of enzyme mixturesontaining laccases in biotechnological applications.
ompeting interests
The authors declare that they have no competing interests.
uthors’ contribution
RFM designed the study, performed the molecular cloningnd heterologous expression of the bacterial enzymes in Pichia,easurement xylanase biochemical characterization, carried out
he statistic experiments, analyzed the results and wrote theanuscript. EB conducted mass spectrometry analyses. LFR partici-
ated in determining the kinetic parameters of laccase and analysisf the results. CBM and GFP participated in measurement of lac-ase thermostability, and GFP analyzed the results. JSA and GFParticipated in circular dichroism spectra measurement. PLM ePMF participated in the planning and execution of the activityeasurements of the enzyme mixtures. LMA participated in plan-
ing and execution of protein purification, PT and RJW revised theanuscript and coordinated the study. All authors have read and
pproved the final manuscript.
cknowledgments
We thank George Jackson de Moraes Rocha, Edgardo Olivaresomez and Roberto Ruller (CTBE, Campinas, Brazil) for providingamples of milled sugar cane bagasse. This work was supported byAPESP grants 2007/01623-0, 2008/05181-5 (CBM), 2009/50838-
(DSV), 2010/07133-8 (LFR) and 2010/18850-2, CNPq grants04807/2009-9 and 307795/2009-8, and CAPES. IRIC is supported
n part by the Canadian Center of Excellence in Commercializationnd Research (CECR), the Canadian Foundation for Innovation andy the Fonds de Recherche du Québec en Santé (FRQS).
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.procbio.2014.03.027.
[
PRESSiochemistry xxx (2014) xxx–xxx 9
References
[1] Hasunuma T, Kondo A. Development of yeast cell factories for consolidatedbioprocessing of lignocellulose to bioethanol through cell surface engineering.Biotechnol Adv 2010;30(6):1207–18.
[2] Leonowicz A, Cho NS, Luterek J, Wilkolazka A, Wojtas-Wasilewska M,Matuszewska A, et al. Fungal laccase: properties and activity on lignin. J BasicMicrobiol 2001;41:185–227.
[3] Coughlan MP, Hazlewood GP. Beta-1,4-d-xylan-degrading enzyme systems:biochemistry, molecular biology and applications. Biotechnol Appl Biochem1993;17(Pt 3):259–89.
[4] Ribeiro LF, Furtado GP, Lourenzoni MR, Costa-Filho AJ, Santos CR, NogueiraSC, et al. Engineering bifunctional laccase–xylanase chimeras for improvedcatalytic performance. J Biol Chem 2011;286:43026–38.
[5] Sun Y, Cheng J. Hydrolysis of lignocellulosic materials for ethanol production:a review. Bioresour Technol 2002;83:1–11.
[6] Messerschmidt A, Huber R. The blue oxidases, ascorbate oxidase, laccaseand ceruloplasmin: modelling and structural relationships. Eur J Biochem1990;187:341–52.
[7] Kersten PJ, Kalyanaraman B, Hammel KE, Reinhammar B, Kirk TK. Comparisonof lignin peroxidase, horseradish peroxidase and laccase in the oxidation ofmethoxybenzenes. Biochem J 1990;268:475–80.
[8] Martins LO, Soares CM, Pereira MM, Teixeira M, Costa T, Jones GH, et al. Molec-ular and biochemical characterization of a highly stable bacterial laccase thatoccurs as a structural component of the Bacillus subtilis endospore coat. J BiolChem 2002;277:18849–59.
[9] Biely P. Microbial xylanolytic systems. Trends Biotechnol 1985;3:286–90.10] Kulkarni N, Shendye A, Rao M. Molecular and biotechnological aspects of
xylanases. FEMS Microbiol Rev 1999;23:411–56.11] Cantarel BL, Coutinho PM, Rancurel C, Bernard T, Lombard V, Henrissat B.
The Carbohydrate-Active EnZymes database (CAZy): an expert resource forglycogenomics. Nucleic Acids Res 2009;37:D233–8.
12] Murakami MT, Arni RK, Vieira DS, Degreve L, Ruller R, Ward RJ. Correlation oftemperature induced conformation change with optimum catalytic activity inthe recombinant G/11 xylanase A from Bacillus subtilis strain 168 (1A1). FEBSLett 2005;579:6505–10.
13] Ruller R, Rosa JC, Faca VM, Greene LJ, Ward RJ. Efficient constitutive expressionof Bacillus subtilis xylanase A in Escherichia coli DH5alpha under the control ofthe Bacillus BsXA promoter. Biotechnol Appl Biochem 2006;43:9–15.
14] Daly R, Hearn MT. Expression of heterologous proteins in Pichia pastoris: a use-ful experimental tool in protein engineering and production. J Mol Recognit2005;18:119–38.
15] Cereghino JL, Cregg JM. Heterologous protein expression in the methylotrophicyeast Pichia pastoris. FEMS Microbiol Rev 2000;24:45–66.
16] Ruller R, Deliberto L, Ferreira TL, Ward RJ. Thermostable variants of the recom-binant xylanase A from Bacillus subtilis produced by directed evolution showreduced heat capacity changes. Proteins 2008;70:1280–93.
17] Leone FA, Baranauskas JA, Furriel RP, Borin IA, Sigraf W. An easy-to-use programfor fitting enzyme kinetic data. Biochem Mol Biol Educ 2005;33:399–403.
18] Miller GL. Use of dinitrosalicylic acid reagent for determination of reducingsugar. Anal Biochem 1959;31:426–8.
19] Fonseca-Maldonado R, Vieira DS, Alponti JS, Bonneil E, Thibault P, Ward RJ.Engineering the pattern of protein glycosylation modulates the thermostabilityof a GH11 xylanase. J Biol Chem 2013;288:25522–34.
20] Furtado GP, Ribeiro LF, Lourenzoni MR, Ward RJ. A designed bifunctionallaccase/beta-1,3-1,4-glucanase enzyme shows synergistic sugar release frommilled sugarcane bagasse. Protein Eng Des Sel 2012;26:15–23.
21] Coll PM, Fernandez-Abalos JM, Villanueva JR, Santamaria R, Perez P. Purificationand characterization of a phenoloxidase (laccase) from the lignin-degradingbasidiomycete PM1 (CECT 2971). Appl Environ Microbiol 1993;59:2607–13.
22] Eggert C, Temp U, Eriksson KE. The ligninolytic system of the white rot fungusPycnoporus cinnabarinus: purification and characterization of the laccase. ApplEnviron Microbiol 1996;62:1151–8.
23] Bento I, Martins LO, Gato Lopes G, Armenia Carrondo M, Lindley PF. Dioxy-gen reduction by multi-copper oxidases: a structural perspective. Dalton Trans2005:3507–13.
24] Gao D, Uppugundla N, Chundawat SP, Yu X, Hermanson S, Gowda K, et al. Hemi-cellulases and auxiliary enzymes for improved conversion of lignocellulosicbiomass to monosaccharides. Biotechnol Biofuels 2011;4:5.
25] Dwivedi P, Vivekanand V, Pareek N, Sharma A, Singh RP. Bleach enhancement ofmixed wood pulp by xylanase–laccase concoction derived through co-culturestrategy. Appl Biochem Biotechnol 2009;160:255–68.
26] Ahmed S, Riaz S, Jamil A. Molecular cloning of fungal xylanases: an overview.Appl Microbiol Biotechnol 2009;84:19–35.
27] Yang SS, Liu ZW, Yi XP, Zhang AL, Zhang TY, Luo JX, et al. Isolation of laccasegene from Bacillus subtilis and analysis of its physicochemical properties. Gene2012;491:49–52.
28] Lu L, Wang TN, Xu TF, Wang JY, Wang CL, Zhao M. Cloning and expression ofthermo-alkali-stable laccase of Bacillus licheniformis in Pichia pastoris and itscharacterization. Bioresour Technol 2013;2013:81–6.
29] Blackburn NJ, Ralle M, Hassett R, Kosman DJ. Spectroscopic analysis of the trin-
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
uclear cluster in the Fet3 protein from yeast, a multinuclear copper oxidase.Biochemistry 2000;39:2316–24.
30] Pita M, Shleev S, Ruzgas T, Fernández VM, Yaropolov AI, Gorton L. Directheterogeneous electron transfer reactions of fungal laccases at bare and thiol-modified gold electrodes. Electrochem Commun 2006;8:747–53.

ING ModelP
1 cess B
[
[
[
[
[
ARTICLERBI-10109; No. of Pages 10
0 R. Fonseca-Maldonado et al. / Pro
31] Bukh C, Bjerrum MJ. The reversible depletion and reconstitution of a copperion in Coprinus cinereus laccase followed by spectroscopic techniques. J InorgBiochem 2000;104:1029–37.
32] Rodgers CJ, Blanford CF, Giddens SR, Skamnioti P, Armstrong FA, Gurr SJ.
Please cite this article in press as: Fonseca-Maldonado R, et al. Synemilled sugar cane bagasse. Process Biochem (2014), http://dx.doi.org/
Designer laccases: a vogue for high-potential fungal enzymes. Trends Biotech-nol 2009;28:63–72.
33] Macauley-Patrick S, Fazenda ML, McNeil B, Harvey LM. Heterologous pro-tein production using the Pichia pastoris expression system. Yeast 2005;22:249–70.
[
PRESSiochemistry xxx (2014) xxx–xxx
34] Hong F, Meinander NQ, Jonsson LJ. Fermentation strategies for improvedheterologous expression of laccase in Pichia pastoris. Biotechnol Bioeng2002;79:438–49.
35] Li K, Xu F, Eriksson KE. Comparison of fungal laccases and redox mediators in
rgistic action of co-expressed xylanase/laccase mixtures against10.1016/j.procbio.2014.03.027
oxidation of a nonphenolic lignin model compound. Appl Environ Microbiol1999;65:2654–60.
36] de Souza AP, Leite DCC, Pattathil S, Hahn MG, Buckeridge MS. Compositionand structure of sugarcane cell wall polysaccharides: implications for second-generation bioethanol production. Bioenergy Res 2013;6:564–79.
Related Documents











