Sustainability and cyanobacteria (blue-green algae): facts and challenges Naveen K. Sharma & Sri Prakash Tiwari & Keshwanand Tripathi & Ashwani K. Rai Received: 4 November 2009 / Revised and accepted: 22 September 2010 / Published online: 18 November 2010 # Springer Science+Business Media B.V. 2010 Abstract Cyanobacteria (blue-green algae) are widely distributed Gram-negative oxygenic photosynthetic prokar- yotes with a long evolutionary history. They have potential applications such as nutrition (food supplements and fine chemicals), in agriculture (as biofertilizer and in reclama- tion of saline USAR soils) and in wastewater treatment (production of exopolysaccharides and flocculants). In addition, they also produce wide variety of chemicals not needed for their normal growth (secondary metabolites) which show powerful biological activities such as strong antiviral, antibacterial, antifungal, antimalarial, antitumoral and anti-inflammatory activities useful for therapeutic purposes. In recent years, cyanobacteria have gained interest for producing biofuels (both biomass and H 2 production). Because of their simple growth needs, it is potentially cost-effective to exploit cyanobacteria for the production of recombinant compounds of medicinal and commercial value. Recent advances in culture, screening and genetic engineering techniques have opened new ways to exploit the potential of cyanobacteria. This review analyses the sustainability of cyanobacteria to solve global problems such as food, energy and environmental degrada- tion. It emphasizes the need to adopt multidisciplinary approaches and a multi-product production (biorefinery) strategy to harness the maximum benefit of cyanobacteria. Keywords Agriculture . Aquaculture . Bioactive compounds . Cyanobacteria . Energy . Human health . Nutraceuticals . Secondary metabolites . Toxins Introduction Issues related to environment, food and energy have presented serious challenge to the stability of nation-states (Tilman et al. 2009). Increasing global population, dwin- dling agriculture and industrial production, and inequitable distribution of resources and technologies have further aggravated the problem. They are entwined in such a way that it becomes almost impossible to grade their severity. The burden placed by increasing population on environ- ment and especially on agricultural productivity is phe- nomenal. To feed such a massive population, it becomes imperative to find new ways and means to increase the production giving due consideration to biosphere’ s ability to regenerate resources and provide ecological services. Cyanobacteria (blue-green algae) are Gram-negative oxygenic photosynthetic autotrophs, and are amongst the most successful and oldest life forms present (Schopf 2000; Gademan and Portman 2008). Globally, they are important primary producers and play significant roles in biogeo- chemical cycles of nitrogen, carbon and oxygen (30% of the annual oxygen production on earth) (Karl et al. 2002; DeRuyter and Fromme 2008). They are the organisms responsible for bringing oxygen on the earth therefore played a key role in the evolution of life. Some cyanobac- teria perform unique biological process (combine N 2 - fixation with oxygenic photosynthesis) and can be a model N. K. Sharma Department of Botany, Postgraduate College, Ghazipur, Uttar Pradesh 233001, India S. P. Tiwari Department of Applied Microbiology, VBS Poorvanchal University, Jaunpur, Uttar Pradesh 221001, India K. Tripathi : A. K. Rai (*) Department of Botany, Banaras Hindu University, Varanasi, Uttar Pradesh 221005, India e-mail: [email protected] J Appl Phycol (2011) 23:1059–1081 DOI 10.1007/s10811-010-9626-3

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Sustainability and cyanobacteria (blue-green algae): factsand challenges
Naveen K. Sharma & Sri Prakash Tiwari &Keshwanand Tripathi & Ashwani K. Rai
Received: 4 November 2009 /Revised and accepted: 22 September 2010 /Published online: 18 November 2010# Springer Science+Business Media B.V. 2010
Abstract Cyanobacteria (blue-green algae) are widelydistributed Gram-negative oxygenic photosynthetic prokar-yotes with a long evolutionary history. They have potentialapplications such as nutrition (food supplements and finechemicals), in agriculture (as biofertilizer and in reclama-tion of saline USAR soils) and in wastewater treatment(production of exopolysaccharides and flocculants). Inaddition, they also produce wide variety of chemicals notneeded for their normal growth (secondary metabolites)which show powerful biological activities such as strongantiviral, antibacterial, antifungal, antimalarial, antitumoraland anti-inflammatory activities useful for therapeuticpurposes. In recent years, cyanobacteria have gainedinterest for producing biofuels (both biomass and H2
production). Because of their simple growth needs, it ispotentially cost-effective to exploit cyanobacteria for theproduction of recombinant compounds of medicinal andcommercial value. Recent advances in culture, screeningand genetic engineering techniques have opened new waysto exploit the potential of cyanobacteria. This reviewanalyses the sustainability of cyanobacteria to solve globalproblems such as food, energy and environmental degrada-tion. It emphasizes the need to adopt multidisciplinary
approaches and a multi-product production (biorefinery)strategy to harness the maximum benefit of cyanobacteria.
Keywords Agriculture . Aquaculture . Bioactivecompounds . Cyanobacteria . Energy . Human health .
Nutraceuticals . Secondary metabolites . Toxins
Introduction
Issues related to environment, food and energy havepresented serious challenge to the stability of nation-states(Tilman et al. 2009). Increasing global population, dwin-dling agriculture and industrial production, and inequitabledistribution of resources and technologies have furtheraggravated the problem. They are entwined in such a waythat it becomes almost impossible to grade their severity.The burden placed by increasing population on environ-ment and especially on agricultural productivity is phe-nomenal. To feed such a massive population, it becomesimperative to find new ways and means to increase theproduction giving due consideration to biosphere’s abilityto regenerate resources and provide ecological services.
Cyanobacteria (blue-green algae) are Gram-negativeoxygenic photosynthetic autotrophs, and are amongst themost successful and oldest life forms present (Schopf 2000;Gademan and Portman 2008). Globally, they are importantprimary producers and play significant roles in biogeo-chemical cycles of nitrogen, carbon and oxygen (30% ofthe annual oxygen production on earth) (Karl et al. 2002;DeRuyter and Fromme 2008). They are the organismsresponsible for bringing oxygen on the earth thereforeplayed a key role in the evolution of life. Some cyanobac-teria perform unique biological process (combine N2-fixation with oxygenic photosynthesis) and can be a model
N. K. SharmaDepartment of Botany, Postgraduate College,Ghazipur, Uttar Pradesh 233001, India
S. P. TiwariDepartment of Applied Microbiology,VBS Poorvanchal University,Jaunpur, Uttar Pradesh 221001, India
K. Tripathi :A. K. Rai (*)Department of Botany, Banaras Hindu University,Varanasi, Uttar Pradesh 221005, Indiae-mail: [email protected]
J Appl Phycol (2011) 23:1059–1081DOI 10.1007/s10811-010-9626-3

to study important biological activities or capabilities(Herrero and Flores 2008).
In the early atmosphere the bulk of oxygen came fromoxygenic photosynthesis by cyanobacteria (Schopf 2000).Geological records indicate that this transforming eventoccurred at least some 2.4–2.3 Ga ago or even earlier(Knoll 2008). However, the question of when oxygenicphotosynthesis evolved still remains a contentious issue(Knoll 2008). The stromatolites of fossilised oxygenproducing cyanobacteria have been reported from 2.8 Gaago (Olson 2006) and Knoll (2008) considered cyanobac-teria as the principal primary producers throughout theProterozoic Eon (2.5–0.5 Ga).
The cyanobacteria consist of a heterogeneous assemblageof oxygen evolving photosynthetic prokaryotes, and arecomposed of about 150 genera and 2,000 species (Pulz andGross 2004) including unicellular, colonial, filamentous tobranched filamentous forms (Thajuddin and Subramanian2005). They are divided into five subsections (Rippka et al.1979; Castenholz 2001). Cyanobacteria are widely distribut-ed in habitats ranging from aquatic to terrestrial environ-ments as well as extreme habitats such as hot springs,hypersaline waters, deserts, and Polar Regions (Whitton andPotts 2000).
During their long evolutionary history, cyanobacteriahave undergone several structural and functional modifica-tions responsible for their versatile physiology and wideecological tolerance. Their abilities to tolerate high temper-ature, UV radiation, desiccation, water and saline stressescontribute to their competitive success in a wide range ofenvironments (Gröniger et al. 2000; Whitton and Potts2000; Herrero and Flores 2008). They can photosynthesiseat low photon densities and use bicarbonate ion forphotosynthesis at high pH (Shapiro 1972). Many speciesfix atmospheric N2 in usable (soluble) form. Also, they canuse diverse sulphur sources (Wolk 1973), and showefficient phosphate acquisition mechanisms (Rai andSharma 2006). Other factors include their resistance topredation pressure and ability to regulate cellular buoyancy.Ray and Bagachi (2001) reported that some cyanobacteriaproduce allelopathic compounds that may assist in theirdominance in the phytoplanktonic community.
Cyanobacteria have mostly been studied for their prokary-otic organisation, mechanism of photosynthesis, especially thestructure of photosynthetic complexes (PSI and II) (DeRuyterand Fromme 2008), N2-fixing ability of certain forms,genetic make-up and structural aspects such as gas vacuolesand akinetes (spores). Credit for origin of eukaryotic plant lifeover the planet also goes to cyanobacteria, as the chloroplastof eukaryotic cell is derived from a cyanobacterial ancestor(Delwiche and Palmer 1997; Tomitani et al. 2006).
The economic importance of cyanobacteria primarily liesin their agronomic importance as biofertilizers due to N2-
fixation that helps them to grow successfully in habitatswhere little or no combined N is available. In recent times,their ability to produce structurally novel and biologicallyactive natural products has been recognised (Ordog et al.2004; Gademan and Portman 2008; Sielaff et al. 2008;Wase and Wright 2008; Rosenberg et al. 2008; Tan 2010).They produce a wide variety of chemically uniquesecondary metabolites that include toxins, hormones, ironchelators, antibiotics, antifungal, antitumor, inflammatoryand anti-inflammatory compounds. Chemically, these com-pounds may be peptides, alkaloids and indole alkaloids,polyketide and terpenes (Gademan and Portman 2008).Many of these compounds display significant pharmaceu-tical potential.
Use of cyanobacteria as a food supplement has a longhistory (Gantar and Svircev 2008). Spirulina (now known asArthrospira) had been collected and used by Aztec popula-tion (Pulz and Gross 2004). Even today, malnutrition,especially due to a protein-poor diet is widespread in manyparts of the world. The use of cyanobacteria as a non-conventional source of food and protein seems promising(Pulz and Gross 2004; Gantar and Svircev 2008; Rosenberget al. 2008). Also, they may be used as a source of naturalchemicals to substitute synthetic cosmetics (Burja et al.2001; Singh et al. 2005) and conventional energy resources(Deng and Coleman 1999; Dutta et al. 2005). Cyanobacteriaare a highly diverse group, largely unexplored and untappedtherefore, present an opportunity for discovery of novelbiochemicals and their use by humankind (Skulberg 2000;Rosenberg et al. 2008). Certain cyanobacteria, known asextremophyles, inhabit extreme environments, e.g. Spirulina(alkalophilic), Mastigocladus laminosus (thermophilic),Aphanothece halophytica (halophilic) etc. Because of theirextreme requirements, mass cultures of extremophyles islikely to be free from microbial contamination thus, avoidinga serious problem in outdoor cultures (Pulz and Gross 2004).Extremophilic cyanobacteria represent a potential source ofbiotechnologically important molecules and enzymes.
Toxic secondary metabolites (Cyanotoxins)
Cyanobacteria occupy nearly all types of aquatic bodies(fresh, brackish and marine ecosystems), with possibleexception of acidic waters. Under favourable growthconditions, cyanobacteria often dominate aquatic bodiesresulting in cyanobacterial blooms. In the last 20 years,excessive nutrient inputs from agricultural fertiliser runoff, and/or domestic and industrial effluents, as well asclimatic change have greatly enhanced the incidence ofcyanobacterial blooms globally (Paerl and Huisman2009). Many of the bloom-forming cyanobacteria producetoxic secondary metabolites (cyanotoxins), mostly active
1060 J Appl Phycol (2011) 23:1059–1081
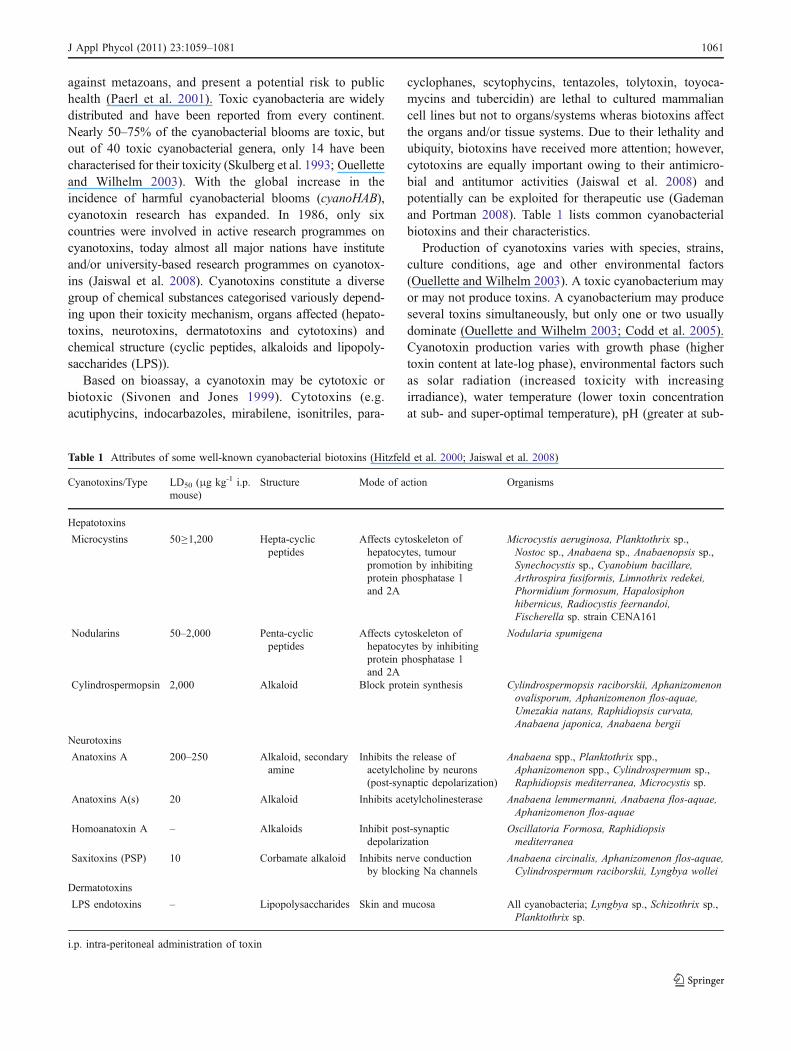
against metazoans, and present a potential risk to publichealth (Paerl et al. 2001). Toxic cyanobacteria are widelydistributed and have been reported from every continent.Nearly 50–75% of the cyanobacterial blooms are toxic, butout of 40 toxic cyanobacterial genera, only 14 have beencharacterised for their toxicity (Skulberg et al. 1993; Ouelletteand Wilhelm 2003). With the global increase in theincidence of harmful cyanobacterial blooms (cyanoHAB),cyanotoxin research has expanded. In 1986, only sixcountries were involved in active research programmes oncyanotoxins, today almost all major nations have instituteand/or university-based research programmes on cyanotox-ins (Jaiswal et al. 2008). Cyanotoxins constitute a diversegroup of chemical substances categorised variously depend-ing upon their toxicity mechanism, organs affected (hepato-toxins, neurotoxins, dermatotoxins and cytotoxins) andchemical structure (cyclic peptides, alkaloids and lipopoly-saccharides (LPS)).
Based on bioassay, a cyanotoxin may be cytotoxic orbiotoxic (Sivonen and Jones 1999). Cytotoxins (e.g.acutiphycins, indocarbazoles, mirabilene, isonitriles, para-
cyclophanes, scytophycins, tentazoles, tolytoxin, toyoca-mycins and tubercidin) are lethal to cultured mammaliancell lines but not to organs/systems wheras biotoxins affectthe organs and/or tissue systems. Due to their lethality andubiquity, biotoxins have received more attention; however,cytotoxins are equally important owing to their antimicro-bial and antitumor activities (Jaiswal et al. 2008) andpotentially can be exploited for therapeutic use (Gademanand Portman 2008). Table 1 lists common cyanobacterialbiotoxins and their characteristics.
Production of cyanotoxins varies with species, strains,culture conditions, age and other environmental factors(Ouellette and Wilhelm 2003). A toxic cyanobacterium mayor may not produce toxins. A cyanobacterium may produceseveral toxins simultaneously, but only one or two usuallydominate (Ouellette and Wilhelm 2003; Codd et al. 2005).Cyanotoxin production varies with growth phase (highertoxin content at late-log phase), environmental factors suchas solar radiation (increased toxicity with increasingirradiance), water temperature (lower toxin concentrationat sub- and super-optimal temperature), pH (greater at sub-
Table 1 Attributes of some well-known cyanobacterial biotoxins (Hitzfeld et al. 2000; Jaiswal et al. 2008)
Cyanotoxins/Type LD50 (μg kg-1 i.p.mouse)
Structure Mode of action Organisms
Hepatotoxins
Microcystins 50≥1,200 Hepta-cyclicpeptides
Affects cytoskeleton ofhepatocytes, tumourpromotion by inhibitingprotein phosphatase 1and 2A
Microcystis aeruginosa, Planktothrix sp.,Nostoc sp., Anabaena sp., Anabaenopsis sp.,Synechocystis sp., Cyanobium bacillare,Arthrospira fusiformis, Limnothrix redekei,Phormidium formosum, Hapalosiphonhibernicus, Radiocystis feernandoi,Fischerella sp. strain CENA161
Nodularins 50–2,000 Penta-cyclicpeptides
Affects cytoskeleton ofhepatocytes by inhibitingprotein phosphatase 1and 2A
Nodularia spumigena
Cylindrospermopsin 2,000 Alkaloid Block protein synthesis Cylindrospermopsis raciborskii, Aphanizomenonovalisporum, Aphanizomenon flos-aquae,Umezakia natans, Raphidiopsis curvata,Anabaena japonica, Anabaena bergii
Neurotoxins
Anatoxins A 200–250 Alkaloid, secondaryamine
Inhibits the release ofacetylcholine by neurons(post-synaptic depolarization)
Anabaena spp., Planktothrix spp.,Aphanizomenon spp., Cylindrospermum sp.,Raphidiopsis mediterranea, Microcystis sp.
Anatoxins A(s) 20 Alkaloid Inhibits acetylcholinesterase Anabaena lemmermanni, Anabaena flos-aquae,Aphanizomenon flos-aquae
Homoanatoxin A – Alkaloids Inhibit post-synapticdepolarization
Oscillatoria Formosa, Raphidiopsismediterranea
Saxitoxins (PSP) 10 Corbamate alkaloid Inhibits nerve conductionby blocking Na channels
Anabaena circinalis, Aphanizomenon flos-aquae,Cylindrospermum raciborskii, Lyngbya wollei
Dermatotoxins
LPS endotoxins – Lipopolysaccharides Skin and mucosa All cyanobacteria; Lyngbya sp., Schizothrix sp.,Planktothrix sp.
i.p. intra-peritoneal administration of toxin
J Appl Phycol (2011) 23:1059–1081 1061

and super-optimal pH), nitrogen and carbon (their limitationcauses 5–6-fold decrease), iron concentration and N/P ratio(see for example, Veize et al. 2002; Jiang et al. 2008;Jaiswal et al. 2008; Li et al. 2009). The ecological andphysiological significance of cyanotoxin production is notyet clear (Paerl et al. 2001). They are supposed to act asdeterrent against zooplankton grazing, in metal ion chela-tion, intraspecific signalling and in allelopathatic interac-tions (Dittmann et al. 2001; Rohrlack et al. 2001;Pflugmacher 2002; Prince et al. 2008).
Cyanotoxins are either membrane-bound or occur freewithin the cells. They are released into the medium (naturalecosystems) with the ageing and death of the cyanobacterialpopulation and/or during water treatment processes where-in, they undergo photo- and bacterial degradation. Conse-quently, a significant fraction of the released cyanotoxinbecomes unavailable. They are also adsorbed on soilsurfaces depending upon environmental factors, soil prop-erty and total organic content of the soil (Tsuji et al. 2006;Edwards et al. 2008).
Cyanotoxins have adverse effect on other aquatic biota.Use of cyanobacteria infested waters for irrigation couldbe a threat to the yield and quality of crops (Zurawell et al.2005; Bibo et al. 2008). Primary symptoms associatedwith cyanotoxin-exposed plants include oxidative damageand growth inhibition (Chen et al. 2004; Järvenpää et al.2007; Pflugmacher et al. 2007; Saqrane et al. 2008). Inanimals (including humans), exposure to cyanotoxins ispossible through ingestion of contaminated food andwater, inhalation of the aerosolized toxins or living cells,dermal contact with toxic cyanobacteria and the toxinsreleased in milieu (Funari and Testai 2008). It was foundthat sensitivity to inhaled toxins is higher than toingestion, and differences in the route of exposuremanifest different clinical symptoms (Kirkpatrick et al.2004). Ingestion of cyanotoxin-contaminated products hasfar greater health significance than the exposure throughwater consumption or recreational activities (Funari andTestai 2008).
Cyclic peptides (microcystins and nodularins)
Cyclic peptides (esp., microcystins) are the most widelydistributed form of cyanotoxins. They are relatively small(MW 800–1,100 Da) in comparison to other cell oligopep-tides and polypeptides, and are synthesised non-ribosomally by a peptide synthetase (Tillett et al. 2000;Kaebernick et al. 2002). Hydrophilic in nature, cyclicpeptides enter cells through membrane transporters. Theyare extremely potent tumour promoters inhibiting proteinphosphatases (PP) 1 and 2A (Nishiwaki-Matsushima et al.1992) and the liver is the main organ affected by micro-cystins and nodularin.
Microcystins (MCs) are amongst the most studiedclass of cyanotoxins. They are cyclic heptapeptidedescribed as cyclo (-D-Alap1-Xaa2-D-MeAsp3-Yaa4-Adda5-D-Glu6-Mdha7-). The amino acids Xaa2 and Yaa4
are highly variable, and are responsible for a number ofMC variants. For example, MC-LR has amino acidsleucine (L) at Xaa2 and arginine (R) at Yaa4 positions(Gulledge et al. 2002). To date, more than 70 variants ofMCs are known (Jayaraj et al. 2006). Toxicity of the MCsresults exclusively due to Adda region [β-amino acid(2S,3S,8S,9S)-3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldecy-4(E), 6(E)-dienoic acid], which is covalentlybound to proteins (Hitzfeld et al. 2000; Gulledge et al.2002). Reports of liver cancer due to consumption ofmicrocystins containing drinking water have increasedsubstantially recently (Ouellette and Wilhelm 2003; Coddet al. 2005). Krakstad et al. (2006) reported that the PP-inhibiting toxins induce morphological alteration compatiblewith apoptosis in different cell types. Ding et al. (1999)found that MC-LR has strong mutagenicity and inducedDNA damage in rat hepatocytes. MCs also enhanced bonemarrow micronucleated polychromatic erythrocytes in mice.The World Health Organization has set a threshold value of1 μg L−1 for MCs in drinking waters. In June 2006, MC-LRwas classified as a possible human carcinogen (group 2B)(Grosse et al. 2006). Chronic consumption of MCs present intap water (low dose) could be a substantial risk factor forliver and colorectal cancer (Hernández et al. 2009).
Nodularin (NODLN) is a cyclic pentapeptide composedof Adda and D-erythro-β-methylaspartic acid as well as N-methyldehydrobutyrine. Saito et al. (2001) reported amodified structure of nodularins (NODLN-Har) fromNodularia PCC 7804 containing homoarginine instead ofarginine. Nodularin acts similar to microcystins except thatit does not bind covalently to PP1 or PP2A (Bagu et al.1997).
Alkaloids
Alkaloids constitute a broad group of heterocyclic nitrog-enous compounds of low to moderate molecular weight(<1 kD). They often undergo spontaneous transformation toform by-products. The cyanobacterial alkaloids includeneurotoxins, cytotoxins and dermatotoxins. Neurotoxicalkaloids are extremely potent toxins resulting in death ofmice within 2–30 min of exposure. Anatoxin A is an aminealkaloid, a structural analogue of cocaine with molecularweight of 165 Da. It causes depolarising neuromuscularblockade leading to respiratory failure (Fawell et al. 1999).Homoanatoxin A (MW 179 Da) is a less toxic ethyl ketoneanalogue of anatoxin A, whereas anatoxin A(s) (MW252 Da) is a phosphate ester of cyclic N-hydroxy guanineacting as a potent inhibitor of cholinesterase. It has the
1062 J Appl Phycol (2011) 23:1059–1081

property of organophosphorus insecticide and causeshypersalivation. Saxitoxins and neosaxitoxins are purinealkaloids. They inhibit nerve conduction by blockingsodium channels (Carmichael 1994). Saxitoxin (tricyclicalkaloid) is structurally related to tetrodotoxin. It blocksneuronal transmission by binding to sodium channels innerve cells leading to muscle paralysis. People consumingsaxitoxin-contaminated shellfish suffer from paralytic shell-fish poisoning (Hitzfeld et al. 2000). Cylindrospermopsis(CYN) contains a uracil fragment to a tricyclic guanidiniummoiety; it acts through DNA intercalation followed bystrand cleavage (Metcalf et al. 2004). According to Bankeret al. (2001), the pyrimidine ring is essential for CYNtoxicity. Furthermore, cytochrome p450 is required for theactivation of CYN toxicity (Froscio et al. 2003).
Cytotoxic alkaloids such as cyclic guanidine (MW415 Da) are potent inhibitors of protein synthesis andcause fatty liver and centralobular necrosis. Aplysiatoxins(dermatotoxic alkaloid) are protein kinase C activatorspromoting inflammation and tumour development (Myn-derse et al. 1977) while, lyngbyatoxin A causes dermati-tis, severe skin, oral and gastrointestinal inflammation(Cardellina et al. 1979).
Lipopolysaccharides (LPS)
LPS are an integral component of the cell wall ofcyanobacteria. They form complexes with proteins andphospholipids. Cyanobacterial LPS are composed of acarbohydrate polymer and an activated glycolipid (Stewartet al. 2006). They differ from bacterial LPS by having avariety of long chain unsaturated fatty acids and hydroxylfatty acids, and lack phosphate. LPS cause skin, gastroin-testinal and respiratory irritations as well as allergenicresponse in human and mammals (Sivonen and Jones 1999;Funari and Testai 2008). Tanikolide 1, is an antifungalcompound isolated from the lipid extract of Lyngbyamajuscula (Singh et al. 1999). Whereas, hierridin B, is anantiplasmodial compound obtained from lipophilic extractof Phormidium ectocarpi (Papendorf et al. 1998).
Some incidences of human toxicoses
In the majority of cases, toxicoses and fatalities related tocyanotoxins are due to ingestion of contaminated waters.Exposed organisms can experience acute to lethal toxicitydepending upon the bloom density, toxin content and thequantity of water ingested. Size of the organisms andspecies sensitivity also influences the severity of intoxica-tion. Monogastric animals are less sensitive than ruminantsand birds. Hitzfeld et al. (2000) have given a list ofincidences related to animal and human poisoning bycyanotoxins.
Poisoning can also result by bioaccumulation of cyano-bacterial toxins in food chains. Prepas et al. (1997) reportedthat mussels (an important food source of water rats,muskrats and birds) inhabiting Microcystis-containing waterbodies accumulated high amount of MCs. Consumption offish from water bodies infested by harmful toxic cyanobac-teria may affect the human population. For instance,Yuksov–Sortlav disease in the erstwhile USSR resultedfrom the consumption of cyanotoxin-containing rainbowtrout and common carp (Codd et al. 1989).
Cyanotoxins have been found to be responsible forgastroenteritis. In Paulo Afonso, Brazil, 88 people died(out of total 2,000 cases) due to of gastroenteritis. Theincidence took place after flooding in Itaparica dam in1988. Clinical investigations of the patients revealed thatcyanotoxin from Anabaena and Microcystis were respon-sible for the casualties (Teixeira et al. 1993). Cases of liverdamage have been reported from Armidale and PalmIsland (Australia) after copper treatment of the watersdominated by toxin producing cyanobacteria (Falconer etal. 1983; Hawkins et al. 1985). The best-documentedincidence of human casualties due to cyanotoxins is fromCaruaru, Brazil where in February 1996, 126 patientsdeveloped illness with acute neurotoxicity and subacutehepatotoxicity after dialysis, and 60 patients died inhospital. Later, it was found that water for dialysis camefrom a local reservoir infested with heavy cyanobacterialbloom (Pouria et al. 1998; Jochimsen et al. 1998). Furtheranalysis of the reservoir water revealed the presence ofmicrocystins YR, LR and AR.
Beneficial use of cyanotoxins
Cyanotoxins show biocidal properties (e.g. algaecides,herbicides and insecticides), and could be used forcommercial development of these compounds (Berry et al.2008). They are short-lived; hence, have little adverse effect(compared with synthetic biocides) on the general health ofecosystems. However, information regarding ecologicalimplications of the potential use of these compounds asbiocides is limited and requires further study. The thera-peutic value of cyanotoxins has been least explored.Compounds exhibiting cytotoxic activity could proveimportant in anticancer compounds (Sirenko et al. 1999).Utilisation of the antiviral, antimicrobial, antifungal prop-erties of cyanotoxins needs investigation.
Other bioactive compounds
Cyanobacteria produce a number of bioactive compoundsof potential therapeutic use (Skulberg 2000; Newman et al.2003; Newman and Cragg 2004a, b; Tan 2007; Gademan
J Appl Phycol (2011) 23:1059–1081 1063

and Portman 2008; Jones et al. 2009; Tan 2010). Crypto-phycins (depsipeptides; peptides with an ester linkage)produced by Nostoc sp. GSV 224, are potent anticancerdrug (Schwartz et al. 1990; Trimurtulu et al. 1995; Barrowet al. 1995). They act as mitotic inhibitor by blocking thecell cycle at the G2/M phase via inhibition of tubulinpolymerization (Chen et al. 1998; Panda et al. 2000). So far,more than 25 different cryptophycins have been discovered(Eggen and Georg 2002). Another promising anticancercompound, borophycine is a boron containing polyketideisolated from Nostoc linkia (Hemscheidt et al. 1994).
Tolyporphin extracted from Tolypothrix nodosa exhibitsphotosensitising activity against tumour cells, and is 5,000-times more effective than the photodynamic treatment(photofrin II) (Morliere et al. 1998). Curacin A, obtainedfrom L. majuscula is an antiproliferative (antimitotic) agentand inhibits the growth of colon, renal and breast cancerderived cell lines (Gerwick et al. 1994; White et al. 1997;Verdier-Pinard et al. 1999; Jones et al. 2009). It is a potentcompetitive inhibitor of colchicine, and binds more tightlyto tubulin than colchicine (Verdier-Pinard et al. 1999).Bauerines A–C, which are indole alkaloids were isolatedfrom Dichtrix baueriana and act against herpes simplexvirus (HSV-2) (Larsen et al. 1994). Norharmane, isolatedform Nodularia harveyana shows anticyanobacterial activ-ity against other cyanobacteria (Volk 2005) thus, could beuseful in controlling harmful cyanobacterial blooms. Hapa-lindoles, fischerindoles and welwitindolinones (indolealkaloids) isolated from Hapalosiphon fontinalis, Fischer-ella musciola and H. weltwitschi, respectively, showedanti-algal, antimycotic and insecticidal properties (videGademan and Portman 2008). Tjipanazoles obtained fromTolypothrix tjipanasensis exhibited antifungal activities,especially against fungi responsible for rice blast andwheat leaf rust (Bonjouklian et al. 1991).
Fischerellin A, obtained from Fischerella muscicolaUTEX 1829 is a potent photosystem II inhibitor and showsactivity against cyanobacteria, eukaryotic algae and herbs(Hagmann and Juettner 1996; Srivastava et al. 1999).Symplostatin 1, isolated from Symploca hydnoides is ananalogue of dolastatin 10, and is active against solid tumour(Harrigan et al. 1998). Aeruginosin 103-A (a linearpeptide), isolated from Microcystis viridis (NIES-103)inhibits thrombin (Kodani et al. 1998). Calothrixins A andB from Calothrix spp. are pentacyclic metabolites with anindol (3, 2-j) phenanthridine ring system. They inhibit thegrowth of a chloroquine-resistant strain of the malariaparasite (Plasmodium falciparum) and human HeLa cancercells (Rickards et al. 1999). The violet pigment nostocineA, produced by Nostoc spongiaeforme TISTR 8169exhibited algicidal, herbicidal and antibiotic properties aswell as cytostatic activity against human tumour cell lines(Hirata et al. 1996). Ypaoamide, a herbivore antifeedant is
reported from L. majuscula (Nagle et al. 1996). Nostocy-clamide with antibiotic and algicidal activities has beenisolated from Nostoc 31 (Todorova et al. 1995). Nostocar-boline isolated from Nostoc 78-12A is a potent cholines-terase inhibitor, which may be helpful in Alzheimertreatment (Becher et al. 2005). Reshef et al. (1997) reportedsulphoquinorosyldiacyl glycerol from cyanobacteria, effec-tive agent against HIV. Besides these, cyanobacteriaproduce a range of protease inhibitors such as cyanopepto-lin A–D (Gademan and Portman 2008). Antillatoxin andkalkitoxin isolated from marine L. majuscula exhibitconcentration-dependent cytotoxicity. Neurotoxicity pro-duced by antillatoxin is acute, while kalkitoxin caused adelayed response (Berman et al. 1999). Sulfolipids, a newclass of anti-HIV compound from Lyngbya lagerheimii andPhormidium tenue inhibit syncytium formation upon HIVinfection (Gustafson et al. 1989). Cyanovirin N isolatedfrom Spirulina (Arthrospira) protects from HIV infectionby inhibiting the process of cell-to-cell and virus-to-cellfusion (Boyd 1997). Cacium spirulan (Ca-Sp; sulphatedpolysaccharide) derived from Spirulina selectively inhibitspenetration of viruses (Herpex Simplex, human cytomega-lovirus, measles, mumps, Influenza A and HIV-1) into hostcell, and reduces lung metastasis of melanoma cellsinhibiting invasion of basal membrane by the tumour cells(Hayashi et al. 1996a, b; Mishima et al. 1998).
Quite a few species of cyanobacteria has been examinedfor secondary metabolites and the number of novelbiologically active compounds indicates their immensepotential to synthesise useful metabolites (Singh et al.2005; Van Wagoner et al. 2007). Some bioactive com-pounds isolated between 2000 and 2009 from cyanobacteriaare listed in Table 2. Recently, Tan (2010) has reviewed theimportance of filamentous marine cyanobacteria in antican-cer drug discovery. A large number of these cyanobacterialsecondary metabolites are synthesised through the integra-tion of non-ribosomal peptide synthetases with polyketidesynthase (Gademan and Portman 2008). Cyanobactins areanother class of bioactive compounds (cyclic peptides)synthesised ribosomally. More than 100 of such compoundshave been identified through gene analysis such aslyngbyatoxin, pettellamides, ultithiacyclamides, lissoclina-mides (Jones et al. 2009).
Cyanobacteria can be used for the production ofrecombinant compounds. Boyd and Shoemaker (1999)reported that Nostoc ellipsosporum transformed withcyanovirins coding nucleotide sequences produced virucideproteins useful in HIV treatment. Synechococcus PCC 6301transformed with mosquitocidal toxin genes of Bacillusthuringiensis and Bacillus sphaericus appeared promisingfor use in the natural mosquito habitat, where it could beingested and digested by mosquito larvae (Sangthongpitaget al. 1996).
1064 J Appl Phycol (2011) 23:1059–1081

UV-screening compounds
Ultraviolet (UV) radiation has harmful effect on biologicalsystems. Owing to their long evolutionary history, cyano-bacteria have evolved protective mechanism against UVradiations. They synthesise UV-absorbing/screening com-pounds such as mycosporine-like amino acids (MAAs) andscytonemin (Fleming and Castenholz 2007). MAAs aresmall (<400 Da), colourless, water-soluble compoundscomposed of cyclohexenone or cyclohexenimine chromo-phores conjugated with the nitrogen substituent of an aminoacid to its imino alcohol. Scytonemins (544 Da) are yellow-brown lipid soluble pigments located in the extracellularpolysaccharide sheath of cyanobacteria. Manipulating en-vironmental conditions such as high temperature, nitrogendepletion, photooxidative stress, and periodic desiccationstress in presence of UV-A could enhance their biosynthesis(Fleming and Castenholz 2007; Fleming and Castenholz2008). Owing to its anti-inflammatory and antiproliferativeactivities, scytonemin has pharmacological potential. Scy-tonemin and MAAs could be used for the production ofsunscreens. Various aspects of cyanobacterial scytonemin
and MAA production have been reviewed by Rastogi andSinha (2009).
Protein and fine chemicals
Cyanobacteria are good source of pigments, vitamins,polysaccharides, proteins, pharmaceuticals and other bio-logically active compounds of high commercial value(Thajuddin and Subramanian 2005). However, this neces-sarily does not mean that cyanobacteria are superior toother producer organisms. Their use as a potential source ofneutraceuticals is likely to have certain advantages such ascost-effectiveness, large-scale production using photobio-bioreactors, and relatively easy genetic manipulation.However, there are no studies to substantiate these argu-ments. Chemical composition of algae is subjected toenvironmental conditions, production, processing, packag-ing and distributional activities, and cultivation techniques(Ortega-Calvo et al. 1993; Campanella et al. 1999).Synechococcus sp. MA (FERM P-15099) produces poly-β-hydroxybutyrate (a granular polyester) in high yield,
Table 2 Some bioactive compounds isolated from cyanobacteria (2000–2009)
Secondary metabolites Organism Bioactivity References
Symplostatin 3 Symploca sp. Anticancer Luesch et al. 2002
Dragonamide C, D Lyngbya polychroa Anticancer Gunasekera et al. 2008
Homodolastatin 16 Lyngbya majuscula Anticancer Davies-Coleman etal. 2003
Curacin A L. majuscula Anticancer Simmons et al. 2005
Pitipeptolides A, B L. majuscula Anticancer Luesch et al. 2001
Grassypeptolide L. majuscula Antiproliferative Kwan et al. 2008
Cyanovirin Nostoc ellipsosporum Anti-HIV Dey et al. 2000
Venturamide A, B Oscillatoria sp. Antimalarial Linington et al. 2007
Dragomabin L. majuscula Antimalarial McPhail et al. 2007
Dragonamide A, B L. majuscula Antimalarial McPhail et al. 2007
Carmabin A, B L. majuscula Antimalarial, anticancer McPhail et al. 2007
Dolastatins Lyngbya sp., Symploca sp. Antimalarials, anticancer Fennell et al. 2003
Nostocarboline Nostoc sp. Antimalarial, cholinesterase inhibitor Barbaras et al. 2008
Symplocamide A Symploca sp. Antimalarial, anticancer Linington et al. 2008
Lyngbyastatin Lyngbya confervoides Serine protease inhibitor Taori et al. 2007
Anabaenopeptins Anabaena 90 Serine protease inhibitor Tonk et al. 2009
Cyanopeptolins Microcystis PCC 7806 Chymotrypsin inhibitor Bister et al. 2004
Norharmane Nodularia harveyana Enzyme inhibitor Volk 2006
Microviridin J Microcystis spp. Trypsin inhibitor Rohrlack et al. 2003
Bis-(χ-butyrolactones) Anabena variabilis Antibiotic Ma and Led 2000
Nostocine A Nostoc spongiaeforme Antibiotic Hirata et al. 2003
Kalkitoxin L. majuscula Block Na+ channels of nerve cells Wu et al. 2000
β-N-methylamino-L alanine
Microcystis, Anabaena, Nostocand Planktothrix spp.
Lateral sclerosis/parkinsonism-dementiacomplex
Cox et al. 2005
Spiroidesin Anabena spiroides Inhibits cell growth Kaya et al. 2002
J Appl Phycol (2011) 23:1059–1081 1065

which might be used in manufacturing biodegradablethermoplastic resins (Asada et al. 1999).
γ-Linolenic acid
The lipid content in cyanobacteria is generally low butSpirulina contains 6–13% lipid on dry weight basis, half ofwhich is fatty acids (Cohen 1996). Gamma-linolenic acid(GLA) is a rare polyunsaturated fatty acid with array ofmedicinal properties. It is 170-fold more effective thanlinoleic acid, the major constituent of most polyunsaturatedoils (Huang et al. 1982). GLA is effective for treatment ofatopic eczema (Biagi et al. 1988), premenstrual syndrome(Horrobin and Huang 1983), heart diseases (Kernoff et al.1977), Parkinson’s disease and multiple sclerosis (Dyerberg1986). GLA lowers low-density lipoprotein in hypercho-lesterolemic patients (Cohen 1996). It also acts as aprecursor for prostaglandins, the master hormones thatcontrol many body functions (Horrobin 1981). For instance,prostaglandin E1 is involved in regulating blood pressure,cholesterol synthesis, inflammation and cell proliferation(Horrobin 1981; Horrobin and Huang 1983; Richmond1988). Spirulina (Arthrospira) is the richest source of GLA,i.e. 20–25% of the lipid fraction compared with only 9% forevening primrose oil (Ciferri 1983). Aphanizomenon flos-aquae contains significant amount of ω-3-α-linoleic acidwhich inhibits the formation of inflammatory prostoglan-dins and arachidonic acid (Kushak et al. 2000). Productionof GLA in cyanobacteria is regulated by temperature, lightintensity, growth phase and nitrogen status of the medium(Piorreck et al. 1984; Cohen et al. 1987) and studiesfocusing on the optimization of GLA and other naturalproducts are much needed.
Vitamins
Cyanobacteria are an excellent source of vitamins. Forexample, Spirulina (Arthrospira) is claimed to be therichest whole-food source of vitamin B12 (two to six timesricher than raw beef liver) and vitamin E, with 20 g ofSpirulina fulfilling the daily body requirements of vitaminB12 and 70% of B1 (thiamine), 50% of vitamin B2
(riboflavin) and 12% of B3 (niacin) (Switzer 1981).Spirulina is also a good source of tocopherol (vitamin E)(Hudson and Karis 1974) with the reported content(190 mg kg−1) beeing three times more than that of purewheat germ. A. flos-aquae, Anabaena hassali, Microcystispulverana, Nostoc punctiforme and Phormidium bijugatumcontain the B-complex vitamins, nicotinic acid and pento-thene (Robbins et al. 1951; Koptera 1970a, b). Phytol, aconstituent of chlorophyll is also a suitable precursor for thesynthesis of several vitamins (A, E, K and K2).
Pigments
Cyanobacteria are a rich source of pigments such aschlorophyll a, carotenoids and phycobiliproteins. Thephycobiliproteins (phycocyanin, allophycocyanin and phy-coerythrin) occur only in the cyanobacteria, red algae andcryptophytes. They are an alternative to synthetic pigmentsand can be used as a natural food colouring agent (candy,ice creams, dairy products and soft drinks), drug andcosmetics, and have economic potential. Buffer extract ofphycocyanin from Spirulina is used in eye shadow, eyelinerand lipsticks. Since the product is water insoluble, it doesnot fade with water or sweat, neither irritates the skin.Besides its free radical scavenging effect, c-phycocyaninalso acts as a selective inhibitor of cytochrome oxydase 2,and has hepatoprotective and anti-inflammatory effect(Reddy et al. 2000). Its fluorescent property is used inanalytical techniques such as immunoassay analysis andflow cytometry (Campanella et al. 1999).
Carbohydrates
Carbohydrates, especially the compatible solutes glucosylglycerol, trehalose and sucrose are synthesised by cyano-bacteria under different osmotic stresses (Mackay et al.1984). Sugars have important industrial application espe-cially in bioethanol production. In Spirulina, carbohydratesaccount for 15% to 22% of dry weight (Santillan 1982),with rhamnose as the main polysaccharide. Borowitzka etal. (1980) found that altering the salinity level of theexternal growth medium increased the amount of osmoreg-ulatory carbohydrates in Synechococcus. Niederholtmeyeret al. (2010) transformed Synechococcus elongatusPCC7942 with transporter genes encoding invertase, glu-cose facilitator, lactate dehydrogenase and lactate transport-er to facilitate the secretion of glucose, fructose and high-value hydrophilic products such as lactate.
Exopolysaccharides
Many cyanobacteria produce exopolymers that are polysac-charidic in nature, known as exopolysaccharides (Cyano-EPS) (De Philippis and Vincenzini 1998). Their ecologicalrole include- protection against desiccation, salinity, UV-irradiation and predation; increase the availability of light,nutrient uptake, nitrogen fixation (prevent entry of oxygen inheterocytes) and movement (gliding) and in establishingsymbiotic association with other organisms (De Philippisand Vincenzini 1998; De Philippis et al. 2003; Pereira et al.2009 and references therein). More than a hundred cyano-bacterial strains, belonging to 22 different genera, are knownto produce EPS (De Philippis et al. 2001).
1066 J Appl Phycol (2011) 23:1059–1081

Cyano-EPSs are categorised into two types; the onesassociated with the cell surface and others which arereleased into the surrounding medium (released poly-saccharides, RPSs). The cyano-EPSs associated with thecell surface are referred as sheaths, capsules and slimes,based on their thickness, consistency and appearance (DePhilippis and Vincenzini 1998). There is no consensus onthe origin of RPSs in the surrounding medium. They canbe derived from the EPS and subsequently released intothe surrounding medium, or they may result frombiosynthetic pathways not directly related to the synthesisof EPS (Pereira et al. 2009). Cyano-RPSs can be easilyrecovered from the medium, and may have variousindustrial applications.
Cyano-EPSs are complex anionic heteropolysaccharideswith high molecular mass. They are mainly composed ofsix to ten different monosaccharides, one or more uronicacids, and non-saccharides (peptides, acetyl, pyruvyl,sulphate moieties etc.). EPSs from other microbial sources(e.g. bacteria or macroalgae) usually contain less than fourdifferent monomers (De Philippis and Vincenzini 1998). Sofar 12 different monosaccharides have been identified in thecyanobacteria. In the majority of cases, glucose is in thehighest concentration (De Philippis et al. 2003; Pereira etal. 2009). In addition, monosaccharides such as N-acetylglucosamine, 2,3-O-methyl rhamnose, 3-O-methyl rham-nose, 4-O-methyl rhamnose and 3-O-methyl glucose havealso been reported from cyano-EPSs (Hu et al. 2003). Sucha large number of different monosaccharides in a polysac-charide make many structures and architecture possible(Atkins 1986). The presence of two uronic acids, sulphate(except Archaea and eukaryotes) and both hydrophilic andhydrobhobic groups are unique to cyano-EPS, unlike thoseproduced by plants and other microorganisms (De Philippiset al. 2003). EPSs produced by Anabaena ATCC 33047,Cyanopira capsulata, Nostoc and Cyanothece are found topossess better rheological properties than the widely usedxanthan gum (De Philippis et al. 2000, 2001).
There are several factors regulating the production ofRPSs in cyanobacteria. They include-C/N and C/P ratios,light intensity, salinity, temperature, aeration and someinorganic molecules (Moreno et al. 2000; De Philippis et al.2001; Pereira et al. 2009). Knowledge about these factorsand other culture conditions are of great use in optimizationof the RPS production. In general, the chemical composi-tion, the type and the amount of the cyano-RPS producedby a particular cyanobacterium remain stable and, dependon the species and the cultivation conditions. However,sugar composition in RPSs may vary with the age of theculture (Pereira et al. 2009).
Their properties such as stabilising, suspending, thick-ening, gelling and water retention capacities have made
EPS a valuable industrial commodity. EPSs are generallyused in food, cosmetic, textile and painting industries asthickening, suspending or emulsifying agent. Furthermore,cyanobacterial polysaccharides with antiviral and immuno-modulatory properties may have application in medicine(Hayashi et al. 1996b) as well as in drug delivery. Theycould also be used to remove heavy metals and oil fromwastewaters and industrial wastes (De Philippis andVincenzini 1998; De Philippis et al. 2003; Pereira et al.2009). De Philippis et al. (2001) reported that residualcyanobacterial biomass (after RPS extraction) could beused as an effective cation-chelating material. Pereira et al.(2009) have reviewed the various aspects of cyano-EPS,which included their production, industrial applications andputative biosynthetic pathways.
Nutritional products
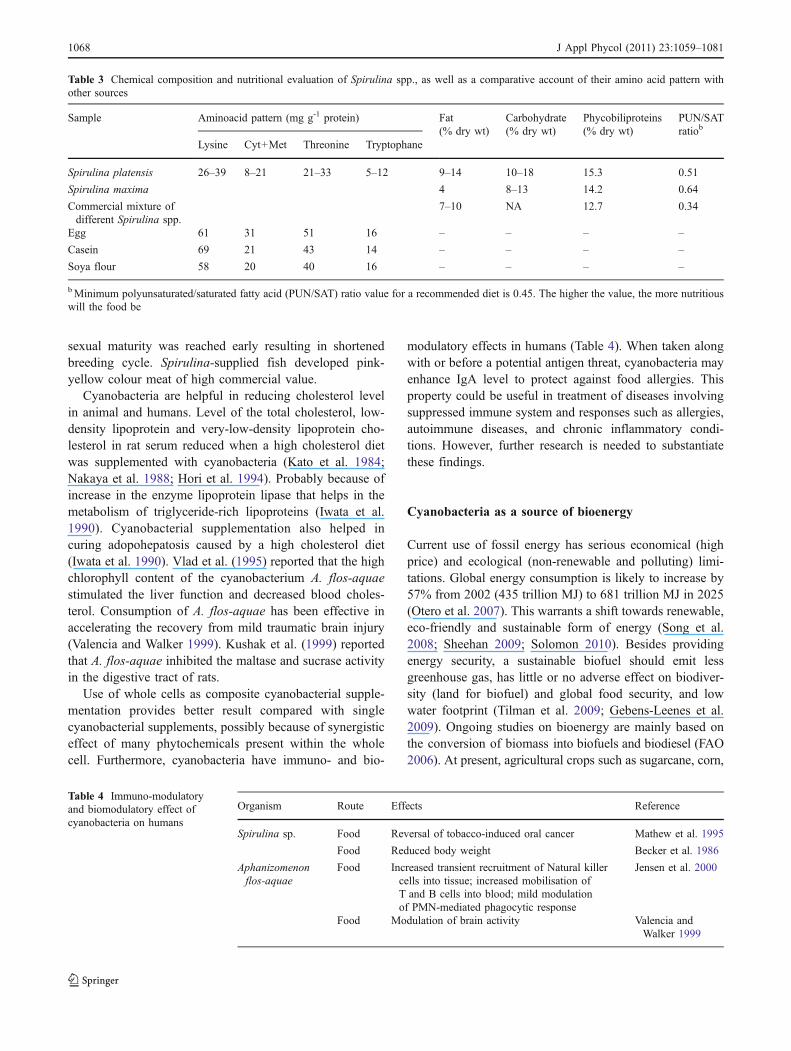
“Dihé” a sun dried hardened mat of Spirulina (Arthrospira)collected from freshwater bodies of Republic of Chad(Africa), is used by the native population to make sauces.The women of the area believe that if used by pregnantwomen, the dark colour of Dihé cakes will screen theunborn baby from the eye of sorcerers. Vonshak (1996)termed Spirulina (Arthrospira) a ‘magic agent’ due to itsutility as food, feed, cosmetics and anticancer agent. Itcontains about 70% of dry weight as protein (comparedwith 22% in beef), highest amongst the cyanobacteria(Table 3). However, Spirulina protein contains less methi-onine, cysteine and lysine, and is inferior to meat or milk(Ciferri 1983) but superior to other plant proteins includinglegumes. In Spirulina platensis, free fatty acids accountfrom 70% to 80% of the total lipids.
The carbohydrate content of S. platensis is 15–20% ofdry weight, composed of glucose and glycogen. It alsocontains all the vitamins, with cyanocobalamin the mostabundant (2 mg kg−1 of dried cells) (Santillan 1982). Totalnucleic acids content is less than 5% of the dry weight,much lower than bacteria and yeast (S. cerevisiae, the mosttypical source of single cell protein). Metabolism of purinesin humans produces uric acid, high level of which causesGout disease. Salazar et al. (1998) assessed the riskassociated with consumption of Spirulina and found noadverse effect on mice after subchronic treatment. Feedingof either the oil extract of Spirulina maxima or its defattedfraction prevented carbon tetrachloride-induced fatty liverdevelopment in rats thus, indicating its hepatoprotectiverole (Torres-Duran et al. 1999). Providing Spirulina to hensstimulated egg production and yolk coloration but not eggsize. Use of Spirulina in aquaculture increased the bodyweight of young shrimp and fish fry while in some species
J Appl Phycol (2011) 23:1059–1081 1067
sexual maturity was reached early resulting in shortenedbreeding cycle. Spirulina-supplied fish developed pink-yellow colour meat of high commercial value.
Cyanobacteria are helpful in reducing cholesterol levelin animal and humans. Level of the total cholesterol, low-density lipoprotein and very-low-density lipoprotein cho-lesterol in rat serum reduced when a high cholesterol dietwas supplemented with cyanobacteria (Kato et al. 1984;Nakaya et al. 1988; Hori et al. 1994). Probably because ofincrease in the enzyme lipoprotein lipase that helps in themetabolism of triglyceride-rich lipoproteins (Iwata et al.1990). Cyanobacterial supplementation also helped incuring adopohepatosis caused by a high cholesterol diet(Iwata et al. 1990). Vlad et al. (1995) reported that the highchlorophyll content of the cyanobacterium A. flos-aquaestimulated the liver function and decreased blood choles-terol. Consumption of A. flos-aquae has been effective inaccelerating the recovery from mild traumatic brain injury(Valencia and Walker 1999). Kushak et al. (1999) reportedthat A. flos-aquae inhibited the maltase and sucrase activityin the digestive tract of rats.
Use of whole cells as composite cyanobacterial supple-mentation provides better result compared with singlecyanobacterial supplements, possibly because of synergisticeffect of many phytochemicals present within the wholecell. Furthermore, cyanobacteria have immuno- and bio-
modulatory effects in humans (Table 4). When taken alongwith or before a potential antigen threat, cyanobacteria mayenhance IgA level to protect against food allergies. Thisproperty could be useful in treatment of diseases involvingsuppressed immune system and responses such as allergies,autoimmune diseases, and chronic inflammatory condi-tions. However, further research is needed to substantiatethese findings.
Cyanobacteria as a source of bioenergy
Current use of fossil energy has serious economical (highprice) and ecological (non-renewable and polluting) limi-tations. Global energy consumption is likely to increase by57% from 2002 (435 trillion MJ) to 681 trillion MJ in 2025(Otero et al. 2007). This warrants a shift towards renewable,eco-friendly and sustainable form of energy (Song et al.2008; Sheehan 2009; Solomon 2010). Besides providingenergy security, a sustainable biofuel should emit lessgreenhouse gas, has little or no adverse effect on biodiver-sity (land for biofuel) and global food security, and lowwater footprint (Tilman et al. 2009; Gebens-Leenes et al.2009). Ongoing studies on bioenergy are mainly based onthe conversion of biomass into biofuels and biodiesel (FAO2006). At present, agricultural crops such as sugarcane, corn,
Table 3 Chemical composition and nutritional evaluation of Spirulina spp., as well as a comparative account of their amino acid pattern withother sources
Sample Aminoacid pattern (mg g-1 protein) Fat(% dry wt)
Carbohydrate(% dry wt)
Phycobiliproteins(% dry wt)
PUN/SATratiob
Lysine Cyt+Met Threonine Tryptophane
Spirulina platensis 26–39 8–21 21–33 5–12 9–14 10–18 15.3 0.51
Spirulina maxima 4 8–13 14.2 0.64
Commercial mixture ofdifferent Spirulina spp.
7–10 NA 12.7 0.34
Egg 61 31 51 16 – – – –
Casein 69 21 43 14 – – – –
Soya flour 58 20 40 16 – – – –
bMinimum polyunsaturated/saturated fatty acid (PUN/SAT) ratio value for a recommended diet is 0.45. The higher the value, the more nutritiouswill the food be
Organism Route Effects Reference
Spirulina sp. Food Reversal of tobacco-induced oral cancer Mathew et al. 1995
Food Reduced body weight Becker et al. 1986
Aphanizomenonflos-aquae
Food Increased transient recruitment of Natural killercells into tissue; increased mobilisation ofT and B cells into blood; mild modulationof PMN-mediated phagocytic response
Jensen et al. 2000
Food Modulation of brain activity Valencia andWalker 1999
Table 4 Immuno-modulatoryand biomodulatory effect ofcyanobacteria on humans
1068 J Appl Phycol (2011) 23:1059–1081

sugar beet, rapseed, soybean, jatropa etc. are mainly used forbiofuel production (Gebens-Leenes et al. 2009). In currentworld bioethanol production, 60% comes from sugar crops,33% from other sources and 7% from chemical synthesis(vide Dexter and Fu 2009, p. 857). However, use of cropplants for biofuel production has invited some criticism(Tilman et al. 2009). Plants such as switchgrass and cornhave energy conversion efficiency of about 1% and 0.25%,respectively (Masukawa et al. 2002). On the other hand,algae can capture up to 10% of solar energy as biomass(Niederholtmeyer et al. 2010).
In biomass-based bioenergy production processes, bio-molecules such as sugars and polysaccharides (fermentationof carbohydrates into ethanol; first-generation technology),or residual parts such as cellulose (also de novo synthes-ised) are converted into biofuels (Second-generation tech-nology). However, these processes are low in efficiency andface constraints due to limited capacity of the organisms tostore the substrates intracellularly (Angermayr et al. 2009).Owing to their simple growth requirements and easygenetic manipulation, cyanobacteria have emerged as aviable option for biofuel production (Nobles et al. 2001;Koksharova and Wolk 2002; Tamagnini et al. 2007;Ananyev et al. 2008; Brown and Nobles 2008; Fu 2009;Angermayr et al. 2009 and references therein). Brown andNobles (2008) have developed strains of cyanobacteria thatproduce biofuel feedstocks (sugar and cellulose) in excess,and secret them into the liquid (growth) medium. This avoidsthe harm to the cells, and facilitates the continuous productionof feedstocks without affected by harvesting process.
Hellingwerf and Teixeira de Mattos (2009) proposed a‘Photanol Approach’ to maximise the production ofbiofuels. In this approach, a photosynthetic organism is tobe genetically modified in such a way (photosyntheticability is coupled with chemotrophy) that it convertsmetabolic intermediates directly into a biofuel productthrough the addition of a heterologous fermentativemetabolic pathway (chemotrophy) to the photoautotroph(Hellingwerf and Teixeira de Mattos 2009; Angermayr etal. 2009) (Third and fourth-generation technology).
Cyanobacteria can also be designed to produce ethanolwith high efficiency by the combination of gene transfor-mation, strain/process development and metabolic model-ling/profiling analysis. Trichodesmium, Synechococcus andSynechocystis are the best-characterised cyanobacteria forbiofuels production. Deng and Coleman (1999) transformedSynechococcus sp. PCC 7942 with pyruvate decarboxylaseand alcohol dehydrogenase genes of Zymomonas mobilis toproduce ethanol. Dexter and Fu (2009) successfullytransferred the pyruvate decarboxylase (pdc) and alcoholdehydrogenase II (adh) genes from Z. mobilis (a prolificethanol producer) into Synechocystis sp. PCC 6803 usinglight-driven psbA2 promoter.
Large-scale cyanobacterial biomass production could beincreased by adopting advanced photobioreacter technology(Gordon and Polle 2007; Lehr and Posten 2009) and supplyof low energy artificial light with optimal property forphotosynthesis (Park et al. 2000). However, the so-produced biofuel is costly and therefore unable to competewith fossil fuels. Mussgnug et al. (2007) increased thephotosynthetic efficiency by modulating the size of anten-na. In contrast, Walker (2009) reported that microalgaeincluding cyanobacteria are intrinsically not more but lessproductive than crop plants. He argued that use of algae forbiofuel production has only marginal contribution to ourpresent energy requirement as well as in carbon sequestra-tion. Besides, ethanolic fuel has certain disadvantages suchas: high solubility in water necessitates energy–intensivedistillation, has reduced energy per gallon content com-pared with petroleum, due to hygroscopic nature difficult indistributing through pipelines, and engine incompatibility athigher blending volume (Otero et al. 2007).
Hydrogen energy is a possible alternative energy source.It is an ecofriendly, efficient, renewable, carbon neutral andcommercially viable option of energy (Dutta et al. 2005).Only microorganisms are capable of producing hydrogen.Microbial hydrogen production is either light mediated (e.g.photosynthetic bacteria and cyanobacteria), or fermentingorganic sources in dark (e.g. members of Enterobacteria-ceae). In dark fermentation, the reaction rate is high, butyield is only 10–20% of hydrogen stored in the substrate,due to incomplete decomposition of the substrates. Fur-thermore, hydrogen yield varies with the type of substrateused. A major problem associated with this process iscontamination of hydrogen consuming microbes such asmethanogene and acetogenic bacteria that convert hydrogeninto methane and acetate.
Cyanobacteria are a source for photo-hydrogen production.The reported cost of photobiologically produced hydrogen ismuch less (US$25 m−3) to that of photovoltaic splitting ofwater (US$170 m−3) (Block and Melody 1992). Cyanobac-teria produce H2 through three main routes - mediated bybidirectional hydrogenase (Ananyev et al. 2008), by nitoge-nase (Lindberg et al. 2004), and through introduction of anefficient hydrogenase from non-cyanobacteria (Tamagnini etal. 2007; Angermayr et al. 2009). However, low rates ofhydrogen production, possibly due to complicated reactionsystem needed to overcome the large free energy i.e. +237 kJmole−1 hydrogen, limited duration of production and diffi-culty in collecting the gas are major drawbacks affectingcommercialization (Miyaki 1998; Chen et al. 2008). Howev-er, cyanobacteria-mediated hydrogen production has manyadvantages over other biological and non-biological processes(Benemann and Weissman 1976; Benemann et al. 1977;Karube et al. 1986; Madamwar et al. 2000; Dutta et al. 2005).Nearly 14 cyanobacterial genera are reported to produce
J Appl Phycol (2011) 23:1059–1081 1069

hydrogen gas (Dutta et al. 2005). According to an estimate, amarine Oscillatoria grown in a chamber of 1.86 m2×0.91 mdeep produced enough hydrogen to generate 1,000 kWelectricity per month (Mitsui 1980).
There are several extrinsic (light, temperature, salinity,micronutrients, carbon and nitrogen sources, oxygen, sulphurand methane) and intrinsic factors (genetic component andsensitive proteins in cyanobacteria) affecting cyanobacterialhydrogen production (Dutta et al. 2005; Angermayr et al.2009). Antal and Lindblad (2005) reported increased produc-tion of hydrogen by Gloeocapsa alpicola under sulphur-starved condition. Likewise, S. platensis and Anabaenacylindrica produced excess hydrogen under anaerobic dark(Aoyama et al. 1997) and light-limited conditions, respec-tively (Jeffries et al. 1978). Hydrogenase-deficient cyanobac-terium N. punctiforme NHM5 when incubated under highlight for a long period (until the culture depleted of CO2)showed increased hydrogen production (Lindberg et al. 2004;Dutta et al. 2005).
Photo-biological hydrogen production in cyanobacteria ismainly associated with nitrogenase rather than hydrogenase.Evolution of oxygen during N2-fixation is a major problemassociated with cyanobacterial hydrogen production. Praba-haran and Subramanian (1996) manipulated the growthconditions of Phormidium valderianum BDU 20041 (amarine, filamentous, non-heterocytous cyanobacterium) toovercome this problem. Under white light (5.5 μmol photonsm−2 s−1, 18:6 h dark/light cycle); temperature 27°C; pH 7.5;salinity 25 gL−1, P. valderianum BDU 20041 producedmaximum of 0.2 μmol hydrogen h−1 mg−1 dry wt in the gasphase without oxygen co-production. This was on a par withthe maximum yield reported in the literature. Masukawa etal. (2002) reported that knocking out the hupL gene inAnabaena sp. 2120 increased hydrogen production by 4 to 7times over wild species. This also increased energyconversion efficiency to 0.59% of total solar radiation.However, hydrogen production lasted only 30–40 h. Thetargeted energy conversion efficiency is 1% of total solarradiation (Masukawa et al. 2002). Madamwar et al. (2000)and Dutta et al. (2005) have reviewed the various aspectsand strategies to improve hydrogen production fromcyanobacteria.
Immobilisation of cyanobacteria in various matrices(polyurethane and polyvinyl foams) could be an interestingsolution to the problem of fuel, energy and chemicals.Markov (1997) patented a method for immobilisation ofcyanobacteria on the outer surfaces of fibres in the photo-reactor. When aerated aqueous salt solution is pumped,hydrogen produced by cyanobacteria seeps inside of thefibres and is collected in a vessel where the solution isheated (80–90°C) to separate the hydrogen gas.
Compared with photo-hydrogen production, much lessfree energy is required when cyanobacteria use organic
substrate. However, it depends on the growth phase.Fructose and sucrose are found to be the preferredsubstrates for hydrogen production (Chen et al. 2008).They also reported that, owing to less oxygen content, cellsat late-log phase produced more hydrogen than at logphase. Addition of fructose after saturation again raiseshydrogen production without initial delay. However, a highconcentration of fructose has inhibitory effect on hydrogenproduction (Chen et al. 2008). The maintenance ofsignificant hydrogen production rate over a long timerequires selection of species/strains for specific hydrogenmetabolism, followed by genetic engineering (if required)and overall improvement in growth conditions. With regardto biodiesel production, it is generally considered that,owing to their high lipid content, diatoms are idealcandidates for biodiesel production. However, a few speciesof cyanobacteria have also shown promise (Chisti 2007;Song et al. 2008).
Flocculants
Flocculants are used for sedimentation of colloidal substancesand cellular materials. Industries utilise them in clarificationof tap water, dredging and industrial processes. Fungi andbacteria are the main source for producing flocculatingsubstances. Cyanobacteria also produce exopolysaccharidesand flocculants (Bar-Or and Shilo 1987; Plude et al. 1991).Choi et al. (1998) reported a polysaccharidal flocculantcontaining neutral sugars, uronic acids and proteins fromAnabaena. Due to broad substrate specificity, rapid floccu-lating activity and thermal stability, it has a potential use inwastewater treatment and clarification of solar ponds.Emulcyan, a sulphated heteropolysacchride from PhormidiumJ-1 increases light availability in water column by flocculat-ing suspended clay particle (Fattom and Shilo 1984).
Aquaculture
In many parts of the world aquaculture is a major methodfor producing protein (crustaceans, molluscs and fishes).Management of algal production (as they are the primaryproducers) is an integral part of all aquacultural operations(Bardach et al. 1972). Algal production is a costly, oftenmajor cost item in aquacultural production (Pulz and Gross2004). Cyanobacteria act as food for molluscs, fishes andcrustaceans. It was reported that cyanobacterium Spirulinanot only enriches the protein content but also enhancescolour of the flesh of fish. Together with bacteria,cyanobacteria play an important role in maintaining O2
and CO2 balance of the water body thus, facilitatingaquaculture (Pruder 1983).
1070 J Appl Phycol (2011) 23:1059–1081

Wastewater treatment
The pollution of natural water bodies with organic wastesand heavy metals has emerged as a serious global problem.Pre-treatment of such wastewaters is essential beforedischarging them into waterways. There are differentphysico-chemical methods to remove these pollutants.However, they are often incomplete, labour and timeintensive, expensive, generate waste products and areunsatisfactory at high concentrations. Use of microalgaeand cyanobacteria provides an interesting alternative forwastewater treatment (Garbisu and Hall 1993; Mehta andGaur 2005). Cyanobacteria help in removing phosphorusand nitrogen from the polluted water with simultaneousproduction of biomass. They enrich the water with oxygenfacilitating further bacterial oxidation of organic matter.Strains such as Anabaena sp. PCC 7120 and N. ellipsospo-rum degrade refractory organic pollutants such as lindane(Kuritz and Wolk 1994). For example, pharmaceuticalindustries often produce troublesome wastes. Antibioticwastes impart objectionable odour as well as adverselyaffect the microflora of the receiving soil and water bodies.The marine cyanobacterium P. valderianum BDU30501degraded antibiotics such as penicillin and ampicillin, anduses them as a nitrogen source (Prabaharan et al. 1994).Furthermore, the strain was capable to grow and toleratehigh phenol concentration, and removed 38 mg L−1 phenolwithin a retention period of 7 days (Shashirekha et al.1997). Van Hille et al. (1999) used alkalinity resulting dueto the growth of Spirulina for precipitating heavy metalsfrom effluents.
Cyanobacteria are promising tool for the removal ofheavy metals from single as well as from multi-metalcontaining wastewaters (Mehta and Gaur 2005). Biosorp-tion involves binding of metals on the cell surfaces and tointracellular ligands, is a two-step process. In initial step,metals are rapidly adsorbed at the surface of the cells. It is apassive (hence, metabolically independent) and short-livedprocess. Followed by much slower but active (metabolicallydependent) uptake of metals into the cytoplasm (accumula-tion or subsequent conversion). The relative contribution toboth steps varies from species to species. In general,adsorption is mainly (80%) responsible for bulk heavy metalremoval (Mehta and Gaur 2005). Biosorption depends uponconcentration of metals and biomass in the environment,pH, temperature, presence of cations and anions, nutrientlevel, illumination and metabolic stage of the organisms(Mehta and Gaur 2005).
Heavy metal adsorption takes place either at cell wall(mainly exopolysachharides, EPS), or binding to cytoplas-mic ligands, phytochelatins and metallothioneins. The cellwall has many functional groups such as hydroxyl (-OH),phosphoryl (-PO3O2), amino (-NH2), carboxyl (-COOH),
sulphydryl (-SH) etc. (Yee et al. 2004; Gong et al. 2005;El-Sheekh et al. 2005). These provide negative charge tothe cell surface. Amongst these, carboxyl (-COOH) iswidely implicated in biosoption. Each functional grouphas a specific dissociation constant and dissociates intocorresponding anion and proton at a specific pH (Mehtaand Gaur 2005 and references therein). In cyanobacteria,the peptidoglycan layer and lipopolysachhides play animportant role in metal binding (Chojnacka et al. 2005).Substantial differences have been reported in the metalbinding capacity of EPS- and non- EPS-producingcyanobacteria; EPS-producers are preferable (De Philippiset al. 2003). The mechanisms behind biosorption includeion exchange, complexation, electrostatic attraction andmicro-precipitation. These vary with species and environ-mental conditions. Ahuja et al. (1999) considered ionexchange (with surface bound protons) as mechanisminvolved in most of the cases. Pre-treatment of biomassincreases their binding capacity. In case of S. maxima, pre-treatment with CaCl2 enhanced Pb2+ sorption capacity by84–92% (Gong et al. 2005). Similarly, introduction ofphosphate group in Lyngbya taylori increased its capacityto sorp Cd2+(Klimmek et al. 2001). Nagase et al. (2005)reported selective adsorption of Cd2+, Zn2+, Cu2+ and Pb2+
by cyanobacteria after the pre-treatment with 0.1 MNaOH.
Use of free cyanobacteria for wastewater treatment isimpractical (Mehta and Gaur 2005) therefore, suitablecyanobacteria are immobilised within or onto inert matrix.For immobilisation, natural (e.g. agar, gelatine, carrageenetc.) or synthetic materials (e.g. silica gel, polyacrylamideetc.) are used (Mehta and Gaur 2005). Furthermore,columns containing immobilised or granulated biomasscan be used for several sorption/desorption cycles, withadsorption capacity of the biomass remaining nearly similarfor many cycles. A suitable eluant (change in pH, organicor mineral acids, salt and metal chelators) can be used torecover metal from the column. Attempts are being made touse mat or biofilm forming (a self-immobilised system)cyanobacteria for large-scale and sustainable sorption.Generally, dead cells show better sorption than living cells.Though cost-effective, use of dead/dried cells has certaindemerits (Mehta and Gaur 2005). Cyanobacteria-mediatedmetal removal can also be used for the recovery of preciousmetals from wastewaters, and subsequently separated byelectrolysis.
Biodegradation of dyes
Synthetic dyes (methyl red, orange II, basic cataionicand basic fuchsin) are widely used in a number ofindustries such as textile, food, cosmetics and paper
J Appl Phycol (2011) 23:1059–1081 1071

printing. Because of their toxicity and carcinogenicity,inappropriate disposal present a potential environmentalconcern. Certain species of cyanobacteria (L. lager-heimii, N. linkia and Oscillatoria rubescens) have theability to degrade these dyes involving enzyme systems(El-Sheekh et al. 2009).
Role in agriculture
Nitrogen is an essential macronutrient limiting agricul-tural productivity, and is the largest and most costly inputin agriculture. Though atmospheric and dissolved dini-trogen (N2) in soil and water is in plenty however, due toits chemical inertia most plants (except those in symbioticassociations) are unable to utilise it. Only two groups oforganisms, free-living and symbiotic eubacteria includingcyanobacteria are capable to utilise N2 through the processof nitrogen fixation. However, this property is notuniversal even in these two groups. About 2×102 Mt ofnitrogen is fixed annually through biological nitrogenfixation (Guerrero et al. 1981).
Paddy fields are amongst the most suitable habitat fornitrogen fixing cyanobacteria. De (1939) for the first timeemphasized the importance of N2-fixing cyanobacteria inpaddy fields. In tropical soils, traditional rice varietiescommonly fulfil their nitrogen needs through cyanobac-teria. Metting (1981, 1988) estimated that total nitrogenfixation is 90 kg N ha−1 y−1. A visible growth ofcyanobacteria usually produces less than 10 kg N ha−1 y−1
whereas dense mats 10–20 kg N ha−1 y−1 (Aiyer et al.1972). Watanabe et al. (1977) estimated this value30 kg N ha−1 per rice crop.
Cyanobacteria are manageable systems, and can beproduced in bulk with ease. Soil-based cyanobacterialbiofertilizers can be produced as described by Venkataraman(1981) (Table 5). Usually mixed cyanobacterial cultures offree-living forms are used for algalization of the paddy fields(Venkataraman 1972; Roger and Kulasooriya 1980). Thefree-floating water fern Azolla is an agronomically importantplant; it harbours cyanobacterium Anabaena azollae in itsfronds. Rice seedlings grown with foam-immobilised A.azollae strains showed release of ammonium into thewater in paddy fields by cyanobacteria (Kannaiyan et al.1997). Mian (1984) using pot cultures showed thatnitrogen from free-living cyanobacteria is more readilyavailable to crops in comparison to cyanobacteria living inassociation.
Total expenditure for preparing the cyanobacterialinoculum is about one third of chemical fertilisers cost(Roger and Kulasooriya 1980). Dubey and Rai (1995)reported that Anabaena fertilissima and A. doliolum alone,or in combination with urea significantly increased the
biomass, grain yield and nutritive value of rice. Bothqualitative and quantitative improvements in rice produc-tion was maximal with 90 kgN ha−1 together with12.5 kg ha−1 cyanobacterial biofertilizer; use of latter savedabout 25% of the chemical N demand of the crop (Dubeyand Rai 1995). The availability of fixed N to the first cropis 23–28%, which increases in succeeding years (Watanabe1962).
Application of cyanobacteria not only provides nitrogenbut also plays an important role in mobilisation of inorganicphosphates by means of extracellular phosphatases andexcretion of organic acids (Bose et al. 1971; Rai andSharma 2006). Cyanobacteria also improve soil structure byproducing mucilage that helps in binding soil particles, andafter death and decay increase humus content of the soil,which leads to strong reducing conditions. Pre-soaking ofrice seeds with cyanobacterial cultures or extractsenhances germination and seedling growth. Possibly,due to the presence of vitamin B12 and amino acids inthe extract. However, no consistent report is available.Sergeeva et al. (2002) identified auxins from Nostoc andPrasanna et al. (2008) reported production of IAA byvarious cyanobacteria. Rodŕiguez et al. (2006) detectedgibberellin-like substances from Scytonema hofmanni.Cyanobacteria are known to produce iron chelators toensure the availability of iron (Gademan and Portman2008). The first report of cyanobacterial siderophoreschizokinen was from Anabaena (Nostoc) PCC 7120(Lankford et al. 1966). Synechbactin was isolated fromSynechococcus PCC 7002 (Ito and Butler 2005) andanachelins 1 and H from A. cylindrica 1403-2a (Beiderbecket al. 2000). Rastogi and Sinha (2009) have reviewed theliterature related to cyanobacterial siderophores and phyto-hormone production.
Table 5 Outdoor-soil-based cyanobacterial biofertilizer production(Venkataraman 1981)
Attribute Condition
Culture basin Shallow trays of galvanised iron sheet (5×1 m)
Nutrients Soil, 20 kg; superphosphate, 50 g and sodiummolybdate, 5 g
Water level 12–15 cm
Starter culture Cyanobacterial biomass
Aeration Medium to be kept in open air after sprinklingof the starter culture on the standing water
pH 7–7.6
Light Open sunlight
Retention time 7–8 days
Contaminationcontrol
To prevent mosquito breeding and other insects,50 g carbofuron (3% granules)
Yield 2 kg soil-based culture per basin
1072 J Appl Phycol (2011) 23:1059–1081

Green manuring
Some plants establish symbiotic association with cyanobacteria(Table 6). Amongst these the Azolla–Anabaena association isan effective green manure (due to high nitrogen-fixation rateand very rapid growth) for flooded crops, and has been in usefor centuries in China and Vietnam (Moore 1969; Lumpkinand Plucknett 1982). Its potential as an alternative orsupplemental nitrogen source for rice in tropical as well astemperate regions is well-established (Watanabe et al. 1977;Singh 1977; Talley et al. 1977). The association doubles itsbiomass in 25 days producing a biomass of 2.1 t ha−1 (Rogerand Watanabe 1984; Watanabe 1987). Under ideal condi-tions, it fixes more than 10 kg N ha−1 day−1 but much less(1.1 kg N ha−1 day−1) in field conditions (Watanabe 1977).This amount is sufficient to meet the entire N-requirement ofrice crop within a few weeks (Lumpkin 1987).
Because of the growing environment concerns, there is aneed to use non-polluting renewable resources. In this contest,application of Azolla as a biofertilizer is of much importance(Rai and Rai 1999). Three different methods are commonlypracticed in the use of Azolla as a green manure. Firstly, Azollais grown as a monocrop and incorporated into the paddy mudas a basal green manure before the rice is transplanted.Secondly, it is intercropped with rice, incorporated as a topdressing manure or allowed to die naturally without incorpo-ration. Thirdly, both monocropping and an intercropping areapplied simultaneously. Symbiotic Azolla reduces acetyleneat rates four to 18 times greater than that observed for free-living A. cylindrica (Tang et al. 1981). Azolla pinnata fixes75 mg N g dry wt−1 day−1 and produces biomass of 347tones fresh wt ha−1 y−1 containing 868 kg N (Singh 1988).
Restoration of soil fertility
“USAR” means unfertile soil containing low organicmatter, N and P. It includes all saline and alkaline soils
with general aridity of land, and canal irrigation as mainfactors for their development. A vast land area in India isUSAR. Singh (1950) had successfully reclaimed USARsoils through algalization for cultivation of the crops suchas rice and sugarcane in rotation. A sequential growth ofdifferent cyanobacteria was observed in rice fields. Out ofseveral genera involved in the process, Nostoc communeand A. fertilissima appear to be the most important (Singh1961). Soil cyanobacteria conserve organic matter, nitro-gen, phosphorus, moisture, and convert sodium clay intocalcium type probably through secretion of oxalic acid(Singh 1961).
Biological crusts occurring in semiarid and arid regionsof the world play an important role in maintaining andrestoring the ecosystem of the area (Belnap 2003). Naturaland anthropogenic disturbances result in the degradation ofbiological soil crusts. Under natural conditions, fullrecovery of such crusts may take decades. However,cyanobacterial inoculation greatly speeds up the recoveryprocess, and may further restore the ecological system(Wang et al. 2009)
Cyanobacterial biotechnology for useful productsand future research
Cyanobacteria are solar-powered natural cell factoriessynthesising an array of useful natural compounds forhuman welfare. We have just started to tap this resource.According to a conservative estimate, microalgal biomassmarket has a size of about 5,000 t y−1 of dry matteraccounting ca. US$1.25×109 y−1 (Pulz and Gross 2004).This estimate does not include processed products, biofuelmarket and environmental services. Authentic data oncyanobacteria-based market is lacking, which is mainlydominated by Spirulina (Arthrospira) (Gershwin and Belay2008). A successful biotechnology mainly depends on
Table 6 Some plants with N2-fixing cyanobacteria as symbiont
Plant Cyanobacteria
Type Genus Genus Location in plant
Algae (diatoms) Rhizosolenia, Chaetoceros Climacodium Richelia, Calothrix Intracellular
16S rDNA sequence is closely related to theCyanothece sp. ATCC 51142
Intracellular
Lichen Peltigera, Collema Nostoc, Calothrix, Scytonema, Fischerella Extracellular
Liverworts Blazia, Anthoceros, Nostoc Extracellular
Moss Sphagnum Haplosiphon Extracellular
Fern Azolla Anabaena azollae Extracellular
Cycads Cycas, Bowenia, Ceratozamia,Macrozamia
Anabaena, Nostoc, Calothrix Extracellular
Angiosperm Gunnera Nostoc punctiforme Intracellular
J Appl Phycol (2011) 23:1059–1081 1073

getting a suitable organism with desired property. As rawmaterials comprise the most significant cost-fraction inindustrial biotechnology (also White Biotechnology). In-dustrial biotechnology is a conversion of biomass viabiocatalysis, microbial fermentation or cell culture toprovide material, chemicals and energy. It has cost-competitiveness, environmental favourability and self-sustainability as essential characteristics. By 2010, indus-trial biotechnology is likely to have a 20% share in totalchemical production (Otero et al. 2007).
Cyanobacteria could contribute significantly in industrialbiotechnology. Formulation of appropriate culture conditionsand suitable culture media are the foremost requirements,followed by extensive screening programmes. Modern genet-ic techniques are capable of increasing the content of valuableproducts in a desired organism. Lack of cell differentiationand absence of allelic genes make cyanobacteria a muchsimpler system for genetic manipulation. However, theprogress is slow, and has drawbacks (Pulz and Gross 2004).To make production cost-effective, there is a need to adopt abiorefineries approach that includes simultaneous productionof other co-products (e.g. fine chemicals) with a mainproduct (say biofuel) (Otero et al. 2007).
An efficient and accurate method to identify toxic andnon-toxic cyanobacteria (nutritional vs. toxicological eval-uation) will enhance their commercial exploitation. Forexample, in the aquaculture industry lack of informationregarding nutritional value and toxicity of cyanobacteriagreatly affects the production. Furthermore, use of cyano-bacteria in rice fields has serious limitations. Normallygrowing cyanobacteria utilise much fixed N for their ownuse releasing only a small fraction of fixed nitrogen to theplants. Whereas, in fertilised fields, instead of fixingnitrogen, cyanobacteria start using the available nitrogenand become like weeds for the crop. Therefore, modifiedstrains capable to compete in field conditions could proveuseful. Studies indicate that nitrate assimilation defectivemutants have the property to fix N2 even in the presence ofhigh levels of nitrate. In such strains, nitrogenase is notrepressed by nitrate availability. Similarly, glutaminesynthetase defective mutants releasing ammonium contin-uously would be a better biofertilizer. If attempts tointroduce these mutants in natural habitats like floodedpaddy fields succeed, it will help agricultural productivityimmensely. Development of strains resistant to herbicidesand insecticides is also needed.
Conclusions
Cyanobacteria have the potential to help humankind innumber of areas including nutrition, agriculture, pharma-ceuticals, neutaceticals and biofuels. They have the unique
combination of plant-like photosynthesis and microbialability to yield high amount of desired products undercontrolled conditions. Whilst the rate of discovery ofnatural products from other microbes is decreasing, thepotential of cyanobacteria still remains largely unexplored.For optimised production of these compounds, there is aneed to combine metabolic engineering with advancesphtobioreactor technology. In the present global socio-economic setup, a move from academic conception (evensome patents) to industrial reality is desired. Despite allconceptual frameworks and positive laboratory findings, todate, we are unable to produce any of these products at acommercial level. Curently there are many claims, but veryfew industrial products on the market; mainly health foodsor niche products. A combination of rigorous research anddevelopmental planning is needed to compete in a globalmarket full of alternatives although it is tough consideringthe complicated establishing procedures and other financialconstrains. Favourable environmental and economicaspects, and largely unharnessed potentials make cyano-bacteria a promising future resource.
Acknowledgements NKS is thankful to the Indo-US Science andTechnology Forum, New Delhi for financial assistance in form of afellowship (IUSSTF Fellowship-2010).
References
Ahuja P, Gupta R, Saxena RK (1999) Zn2+ biosorption by Oscillatoriaanguistissima. Process Biochem 34:77–85
Aiyer RS, Sulahudean S, Venkataraman GS (1972) Long-termalgalization field trial with high yielding rice varieties. Indian JAgric Sci 42:380–383
Ananyev G, Carrieri D, Dismukes GC (2008) Optimization of metaboliccapacity and flux through environmental clues to maximizehydrogen production by the cyanobacterium “Arthrospira (Spir-ulina) maxima”. Appl Environ Microbiol 74:6102–6113
Angermayr SA, Hellingwerf KJ, Lindblad P, Teixera de Mattos MJ(2009) Energy biotechnology with cyanobacteria. Curr OpinBiotechnol 20:257–263
Antal TK, Lindblad P (2005) Production of H2 by sulphur-deprivedcells of the unicellular cyanobacteria Gloeocapsa alpicola andSynechocystis sp. PCC 6803 during dark incubation with methaneor at various extracellular pH. J Appl Microbiol 98:114–120
Aoyama K, Uemura I, Miyake J et al (1997) Fermentative metabolismto produce hydrogen gas and organic compounds in a cyanobac-terium, Spirulina platensis. J Ferment Bioeng 83:17–20
Asada Y, Miyake M, Miyake J et al (1999) Photosyntheticaccumulation of poly-(hydroxybutyrate) by cyanobacteria—themetabolism and potential for CO2 recycling. Int J Biol Macromol25:37–42
Atkins EDT (1986) Biomolecular structures of naturally occurringcarbohydrate polymers. Int J Biol Macromol 8:323–329
Bagu JR, Sykes BD, Craig MM, Holmes CFB (1997) A molecularbasis for different interactions of marine toxins with proteinphosphatase-1, molecular modes for bound motuporin, micro-cystins, okadaic acid and calyculin A. J Biol Chem 272:5087–5089
1074 J Appl Phycol (2011) 23:1059–1081

Banker R, Carmeli S, Werman M, Teltsch B, Porat R, Sukenik A(2001) Uracil moiety is required for toxicity of the cyanobacterialhepatotoxin cylindrospermopsin. J Toxicol Environ Health62:281–288
Barbaras D, Kaiser M, Brunb R, Gademann K (2008) Potent andselective antiplasmodial activity of the cyanobacterial alkaloidnostocarboline and its dimers. Bioorg Med Chem Lett 18:4413–4415
Bardach JE, Ryther JH, McLarney WO (1972) Aquaculture: thefarming and husbandry of fresh water and marine organisms.Wiley, New York
Bar-Or Y, Shilo M (1987) Characterization of macromolecularflocculants produced by Phormidium sp. strain J1 and byAnabaenopsis circularis. Appl Environ Microbiol 53:2226–2230
Barrow RA, Hemscheidt T, Liang J, Paik S, Moore RE, Tius MA(1995) Total synthesis of cryptophycins. Revision of thestructures of cryptophycins A and C. J Am Chem Soc117:2479–2490
Becher PG, Beuchat J, Gademann K, Jüttner F (2005) Nostocarboline:isolation and synthesis of a new cholinesterase inhibitor fromNostoc 78-12A. J Nat Prod 68:1793–1795
Becker E, Jakover B, Luft D, Schmuclling RM (1986) Clinical andbiochemical evaluations of the alga Spirulina with regard to itsapplication in the treatment of obesity: a double blind cross-overstudy. Nutr Rep Int 33:565–574
Beiderbeck H, Taraz K, Budzikiewicz H, Walsby AEZ (2000)Anachelin, the siderophore of the cyanobacterium Anabaenacylindrica CCAP 1403/2A. Z Naturforsch C 55:681–687
Belnap J (2003) The world at your feet: desert biological soil crusts.Front Ecol Environ 1:181–189
Benemann JR, Weissman JC (1976) Biophotolysis: problems andprospects. In: Schlegel HG, Bamen J (eds) Microbial energyconversion. Erich Goltze KG, Göttingen, pp 413–426
Benemann JR, Weissman JC, Koopman BL, Oswald WJ (1977)Energy production by microbial photosynthesis. Nature 268:19–23
Berman FW, Gerwick WH, Murray TF (1999) Antillatoxin andkalkitoxin, ichthyotoxins from the tropical cyanobacteriumLyngbya majuscula, induce distinct temporal patterns ofNMDA receptor-mediated neurotoxicity. Toxicon 37:1645–1648
Berry JP, Gantar M, Perez MH, Berry G, Noriega FG (2008)Cyanobacterial toxins as allelochemicals with potential applica-tions as algaecides, herbicides and insecticides. Mar Drugs6:117–146
Biagi PL, Bordoni A, Masi M, Ricci G, Fanelli C, Patrizi A, CeccoliniE (1988) A long term study on the use of evening primrose oil(Efamol) in atopic children. Drugs Exp Clin Res 14:285–290
Bibo L, Yan G, Bangding X, Jiantong L, Yongding L (2008) Alaboratory study on risk assessment of microcystin-RR incropland. J Environ Manage 86:556–574
Bister B, Keller S, Baumann HI, Nicholson G, Weist S, Jung G,Süssmuth RD, Juüttner FJ (2004) Cyanopeptolin 963A, achymotrypsin inhibitor of Microcystis PCC 7806. J Nat Prod67:1755–1757
Block DL, Melody I (1992) Efficiency and cost goals for photo-enhanced hydrogen production processes. Int J Hydrogen Energy17:853–861
Bonjouklian R, Smitka TA, Doolin LE, Molloy RM, Debono M,Shaffer SA, Moore RE, Stewart JB, Patterson GML (1991)Tjipanazoles, new antifungal agents from the blue-green algaTolypothrix tjipanasensis. Tetrahedron 47:7739
Borowitzka LJ, Demmerle S, Mackay MA, Norton RS et al (1980)Carbon-13 nuclear magnetic resonance study of osmoregulationin a blue-green alga. Science 210:650–651
Bose P, Nagpal US, Venkataraman GS, Goyal SK et al (1971)Solubilization of tricalcium phosphate by blue-green algae. CurrSci 40:165–166
Boyd M (1997) Protein isolated from blue-green algae inactivatesHIV.Antimicrob Agents Chemother 41:1521–1530
Boyd MR, Shoemaker RH (1999) Nucleic acids encoding antiviralcyanovirin proteins isolated from cyanobacteria such as Nostocellipsosporum useful for treating virus infections (especiallyhuman immune deficiency virus). Patent PN 5962668, USDepartment of Health-Human Service, Washington, DC, USA
Brown RM, Nobles DR (2008) The future of biofuels in renewableenergy and reduction of global warming. Available at: www.botany.utexas.edu
Burja AM, Banaigs EB, Abou-Mansour et al (2001) Marinecyanobacteria—a prolific source of natural products. Tetrahedron57:9347–9377
Campanella L, Crescentini G, Avino P (1999) Chemical compositionand nutritional evaluation of some natural and commercial foodproducts based on Spirulina. Analusis 27:533–540
Cardellina JH II, Marner FJ, Moore RE (1979) Seaweed dermatitis:structure of lyngbyatoxin A. Science 204:193–195
Carmichael WW (1994) The toxins of cyanobacteria. Sci Am 270:64–70
Castenholz RW (2001) Bergey’s manual of systematic bacteriology.Boone DR, Castenholz RW (eds), vol I, 2nd edn. Springer,Berlin. pp. 474-487
Chen BD, Nakeff A, Valeriote F (1998) Cellular uptake of a novelcytotoxic agent, cryptophycin-52, by human THP-1 leukaemiacells and H-125 lung tumor cells. Int J Cancer 77:869–873
Chen J, Son L, Dai J, Ganb N, Liua Z (2004) Effects of microcystinson the growth and the activity of superoxide dismutase andperoxidase of rape (Brassica napus L.) and rice (Oryza sativaL.). Toxicon 43:393–400
Chen PC, Fan SH, Chiang CL, Lee CM (2008) Effect of growthconditions on the hydrogen production with cyanobacteriumAnabaena sp. Strain CH3. Int J Hydrogen Energy 33:1460–1464
Chisti Y (2007) Biodiesel from microalgae. Biotechnol Adv 25:294–306
Choi CW, Yoo SA, Oh I-H et al (1998) Characterisation of anextracellular flocculating substance produced by a planktoniccyanobacterium, Anabaena sp. Biotechnol Lett 20:643–646
Chojnacka K, Chojnacki A, Górecka H (2005) Biosorption of Cr3+,Cd2+ and Cu2+ ions by blue-green algae Spirulina sp.: kinetics,equilibrium and the mechanism of the process. Chemosphere59:75–84
Ciferri O (1983) Spirulina, the edible microorganism. Microbiol Rev47:551–578
Codd GA, Bell SG, Brooks WP (1989) Cyanobacterial toxins in water.Water Sci Technol 21:1–13
Codd GA, Morrison LF, Metcalf JS (2005) Cyanobacterial toxins: riskmanagement for health protection. Toxicol Appl Pharmacol203:264–272
Cohen Z (1996) The chemicals of Spirulina. In: Vonshak A (ed)Spirulina platensis (Arthrospira): physiology, cell-biology, andbiotechnology. Taylor and Francis, London, pp 175–204
Cohen Z, Vonshak A, Richmond A (1987) Fatty acid composition ofSpirulina strains grown under various environmental conditions.Phytochemistry 26:2255–2258
Cox PA, Banack SA, Murch SJ, Rasmussen U, Tien G, Bidigare RR,Metcalf JS, Morrison LF, Codd GA, Bergman B et al (2005)Diverse taxa of cyanobacteria produce beta-N-methylamino-L-alanine, a neurotoxic amino acid. Proc Natl Acad Sci USA102:5074–5078
Davies-Coleman M, Dzeha TM, Gray CA, Hess S, Pannell LK,Hendricks DT, Arendse CE (2003) Isolation of homodolastatin
J Appl Phycol (2011) 23:1059–1081 1075

16, a new cyclic depsipeptide from a Kenyan collection ofLyngbya majuscula. J Nat Prod 66:712–715
De PK (1939) The role of blue-green algae in nitrogen fixation in ricefields. Proc R Soc Lond, B Biol Sci 127:121–129
De Philippis R, Vincenzini M (1998) Exocellular polysaccharidesfrom cyanobacteria and their possible applications. FEMSMicrobiol Rev 22:151–175
De Philippis R, Ena A, Paperi R, Sili C, Vincenzini M (2000)Assessment of the potential of Nostoc strains from PasteurCulture Collection for the production of polysaccharides ofapplied interest. J Appl Phycol 12:401–407
De Philippis R, Sili C, Paperi R, Vincenzini M (2001)Exopolysaccharide-producing cyanobacteria and their possi-bleexploitation: a review. J Appl Phycol 13:293–299
De Philippis R, Paperi R, Sili C, Vincenzini M (2003) Assessment ofthe metal removal capability of two capsulated cyanobacteria,Cyanospira capsulata and Nostoc PCC7936. J Appl Phycol15:155–161
Delwiche CF, Palmer JD (1997) The origin of plastids and their spreadvia secondary symbiosis. In: Bhattacharya D (ed) Origin of algaeand their plastids. Springer, Berlin, pp 53–96
Deng M-D, Coleman JR (1999) Ethanol synthesis by geneticengineering in cyanobacteria. Appl Environ Microbiol 65:523–528
DeRuyter YS, Fromme P (2008) Molecular structure of the photo-synthetic apparatus. In: Herrero A, Flores E (eds) The cyanobac-teria: molecular biology, genomics and evolution. CaisterAcademic Press, Norfolk, pp 217–270
Dexter J, Fu P (2009) Metabolic engineering of cyanobacteria forethanol production. Energy Environ Sci 2:857–864
Dey B, Lerner DL, Lusso P, Boyd MR, Elder JH, Berger EA (2000)Multiple antiviral activities of cyanovirin-N: blocking of HumanImmunodeficiency Virus Type I gp 120 interaction with CD4 andcoreceptor and inhibition of diverse enveloped viruses. J Virol74:4562–4569
Ding WX, Shen HM, Zhu HG, Ong CN (1999) Genotoxicity ofmicrocystic cyanobacteria extract of a water source in China.Mutat Res 442:69–77
Dittmann E, Erhard M, Kaebernick M, Scheler C, Neilan BA, vonDöhren H, Börner T (2001) Altered expression of two light-dependent genes in a microcystin-lacking mutant of Microcystisaeruginosa PCC 7806. Microbiol 147:3113–3119
Dubey AK, Rai AK (1995) Application of algal biofertilizers(Aulosira fertilissima tenuis and Anabaena doliolum Bhardwaja)for sustained paddy cultivation in Northern India. Isr J Plant Sci43:41–51
Dutta D, De D, Chaudhuri S, Bhattacharaya SK (2005) Hydrogenproduction by cyanobacteria. Microb Cell Fact 4:1–11
Dyerberg J (1986) Linoleate-derived polyunsaturated fatty acids andprevention of atherosclerosis. Nutr Res 44:125–128
Edwards C, Graham D, Fowler N, Lawton LA (2008) Biodegradationof microcystins and nodularin in freshwaters. Chemosphere73:1315–1321
Eggen MJ, Georg GI (2002) The cryptophycins: their synthesis andanticancer activity. Med Res Rev 22:85–101
El-Sheekh MM, El-Shouny WA, Osman MFH, El-Gammal WE(2005) Growth and heavy metals removal affinity of Nostocmuscorum and Anabaena subcylindrica in sewage and indus-trial wastewater effluent. Environ Toxicol Pharmacol 19:357–365
El-Sheekh MM, Gharieb MM, Abou-El-Souod GW (2009) Biodegra-dation of dyes by some green algae and cyanobacteria. IntBiodeterior Biodegrad 63:699–704
Falconer IR, Beresford AM, Runnegar MTC (1983) Evidence of liverdamage by toxin from a bloom of the blue-green alga Microcystisaeruginosa. Med J Aust 1:511–514
Fattom A, Shilo M (1984) Phormidium J-1 bioflocculant: productionand activity. Arch Microbiol 139:421–426
Fawell JK, Mitchell RE, Hill RE, Everett DJ (1999) The toxicity ofcyanobacterial toxins in the mouse: II Anatoxin-A. Hum ExpToxicol 18:168–173
Fennell BJ, Carolan S, Pettit GR, Bell A (2003) Effects of theantimitotic natural product dolastatin 10, and related peptides, onthe human malarial parasite Plasmodium falciparum. J Anti-microb Chemother 51:833–841
Fleming ED, Castenholz RW (2007) Effects of nitrogen source on thesynthesis of the UV screening compound, scytonemin, in thecyanobacterium Nostoc punctiforme PCC 73102. FEMS Micro-biol Ecol 63:301–308
Fleming ED, Castenholz RW (2008) Effects of periodic dessication onthe synthesis of the UV screening compound, scytonemin, incyanobacteria. Environ Microbiol 9:1448–1455
Food and Agricultural Organization (2006) Introducing the interna-tional bioenergy platform. FAO, Rome
Froscio SM, Humpage AR, Burcham PC, Falconer IR (2003)Cylindrospermopsin-induced protein synthesis inhibition and itsdissociation from acute toxicity in mouse hepatocytes. EnvironToxicol 18:243–251
Fu P (2009) Genome-scale modelling of Synechocystis sp. PCC 6803and prediction of pathway insertion. J Chem Technol Biotechnol84:473–483
Funari E, Testai E (2008) Human health risk assessment related tocyanotoxins exposure. Crit Rev Toxicol 38:97–125
Gademan K, Portman C (2008) Secondary metabolites from cyano-bacteria: complex structures and powerful bioactivities. Curr OrgChem 12:326–341
Gantar M, Svircev Z (2008) Microalgae and cyanobacteria: food forthought. J Phycol 44:260–268
Garbisu C, Hall DO (1993) Removal of phosphate by foamimmobilised Phormidium laminosum in batch and continuousflow bioreactor. J Chem Biotechnol 57:181–189
Gebens-Leenes W, Hoekstra AY, van der Meer TH (2009) The waterfootprint of bioenergy. Proc Natl Acad Sci USA 106:10219–10223
Gershwin ME, Belay A (2008) Spirulina in human nutrition andhealth. CRC Press, Boca Raton, p 312
Gerwick WH, Proteau PJ, Nagle DG, Hamel E, Blokhin A, SlateDL (1994) Structure of Curacin A, a novel antimitotic,antiproliferative and brine shrimp toxic natural product fromthe marine cyanobacterium Lyngbya majuscula. J Org Chem59:1243–1245
Gong R, Ding Y, Liu H, Chen Q, Liu Z (2005) Lead biosorption anddesorption by intact and pretreated Spirulina maxima biomass.Chemosphere 58:125–130
Gordon JM, Polle JEW (2007) Ultra-high bioproductivity from algae.Appl Microbiol Biotechnol 76:969–975
Gröniger A, Sinha RP, Klisch M, Häder DP (2000) Photoprotectivecompounds in cyanobacteria, phytoplankton and macroalgae—adatabase. J Photochem Photobiol, B Biol 58:115–122
Grosse Y, Baan R, Straif K, Secretan B, Ghissassi F, El Cogliano V(2006) Carcinogenicity of nitrate, nitrite, and cyanobacterialpeptide toxins. Lancet Oncol 7:628–629
Guerrero MG, Vega JM, Losada M (1981) The assimilatory nitratereducing system and its regulation. Annu Rev Plant Physiol32:169–204
Gulledge BM, Aggen JB, Huang HB, Nairn AC, Chamberlin AR(2002) The microcystins and modularins: cyclic polypeptideinhibitors of PP1 and PP2A. Curr Med Chem 9:1991–2003
Gunasekera SP, Ross C, Paul VJ, Matthew S, Luesch H (2008)Dragonamides C and D, linear lipopeptides from the marinecyanobacterium Lyngbya polychroa. J Nat Prod 71:887–890
Gustafson KR, Cardellina JH II, Fuller RW, Weislow OS, Kiser RF,Snader KM, Patterson GM, Boyd MR (1989) AIDS-antiviral
1076 J Appl Phycol (2011) 23:1059–1081

sulfolipids from cyanobacteria (blue green algae). J Natl CancerInst 81:1254–1258
Hagmann L, Juettner F (1996) Fischerellin-A, a novel photosystem-II-inhibiting allelochemical of the cyanobacterium Fischerellamuscicola with antifungal and herbicidal activity. TetrahedronLett 37:6539–6542
Harrigan GG, Luesch H, Yoshida YWY, WY MRE, Nagle DG, PaulVJ, Mooberry SL, Corbett TH, Valeriote FA (1998) Symplostatin1: a dolastatin 10 analogue from the marine cyanobacteriumSymploca hydnoides. J Nat Prod 61:1075–1077
Hawkins PR, Runnegar MTC, Jackson ARB, Falconer IR (1985)Severe hepatotoxicity caused by the tropical cyanobacterium(blue-green alga) Cylindrospermopsis raciborskii (Woloszynska)Seenaya and Subba Raju isolated from a domestic water supplyreservoir. Appl Environ Microbiol 50:1292–1295
Hayashi T, Hayashi K, Maedaa M, Kojima I (1996a) Calciumspirulan, an inhibitor of enveloped virus replication, from ablue-green alga Spirulina platensis. J Nat Prod 59:83–87
Hayashi K, Hayashi T, Kojima I (1996b) A natural sulfatedpolysaccharide, calcium spirulan, isolated from Spirulina platen-sis: in vitro and ex vivo evaluation of anti-Herpes simplex virusand anti-human immunodeficiency virus activities. AIDS ResHum Retrovir 12:1463–1471
Hellingwerf KJ, Teixeira de Mattos MJ (2009) Alternative routes tobiofuels: light-driven biofuel formation from CO2 and waterbased on the “Photanol” approach. J Biotechnol 142:87–89
Hemscheidt T, Puglisi MP, Larsen LK, Patterson GML, Moore RE,Rios JL, Clardy J (1994) Structure and biosynthesis ofborophycin, a new boeseken complex of boric acid from amarine strain of the blue-green alga Nostoc linckia. J Org Chem59:3467–3471
Hernández JM, López-Rodas V, Costas E (2009) Microcystins fromtap water could be a risk factor for liver and colorectal cancer: arisk intensified by global change. Med Hypothesis 72:539–540
Herrero A, Flores E (2008) The cyanobacteria: molecular biology,genomics and evolution. Caister Academic Press, Norfolk, p 484
Hirata K, Takashina J, Nakagami H, Ueyama KM, Kanamori T,Miyamoto K (1996) Growth inhibition of various organisms by aviolet pigment, nostocine-A, produced by Nostoc spongiaeforme.Biosci Biotechnol Biochem 60:1905–1906
Hirata K, Yoshitomi S, Dwi S, Iwabea O, Mahakhantb A, Polchaib J,Miyamoto K (2003) Bioactivities of nostocine A produced by afreshwater cyanobacterium Nostoc spongiaeforme TISTR 8169. JBiosci Bioeng 95:512–517
Hitzfeld BC, Hoeger SJ, Dietrich DR (2000) Cyanobacterial toxins:removal during drinking water treatment, and human riskassessment. Environ Health Perspect 108(S1):113–122
Hori KG, Ishibashi G, Okita T (1994) Hypocholesterolemic effect ofblue-green alga, ishikurage (Nostoc commune) in rats fedatherogenic diet. Plant Foods Hum Nutr 45:63–70
Horrobin DF (1981) The possible roles of prostaglandin E1 and ofessential fatty acids in mania depression and alcoholism. In:Holman RT (ed) Progress in lipid research, vol 20. PergamonPress, New York, pp 539–541
Horrobin DF, Huang YS (1983) Schizophrenia: the role of abnormalessential fatty acid and prostaglandin metabolism. Med Hypothesis10:329–336
Hu C, Liu Y, Paulsen BS, Petersen D, Klaveness D (2003)Extracellular carbohydrate polymers from five desert soil algaewith different cohesion in the stabilization of fine sand grain.Carbohydr Polym 54:33–42
Huang YS, Cunnane SC, Horrobin DF, Davignon J et al (1982) Mostbiological effects of Zn deficiency corrected by γ-linolenic acid18:3 ω6 but not by 18:2 ω6. Atherosclerosis 41:193–208
Hudson BZF, Karis IG (1974) The lipids of the alga Spirulina. J SciFood Agric 25:759–763
Ito Y, Butler A (2005) Structure of synechobactins, new siderophoresof the marine cyanobacterium Synechococcus sp. PCC 7002.Limnol Oceanogr 50:1918–1923
Iwata K, Inayama T, Kato T (1990) Effects of Spirulina platensis onplasma lipoprotein lipase activity in fructose-induced hyper-lipidemic rats. J Nutr Sci Vitaminol 36:165–171
Jaiswal P, Singh PK, Prasanna R (2008) Cyanobacterial bioactivemolecules—an overview of their toxic properties. Can J Micro-biol 54:701–717
Järvenpää S, Lundberg-Niinistö C, Spoof L, Sjövall O, Tyystjärvi E,Meriluoto J (2007) Effects of microcystins on broccoli andmustard, and analysis of accumulated toxin by liquid chroma-tography–mass spectrometry. Toxicon 49:865–874
Jayaraj R, Anand T, Rao PVL (2006) Activity and gene expressionprofile of certain antioxidant enzymes to microcystin-LR inducedoxidative stress in mice. Toxicology 220:136–146
Jeffries TW, Timourien H, Ward RL (1978) Hydrogen productionby Anabaena cylindrical: effect of varying ammonium andferric ions, pH and light. Appl Environ Microbiol 35:704–710
Jensen GS, Ginsberg DI, Huerta P, Citton M, Drapeau C (2000)Consumption of Aphanizomenon flos aquae has rapid effects onthe circulation and function of immune cells in humans. JANA2:50–58
Jiang Y, Ji B, Wong RNS, Wong MH (2008) Statistical study on theeffects of environmental factors on the growth and microcystinsproduction of bloom-forming cyanobacterium-Microcystis aeru-ginosa. Harmful Algae 7:127–136
Jochimsen EM, Carmichael WW, An J et al (1998) Liver failure anddeath after exposure to microcystins at a haemodialysis center inBrazil. N Engl J Med 338:873–878
Jones AC, Gu L, Sorrels CM, Sherman DH, Gerwick WH (2009) Newtricks from ancient algae: natural products biosynthesis in marinecyanobacteria. Curr Opin Chem Biol 13:216–223
Kaebernick M, Dittmann E, Börner T, Neilan BA (2002) Multiplealternate transcripts direct the biosynthesis of microcystin, acyanobacterial nonribosomal peptide. Appl Environ Microbiol68:449–455
Kannaiyan S, Aruna SJ, Kumari SMP, Hall DO (1997) Immobilizedcyanobacteria as a biofertilizer for rice crops. J Appl Phycol9:167–174
Karl D, Michaels A, Bergman B, Capone D, Carpenter E, Letelier R,Lipschultz F, Paerl H, Sigman D, Stal L (2002) Dinitrogenfixation in the world’s oceans. Biogeochemistry 57(58):47–98
Karube I, Ikemoto H, Kajiwawa K, Tamiya E, Matsuoka H (1986)Photochemical energy conversion using immobilised blue-greenalgae. J Biotechnol 4:73–80
Kato T, Takemoto K, Katayama H, Kuwabara Y (1984) Effects ofSpirulina on dietary hypercholesterolemia in rats. J Jpn Soc NutrFood Sci 37:323–332
Kaya K, Mahakhant A, Keovara L (2002) Spiroidesin, a novellipopeptide from the cyanobacterium Anabaena spiroides thatinhibits cell growth of the cyanobacterium Microcystis aerugi-nosa. J Nat Prod 65:920–921
Kernoff PBA, Willis AL, Stone KJ, Davies JA, McNicol GP (1977)Antithrombotic potential of dihomo γ-linolenic acid in man. BrMed J 2:1441–1444
Kirkpatrick B, Fleming LE, Squicciarini D (2004) Literature review ofFlorida red tide: implications for human health effects. HarmfulAlgae 3:99–115
Klimmek S, Stan HJ, Wilke A, Bunke G, Buchholz R (2001)Comparative analysis of the biosorption of cadmium, lead,nickel, and zinc by algae. Environ Sci Technol 35:4283–4288
Knoll AH (2008) Cyanobacteria and earth history. In: Herrero A,Flores E (eds) The cyanobacteria: molecular biology, genomicsand evolution. Caister Academic Press, Norfolk, pp 1–20
J Appl Phycol (2011) 23:1059–1081 1077

Kodani S, Ishida K, Murakami M (1998) Aeruginosin 103-A, athrombin inhibitor from the cyanobacterium Microcystis viridis. JNat Prod 61:1046–1048
Koksharova OA, Wolk CP (2002) Genetic tool for cyanobacteria.Appl Microbiol Biotechnol 58:123–137
Koptera ZP (1970a) Biosynthesis of thiamine, riboflavin andvitamin B12 by some blue-green algae. Microbiol Z (Kiev)32:429–433
Koptera ZP (1970b) Biosynthesis of biotin, pyredoxin, nicotinic acidand pantothenic acids by some blue-green algae. Microbiol Z(Kiev) 32:555–560
Krakstad C, Herfindal L, Gjertsen BT, Bøe R, Vintermyr OK,Fladmark KE, Døskeland SO (2006) CaM-kinaseII-dependentcommitment to microcystin induced apoptosis is coupled to cellbudding, but not to shrinkage or chromatin hypercondensation.Cell Death Differ 13:1191–1202
Kuritz T, Wolk CP (1994) Use of filamentous cyanobacteria forbiodegradation of organic pollutants. Environ Microbiol 61:234–238
Kushak R, Van Cott E, Drapeau C, Winter H et al (1999) Effect ofalgae Aphanizomenon flos-aquae on digestive enzyme activityand polyunsaturated fatty acids level in blood plasma. Gastro-enterol 116:A559
Kushak RI, Drapeau C, Van Cott EM (2000) Favorable effects ofblue-green algae Aphanizomenon flos-aquae on rat plasma lipids.JANA 2:59–65
Kwan JC, Rocca JR, Abboud KA, Paul VJ, Luesch H (2008) Totalstructure determination of grassypeptolide, a new marine cyano-bacterial cytotoxin. Org Lett 10:789–792
Lankford CE, Walker JR, Reeves JB, Nabbut NH, Byers BR (1966)Inoculum-dependent division lag of Bacillus cultures and itsrelation to an endogenous factor(s) (“Schizokinen”). J Bacteriol91:1070–1079
Larsen LK, Moore RE, Patterson GML (1994) Carbolines from theblue-green alga Dichothrix baueriana. J Nat Prod 57:419–421
Lehr F, Posten C (2009) Closed photo-bioreactors as tools for biofuelproduction. Curr Opin Biotechnol 20:280–285
Li H, Murphy T, Guo J, Parr T, Nalewajko C (2009) Iron-stimulatedgrowth and microcystin production of Microcystis novacekiiUAM 250. Limnologica-Ecol Manage In Water 39:255–259
Lindberg P, Lindblad P, Cournac L (2004) Gas exchange in thefilamentous cyanobacterium Nostoc punctiforme strainATCC29133 and its hydrogenase-defficient mutant strainNHM5. Appl Environ Microbiol 70:2137–2145
Linington RG, González J, Ureña LD, Romero LI, Ortega-Barría E,Gerwick WH (2007) Venturamides A and B: antimalarialconstituents of the Panamanian marine cyanobacterium Oscil-latoria sp. J Nat Prod 70:397–401
Linington RG, Edwards DJ, McPhail SCF, KL MT, Gerwick WH(2008) Symplocamide A, a potent cytotoxin and chymotrypsininhibitor from the marine cyanobacterium Symploca sp. J NatProd 71:22–27
Luesch H, Pangilinan R, Yoshida WY, Moore RE, Paul VJ (2001)Pitipeptolides A and B, new cyclodepsipeptides from the marinecyanobacterium Lyngbya majuscula. J Nat Prod 64:304–307
Luesch H, Yoshida WY, Paul MRE, VJ MSL, Corbett TH (2002)Symplostatin 3, a new dolastatin 10 analogue from the marinecyanobacterium Symploca sp. VP452. J Nat Prod 65:16–20
Lumpkin TA (1987) Collection, maintenance and cultivation ofAzolla. In: Elkan GH (ed) Symbiotic nitrogen fixation technol-ogy. Marcel Dekker Inc., New York, pp 55–94
Lumpkin TA, Plucknett C (1982) Azolla as a green manure: use andmanagement in crop production. Westview Press, Boulder, p 230
Ma LX, Led JJ (2000) Determination by high field NMR spectroscopyof the longitudinal electron relaxation rate in Cu(II) plastocyaninform Anabaena variabilis. Am Chem Soc 122:7823–7824
Madamwar D, Garg N, Shah V (2000) Cynobacterial hydrogenproduction. World J Microbiol Biotechnol 16:8–9
Mackay MA, Norton RS, Borowitzka LJ (1984) Organic osmoregu-latory solutes in cyanobacteria. J Gen Microbiol 130:2177–2191
Masukawa H, Mochimaru M, Sakurai H (2002) Hydrogenases andphotobiological hydrogen production utilizing nitrogenase sys-tem in cyanobacteria. Int J Hydrogen Energy 27:1471–1474
Markov SA (1997) Production of hydrogen from blue-green algae.Russian Patent RU 2083481
Mathew B, Sankaranarayanan R, Nair P, Varghese C, Somanathan T(1995) Evaluation of chemo-prevention of oral cancer withSpirulina fusiformis. Nutr Cancer 24:197–202
McPhail KL, Correa J, Linington RG, Gonzalez J, Ortega-Barría E,Capson TL, Gerwick WH (2007) Antimalarial linear lipopeptidesfrom a Panamanian strain of the marine cyanobacterium Lyngbyamajuscula. J Nat Prod 70:984–988
Mehta SK, Gaur JP (2005) Use of algae for removing heavy metalions from wastewater: progress and prospects. Crit Rev Bio-technol 25:113–152
Metcalf JS, Barakate A, Codd GA (2004) Inhibition of plant proteinsynthesis by the cyanobacterial hepatotoxin, cylindrospermopsin.FEMS Microbiol Lett 235:125–129
Metting B (1981) Systematics and ecology of soil algae. Bot Rev147:195–312
Metting B (1988) Microalgae in agriculture. In: Borowitzka MA,Borowitzka LJ (eds) Micro-algal biotechnology. CambridgeUniversity Press, Cambridge, pp 288–304
Mian MH (1984) A N-tracer study to differentiate nitrogen supplyto flooded rice plants by Azolla and Anabaena during theirearly and later stages of decomposition. Indian J Agric Sci54:733–738
Mishima T, Murata J, Toyoshima M, Fujii K, Nakajima M, Hayashi T,Kato T, Saiki I (1998) Inhibition of tumor invasion andmetastasis by calcium spirulan (Ca-SP), a novel sulfatedpolysaccharide derived from a blue-green alga, Spirulina platen-sis. Clin Exp Metastasis 16:541–550
Mitsui A (1980) Saltwater based biological solar energy conversionfor fuel, chemical, fertiliser, food and feed. In: Proceedings ofBioenergy’ 80, Bioenergy Council, Washington DC. pp. 486–491
Miyaki J (1998) The science of biohydrogen: an energetic view.In: Zaborsky OR (ed) Biohydrogen. Plenum Press, NewYork, pp 7–17
Moreno J, Vargas MA, Madiedo JM, Muñoz J, Rivas J, Guerrero MG(2000) Chemical and rheological properties of an extracellularpolysaccharide produced by the cyanobacterium Anabaena sp.ATCC 33047. Biotechnol Bioeng 67:283–290
Moore AW (1969) Azolla: biology and agronomic significance. BotRev 35:17–34
Morliere P, Maziere JC, Santus R et al (1998) Tolyporphin: a naturalproduct from cyanobacteria with potent photosensitizing activityagainst tumor cells in vitro and in vivo. Cancer Res 58:3571–3578
Mussgnug JH, Thomas-Hall S, Rupprecht J, Foo A, Klassen V,McDowall A, Schenk PM, Kruse O, Hankamer B (2007)Engineering photosynthetic light capture: impacts on improvedsolar energy to biomass conversion. Plant Biotechnol J 5:802–814
Mynderse JS, Moore RE, Kashiwagi M, Norton TD (1977) Anti-leukemia activity in Oscillatoriaceae, isolation of debromoaply-siatoxin from Lyngbya. Science 196:538–540
Nagase H, Inthorn D, Oda A, Nishimura J, Kajiwara Y, Park MO,Hirata K, Miyamoto K (2005) Improvement of selective removalof heavy metals in cyanobacteria by NaOH treatment. J BiosciBioeng 99:372–377
Nagle DG, Paul VJ, Roberts MA (1996) Ypaoamide, a new broadlyacting feeding deterrent from the marine cyanobacterium Lyng-bya majuscule. Tetrahedron Lett 37:6263–6266
1078 J Appl Phycol (2011) 23:1059–1081

Nakaya N, Homa Y, Goto Y (1988) Cholesterol lowering effect ofSpirulina. Nutr Rep Int 37:1329–1337
Newman DJ, Cragg GM (2004a) Marine natural products and relatedcompounds in clinical and advanced preclinical trials. J Nat Prod67:1216–1238
Newman DJ, Cragg GM (2004b) Advanced preclinical and clinicaltrials of natural products and related compounds from marinesources. Curr Med Chem 11:1693–1713
Newman DJ, Cragg GM, Snader KM (2003) Natural products assources of new drugs over the period 1981–2002. J Nat Prod66:1022–1037
Niederholtmeyer H, Wolfstadter BT, Savage DF, Silver PA, WayJC (2010) Engineering cyanobacteria to synthesize andexport hydrophilic products. Appl Environ Microbiol76:3462–3466
Nishiwaki-Matsushima R, Ohta T, Nishiwaki S, Suganuma M,Kohyama K, Ishikawa T, Carmichael WW, Fujiki H (1992)Liver tumor promotion by the cyanobacterial cyclic peptide toxinmicrocystin LR. J Cancer Res Clin Oncol 118:420–424
Nobles DR, Romanovicz DK, Brown RM (2001) Cellulose incyanobacteria: origin of vascular plant cellulose synthase. PlantPhysiol 127:529–542
Olson JM (2006) Photosynthesis in the Archean era. Photosynth Res88:109–117
Ördög V, Stirk WA, Lenobel R, Bancírová M, Strnad M, van Staden J,Szigeti J, Németh L (2004) Screening microalgae for somepotentially useful agricultural and pharmaceutical secondarymetabolites. J Appl Phycol 16:309–314
Ortega-Calvo JJ, Mazuelos C, Hermosin B, Saiz-Jimenez C (1993)Chemical composition of Spirulina and eukaryotic algae foodproducts marketed in Spain. J Appl Phycol 1:425–435
Otero JM, Panagiotou G, Olsson L (2007) Fuelling industrialbiotechnology growth with bioethanol. Adv Biochem EngBiotechnol 108:1–40
Ouellette AJA, Wilhelm SW (2003) Toxic cyanobacteria: the evolvingmolecular toolbox. Front Ecol Environ 1:359–366
Paerl HW, Huisman J (2009) Climate change: a catalyst for globalexpansion of harmful cyanobacterial blooms. Environ MicrobiolRep 1:27–37
Paerl HW, Fulton RS III, Moisander PH, Dyble J (2001) Harmfulfreshwater algal blooms, with an emphasis on cyanobacteria. SciWorld 1:76–113
Panda D, Ananthnarayan V, Larson G, Shi C, Jordan MA, Wilson L(2000) Interaction of the antitumor compound cryptophycin-52with tubulin. Biochemistry 39:14121–14127
Papendorf O, Konig GM, Wright AD (1998) Hierridin B and 2, 4-dimethoxy-6-heptadecyl-phenol, secondary metabolites from thecyanobacterium Phormidium ectocarpi with antiplasmodial ac-tivity. Phytochemistry 49:2383–2386
Park K-H, Kim D-II, Lee C-G (2000) Effect of flashing light onoxygen production rates in high-density algal cultures. J Micro-biol Biotechnol 10:817–822
Pereira S, Zille A, Micheletti E, Moradas-Ferreira P, De Philippis R,Tamagnini P (2009) Complexity of cyanobacterial exopolysac-charides: composition, structures, inducing factors and putativegenes involved in their biosynthesis and assembly. FEMSMicrobiol Rev 33:917–941
Pflugmacher S (2002) Possible allelopathic effects of cyanotoxins,with reference to microcystin-LR, in aquatic ecosystems. EnvironToxicol 17:407–413
Pflugmacher S, Hofmann J, Hübner B (2007) Effects on growth andphysiological parameters in wheat (Triticum aestivum L.) grownin soil and irrigated with cyanobacterial toxin contaminatedwater. Environ Toxicol Chem 26:2710–2716
Piorreck M, Baasch K-H, Pohl P (1984) Biomass production, totalprotein, chlorophylls, lipids and fatty acids of fresh water green
and blue-green algae under different nitrogen regimes. Phyto-chemistry 23:207–216
Plude JL, Parker DL, Schommer OJ, Timmerman RJ, Hagstrom SA,Joers JM, Hnasko R (1991) Chemical characterization ofpolysaccharides from the slime layer of the cyanobacteriumMicrocystis flos-aquae C3-40. Appl Environ Microbiol 57:1696–1700
Pouria S, de Andrade A, Barbosa J et al (1998) Fatal microcystinintoxication in haemodialysis unit in Caruaru Brazil. Lancet352:21–26
Prabaharan D, Subramanian G (1996) Oxygen-free hydrogen produc-tion by the marine cyanobacterium Phormidium valderianumBDU 20041. Bioresour Technol 57:111–116
Prabaharan D, Sumathi M, Subramanian G (1994) Ability to useampicillin as a nitrogen source by the marine cyanobacteriumPhormidium valderanum BDU 30501. Curr Microbiol 28:314–320
Prasanna R, Nain L, Tripathi R, Gupta V, Chaudhary V, Middha S,Joshi M, Ancha R, Kaushik BD (2008) Evaluation offungicidal activity of extracellular filterates of cyanobacteria—possible role of hydrolytic enzymes. J Basic Microbiol 48:186–194
Prepas EE, Kotak BG, Campbell LM, Evans JC, Hrudey SE, HolmesCFB (1997) Accumulation and elimination of cyanobacterialhepatotoxins by the fresh water clam Anodonta simpsonina. CanJ Fish Aquat Sci 54:41–46
Prince EK, Myers TL, Naar J, Kubanek J (2008) Competingphytoplankton undermines allelopathy of a bloom-formingdinoflagellate. Proc R Soc Lond B 275:2733–2741
Pruder G (1983) Biological control of gas exchange in intensiveaquatic production system. IEEE 15:1002–1004
Pulz O, Gross W (2004) Valuable products from biotechnology ofMicroalgae. Appl Microbiol Biotechnol 65:635–648
Rai V, Rai AK (1999) Growth behaviour of Azolla pinnata at varioussalinity levels and induction of high salt tolerance. Plant Soil206:79–84
Rai AK, Sharma NK (2006) Phosphate metabolism in the cyanobacte-rium Anabaena doliolum under salt stress. Curr Microbiol 52:6–12
Rastogi RP, Sinha RP (2009) Biotechnological and industrialsignificance of cyanobacterial secondary metabolites. BiotechnolAdv 27:521–539
Ray S, Bagachi SN (2001) Nutrients and pH regulate algicideaccumulation in cultures of the cyanobacterium Oscillatorialaetevirens. New Phytol 149:455–460
Reddy CM, Bhat VB, Kiranmai G, Reddy MN, Reddanna P,Madyastha K (2000) Selective inhibition of cyclooxygenase-2by C-phycocyanin, a biliprotein from Spirulina platensis. Bio-chem Biophys Res Commun 3:599–603
Reshef V, Mizrachi E, Maretzki T, Silberstein C, Loya S, Hizi A,Carmeli S (1997) New acylated sulfoglycolipids and digalacto-lipids and related known glycolipids from cyanobacteria with apotential to inhibit to reverse transcriptase of HIV-1. J Nat Prod60:1251–1260
Richmond A (1988) Spirulina. In: Borowitzka MA, Borowitzka LJ(eds) Microlgal biotechnology. Cambridge University Press,Cambridge, pp 85–121
Rickards RW, Rothschild JM, Willis AC et al (1999)Calothrixins-A and -B, novel pentacyclic metabolites fromCalothrix sp. cyanobacteria with potent activity against malariaparasites and human cancer cells. Tetrahedron 55:13513–13520
Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY (1979)Generic assignments, strain histories and properties of purecultures of cyanobacteria. J Gen Microbiol 111:1–61
Robbins WJ, Hervey A, Stebbins M (1951) Further observations onEuglena and B12. Bull Torrey Bot Club 86:367–373
J Appl Phycol (2011) 23:1059–1081 1079

Rodŕiguez AA, Stella AM, Storni MM, Zulpa G, Zaccaro MC(2006) Effects of cyanobacterial extracellular products andgibberellic acid on salinity tolerance in Oryza sativa L. SalineSyst 2:7
Roger PA, Kulasooriya SA (1980) Blue-green algae and rice. IRRI,Los Baños, p 112
Roger PA, Watanabe I (1984) Algae and aquatic weeds as a source oforganic matter and plant nutrition for wetland rice. In: Organicmatter and rice. IRRI, Los Baños, pp 147–168
Rohrlack T, Dittmann E, Borner T, Christoffersen K (2001) Effects ofcell-bound microcystins on survival and feeding of Daphnia spp.Appl Environ Microbiol 67:3523–3529
Rohrlack T, Christoffersen K, Hansen PE et al (2003) Isolation,characterization, and quantitative analysis of Microviridin J, anew Microcystis metabolite toxic to Daphnia. J Chem Ecol29:1757–1770
Rosenberg JN, Oyler GA, Wikinson L, Betenbaugh MJ (2008) Agreen light for engineered algae: redirecting metabolism tofuel a biotechnology revolution. Curr Opin Biotechnol19:430–436
Saito K, Kondo A, Ishii H, Saito H, Nishida F, Abe T, Chen C (2001)Nodularin-Har: a new nodularin from Nodularia. J Nat Prod64:139–141
Salazar M, Martinez E, Madrigal E, Ruiz LE, Chamorro GA (1998)Subchronic toxicity study in mice fed Spirulina maxima. JEthnopharmacol 62:235–241
Sangthongpitag K, Delaney SF, Rogers PL (1996) Evaluation of fourfresh-water unicellular cyanobacteria as potential hosts formosquitocidal toxins. Biotechnol Lett 18:175–180
Santillan C (1982) Mass production of Spirulina. Experientia 38:40–43
Saqrane S, Ghazali IE, Oudra B et al (2008) Effects ofcyanobacteria producing microcystins on seed germinationand seedling growth of several agricultural plants. J EnvironSci Health B 43:443–451
Schopf JW (2000) The fossil records: tracing the roots of thecyanobacterial lineage. In: Whitton BA, Potts M (eds) Theecology of cyanobacteria. Kluwer, Dordrecht, pp 13–35
Schwartz RE, Hirsch CF, Sesin DF et al (1990) Pharmaceuticals fromcultured algae. J Ind Microbiol 5:113–124
Sergeeva E, Liaimer A, Bergman B (2002) Evidence for production ofthe phytohormone indole-3-acetic acid by cyanobacteria. Planta215:229–238
Shapiro J (1972) Blue-green algae: why they become dominant?Science 179:382–384
Shashirekha S, Uma L, Subramanian G (1997) Phenol degradation bythe marine cyanobacterium Phormidium valderianum BDU30501. J Ind Microbiol Biotechnol 19:130–133
Sheehan JJ (2009) Biofuels and the conundrum of sustainability. CurrOpin Biotechnol 20:318–324
Sielaff H, Christiansen G, Schwecke T (2008) Natural products fromcyanobacteria: exploiting a new source for drug discovery. IDrug9:119–127
Simmons TL, Andrianasolo E, McPhail K, Flatt P, Gerwick WH(2005) Marine natural products as anticancer drugs. Mol CancerTher 4:333–342
Singh RN (1950) Reclamation of usar soils in India through blue-green algae. Nature 165:325–326
Singh RN (1961) Role of blue-green algae in nitrogen economy ofIndian agriculture. ICAR, New Delhi
Singh PK (1977) Multiplication and utilization of fern Azollacontaining nitrogen fixing algal symbiont as green manure inrice cultivation. RISO 26:125–136
Singh PK (1988) Biofertilization of rice crop. In: Sen SP, Palit P (eds)Biofertilizers: potentialities and problems. Plant PhysiologyForum, Calcutta, pp 109–114
Singh IP, Milligan KE, Gerwick WH (1999) Tanikolide, a toxic andantifungal lactone from the marine cyanobacterium Lyngbyamajuscula. J Nat Prod 62:1333–1335
Singh S, Kate BN, Banerjee UC (2005) Bioactive compounds fromcyanobacteria and microalgae: an overview. Crit Rev Biotechnol25:73–95
Sirenko LA, Kirpenko YA, Kirpenko NI (1999) influence ofmetabolites of certain algae on human and animal cell cultures.Int J Algae 1:122–126
Sivonen K, Jones G (1999) Cyanobacterial toxins. In: Chorus I,Bartram J (eds) Toxic cyanobacteria in water: a guide to theirpublic health consequences monitoring and management. E andFN Spon, London, pp 41–111
Skulberg OM (2000) Microalgae as a source of bioactive molecules—experience from cyanophyte research. J Appl Phycol 12:341–348
Skulberg OM, Carmichael WW, Codd GA, Skulberg R (1993)Taxonomy of toxic cyanophyceae (cyanobacteria). In: FalconerIR (ed) Algal toxins in seafood and drinking water. Academic,London, pp 145–164
Solomon BD (2010) Biofuels and sustainability. Ann NY Acad Sci1185:119–134
Song D, Fu J, Shi D (2008) Exploitation of oil-bearing microalgae forbiodiesel. Chin J Biotechnol 24:341–348
Srivastava VC, Manderson GJ, Bhamidimarri R (1999) Inhibitorymetabolites production by the cyanobacterium Fischerella mus-cicola. Microbiol Res 153:309–317
Stewart I, Schluter PJ, Shaw GR (2006) Cyanobacterial lipopolysac-charides and human health—a review. Environ Health GlobAcess Sci Source 5:7
Switzer L (1981) Spirulina the whole food revolution. ProteusCorporation Bantam Books, Toronto
Talley SN, Talley BJ, Rains DW (1977) Nitrogen fixation by Azolla inrice fields. In: Lyons JM, Valentine RC, Phillips DA, Rains DW,Huffaker RC (eds) Genetic engineering for nitrogen-fixation.Plenum Press, New York, pp 363–384
Tamagnini P, Leitao E, Oliveira P, Ferreira D, Pinto F, Harris DJ,Heidorn T, Lindblad P (2007) Cyanobacterial hydrogenases:diversity, regulation and applications. FEMS Microbiol Rev31:692–720
Tan LT (2007) Bioactive natural products from marine cyanobacteriafor drug discovery. Phytochemistry 68:954–979
Tan LT (2010) Filamentous tropical marine cyanobacteria: a richsource of natural products for anticancer drug discovery. J ApplPhycol. doi:10.1007/s10811-010-9506-x
Tang PS, Ding-Ji S, Changzheng H (1981) Regulation of energymetabolism (photosynthesis and nitrogen fixation) in blue-greenalgae. In: Proceeding of Joint China-US Phycology Symposium.p. 339
Taori K, Matthew S, Rocca JR, Paul VJ, Luesch H (2007)Lyngbyastatins 5–7, potent elastase inhibitors from Floridianmarine cyanobacteria, Lyngbya spp. J Nat Prod 70:1593–1600
Teixeira M, Da GLC, Da CN CM, De Carvalho VLP, Dos S, Hage E(1993) Gastroenteritis epidemic in the area of the Itaparica Dam,Bahia, Brazil. Bull Pan Am Health Organ 27:244–253
Thajuddin N, Subramanian G (2005) Cyanobacterial biodiversity andpotential applications in biotechnology. Curr Sci 89:47–57
Tillett D, Dittmann E, Erhard M, von Döhren H, Börner T, Neilan BA(2000) Structural organization of microcystin biosynthesis inMicrocystis aeruginosa PCC7806: an integrated peptide-polyketide synthetase system. Chem Biol 7:753–764
Tilman D, Socolow R, Foley JA et al (2009) Beneficial biofuels—thefood, energy, and environment trilemma. Science 325:270–271
Todorova AK, Juettner F, Linden A, Plüss T, von Philipsborn W(1995) Nostocyclamide: a new macrocyclic, thiazole-containingallelochemical from Nostoc sp. 31 (cyanobacteria). J Org Chem60:7891–7895
1080 J Appl Phycol (2011) 23:1059–1081

Tomitani A, Knoll AH, Cavanaugh CM, Ohno T (2006) Theevolutionary diversification of cyanobacteria: molecular-phylogenetic and paleontological perspectives. Proc Nat AcadSci USA 103:5442–5447
Tonk L, Welker M, Huisman J, Visser PM (2009) Production ofcyanopeptolins, anabaenopeptins, and microcystins by the harm-ful cyanobacteria Anabaena 90 and Microcystis PCC 7806.Harmful Algae 8:219–224
Torres-Duran PV, Miranda-Zamora R, Paredes-Carbajal MC, MascherD, Blé-Castillo J, Díaz-Zagoya JC, Juárez-Oropeza MA (1999)Studies on the preventive effect of Spirulina maxima on fattyliver development induced by carbon tetrachloride, in the rat. JEthnopharmacol 64:141–147
Trimurtulu G, Ogino J, Helzel CE et al (1995) Structure determinationconfirmational analysis chemical stability studies and anti tumorevaluation of the cryptophycins: Isolation of 18 new analoguesfrom Nostoc sp. strain GSV 224. J Am Chem Soc 117:12030–12049
Tsuji K, Asakawa M, Anzai Y, Sumino T, Harada K et al (2006)Degradation of microcystins using immobilized microorganismisolated in a eutrophic lake. Chemosphere 65:117–124
Valencia A, Walker J (1999) A multi-axial treatment paradigm formild traumatic brain injury to achieve reparative functional meta-plasticity. In: 3rd World Congress on Brain Injury, IBIA, QuebecCity
Van Hille RP, Boshoff GA, Rose PD, Duncan JR (1999) A continuousprocess for the biological treatment of heavy metal contaminatedacid mine water. Resour Conserv Recycl 27:157–167
Van Wagoner RM, Drummond AK, Wright JLC (2007) Biogeneticdiversity of cyanobacterial metabolites. Adv Appl Microbiol61:89–217
Venkataraman GS (1972) Algal biofertilizer and rice cultivation.Today and tomorrow’s Printers and Publishers, New Delhi
Venkataraman GS (1981) Algal biofertilizer for rice. IARI, New DelhiVerdier-Pinard P, Sitachitta N, Rossi JV, Sackett DL, Gerwick WH,
Hamel E (1999) Biosynthesis of radiolabeled curacin A and itsrapid and apparently irreversible binding to the colchicine site oftubulin. Arch Biochem Biophys 370:51–58
Véize C, Rapala J, Vaitomaa J, Sivonen K (2002) Effect of nitrogenand phosphorus on growth of toxic and nontoxic Microcystisstrains and on intracellular microcystin concentrations. MicrobEcol 43:443–454
Vlad M, Bordas E, Caseanu E, Uza G, Creteanu E, Polinicenco C(1995) Effect of cuprofilin on experimental atherosclerosis. BiolTrace Elem Res 48:99–109
Volk RB (2005) Screening of microalgal culture media for thepresence of algicidal compounds and isolation and identification
of two bioactive metabolites, excreted by the cyanobacteriaNostoc insulare and Nodularia harveyana. J Appl Phycol17:339–347
Volk RB (2006) Antialgal activity of several cyanobacterial exome-tabolites. J Appl Phycol 18:145–151
Vonshak A (1996) Use of Spirulina biomass. In: Vonshak A (ed)Spirulina platensis (Arthropsira). Taylor and Francis, London,pp 205–212
Walker DA (2009) Biofuels, facts, fantasy, and feasibility. J ApplPhycol 21:509–517
Wang W, Liu Y, Li D, Hu C, Rao B (2009) Feasibility ofcyanobacterial inoculation for biological soil crusts formation indesert area. Soil Biol Biochem 41:926–929
Wase NV, Wright PC (2008) System biology of cyanobacterialsecondary metabolite production and its role in drug discovery.Expert Opin Drug Discov 3:903–929
Watanabe I (1962) Effect of nitrogen-fixing blue-green alga Tolypthrixtenuis on the nitrogenous fertility of paddy soil and on the cropyield of rice plants. J Gen Appl Microbiol 8:85–91
Watanabe I (1977) Azolla utilization in rice culture. Int Rice ResNewsl 2:3–8
Watanabe I (1987) Summary reports of the Azolla programme on theinternational network on soil fertility and fertilizer evaluation forrice. IRRI, Los Baños, pp 197–205
Watanabe I, Lee KK, Alimagno BV, Sato M, Del Rosario DC, DeGuzman M (1977) Biological N2 fixation in paddy field studiesby in situ acetylene reduction assays. IRRI Res Pap Ser 3:1–16
White JD, Kim T-S, Nambu M (1997) Absolute configuration and totalsynthesis of (+)-curacin A, an antiproliferative agent from thecyanobacterium Lyngbya majuscula. J Am Chem Soc 119:103–111
Whitton BA, Potts M (eds) (2000) The ecology of cyanobacteria.Kluwer, Dordrecht. pp. 669
Wolk CP (1973) Physiology and cytological chemistry of blue-greenalgae. Bacteriol Rev 37:32–101
Wu M, Okino T, Nogle LM, Marquez BL, Williamson RT, SitachittaN, Berman FW, Murray TF, McGough K, Jacobs R, Colsen K,Asano T, Yokokawa F, Shioiri T, Gerwick WH (2000) Structure,synthesis, and biological properties of kalkitoxin, a novelneurotoxin from the marine cyanobacterium Lyngbya majuscula.J Am Chem Soc 122:12041–12042
Yee N, Benning LG, Phoenix VR, Ferris FG (2004) Characterizationof metal-cyanobacteria sorption reaction: a combined macroscop-ic and infrared spectroscopic investigation. Environ Sci Technol38:775–782
Zurawell RW, Chen H, Burke JM, Prepas EE (2005) Hepatotoxiccyanobacteria: a review of the biological importance of microcystinsin fresh water environments. J Toxicol Environ Health 8:1–37
J Appl Phycol (2011) 23:1059–1081 1081
Related Documents











