DEVELOPMENTAL BIOLOGY 198, 266-276 [1998} ARTICLE NO. DB988910 Survival of patchwork Melanoblasts Is Dependent upon Their Number in the Hair Follicle at the End of Embryogenesis Genevieve Aubin-Houzelstein, Florence Bernex, Colette Elbaz, and Jean-Jacques Panthier URA-INRA de G~n~tique Mol~culaire, Ecole Nationale V~t~rinaire d'Alfort, 7 avenue du G~n~ral-de-Gau]le, 94704 Maisons-Alfort Cedex, France The recessive patchwork (pwk} mutation in mice is associated with a unique hair follicle phenotype. Mice homozygous for patchwork exhibit a variegated coat containing a mixture of white and fully pigmented hairs, but no partially pigmented hairs. We have investigated the etiology of this mutation. We report here that the white hairs result from the lack of melanocytes in the follicle. As indicated by the coat color pattern of patchwork <-~ albino chimeras, the target cell for the patchwork mutation is the melanocyte and/or its precursor. Examination of these chimeras also suggested that patchwork does not act in a cell-autonomous manner. The colonization of the skin by melanoblasts in patchwork embryos was studied using a lacZ transgene. Melanoblasts die by apoptosis in hair follicles from homozygous pwk/pwk fetuses starting at embryonic day 18.5, indicating that patchwork acts from this stage. The combination of pwk and Kit w'ei, a mutation responsible for a reduced number of melanoblasts in the hair follicle, suggested that pwk gene product is necessary for low numbers of melanoblasts to survive and differentiate in the hair follicle from embryonic day 18.5 onward. We conclude that the pigmented hairs on the coat of pwk/pwk mice may be attributed to a community effect among melanoblasts in the hair follicle at the end of embryogenesis. © 1998 AcademicPress Key Words: mouse; variegation; coat color; melanocyte; apoptosis; community effect. INTRODUCTION Melanocytes are specialized cells responsible for the pigmentation of the skin, the hair, and the eyes of mammals. They are characterized by their dendritic shape and the production of the melanin pigments, which are deposited in modified lysosomes, the melanosomes (Hirobe, 1995; Orlow, 1995). There are two kinds of melanins, the brown-black eumelanins and the yellow- red phaeomelanins. The mixing of eu- and phaeomela- nins in different proportions gives rise to the various coat colors known in mice. Melanocyte precursors arise from neural crest cells, which leave the apical ridges of the neural tube from embryonic day 8.5 (E8.5) onward in the mouse (Rawles, 1947; Serbedzija et al., 1990, 1994). These cells then migrate within the mesenchymal layers, between the dermamyotome and the ectoderm, following a dorsolateral pathway (Serbedzija et al., 1990, 1994; Steel et al., 1992). During their migration, they proliferate actively and colonize the entire mesenchyme of the body. They colonize the epiderm from E12.5 and the developing hair follicles from E14.5 (Mayer, 1973; Kunisada et al., 1996). Hair follicles are skin appendages with a mixed origin: the dermal papilla is of dermal origin, while the hair matrix, which is the germinal part of the hair bulb responsible for the formation of the hair shaft and of the internal epithelial root sheath, derives from the epider- mis (Hardy, 1992). At E16.5, the hair folhcle melano- blasts begin to differentiate into melanocytes (Yoshida et al., 1996). After their migration, the melanocytes are localized in the innermost layer of the matrix. They produce melanin granules which are transported to the surrounding keratinocytes responsible for the formation of the pigmented hair shaft. Melanogenesls takes place exclusively during the anagen, the growing phase of the hair cycle (Chase, 1954). In the mouse, 87 loci which affect the coat color have been identified (Mouse Genome Database,. http://www. informatics.jax.org/, 1998). A number of mutations affect the precursors of the pigment cells, altering their migration, 266 0012-1606/98 $25 00 Copyright ~ 1998 by Academtc Press All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DEVELOPMENTAL BIOLOGY 198, 266-276 [1998} ARTICLE NO. DB988910
Survival of patchwork Melanoblasts Is Dependent upon Their Number in the Hair Follicle at the End of Embryogenesis
Genevieve Aubin-Houzelstein, Florence Bernex, Colette Elbaz, and Jean-Jacques Panthier URA-INRA de G~n~tique Mol~culaire, Ecole Nationale V~t~rinaire d'Alfort, 7 avenue du G~n~ral-de-Gau]le, 94704 Maisons-Alfort Cedex, France
The recessive patchwork (pwk} mutation in mice is associated with a unique hair follicle phenotype. Mice homozygous for patchwork exhibit a variegated coat containing a mixture of white and fully pigmented hairs, but no partially pigmented hairs. We have investigated the etiology of this mutation. We report here that the white hairs result from the lack of melanocytes in the follicle. As indicated by the coat color pattern of patchwork <-~ albino chimeras, the target cell for the patchwork mutation is the melanocyte and/or its precursor. Examination of these chimeras also suggested that patchwork does not act in a cell-autonomous manner. The colonization of the skin by melanoblasts in patchwork embryos was studied using a lacZ transgene. Melanoblasts die by apoptosis in hair follicles from homozygous pwk/pwk fetuses starting at embryonic day 18.5, indicating that patchwork acts from this stage. The combination of pwk and Kit w'ei, a mutation responsible for a reduced number of melanoblasts in the hair follicle, suggested that pwk gene product is necessary for low numbers of melanoblasts to survive and differentiate in the hair follicle from embryonic day 18.5 onward. We conclude that the pigmented hairs on the coat of pwk/pwk mice may be attributed to a community effect among melanoblasts in the hair follicle at the end of embryogenesis. © 1998 Academic Press
Key Words: mouse; variegation; coat color; melanocyte; apoptosis; community effect.
INTRODUCTION
Melanocytes are specialized cells responsible for the pigmentation of the skin, the hair, and the eyes of mammals. They are characterized by their dendritic shape and the production of the melanin pigments, which are deposited in modified lysosomes, the melanosomes (Hirobe, 1995; Orlow, 1995). There are two kinds of melanins, the brown-black eumelanins and the yellow- red phaeomelanins. The mixing of eu- and phaeomela- nins in different proportions gives rise to the various coat colors known in mice. Melanocyte precursors arise from neural crest cells, which leave the apical ridges of the neural tube from embryonic day 8.5 (E8.5) onward in the mouse (Rawles, 1947; Serbedzija et al., 1990, 1994). These cells then migrate within the mesenchymal layers, between the dermamyotome and the ectoderm, following a dorsolateral pathway (Serbedzija et al., 1990, 1994; Steel et al., 1992). During their migration, they proliferate actively and colonize the entire mesenchyme of the body.
They colonize the epiderm from E12.5 and the developing hair follicles from E14.5 (Mayer, 1973; Kunisada et al., 1996). Hair follicles are skin appendages with a mixed origin: the dermal papilla is of dermal origin, while the hair matrix, which is the germinal part of the hair bulb responsible for the formation of the hair shaft and of the internal epithelial root sheath, derives from the epider- mis (Hardy, 1992). At E16.5, the hair folhcle melano- blasts begin to differentiate into melanocytes (Yoshida et al., 1996). After their migration, the melanocytes are localized in the innermost layer of the matrix. They produce melanin granules which are transported to the surrounding keratinocytes responsible for the formation of the pigmented hair shaft. Melanogenesls takes place exclusively during the anagen, the growing phase of the hair cycle (Chase, 1954).
In the mouse, 87 loci which affect the coat color have been identified (Mouse Genome Database,. http://www. informatics.jax.org/, 1998). A number of mutations affect the precursors of the pigment cells, altering their migration,
266
0012-1606/98 $25 00 Copyright ~ 1998 by Academtc Press
All rights of reproduction in any form reserved.

Coat Color Variegation in Mice 267
proliferation, and/or survival during development , e.g., Splotch (Pax3Spl, piebald (EdnrbS), d o m i n a n t spot t ing (KitW), and Steel (Mg[ sl) {Silvers, 1979; Jackson, 1991, 1994; Jackson et a1., 1994; Barsh, 1996). The m u t a n t s are charac- terized by wh i t e spot t ing wi th hair follicles lacking mela- nocytes on an o therwise colored coat. The p igmented areas resul t from an in te rming l ing of more or less p igmented hairs, depending on the n u m b e r of melanocytes in the hai r bulbs. Hence, these m u t a t i o n s often combine a pecul iar p igment pa t t e rn w i th a variegated phenotype.
Othe r m u t a t i o n s exert an effect on me lanocy te t e rmina l differentiat ion. They affect various aspects of the biology of the melanocyte : i ts morphology, the melanocyte-specif ic gene expression machinery , the s t ruc ture and func t ion of the melanosome, or the melanogenic enzymes (Silvers, 1979; Jackson, 1991, 1994; Jackson et al., 1994; Barsh, 19961. These m u t a t i o n s are exemplif ied by di lute (Myo5aa), mi- c roph tha lmia (Mitre'), pink-eyed d i lu t ion (p), and albino (TyrO), respectively. In general, the corresponding pheno- types are not variegated.
In this report, we s tudy the etiology of a spontaneous muta t ion , pa tchwork {pwk), which causes coat color mot- t l ing in mice. Animals homozygous for patchwork are variegated wi th whi t e or fully p igmented hairs, bu t no part ial ly p igmented (gray) hairs (K. Moore and V. Pochman, unpub l i shed results), patchwork has been localized to mouse c h r o m o s o m e 10, in a region tha t lacks any mouse m u t a t i o n w i th a s imi lar phenotype (K. Moore and V. Pochman, unpubl i shed results). If each hair follicle was derived from a single progeni tor cell, one possible explana- t ion for this pheno type could be tha t the m u t a t i o n has an incomple te pene t rance and/or expresslvity. However, all componen t s of a single hair are derived from more than one precursor. This holds true for the melanocytes and for the o ther hai r follicle cells, as demons t ra ted us ing aggregation ch imeras (Mintz, 1967; reviewed m Silvers, 1979). To solve this apparent contradict ion, we have followed the develop- m e n t of melanoblas t s in pa tchwork embryos and adults. We show here tha t the i n w a b d l t y of melanoblas t s in the hair follicles of fetuses by E18.5 is responsible for the whi t e hairs on the coat of pa tchwork mice. The occasional sur- vival of melanoblas t s resul ts in the p igmented hairs. It may occur wheneve r pa tchwork melanoblas t s are above a threshold n u m b e r in the hair follicle. The hypothes is tha t pa tchwork melanoblas t s are subject to a c o m m u n i t y effect is discussed.
M A T E R I A L S A N D M E T H O D S
Mice
The patchwork mutation arose spontaneously in an outbred stock of unknown background. Brother-sister matmgs were per- formed until inbreeding was achieved (F? +24}. Albino BALB/cl and AKR/I mice were obtained from the INRA and Charles River, respectively. (C57BL/6 x CBA/J)F1 mice were obtained from the
INRA. (129/Sv x C57BL/61 hybrid KitWh'~Z/+ mice were produced in our laboratory {Bernex et al.. 1996}.
Whole.Mount Preparations of Hairs
Hairs were plucked, placed in 70% ethanol, and degassed over- night. They were aligned in Mowlol on histological slides and observed using stereoscopic microscopy.
Genotype Analys is of Embryos
Genomic DNA was extracted from the yolk sac. The polymor- phic microsatellite sequence DlOMitl 17 was amplified by PCR as described (Dietrich et al. 1994}, using the primers DlOMIt117f, ACTTCCACACATGAGTCATAGCA, and DIOMItl 17r, CCAGT- TGTCTTTCTTGGTfq-IG. Amplification conditions were 94°C (30 s), 55°C 130 sl, and 72°C 140 s) for 35 cycles.
I3-Galactosidase ([J-Gal) Ac t i v i t y Detect ion
/3-Gal acnvity was revealed on 10-day-old skin cryostat sec- nons using 5-bromo-4-chloro-3-mdolyl /3-D-galactoslde {X-Gal) as a substrate as previously described {Bernex et al., 1996}. Whole-mount /3-Gal detecnon was performed on fixed embryos from E11.5 to bzrth. For embryonic ages E17.5 to E19.5, fetuses were incubated in distilled water at 58°C for 1 ram; then the epidermis was removed and the fetuses were fixed and stained as described (Bernex et al, 1996}.
Ant i .S lO0 Immunoh i s tochemis t ry
Skin samples from 10-day-old mice were fixed for 7 days In 10% formaldehyde in phosphate-buffered saline (PBS}, embedded m paraffin, and sectioned at 5 ~tm. Sections were deparaffined in toluene, rehydrated, and treated as follows at room temperature with PBS washes after each step. The secnons were incubated for 20 mm m a 5% normal goat serum solution (Sigma}. After treat- ment with the avidin/blotln blocking kit from Vector Laboratories, they were incubated for 90 min with a 1:100 dilution of an Ig fraction of rabbit ann-cow S100 antiserum {Dakopatts) m PBS containing 1% bovine serum albumin (BSA) and 1% normal goat serum. They were then incubated for 1 h with a biotin-conjugated, goat F(ab'12 ann-rabbit IgG antiserum (Caltag; 2:1000 dilution in PBS with 1% BSA and 1% normal goat serum}. After the endoge- nous peroxadase activity was blocked with 0.3 % hydrogen peroxade m methanol for 20 mm, the sections were incubated for 1 h with an extravidine-peroxidase conjugate (Sigma; 1:50 dilution in PBS) and for 3 mm with aminoethyl carbazole {AEC) red (Zymed). Slides were counterstamed with Mayer's hematoxyhn (Immunotechl and mounted with Mowiol. Sections were viewed using bright-field microscopy.
Anti-13-Galactosidase fmmunofluorescence and Detect ion o[ Apop to t i c Cells
Fetuses ranging from E17.5 to birth were recovered, fixed, frozen, and cryostat-sectioned as previously described (Bemex et at., 1996}. The sections were fixed for 10 min in acetone and conserved at -80°C. They were postfixed in 4% paraformaldehyde for 10 to 20 min, depending on the developmental stage, and treated as follows with two 10-min PBS washes after each step. They were mcubated
Copyright © 1998 by Academic Press All nghts of reproduction m any form reserved.

268 Aubin-Houzelstein et al.
overnight at 4"C in a 2:1000 dilution of rabbit IgG fraction to /3-~alactosidase antiserum (Cappel) in PBS with 1% normal goat serum. After permeabilization for 2 rain at 4"C in 0.1% Triton X-100 and 0.14% trisodium citrate dihydrate, they were incubated for 1 h at 37°C in a TUNEL reaction mixture (In Situ Cell Death Detection Kit, fluorescein; Boehringer Mannheim). After incuba- tion $or I h at room temperature in a tetramethyl rhodamine isothiocyanate [TR1TC}conjugated goat anti-rabbit IgG 11:200 dilu- tion; Jackson Immuno Research), the sections were mounted in Mowiol and viewed using immunofluorescence microscopy {Zeiss; Axioskop).
At least three embryos of each genotype were examined for each time point.
% r , ' .
Aggregation Chimeras
The method described by Bowman and McLaren (1970) was used with minor modifications. Four- to eight-cell embryos were harvested at E2.5. The zona pellucida was removed with acidi- fied Tyrode's solution (pH 2.5) [Nicolson et al., 1975). The embryos were rinsed and incubated as pairs with stage-matched embryos in Whitten's medium at 37°C in 5% CO 2 in a humidi- fied atmosphere for 24 h. The expanded blastocysts were trans- ferred to the uterus of E2.5 pseudopregnant (C57BL/6J x CBA/ JIF1 female mice. Chimerism was monitored by coat color and progeny testing.
RESULTS
There Are No Melanocytes in p w k / p w k White Hair Follicles
The coat of patchwork mice is a juxtaposition of two kinds of hairs, white hairs (which together make up about 40% of the total number of hairs) and fully pigmented hairs, resulting in a salt-and-pepper phenotype {Fig. 1). This phenotype is similar on agouti (A/A) and nonagouti {a/a} backgrounds, indicating that the mutation acts regardless of the nature of the pigment synthesized in the melanocytes. We examined whole-mount preparations of pwk/pwk hairs. In the fully pigmented hairs, all medul- lary cells contained melanin granules in equivalent pro- portions, as occurred in the wild-type hairs. In the white hairs, there was no detectable pigment deposited in the medullary cells, which appeared entirely white (data not shown). The absence of pigmentation may be due to the complete lack of melanocytes in the hair follicle, as observed for several mutations that affect melanocyte survival. Alternatively, this phenotype may result from a defect in an enzyme involved in the synthesis of melanin. To determine whether pwk affects melano- cyte survival or its function, we took advantage of the Kit waltz allele (Bernex et al., 1996). This allele was created by replacing the first exon of Kit with the Escherichia coli IacZ gene using gene targeting technol- ogy. The lacZ reporter gene allows one to follow the distribution of Kit-expressing ceils, including melano- cytes and their precursors. KitWao~z[+ mice are normally pigmented, except that they have white feet and a white
FIG. 1. patchwork (pwk]pwk) homozygote. Note the salt-and- pepper phenotype due to the mixing of white and black hairs. There is no obvious patterning, even on the coat of younger individuals. Furthermore, there is no spatial difference: the percentage of white and black hairs is the same in head, dorsal, abdominal, and tail r e g i o n s .
tail tip. We produced mice carrying the Kit T M allele in combination with the patchwork [pwkl, albino [TyrOl, and viable dominant spotting [Kit wv} mutations, respectively (Table 11. pwk/pwk KitW~a~z/+ mice do not differ phe- notypically from pwk/pwk mice, except for their white feet and tail tip. As expected, Tyr~/Tyr c KitWaocz/+ mice are white, due to the Cys85 --~ Ser amino acid substitu- tion in the tyrosinase which destroys its enzymatic activity (Yokoyama et al., 1990). Finally, KitWV/KitWa~z double heterozygotes are black-eyed white, since their hair follicles lack melanocytes (Mayer, 1970). We exam- ined the/3-Gal labeling obtained following a histochemi- cal staining with X-Gal in hair follicles of pwk/pwk Ki tW~z/+, 73zrC/Tyr ~ KitW4acz/+, and KitWV/Kit T M
10-day-old mice. In pwk/pwk KitW-J~z/+ pigmented hair follicles (Fig. 2A), as well as in Tyr~/Tyr ~ Kitwl~z/+ follicles (Fig. 2C), /3-Gal-positive cells were found at the top of the hair bulb, in the most internal layer of the matrix, in contact with the dermal papilla. These cells were also detected with an anti-S100 antibody (Figs. 2E and 2G), which stains melanocytes (Smoller et al., 1991; Ito et al., 1993). By contrast, these 13-Gal-, S100- positive cells were found neither in pwk/pwk KitW~o~z/+ white hair follicles (Figs. 2B and 2F) nor in KitW~/Kit T M
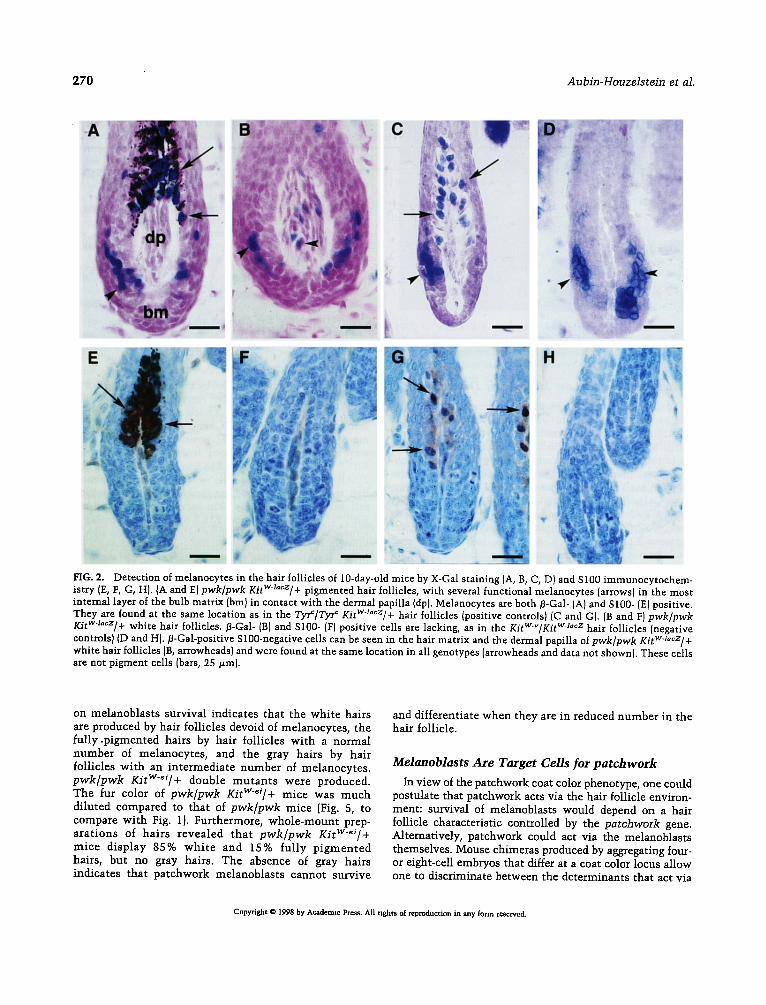
follicles (Figs. 2D and 2H). /3-Gal-positive, S100-negative cells were found in more external layers of the matrix and in the dermal papilla of pwk/pwk KitWl~z/+ mice (Figs. 2A, 2B, 2E, and 2F), as well as in all controls (Figs. 2C, 2D, 2G, and 2H and data not shown); thus, they are not pigment cells. We conclude from theso experiments that the white hair follicles of pwk/pwk mice lack melanocytes.
Copyright O 1998 by Academic Press. All nghcs of reproduction in any form reserved.

Coat Color Variegation in Mice 269
TABLE 1 Mutant Phenotypes and Molecular Defects of the Mice Used in This Study
Coat color phenotype Mutation .Reference
Kit T M K1tW'~¢z/+ heterozygotes have white feet and Insertion of the lacZ gene of (Bernex et al., 1996) a white tail tip. Escherichia coli in place
Kit w'~
KitW'e+
Tyr ~ {albino}
KitW~/+ mice have white feet and a white tail tip. KltWV/Kit T M m i c e a r e black-eyed white.
KitW~'/+ mice have a moderate-sized white belly spot, white feet, and a white tail tip; furthermore, their coat is composed of a mixture of white, gray, and black hairs.
Tyr~/Tyr ~ mice are pink-eyed white.
of the first exon of Kit Missense mutation {Nocka et al., 1990)
replacing threonine 660 of Kit with methionine.
Missense mutation (Panthler et al., 1990; replacing glycine 597 of De Sepulveda et al., 1994) Kit with alanine.
(Jackson and Bennett, 1990) Missense mutation replacing cysteine 85 of tyrosinase with serine.
patchwork Affects Late Melanocyte Development
To de te rmine the deve lopmenta l process affected by the pa tchwork muta t ion , we studied the pat tern of Kit d is t r ibut ion in the m e s e n c h y m e and skin of pwk/pwk Kitw1acz/+ and pwk/+ Kitwzacz/+ embryos from E l l . 5 to birth. D10Mitl17, a marker closely l inked to patch- work, was used to assess the genotype of embryos at the pwk locus. The embryos were stained by whole m o u n t for 13-Gal act ivi ty. The dis t r ibut ion of/3-Gal-posit ive cells in pwk/pwk KitW~cz/+ embryos was ident ical to that of pwk/+ KitW1~cz/+ l i t t e rmates at every stage examined be tween E l l . 5 and E17.5. At E17.5, hair foll icles pro- truded from the epidermis, and melanoblas ts were found in all protuberances in all the pwk/pwk KitWl~z/+ fetuses examined at this stage {Figs. 3A and 3B). The same pat tern was observed in all the control fetuses obtained from each l i t ter (i.e., pwk/+ KitW-l~cz/+ individuals). We compared lacZ expression in skin sect ions of pwk/pwk KitWl~z/+ and pwk/+ Ki tW~z/+ newborn mice. In pwk/+ KitW-l~¢z/+ newborn mice, all hair foll icles were pigmented; they all conta ined /3-Gal-positive melano- cytes. In pwk/pwk KitWZ~¢z/+ newborn mice, the pig- men ted hair follicles conta ined /3-Gal-positive melano- cytes, whi le the whi te hair foll icles lacked these jB-Gal- posi t ive fol l icular cells {data not shown). However , on except ional occasions, a single /3-Gal-positive p igment cell could be detected in follicles of unp igmented hairs (see below). Altogether , our results indicate that the migrat ion, proliferation, and survival of melanoblas ts homozygous for patchwork are not obviously affected unt i l E17.5 and that the disappearance of melanocy tes or their precursors mus t occur in hair foll icles be tween E17.5 and birth.
To assess how and when patchwork melanoblasts die, we looked for apoptotic melanoblasts and/or melanocytes in the hair follicles of pwk/pwk KitW-l~cz/+ and pwk/+ KitWl~cz/+ fetuses from E17.5 to birth. Melanocytes and
their precursors were identified by immunosta in ing with an anti-/3-galactosidase antibody. Apoptotic cells were de- tected after specific staining of nuclear D N A fragmentation using the TUNEL method (Gavrieli et al., 19921. One or more J3-Gal-positive, TUNEL-posi t ive hair follicle cells were found on skin sect ions from 8 of 12 pwk/pwk Kitwl~z/+ individuals from E18.5 to birth (Fig. 4 and Table 21. All the double-stained cells were seen in nonpigmented hair follicles. By contrast, no double-stained hair follicle cell was detected on any skin sections from 12 pwk/+ KitWlacz/+ individuals from E18.5 to birth. The number of fetuses wi th double-stained hair follicles cells was signifi- cantly higher in patchwork individuals than in their control l i t termates (two-tailed Fisher's exact test, P value = 0.00135}. We conclude that the pwk/pwk genotype causes a significant increase in the occurrence of double-stained hair follicle cells starting at E 18.5. These results indicate that in pwk/pwk KitWl~z/+ hair follicles, melanoblasts undergo apoptosis between E18.5 and birth. Altogether, the patch- work mutat ion leads to the premature cell death of mela- noblasts at the end of fetal life; at least a fraction of the melanoblasts die by apoptosis.
There Are No Gray Hairs in p w k / p w k KitW'ei/+ Double Mutants
Muta t ions of tl~e Kit gene affect melanoblas t survival from E10.5 onward (Bernex et al., 1996; MacKenzie et al., 19971. As a result, the coat color of Kit mutan t s is more or less depigmented, depending on the Kit allele. In particular, the coat color of KitW~'/+ mutan ts consists of white , fully pigmented, and gray hairs (De Sepulveda et al., 1994; for an example of KitWe'/+ coat color phenotype , see Pan th ie r et al., 1990). Microscop ica l examina t ion of the gray hairs revealed that their medul- lary cells conta ined a variable amoun t of p igment gran- ules, some of them being comple te ly devoid of melan in {data not shown). Current unders tanding of Kit act ion
Copyright © 1998 by Academic Press All rights ol reproducuon m any form reserved
270 Aubin-Houzelstein et al.
C
FIG. 2. Detection of melanocytes in the hair follicles of 10-day-old mice by X-Gal staining (A, B, C, D) and S100 immunocytochem- istry (E, F, G, H). {A and E) pwk/pwk KitWa"cz]+ pigmented hair follicles, with several functional melanocytes (arrows) in the most internal layer of the bulb matrix (bm) in contact with the dermal papilla (dp). Melanocytes are both fl-Gal- (A) and S100- (E) positive. They are found at the same location as in the Tyr~/Tyr c KitWJacz/+ hair follicles (positive controls) [C and G). [B and F) pwk/pwk Kitw~°cz/+ white hair follicles, fl-Gal- (B) and S100- (F) positive cells are lacking, as in the KitWV/KitW-Ja~z hair follicles (negative controls) (D and Hi. fl-Gal-positive S100-negative cells can be seen in the hair matrix and the dermal papilla of pwk/pwk Kitwzacz/+ white hair follicles (B, arrowheads) and were found at the same location in all genotypes (arrowheads and data not shown). These cells are not pigment cells (bars, 25/zm).
on melanoblasts survival indicates that the whi te hairs are produced by hair follicles devoid of melanocytes , the ful ly .p igmented hairs by hair follicles wi th a normal number of melanocytes , and the gray hairs by hair follicles w i th an in te rmedia te number of melanocytes . pwk/pwk KitWe~/+ double m u t a n t s were produced. The fur color of pwk/pwk KitWe~/+ mice was m u c h di luted compared to that of pwk/pwk mice (Fig. 5, to compare wi th Fig. 1). Furthermore, who le -moun t prep- a ra t ions of hai rs r evea led tha t pwk/pwk KitW-~/+ m i c e display 85% w h i t e and 15% fu l ly p i g m e n t e d hairs, but no gray hairs. The absence of gray hairs indicates that pa tchwork melanoblasts cannot survive
and differentiate when they are in reduced number in the hair follicle.
Melanoblasts Are Target Cells for patchwork
In view of the patchwork coat color phenotype, one could postulate that patchwork acts via the hair follicle environ- ment: survival of melanoblasts would depend on a hair follicle characteristic controlled by the patchwork gene. Alternatively, patchwork could act via the melanoblasts themselves. Mouse chimeras produced by aggregating four- or eight-cell embryos that differ at a coat color locus allow one to discriminate between the determinants that act via
Copyright © 1998 by Academic Press. All rights of reproduction in any form reserved.
Coat Color Variegation in Mice 271
A
hs
4
B C D
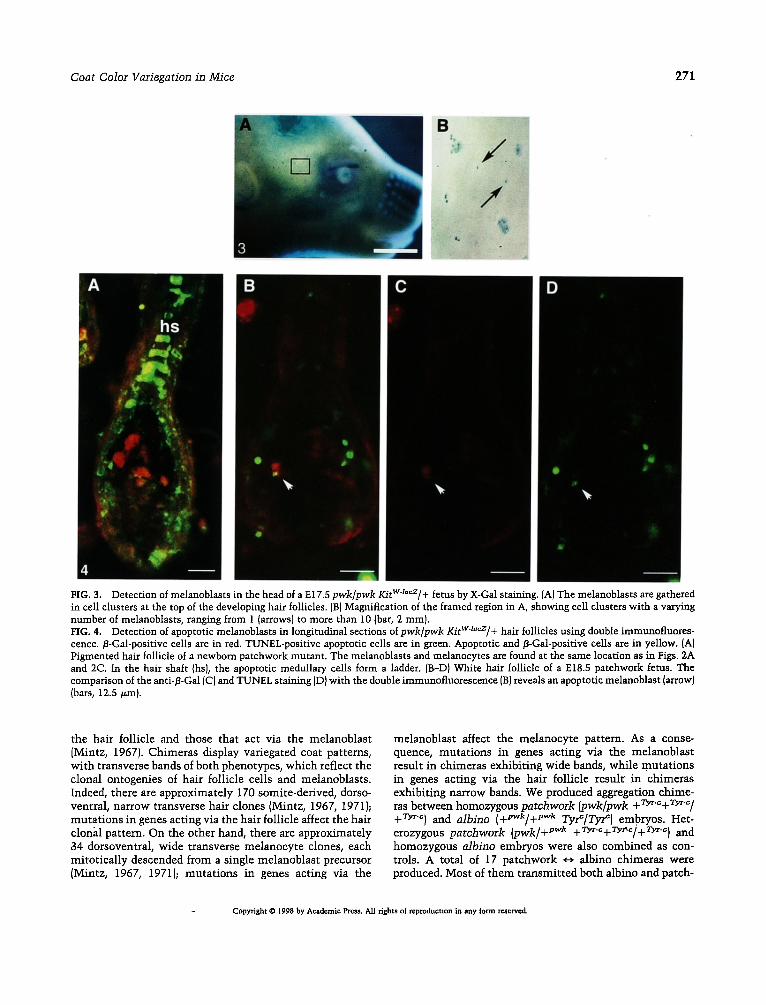
FIG. 3. Detection of melanoblasts in the head of a E17.5 pwk/pwk KitWJacz/+ fetus by X-Gal st~aing. (A) The melanoblasts are gathered in cell clusters at the top of the developing hair follicles. {BI Magnification of the framed region in A, showing cell clusters with a varying number of melanoblasts, ranging from 1 (arrows) to more than 10 (bar, 2 mm). FIG. 4. Detection of apoptotic melanoblasts in longitudinal sections ofpwk/pwk KitWlacz/+ hair follicles using double immunofluores- cence./3-Gad-positive cells are in red. TUNEL-positive apoptotic cells are in green. Apoptotic and/3-Gal-positive cells are in yellow. (A) Pigmented hair follicle of a newborn patchwork mutant. The melanoblasts and melanocytes are found at the same location as in Figs. 2A and 2C. In the hair shaft (hs), the apoptotic medullary cells form a ladder. [B-D) White hair follicle of a E18.5 patchwork fetus. The comparison of the anti-/3-Gal (C) and TUNEL staining (D) with the double immunofluorescence (B) reveals an apoptotic melanoblast (arrow) (bars, 12.5 v.m}.
the hair follicle and those that act via the melanoblast [Mintz, 1967}. Chimeras display variegated coat patterns, wi th transverse bands of both phenotypes, which reflect the clonal ontogenies of hair follicle cells and melanoblasts. Indeed, there are approximately 170 somite-derived, dorso- ventral, narrow transverse hair clones (Mintz, 1967, 1971); mutat ions in genes acting via the hair follicle affect the hair clonal pattern. On the other hand, there are approximately 34 dorsoventral, wide transverse melanocyte clones, each mitot ical ly descended from a single melanoblast precursor (Mintz, 1967, 1971); mutat ions in genes acting via the
melanoblast affect the melanocyte pattern. As a conse- quence, mutat ions in genes acting via the melanoblast result in chimeras exhibiting wide bands, while .mutations in genes acting via the hair follicle result ' in chimeras exhibiting narrow bands. We produced aggregation chime- ras between homozygous patchwork [pwk/pwk + Ty~.~+ ryr-c/ +73,~-c} and albino (+lJwk/+pwk T ~ / T y r el embryos . Het- erozygous patchwork {pwk/ + pwk +73,~-c+73,r-~/+73,~-~) and homozygous albino embryos were also combined as con- trols. A total of 17 patchwork ~ albino chimeras were produced. Most of them transmit ted both albino and patch-
Copyright O 1998 by Academic Press. All rights of reproducnon in any form ~e.senred.

272 Aubin-Houzelstein et al.
TABLE 2 Det-ection of Apoptotic Melanoblasts in Fetuses
Stage Genotype
Number of positive fetuses" [number of apoptotic
melanoblasts detected)
Number of fetuses [total number of hair follicles examined)
E17.5" pwk/pwk Kitwa"cz/+ 0 (0) 4 (22) pwk/+. Kitw'l"cz/+ 0 {0) 4 (19)
E18.5 pwk/pwk KitWa°cz/+ 2 (3J 4 (42) pwk/+ KitWaa~z]+ 0 (0) 4 (451
E19.5 pwk/pwk IO'twa~cz/+ 3 (6) 5{87) pwk/+ Kitw4acz/+ 0 (0J 5 (94)
Newborn pwk/pwk Kit w-la~z/ + 3 b {3) 3 [ 17) pwk/+ KitW'tacZ/+ 0 (0) 3 (24)
Fetuses were considered positive when at least one hair follicle contained a fl-Gal-positive, TUNEL-positive cell. b Although 3/3 mutant fetuses were found positive, a single fl-Gal-positive, TUNEL-positive pigment cell was detected in each of them.
work phenotypes to ,their progeny and thus were germ-line chimeras. All exhibited the patchwork coat color pheno- type to some extent. The chimeras were characterized by a striped coat color, with adjacent bands either white lalbino) or salt-and-pepper {patchwork) {Fig. 6A). The bands were large and well-defined, as those seen in aggregation chime- ras when the two contributing strains differ with respect to a melanocyte marker which acts via the melanocyte (Mintz, 1967). Most importantly, the transverse pigmented bands corresponding to melanoblast clones were devoid of any subclones; such subclones would be formed if the patchwork gene product was able to influence indirectly the survival of melanoblasts through the hair follicle cell components. We conclude that the primary defect in pwk/ pwk mice resides in the melanoblast rather than in the hair follicle.
HG. 5. Coat color phenotype of a pwk[pwk KitW~'[+ mouse. The fur contains no gray hairs. Note the diluted coat color compared with Fig. 1.
patchwork Encodes a Nonautonomous Signal for Melanoblast Survival
Taken together our results suggest that the patchwork pattern arises because the melanocyte precursors cannot differentiate into melanocytes when present in low number in the hair follicle. The survival defect might result from an autonomous genetic defect in single melanoblasts~ such defect would not be rescued by mixing normal and mutant melanoblasts. Alternatively, it could result from deficient cell-cell interactions or abnormal diffusible gradients~ in this case, the presence of wild-type cells in a hair follicle would occasionally rescue the mutant cell phenotype. To investigate whether the pwk/pwk genotype caused an au- tonomous defect, we closely inspected the coat of the aggregation chimeras and we found gray hairs at the bound- aries between patchwork and albino bands (Fig. 6B). The occurrence of mixed hairs in chimeras has been reported; it indicates that more than one melanocyte precursor may colonize a single hair follicle (Mintz, 1969). In this regard, gray hairs in patchwork *-~ albino chimeras must have been produced by melanocytes of both genotypes. We conclude that pwk/pwk melanoblasts, even in small numbers, can survive, differentiate, and participate in the formation of a follicle in conjunction with melanoblasts carrying a wild- type allele at the pwk locus. Thus, the defect associated with patchwork is not strictly melanoblast autonomous (see Discussion).
D I S C U S S I O N
The Survival of pa tchwork Melanoblasts Is Dependent upon Their Number in the Hair Follicle
To identify melanoblasts in pwk/pwk KitWa~cz/+ em- bryos, we have taken advantage of the Kit T M allele, a null mutat ion generated by insertion of the lacZ gene into the first exon of Kit. MacKenzie et al. (1997) recently
Copyright © 1998 by Academic Press All rights of reproducrton in any form reserved.

Coat Color Variegation in Mice 273
FIG. 6. 10-day-old albino ~ patchwork chimera. (A) Coat color phenotype: The transverse bands of both parental phenotypes represent melanoblast clones. The melanoblast clonal pattern with the left-right asymmetry is typical of mutations affecting the melanocyte and/or its precursor. (B} The three hair colors found in the chimera. The black hair (fight) was produced by a hair follicle containing patchwork melanoeytes; it can be found in the patchwork bands. The white hair (left) was produced by a hair follicle either containing nonfunctional melanocytes, as in albino bands, or devoid of melanocytes, as in patchwork bands. The gray hair (middle) can be found only at the boundaries between patchwork and albino bands; it was produced by a hair follicle containing a mixture of albino and patchwork melanocytes. This indicates that the patchwork melanoblasts in reduced number in the hair follicle were rescued by the albino melanoblasts; these patchwork melanoblasts survived and differentiated further into functional melanocytes [bar, 50 gin).
reported that there is a significant decrease in the number of melanoblasts in embryos heterozygous for Kit wv, a point mutat ion exerting a dominant negative effect in comparison to wild type. Thus the embryos heterozygous for the Kit T M null allele may not give an accurate representation of melanoblast distribution in wild-type embryos. Therefore the effect of the patchwork mutat ion on melanoblast development was assayed by comparing the X-Gal staining in KitWaacz/+ embryos that were heterozygous or homozygous for the patchwork mutat ion from within the same litter. No difference in expression of the lacZ reporter was seen unti l E18.5. By E18.5, apoptotic melanoblasts were repeatedly found in hair follicles in Kitw-z"cz/+ fetuses that were homozygous for the patchwork mutation. In contrast, no apoptotic melanoblast has ever been detected in hair follicles in Kitwaacz/+ fetuses heterozygous for the patchwork mu- tation. Furthermore, in KitWa"cz/+ fetuses homozygous for the patchwork mutation, apoptotic melanoblasts were found only in the follicles of unpigmented hairs; no apoptotic melanoblast was detected in the follicles of pigmented hairs. These observations are consistent with a vital role for patchwork by E18.5.
The results obtained with the Kit wei mutation highlight the importance of the melanoblast number in the patch- work phenotype. The follicles of q_pwk/_~pvcl¢ KitW-~i/+ mice contain variable numbers of melanocytes; their hairs are white, gray, or fully pigmented. The gray hairs indicate that melanoblasts carrying wild-type alleles at the pwk locus can survive and differentiate in the hair follicle, even when their number is too low to produce a fully pigmented hair.
By contrast, the coat of pwk/pwk KitW-~i/+ mice contains more white hairs, but no gray hairs. Thus, the survival of patchwork melanoblasts depends on their number within a given hair follicle.
Gray hairs were found in patchwork ~ albino chime- ras, at the limits between albino and patchwork bands. They must have been produced by mixed hair follicles containing both albino and patchwork melanocytes. Thus patchwork melanoblasts in low number survive and differentiate in the mixed hair follicles, indicating that the patchwork defect can be rescued by neighboring normal cells. In other words, the action of the patchwork gene is not cell autonomous. The rescue does not result from the chimeric follicular environment, but from the presence of albino melanoblasts. This is shown by the coat color phenotype of the aggregation chimeras, which indicates that patchwork acts via the melanoblast, rather than via its environment. The albino melanoblasts pre- sumably provide the patchwork melanoblasts with a survival and/or a differentiation signal.
Possible Mechanisms for patchwork
One model to account for our data is that the pwk gene product is secreted by melanoblasts and that its concentra- tion is crucial for melanoblast survival and/or differentia- tion in the hair follicle at E18.5. In pwk/pwk hair follicles, when the concentration of the pwk gene product is below a certain threshold, the melanoblasts fail to differentiate and die. Thus, groups of few pwk/pwk melanoblasts die, but larger groups survive because an above-threshold concen-
Copyright © 1998 by Academic Press All fights of reproduction in any form reserved.

274 Aubin-Houzelstein et al.
tration of the mutant protein is produced. In patchwork albino chimeras, the patchwork protein expressed by albino melanoblasts rescued the viability and differentiating po- tential of the pwk/pwk melanoblasts colonizing the same hair follicle. However, this lack of cell autonomy does not necessarily imply that the patchwork gene encodes a dif- fusible product; indeed, it may control the production of one. Thus, patchwork could encode a protein involved in the expression of a secreted product rather than the product itself.
Another possibility is that pwk encodes a receptor in- volved in an intercellular signal for melanoblast survival and/or differentiation at E18.5. This latter mechanism is known to be responsible for the melanocyte defect associ- ated with the piebald (Ednrb) phenotype. Ednrb acts primar- ily on melanoblast viability in a non-cell-autonomous man- ner, as does pwk (Mayer, 1977; Kapur et al., 1995). However Ednrb acts before El0.5 (Pavan and Tilghman, 1994), whereas pwk acts at E18.5 {our data). As a result, mice homozygous for a severe allele at the Ednrb locus are almost completely white with only small pigmented areas. Both mutations have an all-or-nothing behavior: pigmented areas in Ednrb mutants are fully colored, as are the pig- mented hairs in pwk/pwk mice. Edrzrb encodes the G-protein-coupled receptor-B (EDNRB) for endothelin 3 (EDN3). EDNRB is expressed in melanoblasts, while EDN3 is secreted in their environment (Baynash et al., 1994; Hosoda et al., 1994; Kaput et al., 1996). To account for the cooperation between EDNRB-expressing cells, Kapur et al. (1995) have proposed a model in which autocrine or para- crine stimulation of cells expressing the EDNRB receptor initiates downstream events that alter the extracellular environment encountered by EDNRB-bearing cells and thereby modulate their behavior. As a result, the prolifera- tion, migration, and/or survival of EDNRB-bearing cells would be stimulated. This model could also apply to patchwork-expressing cells.
Variegations in p a t c h w o r k Mice
Individuals with an apparent mixture of two pheno- types, i.e., variegated phenotype, occur in mice. Such variegation can be due to genetic events, such as mitotic recombination (De Sepulveda et al., 1995) or deletion (Gondo et al., 1993). It is also observed for X-linked coat color mutations due to inactivation of one or the other of the X chromosomes (Lyon, 1961). We provide evidence that the variegated phenotype of patchwork mice results from the cooperation of several melanoblasts, allowing any one of them to survive and progress to a differenti- ated state. Interestingly, melanoblasts originating from the epidermis or from adjacent pigmented hair follicles do not colonize the follicles devoid of melanocytes. This indicates that from E18.5 onward, each follicle is an independent developmental unit. Furthermore, the cho- roid of the eyes of pwk/pwk mice, which contains neural crest-derived melanocytes, has a normal pigmentation
(data not shown). Thus, patchwork has no effect on melanoblasts outside the hair follicle, either because the large number of melanoblasts within the choroid pro- duces a sufficient amount of the pwk protein or because pwk is not needed in the eye.
Two-thirds of pwk/pwk mice exhibit patches of wild- type coat (data not shown). The shape and distribution of these phenotypic reversion patches are similar to those previously observed in Kit mutant mice and to the phenoclones of aggregation chimeras (Mintz, 1971; Pan- thier et al., 1990). These phenotypic reversions could result from the clonal development of melanoblasts strongly expressing the mutant allele, although other mechanisms cannot be excluded.
patchwork Melanoblasts Are Subjec t to a C o m m u n i t y Effect
The accumulated data are consistent with survival of patchwork melanoblasts being dependent upon their num- ber in the hair follicle. Such a phenomenon has been described as a community effect (Gurdon, 1988; Gurdon et al., 1992, 1993a, b). This concept implies interactions between cells of the same type in order to respond to inductive signals. Initially described in the Xenopus em- bryo for uncommitted cells, it was later extended to cells already engaged in a specific differentiation pathway. In- deed, committed cells occasionally need to cooperate to respond to differentiation signals (Gurdon et al., 1992, 1993a). Community effects have also been described in Drosophila and in zebrafish (Stuttem and Campos-Ortega, 1991; Ho, 1992). In the mouse, ex vivo experiments have shown that a minimal number of 30-40 cells is required for mesodermal cells to give rise to myogenic clones (Cossu et al., 1995). However, examples illustrating community ef- fects in mammals remain rare, and remarkably few data exist concerning mammalian genes controlling a commu- nity effect. One reason is that mutations in these genes are expected to lead to severe phenotypes, resulting from the loss of a cell lineage or a cell type. The best conditions for identifying these mutants will be met when the commu- nity effect concerns a cell lineage whose absence does not affect the viability and the fertility of the mutant. We think that these conditions are fulfilled in the case of the patch- work mice.
ACKNOWLEDGMENTS
The authors are grateful to Karen Moore for the generous gift of the patchwork stock. They are also indebted to Charles Babinet, Fran~oise Poirier, Fr~d6ric Rosa and Marie-Christine Simmler for critically reading the manuscript. This work was supported by the Institut National de la Recherche Agronomique, the Mmxst~re de l'Agriculture et de la For~t (MAF}, the Fondation d~ France, and the Association pour la Recherche sur le Cancer (ARC). G. A. H. was funded by the MAF and then by the ARC.
Copyright © 1998 by Academic Press All rights of reproducuon m any form reserved.

Coat Color Variegat ion in Mice 275
REFERENCES
Barsh, G. S. (1996). The genetics of pigmentation From fancy genes to complex traits. Trends Genet. 12, 299-305.
Baynash, A. G., Hosoda, K., Giaid, A., Richardson, J. A., Emoto, N., Hammer, R. E., and Yanagisawa, M. (1994). Interaction of endothelin-3 with endothelm-B receptor is essential for develop- ment of epidermal melanocytes and enteric neurons. Cell 79, 1277-1285.
Bemex, F., De Sepulveda, P., Kress, C., Elbaz, C., Delores, C., and Panthier, J. J. (199@ Spatial and temporal patterns of c-kit- expressing cells in W ~ / + and Wl~z/W l'cz mouse embryos. Development 122, 3023-3033.
Chase, H. B. (1954). Growth of the hair. Physiol. Rev 34, 113-126.
Cossu, G., Kelly, R., Di-Donna, S., Vivarelli, E., and Buckingham, M. (1995). Myoblast differentiation during mammalian somito- genesis is dependent upon a community effect. Proc. Natl. Acad. Sci USA 92, 2254-2258.
De Sepulveda, P., Guenet, J. L., and Panthler, J. J. (1995}. Phenotypic reversions at the W/Kit locus mediated by mitotic recombination in mice. Mol. Cell. Biol. 15, 5898-5905.
De Sepulveda, P., Peyrieras, N., and Panthier, J. J. (1994). Instabdity at the W/c-kit locus in mice: Analysis of melanocyte cell lines derived from reversion spots. Oncogene 9, 2655-2661.
Dietrich, W. F., Miller, J. C., Steen, R. G., Merchant, M., Damron, D., Nahf, R., Gross, A., Joyce, D. C., Wessel, M., Dredge, R. D., et al. {1994}. A genetic map of the mouse with 4,006 simple sequence length polymorphisms. Nat. Genet. 7, 220 -245.
Gavneli, Y., Sherman, Y., and Ben-Sasson, S. A. (1992). Identifica- tion of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 119, 493-501.
Gondo, Y., Gardner, J. M., Nakatsu, Y., Durham-Pierre, D., Deveau, S. A., Kuper, C., and Brilliant, M. H. (19931. High-frequency genetic reversion mediated by a DNA duplicatmn: The mouse pink-eyed unstable mutation. Proc. Natl. Acad. Sci. USA 90, 297-301.
Gurdon, J. B. (1988). A community effect in animal development. Nature 336, 772-774.
Gurdon, J. B., Kao, K., Kato, K., and Hopwood, N. D. (1992}. Muscle gene activation in Xenopus requires intercellular communica- tion during gastrula as well as blastula stages. Dev. Suppl. 137-142.
Gurdon, J. B., Lemmre, P., and Kato, K. {1993a}. Community effects and related phenomena in development. Cell 75, 831-834.
Gurdon, J. B., Tiller, E., Roberts, J., and Kato, K. (1993b). A community effect in muscle development. Curr. Biol. 3, 1-11.
Hardy, M. H. (19921. The secret life of the hair follicle. Trends Genet. 8, 55-61.
Hirobe, T. (1995). Structure and function of melanocytes: Micro- scopic morphology and cell biology of mouse melanocytes in the epidermis and hair follicle. H1stol. Hlstopathol. 10, 223-237.
Ho, R. K. (1992). Cell movements and cell fate during zebrafish gastrulation. Dev. Suppl. 65-73.
Hosoda, K., Hammer, R. E., Richardson, J. A., Baynash, A. G., Cheung, J. C., Giaid, A., and Yanagisawa, M. (1994). Targeted and natural (piebald-lethal} mutations of endothelin-B receptor gene produce megacolon associated with spotted coat color in mice. Ceil 79, 1267-1276.
Ito, K., Monta, T., and Sieber-Blum, M. (1993). In vitro clonal analysis of mouse neural crest development. Dev. Biol. 157, 517-525.
Jackson, I. J. (1991). Mouse coat colour mutations: A molecular genetic resource which spans the centuries. Bioessays 13, 439-446.
Jackson, I. J. (1994}. Molecular and developmental genetics of
mouse coat color. Annu. Rev. Genet. 28, 189-217. Jackson, I. J., and Bennett, D. C. (1990). Identification of the albino
mutation of mouse tyrosinase by analysis of an in vitro revertant. Pro& Natl. Acad. Sci. USA 87, 7010-7014.
Jackson, I. J., Budd, P., Horn, J. M., Johnson, R., Raymond, S., and Steel, K. (1994). Genetics and molecular biology of rdouse pig- mentation. Pigment Cell Res. 7, 73-80.
Kapur, R. P., Livingston, R., Doggett, B., Sweetser, D. A., Siebert, J. R., and Palmiter, R. D. {1996). Abnormal microenvironmental signals underlie intestinal aganghonosis in dominant megacolon mutant mice. Dev. Biol. 174, 360-369.
Kapur, R. P., Sweetser, D. A., Doggett, B., Siebert, J. R., and Palmiter, R. D. (1995}. Intercellular signals downstream of endo- thelin receptor-B mediate colonization of the large intestine by enteric neuroblasts. Development 121, 3787-3795.
Kunisada, T., Yoshida, H., Ogawa, M., Shultz, L. D., and Nish- ikawa, S. (199@ Characterization and isolation of melanocyte progenitors from mouse embryos. Dev. Growth Differ. 38, 87-97.
Lyon, M. F. (1961). Gene action in the X-chromosome of the mouse {Mus musculus L.). Nature 190, 372-373.
MacKenzie, M. A. F., Jordan, S. A., Budd, P. S., and Jackson, I. J. (1997}. Activation of the receptor tyrosine kinase Kit is required for the proliferation of melanoblasts in the mouse embryo. Dev Biol. 192, 99-107.
Mayer, T. C. {1970}. A comparison of pigment cell development in albino, steel, and dominant-spotting mutant mouse embryos. Dev. Biol. 23, 297-309.
Mayer, T. C. (1973}. The migratory pathway of neural crest cells into the skin of mouse embryos. Dev. Biol. 34, 39-46.
Mayer, T. C. {1977). Enhancement of melanocyte development from piebald neural crest by a favorable tissue environment. Dev. Biol. 56, 255-262.
Mintz, B. {1967). Gene control of mammalian pigmentary differen- tiation. I. Clonal origin of melanocytes. Proc Natl. Acad. Sci. USA 58, 344-351.
Mintz, B. (1969). Developmental mechanisms found in allophenic mice with sex chromosomal and p~gmentary mosalcism. Birth Dejects Orig. Attic. Ser. 5, 11-22.
Mintz, B. (1971). Control mechanisms of growth and differentia- tion. In "25th Symposium of the Society for Experimental Biology" (D. D. Davies and M. Balls, Eds.L pp. 345-369. Cam- bridge Univ. Press, Cambridge, UK.
Nicolson, G. L., Yanagimachi, R., and Yanagimachi, H. (1975). Ultrastructural localization of lectin-binding sites on the zonae pellucidae and plasma membranes of mammalian eggs. J. Cell Biol. 66, 263-274. '
Nocka, K., Tan, J. C., Chin, E., Chu, T. Y., Ray, P., Traktman, P., and Besmer, P. { 1990). Molecular bases of dominant negative and loss of function mutations at the murine c-kit~white spotting locus: W az, W v, W al and W. EMBO J. 9, 1805-1813.
Orlow, S. J. (1995). Melanosomes are specialized members of the lysosomal lineage of organelles. J. Invest. Dermatol. 105, 3-7.
Panthier, J. J., Guenet, J. L., Condamme, H., and Jacob, F. {1990). Evidence for mitotic recombination in W~'/+ heterozygous mice. Genetics 125, 175-182.
Pavan, W. J., and Tilghman, S. M. {1994}. Piebald lethal [sl) acts early to disrupt the development of neural crest-derived melano- cytes. Proc. Natl. Acad. Sci. USA 91, 7159-7163.
Copyright © 1998 by Academlc Press All rights of reproduction in any form reserved.

2 7 6 Aubin-Houzelstein et al.
Rawles, M. E. {1947). Ongin of pigment cells from the neural crest in the mouse embryo. Physiol. Zool. 20, 248-266.
Serbedzija, G. N., Brormer-Fraser, M., and Fraser, S. E. (1994). Developmental potential of trunk neural crest cells in the mouse. Development 120, 1709-1718.
Serbedzija, G. N., Fraser, S. E., and Brormer-Fraser, M. (1990). Path- ways of trunk neural crest cell migration in the mouse embryo as revealed by vital dye labelling. Development I08, 605-612.
Silvers, W. (1979). "The Coat Colors of Mice: A Model for Mam- malian Gene Action and Interaction," pp. 147-158. Springer- Verlag, New York.
Smoller, B. R., Hsu, A., and Krueger, J. [1991). HMB-45 monoclonal antibody recognizes an inducible and reversible melanocyte cytoplasmic protein. ]. Cutan. Pathol. 18, 315-322.
Steel, K. P., Davidson, D. R., and Jackson, I. J. (1992). TRP-2/DT, a new early melanoblast marker, shows that steel growth
factor [c-kit ligand) is a survival factor. Development 115, 1111-1119.
Stuttem, I., and Campos-Ortega, J. A. (1991}. Cell commitment and cell interactions in the ectoderm of Drosophila melanogaster. Development 2, 39-46.
Yokoyama, T., Silversides, D. W., Waymire, K. G., Kwon, B. S., Takeuchi, T., and Overbeek, P. A. (1990). Conserved cysteine to serine mutation in tyrosinase is responsible for the classical albino mutation in laboratory mice. Nucleic Acids Res. 18, 7293-7298.
Yoshida, H., Kunisada, T., Kusakabe, M., Nishikawa, S., and Nishikawa, S. I. {1996). Dist inct stages of melanocyte differ- entiat ion revealed by analysis of nonuniform pigmentat ion patterns. Development 122, 1207-1214.
Received for publication January 29, 1998 Accepted March 27, 1998
Copyright © 1998 by Academic Press. All nghts of reproducnon m any form reserved
Related Documents