Journal of Leukocyte Biology 48:258-265(1990) © 1990 Wiley-Liss, Inc. Suppression of Late Phase Enhanced Vascular Permeability in Rats by Selective Depletion of Neutrophils With a Monoclonal Antibody Sakae Sekiya, Takao Yamashita, and Fujiro Sendo Department of Parasitology, Yamagata University, School of Medicine, Yamagata, Japan We investigated the role of neutrophils in increased vascular permeability by a selective reduction of neutrophlls usIng a monoclonal antibody, RP-3. An intraperitoneal Injection of RP-3 not only selectively depleted peripheral blood neutrophils, but prevented the neutrophil Infiltration to the tissues. Proteose peptone, zymosan, and BCG induced three different types of Inflammatory edema, showing the early phase only, early plus late phase, and the late phase only, respectively. Only the late phase response of zymosan and BCG was Inhibited by a depletion of neutrophils by RP-3, though the early phase response induced by proteose peptone and zymosan was not affected. Reconstitution of neutrophil-depleted rats by In situ injection of these cells restored the inflammatory edema induced by BCG, depending upon the number of neutrophils injected. Key words: edema, zymosan, BCG INTRODUCTION Neutrophils infiltrate the tissues in the early phase of a serial response from inflammation to immunity. They not only play a major role in defense against bacterial infection, but may modulate inflammation and immunity by producing active oxygens [ 1 ,35] , platelet activating factor [ 1 8] , leukotriene B4 [2] , and interleukin 1 like molecule [7,31]. The possible role of neutrophils in in- creased vascular permeability has been discussed for many years [3, 13, 15,34], but a definite conclusion has not been obtained. Recently, many in vitro experiments have shown the interaction between neutrophils and en- dothelial cells [4,6,8]. Considering these results, we in- tended, in the present study, to clarify the role of neu- trophils in the formation of inflammatory edema. To this end, we used a monoclonal antibody that selectively de- pleted rat peripheral blood neutrophils which we had recently established [27]. In this paper we demonstrate that the late phase increased vascular permeability is de- pendent on the in situ activation of neutrophils. MATERIALS AND METHODS Chemicals The chemicals used were zymosan (Sigma Chemical Co. St. Louis, MO); Bacillus Calmette-Guerin (BCG) (Kyowa Chemical Industries LTD. , Tokyo, Japan); pro- teose peptone (Difco Laboratories, Detroit, MI); diphen- hydramine (Tanabe Pharmaceutical Co., LTD., Osaka, Japan); cimetidine (Fujisawa Pharmaceutical Co. , LTD., Osaka, Japan); Ficoll-Isopaqe gradient (Pharmacia Fine Chemicals, Uppsala, Sweden); Eagle’s minimum essen- tial medium (MEM) (Nissui Pharmaceutical Co. , LTD., Tokyo, Japan); ‘25!-bovine serum albumin (BSA) (ICN Radiochemical, Division of ICN Biochemicals, Inc. Irv- me California); May-Gruenwald’s solution, Giemsa’s solution, and Schiff’s solution (E. Merk, Darmstadt, FRG); and heparin calcium (Shimizu Pharmaceutical Co., LTD., Shizuoka, Japan). Animals Wistar-King-Aptakemann (WKA)/Hok rats were kept and bred in a pathogenic bacteria-free condition by fil- tered air flow in the Yamagata University Animal Center (clean WKA/Hok rats). Specific pathogen free (SPF) WKA/Hkm rats and SPF BALB/c mice were purchased from Funabashi farm (Shizuoka, Japan). Six to nine wk- old male rats were used in the experiments. Cell Preparation Rat neutrophils. Ten milliliters of 3% proteose pep- tone were injected into the peritoneal cavities of rats. Twelve hours later, the same volume of proteose peptone was again administered. Three hours after the last injec- tion, peritoneal exudate cells (PEC) were obtained by Received June 12, 1989; accepted January 22, 1990. Reprint requests: Fujiro Sendo, Department of Parasitology, Yama- gata University School of Medicine, lida-Nishi, Yamagata, 990-23, Japan.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Leukocyte Biology 48:258-265(1990)
© 1990 Wiley-Liss, Inc.
Suppression of Late Phase Enhanced VascularPermeability in Rats by Selective Depletion of
Neutrophils With a Monoclonal Antibody
Sakae Sekiya, Takao Yamashita, and Fujiro SendoDepartment of Parasitology, Yamagata University, School of Medicine, Yamagata, Japan
We investigated the role of neutrophils in increased vascular permeability by a selectivereduction of neutrophlls usIng a monoclonal antibody, RP-3. An intraperitoneal Injectionof RP-3 not only selectively depleted peripheral blood neutrophils, but prevented theneutrophil Infiltration to the tissues. Proteose peptone, zymosan, and BCG induced threedifferent types of Inflammatory edema, showing the early phase only, early plus latephase, and the late phase only, respectively. Only the late phase response of zymosanand BCG was Inhibited by a depletion of neutrophils by RP-3, though the early phaseresponse induced by proteose peptone and zymosan was not affected. Reconstitution ofneutrophil-depleted rats by In situ injection of these cells restored the inflammatoryedema induced by BCG, depending upon the number of neutrophils injected.
Key words: edema, zymosan, BCG
INTRODUCTION
Neutrophils infiltrate the tissues in the early phase of a
serial response from inflammation to immunity. They
not only play a major role in defense against bacterial
infection, but may modulate inflammation and immunity
by producing active oxygens [ 1 ,35] , platelet activating
factor [ 1 8] , leukotriene B4 [2] , and interleukin 1 like
molecule [7,31]. The possible role of neutrophils in in-
creased vascular permeability has been discussed for
many years [3, 13, 15,34], but a definite conclusion has
not been obtained. Recently, many in vitro experiments
have shown the interaction between neutrophils and en-
dothelial cells [4,6,8]. Considering these results, we in-tended, in the present study, to clarify the role of neu-
trophils in the formation of inflammatory edema. To this
end, we used a monoclonal antibody that selectively de-
pleted rat peripheral blood neutrophils which we had
recently established [27]. In this paper we demonstrate
that the late phase increased vascular permeability is de-
pendent on the in situ activation of neutrophils.
MATERIALS AND METHODS
Chemicals
The chemicals used were zymosan (Sigma Chemical
Co. St. Louis, MO); Bacillus Calmette-Guerin (BCG)
(Kyowa Chemical Industries LTD. , Tokyo, Japan); pro-
teose peptone (Difco Laboratories, Detroit, MI); diphen-
hydramine (Tanabe Pharmaceutical Co., LTD., Osaka,
Japan); cimetidine (Fujisawa Pharmaceutical Co. , LTD.,
Osaka, Japan); Ficoll-Isopaqe gradient (Pharmacia Fine
Chemicals, Uppsala, Sweden); Eagle’s minimum essen-
tial medium (MEM) (Nissui Pharmaceutical Co. , LTD.,
Tokyo, Japan); ‘25!-bovine serum albumin (BSA) (ICN
Radiochemical, Division of ICN Biochemicals, Inc. Irv-
me California); May-Gruenwald’s solution, Giemsa’s
solution, and Schiff’s solution (E. Merk, Darmstadt,
FRG); and heparin calcium (Shimizu Pharmaceutical
Co., LTD., Shizuoka, Japan).
Animals
Wistar-King-Aptakemann (WKA)/Hok rats were kept
and bred in a pathogenic bacteria-free condition by fil-
tered air flow in the Yamagata University Animal Center
(clean WKA/Hok rats). Specific pathogen free (SPF)
WKA/Hkm rats and SPF BALB/c mice were purchased
from Funabashi farm (Shizuoka, Japan). Six to nine wk-
old male rats were used in the experiments.
Cell Preparation
Rat neutrophils. Ten milliliters of 3% proteose pep-tone were injected into the peritoneal cavities of rats.
Twelve hours later, the same volume of proteose peptone
was again administered. Three hours after the last injec-
tion, peritoneal exudate cells (PEC) were obtained by
Received June 12, 1989; accepted January 22, 1990.
Reprint requests: Fujiro Sendo, Department of Parasitology, Yama-gata University School of Medicine, lida-Nishi, Yamagata, 990-23,
Japan.

Vascular permeability =
count of the skin
count of the plasma ( 1 ml)
Microscopic Studies
Vascular Hyperpermeability and Neutrophils 259
peritoneal lavage with MEM (pH 7.4) supplementedwith fetal calf serum (FCS) (1%). Neutrophils were pu-
rifled with Ficoll-Isopaque gradient (specific gravity
I .090) according to the method previously described
[12].
The Monoclonal Antibody
We used a monoclonal antibody (RP-3) that selec-
tively depleted peripheral blood neutrophils of rats by
intraperitoneal injection. The details of the establishment
of RP-3 and its nature were described elsewhere [8,27].
Briefly, the peritoneal neutrophils of WKA/Hok rats
were injected into BALB/c mice, and their spleen cells
were hybridized with P3-X63-Ag8.653 myeloma cells
[14] according to the Oi and Herzenberg method [24].
Then a hybridoma (RP-3) was produced. The ascitic
fluid containing the monoclonal antibody (RP-3) was
centrifuged at 1 ,700g, and the supernatant was used in
all of the experiments. It was stored at -40#{176}Cuntil use.
The antibody titer of each lot of RP-3 was examined witha complement mediated cytotoxicity assay , and was
nearly the same every time. The supernatant of the as-
cites of the parental P3-X63-Ag8.653 myeloma cells (X-
63 ascitic fluid) was intraperitoneally injected as a con-
trol ascitic fluid. The ascitic fluid of the hybridoma
(74D1 1 (1gM)) that was produced by hybridization of
P3-X63-Ag8.6S3 and spleen cells of BALB/c mice
which were infected with Schistosoma japonicum [28]
were also used as another control. The monoclonal an-
tibody 74D1 1 reacted with S. japonicum eggs, but not
with the neutrophils of WKA/Hok rats.
Administration of Monoclonal Antibodies
The ascitic fluid of RP-3, 74Dl 1 , or X-63 was in-
jected into the peritoneal cavities of rats. Six hours later,
various reagents were intradermally injected to evoke
inflammatory edema.
Measurement of Vascular Permeability
The method described by Hellewell and Williams [ 10]
was used. First, the hair of the abdominal skin of the rats
was shaved, and 0. 1 ml of solution of the reagents was
intradermally injected in triplicates. The same experi-
ments were repeated at least twice. After from 3 mm to
4.5 hr, the mixtures of PBS added with ‘25I-bovine se-
rum albumin (10 �i Ci/kg) and Evan’s blue (1%) wereinjected intravenously. After 30 mm, heparmnized blood
(10 U/ml) was obtained by heart puncture. After sacri-
fice, the skins of the rats were stripped off and the in-
jected sites were punched out in 16 mm-diameter pieces
by a cork borer. The readioactivity of the plasma and the
skin was counted by a ‘y-counter. The plasma volume of
the skin was calculated from the following formula:
x 1 ,000 (pA/site).
Ten percent formalin-fixed and paraffin-embedded
specimens were used. Hematoxylin-eosin- and PAS-
stained specimens were observed microscopically.
Measurement of Complement Titer
The method described by Mayer [23] was used. First,
sheep red blood cells (SRBC) were sensitized with the
serum of rabbits immunized with SRBC in the optimal
dilution, and were mixed with the test sample. The mix-
ture were incubated at 37#{176}Cfor 90 mm and hemolysis
was determined by absorbance at 541 nm. The amount of
complement required for destruction of 50% of S x 108SRBC in 7.5m1 medium is defined as one CH�0.
Statistical Analysis
The data showed the mean ± SE. The Student’s t test
was employed for statistical analysis.
RESULTS
Inhibition of Zymosan Induced Increase inVascular Permeability by the Administrationof RP-3
One milliliter of the ascitic fluid of RP-3, X-63, or
74D1 1 was injected intraperitoneally. Six hours later,
zymosan was injected intradermally. After another 3.5
hr, 125I-BSA was injected intravenously. The vascular
permeability at 4 hr was examined. The vascular perme-
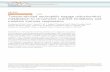
ability induced by zymosan was increased in a dose-
dependent manner in the control rats that had receivedinjections of X-63 , 74D 1 1 , or nothing, but the increase
in vascular permeability was almost completely sup-
pressed in the rats pre-treated with RP-3 within the ex-
perimental condition mentioned above (Fig. 1). Histo-
logical examinations were performed in order toascertain that neutrophils did not infiltrate the connective
tissues, even by stimulation with zymosan in rats of
which peripheral blood neutrophils had been depleted by
RP-3. It was observed that numerous neutrophils had
infiltrated the clump of zymosan in the untreated rats
(Fig. 2A). In the rats pretreated with RP-3, however,
neutrophils hardly infiltrated the zymosan (Fig. 2B). The
number of neutrophils infiltrated in the untreated and
RP-3 treated groups were 70.7±8.5, and 0.7±0.7, re-
spectively, per field (200 times magnification) from the
three randomly selected photographed field from the dif-
ferent untreated and treated rats. This result shows thatintraperitoneal administration of RP-3 not only depleted
a
.0aa,
E
a00
a>
�
500 Saline
260 Sekiya et al.
4).
20
15
10
5
0
50 200
Dose of zymozan ()Jg/O.lmf)
Fig. 1 . Increased vascular permeability induced by zymosanand its suppression by RP-3. The ascitic fluid (1 ml) of RP-3,X-63, and 74D1 1 was i.p. Injected Into rats, and 6 hr later, 50,200, and 500 �.tg/0.1 ml zymosan was intradermally injected.Vascular permeability was examined at 4 hr. The symbols rep-resent: pretreated with RP-3 ., non-treated ., pre-treated withX-63 A, pre-treated with 74D1 1 A, and the control value injectedwith sterilized saline-. The values show the mean ± SE. Thesame examinatIon was repeated 4 times.
the number of peripheral blood neutrophils, but pre-vented the neutrophil infiltration to the tissues.
Time Kinetics of Inhibition by RP-3 of IncreasedVascular Permeability Induced by Zymosan
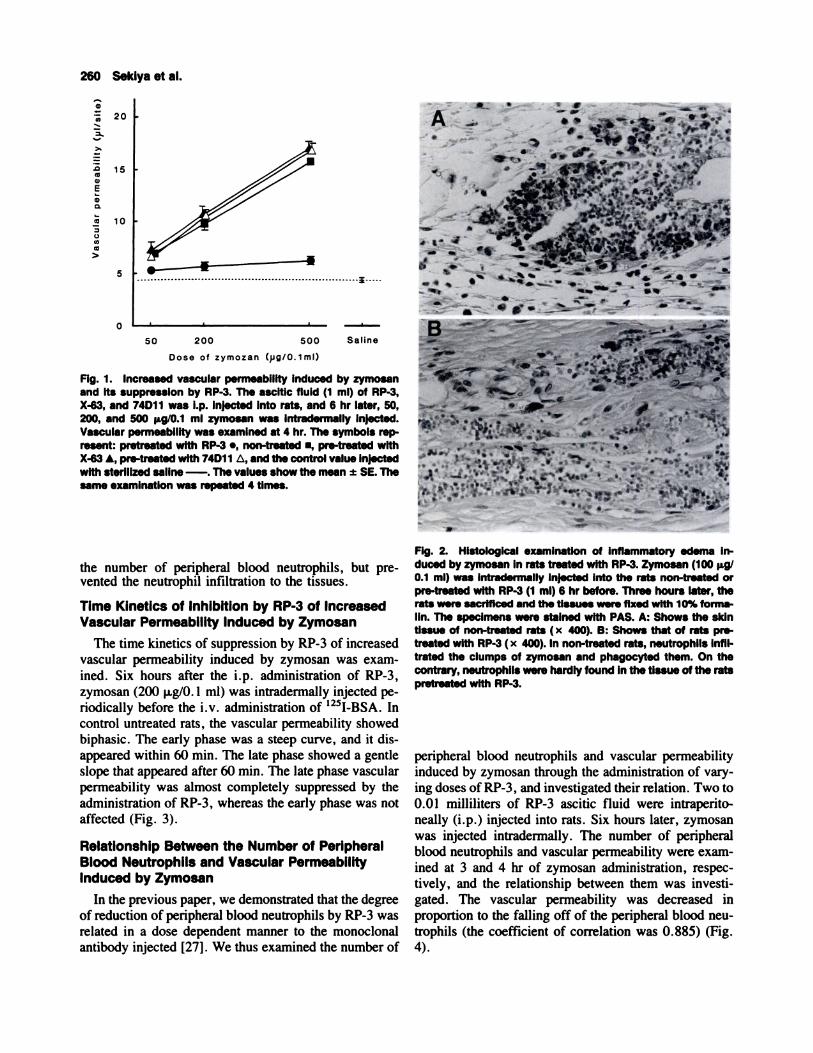
The time kinetics of suppression by RP-3 of increased
vascular permeability induced by zymosan was exam-
med. Six hours after the i.p. administration of RP-3,
zymosan (200 p.g/0. I ml) was intradermally injected pe-
riodically before the i.v. administration of ‘25I-BSA. In
control untreated rats, the vascular permeability showed
biphasic. The early phase was a steep curve, and it dis-
appeared within 60 mm. The late phase showed a gentleslope that appeared after 60 mm. The late phase vascular
permeability was almost completely suppressed by the
administration of RP-3, whereas the early phase was not
affected (Fig. 3).
Relationship Between the Number of PeripheralBlood Neutrophils and Vascular PermeabilityInduced by Zymosan
In the previous paper, we demonstrated that the degree
of reduction of peripheral blood neutrophils by RP-3 was
related in a dose dependent manner to the monoclonal
antibody injected [27] . We thus examined the number of
Fig. 2. Histological examination of inflammatory edema in-duced by zymosan in rats treated with RP-3. Zymosan (100 �tg/0.1 ml) was intradermally Injected into the rats non-treated orpre-treated with RP-3 (1 ml) 6 hr before. Three hours later, therats were sacrificed and the tissues were fixed with 10% forma-lin. The specimens were stained with PAS. A: Shows the skintissue of non-treated rats ( x 400). B: Shows that of rats pie-treated with RP-3 ( x 400). In non-treated rats, neutrophils infil-trated the clumps of zymosan and phagocyted them. On thecontrary, neutrophils were hardly found in the tissue of the ratspretreated with RP-3.
peripheral blood neutrophils and vascular permeability
induced by zymosan through the administration of vary-
ing doses of RP-3, and investigated their relation. Two to
0.01 milliliters of RP-3 ascitic fluid were intraperito-
neally (i.p.) injected into rats. Six hours later, zymosanwas injected intradermally. The number of peripheral
blood neutrophils and vascular permeability were exam-med at 3 and 4 hr of zymosan administration, respec-
tively, and the relationship between them was investi-
gated. The vascular permeability was decreased in
proportion to the falling off of the peripheral blood neu-
trophils (the coefficient of correlation was 0.885) (Fig.
a,
� 25
0�
>%
.0a,a,
Ea,
a
.� 1000C,,
>
5
0 1 2 3 4 5
Time ( hr
-I---
Saline
. .
0 500 10001500 Saline
0
Fig. 3. Time kinetics of increased vascular permeability in-duced by intradermal injection of zymosan. Approximately 6 hrbefore the zymosan Injection, 1 ml RP-3 was i.p. injected. Zy-mosan (200 �ig/0.1 ml) was intradermally injected at varyingtimes (5 hr-33 mm) before sacrifice. The symbols represent:pre-treated with RP-3., non-treated., saline control - - -. Thelate phase vascular permeability was suppressed by adminis-tration of RP-3. The same experiments were repeated 3 times.
15
C,,
0�
.0a 10a,Ea,
aa00
>
0
Number of peripheral neutrophils ( /mm3)
Fig. 4. The vascular permeability induced by zymosan wasproportional to the number of peripheral blood neutrophils.Varying doses of RP-3 were i.p. injected into the rats. Six hourslater, zymosan (200 �g/0.1 ml) was Intradermally injected. Thenumbers of peripheral blood neutrophils and vascular perme-ability were examined at 3 hr and 4 hi, respectively.
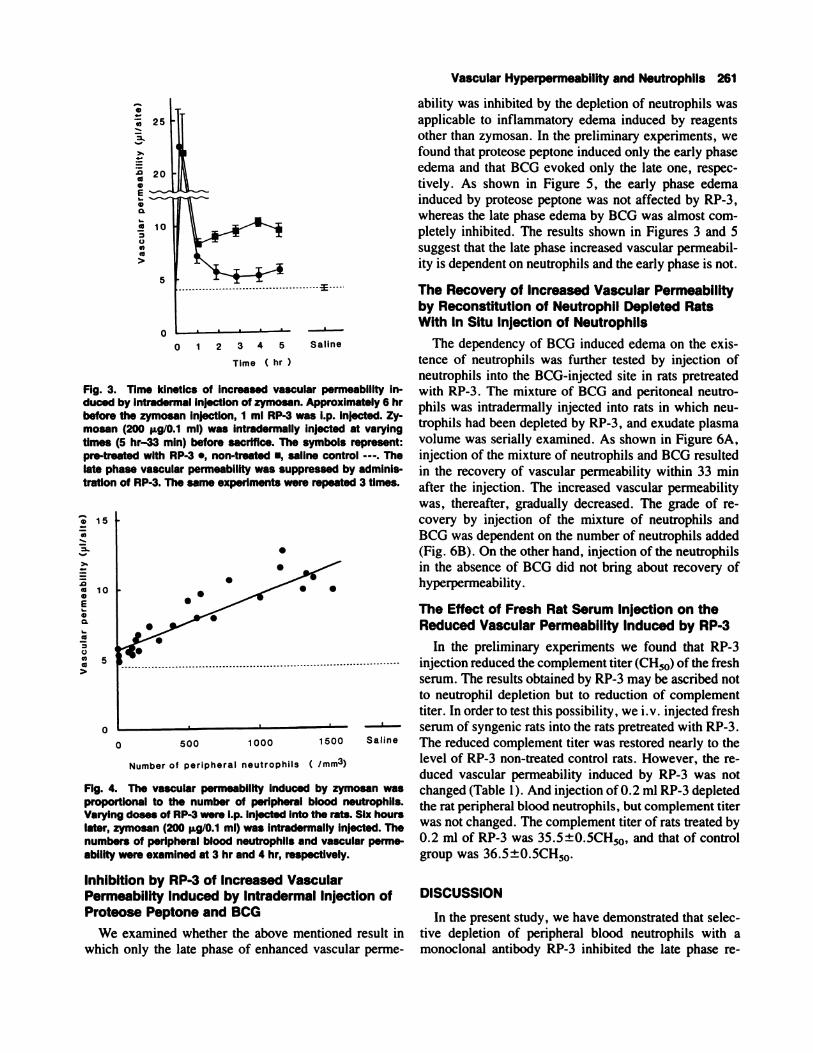
Inhibition by RP-3 of Increased VascularPermeability Induced by Intradermal Injection ofProteose Peptone and BCG
We examined whether the above mentioned result in
which only the late phase of enhanced vascular perme-
Vascular Hyperpermeability and Neutrophils 261
ability was inhibited by the depletion of neutrophils was
applicable to inflammatory edema induced by reagents
other than zymosan. In the preliminary experiments, we
found that proteose peptone induced only the early phase
edema and that BCG evoked only the late one, respec-
tively. As shown in Figure 5, the early phase edema
induced by proteose peptone was not affected by RP-3,
whereas the late phase edema by BCG was almost com-
pletely inhibited. The results shown in Figures 3 and 5
suggest that the late phase increased vascular permeabil-
ity is dependent on neutrophils and the early phase is not.
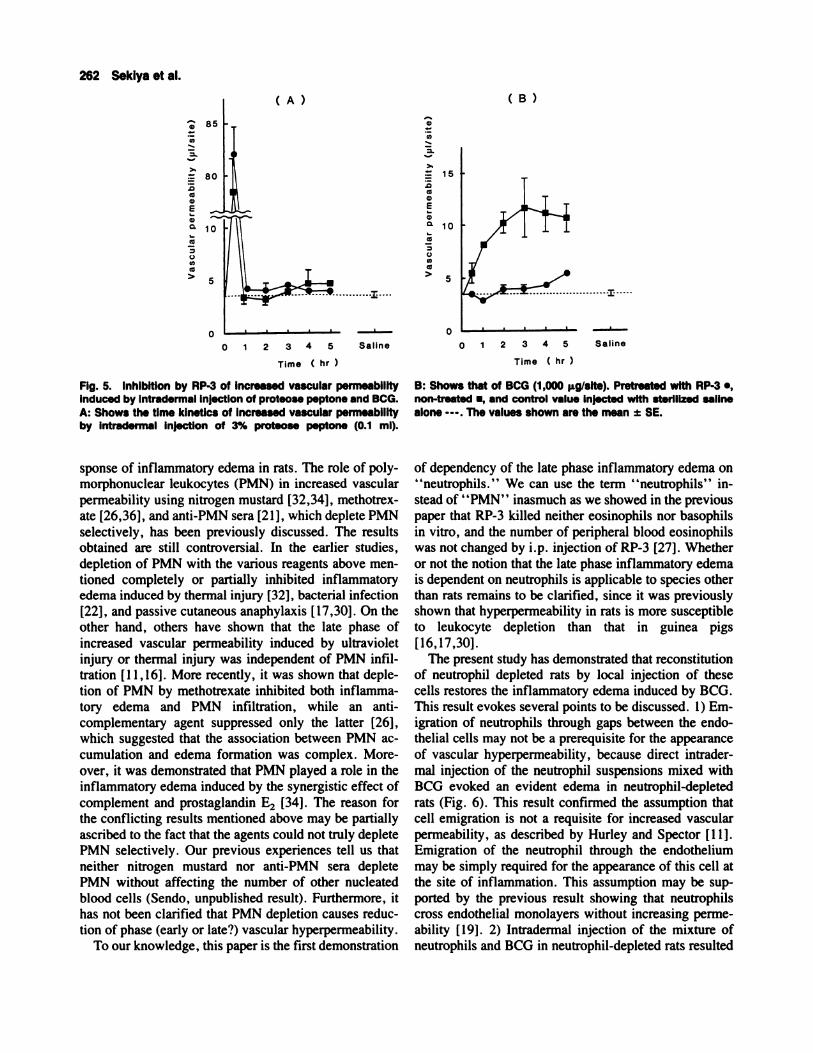
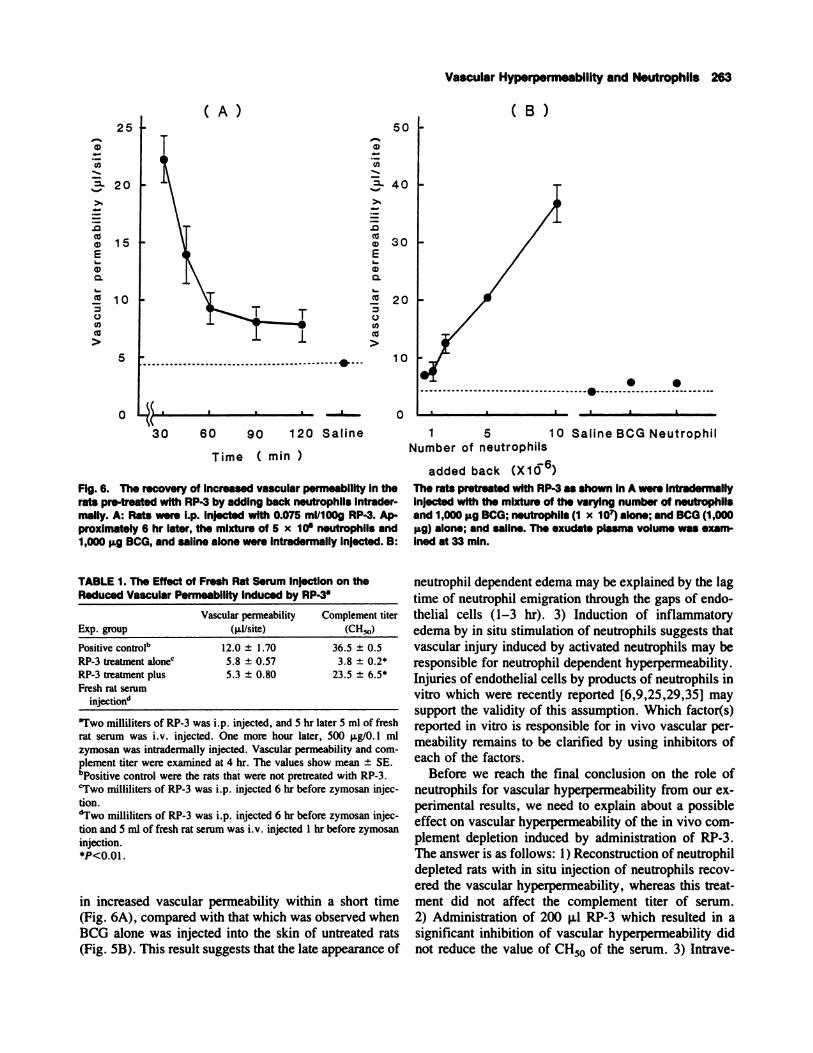
The Recovery of Increased Vascular Permeabilityby Reconstitution of Neutrophil Depleted RatsWith In Situ Injection of Neutrophils
The dependency of BCG induced edema on the exis-
tence of neutrophils was further tested by injection of
neutrophils into the BCG-injected site in rats pretreated
with RP-3. The mixture of BCG and peritoneal neutro-
phils was intradermally injected into rats in which neu-
trophils had been depleted by RP-3, and exudate plasma
volume was serially examined. As shown in Figure 6A,
injection of the mixture of neutrophils and BCG resulted
in the recovery of vascular permeability within 33 mm
after the injection. The increased vascular permeability
was, thereafter, gradually decreased. The grade of re-
covery by injection of the mixture of neutrophils and
BCG was dependent on the number of neutrophils added
(Fig. 6B). On the other hand, injection of the neutrophils
in the absence of BCG did not bring about recovery of
hyperpermeability.
The Effect of Fresh Rat Serum Injection on theReduced Vascular Permeability Induced by RP-3
In the preliminary experiments we found that RP-3
injection reduced the complement titer (CH�#{216})of the fresh
serum. The results obtained by RP-3 may be ascribed not
to neutrophil depletion but to reduction of complementtiter. In order to test this possibility, we iv. injected fresh
serum of syngenic rats into the rats pretreated with RP-3.
The reduced complement titer was restored nearly to the
level of RP-3 non-treated control rats. However, the re-
duced vascular permeability induced by RP-3 was not
changed (Table 1). And injection of0.2 ml RP-3 depleted
the rat peripheral blood neutrophils, but complement titer
was not changed. The complement titer of rats treated by
0.2 ml of RP-3 was 35.5±0.5CH50, and that of control
group was 36.5±0.5CH��.
DISCUSSION
In the present study, we have demonstrated that selec-
tive depletion of peripheral blood neutrophils with a
monoclonal antibody RP-3 inhibited the late phase re-
(A) (B)
85
U,
0�
.0aa,
Ea,
aa00U,
a>
80
10
0
a,
U,
0�
:� 15
.0aa,
Ea,
a
a00U)
a>
0
Time ( hr
B: Shows that of BCG (1 ,000 p.g/slte). Pretreated with RP-3.,non-treated ., and control value injected with sterilized salinealone - - - . The values shown are the mean ± SE.
0 1 2 3 4 5 Saline
Time ( hr
Fig. 5. InhibItion by RP-3 of Increased vascular permeabilityinduced by Intradermal Injection of proteose peptone and BCG.A: Shows the time kinetics of increased vascular permeabilityby intradermal injection of 3% proteose peptone (0.1 ml).
0 1 2 3 4 5 Saline
262 Sekiya et al.
5
sponse of inflammatory edema in rats. The role of poly-
morphonuclear leukocytes (PMN) in increased vascular
permeability using nitrogen mustard [32,34], methotrex-
ate [26,36], and anti-PMN sera [21], which deplete PMN
selectively, has been previously discussed. The results
obtained are still controversial. In the earlier studies,
depletion of PMN with the various reagents above men-
tioned completely or partially inhibited inflammatory
edema induced by thermal injury [32], bacterial infection
[22], and passive cutaneous anaphylaxis [17,30]. On the
other hand, others have shown that the late phase of
increased vascular permeability induced by ultraviolet
injury or thermal injury was independent of PMN infil-
tration [I I , 16]. More recently, it was shown that deple-
tion of PMN by methotrexate inhibited both inflamma-
tory edema and PMN infiltration, while an anti-
complementary agent suppressed only the latter [26],
which suggested that the association between PMN ac-
cumulation and edema formation was complex. More-
over, it was demonstrated that PMN played a role in the
inflammatory edema induced by the synergistic effect of
complement and prostaglandin E2 [34]. The reason for
the conflicting results mentioned above may be partially
ascribed to the fact that the agents could not truly deplete
PMN selectively. Our previous experiences tell us that
neither nitrogen mustard nor anti-PMN sera deplete
PMN without affecting the number of other nucleated
blood cells (Sendo, unpublished result). Furthermore, it
has not been clarified that PMN depletion causes reduc-
tion of phase (early or late?) vascular hyperpermeability.
To our knowledge, this paper is the first demonstration
5
of dependency of the late phase inflammatory edema on
‘ ‘neutrophils. “ We can use the term “neutrophils” in-
stead of “PMN” inasmuch as we showed in the previous
paper that RP-3 killed neither eosinophils nor basophils
in vitro, and the number of peripheral blood eosinophils
was not changed by i.p. injection of RP-3 [27]. Whether
or not the notion that the late phase inflammatory edema
is dependent on neutrophils is applicable to species other
than rats remains to be clarified, since it was previously
shown that hyperpermeability in rats is more susceptible
to leukocyte depletion than that in guinea pigs
[16,17,30].
The present study has demonstrated that reconstitution
of neutrophil depleted rats by local injection of these
cells restores the inflammatory edema induced by BCG.
This result evokes several points to be discussed. 1) Em-igration of neutrophils through gaps between the endo-
thelial cells may not be a prerequisite for the appearance
of vascular hyperpermeability, because direct intrader-
mal injection of the neutrophil suspensions mixed with
BCG evoked an evident edema in neutrophil-depleted
rats (Fig. 6). This result confirmed the assumption that
cell emigration is not a requisite for increased vascular
permeability, as described by Hurley and Spector [1 1].Emigration of the neutrophil through the endothelium
may be simply required for the appearance of this cell at
the site of inflammation. This assumption may be sup-
ported by the previous result showing that neutrophils
cross endothelial monolayers without increasing perme-
ability [ 19] . 2) Intradermal injection of the mixture ofneutrophils and BCG in neutrophil-depleted rats resulted
(A) (B)
U)
>C
(5C,
EC,
0.
CO
0(I)
CO
>
25
20
15
10
5
0
50
C,
C’)
�- 40
>,
.0
� 30EC,
0.
.� 20
0U)(5
>
.... 10
-�---1-cc30
Vascular Hyperpermeability and Neutrophils 263
60 90 120 Saline
Time ( mm
Fig. 6. The recovery of Increased vascular permeability in therats pre-treated with RP-3 by adding back neutrophils intrader.mally. A: Rats were i.p. Injected with 0.075 mI/bOg RP-3. Ap-proximately 6 hr later, the mixture of 5 x 10� neutrophils and1,000 � BCG, and saline alone were Intradermally Injected. B:
TABLE 1 . The Effect of Fresh Rat Serum Injection on theReduced Vascular PermeabIlity Induced by RP-3
Exp. group
Vascular permeability
(p.1/site)
Complement titer
(CH�0)
Positive control” 12.0 ± 1.70 36.5 ± 0.5
RP-3 treatment alonec 5.8 ± 0.57 3.8 ± 0.2*
RP-3 treatment plus 5.3 ± 0.80 23.5 ± 6.5*
Fresh rat seruminjection”
aTwo milliliters of RP-3 was i.p. injected, and 5 hr later 5 ml of fresh
rat serum was iv. injected. One more hour later, 500 p.g/0. 1 ml
zymosan was intradermally injected. Vascular permeability and com-plement titer were examined at 4 hr. The values show mean ± SE.bPositive control were the rats that were not pretreated with RP-3.cTwo milliliters of RP-3 was i.p. injected 6 hr before zymosan injec-
tion.“Two milliliters of RP-3 was i.p. injected 6 hr before zymosan injec-
tion and 5 ml of fresh rat serum was iv. injected 1 hr before zymosan
injection.
*P<0.0l.
in increased vascular permeability within a short time
(Fig. 6A), compared with that which was observed when
BCG alone was injected into the skin of untreated rats
(Fig. SB). This result suggests that the late appearance of
. I
I
C C I I � I
1 5 1 0 Saline BCG Neutrophil
Number of neutrophils
added back (X166)
The rats pretreated with RP-3 as shown in A were Intradermallyinjected with the mixture of the varying number of neutrophilsand 1,000 � BCG; neutrophils (1 x 10w) alone; and BCG (1,000
ILg) alone; and saline. The exudate plasma volume was exam-
med at 33 mm.
neutrophil dependent edema may be explained by the lag
time of neutrophil emigration through the gaps of endo-
thelial cells ( 1-3 hr). 3) Induction of inflammatory
edema by in situ stimulation of neutrophils suggests that
vascular injury induced by activated neutrophils may be
responsible for neutrophil dependent hyperpermeability.
Injuries of endothelial cells by products of neutrophils in
vitro which were recently reported [6,9,25,29,35] may
support the validity of this assumption. Which factor(s)
reported in vitro is responsible for in vivo vascular per-
meability remains to be clarified by using inhibitors of
each of the factors.
Before we reach the final conclusion on the role of
neutrophils for vascular hyperpermeability from our ex-perimental results, we need to explain about a possible
effect on vascular hyperpermeability of the in vivo com-
plement depletion induced by administration of RP-3.
The answer is as follows: 1 ) Reconstruction of neutrophil
depleted rats with in situ injection of neutrophils recov-
ered the vascular hyperpermeability, whereas this treat-
ment did not affect the complement titer of serum.
2) Administration of 200 pA RP-3 which resulted in a
significant inhibition of vascular hyperpermeability did
not reduce the value of CH50 of the serum. 3) Intrave-

264 Seklya et al.
nous injection of fresh serum from syngeneic rats intoWKA rats whose serum complement titer was reduced
by RP-3 treatment restored the value of CH50 of the
serum. However, this treatment did not restore the sup-
pressed vascular hyperpermeability induced by RP-3 in-
jected as shown in the result. These results indicate that
complement depletion induced by RP-3 injection is not
responsible for the decreased vascular hyperpermeability
observed in our experiments.
Finally, we have demonstrated that dependence on the
neutrophils of inflammatory edema is restricted to the
late phase one. The dependence of the late phase vascu-
lar permeability on PMN was also described earlier
[ 1S ,32]. However, it was not shown whether the deple-
tion of neutrophils affects the early response of hyper-
permeability. Here, we have extended previous results
and added a novel result which shows no effect on the
early phase edema of neutrophil depletion.
It has been postulated that inflammatory edema, de-
pendent on neutrophils, may be pathogenic in certain
diseases such as adult respiratory distress syndrome
[20,33]. The experiments using rats, of which neutro-phils are selectively depleted by RP-3, may be helpful
for analysis of the pathogenesis of these diseases. As a
next step the use of other neutrophil stimuli such as CSa,
platelet activating factor, and NAP- 1, a novel chemo-
tactic peptide [5], may be required to ensure that our
conclusion is applicable to vascular hyperpermeability in
general.
REFERENCES
I . Babior, B.M. Oxygen-dependent microbial killing by phago-
cytes. N. EngI. J. Med. 298,659, 1978.2. Borgeat, P. , and Samuelsson, B. Arachidonic acid metabolism
in polymorphonuclear leukocyte. Effect of ionophore A23 187.Proc. Natl. Acad. Sci. USA. 76,2148, 1979.
3. Burke J.F. , and Miles, A.A. The sequence of vascular events inearly infective inflammation. J. Path. Bact. 76, 1 , 1958.
4. Cochrane, C.G. , and Aikin, B.S. Polymorphonuclear leuko-cytes in immunologic reactions. The destruction of vascular
basement membrane in vitro and in vivo. J. Exp. Med. 124,733,1966.
5. Colditz. 1. , Zwahlen, R. , Dewald, B. , and Baggiolini M. Invivo inflammatory activity of neutrophil-activating factor, a
novel chemotactic peptide derived from human monocytes. Am.J. Pathol. 134,755, 1989.
6. Diener, AM. , Beatty, PG. , Ochs, H.D. , and Harlan, J.M. Therole of neutrophil membrane glycoprotein 150 (GP-l50) in neu-
trophil-mediated endothelial cell injury in vitro. J. lmmunol.135,537, 1985.
7. Gotoh, F. , Nakamura, S. , Gotoh, K. , and Yoshinaga, M. Pro-duction of a lymphocyte proliferation promoting factor by puri-fled polymorphonuclear leukocytes from mice and rabbits. Im-
munology 53,683, 1984.
8. Gotoh, S., Itoh, M., Fujii, Y., Arai, S., and Sendo, F. En-hancement of the expression of a rat neutrophil-speciflc cell
surface antigen by activation with phorbol myristate acetate and
concanavalin A. J. Immunol. 137,643, 1986.9. Harlan, J.M. , Killen, PD. , Harker, L.A. , and Striker, G.E.
Neutrophil-mediated endothelial injury in vitro. Mechanisms ofcell detachment. J. Clin. Invest. 68,1394, 1981.
10. Hellewell, PG. , and Williams, T.J. A specific antagonist ofplatelet-activating factor suppresses oedema formation in anArthus reaction but not oedema induced by leukocyte chemoat-
tractants in rabbit skin. J. Immunol. 137,302, 1986.I I . Hurley, J.V. , and Spector, W.G. Delayed leucocytic emigration
after intradermal injections and thermal injury. J. Path. Bat.
82,421, 1961.
12. Inoue, T. , and Sendo, F. In vitro induction of cytotoxic poly-
morphonuclear leukocytes by supernatant from a concanavalinA-stimulated spleen cell culture. J. Immunol. 131 ,2508, 1983.
13. Issekutz, AC. , Movat, K.W. , and Movat, HZ. Enhanced vas-cular permeability and haemorrhage-inducing activity of rabbitC5a,, ,,,: probable role of polymorphonuclear leukocyte lyso-somes. Clin. Exp. Immunol. 41,512, 1980.
14. Kearney, J.F. , Radbruch, A. , Liesegang, B. , and Rajewsky, K.A new mouse myeloma cell line that has lost immunoglobulin
expression but permits the constitution of antibody-secreting hy-brid cell lines. J. Immunol. 123,1548, 1979.
15. Kopaniak, MM. , and Movat, HZ. Kinetics of acute inflam-mation induced by Escherichia coli in rabbits. II. The effect of
hyperimmunization, complement depletion, and depletion ofleukocytes. Am. J. Pathol. 110,13, 1983.
16. Logan, G. , and Wilhelm, DL. The inflammatory reaction in
ultraviolet injury. Brit. J. Exp. Pathol. 47,286, 1966.
17. Lovett, CA. , and Movat, HZ. Role of PMN-leukocyte lyso-somes in tissue injury, inflammation and hypersensitivity. III.Passive cutaneous anaphylaxis induced in the rat with homolo-
gous and heterologous hyperimmune antibody. Proc. Soc. Exp.Biol. Med. 122,991, 1966.
18. Lynch, J.M. , Lotner, G.Z. , Betz, S.J., and Henson, P.M. Therelease of a platelet-activating factor by stimulated rabbit neu-
trophils.J. Immunol. 123,1219, 1979.
19. Meyrick, B. , Hoffman, L.H. , and Brigham, K.L. Chemotaxisof granulocytes across bovine pulmonary artery intimal explants
without endothelial cell injury. Tissue Cell 16,1, 1984.
20. Michael, R.F. , Azriel, P. , and Norman, S.C. Leukocytes arerequired for increased lung microvascular permeability after mi-croembolization in sheep. Circ. Res. 48,344, 1981.
21 . Movat, HZ. Inflammation, Immunity and Hypersensitivity.
Cellular and molecular mechanisms, second edition. Harper andRow, Publisher, Inc. , p. 38, 1979.
22. Movat, HZ. , and Uriuhara, T. Acute inflammation. IV. On themediation of the late phase of increased vascular permeability.
Lab. Invest. 15,1136, 1966.
23. Muller-Eberharb, H.J. , Hoffman, L.G. , and Mayer, MM.
Methods in Immunology and Immunochemistry. AcademicPress, Inc. 4,127, 1977.
24. Oi, VT. , and Herzenberg, J.A. Immunoglobulin-producing hy-brid cell lines. In Selected Methods in Cellular Immunology,(Mishell, B.B., and Shigi, SM. Eds.). San Francisco: W.H.
FreemanCo., p. 351, 1980.
25. Sacks, T. , Moldow, CF. , Craddock, P.R. , Bowers, T.K. , and
Jacob, H.S. Oxygen radicals mediated endothelial cell damageby complement-stimulated granulocytes. An in vitro model of
immune vasculardamage. J. Clin. Invest. 61,1161, 1978.
26. Sedgwick, AD. , Moore, AR. , Al-Duaij, A. , and Willoughby,D.A. Studies into the association between leucocyte accumula-
tion and oedema formation. Agents Actions 17,209, 1985.
27. Sekiya, S. , Gotoh, S. , Yamashita, T. , Watanabe, T. , Saitoh, S.,

and Sendo, F. Selective depletion of rat neutrophils by in vivo
administration of a monoclonal antibody. J. Leukocyte Biol.46,96, 1989.
28. Yamashita, T. , Watanabe, T. , Hosaka, Y. , Asahi, H. , Minai,
M. , Saito, S. , and Sendo, F. Immunological studies on Schis-
tosomiasis japonica using hybridoma technology (1) Character-ization of two monoclonal antibodies obtained from Schistosoma
japonicum infected mice , reactive with a Schistosoma japonicum
egg antigen or a heterophil antigen. Jpn. J. Parasitol. 38,307,1989.
29. Smedly, L.A. , Tonnesen, MG. , Sandhaus, R.A. , Haslett, C.,
Guth rie, L.A. , Johnston, RB. , Jr. , Henson, P.M. , andWorthen, G.S. Neutrophil-mediated injury to endothelial cells.
Enhancement by endotoxin and essential role of neutrophil
elastase. J. Clin. Invest. 77,1233, 1986.30. Taichman, N.S. , and Movat, HZ. Do polymorphonuclear leu-
kocytes play a role in passive cutaneous anaphylaxis of guineapig? Int. Arch. Allergy Appl. Immunol. 30,97, 1966.
31. Tiku, K., Tiku, ML., and Skosey, J.L. Interleukin 1 production
Vascular Hyperpermeability and Neutrophils 265
by human polymorphonuclear neutrophils. J. Immunol.
136,3677, 1986.
32. Uriuhara, T. , and Movat, HZ. Role of PMN-leukocyte lyso-somes in tissue injury, inflammation and hypersensitivity. V.
Partial suppression in leukopenic rabbits of vascular hyperper-meability due to thermal injury. Proc. Soc. Exp. Biol. Med.
124,279, 1967.
33. Powe, J.E. , Short, A. , Sibbald, W.J. , and Driedger, A.A. Pul-
monary accumulation of polymorphonuclear leukocytes in the
adult respiratory distress syndrome. Crit. Care Med. 10,712,1982.
34. Wedmore, CV. , and Williams, T.J. Control of vascular perme-ability by polymorphonuclear leukocytes in inflammation. Na-
ture 289,646, 1981.35. Weiss, S.J. , Young, J. , LoBuglio, A.F. , Slivka, A. , and Nimeh
N.F. Role of hydrogen peroxide in neutrophil-mediated destruc-tion of cultured endothelial cells. J. Clin. Invest. 68,714, 1981.
36. Willoughby, D.A. , and Spector, W.G. Inflammation in agran-ulocytotic rats. Nature 219,1258, 1966.
Related Documents