advances.sciencemag.org/cgi/content/full/7/11/eabf2704/DC1 Supplementary Materials for Small-molecule mimicry hunting strategy in the imperial cone snail, Conus imperialis Joshua P. Torres, Zhenjian Lin*, Maren Watkins, Paula Flórez Salcedo, Robert P. Baskin, Shireen Elhabian, Helena Safavi-Hemami, Dylan Taylor, Jortan Tun, Gisela P. Concepcion, Noel Saguil, Angel A. Yanagihara, Yixin Fang, Jeffrey R. McArthur, Han-Shen Tae, Rocio K. Finol-Urdaneta, B. Duygu Özpolat, Baldomero M. Olivera, Eric W. Schmidt* *Corresponding author. Email: [email protected] (E.W.S.); [email protected] (Z.L.) Published 12 March 2021, Sci. Adv. 7, eabf2704 (2021) DOI: 10.1126/sciadv.abf2704 The PDF file includes: Structure elucidation of conazoliums Tables S1 to S5 Figs. S1 to S6 Legends for movies S1 to S7 References Other Supplementary Material for this manuscript includes the following: (available at advances.sciencemag.org/cgi/content/full/7/11/eabf2704/DC1) Movies S1 to S7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

advances.sciencemag.org/cgi/content/full/7/11/eabf2704/DC1
Supplementary Materials for
Small-molecule mimicry hunting strategy in the imperial cone snail,
Conus imperialis
Joshua P. Torres, Zhenjian Lin*, Maren Watkins, Paula Flórez Salcedo, Robert P. Baskin, Shireen Elhabian, Helena Safavi-Hemami, Dylan Taylor, Jortan Tun, Gisela P. Concepcion, Noel Saguil, Angel A. Yanagihara, Yixin Fang,
Jeffrey R. McArthur, Han-Shen Tae, Rocio K. Finol-Urdaneta, B. Duygu Özpolat, Baldomero M. Olivera, Eric W. Schmidt*
*Corresponding author. Email: [email protected] (E.W.S.); [email protected] (Z.L.)
Published 12 March 2021, Sci. Adv. 7, eabf2704 (2021)
DOI: 10.1126/sciadv.abf2704
The PDF file includes:
Structure elucidation of conazoliums Tables S1 to S5 Figs. S1 to S6 Legends for movies S1 to S7 References
Other Supplementary Material for this manuscript includes the following: (available at advances.sciencemag.org/cgi/content/full/7/11/eabf2704/DC1)
Movies S1 to S7

Structure elucidation of conazoliums Conazolium A (1) was isolated as its TFA salt. A molecular formula C16H25N6O4S+ was assigned based upon a molecular ion at m/z 397.1643 and the 1H and 13C NMR spectra (see Table S2 and Fig. S1). The 1H NMR spectrum in D2O showed signals of 22 protons, including 18 singlet methyl protons, two methylene protons, a methine proton, and an aromatic proton. The 13C NMR spectrum showed 16 carbon signals, including seven quaternary carbons. Analysis of HMBC and COSY correlations revealed the presence of a highly methylated histidinyl moiety in the molecule. The HMBC correlation from the protons H3-10 (9×H) to their methyl carbon indicated a trimethylammonium group. The 13C chemical shift of the trimethylammonium group was consistent with previous data for this functional group (62). HMBC correlations from the histidinyl α-proton H-2 to C-1 and C-10, and from H-10 to C-2, indicated the position of the trimethylammonium group. HMBC correlations from H-6 to C-4 and C-8, H3-11 to C-4 and C-8, and H3-12 to C-6 and C-8 revealed an N,N-dimethyl imidazole unit, which based upon HMBC correlations between H-3 and C-4, C-8, and N-5 indicated the presence of a pentamethylated histidine derivative. Moreover, the 13C chemical shift of quaternary carbon C-8 (δ 128.3 ppm) was reminiscent of that for ovothiol, which is thiol substituted in the same position. The histidine portion of 1 was thus assigned as shown. What remained to assign was a molecular formula of C5H6N3O2, in which all of the carbons were quaternary except for a single methyl group. These methyl protons at H3-18 had three HMBC correlations to C-14, C-17 and N-13, suggesting that C-17 and C-14 were fully substituted and directly connected to an N-methyl group. Additionally, the C5H6N3O2moiety contributed a UV λmax of 269 nm. Taken together, these findings resulted in a tentative proposal that this half of the molecule consisted of 6-amino-1-methyluracil (AmMU). Based upon the molecular formula and all available data, the histidinyl and AmMU moieties should be connected via a thioether bridge from C-15 to C-8, but an absence of any correlations across this bond made the elucidation challenging. AmMU has been previously reported as a synthetic compound, although not as a natural product or a portion of a natural product, to the best of our knowledge. Moreover, thioether derivatives of AmMU coupled to aromatic groups have been synthesized and characterized, providing a good model system for 1. The chemical shift of C-15 (δ 77.9) and the relatively low intensity 13C NMR signal was consistent with that of previously reported 5-thioether-uracil model compounds(15) (FigS1). Thus, previous chemical shift data were consistent with the presence of the thioether bridge linking C-8 and C-15. To ensure that we were not constrained by possibly incorrect assumptions, we used MOLGEN(16)to generate all possible structures. Only AmMU fit within these constraints, supporting the structure 1 as shown. Although these data were compelling, the lack of protonated carbons in AmMU made the hypothesis of the AmMU structure, and its link to imidazole via a thioether, uncertain. We thus employed a chemical method for confirmation. Compound 1 was reduced using nickel boride (63). The resulting reaction mixture was analyzed by HPLC in comparison to a commercial standard of AmMU. The standard eluted at an identical time, and with an identical UV spectrum, to the reaction product (Fig. S1), and it was coincident in coelution experiments, confirming the assignment. Finally, the calculated and measured ECD spectra of 1 matched relatively well, leading to assignment of C-2 as S. Conazolium B (2) was also isolated as its TFA salt with a molecular formula C21H31N5O3S+, based on the analysis of HRESIMS and NMR data (TableS2). The NMR data (TableS2, and Fig. S1) of 2 clearly indicated the presence of the same pentamethyl histidine moiety found in 1. The remaining part of the molecule is composed of C10H11N2OS. A 4-substituted serotonin substructure was deduced based upon COSY, HSQC and HMBC data (TableS2 and Fig. S1). The chemical shift of C-8 in 2 was almost identical to that of 1, indicating the same thioether linkage between the histidine moiety and the serotonin moiety. The structure assignment was fully supported by all available spectroscopic data, including a shift in UV spectra between 1 (λmax 269) and 2 (λmax 327), consistent with a change from AmMU to serotonin. Finally, the ECD spectrum in comparison to calculation was consistent with the S-configuration at C-2.
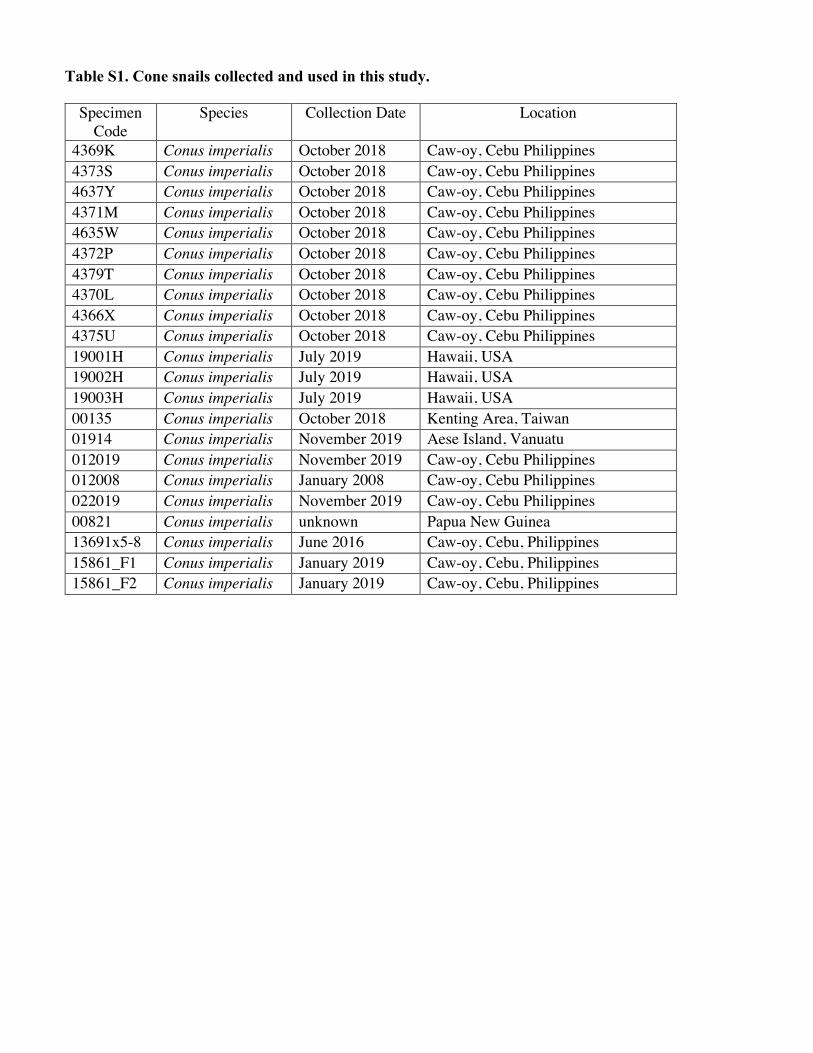
Table S1. Cone snails collected and used in this study.
Specimen Code
Species Collection Date Location
4369K Conus imperialis October 2018 Caw-oy, Cebu Philippines 4373S Conus imperialis October 2018 Caw-oy, Cebu Philippines 4637Y Conus imperialis October 2018 Caw-oy, Cebu Philippines 4371M Conus imperialis October 2018 Caw-oy, Cebu Philippines 4635W Conus imperialis October 2018 Caw-oy, Cebu Philippines 4372P Conus imperialis October 2018 Caw-oy, Cebu Philippines 4379T Conus imperialis October 2018 Caw-oy, Cebu Philippines 4370L Conus imperialis October 2018 Caw-oy, Cebu Philippines 4366X Conus imperialis October 2018 Caw-oy, Cebu Philippines 4375U Conus imperialis October 2018 Caw-oy, Cebu Philippines 19001H Conus imperialis July 2019 Hawaii, USA 19002H Conus imperialis July 2019 Hawaii, USA 19003H Conus imperialis July 2019 Hawaii, USA 00135 Conus imperialis October 2018 Kenting Area, Taiwan 01914 Conus imperialis November 2019 Aese Island, Vanuatu 012019 Conus imperialis November 2019 Caw-oy, Cebu Philippines 012008 Conus imperialis January 2008 Caw-oy, Cebu Philippines 022019 Conus imperialis November 2019 Caw-oy, Cebu Philippines 00821 Conus imperialis unknown Papua New Guinea 13691x5-8 Conus imperialis June 2016 Caw-oy, Cebu, Philippines 15861_F1 Conus imperialis January 2019 Caw-oy, Cebu, Philippines 15861_F2 Conus imperialis January 2019 Caw-oy, Cebu, Philippines
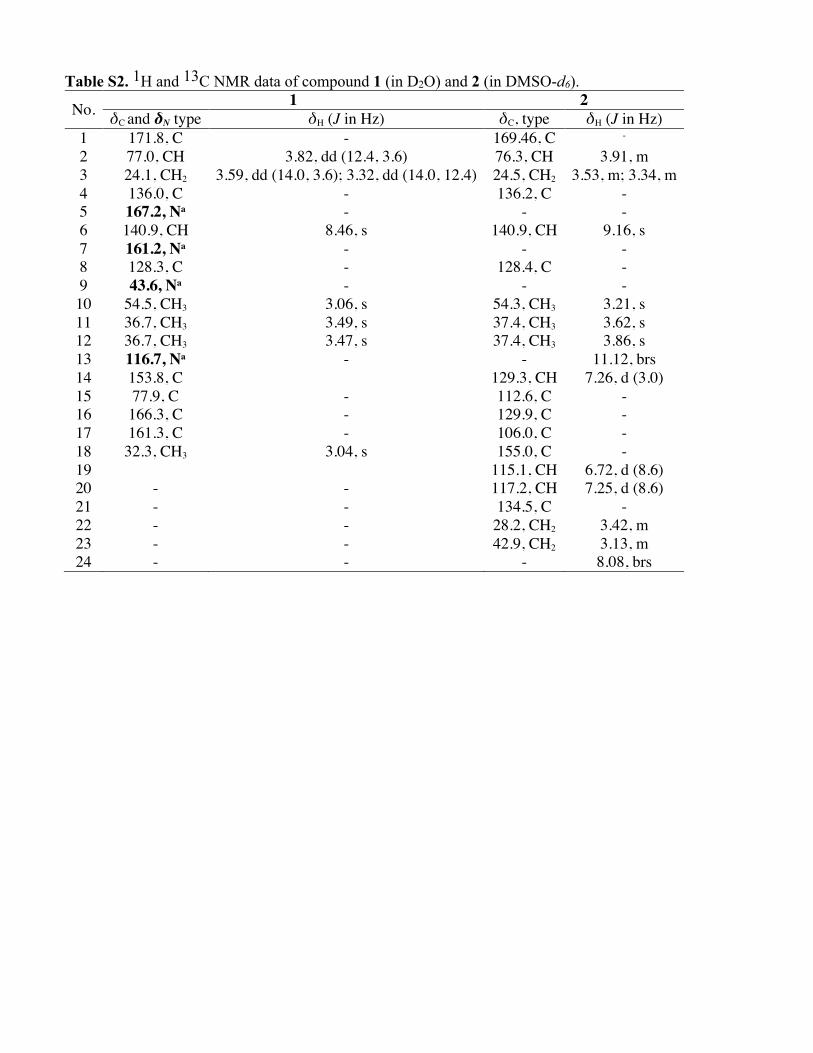
Table S2. 1H and 13C NMR data of compound 1 (in D2O) and 2 (in DMSO-d6).
No. 1 2 δC and δN type δH (J in Hz) δC, type δH (J in Hz)
1 171.8, C - 169.46, C - 2 77.0, CH 3.82, dd (12.4, 3.6) 76.3, CH 3.91, m 3 24.1, CH2 3.59, dd (14.0, 3.6); 3.32, dd (14.0, 12.4) 24.5, CH2 3.53, m; 3.34, m 4 136.0, C - 136.2, C - 5 167.2, Na - - - 6 140.9, CH 8.46, s 140.9, CH 9.16, s 7 161.2, Na - - - 8 128.3, C - 128.4, C - 9 43.6, Na - - - 10 54.5, CH3 3.06, s 54.3, CH3 3.21, s 11 36.7, CH3 3.49, s 37.4, CH3 3.62, s 12 36.7, CH3 3.47, s 37.4, CH3 3.86, s 13 116.7, Na - - 11.12, brs 14 153.8, C 129.3, CH 7.26, d (3.0) 15 77.9, C - 112.6, C - 16 166.3, C - 129.9, C - 17 161.3, C - 106.0, C - 18 32.3, CH3 3.04, s 155.0, C - 19 115.1, CH 6.72, d (8.6) 20 - - 117.2, CH 7.25, d (8.6) 21 - - 134.5, C - 22 - - 28.2, CH2 3.42, m 23 - - 42.9, CH2 3.13, m 24 - - - 8.08, brs
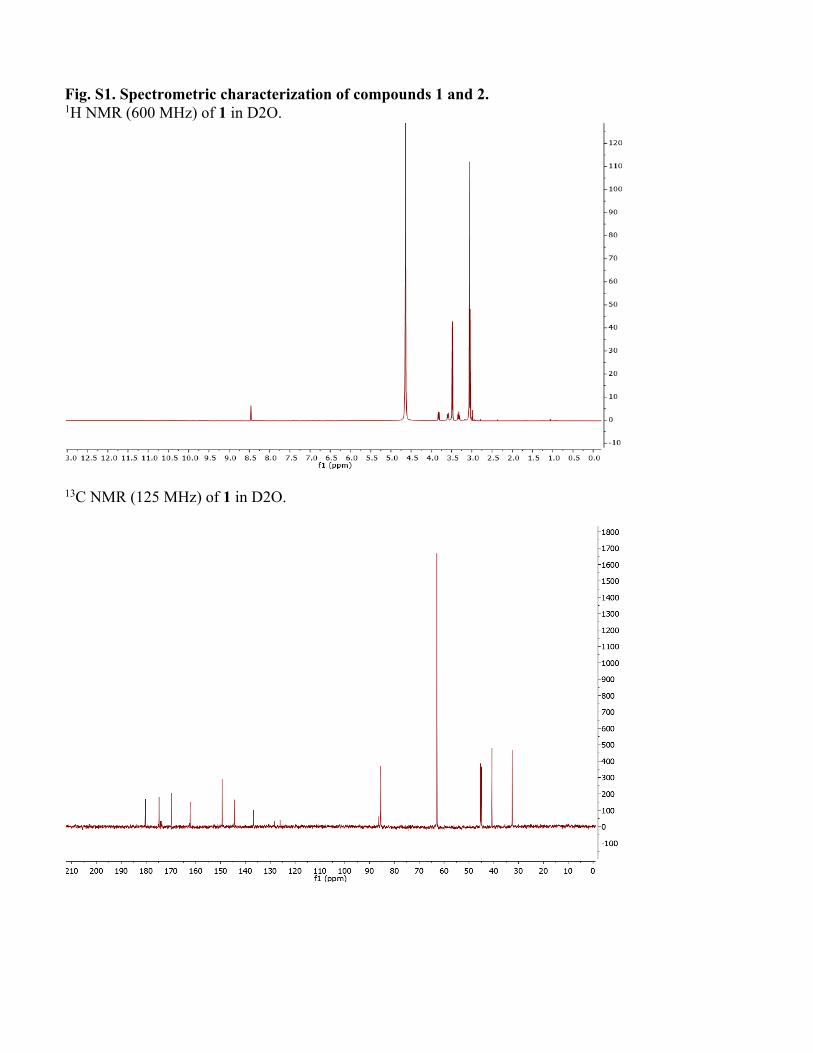
Fig. S1. Spectrometric characterization of compounds 1 and 2. 1H NMR (600 MHz) of 1 in D2O.
13C NMR (125 MHz) of 1 in D2O.
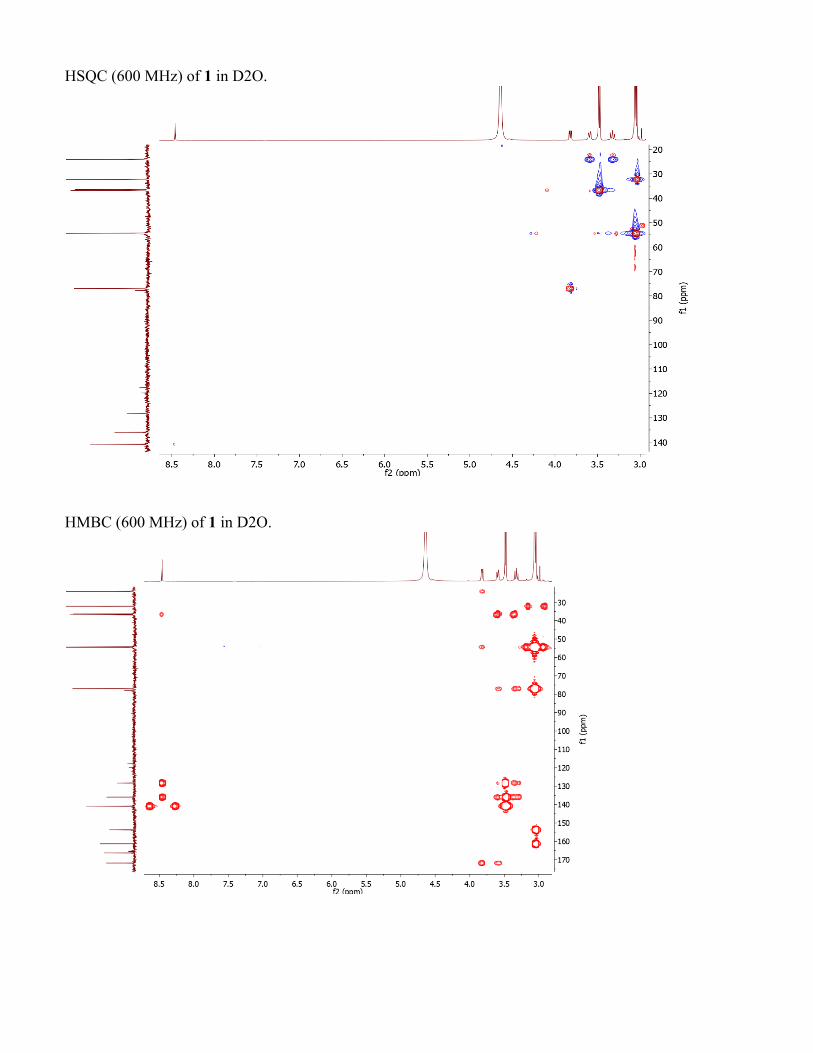
HSQC (600 MHz) of 1 in D2O.
HMBC (600 MHz) of 1 in D2O.
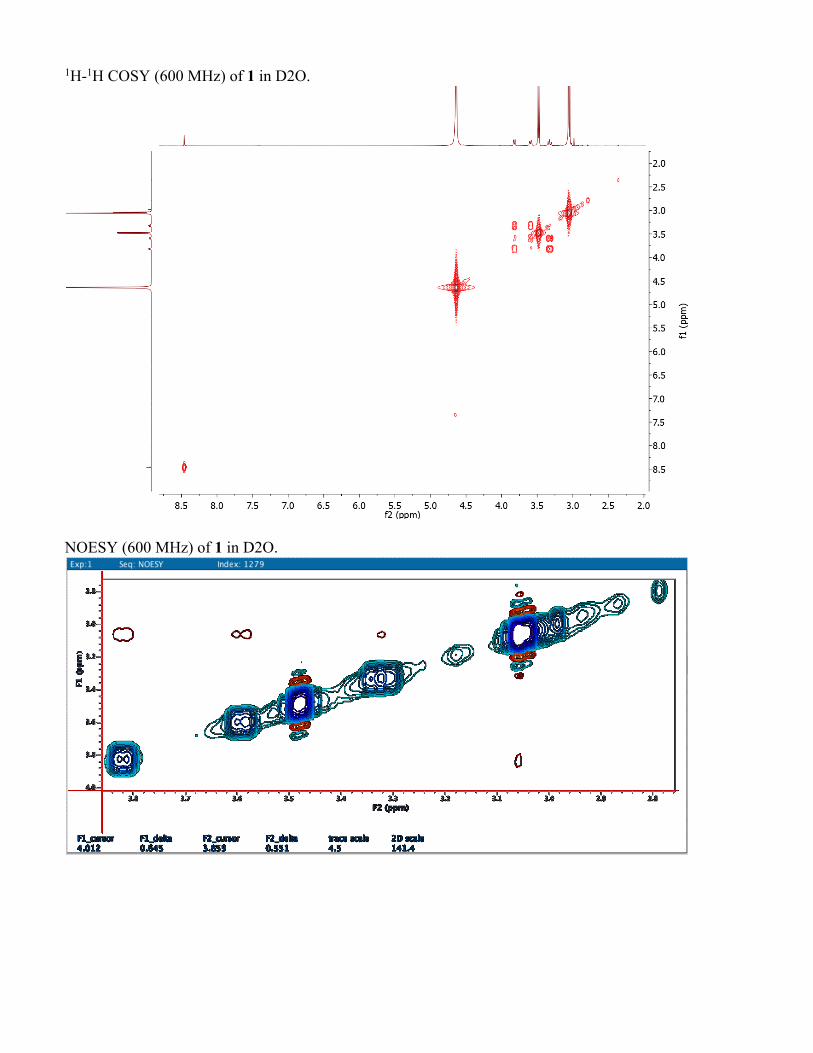
1H-1H COSY (600 MHz) of 1 in D2O.
NOESY (600 MHz) of 1 in D2O.
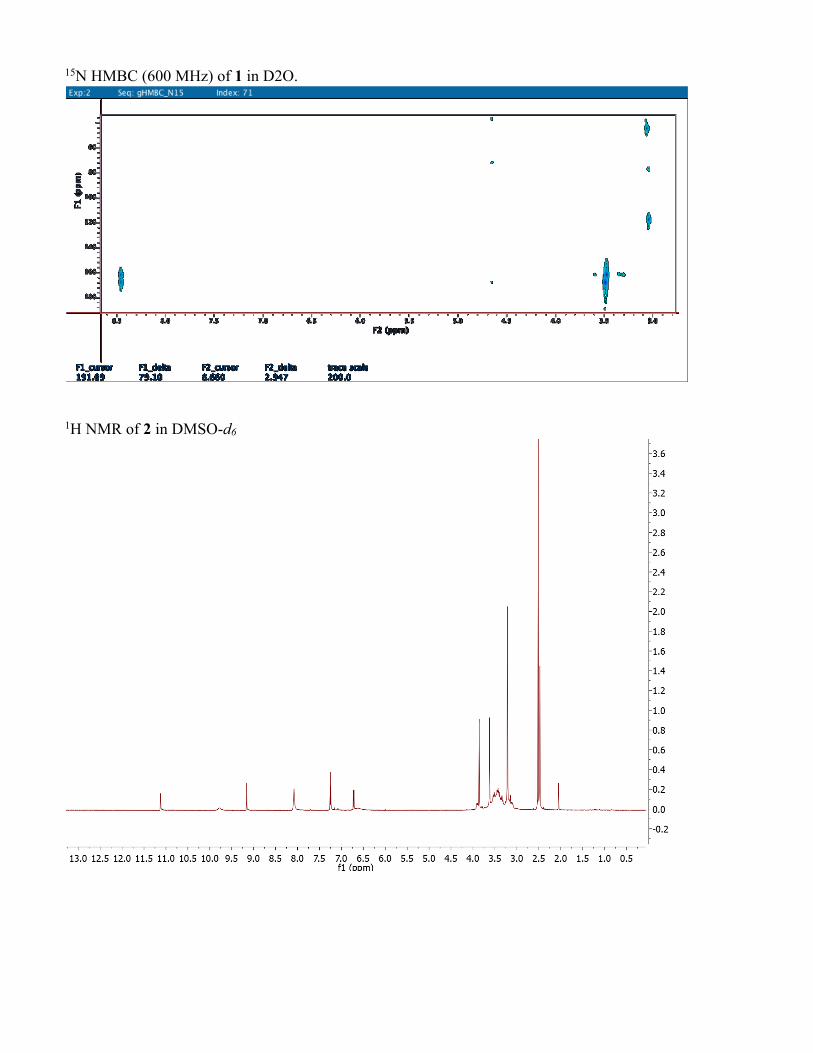
15N HMBC (600 MHz) of 1 in D2O.
1H NMR of 2 in DMSO-d6
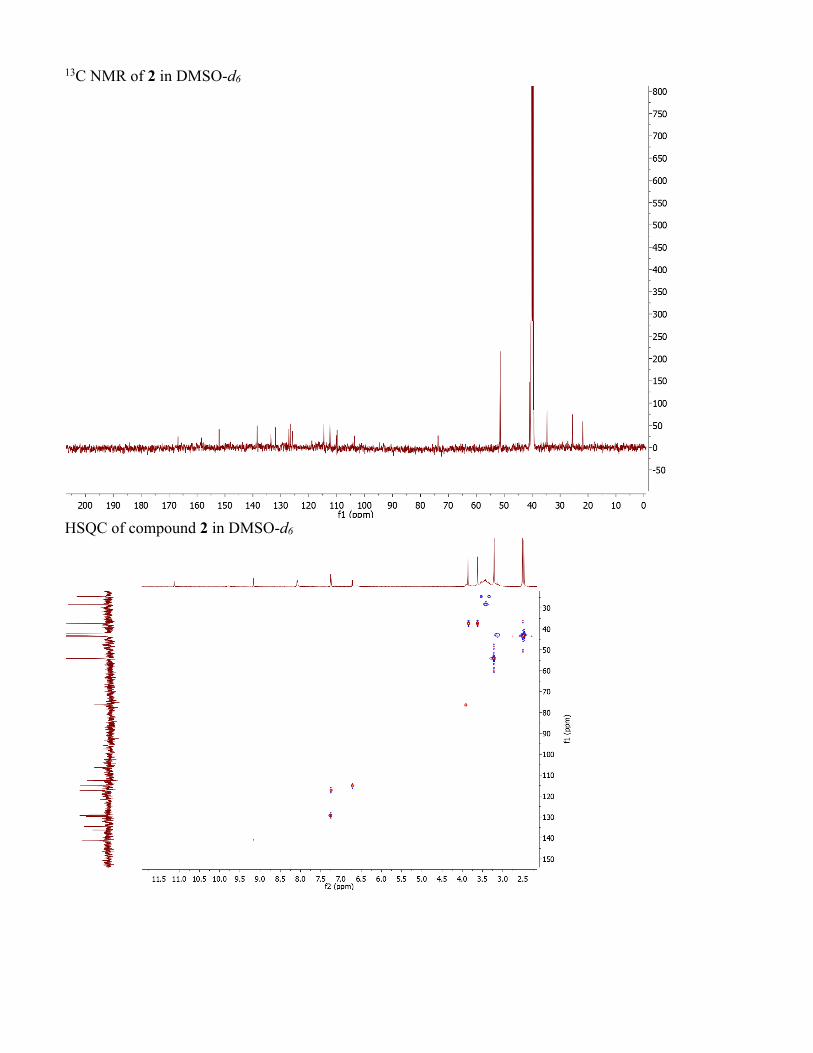
13C NMR of 2 in DMSO-d6
HSQC of compound 2 in DMSO-d6
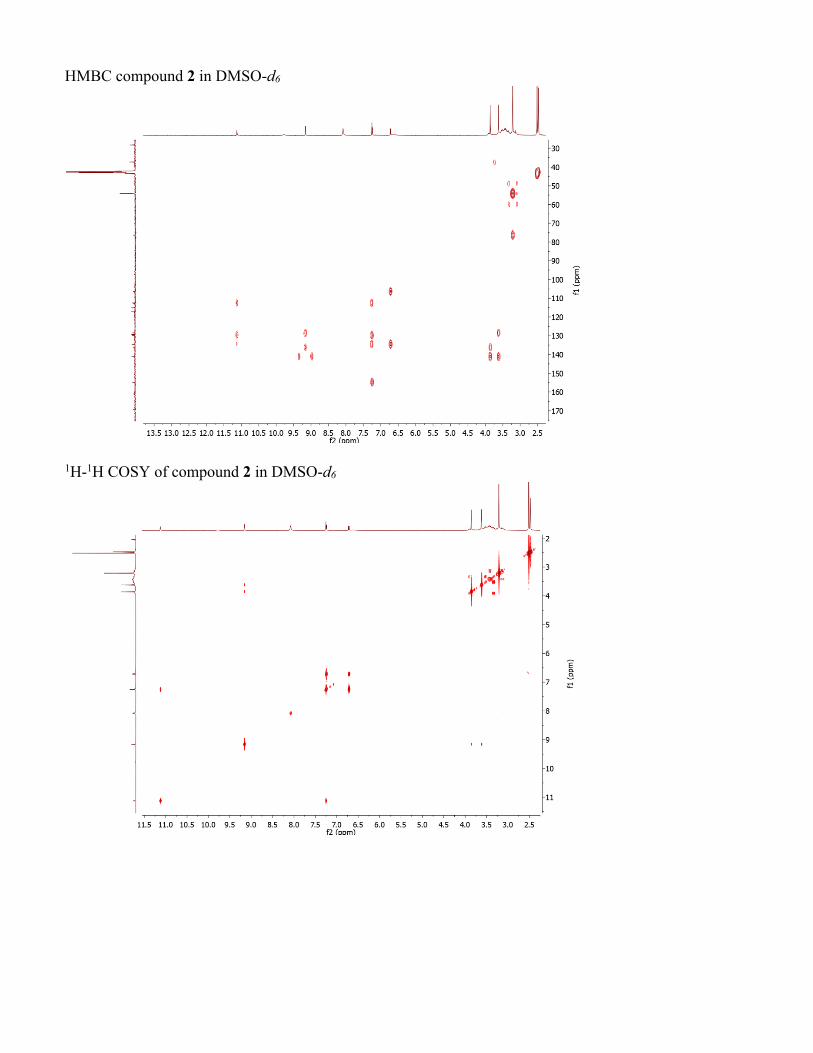
HMBC compound 2 in DMSO-d6
1H-1H COSY of compound 2 in DMSO-d6
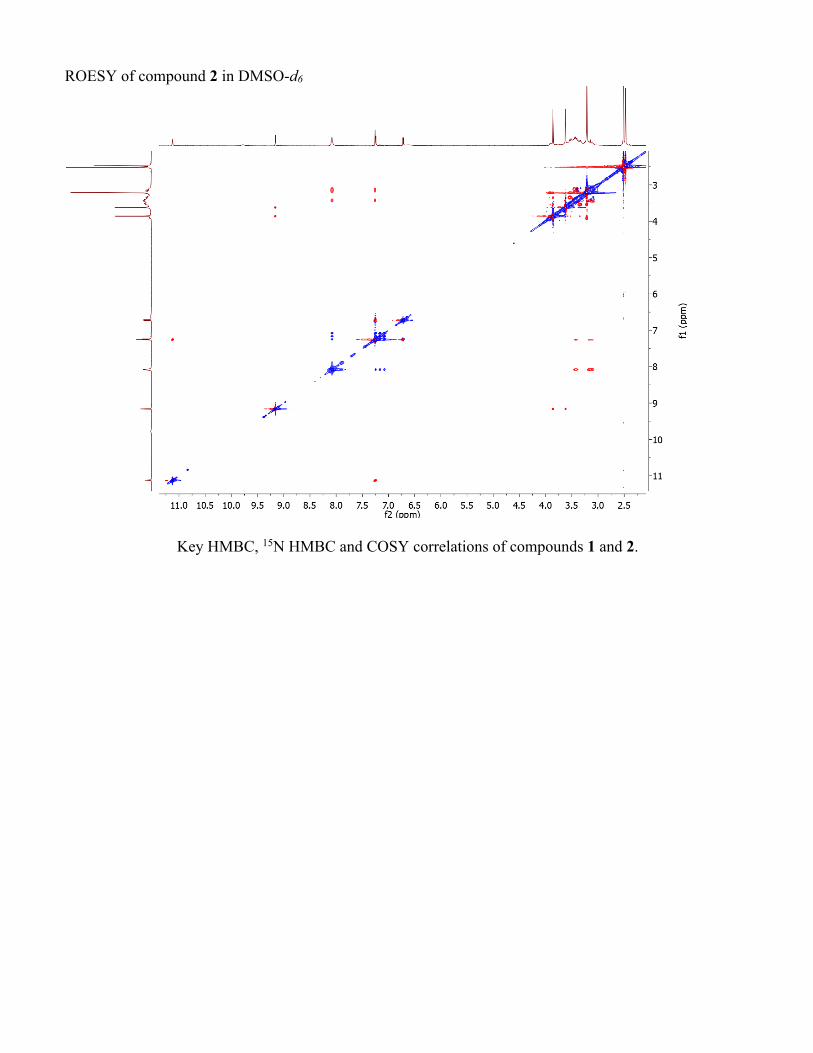
ROESY of compound 2 in DMSO-d6
Key HMBC, 15N HMBC and COSY correlations of compounds 1 and 2.

13C NMR spectra of synthetic 3-thio-uracil derivatives reported previously
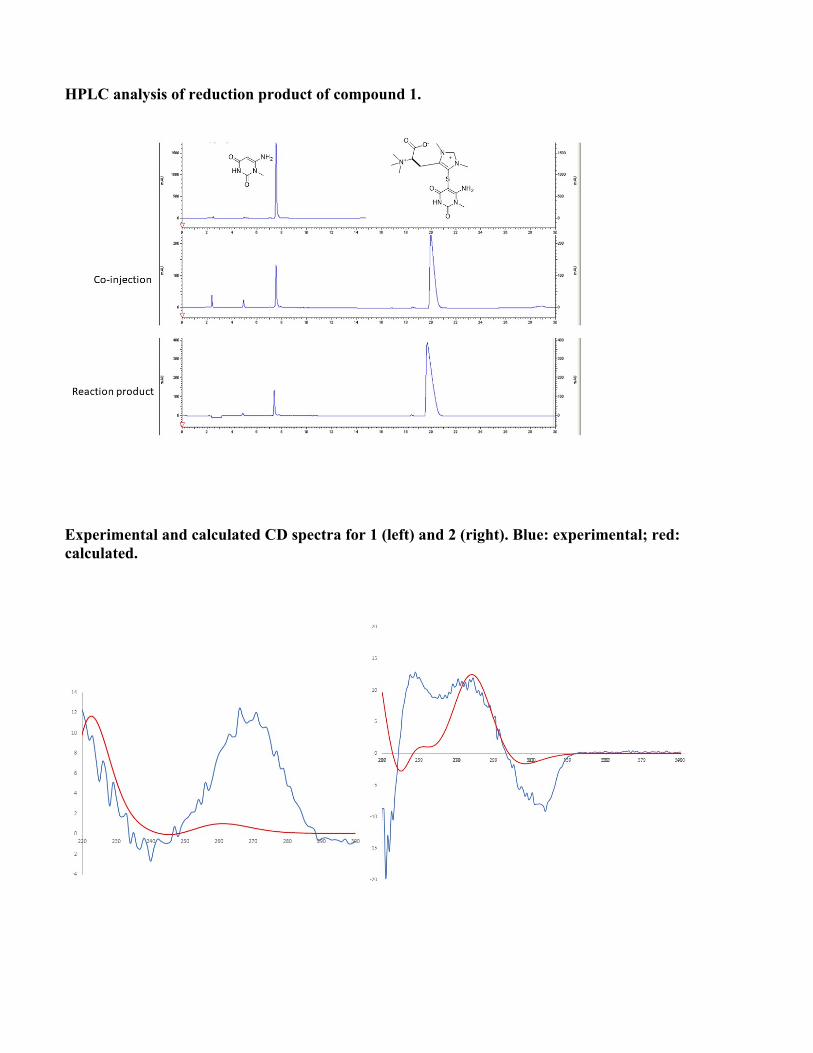
HPLC analysis of reduction product of compound 1.
Experimental and calculated CD spectra for 1 (left) and 2 (right). Blue: experimental; red: calculated.
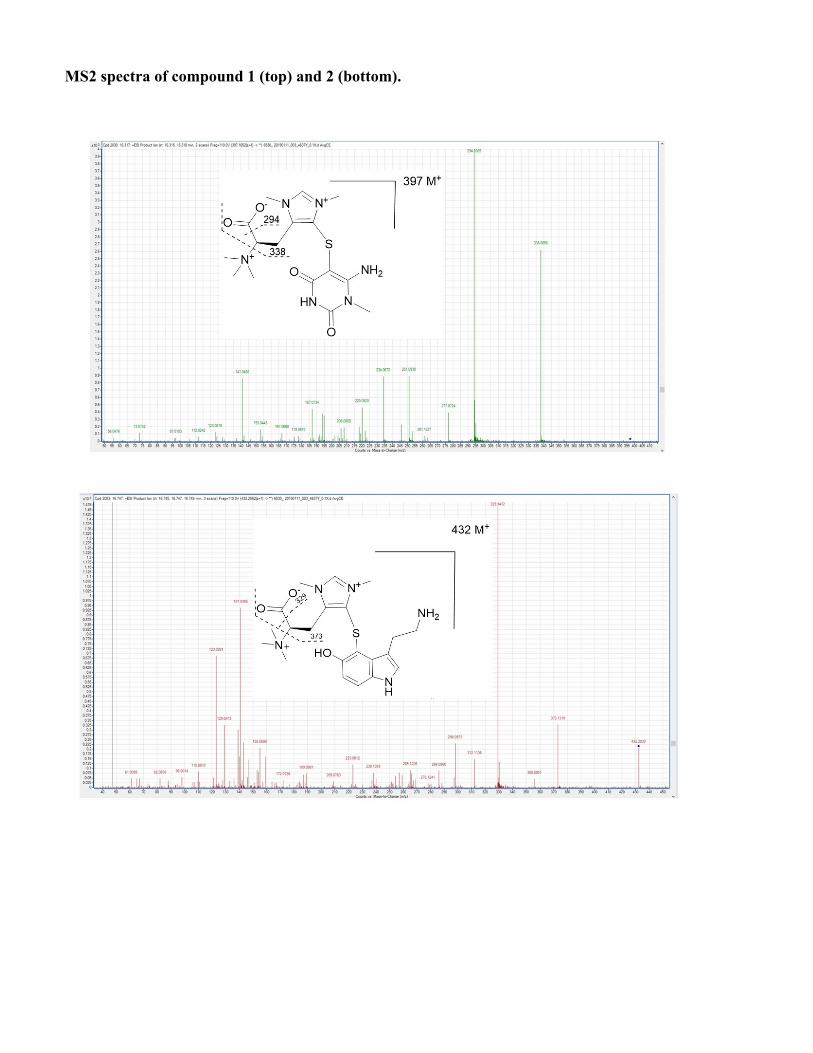
MS2 spectra of compound 1 (top) and 2 (bottom).
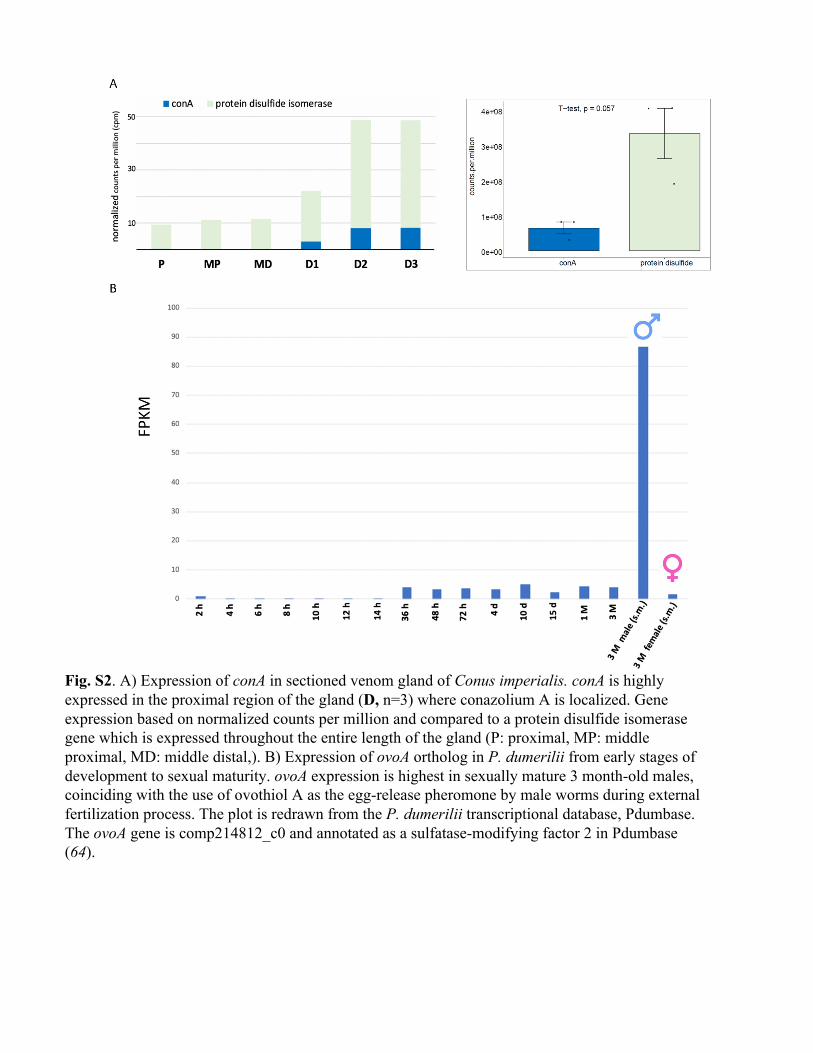
Fig. S2. A) Expression of conA in sectioned venom gland of Conus imperialis. conA is highly expressed in the proximal region of the gland (D, n=3) where conazolium A is localized. Gene expression based on normalized counts per million and compared to a protein disulfide isomerase gene which is expressed throughout the entire length of the gland (P: proximal, MP: middle proximal, MD: middle distal,). B) Expression of ovoA ortholog in P. dumerilii from early stages of development to sexual maturity. ovoA expression is highest in sexually mature 3 month-old males, coinciding with the use of ovothiol A as the egg-release pheromone by male worms during external fertilization process. The plot is redrawn from the P. dumerilii transcriptional database, Pdumbase. The ovoA gene is comp214812_c0 and annotated as a sulfatase-modifying factor 2 in Pdumbase (64).
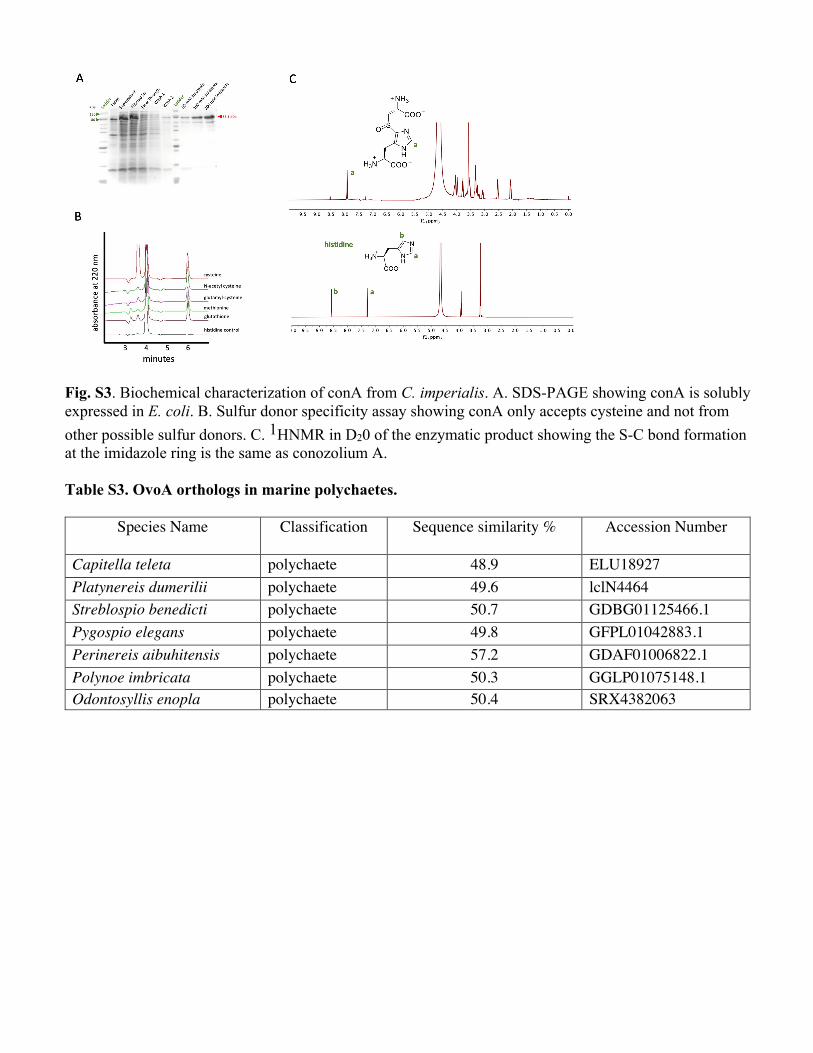
Fig. S3. Biochemical characterization of conA from C. imperialis. A. SDS-PAGE showing conA is solubly expressed in E. coli. B. Sulfur donor specificity assay showing conA only accepts cysteine and not from other possible sulfur donors. C. 1HNMR in D20 of the enzymatic product showing the S-C bond formation at the imidazole ring is the same as conozolium A. Table S3. OvoA orthologs in marine polychaetes.
Species Name Classification Sequence similarity % Accession Number
Capitella teleta polychaete 48.9 ELU18927 Platynereis dumerilii polychaete 49.6 lclN4464 Streblospio benedicti polychaete 50.7 GDBG01125466.1 Pygospio elegans polychaete 49.8 GFPL01042883.1 Perinereis aibuhitensis polychaete 57.2 GDAF01006822.1 Polynoe imbricata polychaete 50.3 GGLP01075148.1 Odontosyllis enopla polychaete 50.4 SRX4382063

Fig. S4. Shallow and deep water Conus imperialis specimens. A Deepwater C. imperialis are generally smaller, free of periostacum and more pronounced spires. B Shallow water variants are bigger, with periostacum and have less pronounced spires. Table S4. GenBank accession numbers for species in COI-based phylogenetic tree.
Species COI Reference Profundiconus teramachii KJ550488 (1) Californiconus californicus FJ868112 (65) Conus distans KJ606037 (11) Conus distans KJ550204 (1) Conus varius KJ550512 (1) Conus monile KJ549959 (1) Conus generalis KJ549916 (1) Conus sugimotonis KJ550462 (1) Conus plinthis KJ550416 (1) Conus hirasei KJ550284 (1) Conus striatellus KJ549994 (1) Conus planorbis FJ868158 (65) Conus ferrugineus KJ550257 (1) Conus crotchii MF491598 (66) Conus fuscoflavus MF491592 (66) Conus cuneolus MF491582 (66) Conus longilineus MF491572 (66) Conus miruchae KY864971 (66) Conus navarroi MF491600 (66) Conus infinitus KY864967 (66) Conus dorotheae KJ550412 (1) Conus zebroides KU892093 (67) Conus ventricosus KJ550006 (1) Conus ateralbus AY588154 (68) Conus marimaris MH243428 (69) Conus pseudonivifer KY864969 (66) Conus trochulus AY588227 (68)
Conus venulatus AY588230 (66) Conus byssinus KY864073 (64) Conus pulcher KY864972 (64) Conus archon KJ549860 (1) Conus cedonulli dominicanus KJ549885 (1) Conus curassaviensis KJ549893 (1) Conus mappa KJ549949 (1) Conus mappa granarius KJ549948 (1) Conus regius GU134385 (70) Conus chiangi GU134381 (70) Conus chiangi KJ550172 (1) Conus genuanus KY864974 (71) Conus brunneus GU134384 (70) Conus bartschii AY588159 (68) Conus zonatus GU134383 (70) Conus imperialis 4637Y MW504291 This work Conus imperialis 01914 KJ550308 (1) Conus imperialis 00821 AY588186 (68) Conus imperialis 4370L MW504292 This work Conus imperialis 4372P MW504293 This work Conus imperialis 00135 KJ549924 (1) Conus imperialis var fuscatus KJ550309 (1) Conus imperialis 012019 MW504294 This work Conus imperialis 012008 GU134382 (70) Conus imperialis 022019 MW504295 This work Conus imperialis 4371M MW504296 This work Conus imperialis 19003H MW504297 This work Conus imperialis 4373S MW504298 This work Conus imperialis 19002H MW504299 This work Conus imperialis 19001H MW504300 This work Table S5. DRG assay (% neurons affected per subclass).
L1 L2 L3 L4 L5 L6 G7 G8 G9 G10 R11 R12 R13 N14 N15 N16 Compounds A B D A B D A B D A B D A B D A B D A B D A B D A B D A B D A B D A B D A B D A B D A B D A B D γ-glutamyl-serotonin 0 0 0 0 0 0 6 0 0 0 0 0 0 0 0 7 0 0 4 0 0 23 0 0 49 0 8 36 0 5 15 0 0 18 0 9 9 0 1 6 0 0 5 1 0 0 0 0
tryptophan 0 0 0 1 10 0 20 10 0 9 0 0 60 0 0 11 0 0 0 0 0 3 0 0 10 0 2 13 0 0 0 0 0 0 0 0 0 0 0 3 0 0 0 0 1 0 0 0 A: application of KCl response, B: blockage of KCl response, D: direct depolarizing the neuronal cells, L1-L6, G7-G10, R11-R13 and N14-N16: subclass of DRG neuronal cells.
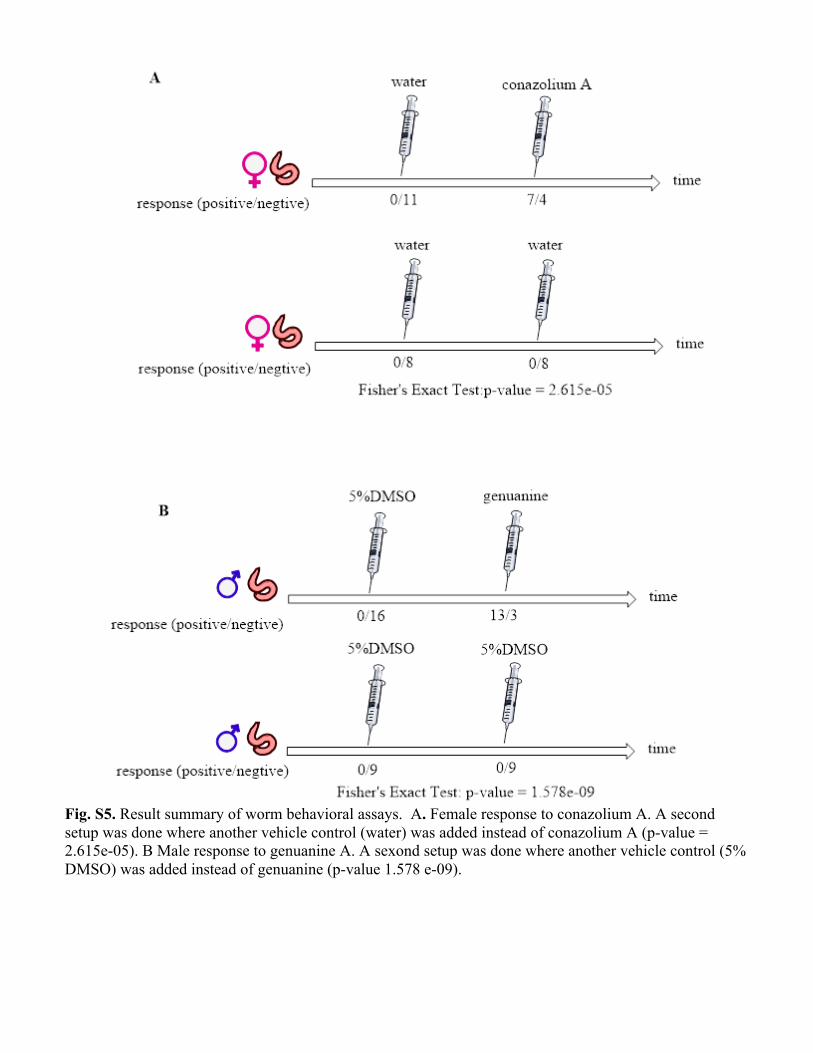
Fig. S5. Result summary of worm behavioral assays. A. Female response to conazolium A. A second setup was done where another vehicle control (water) was added instead of conazolium A (p-value = 2.615e-05). B Male response to genuanine A. A sexond setup was done where another vehicle control (5% DMSO) was added instead of genuanine (p-value 1.578 e-09).

Fig. S6. 16S rRNA gene phylogenetic tree showing placement of prey polychaetes within order Phyllodocida. Bootstrap numbers are given at each node (n = 100). This result expands the known prey range of C. imperialis beyond order Aciculata. Supplementary Movie Captions
Movie S1. Platynereis dumerilii mating. Male and female worms can be observed undergoing the normal nuptial dance, followed by fertilization. The female can be seen swimming in tight circles. The behavior and egg/sperm release is mediated by pheromones ovothiol and uric acid. Move S2. Genuanine induces sperm release. A male worm is treated with vehicle (left) or genuanine (right). When genuanine was added, sperm was released in 5/7 male worms tested. Movie S3. Conazolium A induces mating dance tight swimming phenotype. A female worm is treated with vehicle (left) or conazolium A (right). When conazolium was added, the tight-swimming phenotype was observed in 3/4 worms tested.
Movie S4. Ruby-Tracker video of female worm treated with conazolium A. Tight circle swimming behavior was quantified using this method.
Movie S5. Conus imperialis probing for fireworms. The cone snail is observed discovering a polychaete that is hiding in a crevice in an aquarium.
Movie S6. Conus imperialis feeding on fireworm. The cone snail harpoons a polychaete and pulls it into its mouth. The harpoon is not released.
Movie S7. Conus imperialis injects venom. The colored venom can be observed during injection. In this instance, the harpoon is released from the prey.

REFERENCES AND NOTES
1. N. Puillandre, P. Bouchet, T. F. Duda Jr., S. Kauferstein, A. J. Kohn, B. M. Olivera, M. Watkins, C.
Meyer, Molecular phylogeny and evolution of the cone snails (Gastropoda, Conoidea). Mol.
Phylogenet. Evol. 78, 290–303 (2014).
2. B. Gao, C. Peng, J. Yang, Y. Yi, J. Zhang, Q. Shi, Cone snails: A big store of conotoxins for novel
drug discovery. Toxins 9, 397 (2017).
3. S. W. A. Himaya, R. J. Lewis, Venomics-accelerated cone snail venom peptide discovery. Int. J. Mol.
Sci. 19, 788 (2018).
4. J. W.-H. Li, J. C. Vederas, Drug discovery and natural products: End of an era or an endless frontier?
Science 325, 161–165 (2009).
5. B. M. Olivera, J. Seger, M. P. Horvath, A. E. Fedosov, Prey-capture strategies of fish-hunting cone
snails: Behavior, neurobiology and evolution. Brain Behav. Evol. 86, 58–74 (2015).
6. H. Safavi-Hemami, J. Gajewiak, S. Karanth, S. D. Robinson, B. Ueberheide, A. D. Douglass, A.
Schlegel, J. S. Imperial, M. Watkins, P. K. Bandyopadhyay, M. Yandell, Q. Li, A. W. Purcell, R. S.
Norton, L. Ellgaard, B. M. Olivera, Specialized insulin is used for chemical warfare by fish-hunting
cone snails. Proc. Natl. Acad. Sci. U.S.A. 112, 1743–1748 (2015).
7. P. Ahorukomeye, M. M. Disotuar, J. Gajewiak, S. Karanth, M. Watkins, S. D. Robinson, P. Flórez
Salcedo, N. A. Smith, B. J. Smith, A. Schlegel, B. E. Forbes, B. Olivera, D. Hung-Chieh Chou, H.
Safavi-Hemami, Fish-hunting cone snail venoms are a rich source of minimized ligands of the
vertebrate insulin receptor. eLife 8, e41574 (2019).
8. H. Safavi-Hemami, A. Lu, Q. Li, A. E. Fedosov, J. Biggs, P. Showers Corneli, J. Seger, M. Yandell,
B. M. Olivera, Venom insulins of cone snails diversify rapidly and track prey taxa. Mol. Biol. Evol.
33, 2924–2934 (2016).
9. J. G. Menting, J. Gajewiak, C. A. MacRaild, D. H.-C. Chou, M. M. Disotuar, N. A. Smith, C. Miller,
J. Erchegyi, J. E. Rivier, B. M. Olivera, B. E. Forbes, B. J. Smith, R. S. Norton, H. Safavi-Hemami,

M. C. Lawrence, A minimized human insulin-receptor-binding motif revealed in a Conus
geographus venom insulin. Nat. Struct. Mol. Biol. 23, 916–920 (2016).
10. B. M. Olivera, S. Raghuraman, E. W. Schmidt, H. Safavi-Hemami, Linking neuroethology to the
chemical biology of natural products: Interactions between cone snails and their fish prey, a case
study. J. Comp. Physiol. A 203, 717–735 (2017).
11. J. W. Aman, J. S. Imperial, B. Ueberheide, M.-M. Zhang, M. Aguilar, D. Taylor, M. Watkins, D.
Yoshikami, P. Showers-Corneli, H. Safavi-Hemami, J. Biggs, R. W. Teichert, B. M. Olivera,
Insights into the origins of fish hunting in venomous cone snails from studies of Conus tessulatus.
Proc. Natl. Acad. Sci. U.S.A. 112, 5087–5092 (2015).
12. J. M. McIntosh, D. Yoshikami, E. Mahe, D. B. Nielsen, J. E. Rivier, W. R. Gray, B. M. Olivera, A
nicotinic acetylcholine receptor ligand of unique specificity, alpha-conotoxin ImI. J. Biol. Chem.
269, 16733–16739 (1994).
13. J. L. B. Neves, Z. Lin, J. S. Imperial, A. Antunes, V. Vasconcelos, B. M. Olivera, E. W. Schmidt,
Small molecules in the cone snail arsenal. Org. Lett. 17, 4933–4935 (2015).
14. J. M. McIntosh, T. A. Foderaro, W. Li, C. M. Ireland, B. M. Olivera, Presence of serotonin in the
venom of Conus imperialis. Toxicon 31, 1561–1566 (1993).
15. G. Khalili, A new synthesis of S-aryl uracils from aryl thiols and 6-amino uracils in the presence of
NCS. Mol. Divers. 20, 963–968 (2016).
16. R. Gugisch, A. Kerber, A. Kohnert, R. Laue, M. Meringer, C. Rücker, A. Wassermann, MOLGEN
5.0, A molecular structure generator,in Advances in Mathematical Chemistry and Applications:
Volume 1, S. C. Basak, G. Restrepo, Jose Villaveces, Eds. (Elsevier, 2016), pp. 113–138.
17. S. D. Robinson, Q. Li, P. K. Bandyopadhyay, J. Gajewiak, M. Yandell, A. T. Papenfuss, A. W.
Purcell, R. S. Norton, H. Safavi-Hemami, Hormone-like peptides in the venoms of marine cone
snails. Gen. Comp. Endocrinol. 244, 11–18 (2017).

18. I. Röhl, B. Schneider, B. Schmidt, E. Zeeck, L-Ovothiol A: The egg release pheromone of the
marine polychaete Platynereis dumerilli: Annelida: Polychaete. Zeischrift Naturforsch. C 54, 1145–
1147 (1999).
19. E. Zeeck, T. Harder, M. Beckmann, C. T. Müller, Marine gamete-release pheomones. Nature 382,
214 (1996).
20. E. Zeeck, T. Harder, M. Beckmann, Uric acid: The sperm release pheromone of the marine polchaete
Platynereis dumerilli. J. Chem. Ecol. 24, 13–22 (1998).
21. E. Zeeck, T. Harder, M. Beckmann, Inosine, L-glutamic acid and L-glutamine as components of a
sex pheromone complex of the marine polychaete Nereis succinea (Annelida: Polychaeta).
Chemoecology 8, 77–84 (1998).
22. G. J. Watson, F. M. Langford, S. M. Gaudron, M. G. Bentley, Factors influencing spawning and
pairing in the scale worm Harmothoe imbricata (Annelida: Polychaeta). Biol. Bull. 199, 50–58
(2000).
23. A. J. Kohn, The ecology of Conus in Hawaii. Ecol. Monogr. 29, 47–90 (1959).
24. E. Kuehn, A. W. Stockinger, J. Girard, F. Raible, B. D. Özpolat, A scalable culturing system for the
marine annelid Platynereis dumerilii. PLOS ONE 14, e0226156 (2019).
25. A. J. Kohn, C. Hunter, The feeding process in Conus imperialis. Veliger 44, 232–234 (2001).
26. D. A. Weese, T. F. Duda Jr., Effects of predator-prey interactions on predator traits: Differentiation
of diets and venoms of a marine snail. Toxins 11, 299 (2019).
27. G. Bulaj, B. M. Olivera, Folding of conotoxins: Formation of the native disulfide bridges during
chemical synthesis and biosynthesis of Conus peptides. Antioxid. Redox Signal. 10, 141–156 (2008).
28. M. Gerdol, M. Sollitto, A. Pallavicini, I. Castellano, The complex evolutionary history of sulfoxide
synthase in ovothiol biosynthesis. Proc. Biol. Sci. 286, 20191812 (2019).

29. A. Braunshausen, F. P. Seebeck, Identification and characterization of the first ovothiol biosynthetic
enzyme. J. Am. Chem. Soc. 133, 1757–1759 (2011).
30. N. Naowarojna, P. Huang, Y. Cai, H. Song, L. Wu, R. Cheng, Y. Li, S. Wang, H. Lyu, L. Zhang, J.
Zhou, P. Liu, In vitro reconstitution of the remaining steps in ovothiol A biosynthesis: C-S lyase and
methyltransferase reactions. Org. Lett. 20, 5427–5430 (2018).
31. J. Corradi, C. Bouzat, Understanding the bases of function and modulation of α7 nicotinic receptors:
Implications for drug discovery. Mol. Pharmacol. 90, 288–299 (2016).
32. N. Abraham, R. J. Lewis, Neuronal nicotinic acetylcholine receptor modulators from cone snails.
Mar. Drugs 16, 208 (2018).
33. R. W. Teichert, E. W. Schmidt, B. M. Olivera, Constellation pharmacology: A new paradigm for
drug discovery. Annu. Rev. Pharmacol. Toxicol. 55, 573–589 (2015).
34. E. Monnier, L. Limpalaër, A. Robin, C. Roux, A Taxonomic Iconography of Living Conidae—
Volume 1 (ConchBooks, 2018).
35. E. G. Fofanova, T. D. Mayorova, E. E. Voronezhskaya, Paradoxical effect of sorotonin on ciliary
locomotion of the adult archiannelid worms Dinophilus gyrociliatus and D. taeniatus (Annelida:
Polychaeta). Invert. Zool. 14, 114–120 (2017).
36. B. D. Sloley, γ-Glutamyl conjugation of 5-hydroxytryptamine (serotonin) in the earthworm
(Lumbricus terrestris). Neurochem. Res. 19, 217–222 (1994).
37. A. W. Sangster, S. E. Thomas, N. L. Tingling, Fish attractants from marine invertebrates: Arcamine
from Arca zebra and strombine from Strombus gigas. Tetrahedron 31, 1135–1137 (1974).
38. G. Ortar, M. G. Cascio, L. de Petrocellis, E. Morera, F. Rossi, A. Schiano-Moriello, M. Nalli, V. de
Novellis, D. F. Woodward, S. Maione, V. di Marzo, New N-arachidonoylserotonin analogues with
potential “Dual” mechanism of action against pain. J. Med. Chem. 50, 6554–6569 (2007).

39. A. Fischer, A. Dorresteijn, The polychaete Platynereis dumerilii (Annelida): A laboratory animal
with spiralian cleavage, lifelong segment proliferation and a mixed benthic/pelagic life cycle.
Bioessays 26, 314–325 (2004).
40. W. G. Eberhard, Aggressive chemical mimicry by a bolas spider. Science 198, 1173–1175 (1977).
41. M. K. Stowe, J. H. Tumlinson, R. R. Heath, Chemical mimicry: Bolas spiders emit components of
moth prey species sex pheromones. Science 236, 964–967 (1987).
42. C. Gemeno, K. V. Yeargan, K. F. Haynes, Aggressive chemical mimicry by the bolas spider
Mastophora hutchinsoni: Identification and quantification of a major prey’s sex pheromone
components in the spider’s volatile emissions. J. Chem. Ecol. 26, 1235–1243 (2000).
43. R. R. Jackson, F. R. Cross, A cognitive perspective on aggressive mimicry. J. Zool. 290, 161–171
(2013).
44. G. A. Jamie, Signals, cues and the nature of mimicry. Proc. Biol. Sci. 284, 20162080 (2017).
45. E. Gebel Berg, The chemistry of the pill. ACS Cent. Sci. 1, 5–7 (2015).
46. E. Zeeck, J. Hardege, H. Bartels-Hardege, Sex pheromones and reproductive isolation in two nereid
species, Nereis succinea and Platynereis dumerilii. Mar. Ecol. Prog. Ser. 67, 183–188 (1990).
47. J. Hardege, C. T. Müller, M. Beckmann, H. D. Bartels-Hardege, Timing of reproduction in marine
polychaetes: The role of sex pheromones. Ecoscience 5, 395–404 (1998).
48. M. D. Shultz, Two decades under the influence of the rule of five and the changing properties of
approved oral drugs. J. Med. Chem. 62, 1701–1714 (2019).
49. Z. Lin, S. Phadke, Z. Lu, S. Beyhan, M. H. Abdel Aziz, C. Reilly, E. W. Schmidt, Onydecalins,
fungal polyketides with anti-Histoplasma and anti-TRP activity. J. Nat. Prod. 81, 2605–2611 (2018).
50. M. Wang, J. J. Carver, V. V. Phelan, L. M. Sanchez, N. Garg, Y. Peng, D. D. Nguyen, J. Watrous,
C. A. Kapono, T. Luzzatto-Knaan, C. Porto, A. Bouslimani, A. V. Melnik, M. J. Meehan, W.-T. Liu,
M. Crüsemann, P. D. Boudreau, E. Esquenazi, M. Sandoval-Calderón, R. D. Kersten, L. A. Pace, R.

A. Quinn, K. R. Duncan, C.-C. Hsu, D. J. Floros, R. G. Gavilan, K. Kleigrewe, T. Northen, R. J.
Dutton, D. Parrot, E. E. Carlson, B. Aigle, C. F. Michelsen, L. Jelsbak, C. Sohlenkamp, P. Pevzner,
A. Edlund, J. M. Lean, J. Piel, B. T. Murphy, L. Gerwick, C.-C. Liaw, Y.-L. Yang, H.-U. Humpf, M.
Maansson, R. A. Keyzers, A. C. Sims, A. R. Johnson, A. M. Sidebottom, B. E. Sedio, A. Klitgaard,
C. B. Larson, C. A. Boya P, D. Torres-Mendoza, D. J. Gonzalez, D. B. Silva, L. M. Marques, D. P.
Demarque, E. Pociute, E. C. O’Neill, E. Briand, E. J. N. Helfrich, E. A. Granatosky, E. Glukhov, F.
Ryffel, H. Houson, H. Mohimani, J. J. Kharbush, Y. Zeng, J. A. Vorholt, K. L. Kurita, P. Charusanti,
K. L. McPhail, K. F. Nielsen, L. Vuong, M. Elfeki, M. F. Traxler, N. Engene, N. Koyama, O. B.
Vining, R. Baric, R. R. Silva, S. J. Mascuch, S. Tomasi, S. Jenkins, V. Macherla, T. Hoffman, V.
Agarwal, P. G. Williams, J. Dai, R. Neupane, J. Gurr, A. M. C. Rodríguez, A. Lamsa, C. Zhang, K.
Dorrestein, B. M. Duggan, J. Almaliti, P.-M. Allard, P. Phapale, L.-F. Nothias, T. Alexandrov, M.
Litaudon, J.-L. Wolfender, J. E. Kyle, T. O. Metz, T. Peryea, D.-T. Nguyen, D. Van Leer, P. Shinn,
A. Jadhav, R. Müller, K. M. Waters, W. Shi, X. Liu, L. Zhang, R. Knight, P. R. Jensen, B. Ø.
Palsson, K. Pogliano, R. G. Linington, M. Gutiérrez, N. P. Lopes, W. H. Gerwick, B. S. Moore, P.
C. Dorrestein, N. Bandeira, Sharing and community curation of mass spectrometry data with Global
Natural Products Social Molecular Networking. Nat. Biotechnol. 34, 828–837 (2016).
51. P. Shannon, A. Markiel, O. Ozier, N. S. Baliga, J. T. Wang, D. Ramage, N. Amin, B. Schwikowski,
T. Ideker, Cytoscape: A software environment for integrated models of biomolecular interaction
networks. Genome Res. 13, 2498–2504 (2003).
52. A. Bankevich, S. Nurk, D. Antipov, A. A. Gurevich, M. Dvorkin, A. S. Kulikov, V. M. Lesin, S. I.
Nikolenko, S. Pham, A. D. Prjibelski, A. V. Pyshkin, A. V. Sirotkin, N. Vyahhi, G. Tesler, M. A.
Alekseyev, P. A. Pevzner, SPAdes: A new genome assembly algorithm and its applications to
single-cell sequencing. J. Comput. Biol. 19, 455–477 (2012).
53. D. Hyatt, G.-L. Chen, P. F. LoCascio, M. L. Land, F. W. Larimer, L. J. Hauser, Prodigal:
Prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 11,
119 (2010).
54. B. Langmead, C. Trapnell, M. Pop, S. L. Salzberg, Ultrafast and memory-efficient alignment of
short DNA sequences to the human genome. Genome Biol. 10, R25 (2009).

55. N. M. Davidson, A. Oshlack, Corset: Enabling differential gene expression analysis for de
novoassembled transcriptomes. Genome Biol. 15, 410 (2014).
56. M. D. Robinson, D. J. McCarthy, G. K. Smyth, edgeR: A Bioconductor package for differential
expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
57. O. Folmer, M. Black, W. Hoeh, R. Lutz, R. Vrijenhoek, DNA primers for amplification of
mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol.
Biotechnol. 3, 294–299 (1994)
58. M. A. Larkin, G. Blackshields, N. P. Brown, R. Chenna, P. A. McGettigan, H. McWilliam, F.
Valentin, I. M. Wallace, A. Wilm, R. Lopez, J. D. Thompson, T. J. Gibson, D. G. Higgins, Clustal
W and Clustal X version 2.0. Bioinformatics 23, 2947–2948 (2007).
59. A. Dereeper, V. Guignon, G. Blanc, S. Audic, S. Buffet, F. Chevenet, J.-F. Dufayard, S. Guindon, V.
Lefort, M. Lescot, J.-M. Claverie, O. Gascuel, Phylogeny.fr: Robust phylogenetic analysis for the
non-specialist. Nucleic Acids Res. 36, W465–W469 (2008).
60. R. W. Teichert, N. J. Smith, S. Raghuraman, D. Yoshikami, A. R. Light, B. M. Olivera, Functional
profiling of neurons through cellular neuropharmacology. Proc. Natl. Acad. Sci. U.S.A. 109, 1388–
1395 (2012).
61. M. J. Giacobassi, L. S. Leavitt, S. Raghuraman, R. Alluri, K. Chase, R. K. Finol-Urdaneta, H.
Terlau, R. W. Teichert, B. M. Olivera, An integrative approach to the facile functional classification
of dorsal root ganglion neuronal subclasses. Proc. Natl. Acad. Sci. U.S.A. 117, 5494–5501 (2020).
62. H. Gross, J. Reitner, G. M. König, Isolation and structure elucidation of azoricasterol, a new sterol of
the deepwater sponge Macandrewia azorica. Naturwissenschaften 91, 441–446 (2004).
63. C. D. Smith, A. Wang, K. Vembaiyan, J. Zhang, C. Xie, Q. Zhou, G. Wu, S. R. W. Chen, T. G.
Back, Novel carvedilol analogues that suppress store-overload-induced Ca2+ release. J. Med. Chem.
56, 8626–8655 (2013).

64. H.-C. Chou, N. Acevedo-Luna, J. A. Kuhlman, S. Q. Schneider, PdumBase: A transcriptome
database and research tool for Platynereis dumerilii and early development of other metazoans. BMC
Genomics 19, 618 (2018).
65. J. S. Biggs, M. Watkins, N. Puillandre, J.P. Ownby, E. Lopez-Vera, S. Christensen, K. J. Moreno, J.
Bernaldez, A. Licea-Navarro, P. S. Corneli, B. M. Olivera, Evolution of Conus peptide toxins:
Analysis of Conus californicus Reeve, 1844. Mol. Phylogenet. Evol. 56, 1–12 (2010)
66. S. Albade, M. J. Tenorio, C. M. L. Afonso, J. E. Uribe, A. M. Echeverry, R. Zardoya, Phylogenetic
relationships of cone snails endemic to Cabo Verde based on mitochondrial genomes. BMC Evol.
Biol. 17 231 (2017).
67. M. J. Tenorio, S. Abalde, J. R. Pardos-Blas, R. Zardoya, Taxonomic revision of West African cone
snails (Gastropoda: Conidae) based upon mitogenomic studies: Implications for conservation. Eur. J.
Taxon. 663, 1–89 (2020).
68. T. F. Duda Jr., E. Rolán, Explosive radiation of Cape Verde Conus, a marine species flock. Mol.
Ecol. 14, 267–272 (2005).
69. M. J. Tenorio, S. Abalde, R. Zardoya, Identification of new species of Kalloconus and Africonus
(Gastropoda, Conidae) from the Cabo Verde Islands through mitochondrial genome comparison.
Festivus 50, 73–88 (2018).
70. M. Watkins, P. S. Cornell, D. Hillyard, B. M. Olivera, Molecular phylogeny of Conus chiangi
(Azuma, 1972) (Gastropoda: Conidae). Nautilus 124, 129–136 (2010).
71. S. Albade, M. J. Tenorio, J. E. Uribe, R. Zardoya, Conidae phylogenomics and evolution. Zool. Scr.
48, 119–214 (2019).
Related Documents











