www.sciencemag.org/cgi/content/full/312/5780/1640/DC1 Supporting Online Material for A Nearly Modern Amphibious Bird from the Early Cretaceous of Northwestern China Hai-lu You, Matthew C. Lamanna,* Jerald D. Harris,* Luis M. Chiappe, Jingmai O’Connor, Shu-an Ji, Jun-chang Lü, Chong-xi Yuan, Da-qing Li, Xing Zhang, Kenneth J. Lacovara, Peter Dodson, Qiang Ji *To whom correspondence should be addressed. E-mail: [email protected] (M.C.L); [email protected] (J.D.H.) Published 16 June 2006, Science 312, 1640 (2006) DOI: 10.1126/science.1126377 This PDF file includes: Materials and Methods SOM Text Figs. S1 to S6 Tables S1 and S2 References

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.sciencemag.org/cgi/content/full/312/5780/1640/DC1
Supporting Online Material for
A Nearly Modern Amphibious Bird from the Early Cretaceous of Northwestern China
Hai-lu You, Matthew C. Lamanna,* Jerald D. Harris,* Luis M. Chiappe, Jingmai O’Connor, Shu-an Ji, Jun-chang Lü, Chong-xi Yuan, Da-qing Li, Xing Zhang, Kenneth J.
Lacovara, Peter Dodson, Qiang Ji
*To whom correspondence should be addressed. E-mail: [email protected] (M.C.L); [email protected] (J.D.H.)
Published 16 June 2006, Science 312, 1640 (2006)
DOI: 10.1126/science.1126377
This PDF file includes:
Materials and Methods SOM Text Figs. S1 to S6 Tables S1 and S2 References

Supporting Online Material for:
A Nearly Modern Amphibious Bird from the Early Cretaceous of Northwestern China
Hai-lu You, Matthew C. Lamanna*, Jerald D. Harris*, Luis M. Chiappe, Jingmai O’Connor, Shu-an Ji, Jun-
chang Lü, Chong-xi Yuan, Da-qing Li, Xing Zhang, Kenneth J. Lacovara, Peter Dodson, & Qiang Ji
*To whom correspondence should be addressed. E-mail: [email protected] and [email protected]
This PDF file includes: SOM text
1. Materials and Methods
2. Age of the Xiagou Formation
3. Comment on the Contrasting Avifaunas of the Xiagou Formation and Jehol Group
4. Systematic Terminology and Groupings
5. Results of the Phylogenetic Analysis
6. Character States for Ambiortus, Gansus, Hongshanornis, Yanornis, and Yixianornis
7. Comments on Taxa Not Included in the Phylogenetic Analysis
References
Tables S1, S2
Figures S1-S6

1. Materials and Methods All fossil bird specimens described herein were recovered from the Xiagou Formation near Changma, all from very close to the holotype locality of Gansus yumenensis. In addition to specimens of G. yumenensis, our localities have also thus far produced a salamander, a turtle, numerous osteichthyans, insects, ostracodes, conchostracans, and plant megafossils, none of which has yet been formally studied and thus cannot be assigned to any particular taxon. All of the specimens of G. yumenensis discussed herein are reposited at the Chinese Academy of Geological Sciences, Institute of Geology (CAGS-IG) in Beijing. Most of the avian specimens are currently undergoing preparation; however, five completely prepared exemplars (Figs. 2A-J, S1-5), CAGS-IG-04-CM-001, -002, -003, -004, and -008, were utilized to code Gansus for phylogenetic analysis in the character matrix of (S1). Codings were also added for Liaoningornis longidigitrus, Hongshanornis longicresta, Yanornis martini, and Yixianornis grabaui from (S2) with emendations for the latter three taxa based on personal observation by L.M.C. and J.O. We did not incorporate a separately-described specimen referred to Yanornis (S3). Using data from published sources (S4-S6), we added the Early Cretaceous Mongolian putative ornithuran Ambiortus dementjevi to our matrix as well (see below for codings). The resultant data matrix was analyzed in PAUP* 4.0b10 (S7) on a Macintosh G5.
Because the specimens described herein pertain to birds, osteological terminology used follows the Nomina Anatomica Avium (S8). The term “enantiornithean,” rather than “enantiornithine,” is used herein as an informal shorthand for a member of the clade Enantiornithes because the latter implies the existence of, and membership in, a clade “Enantiornithinae” despite the fact that no such clade has ever been recognized. Similarly, “ornithothoracean” is used instead of “ornithothoracine” because there is no clade “Ornithothoracinae,” only Ornithothoraces. The same logic applies to “ornithuran” versus “ornithurine” for Ornithurae, “hesperornithean” vs. “hesperornithine” for Hesperornithes, and “neornithean” versus “neornithine” for Neornithes. This pattern conforms to prevalent usage for other avian and non-avian theropod clade names with similar suffixes (e.g., “avian” rather than “avine” for Aves, “avialan” rather than “avialine” for Avialae, and “tetanuran” rather than “tetanurine” for Tetanurae) and brings paleornithological terminology into congruence with the nomenclature of other organisms (e.g., “gnetalean” for Gnetales, “aranean” for Araneae, “schizacean” for Schizaceae, etc.).
2. Age of the Xiagou Formation The Xiagou Formation is the middle unit of the Xinminpu Group, which is considered Early Cretaceous in age (S9, S10). A more precise age assignment cannot yet be stated with certainty, but the underlying Chijinpu Formation has produced fossils characteristic of the Jehol Biota (S11); the Xiagou Formation produces some identical insect species and taxa similar at higher taxonomic levels, but lacks key taxa such as the fish Lycoptera (S10). The Xiagou Formation thus appears to be slightly younger than the Barremian-Aptian (S12, S13) Jehol Group of Liaoning, Hebei, and Nei Mongol provinces, rendering Gansus younger in age than probable non-ornithuran ornithuromorphs from the Yixian and Jiufotang formations (Chaoyangia, Hongshanornis, Liaoningornis, Songlingornis, Yanornis, and Yixianornis) and the Unduruh Formation (“Khurilt beds”) of Mongolia (Ambiortus, the age of which was discussed in (S5)).
However, while no biostratigraphic analyses have been performed on fossils from the Xiagou Formation in the Changma Basin itself, studies of taxa from other outcrops referred to

this unit have led to other age assessments: ostracodes (S14) and charophytes (S15, S16) both appear to indicate a Barremian age, while a palynoflora has been interpreted as Hauterivian (S17). Either age would render Gansus as old as or older than the Jehol Biota. Although we cannot comment on the validity of these results, we note that previous, biostratigraphically-determined age assessments for the Yixian Formation based on palynomorphs and ostracodes (often employing “stage of evolution” arguments) tended to overestimate the age of that unit with respect to the radiometrically-determined dates recovered later (S18). Therefore, we regard the results of (S14-S17) with caution.
3. Comment on the Contrasting Avifaunas of the Xiagou Formation and Jehol Group The Xiagou Formation is currently the only known Lower Cretaceous, fossil bird-producing unit dominated by ornithuromorphs rather than non-ornithuromorph (enantiornithean and more basal, non-ornithothoracean) taxa. Enantiornitheans are not absent, however (S19-S21). Nevertheless, the inverse ratios of ornithuromorph to non-ornithuromorph birds between the Xiagou Formation and Jehol Group of northeastern China further distinguishes the avifaunas of these strata. Given that both units consist largely of lacustrine deposits that lay at similar paleolatitudes at the times of their deposition, the probable age differences are more likely than other factors (e.g., paleoclimate, paleogeographic distance, etc.) to explain these inverse ratios (although it must be noted that the Xiagou Formation was deposited much further inland than the Jehol Group and thus might preserve a more continental paleoclimatic setting). The only hypothesis previously proposed to explain the greater diversity and abundance of specimens of non-ornithuromorph birds in the Jehol Group is that ornithuromorphs were genuinely rarer and/or less speciose than these birds (S22).
No statistics are available as to the comparative diversity of non-ornithuromorph and ornithuromorph birds in the Late Cretaceous, but it is noteworthy that there are many more recognized ornithuromorph taxa from the Late Cretaceous than the Early Cretaceous. Although not all Late Cretaceous avian assemblages are ornithuromorph-dominated [the Maastrichtian El Brete assemblage of Argentina, for example, does not include ornithuromorphs (S23-S25)], many Late Cretaceous assemblages are composed primarily or exclusively of ornithuromorphs, including those of the Coniacian-Santonian Niobrara Formation of Kansas (S26-S28), the Maastrichtian Lance Formation of Wyoming (S29-S31), the Maastrichtian Navesink and (at least lowermost) Hornerstown formations of New Jersey (S32-S36), and the Campanian-Maastrichtian López de Bertodano Formation and related deposits of Antarctica (S37-S39). Many terrestrial, Upper Cretaceous formations also produce diving ornithuran birds, including the Nemegt Formation of Mongolia (S40), the Dinosaur Park Formation of Alberta (S41), and the Lance Formation of Wyoming (S31).
How many of the differences between the Xiagou Formation and Jehol Group avifaunas can be attributed to taphonomic biases is unknown. However, these differences may not be purely taphonomic, since the lacustrine setting of the Yixian and Jiufotang Formations of northeastern China (which are paleoenvironmentally at least broadly similar to the Xiagou Formation) produce more terrestrial (non-amphibious) than amphibious birds despite the definite presence of the latter (S2, S3, S22) and the predominance of aquatic, non-avian taxa (e.g., fish, choristoderes) compared to other vertebrates. If there was a genuine increase in ornithuromorph diversity and abundance through the Cretaceous, and a concomitant transition of most avifaunas

from non-ornithuromorph-dominated to ornithuromorph-dominated, then the Changma assemblage represents the earliest known stage of this transition.
4. Systematic Terminology and Groupings How the systematic position of Gansus is perceived depends on which clade definitions are used. We resolve Gansus within the Ornithuromorpha sensu (S6), which Chiappe [(S6): 457)] explicitly defined as “the common ancestor of Patagopteryx and Ornithurae plus all its descendants.” We further resolve Gansus within the Ornithurae sensu (S42), itself sensu (S43). Using the definitions of (S1) and (S44), Gansus resides within the Ornithurae based solely on its possession of the diagnostic character of that clade (caudal vertebral series shorter than the femur). However, the definitions of (S1) and (S44) utilize a nomenclatural protocol that explicitly includes a diagnostic character as part of the definition, a practice that has not yet been widely adopted and remains somewhat controversial. If this classification were adopted here, Gansus would also reside in the Carinatae, a substantially different use of this clade name than currently recognized. We understand the utility of the system promoted by (S1) and (S44) and would accept this alternative phylogenetic definition if and when their system gains more widespread acceptance. However, the phylogenetic topology of the present analysis would not be altered by such changes in higher taxonomic nomenclature.
Other clade names employed in the paper (e.g., Fig. 3) are used as follows:
Aves Linnaeus, 1758 [= Avialae of (S45)]
Pygostylia Chatterjee, 1997 (S46) [sensu (S6)]
Ornithothoraces Chiappe, 1996 (S47) [sensu (S42)]
Enantiornithes Walker, 1981 (S23) [sensu (S6)]
Ornithuromorpha Chiappe, 2002 (S6)
Ornithurae Haeckel, 1866 (S48) [sensu (S42)]
Carinatae Merrem, 1816 (S49) [sensu (S42)]
Neornithes Gadow, 1893 (S50) [= Aves of (S45)] [sensu (S42)].
5. Results of the Phylogenetic Analysis The analysis generated 24 equally parsimonious trees, all with length = 538, CI = 0.643, and RI = 0.762. In six of these trees, Gansus occurs as the sister taxon of the Ornithurae [Hesperornithes + Carinatae]; in the remaining 18 trees, Gansus is nested within the Ornithurae, but in two different positions. In nine of the 18 trees, the Ornithurae consists of [Hesperornithes + [Ichthyornis + [Gansus + Neornithes]]], which would make Gansus not only an ornithuran but a member of the Carinatae as well. In the remaining nine, the positions of Gansus and

Ichthyornis dispar are reversed such that Gansus becomes the sister taxon of the Carinatae. The large number of trees was due, in part, to rearrangement within the Enantiornithes and the presence of wildcard taxa such as the very fragmentary Vorona berivotrensis, which alternated between being the sister taxon of the Ornithuromorpha, within or the sister taxon of the Enantiornithes, and the sister taxon of the Ornithothoraces.
The non-ornithuran Ornithuromorpha, including other wildcard taxa coded with large quantities of missing data (Ambiortus, Apsaravis ukhaana, Yanornis, and Yixianornis), were recovered in several different configurations, including:
[Patagopteryx + [Hongshanornis +[Ambiortus + [[Yanornis + Yixianornis] + [Apsaravis +…]]]]] (18 trees)
[Patagopteryx + [Hongshanornis + [[Apsaravis + [Yanornis + [Ambiortus + Yixianornis]]] + …]]] (2 trees)
[Patagopteryx + [Hongshanornis +[Ambiortus + [[Apsaravis + [Yanornis + Yixianornis]] + …]]]] (2 trees)
[Patagopteryx + [Hongshanornis +[Ambiortus + [Yixianornis + [[Apsaravis + Yanornis] + …]]]]] (2 trees)
All consensus trees also recovered Gansus within the Ornithurae. Identical strict and semi-strict consensus trees recover a basal ornithuran polytomy consisting of Ichthyornis, Gansus, Hesperornithes, and Neornithes. An Adams consensus recovered the Ornithurae with a smaller basal polytomy consisting of Gansus, Hesperornithes, and the Carinatae. A 50% majority rule consensus (Fig. 3) recovered no polytomies and instead constructed the Ornithurae as [Hesperornithes + [Gansus + Carinatae]]. In this tree, the Ornithurae had a 100% frequency occurrence, though the clade [Gansus + Carinatae] had 75% occurrence and the Carinatae only 62%. A bootstrap analysis, again using the methodology of (S1), however, recovered a massive polytomy (with a value of 57) in which the Ornithuromorpha and Ornithurae have the same contents (in which case Ornithurae has priority over Ornithuromorpha): Ambiortus, Apsaravis, Gansus, Hongshanornis, Ichthyornis, Patagopteryx deferrariisi, Yanornis, Yixianornis, the Hesperornithes, and the Neornithes.
None of the Jehol Group taxa that have often been referred to as ornithurans occur within the Ornithurae as recovered by our analysis. Hongshanornis, Yanornis, and Yixianornis are, however, all ornithuromorphs (see above for recovered positions). Unexpectedly, Liaoningornis never fell as an ornithuromorph: in 20 of the 24 trees, it occurs as the second-most basal bird in the analysis, between Archaeopteryx and Confuciusornis, a position also recovered in the 50% majority rule consensus tree (Fig. 3). In the remaining four trees, it occurs as the sister taxon of Vorona, in a clade that is itself the sister taxon of the Enantiornithes. Either of these results would require that features such a long sternal carina and an extensively coossified tarsometatarsus evolved convergently in Liaoningornis and the Ornithuromorpha. Strict and semi-strict consensus trees recovered broadly-encompassing polytomies below the Ornithuromorpha that include all enantiornithean taxa, Liaoningornis, Confuciusornis, and

Vorona. The Adams consensus recovered a polytomous clade consisting of Confuciusornis, Liaoningornis, and another polytomous clade including Vorona, the Enantiornithes, and the Ornithuromorpha.
The following synapomorphies for various ornithuromorph clades recovered by the present analysis [using the 50% majority rule consensus] were determined using MacClade 4.06 (S51):
Under the default, “soft polytomy” option, MacClade could not produce a list of synapomorphies for the Ornithuromorpha because of its inclusion in a polytomy also involving the Enantiornithes and Vorona. Under the “hard polytomy” option, however, the Ornithuromorpha is united by eight unambiguous synapomorphies: scapula curved dorsoventrally (101:1), scapula as long or longer than the humerus (102:1), semilunate carpal and metacarpals completely fused into carpometacarpus (139:3), ilium, ischium, and pubis completely fused proximally (153:2), M. iliofemoralis internus (sensu (S52); = “cuppedicus” of (S1, S53)) fossa not demarcated by broad, mediolaterally oriented surface cranioventral to acetabulum (164:1), cranial trochanter of femur absent (170:2), distal tarsals and metatarsals completely fused and metatarsals fused distally to enclose a distal vascular foramen (188:3), and hypotarsus with a flat caudal surface developed as caudal projection of tarsometatarsus (192:1).
The clade [Hongshanornis + [Ambiortus + [[Yanornis + Yixianornis] + [Apsaravis + Ornithurae]]]] is supported by two unambiguous synapomorphies: humeral head domed proximally (106:1), first phalanx of manual major digit strongly compressed dorsoventrally and flat caudally (150:1).
The clade [Ambiortus + [[Yanornis + Yixianornis] + [Apsaravis + Ornithurae]]] is united by two unambiguous synapomorphies: furcula lacking a hypocleidum (80:1 0) and extensor process of metacarpal I present and barely surpasses the distal articular facet for phalanx I-1 cranially (142:1).
The clade [[Yanornis + Yixianornis] + [Apsaravis + Ornithurae]] is supported by two unambiguous synapomorphies: craniocaudal diameter of metacarpal III less than 50% that of metacarpal II (141:1) and phalanx II-1 longer than II-2 (151:1).
The clade [Apsaravis + Ornithurae] is supported by five unambiguous synapomorphies: 10 ankylosed sacral vertebrae (61:4), pit-like scar for muscle attachment present on craniodistal, distal, or caudodistal surface of humeral bicipital crest (115:1), radius with muscle scar along most of surface (135:1), pubes compressed mediolaterally (166:1), and pubes not in contact distally (167:1).
The Ornithurae [Hesperornithes + [Gansus + Carinatae] is supported by four unambiguous synapomorphies: dorsal surface of coracoid flat to convex (88:1), extensor canal of tibiotarsus comprised of an emarginate groove (180:1), fossa for metatarsal I on metatarsal II a conspicuous, ovoid fossa (196:1), and metatarsal II shorter than metatarsal IV (202:2).
The clade [Gansus + Carinatae] is united by four unambiguous synapomorphies: three sacral vertebrae with dorsally-directed costolateral eminences (62:1), lateral process of coracoid present (92:1), medial surface of coracoid in vicinity of N. supracoracoideus foramen flat to convex (98:1), and ischium straight, with no dorsal process (155:1 0).
Gansus itself is supported by 10 autapomorphic states, seven of which constitute reversals: cervical vertebrae not heterocoelous (52:1/2 0) (note that this character is based

solely on the caudal cervicals; the condition of the cranial-mid cervicals is presently unknown), uncinate processes absent (69:1 0), length of humeral ventral condyle equal to or greater than that of the dorsal condyle (125:1), dorsal cotyle of ulna not convex (130:1 0), dorsal process of ischium contacts ilium (156:1), obturator foramen absent (165:1 0), pubis suboval in cross section (166:1 0), pubes contact distally (167:1 0), distal end of pubis expanded (168:1 0), and femur lacks fossa for insertion of capitular ligament (169:1 0). Three of the reversed characters, 166-168, were coded conservatively in this analysis: 166-167 may be taphonomic (due to coincidental orientation and positions of the pubes), and 168 is fairly slight, comparable to what is seen in some neornitheans (e.g., Gallus).
The Carinatae [Ichthyornis + Neornithes] is united by five unambiguous synapomorphies: thoracic vertebrae with ossified connective tissue bridging the transverse processes (59:1), intermuscular line present on ventral surface of coracoid (93:1); acrocoracoid process of coracoid hooked medially (95:1), ulnare V-shaped with well-developed dorsal and ventral rami (137:1), and postacetabular portion of ilium oriented medially (162:1).
6. Character States for Ambiortus, Gansus, Hongshanornis, Yanornis, and Yixianornis Character 1 2 3 4 5 6
0 0 0 0 0 0 Ambiortus ?????????? ?????????? ?????????? ?????????? ?????????? ?20???????
Gansus ?????????? ?????????? ?????????? ?????????? ?????????? ?012001100 Hongshanornis ?11??????? ?1???????? ?????????? ?????????? ?0?0????01 ???????1??
Yanornis a00010001? ?1???????? ?????????? ?????????? ?000???0?1 ????0?110? Yixianornis 10?010001? ?????????? ?????????? ????11???? ?1?0??1?0? ?a?b0011?0
Character 1 1 1 7 8 9 0 1 2 0 0 0 0 0 0
Ambiortus ?????????? 31?????010 0000110?0? ?1?1010?1? 1?00?111?0 10b00?00?? Gansus e11?011101 31??1?1010 00?01101?? ?1010101?1 10??111100 101?121?10
Hongshanornis ??????1??0 c??????011 ??1??10??? ???10????1 11?00111?? ??10?????? Yanornis f???????10 31?????010 ??10110??? ?0???????1 10??011??? 11100??011
Yixianornis c?????11b0 31???????? 0110111000 ?1010001?1 110011110? ???????01?
Character 1 1 1 1 1 1 3 4 5 6 7 8 0 0 0 0 0 0
Ambiortus ?????????? ?????1??c? 01???????1 00???????? ?????????? ?????????? Gansus 00011???00 1?21?10?c1 1101??1001 102?011110 100100000? 111011210b
Hongshanornis ?0???????? ?????????1 00????00a1 00???????? ??????01?? ?????????? Yanornis 001?0????0 ???1?1??31 ?0?10?0001 1?201?110? ?0??0000?? 1?????????
Yixianornis 00???01000 ??2101??31 1101110?01 10201010?? ?0?10000?2 1?10112???
Character 1 2 9 0 0 0
Ambiortus ?????????? ?????????? ?? Gansus 1b00211311 01122?000? 02
Hongshanornis ?????11??0 0??a???00? ?2 Yanornis ???????311 0?102???a? 01
Yixianornis ????b1131? 01?1?0?0?0 ?1 a = 0/1; b = 1/2; c = 2/3; e = 4/5; f = 2/3/4
Highlighting indicates changes from codings of (S2). Codings for Yanornis based on Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) V13558.

7. Comments on Taxa Not Included in the Phylogenetic Analysis Although other proposed Early Cretaceous ornithuromorphs (typically referred to in their respective descriptions as members of the Ornithurae) have been reported, including Chaoyangia beishanensis (S54) and Songlingornis linghensis (S55), we specifically excluded these taxa from our phylogenetic analysis because they have so far received only preliminary descriptions that do not permit evaluation of many character states in these forms. Moreover, Chaoyangia and, especially, Songlingornis are currently represented only by incomplete material. However, based on the published information available on these taxa, we can briefly comment on their anatomies and possible systematic positions.
Chaoyangia retains an elongate and well-developed pubic symphysis, an apparently plesiomorphic character that is absent in Gansus and the Neornithes. In Gansus, the unfused distal pubes are in contact only for ~5-6% the length of the pubis, whereas in Chaoyangia, the pubic symphysis spans 20% or more of pubic length (S54). Similarly, the retention of gastralia and unfused tarsometatarsal shafts in Chaoyangia (S54) all constitute probable plesiomorphies that are lacking in Gansus and modern birds. Unlike that of Gansus, the scapula of Songlingornis plesiomorphically does not taper distally (S55). The furcula of Songlingornis is primitively thick and heavy (S55), dissimilar to the gracile element of Gansus. Based on the characters described above, we predict that Chaoyangia and Songlingornis will ultimately be resolved as outside the Ornithurae and thus basal to Gansus. An evaluation of this prediction must await further description of known material and/or the documentation of additional specimens of these taxa, either or both of which will facilitate more detailed assessments of their phylogenetic affinities.
Enaliornis spp. were not included in the present phylogenetic analysis because they exhibit hesperornithean autapomorphies (S56) that are absent in Gansus, and consequently would presumably cluster with Hesperornis regalis and Baptornis advenus, both of which were included in the analysis. Although its exact age is uncertain (S56), Enaliornis may represent the only other, currently-recognized, Early Cretaceous ornithuran, so it is worthwhile to present some preliminary comparisons of this taxon to Gansus. As far as can be currently determined, Enaliornis is somewhat younger than Gansus: though recovered from the Cenomanian (99.6-93.5 Mya) Cambridge Greensand of England, its fossils may have been reworked from older, Upper Albian sediments (S56); it thus seems to be between 5-20 million years younger than Gansus (Fig. 3).
Gansus appears less derived than Enaliornis because it lacks fully heterocoelous cervical and thoracic vertebrae. However, Enaliornis is similar to Gansus in the morphology of its cnemial crests: in both taxa, the cnemial crests are conjoined into a proximocranially projecting, triangular process (S56), though this structure is larger in Gansus than Enaliornis. In Gansus, the hypotarsus lacks the low divisory ridge reported in Enaliornis (S56). Though the tarsometatarsus of Enaliornis possesses a proximal vascular foramen on its dorsal face, it is not expressed on the plantar surface, whereas this opening completely penetrates the tarsometatarsus in Gansus. Metatarsal IV of Gansus is neither longer nor significantly larger than metatarsals II or III, unlike Enaliornis and other hesperornitheans. The degree of plantarolateral migration of the trochlea of metatarsal II in the new specimens of Gansus is less than in Enaliornis (S56), but is greater than that figured in the holotype of the former [(S57): fig. 3, no. 6]; this is possibly the result of diagenetic distortion of that specimen.

Supplementary Online Information References S1. J. A. Clarke, M. A. Norell, Am. Mus. Novit. 3387, 1 (2002). S2. Z. Zhou, F. Zhang, Proc. Nat. Acad. Sci. 102, 18998 (2006). S3. C. Yuan, Acta Geol. Sin. 78, 464 (2004). S4. E. N. Kurochkin, in Acta XVIII Congressus Internationalis Ornithologici V. D. Ilyichev,
V. M. Gavrilov, Eds. (Academy of Sciences of the USSR, Moscow), 191-200 (1985). S5. E. N. Kurochkin, in Avian Paleontology at the Close of the 20th Century: Proceedings of
the 4th International Meeting of the Society of Avian Paleontology and Evolution, Washington, D.C., 4-7 June 1996 S. L. Olson, P. Wellnhofer, C. Mourer-Chauviré, D. W. Steadman, L. D. Martin, Eds. (Smithsonian Inst. Press, Washington, D.C.), Smithson. Contrib. Paleobiol. 89, 275-284 (1999).
S6. L. M. Chiappe, in Mesozoic Birds: Above the Heads of Dinosaurs L. M. Chiappe, L. M. Witmer, Eds. (Univ. of California Press, Berkeley), 448-472 (2002).
S7. D. L. Swofford, PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods) (Sinauer Assoc., Sunderland, 2002), pp.
S8. J. J. Baumel, L. M. Witmer, in Handbook of Avian Anatomy: Nomina Anatomica Avium, Second Edition J. J. Baumel, A. S. King, J. E. Breazile, H. E. Evans, J. C. Vanden Berge, Eds. (Nuttall Ornithol. Club, Cambridge), Publ. Nuttall Ornithol. Club 23, 45-132 (1993).
S9. Bureau of Geology and Mineral Resources of Gansu Province, Regional Geology of Gansu Province (Geological Publishing House, Beijing, 1989), pp. 692.
S10. Editorial Committee of Chinese Stratigraphic Standard: Cretaceous, Chinese Stratigraphic Standard: Cretaceous (Geological Publishing House, Beijing, 2000), pp. 124.
S11. Y.-B. Shen, Acta Palaeontol. Sin. 20, 266 (1981). S12. C. C. I. Swisher et al., Chin. Sci. Bull. 47, 135 (2002). S13. H. Y. He et al., Geophys. Res. Lett. 31, L12605 (2004). S14. Y.-X. Hu, Acta Micropalaeontol. Sin. 21, 439 (2004). S15. Q.-F. Wang, J.-L. Yang, H.-N. Lu, Acta Micropalaeontol. Sin. 20, 187 (2003). S16. J.-H. Fu, H.-N. Lu, Acta Palaeontol. Sin. 36, 350 (1997). S17. Z.-S. Liu, Acta Micropalaeontol. Sin. 17, 73 (2000). S18. J. B. Smith, J. D. Harris, G. I. Omar, P. Dodson, H. You, in New Perspectives on the
Origin and Early Evolution of Birds J. Gauthier, L. F. Gall, Eds. (Peabody Mus. Nat. Hist., New Haven), 549-589 (2001).
S19. J. D. Harris, M. C. Lamanna, H.-L. You, S.-A. Ji, Q. Ji, Can. J. Earth Sci. (in press). S20. H. You, J. O'Connor, L. M. Chiappe, Q. Ji, Hist. Biol. 17, 7 (2005). S21. M. C. Lamanna, H.-L. You, S.-A. Ji, J. Lü, Q. Ji, L. M. Chiappe, J. Vertebr. Paleontol.
25, 81A (2005). S22. Z. Zhou, F. Zhang, Chin. Sci. Bull. 46, 1258 (2001). S23. C. A. Walker, Nature 292, 51 (1981). S24. L. M. Chiappe, Am. Mus. Novit. 3083, 1 (1993). S25. L. M. Chiappe, Mem. Queensl. Mus. 39, 533 (1996). S26. J. A. Clarke, Bull. Am. Mus. Nat. Hist. 286, 1 (2004). S27. L. D. Martin, J. D. Stewart, Can. J. Earth Sci. 19, 324 (1982). S28. O. C. Marsh, Prof. Pap. Eng. Dept., U.S. Army 18, 1 (1880).

S29. S. Hope, in Mesozoic Birds: Above the Heads of Dinosaurs L. M. Chiappe, L. M. Witmer, Eds. (Univ. of California Press, Berkeley), 339-388 (2002).
S30. P. Brodkorb, in Proceedings of the 13th Ornithological Congress C. G. Sibley, Ed. 55-70 (1963).
S31. R. Estes, Univ. Calif. Publ. Geol. Sci. 49, 1 (1964). S32. D. C. Parris, S. Hope, in Proceedings of the 5th Symposium of the Society of Avian
Paleontology and Evolution, Beijing, 1-4 June 2000 Z. Zhou, F. Zhang, Eds. (Science Press, Beijing), 113-124 (2002).
S33. O. C. Marsh, Am. J. Sci. (Second Ser.) 146, 205 (1870). S34. S. L. Olson, D. C. Parris, Smithson. Contrib. Paleobiol. 63, 1 (1987). S35. S. L. Olson, in Avian Paleontology at the Close of the 20th Century: Proceedings of the
4th International Meeting of the Society of Avian Paleontology and Evolution, Washington, D.C., 4-7 June 1996 S. L. Olson, P. Wellnhofer, C. Mourer-Chauviré, D. W. Steadman, L. D. Martin, Eds. (Smithson. Inst. Press, Washington, D.C.), Smithson. Contrib. Paleobiol. 89, 231-243 (1999).
S36. J. Cracraft, Auk 89, 36 (1972). S37. J. A. Clarke, C. P. Tambussi, J. I. Noriega, G. M. Erickson, R. A. Ketcham, Nature 433,
305 (2005). S38. S. Chatterjee, in Proceedings of the 5th Symposium of the Society of Avian Paleontology
and Evolution, Beijing, 1-4 June 2000 Z. Zhou, F. Zhang, Eds. (Science Press, Beijing), 125-155 (2002).
S39. J. J. Noriega, C. P. Tambussi, Ameghiniana 32, 57 (1995). S40. J. A. Clarke, M. A. Norell, Am. Mus. Novit. 3447, 1 (2004). S41. N. Longrich, Can. J. Earth Sci. 43, 1 (2006). S42. K. Padian, J. R. Hutchinson, T. R. Holtz, Jr., J. Vertebr. Paleontol. 19, 69 (1999). S43. L. D. Martin, in Perspectives in Ornithology: Essays presented for the Centennial of the
American Ornithological Union A. H. Brush, G. A. Clark, Jr., Eds. (Cambridge Univ. Press, Cambridge), 291-338 (1983).
S44. J. Gauthier, K. de Quieroz, in New Perspectives on the Origin and Early Evolution of Birds J. Gauthier, L. F. Gall, Eds. (Peabody Mus. Nat. Hist., New Haven), 7-41 (2001).
S45. J. Gauthier, in The Origin of Birds and the Evolution of Flight K. Padian, Ed. (California Acad. of Science, San Francisco), Mem. Calif. Acad. Sci. 8, 1-55 (1986).
S46. S. Chatterjee, The Rise of Birds (The Johns Hopkins Univ. Press, Baltimore, 1997), pp. 312.
S47. L. M. Chiappe, Münch. Geowiss. Abh. 30, 203 (1996). S48. E. Haeckel, Generelle Morphologie der Organismen (Verlag von Georg Reimer, Berlin,
1866), pp. 462. S49. B. Merrem, Abhandlungen der Königlichen Akademie der Wissenschaften in Berlin 1812-
1813, 237 (1816). S50. H. Gadow, Dr. H.G. Bronn's Klassen und Ordnungen des Thier-Reichs, wissenschaftlich
dargestellt in Wort und Bild. Sechster Band. Vierte Abtheilung: Vögel (C.F. Winter'sche Verlagshandlung, Leipzig, 1893), pp. 303.
S51. D. R. Maddison, W. P. Maddison. (Sinauer Assoc., Sunderland, 2003). S52. J. R. Hutchinson, Zool. J. Linn. Soc. 131, 123 (2001). S53. T. Rowe, J. Morphol. 189, 327 (1986). S54. L. Hou, J. Zhang, Vertebr. PalAs. 31, 217 (1993).

S55. L. Hou, Mesozoic Birds of China (Feng-huang-ku Bird Park, Taiwan, 1997), pp. 221. S56. P. M. Galton, L. D. Martin, in Mesozoic Birds: Above the Heads of Dinosaurs L. M.
Chiappe, L. M. Witmer, Eds. (Univ. of California Press, Berkeley), 317-338 (2002). S57. L. Hou, Z. Liu, Sci. Sin., Ser. B 27, 1296 (1984).

Table S1: Measurements of selected elements of Gansus yumenensis based on new specimens. For paired elements, all measurements represent elements of the left side unless noted with an (r) indicating that the right side was measured. All measurements reported in mm.
Measurement CAGS-IG-04-
CM-001 CAGS-IG-04-
CM-002 CAGS-IG-04-
CM-003 CAGS-IG-04-
CM-004 CAGS-IG-04-
CM-008
Thoracic vertebral column length — 37 38e 33.3 —
Sacrum length — 26.6 26.9 29.2 —
Tail length — 15.6 — — —
Ver
tebr
ae
Pygostyle length — 5.9 — — —
Scapula length — — — 41.7 —
Coracoid length — — 21.7 19.2 —
Coracoid distal width — — 14.5 14.3 —
Sternum length (maximum) — — 43.4 — —
Sternum width (maximum) — — 30.1 — —
Sternal carina height — — 7.5e — —
Furcula length — — 16.7e — —
Thor
acic
Gird
le
Furcula width (maximum) — — 13.6 13.7e —
Humerus length — — 48.4e 48.0 —
Ulna length — — 52.8 48.8 —
Radius length — — — 46.8 —
Carpometacarpus length — — 25.2 23.4 —
Manual phalanx I-1 length — — — 9.0 —
Manual phalanx I-2 length — — — 3.6 —
Manual phalanx II-1 length — — 11.0 9.8 —
Manual phalanx II-2 length — — 9.6 9.1 —
Thor
acic
Lim
b
Manual phalanx II-3 length — — — 3.4 —
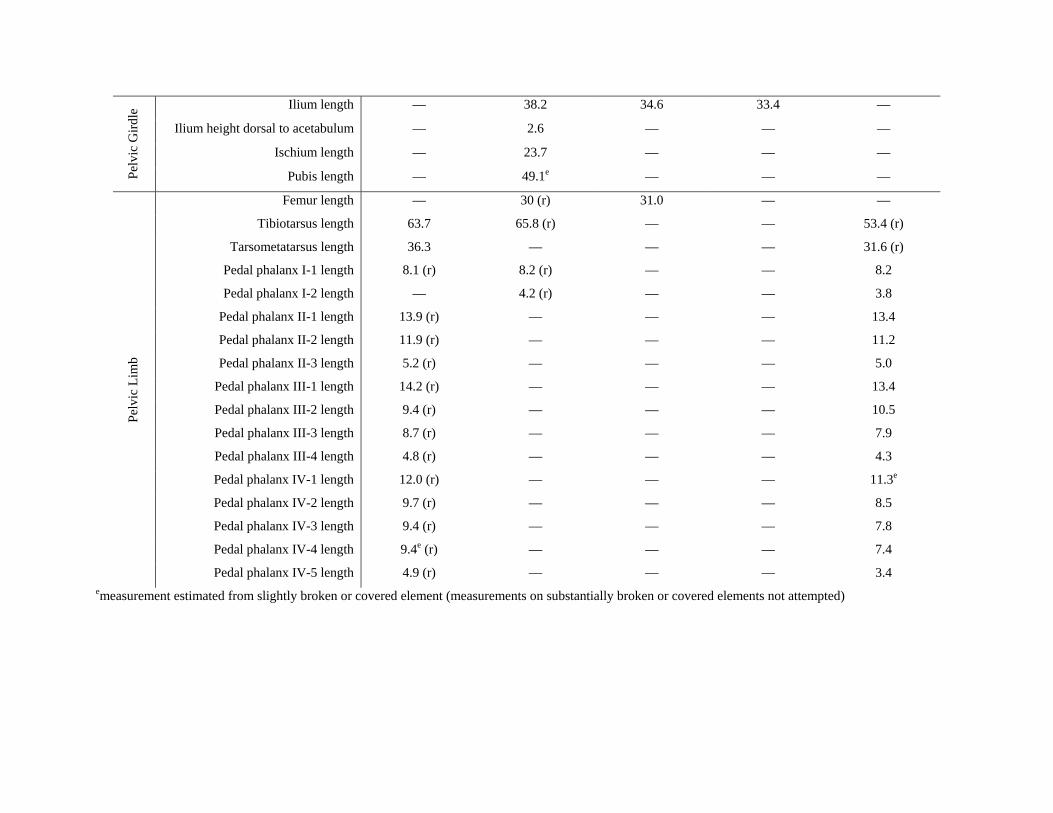
Ilium length — 38.2 34.6 33.4 —
Ilium height dorsal to acetabulum — 2.6 — — —
Ischium length — 23.7 — — — Pe
lvic
Gird
le
Pubis length — 49.1e — — —
Femur length — 30 (r) 31.0 — —
Tibiotarsus length 63.7 65.8 (r) — — 53.4 (r)
Tarsometatarsus length 36.3 — — — 31.6 (r)
Pedal phalanx I-1 length 8.1 (r) 8.2 (r) — — 8.2
Pedal phalanx I-2 length — 4.2 (r) — — 3.8
Pedal phalanx II-1 length 13.9 (r) — — — 13.4
Pedal phalanx II-2 length 11.9 (r) — — — 11.2
Pedal phalanx II-3 length 5.2 (r) — — — 5.0
Pedal phalanx III-1 length 14.2 (r) — — — 13.4
Pedal phalanx III-2 length 9.4 (r) — — — 10.5
Pedal phalanx III-3 length 8.7 (r) — — — 7.9
Pedal phalanx III-4 length 4.8 (r) — — — 4.3
Pedal phalanx IV-1 length 12.0 (r) — — — 11.3e
Pedal phalanx IV-2 length 9.7 (r) — — — 8.5
Pedal phalanx IV-3 length 9.4 (r) — — — 7.8
Pedal phalanx IV-4 length 9.4e (r) — — — 7.4
Pelv
ic L
imb
Pedal phalanx IV-5 length 4.9 (r) — — — 3.4 emeasurement estimated from slightly broken or covered element (measurements on substantially broken or covered elements not attempted)
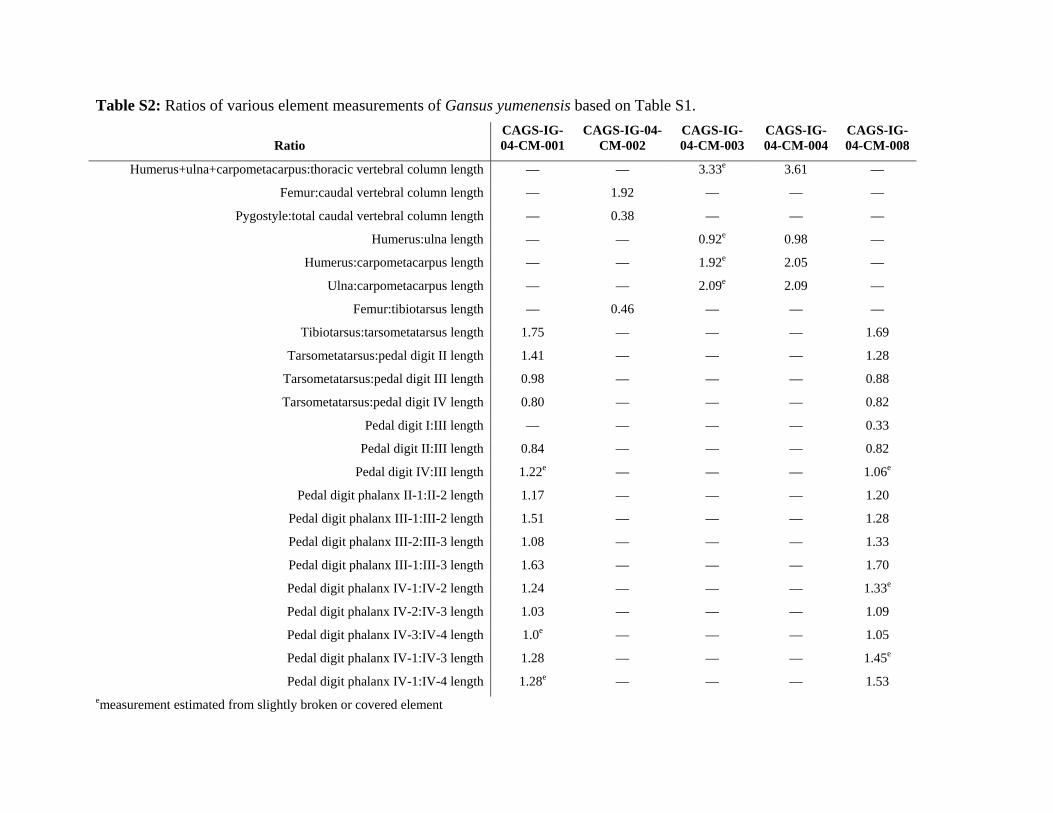
Table S2: Ratios of various element measurements of Gansus yumenensis based on Table S1.
Ratio CAGS-IG-04-CM-001
CAGS-IG-04-CM-002
CAGS-IG-04-CM-003
CAGS-IG-04-CM-004
CAGS-IG-04-CM-008
Humerus+ulna+carpometacarpus:thoracic vertebral column length — — 3.33e 3.61 —
Femur:caudal vertebral column length — 1.92 — — —
Pygostyle:total caudal vertebral column length — 0.38 — — —
Humerus:ulna length — — 0.92e 0.98 —
Humerus:carpometacarpus length — — 1.92e 2.05 —
Ulna:carpometacarpus length — — 2.09e 2.09 —
Femur:tibiotarsus length — 0.46 — — —
Tibiotarsus:tarsometatarsus length 1.75 — — — 1.69
Tarsometatarsus:pedal digit II length 1.41 — — — 1.28
Tarsometatarsus:pedal digit III length 0.98 — — — 0.88
Tarsometatarsus:pedal digit IV length 0.80 — — — 0.82
Pedal digit I:III length — — — — 0.33
Pedal digit II:III length 0.84 — — — 0.82
Pedal digit IV:III length 1.22e — — — 1.06e
Pedal digit phalanx II-1:II-2 length 1.17 — — — 1.20
Pedal digit phalanx III-1:III-2 length 1.51 — — — 1.28
Pedal digit phalanx III-2:III-3 length 1.08 — — — 1.33
Pedal digit phalanx III-1:III-3 length 1.63 — — — 1.70
Pedal digit phalanx IV-1:IV-2 length 1.24 — — — 1.33e
Pedal digit phalanx IV-2:IV-3 length 1.03 — — — 1.09
Pedal digit phalanx IV-3:IV-4 length 1.0e — — — 1.05
Pedal digit phalanx IV-1:IV-3 length 1.28 — — — 1.45e
Pedal digit phalanx IV-1:IV-4 length 1.28e — — — 1.53 emeasurement estimated from slightly broken or covered element

cc
sy
dp
B
A
C
Fig. S1. (A) Enlarged photograph of specimen CAGS-IG-04-CM-002. (B)Close-up of synsacrum and pelvis. (C) Close-up of proximal right tibiotarsus(arrow in A). Abbreviations as in Fig. 2.
5 cm
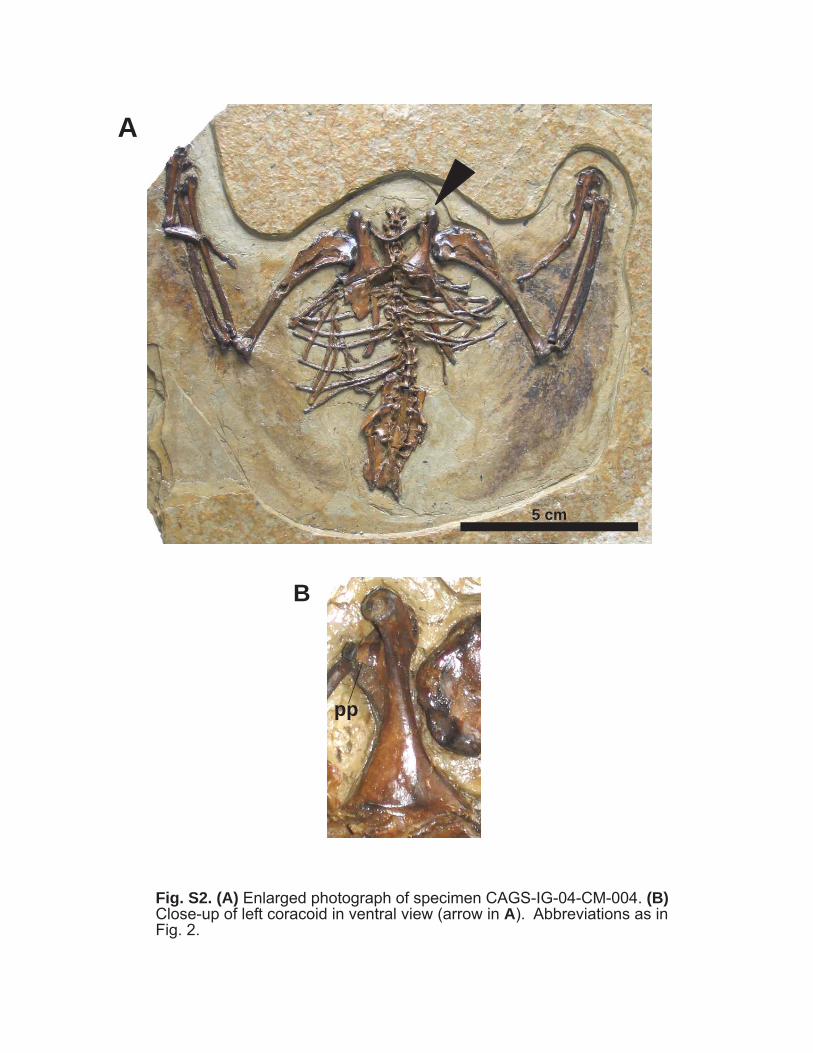
pp
B
Fig. S2. (A) Enlarged photograph of specimen CAGS-IG-04-CM-004. (B)Close-up of left coracoid in ventral view (arrow in A). Abbreviations as inFig. 2.
A
5 cm

Fig. S3. Enlarged photograph of specimen CAGS-IG-04-CM-001.
5 cm
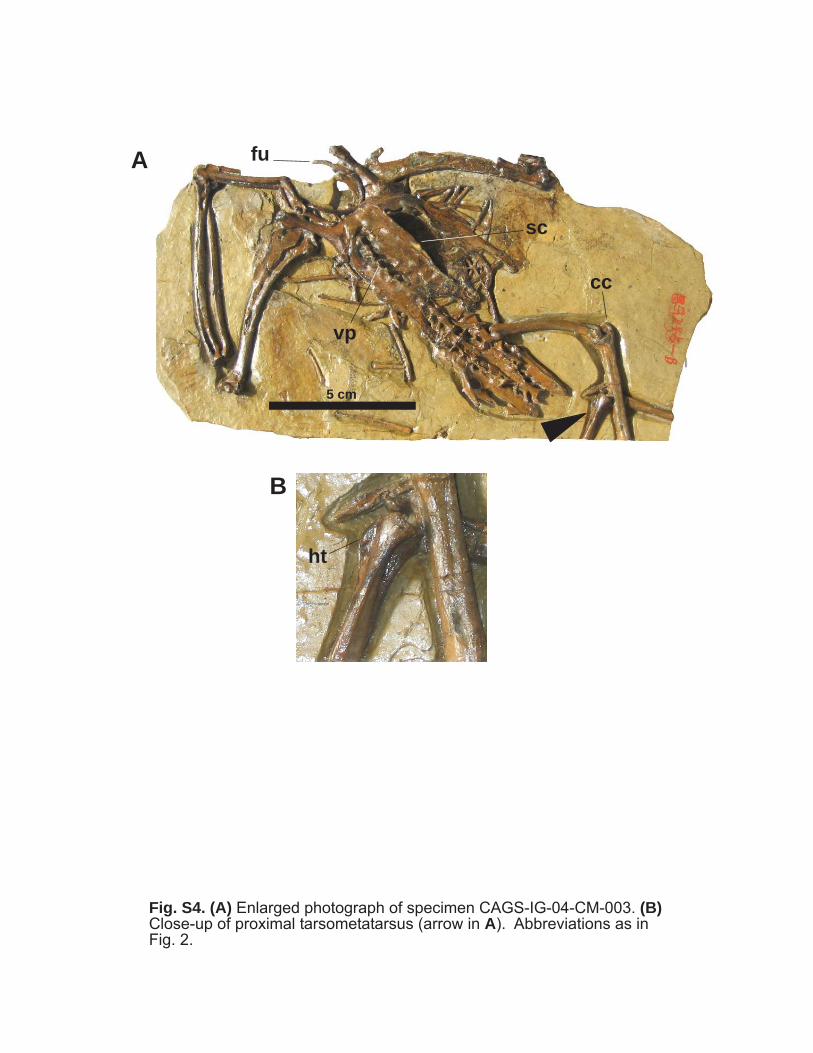
vp
sc
fu
cc
ht
A
B
Fig. S4. (A) Enlarged photograph of specimen CAGS-IG-04-CM-003. (B)Close-up of proximal tarsometatarsus (arrow in A). Abbreviations as inFig. 2.
5 cm

A
B
Fig. S5. (A) Enlarged photograph of specimen CAGS-IG-04-CM-008. (B)Close-up of tubercular soft-tissue preserved around toes (arrow in A).Abbreviations as in Fig. 2.
ft
5 cm
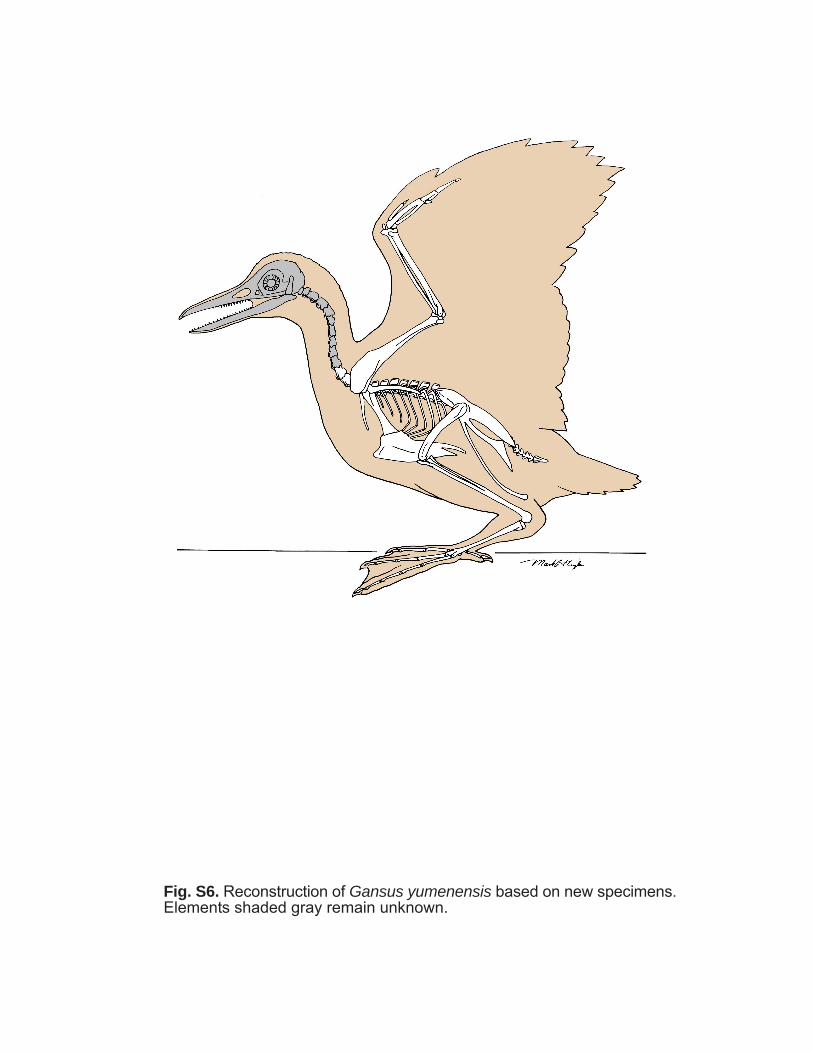
Fig. S6. Reconstruction of Gansus yumenensis based on new specimens.Elements shaded gray remain unknown.
Related Documents











