International Journal of Bifurcation and Chaos, Vol. 14, No. 2 (2004) 453–491 c World Scientific Publishing Company SUPER-SYNERGY IN THE BRAIN: THE GRANDMOTHER PERCEPT IS MANIFESTED BY MULTIPLE OSCILLATIONS EROL BAS ¸AR *,† and MURAT ¨ OZG ¨ OREN † Department of Biophysics, Dokuz Eyl¨ ul University, Faculty of Medicine, 35340 Izmir, Turkey * [email protected] SIREL KARAKAS ¸ † Department of Experimental Psychology, Hacettepe University, Ankara, Turkey † TUBITAK Brain Dynamics Multidisiplinary Research Network, Ankara, Turkey CANAN BAS ¸AR-ERO ˘ GLU Institute of Psychology and Cognition Research, University of Bremen, Bremen, Germany Received May 20, 2002; Revised November 1, 2002 The present report describes the dynamic foundations of long-standing experimental work in the field of oscillatory dynamics in the human and animal brain. It aims to show the role of multiple oscillations in the integrative brain function, memory, and complex perception by a recently introduced conceptional framework: the super-synergy in the whole brain. Results of recent experiments related to the percept of the grandmother-face support our concept of super- synergy in the whole brain in order to explain manifestation of Gestalts and Memory-Stages. This report may also provide new research avenues in macrodynamics of the brain. Keywords : EEG; ERP; oscillations; event related oscillations; alpha; beta; gamma; delta; theta; binding; superbinding; perception; memory; brain theory; coherence; wavelet analysis; wavelet entropy; grandmother percept. 1. Introduction ..................................... 455 1.1. Multiple oscillations in brain function ...................... 455 1.2. What is the role of oscillations in memory processing? .............. 455 1.3. Super-synergy in the whole brain and the grandmother percept ......... 456 2. Selectively Distributed Oscillatory Systems in Brain Function: Distributed Multiple Oscillations in Brains ................................ 457 2.1. Concept, definition, methods .......................... 457 2.2. Gamma oscillations in sensory, cognitive, and motor processes .......... 458 2.3. Alpha oscillations in perception and cognition: The alphas ............ 461 2.3.1. Sensory components ........................... 462 2.3.2. Cognitive components .......................... 464 2.4. Theta oscillations in perception and cognition .................. 466 2.5. Delta oscillations in cognition .......................... 468 * Author for correspondence. 453

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

March 5, 2004 9:23 00927
International Journal of Bifurcation and Chaos, Vol. 14, No. 2 (2004) 453–491c© World Scientific Publishing Company
SUPER-SYNERGY IN THE BRAIN: THEGRANDMOTHER PERCEPT IS MANIFESTED BY
MULTIPLE OSCILLATIONS
EROL BASAR∗,† and MURAT OZGOREN†
Department of Biophysics, Dokuz Eylul University, Faculty of Medicine, 35340 Izmir, Turkey∗[email protected]
SIREL KARAKAS†
Department of Experimental Psychology, Hacettepe University, Ankara, Turkey†TUBITAK Brain Dynamics Multidisiplinary Research Network, Ankara, Turkey
CANAN BASAR-EROGLUInstitute of Psychology and Cognition Research, University of Bremen, Bremen, Germany
Received May 20, 2002; Revised November 1, 2002
The present report describes the dynamic foundations of long-standing experimental work inthe field of oscillatory dynamics in the human and animal brain. It aims to show the role ofmultiple oscillations in the integrative brain function, memory, and complex perception by arecently introduced conceptional framework: the super-synergy in the whole brain. Results ofrecent experiments related to the percept of the grandmother-face support our concept of super-synergy in the whole brain in order to explain manifestation of Gestalts and Memory-Stages.This report may also provide new research avenues in macrodynamics of the brain.
Keywords : EEG; ERP; oscillations; event related oscillations; alpha; beta; gamma; delta; theta;binding; superbinding; perception; memory; brain theory; coherence; wavelet analysis; waveletentropy; grandmother percept.
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 455
1.1. Multiple oscillations in brain function . . . . . . . . . . . . . . . . . . . . . . 455
1.2. What is the role of oscillations in memory processing? . . . . . . . . . . . . . . 455
1.3. Super-synergy in the whole brain and the grandmother percept . . . . . . . . . 456
2. Selectively Distributed Oscillatory Systems in Brain Function: Distributed Multiple
Oscillations in Brains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 457
2.1. Concept, definition, methods . . . . . . . . . . . . . . . . . . . . . . . . . . 457
2.2. Gamma oscillations in sensory, cognitive, and motor processes . . . . . . . . . . 458
2.3. Alpha oscillations in perception and cognition: The alphas . . . . . . . . . . . . 461
2.3.1. Sensory components . . . . . . . . . . . . . . . . . . . . . . . . . . . 462
2.3.2. Cognitive components . . . . . . . . . . . . . . . . . . . . . . . . . . 464
2.4. Theta oscillations in perception and cognition . . . . . . . . . . . . . . . . . . 466
2.5. Delta oscillations in cognition . . . . . . . . . . . . . . . . . . . . . . . . . . 468
∗Author for correspondence.
453

March 5, 2004 9:23 00927
454 E. Basar et al.
3. Superposition Principle and Superimposed Multiple Oscillations in Theta and Delta
Frequency Windows in Cognitive Processes . . . . . . . . . . . . . . . . . . . . . . 470
4. Selectively Distributed and Selectively Coherent Oscillatory Networks . . . . . . . . . 472
5. Distributed Oscillatory Systems in the Brain and Distributed Memory . . . . . . . . . 476
5.1. Event-processing in distributed systems . . . . . . . . . . . . . . . . . . . . . 476
5.2. Memory and information: Working memory systems and long term memory
system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 476
5.2.1. fMRI studies related to memory function . . . . . . . . . . . . . . . . . 476
5.3. EEG-oscillations and memory . . . . . . . . . . . . . . . . . . . . . . . . . . 477
5.4. Multiple functions of event-related oscillations and multiple functions of memory:
Convergence of concepts . . . . . . . . . . . . . . . . . . . . . . . . . . . . 478
6. The “Super-Synergy” is a Spatiotemporal and Functional Organization of Multiple and
Distributed Oscillations in the Brain . . . . . . . . . . . . . . . . . . . . . . . . . 479
6.1. Preliminary hypothesis for “Grandmother cell or grandmother cell assemblies” . . 481
7. Grandmother-Paradigm and Gestalt-Experiments . . . . . . . . . . . . . . . . . . . 483
7.1. Experimental strategy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 483
7.1.1. Electrophysiological recording . . . . . . . . . . . . . . . . . . . . . . 483
7.2. Event related oscillations to all three stimulations: Simple-light, anonymous face and
the grandmother picture . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485
7.2.1. Topology of Delta responses . . . . . . . . . . . . . . . . . . . . . . . 485
7.2.2. Topology of Theta responses . . . . . . . . . . . . . . . . . . . . . . . 485
7.2.3. Topology of Alpha responses . . . . . . . . . . . . . . . . . . . . . . . 485
7.3. Distributed Beta and Gamma responses . . . . . . . . . . . . . . . . . . . . . 486
7.4. What are the grandmother experiments saying? A distributed template of memory
reactivation? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 486
7.4.1. Selectively distributed enhancements in the whole cortex . . . . . . . . . 486
7.4.2. The distinction between episodic and semantic memory . . . . . . . . . . 486
7.5. The efficiency of the grandmother paradigm for differentiation of memory components
or stages . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 487
8. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 487
Acknowledgment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 487
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 487
Abbreviations
AFC: Amplitude Frequency Characteristics STM: Short Term MemoryEP: Evoked potential TRFC: Transient Response-Frequency Character-ERP: Event related potential istics MethodERO: Event related Oscillation VEP: Visual Evoked PotentialfMRI: functional Magnetic Resonance Imaging WMS : Working Memory SystemLTM: Long Term Memory 3.ATT: The third attended signal in the omittedMEG: Magnetoencephalography signal paradigm. (last auditory stimulationMMN: Mismatch Negativity before omitted one)OB: Oddball

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 455
Anatomical Abbreviations
GEA: Gyrus Ectosylvian AnteriorHI: HippocampusIC: Inferior ColliculusLG: Lateral Geniculate NucleusMG: Medial geniculate NucleusOC: Occipital CortexRF: Reticular FormationSC: Superior ColliculusTH: Thalamus
1. Introduction
The present report aims to show the role ofmultiple oscillations in the integrative brain func-tion, memory, and complex perception through arecently introduced conceptional framework: thesuper-synergy in the whole brain. The contentsof this paper comprehend presentation and newsynthesis of experimental work, covering 30 yearsof experience of our research groups. Results ofrecent experiments related to the percept of thegrandmother-face support our concept of super-synergy in the whole brain and may provide newresearch avenues in macrodynamics of the brain.The tutorial companion report [Basar, 2004], whichdescribes some of the important developments forthe understanding of integrative brain function,may also provide a useful reading to explain therationale of starting with the type of experimentspresented in this report.
1.1. Multiple oscillations in brain
function
The new trend towards the treatise of brain os-cillations imply the following immediate thoughts:(1) Not only single neurons, but neurons assem-blies, (2) not only spikes of single neurons, but os-cillatory activity of neurons and neural assemblies,(3) not only movements but cognitive processes andmemory processes are interwoven in the integrativebrain function. These concepts have not been in-cluded in Sherrington’s description of integrativebrain activity, and neuroscience needs a new frame-work or theories and possibly new rules to analyzethe integrative brain function.
The functional importance of distributed mul-tiple oscillations in the brain was first emphasizedin a series of reports of our research group [Basar
& Ungan, 1973; Basar et al., 1975; Basar, 1992].Further, the relevance of the superposition princi-ple and long distance synchronization in the brainwas also shown already in 1970s [Basar et al., 1979;Basar, 1980]. The concerted activity of alpha, theta,delta, beta and gamma oscillations was measured instructures as reticular formation (RF), Hippocam-pus (HI), thalamus and sensory cortices, and itwas assumed that this fact is one of the impor-tant frameworks in brain’s signal processing. Thetrend in analyzing the integrative brain functionsby means of multiple oscillations and long distancecoherences is rapidly increasing [Klimesch et al.,2000; Gruzelier et al., 1996; Bullock, 1992].
A fundamental unsolved process in neuro-science concerns the manner in which the vastarray of parallel processing occurs in the brain atany given time, the diverse neural activities arebound together or integrated [Haig et al., 2000].For instance, a visual image of an object containsa collection of features which must be identifiedand segregated from those comprising other objects.Generally it is assumed that different features ofthe image are processed by different areas of thebrain. How then is the spatially distributed process-ing relating to one percept integrated? This processis known as the binding problem [Singer & Gray,1995].
1.2. What is the role of oscillations
in memory processing?
According to Fuster’s [1997] view memory reflectsa distributed property of cortical systems. Animportant part of higher nervous function, as per-ception, recognition, language, planning, problemsolving and decision-making, is interwoven withmemory. Memory is a property of the neurobio-logical systems it serves and inseparable from theirother functions. By surveying the data presented inthis review it can be hypothesized that the selec-tively distributed oscillatory systems (or networks)may provide a general communication frameworkand can be a useful concept for functional map-ping of the brain [Mesulam, 1990, 1994]. As earlyas 1985 we have used the expression “dynamic mem-ory” for cognitive performances of short states thatare manifested with short regular oscillatory states.Experimental designs by Klimesch and Gruzelierand co-workers [Klimesch et al., 1994; Klimesch,1999; Burgess & Gruzelier, 2000; Haenschel et al.,2000; Egner & Gruzelier, 2001] are extremely

March 5, 2004 9:23 00927
456 E. Basar et al.
n sweeps(e.g. n = 100)
selectiveaveraging
average EP
averagingacross subjects(in time domain)
grandaverage
FFT
digitalfiltering
FFT
digitalfiltering
FFT
digitalfiltering
frequencycomponents
of grand average
AFC of grand average
frequencycomponents
of average EP / ERP
AFC of averagedEP / ERP
frequency componentsof single sweeps
enhancementfactors etc.
power spectrumof pre- and
poststimulus EEG
averagingacross subjects
(in frequencydomain)
mean value of AFCs(spectrum average)
Fig 1 Basar et al
Fig. 1. Schematical illustration of showing the stepwise application of system-theoretical methods used in this report. For thesake of simplicity evaluation of wavelet decomposition and coherence functions are not included here. Details of the methodsare in [Basar, 1980, 1998; Schurmann et al., 2000; Basar et al., 2001].
important by adding new steps to the general frame-work of function-related oscillations. This view isin accordance of Basar et al. [1999, 2000] em-phasizing, in the line of Fuster-statements, thatevery oscillation-response is perception and memoryrelated.
1.3. Super-synergy in the whole brain
and the grandmother percept
The results of a broad range of papers surveyed inthe present report suggest that the binding prob-
lem cannot be solved solely with the oscillatorydynamics in the narrow gamma band. Accordingly,we suggest that the super-synergy of at least sixsubmechanisms in the whole brain may present amore plausible approach for integration in the brain.Our recent experiments introducing a new strategyof measurements indicate that the percept of grand-mother is manifested with multiple oscillations inthe whole brain, thus supporting the newly intro-duced super-synergy concept [Basar et al., 2001a,2001d].
March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 457
2. Selectively Distributed Oscillatory
Systems in Brain Function:
Distributed Multiple Oscillations
in Brains
2.1. Concept, definition, methods
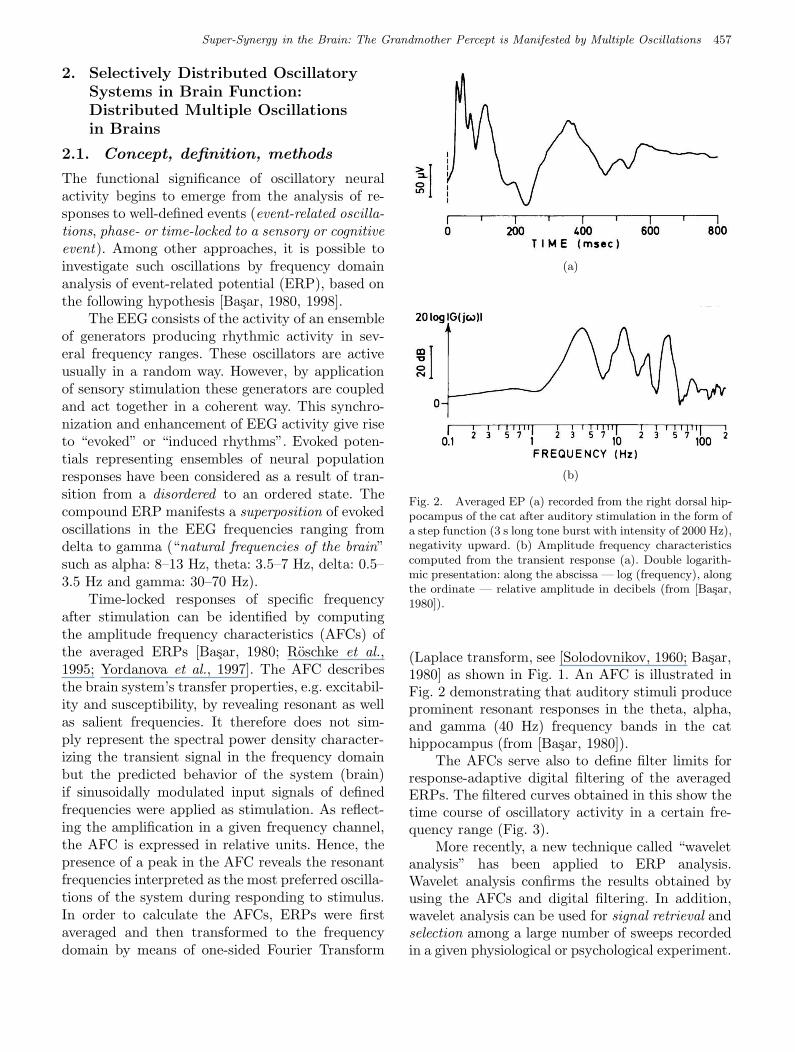
The functional significance of oscillatory neuralactivity begins to emerge from the analysis of re-sponses to well-defined events (event-related oscilla-tions, phase- or time-locked to a sensory or cognitiveevent). Among other approaches, it is possible toinvestigate such oscillations by frequency domainanalysis of event-related potential (ERP), based onthe following hypothesis [Basar, 1980, 1998].
The EEG consists of the activity of an ensembleof generators producing rhythmic activity in sev-eral frequency ranges. These oscillators are activeusually in a random way. However, by applicationof sensory stimulation these generators are coupledand act together in a coherent way. This synchro-nization and enhancement of EEG activity give riseto “evoked” or “induced rhythms”. Evoked poten-tials representing ensembles of neural populationresponses have been considered as a result of tran-sition from a disordered to an ordered state. Thecompound ERP manifests a superposition of evokedoscillations in the EEG frequencies ranging fromdelta to gamma (“natural frequencies of the brain”such as alpha: 8–13 Hz, theta: 3.5–7 Hz, delta: 0.5–3.5 Hz and gamma: 30–70 Hz).
Time-locked responses of specific frequencyafter stimulation can be identified by computingthe amplitude frequency characteristics (AFCs) ofthe averaged ERPs [Basar, 1980; Roschke et al.,1995; Yordanova et al., 1997]. The AFC describesthe brain system’s transfer properties, e.g. excitabil-ity and susceptibility, by revealing resonant as wellas salient frequencies. It therefore does not sim-ply represent the spectral power density character-izing the transient signal in the frequency domainbut the predicted behavior of the system (brain)if sinusoidally modulated input signals of definedfrequencies were applied as stimulation. As reflect-ing the amplification in a given frequency channel,the AFC is expressed in relative units. Hence, thepresence of a peak in the AFC reveals the resonantfrequencies interpreted as the most preferred oscilla-tions of the system during responding to stimulus.In order to calculate the AFCs, ERPs were firstaveraged and then transformed to the frequencydomain by means of one-sided Fourier Transform
Fig 2 Basar et al.
(a)
Fig 2 Basar et al.
(b)
Fig. 2. Averaged EP (a) recorded from the right dorsal hip-pocampus of the cat after auditory stimulation in the form ofa step function (3 s long tone burst with intensity of 2000 Hz),negativity upward. (b) Amplitude frequency characteristicscomputed from the transient response (a). Double logarith-mic presentation: along the abscissa — log (frequency), alongthe ordinate — relative amplitude in decibels (from [Basar,1980]).
(Laplace transform, see [Solodovnikov, 1960; Basar,1980] as shown in Fig. 1. An AFC is illustrated inFig. 2 demonstrating that auditory stimuli produceprominent resonant responses in the theta, alpha,and gamma (40 Hz) frequency bands in the cathippocampus (from [Basar, 1980]).
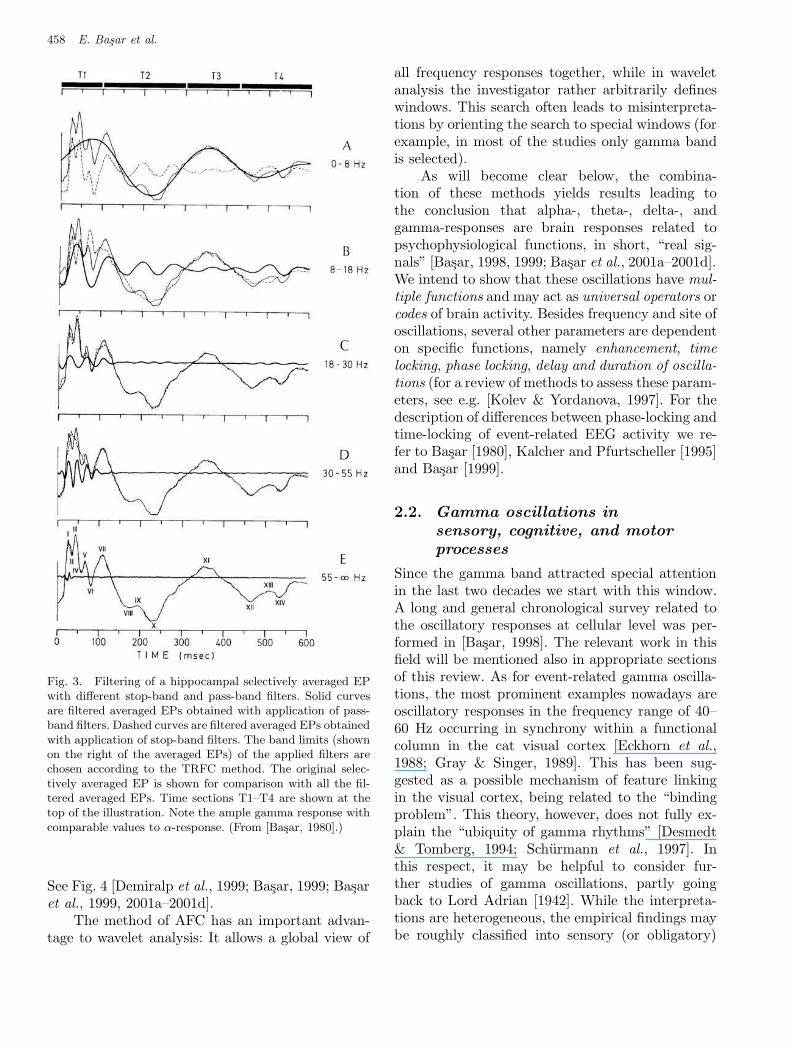
The AFCs serve also to define filter limits forresponse-adaptive digital filtering of the averagedERPs. The filtered curves obtained in this show thetime course of oscillatory activity in a certain fre-quency range (Fig. 3).
More recently, a new technique called “waveletanalysis” has been applied to ERP analysis.Wavelet analysis confirms the results obtained byusing the AFCs and digital filtering. In addition,wavelet analysis can be used for signal retrieval andselection among a large number of sweeps recordedin a given physiological or psychological experiment.
March 5, 2004 9:23 00927
458 E. Basar et al.
Fig 3 Basar et al.
Fig. 3. Filtering of a hippocampal selectively averaged EPwith different stop-band and pass-band filters. Solid curvesare filtered averaged EPs obtained with application of pass-band filters. Dashed curves are filtered averaged EPs obtainedwith application of stop-band filters. The band limits (shownon the right of the averaged EPs) of the applied filters arechosen according to the TRFC method. The original selec-tively averaged EP is shown for comparison with all the fil-tered averaged EPs. Time sections T1–T4 are shown at thetop of the illustration. Note the ample gamma response withcomparable values to α-response. (From [Basar, 1980].)
See Fig. 4 [Demiralp et al., 1999; Basar, 1999; Basaret al., 1999, 2001a–2001d].
The method of AFC has an important advan-tage to wavelet analysis: It allows a global view of
all frequency responses together, while in waveletanalysis the investigator rather arbitrarily defineswindows. This search often leads to misinterpreta-tions by orienting the search to special windows (forexample, in most of the studies only gamma bandis selected).
As will become clear below, the combina-tion of these methods yields results leading tothe conclusion that alpha-, theta-, delta-, andgamma-responses are brain responses related topsychophysiological functions, in short, “real sig-nals” [Basar, 1998, 1999; Basar et al., 2001a–2001d].We intend to show that these oscillations have mul-tiple functions and may act as universal operators orcodes of brain activity. Besides frequency and site ofoscillations, several other parameters are dependenton specific functions, namely enhancement, timelocking, phase locking, delay and duration of oscilla-tions (for a review of methods to assess these param-eters, see e.g. [Kolev & Yordanova, 1997]. For thedescription of differences between phase-locking andtime-locking of event-related EEG activity we re-fer to Basar [1980], Kalcher and Pfurtscheller [1995]and Basar [1999].
2.2. Gamma oscillations in
sensory, cognitive, and motor
processes
Since the gamma band attracted special attentionin the last two decades we start with this window.A long and general chronological survey related tothe oscillatory responses at cellular level was per-formed in [Basar, 1998]. The relevant work in thisfield will be mentioned also in appropriate sectionsof this review. As for event-related gamma oscilla-tions, the most prominent examples nowadays areoscillatory responses in the frequency range of 40–60 Hz occurring in synchrony within a functionalcolumn in the cat visual cortex [Eckhorn et al.,1988; Gray & Singer, 1989]. This has been sug-gested as a possible mechanism of feature linkingin the visual cortex, being related to the “bindingproblem”. This theory, however, does not fully ex-plain the “ubiquity of gamma rhythms” [Desmedt& Tomberg, 1994; Schurmann et al., 1997]. Inthis respect, it may be helpful to consider fur-ther studies of gamma oscillations, partly goingback to Lord Adrian [1942]. While the interpreta-tions are heterogeneous, the empirical findings maybe roughly classified into sensory (or obligatory)

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 459
Fig 4 Basar et al.
Fig. 4. Results of wavelet decompostion in a typical animal. (Left) Auditory stimulation; (right) visual stimulation. Eachcolumn refers to an electrode site (auditory cortex: GEA; visual cortex: OC; hippocampus: HI). The uppermost row shows thewideband filtered curve. Further rows show the frequency components gamma (32–64 Hz), beta (16–32 Hz), alpha (8–16 Hz),theta (4–8 Hz), and delta (0.5–4 Hz). (Modified from [Basar et al., 1999].)
versus cognitive gamma responses. Some examplesof sensory functions follow here:
Auditory and visual gamma EEG responses areselectively distributed in different cortical and sub-cortical structures: They are phase-locked stablecomponents of EPs in cortex, hippocampus, brainstem, and cerebellum of cats occurring 100 ms af-ter the sensory stimulation with a second windowof approx. 300 ms latency (see Fig. 5) [Basar, 1980;Basar et al., 1987; Basar, 1999]. The earliest studyof gamma auditory response has been recorded byBasar and Ozesmi [1972] and Basar and Ungan[1973] in cat hippocampus and later confirmed byLeung et al. [1992]. A recent review [Keil et al.,2001] presents evidences supporting the fact thatgamma oscillations are involved in several aspectsof visual processing.
(1) Sensory processes: A phase-locked gammaoscillation is also a component of the human au-ditory and visual response as Fig. 6 shows [Basaret al., 1987; Gurtubay et al., 2001]. A new strategyby application of six cognitive paradigms showedthat the 40 Hz response in the 100 ms after stim-ulations has a sensory origin, being independent ofcognitive tasks [Karakas & Basar, 1998].
(2) The auditory MEG gamma response issimilar to human EEG responses with a close re-lationship to the middle latency auditory evokedresponse [Pantev et al., 1991].
(3) Cognitive processes: Several investigationsdealt with cognitive processes related to gammaresponses, some of them based on measuring theP300 wave. This positive deflection typically occursin human ERPs in response to “oddball” stimuli

March 5, 2004 9:23 00927
460 E. Basar et al.
Fig 5 Basar et al.
Fig. 5. Responses to auditory stimuli in single cats, filtered in gamma band (30–70 Hz) auditory responses of single cats inGEA, RF, HI and CE. The gamma band filtered grand averages of responses are shown in the bottom row. Note the largegamma responses also in the cerebellum. (Modified from [Basar et al., 1995].)
- 300 -200 -100 0 100 200 ms
4µV-
+
A
30 - 50 Hz
*
4µV-
+
B
**- 300 -200 -100 0 100 200 ms
30 - 50 Hz
C
4µV-
+
4µV-
+
4µV-
+
4µV-
+
- 50 0 50 100 150 200 ms
30 - 50 Hz
30 - 50 Hz
1 - 200 Hz
30 - 50 Hz
1 - 200 Hz
30 - 50 Hz
1 - 200 Hz
1 - 200 Hz
(a)
(b)
(c)
(d)
(e)
(f)
(g)
(h)
Fig 6 Basar et al.
Fig. 6. Ten randomly selected single EEG-EP trials filtered with digital filters of 30–50 Hz. ∗, average of these trials.Stimulation is applied at time “0 ms”. (B) Ten single EEG-EP trials, digitally filtered (30–50 Hz) and selected for highenhancement, i.e. high amplitude increase after stimulus (in comparison with prestimulus EEG amplitude). ∗∗, average ofthese trials. (C) Selectively averaged evoked potentials (EPs); averages of 40 single trials each, with different selection of trials(human vertex recordings). (a) EP averaged from randomly selected single trials (“conventional” averaged EP); (b) sameEP, filtered 30–50 Hz. (c) EP averaged from single trials selected for specific criteria (marked amplitude enhancement
in the 40 Hz range after stimulation); (d) same EP, filtered with band limits 30–50 Hz. (e) EP averaged from single trialswith low amplitude enhancement; (f) same EP, filtered with band limits 30–50 Hz. (g) Identical with (e) — average of lowenhancement trials; (h) result of applying a 30–50 Hz stop-band filter (which theoretically rejects the 40 Hz response) to theconventional averaged EP of (a). Note the similarity of (h), obtained with stop band filtering, and (g), obtained with lowenhancement trials. (Modified from [Basar et al., 1987] Int. J. Neurosci. 33, 103–117.)

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 461
P300 - 40 Hz RESPONSE IN HIPPOCAMPUS
50
45
40
.
.
5
10
15
.
.
0 100 300 500time (ms)
0 100 300 500time (ms)
I
I
I
+
+
+
-
-
-
-
-
-
-
-
-
10µV
2.5µV
25µV
Single Sweeps(30 - 50) Hz
Averaged ERP(30 - 50) Hz
AveragedERP(1 - 100 Hz)
Fig 7 Basar et al.
Fig. 7. Event-related potentials of lower pyramidal layer(CA3) of hippocampus (one cat). (Top) Single ERP sweeps(epochs) filtered at 30–50 Hz. (Middle) Averaged ERP fil-tered at 30–50 Hz. (Bottom) Unfiltered ERP, average of 50artefact-free epochs. (Modified from [Basar-Eroglu & Basar,1991] Int. J. Neurosci. 60, 239–248.)
or omitted stimuli interspersed as “targets” into aseries of standard stimuli:
A P300-40 Hz component has been recordedin the cat hippocampus, reticular formation andcortex (with omitted auditory stimuli as targets).This response occurs approx. 300 ms after stimula-tion, being superimposed with a slow wave of 4 Hzillustrated in Fig. 7 [Basar-Eroglu & Basar, 1991].Preliminary data indicate similar P300-40 Hz re-sponses to oddball stimuli in humans [Basar-Erogluet al., 1992]. However, a suppression of 40 Hz ac-tivity after target stimuli has also been reported[Fell et al., 1997]. In a recent study the gamma
band activity in an auditory oddball paradigm wasanalyzed with the wavelet transform. A late os-cillatory peaking at 37 Hz with a latency around360 ms was observed appearing only for targetstimuli [Gurtubay et al., 2001]. The superimposedtheta-gamma complex pattern is analyzed recentlyalso in modeling studies [Baird, 1999; Kiss et al.,2001]. A study in the human limbic system alsoconfirms the synchronization of gamma rhythmsduring cognitive tasks [Fell et al., 2001].
During visual perception of reversible or am-biguous figures a significant increase (almost 50%)in human frontal gamma EEG activity has beenrecorded [Basar-Eroglu et al., 1996b]. The spa-tiotemporal magnetic field pattern of gamma bandactivity has been interpreted as a coherent rostro-caudal “sweep of activity” [Llinas & Ribary, 1992].Sakowitz et al. [2001] reported significant gammaresponse amplitude increases in distributed areasof the brain and a 100% frontal theta enhance-ment by bimodal stimulation compared to unimodalone. Tallon-Baudry et al. [1998] reported that re-trieval of visual experience from short-term memoryis associated with 40 Hz activity.
Further, experimental data indicates that elec-trical stimulation evokes gamma oscillations also inisolated invertebrate ganglia. This finding clearlyshows that, in absence of perceptual binding orhigher cognitive processes the gamma response isstill present [Schutt et al., 1999]. In conclusion,gamma oscillations possibly represent a universalcode of CNS communication [Basar, 1998, 1999].This view might also serve as a synthesis overcom-ing controversies in earlier reports. See also [Basar& Bullock, 1992; Basar-Eroglu et al., 1996a; Basaret al., 2001a–2001d].
2.3. Alpha oscillations in
perception and cognition: The
alphas
Alpha oscillations in functional EEG gained im-portance in the last decade as we indicated as a“renaissance of alpha” [Basar et al., 1997]. Obser-vations at the cellular level are noteworthy: Evokedoscillations in the 8–10 Hz frequency range in vi-sual cortex neurons upon visual stimulation sug-gest a relation to scalp-recordable alpha responses[Silva et al., 1991; Dinse et al., 1997]. Thalamo-cortical neurons are discharged in the alpha band[Steriade et al., 1990]. Alpha responses from dif-ferent structures of the cat brain are presented in
March 5, 2004 9:23 00927
462 E. Basar et al.
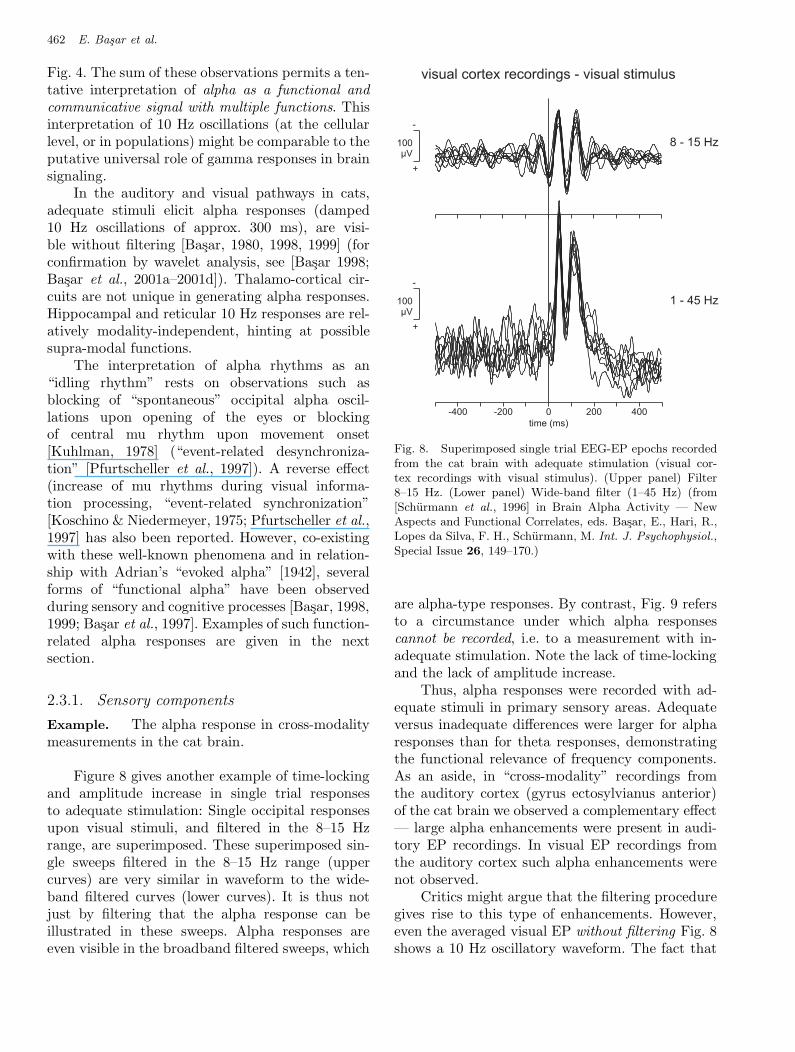
Fig. 4. The sum of these observations permits a ten-tative interpretation of alpha as a functional andcommunicative signal with multiple functions. Thisinterpretation of 10 Hz oscillations (at the cellularlevel, or in populations) might be comparable to theputative universal role of gamma responses in brainsignaling.
In the auditory and visual pathways in cats,adequate stimuli elicit alpha responses (damped10 Hz oscillations of approx. 300 ms), are visi-ble without filtering [Basar, 1980, 1998, 1999] (forconfirmation by wavelet analysis, see [Basar 1998;Basar et al., 2001a–2001d]). Thalamo-cortical cir-cuits are not unique in generating alpha responses.Hippocampal and reticular 10 Hz responses are rel-atively modality-independent, hinting at possiblesupra-modal functions.
The interpretation of alpha rhythms as an“idling rhythm” rests on observations such asblocking of “spontaneous” occipital alpha oscil-lations upon opening of the eyes or blockingof central mu rhythm upon movement onset[Kuhlman, 1978] (“event-related desynchroniza-tion” [Pfurtscheller et al., 1997]). A reverse effect(increase of mu rhythms during visual informa-tion processing, “event-related synchronization”[Koschino & Niedermeyer, 1975; Pfurtscheller et al.,1997] has also been reported. However, co-existingwith these well-known phenomena and in relation-ship with Adrian’s “evoked alpha” [1942], severalforms of “functional alpha” have been observedduring sensory and cognitive processes [Basar, 1998,1999; Basar et al., 1997]. Examples of such function-related alpha responses are given in the nextsection.
2.3.1. Sensory components
Example. The alpha response in cross-modalitymeasurements in the cat brain.
Figure 8 gives another example of time-lockingand amplitude increase in single trial responsesto adequate stimulation: Single occipital responsesupon visual stimuli, and filtered in the 8–15 Hzrange, are superimposed. These superimposed sin-gle sweeps filtered in the 8–15 Hz range (uppercurves) are very similar in waveform to the wide-band filtered curves (lower curves). It is thus notjust by filtering that the alpha response can beillustrated in these sweeps. Alpha responses areeven visible in the broadband filtered sweeps, which
visual cortex recordings - visual stimulus
100µV
100µV
8 - 15 Hz
1 - 45 Hz
-400 -200 0 200 400
time (ms)
-
+
+
-
Fig 8 Basar et al.
Fig. 8. Superimposed single trial EEG-EP epochs recordedfrom the cat brain with adequate stimulation (visual cor-tex recordings with visual stimulus). (Upper panel) Filter8–15 Hz. (Lower panel) Wide-band filter (1–45 Hz) (from[Schurmann et al., 1996] in Brain Alpha Activity — NewAspects and Functional Correlates, eds. Basar, E., Hari, R.,Lopes da Silva, F. H., Schurmann, M. Int. J. Psychophysiol.,Special Issue 26, 149–170.)
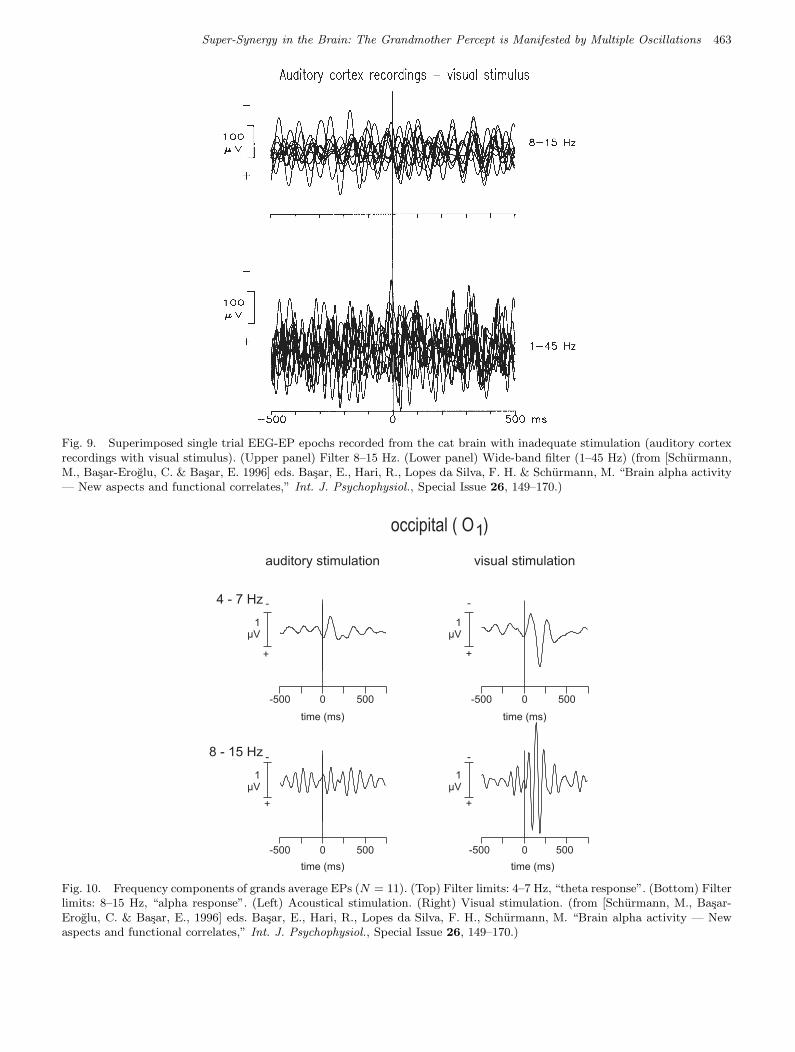
are alpha-type responses. By contrast, Fig. 9 refersto a circumstance under which alpha responsescannot be recorded, i.e. to a measurement with in-adequate stimulation. Note the lack of time-lockingand the lack of amplitude increase.
Thus, alpha responses were recorded with ad-equate stimuli in primary sensory areas. Adequateversus inadequate differences were larger for alpharesponses than for theta responses, demonstratingthe functional relevance of frequency components.As an aside, in “cross-modality” recordings fromthe auditory cortex (gyrus ectosylvianus anterior)of the cat brain we observed a complementary effect— large alpha enhancements were present in audi-tory EP recordings. In visual EP recordings fromthe auditory cortex such alpha enhancements werenot observed.
Critics might argue that the filtering proceduregives rise to this type of enhancements. However,even the averaged visual EP without filtering Fig. 8shows a 10 Hz oscillatory waveform. The fact that
March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 463
Fig 9 Basar et al.
Fig. 9. Superimposed single trial EEG-EP epochs recorded from the cat brain with inadequate stimulation (auditory cortexrecordings with visual stimulus). (Upper panel) Filter 8–15 Hz. (Lower panel) Wide-band filter (1–45 Hz) (from [Schurmann,M., Basar-Eroglu, C. & Basar, E. 1996] eds. Basar, E., Hari, R., Lopes da Silva, F. H. & Schurmann, M. “Brain alpha activity— New aspects and functional correlates,” Int. J. Psychophysiol., Special Issue 26, 149–170.)
occipital ( O )1
auditory stimulation visual stimulation
4 - 7 Hz
8 - 15 Hz
- -
- -
+ +
+ +
-500
-500
-5000
0
0500
500
500
-500 0 500
time (ms)
time (ms)
time (ms)
time (ms)
1µV
1µV
1µV
1µV
Fig 10 Basar et al.
Fig. 10. Frequency components of grands average EPs (N = 11). (Top) Filter limits: 4–7 Hz, “theta response”. (Bottom) Filterlimits: 8–15 Hz, “alpha response”. (Left) Acoustical stimulation. (Right) Visual stimulation. (from [Schurmann, M., Basar-Eroglu, C. & Basar, E., 1996] eds. Basar, E., Hari, R., Lopes da Silva, F. H., Schurmann, M. “Brain alpha activity — Newaspects and functional correlates,” Int. J. Psychophysiol., Special Issue 26, 149–170.)

March 5, 2004 9:23 00927
464 E. Basar et al.
this is absolutely not the case is shown by severalstudies. The reader is referred to different reviewsby Basar [1998] and Basar et al. [2001].
Examples. Alpha responses in human EEG andMEG in cross-modality experiments.
It is useful to compare the cat data to EEGand MEG recordings in humans. EEG measure-ments were performed in N = 11 subjects [Basar &Schurmann, 1994]. Figure 10 shows filtered curvescomputed from grand averages of occipital record-ings (O1). The upper half of the figure shows thetaresponses whereas the lower half shows alpha re-sponses. The alpha response to auditory stimulation(inadequate for the visual cortex, occipital located)is on the left, where the response is of low ampli-tude. The response to visual stimulation, however,is on the right, with a distinct alpha response. Notethat the adequate–inadequate difference is less forthe theta response. The hypothesis as given previ-ously is thus supported: As observed in cats it ismainly the alpha response, which is dependent onwhether or not a stimulus is adequate. A correlationbetween the alpha response and primary sensoryprocessing is thus plausible both for human and forcat EEG-EP data.
MEG measurements were performed both witha BTI 7 channel MEG system [Saermark et al.,1992] and with a PHILIPS 19-channel MEG sys-tem [Schurmann et al., 1992; Basar et al., 1992].The methods used were similar to those used forEEG recordings where possible. We used auditorystimuli (2000 Hz; 80 dB sound pressure level) andselected sensor positions close to the auditory cor-tex and close to the visual cortex.
The data shown in Fig. 11 were obtained withthe 7-channel system where the different positionsrequired two experimental sessions. Panel A showstemporal recordings, Panel B occipital recordings,in both cases with auditory stimuli. The underly-ing cortical areas being the primary auditory cor-tex and the primary visual cortex, auditory stimuliare regarded as adequate in the first case (Fig. 11,panel A) and as inadequate in the second case(Fig. 11, panel B). High amplitude alpha responsesare visible in panel A with adequate stimulation. Incontrast, panel B with inadequate stimulation doesnot show such alpha responses.
2.3.2. Cognitive components
Cognitive targets significantly influence the alpharesponses in P300: Using an oddball paradigm, pro-
January 7, 2004 9:48 WSPC/102-IDAQPRT 00150
Logarithmic Sobolev Inequality forHs
0-Metric on Pinned Loop Groups 13
A
B
8 - 15 Hz
80fT
-800 0 800
time (ms)
80fT
-800 0 800
time (ms)
Fig 11 Basar et al.
Here gx = g is the process defined by (4.2). We set Mx(t, τ) = ddε
∣
∣
∣
∣
ε=0
gx,ε(t, τ) and
qz(t)X = −[Utzt, X ]− DX(Utzt)
for X ∈ Hs0 . Then qz(t) is a bounded linear operator on Hs
0 . We denote the eval-
uation at τ ∈ S1 by δτ ∈ L0(R)∗ ⊂ Hs0(R)∗ = Hs
0(R). Then we may consider
δτ ⊗ ei ∈ Hs0 .
The following proposition is due to Fang.?,?
(a)
A
B
8 - 15 Hz
80fT
-800 0 800
time (ms)
80fT
-800 0 800
time (ms)
Fig 11 Basar et al.
(b)
Fig. 11. Human MEG responses to auditory stimulation:averaged evoked fields recorded in a typical subject (filterlimits: 8–15 Hz). (a) 7 channels with “pure temporal” lo-cation. (b) 7 channels with “pure occipital” location (from[Schurmann, M., Basar-Eroglu, C. & Basar, E., 1996] eds.Basar, E., Hari, R., Lopes da Silva, F. H. & Schurmann, M.“Brain alpha activity — New aspects and functional corre-lates,” Int. J. Psychophysiol., Special Issue 26, 149–170.)
longed event-related alpha oscillations up to 400 mswere observed as firstly published by Stampfer andBasar [1985] and later with a clear demonstra-tion by means of single sweep analysis by Kolevet al. [1999] (see also [Basar, 1998, 1999]). Memoryrelated event-related alpha oscillations can be ob-served in well-trained subjects one second beforean expected target [Maltseva, 2000]. New results[Klimesch et al., 1994; Basar et al., 1997] demon-strate that alpha activity is strongly correlatedwith working memory and probably with long termmemory engrams.
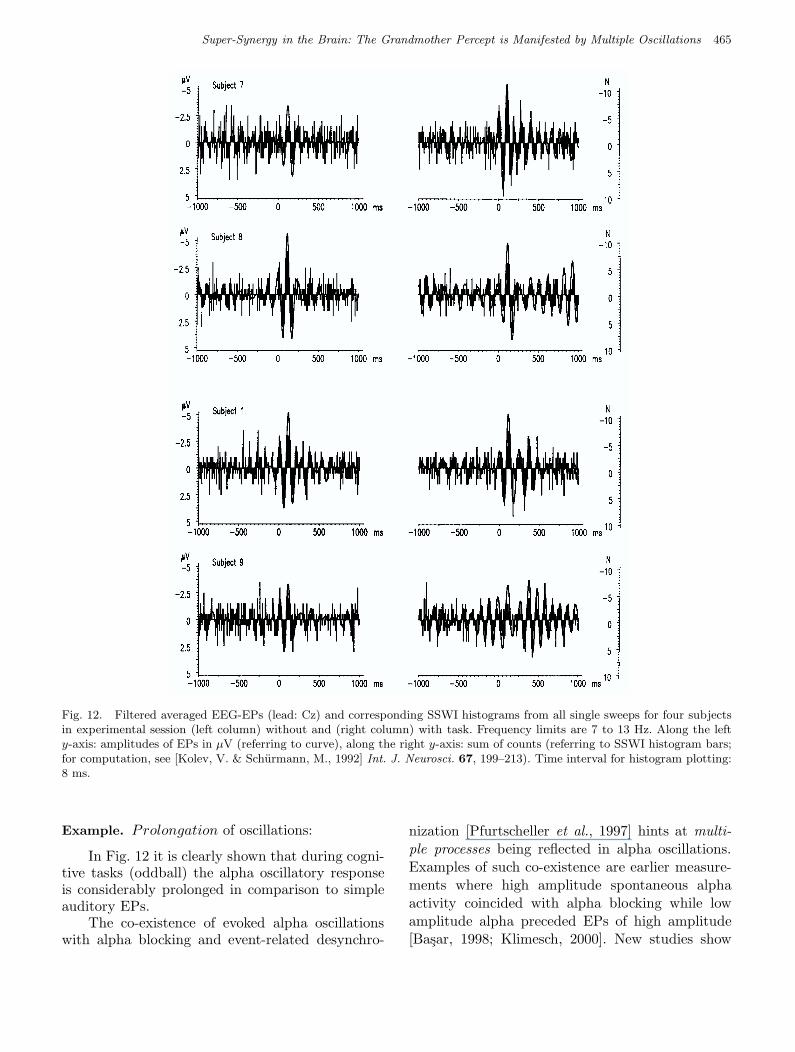
March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 465
Fig 12 Basar et al.
Fig. 12. Filtered averaged EEG-EPs (lead: Cz) and corresponding SSWI histograms from all single sweeps for four subjectsin experimental session (left column) without and (right column) with task. Frequency limits are 7 to 13 Hz. Along the lefty-axis: amplitudes of EPs in µV (referring to curve), along the right y-axis: sum of counts (referring to SSWI histogram bars;for computation, see [Kolev, V. & Schurmann, M., 1992] Int. J. Neurosci. 67, 199–213). Time interval for histogram plotting:8 ms.
Example. Prolongation of oscillations:
In Fig. 12 it is clearly shown that during cogni-tive tasks (oddball) the alpha oscillatory responseis considerably prolonged in comparison to simpleauditory EPs.
The co-existence of evoked alpha oscillationswith alpha blocking and event-related desynchro-
nization [Pfurtscheller et al., 1997] hints at multi-ple processes being reflected in alpha oscillations.Examples of such co-existence are earlier measure-ments where high amplitude spontaneous alphaactivity coincided with alpha blocking while lowamplitude alpha preceded EPs of high amplitude[Basar, 1998; Klimesch, 2000]. New studies show

March 5, 2004 9:23 00927
466 E. Basar et al.
a plausible superposition of several types of al-pha oscillations together in a schematical form.[Krause et al., 2001] also reported event-relatedsynchronizations and desynchronization together.For more complete descriptions of function-relatedalpha the reader is referred to [Basar et al., 1997];some examples follow in the next paragraph.
The experiments showed that damped alphaactivity is not present in all parts of the brainor elicited by all types of stimuli: Only by thecombination of EP frequency analysis, of adequatestimuli and of appropriate electrode positions suchactivities can be demonstrated. The results under-line the following properties of the neural tissuesunder study: In the 10 Hz frequency range (filterlimits: 8–14 Hz) we recorded large enhancements ofsingle visual EPs in the visual cortex (also reflectedin the amplitude frequency characteristics in theshape of a dominant 12 Hz peak). In the languageof systems theory significant (sharp) peaks in theamplitude characteristics of the transfer functioncharacterize resonant behavior of the studied sys-tem. One may also express this behavior as tuningof the “device”, or one might express the resonantfrequency channels as the “natural frequencies”of the system.
Similar to the gamma band the selectivelydistributed alpha system in the brain is interwovenwith multiple functions and control functions:
(a) The 10 Hz processes may facilitate, associationmechanisms in the brain: When a sensory orcognitive input elicits “10 Hz wave-trains” inseveral brain structures then it can be expectedthat this general activity can serve as a resonat-ing signal “par excellence” [Basar, 1980].
(b) Alpha activity controls EPs. Experiments ofseveral authors point out, that the ampli-tude, time course, and frequency responses inEPs strongly depend on the amplitude of theprestimulus alpha activity [Basar et al., 1997,1998].
Again, parallel observations at the cellular levelare noteworthy: Evoked oscillations in the 8–10 Hzfrequency range in visual cortex neurons uponvisual stimulation suggest a relation to scalp-recordable alpha responses [Silva et al., 1991; Dinseet al., 1997]. The sum of these observations permitsa tentative interpretation of alpha as a functionaland communicative signal with multiple functions.This interpretation of 10 Hz oscillations (at the cel-
lular level, or in populations) might be comparableto the putative universal role of gamma responsesin brain signaling.
2.4. Theta oscillations in
perception and cognition
Theta discharges are recorded from hippocampalneurons [Miller, 1991], neurons in n. accumbens,as well as cortical neurons. Hippocampal-cortical(frontal) networks operate in the theta frequencyrange. Theta cells in the hippocampus with multi-ple sensory behavioral correlates are described byBest and Ranck [1982]. Theta EEG response arerecorded in rats [Miller, 1991], cats, humans [Basar,1999].
In the following there are some examplesrelated to theta oscillations in perception andcognition:
(1) Experimental data suggests that event-relatedtheta oscillations are related to cognitive pro-cessing and cortico-hippocampal interaction[Miller, 1991; Klimesch et al., 1994; Basar,1999].
(2) Theta is the most stable component of thecat P300-like response [Basar, 1998, 1999;Sakowitz et al., 2000].
(3) Bimodal sensory stimulation induces large in-creases in frontal theta response thus demon-strating that complex events require frontaltheta processing [Basar, 1989, 1999; Sakowitzet al., 2000].
(4) Event-related theta oscillations are prolongedand/or have a second time window approx.300 ms after target stimuli in oddball exper-iments. Prolongation of theta is interpretedas being correlated with selective attention[Basar & Stampfer, 1985; Basar-Eroglu et al.,1992].
(5) Event-related theta oscillations are also ob-served after an inadequate stimulation whereasevent-related alpha oscillations are not ex-istent if the stimulation is an inadequateone. Accordingly, the associative characterfor event-related theta oscillations is morepronounced than for higher frequency event-related oscillations [Basar-Eroglu et al., 1992].
(6) “Orienting” — a coordinated response indicat-ing alertness, arousal or readiness to processinformation — is related to theta oscillationsand manifested in cat experiments during ex-
March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 467
0 250 5000 250 500
0 250 500 0 250 500 0 250 500
0 250 500
time (ms) time (ms)
time (ms) time (ms)time (ms)
time (ms)
F3 O1P3
5µV
F3F3F3
5µV
5µV
5µV
5µV
5µV
3.ATT.
VEP
Visual Stimulation
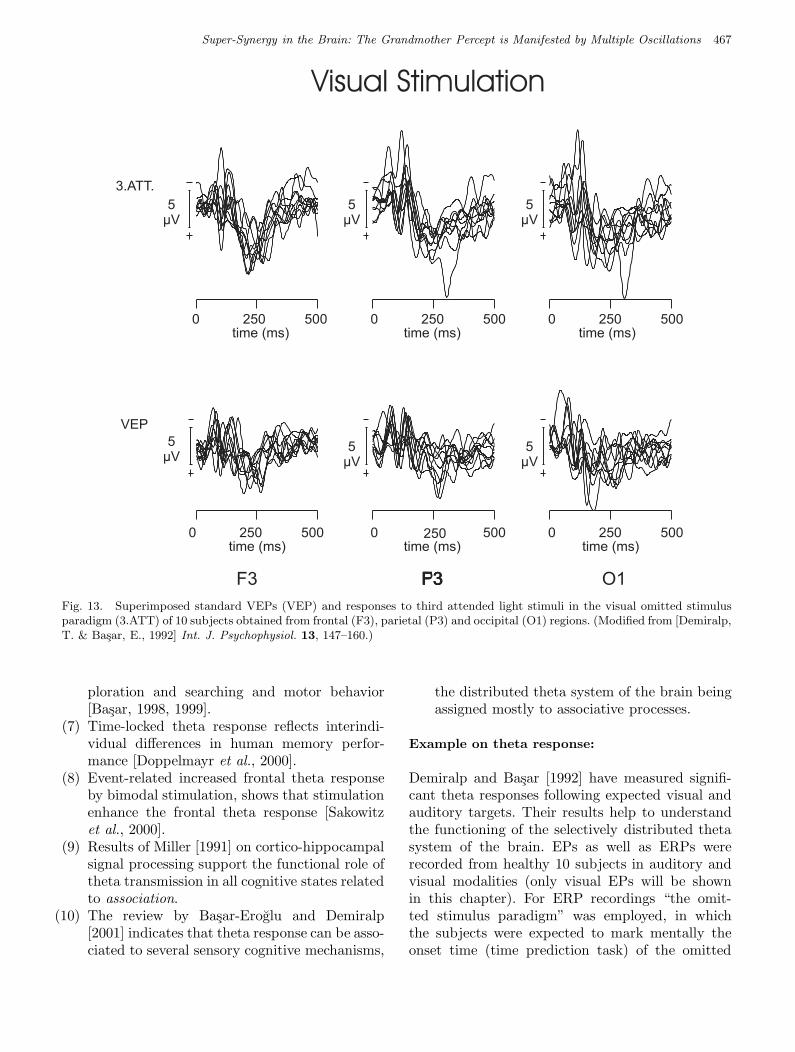
Fig 13 Basar et al.
Fig. 13. Superimposed standard VEPs (VEP) and responses to third attended light stimuli in the visual omitted stimulusparadigm (3.ATT) of 10 subjects obtained from frontal (F3), parietal (P3) and occipital (O1) regions. (Modified from [Demiralp,T. & Basar, E., 1992] Int. J. Psychophysiol. 13, 147–160.)
ploration and searching and motor behavior[Basar, 1998, 1999].
(7) Time-locked theta response reflects interindi-vidual differences in human memory perfor-mance [Doppelmayr et al., 2000].
(8) Event-related increased frontal theta responseby bimodal stimulation, shows that stimulationenhance the frontal theta response [Sakowitzet al., 2000].
(9) Results of Miller [1991] on cortico-hippocampalsignal processing support the functional role oftheta transmission in all cognitive states relatedto association.
(10) The review by Basar-Eroglu and Demiralp[2001] indicates that theta response can be asso-ciated to several sensory cognitive mechanisms,
the distributed theta system of the brain beingassigned mostly to associative processes.
Example on theta response:
Demiralp and Basar [1992] have measured signifi-cant theta responses following expected visual andauditory targets. Their results help to understandthe functioning of the selectively distributed thetasystem of the brain. EPs as well as ERPs wererecorded from healthy 10 subjects in auditory andvisual modalities (only visual EPs will be shownin this chapter). For ERP recordings “the omit-ted stimulus paradigm” was employed, in whichthe subjects were expected to mark mentally theonset time (time prediction task) of the omitted
March 5, 2004 9:23 00927
468 E. Basar et al.
Visual Stimulation Filter: 3 - 6 Hz
5 55
555
µV
µVµVµV
µVµV
µVµV
µV
µV
µV µV
2.5
2.52.52.5
2.52.5
3.ATT.
VEP
individual
individualexperiments
experiments
n = 10
n = 10
0 250 500 0 250 500
0 250 500 0 250 500
time (ms)
time (ms)
0 250 500
0 250 500
F3 P3 O1
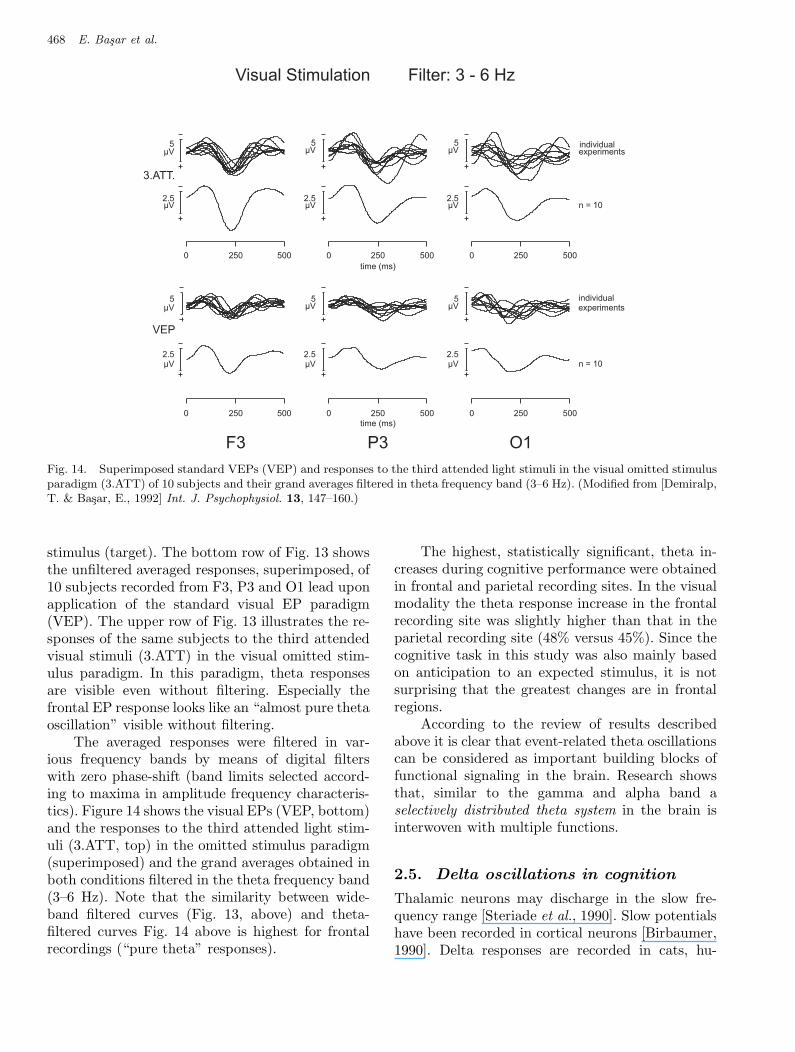
Fig 14 Basar et al.
Fig. 14. Superimposed standard VEPs (VEP) and responses to the third attended light stimuli in the visual omitted stimulusparadigm (3.ATT) of 10 subjects and their grand averages filtered in theta frequency band (3–6 Hz). (Modified from [Demiralp,T. & Basar, E., 1992] Int. J. Psychophysiol. 13, 147–160.)
stimulus (target). The bottom row of Fig. 13 showsthe unfiltered averaged responses, superimposed, of10 subjects recorded from F3, P3 and O1 lead uponapplication of the standard visual EP paradigm(VEP). The upper row of Fig. 13 illustrates the re-sponses of the same subjects to the third attendedvisual stimuli (3.ATT) in the visual omitted stim-ulus paradigm. In this paradigm, theta responsesare visible even without filtering. Especially thefrontal EP response looks like an “almost pure thetaoscillation” visible without filtering.
The averaged responses were filtered in var-ious frequency bands by means of digital filterswith zero phase-shift (band limits selected accord-ing to maxima in amplitude frequency characteris-tics). Figure 14 shows the visual EPs (VEP, bottom)and the responses to the third attended light stim-uli (3.ATT, top) in the omitted stimulus paradigm(superimposed) and the grand averages obtained inboth conditions filtered in the theta frequency band(3–6 Hz). Note that the similarity between wide-band filtered curves (Fig. 13, above) and theta-filtered curves Fig. 14 above is highest for frontalrecordings (“pure theta” responses).
The highest, statistically significant, theta in-creases during cognitive performance were obtainedin frontal and parietal recording sites. In the visualmodality the theta response increase in the frontalrecording site was slightly higher than that in theparietal recording site (48% versus 45%). Since thecognitive task in this study was also mainly basedon anticipation to an expected stimulus, it is notsurprising that the greatest changes are in frontalregions.
According to the review of results describedabove it is clear that event-related theta oscillationscan be considered as important building blocks offunctional signaling in the brain. Research showsthat, similar to the gamma and alpha band aselectively distributed theta system in the brain isinterwoven with multiple functions.
2.5. Delta oscillations in cognition
Thalamic neurons may discharge in the slow fre-quency range [Steriade et al., 1990]. Slow potentialshave been recorded in cortical neurons [Birbaumer,1990]. Delta responses are recorded in cats, hu-

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 469
Table 1. The medians of maximum amplitudes of delta, thetaand alpha frequency components of auditory evoked potentials(AEP), responses to third attended stimuli in the omitted stimu-lus paradigm (3.ATT) and responses to nonfrequent target tones inthe oddball paradigm (ODDBALL) obtained in frontal (F3), vertex(Cz), parietal (P3) and occipital (O1) recording sites. The percentchanges of amplitudes in (3.ATT) and (ODDBALL) conditions asthe percent of the standard AEP amplitudes are given in paren-theses. Statistically significant differences are marked with symbolsrepresenting the significance levels (∗p < 0.05; ∗∗p < 0.01). (Modi-fied from [Basar-Eroglu, C. et al., 1992] Int. J. Psychophysiol. 13,161–179.)
Theta (3–6 Hz)
Delta Window 1 Window 2(1–3 Hz) (0–250 ms) (250–300 ms)
AEP 1.8 4.7 2.0
F3 3.ATT 3.0 6.7 2.0(66%) (43%)∗∗ (0%)
P300 10.6 4.5 6.5(489%)∗∗ (−4%) (225%)∗∗
AEP 5.3 8.2 3.0
Cz 3.ATT 7.3 9.5 2.9(37%) (16%) (−3%)
P300 10.9 7.2 9.7(106%)∗∗ (−12%) (223%)∗∗
AEP 2.4 4.0 1.6
P3 3.ATT 3.2 4.4 1.8(33%) (10%) (13%)
P300 9.7 2.7 5.8(304%)∗∗ (−33%) (263%)∗∗
AEP 1.6 2.3 1.3
O1 3.ATT 1.9 2.3 1.5(19%) (0%) (15%)
P300 8.2 2.8 3.9(413%)∗∗ (22%) (200%)∗∗
mans [Basar & Stampfer, 1985; Schurmann et al.,1995]. Experimental data hint at functional corre-lates roughly similar to those mentioned for thetaoscillations, i.e. mainly in cognitive processing:
(1) The responses to visual oddball targets havetheir highest response amplitude in parietal lo-cations, whereas for auditory target stimuli thehighest delta response amplitudes are observedin central and frontal areas [Basar, 1998, 1999;Schurmann et al., 1995].
(2) Cognitive functions: The amplitude of the deltaresponse is considerably increased during odd-ball experiments. Accordingly, it was concludedthat the delta response is related to signal
detection and decision making [Basar-Erogluet al., 1992].
(3) In response to stimuli at the hearing thresholddelta oscillations are observed in human sub-jects, consistent with the hypothetical relationto signal detection and decision-making [Basar,1989, 1999].
(4) A waveform observed in response to deviantstimuli not attended by the subject, the mis-match negativity [Naatanen, 1992] is shaped bya delayed delta response superimposed witha significant theta response [Karakas et al.,2000a, 2000b].
(5) Phase-locked delta responses are probably themajor processing signals in the sleeping cat

March 5, 2004 9:23 00927
470 E. Basar et al.
Fig 15 Basar et al.
Fig 15 Basar et al.
Fig. 15. Time-domain and frequency-domain representations of grand averages of auditory evoked potentials, responses tothird attended tones in the omitted stimulus paradigm (3.ATT) and responses to nonfrequent target tones in the oddballparadigm (ODDBALL) obtained in frontal, vertex, parietal and occipital (F3, Cz, P3, O1) recording sites. (Modified from[Basar-Eroglu et al., 1992] Int. J. Psychophysiol. 13, 161–179.)
and human brain [Basar, 1980, 1999; Roschke,1995].
The topographic distribution of the results isagain consistent with a distributed response system.The delta response obtained during a typical P300experiment will be described in the following to givea good support about the cognitive nature of thedelta responses. Details are given in [Basar, 1998,1999]; some examples follow in the next paragraph.
Example on auditory P300:
The P300 human response to a special type of au-ditory stimuli shows that delta responses can beconsidered as “real brain responses” with precisefunctional correlates. This was demonstrated in astudy using an auditory oddball paradigm [Basar-Eroglu et al., 1992]. Standard auditory EPs (deltaresponse amplitude set to 100%) were comparedwith responses to oddball stimuli where the nor-malized delta amplitude was approximately 500%(see Fig. 15 and Table 1). This remarkable increaseis an example of a major change in the frequencycontents of an EP as mentioned in the beginning ofthis chapter. Taking into account the psychophysi-ological foundation of the P300 paradigm this hints
at cognitive processing as a functional correlate ofthe delta response. The same conclusion was drawnfrom a study employing a visual oddball paradigmwith standard versus target checkerboard stimuli[Schurmann et al., 1995].
3. Superposition Principle and
Superimposed Multiple
Oscillations in Theta and Delta
Frequency Windows in Cognitive
Processes
In previous sections we have chosen different fre-quency windows in order to describe functionalcorrelates of oscillations. We will emphasize now therole of multiple oscillations during cognitive pro-cesses. According to the principle of superposition,the existence of different peaks does not necessitatethe existence of different functional structures; sim-ilarly, disappearance of peaks does not necessarilyshow that the functional groups have ceased theiractivity [Basar & Ungan, 1973; Basar, 1980]. Theapplication of the superposition principle is shownin the example of the auditory evoked potential ofthe cat hippocampus of (Fig. 3).
Karakas et al. [2000b] investigated the contribu-tion of delta and theta responses to N200 and P300
March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 471
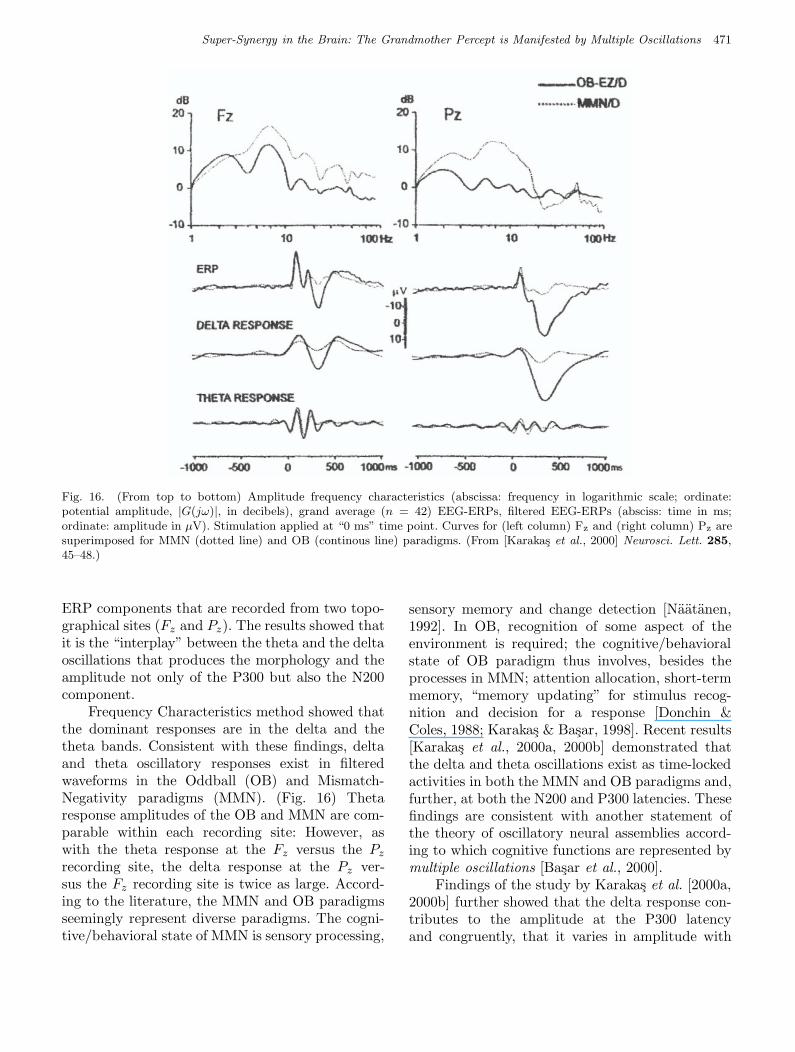
Fig 16 Basar et al.
Fig. 16. (From top to bottom) Amplitude frequency characteristics (abscissa: frequency in logarithmic scale; ordinate:potential amplitude, |G(jω)|, in decibels), grand average (n = 42) EEG-ERPs, filtered EEG-ERPs (absciss: time in ms;ordinate: amplitude in µV). Stimulation applied at “0 ms” time point. Curves for (left column) Fz and (right column) Pz aresuperimposed for MMN (dotted line) and OB (continous line) paradigms. (From [Karakas et al., 2000] Neurosci. Lett. 285,45–48.)
ERP components that are recorded from two topo-graphical sites (Fz and Pz). The results showed thatit is the “interplay” between the theta and the deltaoscillations that produces the morphology and theamplitude not only of the P300 but also the N200component.
Frequency Characteristics method showed thatthe dominant responses are in the delta and thetheta bands. Consistent with these findings, deltaand theta oscillatory responses exist in filteredwaveforms in the Oddball (OB) and Mismatch-Negativity paradigms (MMN). (Fig. 16) Thetaresponse amplitudes of the OB and MMN are com-parable within each recording site: However, aswith the theta response at the Fz versus the Pz
recording site, the delta response at the Pz ver-sus the Fz recording site is twice as large. Accord-ing to the literature, the MMN and OB paradigmsseemingly represent diverse paradigms. The cogni-tive/behavioral state of MMN is sensory processing,
sensory memory and change detection [Naatanen,1992]. In OB, recognition of some aspect of theenvironment is required; the cognitive/behavioralstate of OB paradigm thus involves, besides theprocesses in MMN; attention allocation, short-termmemory, “memory updating” for stimulus recog-nition and decision for a response [Donchin &Coles, 1988; Karakas & Basar, 1998]. Recent results[Karakas et al., 2000a, 2000b] demonstrated thatthe delta and theta oscillations exist as time-lockedactivities in both the MMN and OB paradigms and,further, at both the N200 and P300 latencies. Thesefindings are consistent with another statement ofthe theory of oscillatory neural assemblies accord-ing to which cognitive functions are represented bymultiple oscillations [Basar et al., 2000].
Findings of the study by Karakas et al. [2000a,2000b] further showed that the delta response con-tributes to the amplitude at the P300 latencyand congruently, that it varies in amplitude with
March 5, 2004 9:23 00927
472 E. Basar et al.
task-relevant responding that necessitates consciousstimulus evaluation and memory updating.
The work of groups with Klimesch, Gruzelierand Chen confirm and elegantly extend functionalrelevance of function related multiple oscillations[Gruzelier, 1996; Klimesch et al., 2000; Chen &Hermann, 2001].
4. Selectively Distributed and
Selectively Coherent Oscillatory
Networks
The description of integration needs morphologi-cal, functional interrelation in defined durations inthe time space; the degree of interactions betweentwo signals can be measured by coherence [vonStein & Sarnthein, 2000]. Coherence is a statisti-cal measure: the value of coherence depends on theamount of repeated correlations between events inthe frequency domain. The phase relationship be-tween the two signals is less relevant, however it
must be stable. Since the signal at each electrode inintracranial recordings mostly reflects the networkactivity under the electrode, coherence between twoelectrodes should measure interactions between twoneural populations. The statistical nature of coher-ence helps to unravel them from noise if they re-peat consistently [von Stein & Sarnthein, 2000]. Iftwo brain locations are coherent, one of the loca-tions drives the other or they reciprocally coop-erate. They can be also coherently activated by acommon driver [Bullock & McClune, 1989]. For in-terpretations of measurements with scalp electrodesthe view of Srinivasan et al. [1998] has to be takeninto consideration.
Findings of our research groups [Basar et al.,1979; Basar, 1980] demonstrated long distance co-herences in alpha, beta, theta and delta frequencyranges in structures such as sensory cortices, hip-pocampus, and brain stem in waking and freelymoving or sleeping cats. Their strength depends
Fig 17 Basar et al.
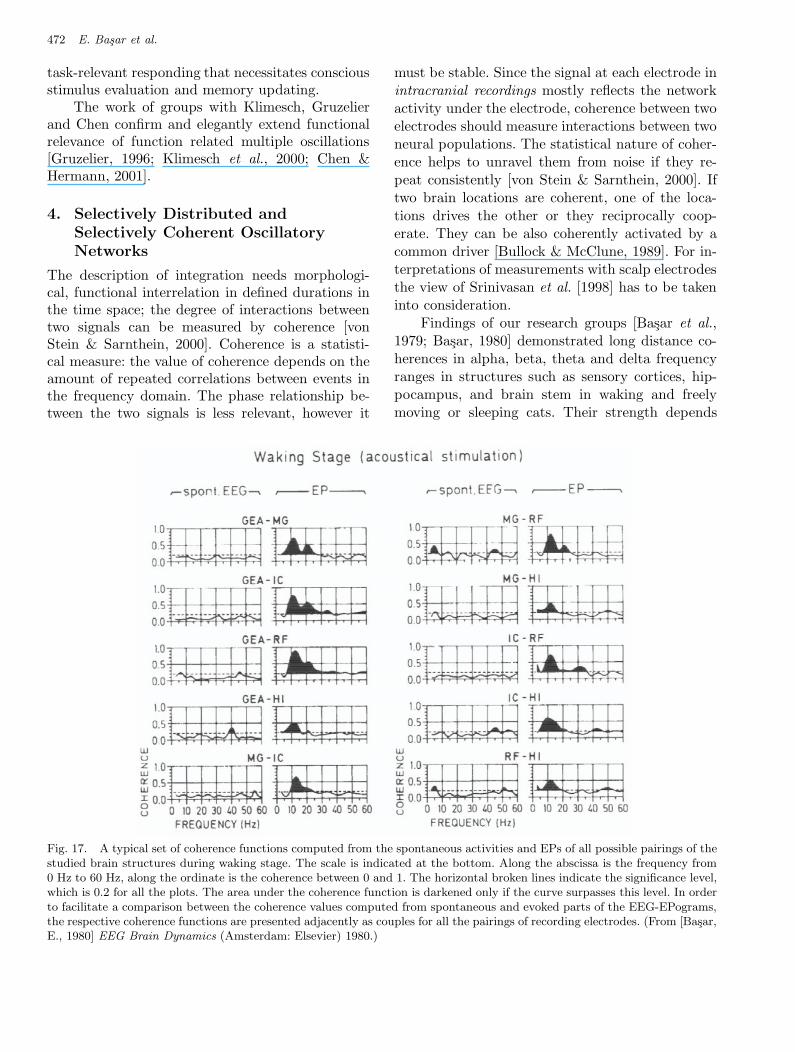
Fig. 17. A typical set of coherence functions computed from the spontaneous activities and EPs of all possible pairings of thestudied brain structures during waking stage. The scale is indicated at the bottom. Along the abscissa is the frequency from0 Hz to 60 Hz, along the ordinate is the coherence between 0 and 1. The horizontal broken lines indicate the significance level,which is 0.2 for all the plots. The area under the coherence function is darkened only if the curve surpasses this level. In orderto facilitate a comparison between the coherence values computed from spontaneous and evoked parts of the EEG-EPograms,the respective coherence functions are presented adjacently as couples for all the pairings of recording electrodes. (From [Basar,E., 1980] EEG Brain Dynamics (Amsterdam: Elsevier) 1980.)

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 473
on stimulation modality and recording sites. Duringthe waking stage the coupling (and/or synchro-nization) of resonant responses from various nu-clei in the alpha and beta frequency ranges canalso be demonstrated by using the coherence func-tions between all possible pairings of spontaneousand evoked activities in the structures as GEA,MG, IC, RF HI. (Fig. 17), which presents resultsof a typical experiment, the coherences in a fre-quency range of 3–60 Hz for spontaneous activi-ties and evoked potentials of all possible pairingsof the studied brain structures are illustrated. Dueto the reasonably large number of single sweepsincluded in the computation of averages and dueto the spectral window used, the auto and cross-spectral amplitudes have been adequately smoothedand a significance value of 0.2 has been attained forall the curves. Therefore, the area under the co-herence function is darkened only if the curve sur-passes this value in order to give emphasis to thoseparts of the curves above the significance level.One recognizes immediately, that in the alpha (8–14 Hz) and beta (14–25 Hz) frequency ranges thecoherency has usually high values between 0.5 and0.9 for evoked responses. However, the coherencebetween spontaneous activities of the same pair-ings of nuclei has definitely lower values. In the al-pha and beta frequency ranges, the coherence ofthe spontaneous activity barely reaches 0.3 in afew cases. In other words, there exists an impor-tant coherency increase upon stimulation. Duringthe slow sleep stage the evoked coherences be-tween all brain structures were shifted to slowerdelta frequency range [Basar et al., 1979; Basar,1980].
The term “Theory” may be used to signify anyhyphothesis whether confirmed or not, or may berestricted to hyphotheses that have been stronglyconfirmed as to become part of the accepted doc-trine of a particular science. In its best use, itsignifies a systematic account of some field ofstudy, derived from a set of general propositions.These propositions may be taken as postulates,as in pure mathematics, or they may be princi-ples more or less strongly confirmed by experiences,as in natural science [from Encyclopedia Britani-cae].
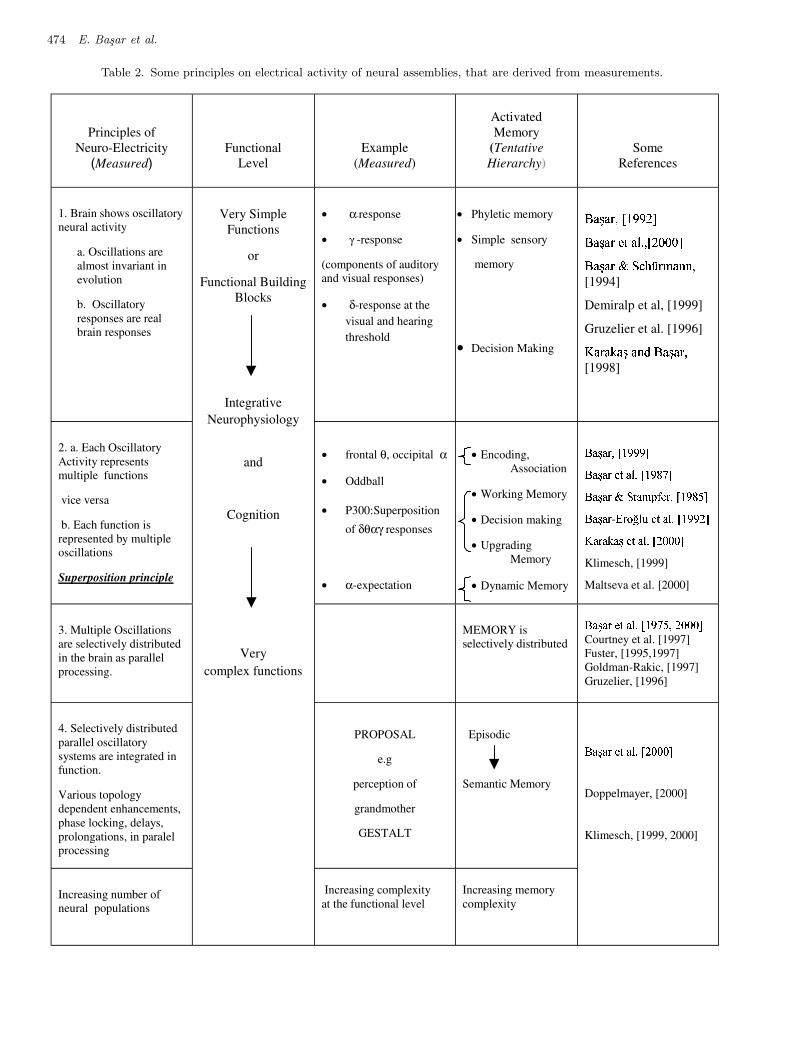
The neurons-brain theory decribes a systemat-ical account of measurements on electrical activityof neural assemblies, and incorporates also propos-als derived immediately from measurements. Ac-cordingly, in Table 2 we underline some principles
on electrical activity of neural assemblies, that arederived from measurements.
(1) The Brain has NATURAL FREQUENCIES oroscillations measurable at the cellular and pop-ulation levels.
(2) Brain Oscillatory Responses are real responsesrelated to function [Basar et al., 2001].
(3) Oscillations are selectively distributed and se-lectively coherent.
(4) Oscillations are related to multiple functionsand a given function is often manifested bymeans of multiple oscillations [Klimesch, 1999,2000]. Principle of superposition is confirmed byseveral publications [Karakas, 2000a, 2000b].
A natural consequence of the Neurons-BrainTheory (or Brain Assemblies Theory) is the super-synergy in oscillatory dynamics. The proposi-tion or a model for the electrical manifestation ofthe grandmother percept is based on evident facts:Research shows that the increasing complexity ofpercepts are accompanied with increased numberof multiple oscillations in parallel with increasednumber of activated neural populations. Althoughthe proposal related to the perception of the grand-mother is a fictive one, it is anchored on tenablearguments: May the grandmother percept involvemultiple and distributed oscillations instead of thefiring of a unique cell [Stryker, 1989]? A semi-empirical approach to the explanation of the grand-mother percept would be through the concept ofsuper-synergy.
According to results described in the previ-ous reviews selectively distributed and selectivelycoherent oscillatory networks in the delta, theta,alpha, beta and gamma bands play a major rolein brain functioning. Sensory and cognitive eventsevoke superimposed multiple oscillations that aretransferred to spatially distributed tissues almost inparallel with various degrees of amplitude, latency,duration, synchronization and coherence. Sakowitzet al. [2001] reported significant gamma responseamplitude increases in distributed areas of thebrain and a 100% frontal theta enhancement bybimodal stimulation compared to unimodal one.[Basar et al., 1993] reported that retrieval of visualexperience from short-term memory is associatedwith 40 Hz activity. The coherences between dif-ferent spatial locations of the brain vary as theseareas are activated with different classes of stimuli(haptic and visual) in an associative learning task
March 5, 2004 9:23 00927
474 E. Basar et al.
Table 2. Some principles on electrical activity of neural assemblies, that are derived from measurements.
Principles of
Neuro-Electricity (Measured)
Functional
Level
Example
(Measured)
Activated
Memory
(Tentative
Hierarchy)
Some
References
1. Brain shows oscillatory
neural activity
a. Oscillations are
almost invariant in
evolution
b. Oscillatory
responses are real
brain responses
• α-response
• γ -response
(components of auditory
and visual responses)
• δ-response at the
visual and hearing
threshold
• Phyletic memory
• Simple sensory
memory
• Decision Making
��������������� �����
������������������� ������������
���������! #"�$�%�&��(')��*�*��[1994]
Demiralp et al, [1999]
Gruzelier et al. [1996] +,���-��./���0�1*/2,���3�����4�[1998]
2. a. Each Oscillatory
Activity represents
multiple functions
vice versa
b. Each function is
represented by multiple
oscillations
Superposition principle
• frontal θ, occipital α
• Oddball
• P300:Superposition
of δθαγ responses
• α-expectation
• Encoding,
Association
• Working Memory
• Decision making
• Upgrading
Memory
• Dynamic Memory
56�7(6�8�9�:<;�=3=�=1>
56�7(6�8@?-A�6CB�D�:(;�=3E�F1>
56�7(6�8GIH�A�64JLKNMO?�8�9�:<;�=NE�P�>
56�7(6�8
- Q 8�R�S3B TU?�A�6VB�D :<;4=3=�W3> X 648�6�Y�6C7Z?-A�6CB�D�:[WN\�\�\�>
Klimesch, [1999]
Maltseva et al. [2000]
3. Multiple Oscillations
are selectively distributed
in the brain as parallel
processing.
MEMORY is
selectively distributed
56�7(6�8@?-A�6CB�D�:(;�=�F3P�9�W3\3\�\�>
Courtney et al. [1997]
Fuster, [1995,1997]
Goldman-Rakic, [1997]
Gruzelier, [1996]
4. Selectively distributed
parallel oscillatory
systems are integrated in
function.
Various topology
dependent enhancements,
phase locking, delays,
prolongations, in paralel
processing
PROPOSAL
e.g
perception of
grandmother
GESTALT
Episodic
Semantic Memory
Increasing number of
neural populations
Very Simple
Functions
or
Functional Building
Blocks
Integrative
Neurophysiology
and
Cognition
Very
complex functions
Increasing complexity
at the functional level
Increasing memory
complexity
56�7(6�8@?-A�6CB�D�:]W�\�\�\1>
Doppelmayer, [2000]
Klimesch, [1999, 2000]

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 475
[Basar, 1988]. Such findings experimentally sub-stantiate the submechanisms that have been out-lined above. Accordingly, we suggest that com-plex percepts (such as the visual image of one’s“grandmother”) are formed and/or manifested bymeans of the ensemble of oscillatory superbindingdynamics.
In the adjacent column to the proposal toGrandmother or to Perception of Gestalt, the com-plex memory function (episodic and semantic mem-ory) is placed. This arrangement of the table is inaccordance with the references indicating the in-creasing number of involved populations and fre-quency windows with the increased complexity atthe functional level.
In search of experimental support for thehypothesized distributed alpha response system[Basar, 1999] we measured EEG responses to vi-sual stimuli in the cat brain (with intracranialelectrodes in cortical, thalamic and hippocampalsites). Alpha responses (10 Hz oscillations of 200–300 ms duration) were observed not only in occipi-tal cortex and thalamus, but also in hippocampus.Remarkably, hippocampal alpha amplitudes werehigher in amplitude than thalamic ones, and thecoherence of hippocampus-cortex was higher than
the evoked coherence between thalamus and cortex[Schurmann et al., 2000].
Coherences increased significantly upon visualstimulation in delta, alpha, beta and gamma fre-quency ranges; (2) hippocampo-cortical coherenceswere significantly higher than thalamo-cortical onesfor the alpha (HI-OC: 0.28 versus LG-OC: 0.15),beta and gamma frequency ranges; (3) the effect ofthe interaction of channel and experiment fac-tors, which refers to the differences of coherencechanges upon stimulation between the two elec-trode pairs, was only significant in alpha and betafrequency ranges. Upon visual stimulation, the al-pha coherence HI-OC increased to 0.41, whereasthe coherence LG-OC increased only to 0.18 (seeFig. 18).
These findings demonstrated also that duringsensory stimulation the recordings between variousstructures showed varied degrees of coherence, thusindicating the interaction between two structures[Schurmann et al., 2000; Kocsis et al., 2001]. “Var-ied degrees of coherences” lead to the concept ofselectively coherent oscillatory networks. The im-portant role of functional integration and frequencyresponse of long-range interactions in the alpha andtheta ranges in processing of the mental context is
Fig 18 Basar et al.
Fig. 18. Coherence values (dimensionless) for the electrode pairs thalamus-visual cortex (TH–OC, top) and hippocampus-visual cortex (HI–OC, bottom). Each pair of dots represents coherence for corresponding (left) EEG and (right) EP segmentsin an individual cat. Note: The marked stimulus-induced increase in coherence for HI–OC. (From [Schurmann et al., 2000]Neurosci. Lett. 292, 175–178.)

March 5, 2004 9:23 00927
476 E. Basar et al.
also emphasized by confirming our long-standingview [Basar, 1980, 1999; von Stein & Sarnthein,2000]. In the gamma frequency range coherencesbetween different spatial locations of the brain varyas these areas are activated with different classes ofstimuli (haptic and visual) in an associative learningtask [Miltner, 1999].
5. Distributed Oscillatory Systems
in the Brain and Distributed
Memory
5.1. Event-processing in distributed
systems
The synchrony of selectivities described earlier byour group could have a conceptual parallel in“selectively distributed processing” in neurocogni-tive networks [Mesulam, 1990, 1994]. In Mesulam’sneurological model of cognition, the unimodal areasof cortex provide the most veridical building blocksof experience. Transmodal nodes bind informationin a way that introduces temporal and contex-tual coherence. The formation of specific templatesbelonging to objects and memories occurs in dis-tributed form but with considerable specialization.This arrangement leads to a highly flexible andpowerful computational system, which underliesthe selectively distributed processing. In our ear-lier work, we often used the expression or conceptof distributed oscillatory systems and their reso-nance as selective activities. According to Mesulam,functional selectivities exist in distributed functionsthat are based on the anatomy. The electrophysi-ological activity of selectively distributed systemsmust be also of selective behavior. Accordingly, os-cillatory response susceptibility of the sensory cor-tices, of the hippocampus, thalamus or cerebellumshould also be differentiated, depicting selective be-havior to stimulation from the milieu interieur orexterieur.
5.2. Memory and information:
Working memory systems and
long term memory system
In this report it is impossible to describe all currenttheories related to memory functions. Therefore,we issue only short statements by describing someof the current developments only pertinent tobrain dynamics electrophysiology and fMRI studiesundertaken; in order to explain the relation of selec-
tively distributed oscillatory systems with memoryprocessing.
Two basic aspects of memory processes will bedistinguished [Klimesch et al., 1994]. The first refersto processes of the working memory system (WMS),the second to that of the long-term memory system(LTMS). Probably any cognitive process dependson the resources of both systems. As an example,let us consider an every day cognitive process suchas recognizing a familiar object. The basic idea hereis that after a sensory code is established, semanticinformation in long-term memory (LTM) is accessedwhich is used to identify the perceived object. If thematching process yields a positive result, the objectis recognized which in turn leads to the creation ofa short-term memory (STM) code. It is also im-portant to mention Baddeley’s concept of workingmemory, which comprises an attentional controller,the central executive and subsidiary slave systems[Baddeley, 1986, 1992].
5.2.1. fMRI studies related to memory
function
For fMRI investigations in humans, Courtney et al.[1997] presented subjects with pictures of humanfaces, and asked them to recall whether the pic-ture being shown was the same, or different, fromone that had been presented eight seconds ear-lier. The authors found that activations in theprefrontal areas correlated most strongly with de-lay periods, compared with activations in the visualareas, which were more strongly correlated withsensory stimulation.
Cohen et al. [1997] presented subjects withwritten consonants, one at a time every ten seconds,and asked them to judge whether each consonantwas the same as a letter presented one, two or threetrials back in the sequence. This task requires thatsubjects remember the order of consonants, as wellas their identity. The farther back in the sequencethe consonant to be recalled occurs, the greaterthe “load” on working memory. These authorsshowed that activations in the prefrontal cortex aremaintained throughout the ten-second interstimu-lus interval and, importantly, that the degree of pre-frontal activation is higher for the conditions withthe greatest memory load. By contrast activationsby the primary visual, somatosensory and motorcortices, as well as in several secondary regions, arenot sustained across the ten-second-interval, andthey are not related to memory demand. They are

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 477
probably responsive to the sensory or perceptual,but not memory-aiding, aspects of working mem-ory tasks. Further, according to the fMRI results in[Courtney et al., 1997] early extrastriate visual ar-eas demonstrate transient, relatively non-selectiveresponses to complex visual stimuli and later extras-triate visual areas demonstrate transient, selectiveresponses to faces, indicating a more specializedrole in the processing of meaningful images, andboth extrastriate visual and prefrontal cortical ar-eas demonstrate sustained activity during memorydelays, indicating a role in maintaining an activerepresentation of the face in working memory.
According to Fuster’s view stating that memoryreflects a distributed property of cortical systemsand to our view outlined in the present paper itcan be hypothesized that the selectively distributedoscillatory systems (or networks) may provide ageneral communication framework that can be auseful concept for functional mapping of the brain[Mesulam, 1990, 1994].
Communication in these networks might con-tribute to the formation of specific templatesbelonging to objects and memories. According toa model of cognition, this formation occurs asselectively distributed processing with considerablespecialization and in anatomically differentiated lo-calizations [Mesulam, 1990, 1994] (for details aboutmemory as a distributed property of cortical sys-tems see also Fuster [1997]). In particular, analysisof the hypothetical distributed oscillatory systemsmay lead to fundamental functional mapping of thebrain, complementary to morphological studies.
5.3. EEG-oscillations and memory
As one can extract from Mesulam’s and Fuster’swork, there must be common codes for perpetualsignal transfer between neural networks for paralleland serial processing and also for possible reverber-ation circuits and loops between neural networks.Oscillations might serve as adequate codes for thisgeneral communication by putting the networks toresonate.
A more general view is that functional or os-cillatory network modules are distributed not onlyin the cortex but also in the whole brain [Basar,1998]. In the following, we will try to discuss an elec-trophysiological parallel between Fuster’s “memorynetwork” and the distributed oscillatory systemsmentioned earlier.
When analyzing the field potentials it is diffi-cult to define boundaries of brain nuclei and their
electrical activity. Nevertheless, this approach isuseful, since a great amount of data can be collectedand interpreted from several electrodes distributedin the brain. Furthermore, it is possible to performmeasurements during continuously changing cogni-tive states. This way, EPs or EEG segments arerecorded in the cortex, limbic system, and thalamus,and cerebellum. They can be compared in wakingand freely behaving animals.
This type of recording during behavioral statescannot possibly be managed with recordings of sin-gle cell electrodes. Studies on functional correlatesof structures like sensory cortices, hippocampus,and thalamic relay nuclei are mostly based on ex-periments using unit recordings. A major difficultyconcerning the interpretation of experiments withsingle unit recordings (for example, experiments oncortico-thalamic information transfer) is that theresults are limited to a few neurons. Accordingly,we assume that every hypothesis on the localiza-tion of “thalamo-cortical circuit as 10 Hz generator”is restricted and not acceptable with regard to theresults of experiments described in this article: Thealpha, theta and gamma generators are selectivelydistributed in the brain.
Basar [1999] reviewed several classifications ofmemories given by distinguished memory inves-tigators in order to find a strategy to explorethe electrophysiology of distributed memories. Heconcluded that the brain oscillations in EEG andERPs could provide a good foundation to attackthe problem both at the neurophysiological andpsychological levels since brain oscillations havesimilar frequency codes in the whole brain. Re-membering and memory are manifestations of var-ious and multiple functional processes dependingon the complexity of the input to the CNS. Al-ready the brain’s electrical response elicited bymeans of a simple light stimulation is related tomemory processes at the lowest hierarchical order.When we talk about a memory process — eithera short- or long-term one — then we have in ourmind the perception of a sensory input, which ismatched with information already stored in the neu-ral tissue. If a simple light evokes alpha and gammaresponses then it is almost obligatory to assumethat elementary oscillatory responses are also man-ifestations of several memory processes at differenthierarchical levels. The topology of the memoriesdepending on the modality of the input must bedifferent (see examples given above: cross modality

March 5, 2004 9:23 00927
478 E. Basar et al.
experiments; measurements in cortical and subcor-tical structures).
So far, such studies have rarely been per-formed, the multiple distributed memories cannotbe treated in detail and a classification on all lev-els of distributed memories cannot be providedyet.
In performing many complex tasks, it is nec-essary to hold information in temporary storageto complete the task. The system used for thisis referred to as working memory [Baddeley,1996]. Working memory is the temporary, ad hoc,activation of an extensive network of short- or long-term perceptual component of that network as anyother perceptual memory, would be, retrievable andexpandable by a new stimulus or experience. Fusterstates that working memory has the same corticalsubstrate as the kind of short-term memory tra-ditionally considered as the gateway to long-termmemory.
According to the functional descriptions above,a simple light stimulation, or a complex light stimu-lation or a light stimulation with some tasks (event)should evoke oscillatory responses with differenttime hierarchies. Our view is that functional or os-cillatory networks modules are distributed not onlyin the cortex but also in the whole brain [Basar,1999].
The detection of a target signal during a P300type of experiment requires also a type of workingmemory. The subject has to sustain knowledge re-lated to the target (nature of the target, frequency,shape, color, etc, depending on the type of the ex-periment) during the experiment. The linkage be-tween P300 amplitude and latency measures andworking memory processes has been already estab-lished [Sanquist et al., 1980; Howard & Polich, 1985;Pratt et al., 1989; Fabiani et al., 1990]. The match-ing process after the detection of the target shouldrely in this case on working memory.
5.4. Multiple functions of
event-related oscillations and
multiple functions of memory:
Convergence of concepts
To see something, even the simplest light signal, isalready a memory process related to a fundamen-tal in-born retrieval process: a baby perceives thelight and show reflex responses to light before goingthrough learning processes. This is probably a basicdecoding process.
In Fig. 19 responses to target and non-targetstimulation (upon checkerboard stimulation) in al-pha and delta frequencies are shown. The occipital10 Hz response is large in posterior areas (related tovision). The delta response however is distributed,being most marked in posterior areas upon targetstimuli. As explained above, the target signal alsorequires working memory processes. So the fact thatdelta responses are most marked in posterior ar-eas hints at selective distribution of “memory oscil-lations”. Ample occipital 10 Hz responses are notrecorded in frontal locations indicating that thefrontal lobes are not involved in primary visualprocessing. This response is a sign of perceptualmemory: When no visual perception occurs, thenthere are no 10 Hz responses.
As to the delta response in the auditory P300paradigm, a distributed highly enhanced responsein the whole cortex is observed [Basar-Eroglu et al.,1992], the maxima being in frontal and parietal ar-eas. The auditory 10 Hz signal to auditory signalis missing. This findings again indicate a functionalselective distribution 10 Hz responses are recordedin primary sensory areas, frontal and parietal deltaareas.
As to the theta response the findings are yetmore complicated to interpret: In the auditoryP300 paradigm only target signals have a prolongedtheta oscillation (second window) marked in pari-etal and frontal recording indicating a correlationto working memory. In experiments where subjectspay attention to the third applied signal in anevoked potential experiment, the third attended sig-nal show ample theta increases again especially infrontal location. In this report it is not our aim todifferentiate the functional roles of theta and deltaresponses in working memory processes; we solelyindicate that this oscillatory responses are selec-tively distributed depending on the memory loadrequired during the experiments.
A P300-40 Hz response is visible in Fig. 7.This figure illustrates the superposition of theta andgamma response following omitted stimuli as tar-gets. The combination of gamma and delta responseoscillations are here again involved with workingmemory.
These findings have remarkable parallelitiesto fMRI experiments showing topographical func-tional selectivity [Goldman-Rakic, 1997]. Moreover,it is possible to define the working and percep-tual memory components in a time window already

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 479
Fig 19 Basar et al.
(a)
Fig 19 Basar et al.
(b)
Fig. 19. Grand average ERPs (50′ checkerboards; N = 9 subjects), filtered (a) 1–5 Hz and (b) 8–15 Hz, respectively.(Left) Visual evoked potentials (VEP), (middle) responses to non-target stimuli; (right) responses to target stimuli.
100 ms upon a memory load. At least two typesof memory responses with a time hierarchy is ob-served (1) sensory (perceptual) memory oscillatoryresponse and (2) late window responses related toworking memory. This time hierarchy occurring inthe 1 sec period following stimuli, are not reflectedin fMRI studies, and not found in conventionalERPs that, in the best case, allow analysis of tworelevant peaks.
6. The “Super-Synergy” is a
Spatiotemporal and Functional
Organization of Multiple and
Distributed Oscillations in the
Brain
In current neuroscience literature an importantquestion is how the activities within and be-
tween those separate processing areas are unifiedin momentary association to form a coherent repre-sentational state — the neural basis of a percep-tion. “Binding” is assumed to get delineated byrecombination of a neural image of the stimulus as awhole. Synchronized activity is linked to the stim-uli that evoke it; only those activities evoked bystimuli with common stimulus features are boundtogether in a dynamically formed ensemble. Thecurrent literature in favor of the so-called “bind-ing theory” considers the binding as the uniquefunctionale correlate of only gamma activity and isbased on experiments with multiple microelectrodesin small portions of cortical areas [Eckhorn, 1988;Singer, 1995]. Although this view is interesting, theresults of many laboratories presented in the presentreview clearly show that even the most simple per-cept is manifested with multiple oscillations in a

March 5, 2004 9:23 00927
480 E. Basar et al.
large amount of selectively distributed oscillatorynetworks and long distance coherences [Basar et al.,1979; Basar, 1980; Schurmann et al., 2000; Koc-sis et al., 2001]. The binding of percepts cannotbe solely explained with the binding of 40 Hz os-cillations evoked by the complex features of per-ceptual targets processed in separate neocorticalareas. The neurons-brain theory is a consequenceof measurements related to the oscillatory activ-ity of neural populations, in both human and ani-mal experiments (see the companion report [Basar,2003]). Our research group introduced already in1970s the notion of distributed and parallel process-ing and multiple coherent oscillations [Basar, 1980,1983, 1992; Basar et al., 1999, 2000, 2001a–2001d].The consequence is that extremely large numberof combinations of oscillatory parameters is man-ifested by sensory-cognitive information processing.The crucial implication of all referred results inthis report is that the explanation of complex per-cepts with 40 Hz binding leads to a reductionism.Because long distance event-related coherence in-creases in alpha, beta, theta, delta and gamma indistributed networks are also selective [Basar et al.,1979; Basar, 1980; Schurmann et al., 2000; Kocsiset al., 2001]. Accordingly the consideration of nu-merous selectively distributed oscillatory networkswith varied degrees of event-related enhancementsand varied degree of coherency as manifestation ofcomplex percepts is a much more tenable proposi-tion. At every cognitive and sensory event not onlyprimary sensory areas but also at least frontal areasare also activated as fMRI and electrophysiologicaldata clearly demonstrated [Courtney et al., 1997;Basar et al., 2000].
It was shown that complex signals and asso-ciations (also remembering) are accompanied withlarge increase of frontal theta and delta, not onlyin occipital theta or gamma. Instead, the huge al-pha increase in occipital regions is not registeredin frontal alpha. In animals, complex events are ac-companied with increase in frontal and hippocam-pal theta oscillations additional to gamma andalpha.
In the analysis of distributed and parallel pro-cessing it was assumed that EEG goes from a disor-dered state (chaotic state with large dimensions) toan ordered state upon application of sensory cogni-tive signals [Basar, 1980]. For short EEG segments,calculation of wavelet entropy — a new emergingmethod — demonstrates that, upon application ofsensory cognitive signals, the EEG goes from a dis-
ordered state to an ordered state. The distributionof entropy changes are also a part of differenti-ated parallel processing. However, preliminary re-sults show that the entropy distribution does notnecessarily coincide with the distribution of brainoscillations [Quiroga et al., 1999, 2001; Rosso et al.,2001]. The results show that the distribution oftheta entropy is also selective upon complex stimuli,the frontal areas showing the largest entropy de-crease following complex stimuli ([Yordanova et al.,2002] also see [beim Graben, 2000]).
Taking together all these results on paral-lel processing we tentatively hypothesize that thecomplex percepts are manifested with a super-synergy consisting of an ensemble of (at least)six submechanisms acting parallel or in synchro-nization upon sensory-cognitive input. It is pro-posed that the coexistence and cooperative actionof these interwoven and interacting submechanismspartly explain the mechanisms of integrative brainfunctions:
A natural consequence of the stated rules ofthe neurons-brain theory is explained in the for-mer sections as “super-synergy”. The long distanceevent-related coherence increases in alpha, beta,theta, delta and gamma in distributed networks arealso selective. Thus, selective distribution appliesnot only to the amplitudes of oscillations or theirfrequency components but also to their coherences[Basar, 1980, 1983, 1988, 1998; Basar et al., 1975,1979; Schurmann et al., 2000; Kocsis et al., 2001].Consequently, sensory and/or cognitive informationprocessing is manifested in a complex signal that isformed from the combination of an extremely largenumber of oscillatory parameters; thus the concept,“oscillatory superbinding dynamics”.
The super-synergy comprehends an ensemble ofat least five mechanisms that act in synergy uponsensory-cognitive input. It is proposed that the co-existence and cooperative action of these interwovenand interacting submechanisms is a manifestationof integrative brain functions. The submechanismsand/or related processes are as follows:
(1) The superposition principle that includes thealpha, beta, gamma, theta and delta bands[Basar et al., 1999; Karakas et al., 2000a, 2000b;Klimesch et al., 2000; Chen & Hermann 2001].
(2) Activation of more than one selectivelydistributed oscillations in gamma, alpha,theta and delta bands upon exogenous orendogenous input. These activities are mani-

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 481
fested with a multifold occurrence of param-eters such as enhancement, delay, blocking(desynchronization), and prolongation [Basar,1980, 1999; Basar et al., 1999, 2000, 2001a–2001d]. The ensemble of oscillations andamplitude of oscillations usually increase asthe complexity of the stimulation increases orthe recognition of the stimulation becomes moredifficult.
(3) Temporal and spatial changes of entropy in thebrain [Quiroga et al., 1999; Yordanova et al.,2002].
(4) Temporal coherence between cells in corticalcolumns for simple binding [Eckhorn et al.,1988; Gray & Singer, 1989].
(5) Varying degrees of spatial coherence that oc-cur over long-distances as parallel processing[Basar, 1980, 1983; Basar et al., 1999; Miltner,1999; Schurmann et al., 2000; Kocsis et al.,2001].
(6) Inverse relation between EEG and Event Re-lated Potentials, EEG being a control parame-ter for responsiveness of the brain.
Thus, integrative brain function occurs overa spatiotemporal organization. The principle ofsuperposition describes integration over the tem-poral axis, consisting of temporal relationship be-tween the amplitude and phase of oscillations invarious frequency bands. Furthermore, selectivelydistributed and selectively coherent oscillatory neu-ral populations describe integration over the spatialaxis. Consequently, integrative activity is a functionof the coherences between spatial locations of thebrain; these coherences vary according to the typeof sensory and or cognitive event and possibly theconsciousness state of the species.
The empirically founded submechanisms areaccompanied by the formation of a percept orin general of all cognitive events. This is real-ized through combinations of the response pa-rameters among which are the varying strengthsin the associative links between brain structures.The number of combination that these variablesform leads to an enormous amount of neuroelectricconfigurations. Spatiotemporal and functional or-ganization of multiple and distributed oscillationsin the brain inevitably leads to superbinding as away to explain the integrative activity of the brainduring the formation of percepts and other cogni-tive events.
We denote tentatively “superbinding”, the com-bination of mechanisms of (1) superposition, (2) ac-
tivation of selectively distributed oscillatory systemsand (3) the selectively distributed long distance co-herence. All these processes are interwoven alongthe time axis as large numbers of data demonstrated.
Consequently, sensory and/or cognitive infor-mation processing is manifested in a complex signalthat is formed from the combination of an extremelylarge number of oscillatory parameters; thus theconcept, “oscillatory superbinding dynamics”.
6.1. Preliminary hypothesis for
“Grandmother cell or
grandmother cell assemblies”
The grandmother neuron is a neuron that respondsto nothing else but the face of one’s grandmother.Barlow [1972], stated that the sensory system isorganized to achieve as completely as possible arepresentation of the sensory stimulus with a min-imum of active neurons. According to his view,perception consists of achieving a selection from thevery numerous high-level neurons corresponding toa pattern of external events of the order of com-plexity of the event that is symbolized by the word“grandmother” [Lettvin et al., 1959].
According to what Mountcastle [1992] called a“paradigm change in neuroscience”, the role of neu-ral assemblies gains an important rank in the func-tional studies. Damasio and Damasio [1994] statethat our brains use dynamic records, rather thanstatic, immutable memory traces. For example therecord of the face of a person is a set of neuroncircuit changes which can be reactivated, ratherthan the “picture” that is stored somewhere in thebrain.
The theory on the existence of the selectivelydistributed alpha and theta and 40 Hz systemsof the brain has been formulated on the basisof progressive developing results and publications(references). Further, the existence of EEG-modules(active in parallel and selectively — or diffusely— distributed in the brain) was proposed. EEGoscillation-modules, in this context, are neural pop-ulations giving rise to coherent EEG activities indifferent frequency channels [Basar, 1999].
We assume that the EEG oscillations which canattain coherent states belong to functional reper-toires of the higher brain function. The superpo-sition of several of these signals makes it possibleto connect behavioral events with an ensemble ofevent-related oscillations. Since the ERPs are nowvery popular for scientists of psychophysiology who

March 5, 2004 9:23 00927
482 E. Basar et al.
use a diversity of psychological paradigms, the con-sideration of EEG dynamics can bring a deeperunderstanding to the correlation between cognitiveperformance and physiological states.
According to Szentagothai, connections areestablished between vertical columns and mod-ules. These connections can also link frontal ar-eas to occipital ones. Now the question is whetherthe hypothesized EEG generating modules in thefrontal and occipital areas are also connected. Ifevent-related 10 Hz oscillations are measured inoccipital and frontal areas, then it is certainlylegitimate to postulate such modules and neuralconnections between these modules. The same pos-sibility also exists in theta, delta, and gammafrequency bands. With the knowledge presentedin Basar [1998] it is not possible to clearly in-dicate that the modules described by Szentago-tai also serve for processing information in theEEG frequency channels. Volume conduction be-tween modules as distant as occipital and frontalcortices is unlikely. Accordingly, the high ampli-tude delta responses following target signals in P300experiments must have different local generators.Furthermore, it is probable that neural impulsescan reach several parts of the cortex following anevent and generate in cortical areas oscillatory re-sponses in various frequency channels depending onthe sensory information or task.
According to experiments described in Basar[1980] gamma activity is not recorded only in asmall given brain population but in widely dis-tributed neural populations. The 10 Hz resonancesare also widely distributed and depend on functionsprocessed in the brain. Depending on “regimes”or “states” of the brain, the limbic system, brain-stem, thalamus, and cortex are all involved with2 Hz, 4 Hz, 10 Hz, and 40 Hz firing or with all ofthem. The distribution of coherently working EEGmodules does not take place only in the cortex; weassume that they are selectively distributed in theentire system.
One of the examples is the frontal-hippocampal-parietal-system, which gains a highlyorganized theta state during attentive processes. Bycombining examples and concepts arising from theanalysis of EEG and evoked potential responses, weaim to extend from modular concepts to modularfrequency systems operating in the entire brain.This is a new approach based on measurement offield potentials that reflect the activity of large neu-
ral populations [Basar, 1998, 1999]. These neuralpopulations can also be interpreted as functionallyactive modules. These explanations are based onthe different frequency systems of the brain whichare not defined in space and time precisely. Buteven then, such explanations seem more efficient inexplaining the dynamic aspect of the percepts.
In this report we have described numeroustypes of oscillatory activities with definitive ortentative explanations of their functional rela-tions. The results from several laboratories clearlydemonstrated that it is not possible to assign asingle function to a given type of oscillatory activ-ity. These oscillations have multiple functions andmay act as universal operators or codes of brainactivity. Besides frequency and site of oscillations,several other parameters are dependent on specificfunctions, namely enhancement, time locking, phaselocking, delay and duration of oscillations.
Stryker [1989] raised the question “Is grand-mother an oscillation?” by commenting that neu-rons in the visual cortex activated by the sameobject in the world, tend to discharge rhythmicallyand in unison. Concerning the question at the be-ginning of this section, our hypothesis about thefunctional role of event-related brain oscillationsis as follows: Complex and integrative brain func-tions are manifested in the superposition of severaldistributed multiple oscillations. According to thestudies reviewed above, [Basar et al., 2001] the ob-servation of the grandmother picture would activateoscillations, not only in the visual cortices,
but in all parts of the brain (probably includ-ing fronto-parietal delta, occipital alpha, theta andgamma oscillations). It was shown that complex sig-nals and associations (also remembering) are ac-companied with large increase of frontal theta anddelta and not only in occipital theta or gamma.Instead the huge alpha increase in occipital re-gions is not registered in frontal alpha. In animals,complex events are accompanied with increase infrontal and hippocampal oscillations. According tothese considerations, Fig. 20 presents the hypothet-ical distributed oscillations in the human cortex.Accordingly, distributed neural groups of all fre-quencies have to be involved in the processing of thiscomplicated percept [Desmedt et al., 1994; Dinseet al., 1997].
The changes in the duration of oscillations(delays or prolongation) were not presented in thishypothetical description. This illustration was a

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 483
Fig 20 Basar et al.
Fig. 20. A hypothetical description of selectively distributedoscillations manifested in the brain by vision (or imagination)of the grandmother-face. α, θ, γ, δ oscillations are illustratedwith large or small letters according to possible enhancementof oscillations. (Modified from [Basar et al., 2001].)
tentative approach, a design based on results thatalready exist in references. For example (1) Increaseof frontal theta and/or gamma by complex stimu-lation, (2) the increase of occipital alpha by visualstimulation while θ remains small, (3) increase ofθ in association areas during cognitive inputs areexplained in references (see [Basar et al., 1999]).
The illustration of Fig. 20 gave a rough idea ofthe probable superposition of multiple oscillationsthat are selectively distributed in various parts ofthe cortex. In the meantime, we started preliminaryexperiments to measure event-related oscillationsupon presentation of the grandmother face. We willdescribe the preliminary results in the next Sec. 7.We reproduced this hypothetical illustration in or-der to demonstrate the validity of the neurons-braintheory and the prediction-power of the concept of“super-synergy-in the whole brain”.
7. Grandmother-Paradigm and
Gestalt-Experiments
According to our hypothesis of “superbinding” inintegrative brain function the grandmother pic-ture would activate selectively distributed and co-herent oscillations in “the whole brain”, frontaltheta response, occipital alpha response and dis-tributed gamma, being major components [Basar,2001a–2001d]. This hypothesis was recently testedby recording Event Related Oscillations of 10 se-lected subjects by random sequence of presenta-tions of three stimulations: (1) the picture of an
elderly anonymous lady, (2) the picture of owngrandmother and (3) simple light.
7.1. Experimental strategy
We designed a strategy with the application of threeexperimental data recording-sets and three differenttypes of stimulations.
The stimulations consisted of,
(1) A simple light signal, its luminance being ap-prox at the same level of the stimulations 2 and3 described in the following.
(2) A stimulation signal containing the face of ananonymous elder lady.
(3) The face of own grandmother. The subjectswere known, healthy, young people, in the ageof 15–32 years. The pictures of the face ofown (genuine) grandmother of the subjects wereprepared prior to measurements.
7.1.1. Electrophysiological recording
The EEG was recorded from Ag/AgCl electrodesat F3, F4, Cz, C3, C4, P3, P4, O1 and O2 locationsaccording to the 10–20 system [Jasper, 1958]. Theelectrodes were affixed with Grass electrode paste.Linked earlobe electrodes served as reference. Elec-trode impedance did not exceed 5 kV. EOG medialupper and lateral orbital rim of the right eye wasalso registered. The EEG was amplified by means ofa Nihon Kohden EEG-4421 G EEG apparatus withband limits 0.1–70 Hz 24 dB/octave. The EEG wasdigitized online with a sampling rate of 250 sam-ples per second and stored on the hard disc of thecomputer for offline EEG analysis. An additionalnotch filter 36 dB octave was also applied to re-move the mains interference. For the recording ofEOG, the time constant 0.3 s with a low pass fil-ter at 70 Hz was used. All channels were displayedon paper and online by monitor scope in order toobserve both single trials and averaged signals. Anadditional EMG recording was carried out in orderto estimate the contribution of the motor potentialsto the main recording.
The first data recording set (random set): Allthree stimulations described above have been ap-plied in a random sequence with intervals varyingbetween 3.5 and 4.5 seconds. 75 stimulation signalshave been applied with approximately the same dis-tribution of each type of stimulation. Following therecordings, grandmother, face and light (controls)responses have been divided in subsets.
March 5, 2004 9:23 00927
c
FD
elta
4F
The
ta4
FA
lpha
4
OD
elta
2O
The
ta2
OA
lpha
2
Gre
en
Lig
ht
-Blu
eFa
ce
-Re
dG
rand
mo
the
r-
20-2 �V
20-2 �V
20-2 �V
20-2 �V
800
1000
msec
600
400
20
0-2
00
-60
0-8
00
-40
0-1
00
00
20-2 �V
800
1000
msec
600
400
20
0-2
00
-60
0-8
00
-40
0-1
00
00
800
1000
msec
600
400
20
0-2
00
-60
0-8
00
-40
0-1
00
00
800
1000
msec
600
400
20
0-2
00
-60
0-8
00
-40
0-1
00
00
20-2 �V
800
1000
msec
600
400
20
0-2
00
-60
0-8
00
-40
0-1
00
00
800
1000
msec
600
400
20
0-2
00
-60
0-8
00
-40
0-1
00
00
Fig
21
Ba
sar
et
al.
Fig
.21.
The
osc
illa
tory
resp
onse
sin
del
ta(0
.5–3.5
Hz)
,th
eta
(4–7
Hz)
,alp
ha
(8–13
Hz)
freq
uen
cyw
indow
sfo
rth
ree
types
of
stim
ulu
spre
senta
tions:
sim
ple
light,
face
,gra
ndm
oth
er(o
nly
fronta
l-F
4and
occ
ipital-O
2re
cord
ings)
.
484

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 485
The second data recording set (regularset): In the first subset of recordings 30 light sig-nals, in the second subset the anonymous face, andin the third subset picture of own grandmotherstimulation have been applied.
In all experiments subjects reported that they hadclearly recognized and differentiated the face oftheir own grandmother. The results were analyzedby digital filtering of data firstly in five frequencywindows of the EEG-convention. The unfilteredconventional EPs showed important topological dif-ferences in all type of stimulations. However, in thisreport for the sake of simplicity we will describeglobally only the filtered EPs that showed moredifferentiated results. This is important in order tounderstand the distributed nature of manifestationsto grandmother percept.
7.2. Event related oscillations to all
three stimulations:
Simple-light, anonymous face
and the grandmother picture
In this preliminary description we analyze grandaverage results consisting of 20 experiments with10 subjects. In these preliminary results, for thesake of simplicity, we describe only responses of 5locations and illustrate only event-related oscilla-tions in frontal and occipital areas. All individualexperiments showed very close results to that ofthe grand averages. Therefore, in these preliminaryresults manifesting very stable and clear differenti-ations in response to various stimuli, we do not yettackle with statistical analysis, in order to avoid themasking of the relevant preliminary findings.
7.2.1. Topology of delta responses
Delta responses are progressively increased fromlight experiments to face and grandmother. Proba-bly a phase change takes place between light andface/grandmother. Significant delta increases areobserved in posterior delta responses to Face andGrandmother.
In comparison to frontal delta responses pos-terior (P1, P2, O1 and O2) delta responses showincreases of 200%. The delta response to simplelight do not show at all this important and signif-icant increases of dominant delta response in faceand grandmother responses.
Accordingly, (1) the phase delay, (2) the in-crease of delta in posterior areas in face and
grandmother in comparison to light stimulationbelong to important findings in this strategy ofexperiments.
7.2.2. Topology of theta responses
Frontal theta responses also show important differ-ences for three types of stimuli: Theta response tosimple light stimulation is significantly smaller butmore prolonged to Face stimulation. Frontal thetais largest in the grandmother response (approx 8microvolts pp). Second largest is the frontal theta,which is only 5% lower. Theta response is decreas-ing from frontal to posterior areas.
In frontal area the grandmother response is 15%larger in comparison to simple light and 70% largerin comparison to face response. However we haveto note that frontal face theta response is moreprolonged in comparison to other stimulations. Incentral, parietal and occipital areas theta responseshave also more prolonged character in face-responseand grandmother-response in comparison to light-response. There is a second time window peaking ataround 300 ms in face and grandmother responses.Occipital light response is significantly higher incomparison to face and grandmother (60%). Themost important finding in this frequency range isthe significant dominance of frontal response. Notealso that the frontal theta response is the highest,and the second large one is the Cz response.
7.2.3. Topology of alpha responses
Frontal alpha responses to light-stimulation arenot phase-locked to stimulus, they are delayed andlow in amplitude also showing longer duration infrontal light response. However, phase-locked andlarge alpha responses are present in responses tograndmother and face stimulations. F3 and Cz
alpha responses to phase face and grandmotherstimulations are approx 250% larger. Occipital al-pha responses to grandmother face were very highand phase-locked at the signal application (0 ms).The results of light stimulation confirm our ear-lier data with significant enhancements (see Sec. 2).Upon face stimulation alpha response had lower am-plitudes and are not phase-locked in contrast tolight experiments, however the oscillation is pro-longed (possibly second window). In F4, C4, P4
and O2 less distinguished alpha responses wereobserved.
Occipital alpha responses to grandmothershowed approx 9 µV pp at the application of

March 5, 2004 9:23 00927
486 E. Basar et al.
stimulation and phase-locked, almost congruentwith occipital light alpha response. Note thesimilarity of alpha in light and grandmother stim-ulation in contrast to face stimulation. Occipitalalpha response to grandmother face is approx 300%larger in comparison to anonymous face response.Occipital high alpha response reflects the processof iconic memory (phyletic and built-in, persistentmemory). The occipital low and prolonged alpha re-sponse reflects the activation of semantic memory, atype of memory more abstract and in the case of anunknown face more difficult to identify and recog-nize. The low alpha amplitudes response may meanthat the numbers of neural populations activatedare smaller and have less synchrony in comparisonto grandmother activations. The prolongation of al-pha response possibly means that the occipital areais engaged or works for a longer period to identifythe yet unknown face. The same interpretations arealso valid for frontal theta response described in theformer section.
7.3. Distributed beta and gamma
responses
The preliminary findings in the beta and gammafrequency ranges also manifest selective topologi-cal distribution similar to those in lower frequencyranges. However, the results show larger individualfluctuations, accordingly, we shortly mention im-portant features without illustrations.
Frontal and C4 beta responses upon light werenot significant, however in P4 and O2 1 µV ppresponses were seen, P4 beta response was alsoobserved, but was less clear in comparison to C4
and O2. Gamma response to grandmother pictureshowed a clear pp 0.7 µV response in the grand aver-age of C4, P4 gamma-response being also a distinctone. Cz, C4 and P4 gamma responses had highestamplitudes upon face stimulation.
7.4. What are the grandmother
experiments saying? A
distributed template of
memory reactivation?
The preliminary, however tenable consequences ofthe so-called “Grandmother experiments” cannotbe fully described in this report. However we willindicate the most important and relevant conse-quence: The “Whole Brain” and “All Oscillations”are activated during recognition or remembering of
grandmother and anonymous face (at the beginningof experiments being an unknown face). The en-semble of responses behave as a three-dimensionalconstruct, consisting of temporal, spatial and fre-quency spaces. The responses to anonymous faceor to grandmother face are not represented solelyby one location and a unique frequency or at thesame position along the temporal axis, as differenti-ated delay and prolongation of multiple oscillationsthat are selectively distributed on the whole cortexshow.
7.4.1. Selectively distributed
enhancements in the whole cortex
The pictures of the grandmother and the anony-mous face elicit in the whole cortex (and accordingto the data and interpretations in previous sectionsof this paper) varied degrees of enhancements invarious frequency windows and recording sites. Ac-cordingly, the distributed alpha, beta, delta, theta,beta and gamma systems are selectively activated,thus again demonstrating one of the importantstatements of neurons-brain theory [Basar, 2003].Globally seen, selectively distributed enhancementsof amplitudes in response to grandmother pictureoriginate from a greater number of populations ofactivated neurons. This greater involvement of neu-ral populations in all frequencies require probablya more ample distribution of memory activation.
7.4.2. The distinction between episodic
and semantic memory
The distinction between episodic and semanticmemory is appealing and suggestive of two sep-arate memory systems, but the empirical evi-dence for corresponding neural systems is weak.Fuster [1995] states that this may be becausethese two implied memory systems are neu-rally inseparable. The phenomenology is somewhatobvious: The two kinds of declarative memory de-rive from, and blend into, each other without sharptransition.
Even if they can be separated as distinct en-tities they maintain a dynamic interaction throughlife. Fuster emphasizes also the neural level: “Thetwo memories may be based on the same corti-cal network, although perhaps different parts ofit, different levels of representational hierarchy forexplicit information. Nonetheless, it seems unwar-ranted to claim a separate cortical system or processfor each”.

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 487
7.5. The efficiency of the
grandmother paradigm for
differentiation of memory
components or stages
This paradigm show that it can serve as a goodstrategy to check mental disorders due to memoryreduction or complete memory-lost. The refinementof results and possible interpretations cannot beachieved by simple application of ERPs. Combina-tion and comparison of results of the whole strategy,different recording areas and all frequency win-dows, consideration of enhancement, delays andprolongations are most useful parameters as statedin neurons-brain theory. They are all needed fora deeper understanding of memory activation orreactivated memory components.
The components of memories are working ina given template comprehending the brain. Theyare major activation areas similar to major oper-ating rhythms described by Basar [1999]: Frontaltheta is the major operating rhythm, frontal thetaresponse plays a major role in differentiating se-mantic and episodic memories. Alpha activity isthe major operating occipital rhythm, it plays amajor role differentiating the memory related tograndmother-episodic memory and to the semanticmemory activated by faces.
8. Conclusion
The data obtained with the strategy of grand-mother experiments do not favor the idea of simplebinding as discussed in several papers and es-pecially in the previous Sec. 6. The theory ofsuper-synergy and superbinding predicted multi-ple frequency components selectively distributed inthe brain. Our data fit clearly with the frameworkof super-synergy and superbinding in the brain.Therefore the shortcoming of the simple bindingtheory to explain complex percepts has been clearlyshown. Two important points favor Hebb [1949]:(1) Increased electrical activity during learning andmemory performance and (2) cooperation of largerlong-distant neural populations.
The results and their differences are clear andshow important differentiation even between theunknown and grandmother face. Accordingly thestrategies of the Grandmother paradigm may opennew horizons in search of memories or evolvingmemories, and possibly provide relevant clinicalapplications.
Acknowledgments
The grandmother experiment and analysis de-scribed in the paper were performed with the closecollaboration of the University of Bremen, Instituteof Psychology and Cognition Research, Bremen,Germany and Dokuz Eylul University, BiophysicsDepartment, Izmir, Turkey and supported byTUBITAK-DFG grant 446 TUR 112/14/01.
References
Adrian, E. D. [1942] “Olfactory reactions in the brain ofthe hedgehog,” J. Physiol. (Lond.) 100, 459–473.
Baddeley, A. (ed.) [1986] Working Memory (ClarendonPress, Oxford).
Baddeley, A. [1992] “Working memory,” Science 255,556–559.
Baddeley, A. [1996] “The fractionation of workingmemory,” Proc. Natl. Acad. Sci. USA 93, 13468–13472.
Baird, B. [1999] “An oscillating cortical model ofauditory attention and electrophysiology,” Neurocom-puting 26 & 27, 319–328.
Barlow, H. B. [1972] “Single units and sensation: A neu-ron doctrine for perceptual psychology,” Perception1, 371–394.
Basar, E. & Ozesmi, C. [1972] “The hippocampal EEG-activity and a systems analytical interpretation ofaveraged evoked potentials of the brain,” Kybernetik.12, 45–54.
Basar, E. & Ungan, P. [1973] “A component analysis andprinciples derived for the understanding of evoked po-tentials of the brain: Studies in the hippocampus,”Kybernetik. 12, 133–140.
Basar, E., Gonder, A., Ozesmi, C. & Ungan, P. [1975]“Dynamics of brain rhythmic and evoked potentials.III, Studies in the auditory pathway, reticular forma-tion, and hippocampus during sleep,” Biol. Cybern.20, 161–169.
Basar, E. [1980] EEG-Brain Dynamics (Elsevier,Amsterdam).
Basar, E. [1983] “Toward a physical approach to inte-grative physiology, I. Brain dynamics and physicalcausality,” Am. J. Physiol. 245, 510–533.
Basar, E. & Stampfer, H. G. [1985] “Important associa-tions among EEG-dynamics, event-related potentials,short-term memory and learning,” Int. J. Neurosci.26, 161–80.
Basar, E., Rosen, B., Basar-Eroglu, C. & Greitschus, F.[1987] “The associations between 40-Hz EEG and themiddle latency response of the auditory evoked po-tential,” Int. J. Neurosci. 33, 103–117.
Basar, E. [1988] “EEG-dynamics and evoked potentialsin sensory and cognitive processing by the brain,”

March 5, 2004 9:23 00927
488 E. Basar et al.
in Dynamics of Sensory and Cognitive Processing bythe Brain, ed. Basar, E. (Springer, Berlin Heidelberg,NY), pp. 30–55.
Basar, E. [1992] “Brain natural frequencies are causalfactors for resonances and induced rhythms (Epi-logue),” in Induced Rhythms in the Brain, eds. Basar,E. & Bullock, T. H. (Birkhauser, Boston, MA),pp. 425–467.
Basar, E. & Bullock, T. H. (eds.) [1992] Induced Rhythmsin the Brain (Birkhauser, Boston, MA).
Basar, E. & Schurmann, M. [1994] “Functional aspectsof evoked alpha and theta responses in humans andcats. Occipital recordings in ‘cross modality’ experi-ments,” Biol. Cybern. 72, 175–183.
Basar, E., Hari, R., Lopes da Silva, F. H. & Schurmann,M. (eds.) [1997] “Brain alpha activity — new aspectsand functional correlates,” Int. J. Psychophysiol. 26,1–482.
Basar, E. [1998] Brain Function and Oscillations. I.Brain Oscillations: Principles and Approaches (BerlinHeidelberg, Springer), pp. 153–167.
Basar, E. [1999] Brain Function and Oscillations.II. Integrative Brain Function. Neurophysiology andCognitive Processes (Springer, Berlin Heidelberg),pp. 1–474.
Basar, E., Basar-Eroglu, C., Karakas, S. & Schurmann,M. [1999] “Are cognitive processes manifested inevent-related gamma, alpha, theta and delta oscilla-tions in the EEG?” Neurosci. Lett. 259, 165–168.
Basar, E., Basar-Eroglu, C., Karakas, S. & Schurmann,M. [2000] “Brain oscillations in perception and mem-ory,” Int. J. Psychophysiol. 35, 95–124.
Basar, E., Basar-Eroglu, C., Karakas, S. & Schurmann,M. [2001a] “Gamma, alpha, delta, and theta oscilla-tions govern cognitive processes,” Int. J. Psychophys-iol. 39, 241–248.
Basar, E., Schurmann, M., Demiralp, T., Basar-Eroglu,C. & Ademoglu, A. [2001b] “Event-related oscillationsare ‘real brain responses’ — wavelet analysis and newstrategies,” Int. J. Psychophysiol. 39, 91–127.
Basar, E., Ozgoren, M. & Karakas, S. [2001c] “A braintheory based on neural assemblies and superbind-ing,” in Wahrnehmen und Erkennen, eds. Reuter, H.,Schwab, P., Kleiber, D. & Gniech (PABST SciencePublishers, Lengerich), pp. 11–24.
Basar, E., Ozgoren, M. & Karakas, S. [2001d] “Su-perbinding in integrative brain function and mem-ory,” Proc. 23rd Annual Int. Conf. IEEE Engineeringin Medicine and Biology Society.
Basar, E. [2004] “Macrodynamics of electrical activity inthe whole brain: A review and tutorial report,” Int.J. Bifurcation and Chaos 14, 363–381.
Basar-Eroglu, C. & Basar, E. [1991] “A compound P300-40 Hz response of the cat hippocampus,” Int. J. Neu-rosci. 60, 227–237.
Basar-Eroglu, C., Basar, E., Demiralp, T. & Schurmann,
M. [1992] “P300-response: Possible psychophysiologi-cal correlates in delta and theta frequency channels,A review,” Int. J. Psychophysiol. 13, 161–179.
Basar-Eroglu, C., Struber, D., Schurmann, M., Stadler,M. & Basar, E. [1996a] “Gamma-band responses inthe brain: A short review of psychophysiological corre-lates and functional significance,” Int. J. Psychophys-iol. 24, 101–112.
Basar-Eroglu, C., Struber, D., Kruse, P., Basar, E. &Stadler, M. [1996b] “Frontal gamma-band enhance-ment during multistable visual perception,” Int. J.Psychophysiol. 24, 113–125.
Basar-Eroglu, C. & Demiralp, T. [2001] “Event-relatedtheta oscillations: An integrative and comparativeapproach in the human and animal brain,” Int. J.Psychophysiol. 39, 167–195.
beim Graben, P., Saddy, J. D. & Schlesewsky, M. [2000]“Symbolic dynamics of event-related brain poten-tials,” Phys. Rev. E62, p. 4.
Best, P. J. & Ranck, J. B. Jr. [1982] “Reliability of therelationship between hippocampal unit activity andsensory-behavioral events in the rat,” Exp. Neurol.75, 652–664.
Birbaumer, N., Elbert, T., Canavan, A. G. M. &Rockstroh, B. [1990] “Slow potentials of the cerebralcortex and behavior,” Physiol. Rev. 70, 1–41.
Bullock, T. H. & McClune, M. C. [1989] “Lateralcoherence of the electrocorticogram: A new measureof brain synchrony,” EEG Clin. Neurophysiol. 73,479–498.
Bullock, T. H. [1992] “Introduction to induced rhythms:A widespread, heterogeneous class of oscillations,”in Induced Rhythms in the Brain, eds. Basar, E. &Bullock, T. H. (Birkhauser, Boston, MA), pp. 1–26.
Burgess, A. P. & Gruzelier, J. H. [2000] “Short durationpower changes in the EEG during recognition memoryfor words and faces,” Psychophysiol. 37, 596–606.
Chen, A. C. N. & Hermann, C. S. [2001] “Per-ception of pain coincides with the spatial expan-sion of electroencephalographic dynamics in humansubjects,” Neurosci. Lett. 297, 183–186.
Cohen, J. D., Perlstein, W. M., Braver, T. S., Nystrom,L. E., Noll, D. C. & Smith, E. E. [1997] “Temporal dy-namics of brain activation during a working memorytask,” Nature 386, 604–608.
Courtney, S. M., Ungerleider, L. G., Keil, K. & Haxby,J. V. [1997] “Transient and sustained activity in a dis-tributed neural system for human working memory,”Nature 386, 608–611.
Damasio, A. R. & Damasio, H. [1994] “Cortical systemsfor retrieval of concrete knowledge: The convergencezone framework,” in Large Scale Neuronal Theories ofthe Brain, eds. Koch, C. & Davis, J. L. (MIT Press,Cambridge, MA), pp. 61–74.
Demiralp, T. & Basar, E. [1992] “Theta rhythmicities

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 489
following expected visual and auditory targets,” Int.J. Psychophysiol. 13, 147–160.
Demiralp, T., Ademoglu, A., Schurmann, M., Basar-Eroglu, C. & Basar, E. [1999] “Detection of P300waves in single trials by the wavelet transform (WT),”Brain Lang. 66, 108–128.
Desmedt, J. E. & Tomberg, C. [1994] “Transient phase-locking of 40 Hz electrical oscillations in prefrontaland parietal human cortex reflects the process ofconscious somatic perception,” Neurosci. Lett. 168,126–129.
Dinse, H. R., Kruger, K., Akhavan, A. C., Spengler, F.,Schoner, G. & Schreiner, C. E. [1997] “Low- frequencyoscillations of visual, auditory and somatosensory cor-tical neurons evoked by sensory stimulation,” Int. J.Psychophysiol. 26, 205–227.
Donchin, E. & Coles, M. G. H. [1988] “Is the P300 com-ponent a manifestation of context updating?” BrainBehav. Sci. 11, 357–374.
Doppelmayr, M., Klimesch, W., Schwaiger, J., Stadler,V. & Rohm, D. [2000] “The time locked theta responsereflects interindividual differences in human memoryperformance,” Neurosci. Lett. 278, 141–144.
Eckhorn, R., Bauer, R., Jordan, R., Brosch, W., Kruse,M., Munk, M. & Reitboeck, H. J. [1988] “Coherentoscillations: A mechanism of feature linking in the vi-sual cortex?” Biol. Cybern. 60, 121–130.
Egner, T. & Gruzelier, J. H. [2001] “Learned self-regulation of EEG frequency components affectsattention and event-related brain potentials inhumans,” Neuroreport. 12, 4155–4159.
Fabiani, M., Karis, D. & Donchin, E. [1990] “Effects ofmnemonic strategy manipulation in a Van Restorffparadigm,” Electroencephalogr. Clin. Neurophysiol.75, 22–35.
Fell, J., Hinrichs, H. & Roschke, J. [1997] “Time courseof 40 Hz EEG activity accompanying P3 responses inan auditory oddball paradigm,” Neurosci. Lett. 235,121–124.
Fell, J., Klaver, P., Lehnertz, K., Grunwald, T.,Schaller, C., Elger, C. E. & Fernandez, G. [2001]“Human memory formation is accompanied byrhinal-hippocampal coupling and decoupling,” NatureNeurosci. 4, 1259–1264.
Fuster, J. M. [1995] “Memory in the cortex of theprimate,” Biol. Res. 28, 59–72.
Fuster, J. M. [1997] “Network memory,” TrendsNeurosci. 20, 451–459.
Goldman-Rakic, P. [1997] “Space and time in the mentaluniverse,” Nature 386, 559–560.
Gray, C. M. & Singer, W. [1989] “Stimulus-specific neu-ronal oscillations in orientation columns of cat visualcortex,” Proc. Natl. Acad. Sci. USA 86, 1698–1702.
Gruzelier, J. (ed.) [1996] “New advances in EEG andcognition,” Int. J. Psychophysiol. 24, 1–187.
Gurtubay, I. G., Alegre, M., Labarga, A., Malanda,A., Iriarte, J. & Artieda, J. [2001] “Gamma bandactivity in an auditory oddball paradigm studiedwith the wavelet transform,” Clin. Neurophysiol. 112,1219–1228.
Haenschel, C., Baldeweg, T., Croft, R. J., Whittington,M. & Gruzelier, J. [2000] “Gamma and beta frequencyoscillations in response to novel auditory stimuli: Acomparison of human electroencephalogram (EEG)data with in vitro models,” Proc. Nat. Acad. Sci. USA13, 7645–7650.
Hebb, D. O. (ed.) [1949] The Organization of Behavior(Wiley, NY).
Howard, L. & Polich, J. [1985] “P300 latency and mem-ory span development,” Dev. Psychol. 21, 283–289.
Kalcher, J. & Pfurtscheller, G. [1995] “Discrimina-tion between phase-locked and non-phase-lockedevent related EEG activity,” Electroencephalogr. Clin.Neurophysiol. 5, 381–384.
Karakas, S. & Basar, E. [1998] “Early gamma responseis sensory in origin, A conclusion based on crosscomparison of results from multiple experimentalparadigmes,” Int. J. Psychophysiol. 31, 13–31.
Karakas, S., Erzengin, O. U. & Basar, E. [2000a] “Anew strategy involving multiple cognitive paradigmsdemonstrates that ERP components are determinedby the superposition of oscillatory responses,” Clin.Neurophysiol. 111, 1719–1732.
Karakas, S., Erzengin, O. U. & Basar, E. [2000b] “Thegenesis of human event-related responses explainedthrough the theory of oscillatory neural assemblies,”Neurosci. Lett. 285, 45–48.
Keil, A., Gruber, T. & Muller, M. M. [2001] “Functionalcorrelates of macroscopic high-frequency brain activ-ity in the human visual system,” Neurosci. Biobehav.Rev. 25, 527–534.
Kiss, T., Orban, G., Lengyel, M. & Erdi, P. [2001]“Intrahippocampal gamma and theta rhythm gener-ation in a network model of inhibitory interneurons,”Neurocomputing 38 40, 713–719.
Klimesch, W. [1999] “EEG alpha and theta oscillationsreflect cognitive and memory performance: A reviewand analysis,” Brain Res. Rev. 29, 169–195.
Klimesch, W., Doppelmayr, M., Schwaiger, J., Winkler,T. & Gruber, W. [2000] “Theta oscillations and theERP old/new effect: Independent phenomena?” Clin.Neurophysiol. 111, 781–793.
Klimesch, W., Schimke, H. & Schwaiger, J. [1994]“Episodic and semantic memory: An analysis in theEEG theta and alpha band,” Electroencephalogr. Clin.Neurophysiol. 91, 428–441.
Kocsis, B., Viana Di Prisco, G. & Vertes, R. P. [2001]“Theta synchronization in the limbic system: The roleof Gudden’s tegmental nuclei,” Europ. J. Neurosci.13, 381–388.

March 5, 2004 9:23 00927
490 E. Basar et al.
Kolev, V. & Yordanova, J. [1997] “Analysis of phase-locking is informative for studying event-related EEGactivity,” Biol. Cybern. 76, 229–235.
Kolev, V., Yordanova, J., Schurmann, M. & Basar, E.[1999] “Event-related alpha oscillations in task pro-cessing,” Clin. Neurophysiol. 110, 1784–1792.
Koshino, Y. & Niedermeyer, E. [1975] “Enhancementof rolandic mu rhythm by pattern vision,” Electroen-cephalogr. Clin. Neurophysiol. 38, 535–538.
Krause, C. M., Sillanmaki, L., Haggqvist, A. & Heino,R. [2001] “Test-retest consistency of the event-relateddesynchronization/event-related synchronization ofthe 4–6, 6–8, 8–10 and 10–12 Hz frequency bandsduring a memory task,” Clin. Neurophysiol. 112,750–757.
Kuhlman, W. N. [1978] “Functional topography of thehuman mu rhythm,” Electroencephalogr. Clin. Neuro-physiol. 44, 83–93.
Lettvin, J. Y., Maturana, H. R., McCulloch, W. S. &Pitts, W. H. [1959] “What the frog’s eye tells the frog’sbrain,” Proc. Inst. R.E. NY 47, 1940–1951.
Leung, L. S. [1992] “Fast (beta) rhythms in the hip-pocampus: A review,” Hippocampus 2, 93–98.
Llinas, R. R. & Ribary, U. [1992] “Rostrocaudal scanin the human brain: A global characteristic of the40 Hz response during sensory input,” in InducedRhythms in the Brain, eds. Basar, E. & Bullock, T. H.(Birkhauser, Boston), pp. 147–154.
Maltseva, I., Geissler, H. G. & Basar, E. [2000] “Alphaoscillations as an indicator of dynamic memory op-erations — anticipation of omitted stimuli,” Int. J.Psychophysiol. 36, 185–197.
Mesulam, M. M. [1990] “Large scale neurocognitivenetworks and distributed processing for attention,language, and memory,” Ann. Neurol. 28, 597–613.
Mesulam, M. M. [1994] “Neurocognitive networksand selectively distributed processing,” Rev. Neurol.(Paris) 150, 564–569.
Miller, R. (ed.) [1991] Cortico-Hippocampal Interplayand the Representation of Contexts in the Brain(Berlin Heidelberg, Springer).
Miltner, W., Braun, C., Arnold, M., Witte, H. & Taub,E. [1999] “Coherence of gamma-band EEG activity asa basis for associative learning,” Nature 397, 434–436.
Mountcastle, V. B. [1992] “Preface,” in Induced Rhythmsin the Brain, eds. Basar, E. & Bullock, T. H.(Birkhauser, Boston, MA), pp. 217–231.
Naatanen, R. (ed.) [1992] Attention and Brain Function(Hillsdale, NJ, Erlbaum).
Pantev, C., Makeig, S., Hoke, M., Galambos, R., Hamp-son, S. & Gallen, C. [1991] “Human auditory evokedgamma-band magnetic fields,” Proc. Natl. Acad. Sci.USA 88, 8996–9000.
Pfurtscheller, G., Neuper, C., Andrew, C. & Edlinger,G. [1997] “Hand and foot area mu rhythms,” Int. J.Psychophysiol. 26, 121–135.
Pratt, H., Michalewski, H. J., Barrett, G. & Starr, A.
[1989] “Brain potentials in a memory-scanning task. I.Modality and task effects on potentials to the probes,”Electroencephalogr. Clin. Neurophysiol. 72, 407–421.
Quiroga, R. Q., Rosso, O. A. & Basar, E. [1999] “Waveletentropy: A measure of order in evoked potentials,”Electrenceph. Clin. Neurophysiol. Suppl. 49, 299–303.
Quiroga, R. Q., Rosso, O. A., Basar, E. & Schurmann,M. [2001] “Wavelet entropy in event-related poten-tials: a new method shows ordering of EEG oscilla-tions,” Biol. Cybern. 84, 291–299.
Roschke, J., Mann, K., Riemann, D., Frank, C. & Fell,J. [1995] “Sequential analysis of the brain’s transferproperties during consecutive REM episodes,” Elec-troencephalogr. Clin. Neurophysiol. 96, 390–397.
Rosso, O. A., Blanco, S., Yordanova, J., Kolev, V., Figli-ola, A., Schurmann, M. & Basar, E. [2001] “Waveletentropy: A new tool for analysis of short durationbrain electrical signals,” J. Neurosci. Meth. 105,65–75.
Saermark, K., Mikkelsen, K. B. & Basar, E. [1992] “Mag-netoencephalographic evidence for induced rhythms,”in Induced Rhythms in the Brain, eds. Basar,E. & Bullock, T. H. (Birkhauser, Boston, MA),pp. 129–145.
Sakowitz, O. W., Schurmann, M. & Basar, E. [2000]“Oscillatory frontal theta responses are increasedupon bisensory stimulation,” Clin. Neurophysiol. 111,884–893.
Sakowitz, O. W., Quiroga, R. Q., Schurmann, M. &Basar, E. [2001] “Bisensory stimulation increasesgamma-responses over multiple cortical regions,”Cogn. Brain Res. 11, 267–279.
Sanquist, Th. F., Rohrbaugh, J. W., Syndulko, K. &Lindsley, D. B. [1980] “Electrocortical signs of lev-els of processing: Perceptual analysis and recognitionmemory,” Psychophysiol. 17, 568–576.
Schurmann, M., Basar-Eroglu, C., Rahn, E., Braasch,M., Dossel, O., Fuchs, M. & Basar, E. [1992] “Acomparative study of alpha responses in humanMEG temporoparietal and occipital recordings andcat intracranial EEG recordings,” Proc. IEEE Symp.Neuroscience and Technology (Lyon), pp. 132–137.
Schurmann, M., Basar-Eroglu, C., Kolev, V. & Basar, E.[1995] “A new metric for analyzing single-trial event-related potentials (ERPs) application to human visualP300 delta response,” Neurosci. Lett. 197, 167–170.
Schurmann, M., Basar-Eroglu, C. & Basar, E. [1997]“Gamma responses in the EEG: Elementary signalswith multiple functional correlates,” Neuroreport 8,531–534.
Schurmann, M., Demiralp, T., Basar, E. & Basar-Eroglu, C. [2000] “Electroencephalogram alpha (8–15 Hz) responses to visual stimuli in cat cortex,thalamus, and hippocampus: A distributed alphanetwork?” Neurosci. Lett. 292, 175–178.
Schutt, A., Basar, E. & Bullock, T. H. [1999] “Powerspectra of ongoing activity of the snail brain can

March 5, 2004 9:23 00927
Super-Synergy in the Brain: The Grandmother Percept is Manifested by Multiple Oscillations 491
discriminate odorants, Comparative Biochemistryand Physiology — Part A,” Molecul. Integr. Physiol.123, 95–110.
Silva, L. R., Amitai, Y. & Connors, B. G. [1991] “In-trinsic oscillations of neocortex generated by layer 5pyramidal neurons,” Science 251, 432–435.
Singer, W. & Gray, C. M. [1995] “Visual feature integra-tion and the temporal correlation hypothesis,” Ann.Rev. Neurosci. 18, 555–586.
Solodovnikov, V. V. [1960] Introduction to the Statisti-cal Dynamics of Automatic Control Systems (Dover,NY).
Srinivasan, R., Nunez, P. L. & Silberstein, R. B. [1998]“Spatial filtering and neocortical dynamics: Estimatesof EEG coherence,” IEEE Trans. Biomed. Engin. 45,814–826.
Stampfer, H. G. & Basar, E. [1985] “Does frequencyanalysis lead to better understanding of human eventrelated potentials,” Int. J. Neurosci. 26, 181–196.
Stein, A. Von & Sarnthein, J. [2000] “Different frequen-cies for different scales of cortical integration: From
local gamma to long range alpha/theta synchroniza-tion,” Int. J. Psychophysiol. 38, 301–313.
Steriade, M., Gloor, P., Llinas, R. R., Lopes da Silva,F. H. & Mesulam, M. [1990] “Basic mechanisms ofcerebal rhythmic activities,” Electroencephalogr. Clin.Neurophysiol. 76, 481–508.
Stryker, M. P. [1989] “Is grandmother an oscillation?”Nature 338, 297–298.
Tallon-Baudry, C., Bertrand, O., Peronnet, F. & Pernier,J. [1998] “Induced gamma-band activity during thedelay of a visual short-term memory task in humans,”J. Neurosci. 18, 4244–4254.
Yordanova, J., Kolev, V. & Demiralp, T. [1997] “Thephase-locking of auditory gamma band responses inhumans is sensitive to task processing,” Neuroreport8, 3999–4004.
Yordanova, J., Kolev, V., Rosso, O. A., Schurmann,M., Sakowitz, O.W., Ozgoren, M. & Basar, E. [2002]“Wavelet entropy analysis of event-related potentialsindicates modality-independent theta dominance,” J.Neurosci. Meth. 117, 99–109.
Related Documents











