Study of the effect of lethal and sublethal pH and a w stresses on the inactivation or growth of Listeria monocytogenes and Salmonella Typhimurium Alexandros Tiganitas a , Nikoleta Zeaki a , Antonia S. Gounadaki b , Eleftherios H. Drosinos a , Panagiotis N. Skandamis a, ⁎ a Agricultural University of Athens, Department of Food Science & Technology, Laboratory of Food Hygiene and Quality Control, Iera Odos 75, Athens,11855, Greece b Agricultural University of Athens, Department of Food Science & Technology, Laboratory of Microbiology & Biotechnology of Foods, Greece abstract article info Available online xxxx Keywords: Stress Acid Osmotic Listeria monocytogenes Sequence Salmonella During food processing, microorganisms are commonly exposed to multiple sublethal or lethal stresses (commonly a w , pH) sequentially or simultaneously. The objectives of the present study were: (i) to comparatively evaluate the survival of Listeria monocytogenes and Salmonella Typhimurium in lethal acid (pH 4.0 and 4.5 with lactic acid) or osmotic conditions (15 and 20% NaCl), applied singly, sequentially (pH then NaCl or NaCl then pH), or simultaneously at 5 and 10 °C; and, (ii) to quantify the effect of osmotic shifts at pH 7.0, 6.0 or 5.0 (adjusted with lactic acid) on the lag phase and growth rate of L. monocytogenes at 10 °C. In sequential lethal stress applications, the second stress was applied 2 or 3 days after the first for Salmonella and L. monocytogenes, respectively. Acid tolerance of L. monocytogenes was higher than osmotic tolerance and the opposite was observed for Salmonella. Higher inactivation was observed after exposure to pH 4.0 compared to pH 4.5 as well as after exposure to 20% NaCl compared to 15% NaCl. Exposure to stresses sequentially resulted in faster (P b 0.05) reductions than the exposure to single or double stresses applied simultaneously. The pH then NaCl sequence was more detrimental for pathogens than the reverse sequence. Incubation temperature (5 and 10 °C) did not show any profound (P b 0.05) effect on microbial inactivation. When L. monocytogenes was incubated at a w 0.930 or 0.995 at 30 °C, then the lag phase increased both in subsequent osmotic downshift and upshift, respectively, at 10 °C. Shorter lag phase and higher ability to initiate growth at lower a w was observed after pre-adaptation at pH 6.0 or 5.0 compared to neutral pH. The results may contribute to the review of critical limits in low pH (with lactic acid) and water activity products, considering the risk of L. monocytogenes and Salmonella survival. In addition, the present indications may address the points in processing where stricter sanitation procedures should be applied in order to minimize the risk of survivors. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Food safety relies primarily on the use of multiple hurdle technology (Leistner, 2000). Therefore, depending on processing, pathogens may be exposed to various stresses simultaneously (e.g., on ripened cheese or meat) or sequentially (e.g., during fermentation and ripening of meat, milk or plant products as well as drying of marinated meat; Niksic et al., 2005; Yoon et al., 2006). Of the well-established hurdles, acid and osmotic stresses, are most commonly applied in food processing (e.g., by acidification or addition of NaCl), or naturally encountered in foods (e.g., low pH of fruits, water evaporation, etc.). The sequence of hurdles has been shown to be crucial for the survival of pathogens and studies taking into account this concept may contribute to assess the risk of pathogen survival or growth depending on the sequence of hurdles that has or will be exposed to. For instance, Shadbolt et al. (2001) demonstrated that exposure of Escherichia coli to lethal pH (pH 3.5) followed by exposure to lethal NaCl (10%, a w 0.90), a sequence, which simulates the order of stresses encountered during fermentation and ripening, is more deleterious than the reverse sequence, or the simultaneous exposure to both stresses, a situation that simulates post-fermentation inoculation. However, the above study used HCl to reduce the pH to 3.5. Therefore, approaches using an organic acid, such as lactic acid, which is commonly present in meat products, at the targeted pH values in meat or cheese fermentations (e.g., 4.6–5.0) would also be useful and are currently missing. Pathogens have a variable resistance to the processes involving acid and/or osmotic stress (Adams and Nicolaides, 1997; Nightingale et al., 2006). For instance, Listeria monocytogenes was found to survive fermentation (pH 4.39–4.63), drying (a w 0.76–0.80) and subsequent storage of dry fermented sausage for 49 days at 17–23 °C, with less than 3 log 10 CFU/g reductions (Lahti et al., 2001; Nissen and Holck, 1998; Thévenot et al., 2005). Conversely, 4 log units of Salmonella International Journal of Food Microbiology xxx (2009) xxx–xxx ⁎ Corresponding author. Tel.: +30 210 5294684; fax: +30 210 5294683. E-mail address: [email protected] (P.N. Skandamis). FOOD-04692; No of Pages 9 0168-1605/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.ijfoodmicro.2009.02.016 Contents lists available at ScienceDirect International Journal of Food Microbiology journal homepage: www.elsevier.com/locate/ijfoodmicro ARTICLE IN PRESS Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal and sublethal pH and a w stresses on the inactivation or growth of Listeria monocytogenes and Salmonella Typhimurium, International Journal of Food Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

International Journal of Food Microbiology xxx (2009) xxx–xxx
FOOD-04692; No of Pages 9
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
ARTICLE IN PRESS
Study of the effect of lethal and sublethal pH and aw stresses on the inactivation orgrowth of Listeria monocytogenes and Salmonella Typhimurium
Alexandros Tiganitas a, Nikoleta Zeaki a, Antonia S. Gounadaki b,Eleftherios H. Drosinos a, Panagiotis N. Skandamis a,⁎a Agricultural University of Athens, Department of Food Science & Technology, Laboratory of Food Hygiene and Quality Control, Iera Odos 75, Athens, 11855, Greeceb Agricultural University of Athens, Department of Food Science & Technology, Laboratory of Microbiology & Biotechnology of Foods, Greece
⁎ Corresponding author. Tel.: +30 210 5294684; fax:E-mail address: [email protected] (P.N. Skandamis).
0168-1605/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.ijfoodmicro.2009.02.016
Please cite this article as: Tiganitas, A., et al., Smonocytogenes and Salmonella Typhimurium
a b s t r a c t
a r t i c l e i n f oAvailable online xxxx
Keywords:
During food processing, m(commonly aw, pH) sequecomparatively evaluate the
StressAcidOsmoticListeria monocytogenesSequenceSalmonella
survival of Listeria monocytogenes and Salmonella Typhimurium in lethal acid (pH4.0 and 4.5 with lactic acid) or osmotic conditions (15 and 20% NaCl), applied singly, sequentially (pH thenNaCl or NaCl then pH), or simultaneously at 5 and 10 °C; and, (ii) to quantify the effect of osmotic shifts at pH7.0, 6.0 or 5.0 (adjusted with lactic acid) on the lag phase and growth rate of L. monocytogenes at 10 °C. Insequential lethal stress applications, the second stress was applied 2 or 3 days after the first for Salmonellaand L. monocytogenes, respectively. Acid tolerance of L. monocytogenes was higher than osmotic tolerance
icroorganisms are commonly exposed to multiple sublethal or lethal stressesntially or simultaneously. The objectives of the present study were: (i) to
and the opposite was observed for Salmonella. Higher inactivation was observed after exposure to pH 4.0compared to pH 4.5 as well as after exposure to 20% NaCl compared to 15% NaCl. Exposure to stressessequentially resulted in faster (Pb0.05) reductions than the exposure to single or double stresses appliedsimultaneously. The pH then NaCl sequence was more detrimental for pathogens than the reverse sequence.Incubation temperature (5 and 10 °C) did not show any profound (Pb0.05) effect on microbial inactivation.When L. monocytogenes was incubated at aw 0.930 or 0.995 at 30 °C, then the lag phase increased both insubsequent osmotic downshift and upshift, respectively, at 10 °C. Shorter lag phase and higher ability toinitiate growth at lower aw was observed after pre-adaptation at pH 6.0 or 5.0 compared to neutral pH. Theresults may contribute to the review of critical limits in low pH (with lactic acid) and water activity products,considering the risk of L. monocytogenes and Salmonella survival. In addition, the present indications mayaddress the points in processing where stricter sanitation procedures should be applied in order to minimizethe risk of survivors.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Food safety relies primarily on the use of multiple hurdletechnology (Leistner, 2000). Therefore, depending on processing,pathogensmay be exposed to various stresses simultaneously (e.g., onripened cheese ormeat) or sequentially (e.g., during fermentation andripening of meat, milk or plant products as well as drying of marinatedmeat; Niksic et al., 2005; Yoon et al., 2006). Of the well-establishedhurdles, acid and osmotic stresses, aremost commonly applied in foodprocessing (e.g., by acidification or addition of NaCl), or naturallyencountered in foods (e.g., low pH of fruits, water evaporation, etc.).The sequence of hurdles has been shown to be crucial for the survivalof pathogens and studies taking into account this concept maycontribute to assess the risk of pathogen survival or growth dependingon the sequence of hurdles that has or will be exposed to. For instance,
+30 210 5294683.
l rights reserved.
tudy of the effect of lethal an, International Journal of Fo
Shadbolt et al. (2001) demonstrated that exposure of Escherichia colito lethal pH (pH 3.5) followed by exposure to lethal NaCl (10%, aw0.90), a sequence, which simulates the order of stresses encounteredduring fermentation and ripening, is more deleterious than thereverse sequence, or the simultaneous exposure to both stresses, asituation that simulates post-fermentation inoculation. However, theabove study used HCl to reduce the pH to 3.5. Therefore, approachesusing an organic acid, such as lactic acid, which is commonly presentin meat products, at the targeted pH values in meat or cheesefermentations (e.g., 4.6–5.0) would also be useful and are currentlymissing.
Pathogens have a variable resistance to the processes involvingacid and/or osmotic stress (Adams and Nicolaides, 1997; Nightingaleet al., 2006). For instance, Listeria monocytogeneswas found to survivefermentation (pH 4.39–4.63), drying (aw 0.76–0.80) and subsequentstorage of dry fermented sausage for 49 days at 17–23 °C, with lessthan 3 log10 CFU/g reductions (Lahti et al., 2001; Nissen and Holck,1998; Thévenot et al., 2005). Conversely, 4 log units of Salmonella
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016

2 A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
Typhimurium were completely eliminated within 40 days of manu-facture and storage of pepperoni sausage at pH 4.6 and aw 0.798–0.828 (Nissen and Holck, 1998; Smith et al., 1975). It needs to be notedhowever, that the resistance of pathogens to food processes (andespecially fermentations) has been comparatively evaluated inproducts with marked variability in their recipes and manufacturingconditions and the results are not directly comparable. Moreover,available in vitro studies, commonly performed in laboratory media,usually lack of the comparative evaluation of the resistance ofpathogens to acid and osmotic stress applied singly or sequentiallyand in multiple combinations that simulate food processes. It wouldbe of significance for assessment of pathogen survival in foods to takeinto account the hurdles applied at different food manufacturingstages, where contamination occurs. Indeed, apart from the studies byShadbolt et al. (2001) and Shabala et al. (2008) on the resistance of E.coli and L. monocytogenes, respectively, to acid and osmotic stressapplied singly, simultaneously or sequentially, knowledge of suchresistance on other pathogens, such as Salmonella, is lacking.
In addition to the pH at the end of fermentation, the levels of awachieved by drying (ripening) are also known to affect the survival ofpathogens (Hew et al., 2005). Shabala et al. (2008) and Faleiro et al.(2003) reported on the inactivation rate of L. monocytogenes at 14%and 20% NaCl, respectively, whereas Koutsoumanis et al. (2003)monitored the death of L. monocytogenes at 10.5 and 20.5% NaCl. All ofthem observed increase of inactivation rate with NaCl level. However,detailed knowledge on osmotolerance of pathogens as well as incombination with acid stress is limited, especially for lengthy storage(e.g., N25 days), which is commonly applied in ripened products.
In addition to combinations of pH and aw, which cause bacterialdeath, a desirable food safety target is also the delay (or cessation) ofbacterial growth in sub-optimal osmotic or pH environments. Delay ingrowth is translated into extension of bacterial lag phase until bacteriarestore their physiological equilibrium upon transferring to differentenvironments and initiate growth. Mellfont et al. (2004) found thatwhen Gram negative bacteria, such as S. Typhimurium and E. coli aretransferred from high (0.993) to low (0.947) aw environments, theiradaptationwork, expressed as ‘relative lag time’, increases. Studies onGram positive bacteria, such as L. monocytogenes, have mainlyconcentrated on the effect of temperature shifts (Bovill et al., 2001;Delignette-Muller et al., 2005) or variable precultural conditions suchas pH (e.g. 5.7 or 7.4; Francois et al., 2007) or other single stresses, (i.e.,chlorine, starvation, biofilm formation, or NaCl stress; Guillier et al.,2005; Jacobsen and Koch, 2006) on the lag time of L. monocytogenes.Thus, the effect of osmotic up- or down-shifts on the lag time of thelatter bacterium needs further investigation.
The objectives of the present study were: (i) to comparativelyevaluate the effect of levels and sequence of acid and osmotic stress onL. monocytogenes and S. Typhimurium; and, (ii) to evaluate the effectof osmotic up- and down-shifts on the lag phase of L. monocytogenes.
2. Materials and methods
2.1. Bacterial cultures
L. monocytogenes Scott A (serotype 4b) and S. Typhimurium weremaintained in tryptic soy broth (TSB; LABM, International DiagnosticsGroup Plc., Bury, UK) with 20% glycerol at −20 °C. Strains wereactivated by transferring 100 µl of the stock cultures to 10 ml of TSBand incubating at 30 °C (L. monocytogenes) and 37 °C (S. Typhimur-ium) for 24 h. Then, for preparing cultures for survival experiments,aliquots (100 µl) were transferred to fresh glucose-free TSB (TSB-G)and incubated for 24 h at 30 or 37 °C for L. monocytogenes and S.Typhimurium, respectively. Aliquots (10 ml) of activated cultures(approx. 108.5 CFU ml−1) were centrifuged at 5000 ×g for 10 min andwashed twice in 10 ml quarter strength Ringer's solution (Merck;Germany) to be further used as inoculum. L. monocytogenes cultures
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
intended to be used for growth studies were grown in TSB at pH 7.2and aw of 0.995 at 30 °C.
2.2. Exposure to lethal acid and osmotic stress conditions
Both microorganisms were exposed to lethal acid (pH 4.0 and 4.5)and osmotic (15% and 20% NaCl at pH 7.2) stresses, singly, sequentially(first to pH and then to NaCl, and in the reverse order), orsimultaneously at 5 and 10 °C. Specifically, three series of TSB(90 ml; pH 7.2 and 0.5% NaCl) were prepared in glass containers(250 ml) by adding 14.5 or 19.5% of NaCl, or its pH was adjusted to 4.5or 4.0 with 85% DL-lactic acid (Sigma). All flasks were autoclaved at121 °C for 15 min and after cooling; they were inoculated with 0.5 mlof the washed cultures of both microorganisms in order to yield initiallevels of 106–7 CFU ml−1 and incubated at 5 and 10 °C. Based onpreliminary results, showing that Salmonella died markedly fasterthan L. monocytogenes within 2 days of exposure to the above singlepH stresses, we decided to apply the second lethal stress after 2 daysfor flasks inoculated with Salmonella and after 3 days of incubation offlasks inoculated with L. monocytogenes. The second stress (i.e.,reduction of pH, or increase of NaCl) was applied to 4 out of the 6 totalseries of flasks of each microorganism, in order to simulate sequentialexposure from low pH to high NaCl and the opposite, as follows; thepH of flasks containing 15 or 20% NaCl were aseptically adjusted to 4.0and 4.5, whereas flasks with 0.5% NaCl and pH 4.0 or 4.5 weresupplemented with 14.5 and 19.5% NaCl. In order to evaluate thesimultaneous exposure to acid and osmotic stress, two additionalseries of flasks containing 90 ml of TSB were prepared with 20% NaCland their pH adjusted to 4.0 or 4.5 by adding lactic acid. Flasks wereinoculated with 0.5 ml of the washed cultures and incubated at 5 and10 °C. Totally, each stress treatment was examined in at least twoindependent experiments and duplicate flasks were used in eachexperiment.
2.3. Effect of osmotic shifts on lag phase of L. monocytogenes
TSB supplemented with 0.6% yeast extract (YE powder; LAB M,International Diagnostics Group Plc., Bury, UK) was prepared (TSBYE;90ml) in250-ml glass containers andNaClwas added toyieldawvaluesof0.995 or 0.930. The pH of flasks with TSBYE at 0.995was further adjustedby adding lactic acid to the final values of 7.2, 6.0 and 5.0, whereas the pHof flasks with aw of 0.930 was 7.2. Duplicate flasks containing the growthmedia were autoclaved (121 °C for 15 min) and after cooling they wereinoculated with 0.5 ml of washed culture of L. monocytogenes. Theinoculated flasks were incubated at 30 °C until L. monocytogenes reachedthe late stationary phase (8.8±0.5 log CFU ml−1). Then, cells wereharvested by centrifugation as described in Section 2.1, serially diluted inquarter strength Ringer's solution and aliquots (0.1 ml) were used toinoculate a second series of flasks containing 90 ml of autoclaved TSBYEwith aw adjusted to 0.940, 0.950, 0.960, 0.970 and 0.980 by adding NaCl.These flasks were further incubated at 10 °C and growth of L.monocytogenes was monitored by plating on TSAYE as described in thenext paragraph. The above procedure was carried out twice.
2.4. Enumeration of bacteria
At regular time intervals, during incubation of the inoculatedflasks, aliquots (1 ml) of TSB or TSBYE were serially diluted in quarterstrength Ringer's solution and surface-plated on duplicate plates ofTryptic Soy Agar (TSA; Merck), supplemented with 0.6% yeast extract(TSAYE). Plates were incubated at 30 °C for 48 h for enumeration of L.monocytogenes and at 37 °C for 24 h for enumeration of S.Typhimurium. The latest endpoint of sampling time was after 649 or719 h of incubation, depending on the microorganism and tempera-ture of incubation. However, sampling for specific treatmentsinvolving double-stress (applied sequentially or simultaneously) was
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016
3A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
stopped earlier, when undetectable survivors (i.e., b0 log CFU ml−1)were observed in one of the two treatments of the following pairs: (i)two sequential treatments differing only in the order of NaCl and pHstress, or in one of the two stresses (e.g., 15% NaCl then pH 4.0compared to 20% NaCl then pH 4.0); (ii) one sequential and therespective simultaneous treatment; or, (iii) two identical treatmentsfor different pathogen.
2.5. Measurements of pH and aw
The pH of media was measured with a digital pH meter (pH 691,Metrohm). The aw was determined with a calibrated electrichygrometer (HygroLab, Rotronic, Bassersdorf, Switzerland) accordingto the manufacturer's instructions.
2.6. Fitting of data and statistical analysis
The log10 transformed data were fitted to the Baranyi model or thebiphasic model of DMFit ver 2.1 (available at http://www.ifr.ac.uk/Safety/DMfit/default.html), in order to estimate the survival andgrowth kinetics of bacteria tested. Given that the inactivation curveswere biphasic with no shoulder, the death rates (log10 h−1) of eachinactivation phase, as well as the time to the end of the first and thebeginning of the second inactivation phase (t1st; h) were estimated bythe fitting. For each growth curve, the maximum specific growth rate(μ; days−1), the lag time (tlag; days) and the work-to-be doneparameter (Eq. (1)) were estimated by the fitting.
ho = μ × tlag ð1Þ
The inactivation kinetics of Table 1 were analyzed by the generallinear model procedures of SPSS (release 10.0.1 © SPSS Inc.).Descriptive statistics (means and standard deviations) were com-
Table 1Parameter estimates of the biphasic inactivation kinetics of Listeria monocytogenes and Salmsimultaneously at 10 °C.
Microorganism Stresses a (log·h−1)a
L. monocytogenes Single 15% NaCl −0.003 AB (0.020% NaCl −0.009 ABC (0.pH 4.5 −0.002 A (0.00pH 4 −0.004 ABC (0.
Sequential 15% NaCl then pH 4.5 −0.013 ABC (0.015% NaCl then pH 4 −0.002 A (0.0020% NaCl then pH 4.5 −0.022 CD (0.020% NaCl then pH 4 −0.020 BCD (0.pH 4.5 then 15% NaCl −0.009 ABC (0.pH 4.5 then 20% NaCl −0.032 DE (0.01pH 4 then 15% NaCl −0.009 ABC (0.pH 4 then 20%NaCl −0.005 ABC (0.
Simultaneous pH 4.5+20% NaCl −0.036 DE (0.01pH 4+20% NaCl −0.040 E (0.007
S. Typhimurium Single 15% NaCl −0.029 A (0.02820% NaCl −0.030 A (0.021pH 4.5 −0.002 A (0.001pH 4 0.000 A (0.000)
Sequential 15% NaCl then pH 4.5 −0.020 A (0.0015% NaCl then pH 4 −0.017 A (0.02420% NaCl then pH 4.5 −0.019 A (0.00220% NaCl then pH 4 −0.039 AB (0.01pH 4.5 then 15% NaCl −0.015 A (0.006pH 4.5 then 20% NaCl −0.019 A (0.001pH 4 then 15% NaCl −0.080 B (0.010pH 4 then 20%NaCl −0.039 AB (0.05
Simultaneous pH 4.5+20% NaCl −0.023 A (0.015pH 4+20% NaCl −0.025 A (0.003
a Death rate of the first phase of inactivation curve.b Duration of the first phase of the inactivation curve.c Death rate of the second phase of the inactivation curve.d Adjusted R2 statistics of the fitting (minimum, maximum).e ABCDE: For each microorganism, means within a column sharing at least a common let
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
puted, and analysis of variance (ANOVA) was performed withpairwise t tests for statistical differences (Pb0.05) of individualkinetic estimates of L. monocytogenes and S. Typhimurium betweendifferent treatments. The least significant difference procedure wasused to perform mean separation analyses at significance level ofalpha=0.05.
3. Results and discussion
The majority of the inactivation curves for the treatments of thepresent study were biphasic, comprised of two distinct death phases(Figs. 1–3; Table 1). The parameters a, t1st, and b of the inactivationcurves (Table 1) were used to quantify the effect of storagetemperature and type (including sequence) of stress on the reductionof both pathogens. Analysis of variance indicated that incubationtemperature did not significantly (PN0.05) affect the inactivation ofpathogens, whereas the type of stress had a significant (Pb0.05)impact on the inactivation kinetics of bacteria (Table 1).
Exposure to single acid and osmotic stresses showed that bothpathogens were more sensitive to pH 4.0 than 4.5 or any of theosmotic stresses (i.e., 15 and 20% NaCl) tested (Figs. 1A and 2A;Table 1). More specifically, exposure to pH 4.0 totally inactivated L.monocytogenes and S. Typhimurium (approx. 7 log CFU ml−1
reduction) at both incubation temperatures (Figs. 1A, 2A and 3),whereas exposure to pH 4.5 for up to 719 h caused approximately 1.8and 4.3 log reductions at 5 °C and 1.2 and 4.0 log reductions at 10 °C forL. monocytogenes and S. Typhimurium, respectively (Figs. 1A, 2A and3). The increased resistance of microorganisms to pH 4.5 compared topH 4.0 was also evident by the longer duration of the first phase of theinactivation (Table 1). However, L. monocytogenes seemed to toleratelow pH more than S. Typhimurium, whereas both microorganismswere similarly resistant to 15% NaCl regardless of the incubationtemperature (Figs. 1A and 2A; Table 1). The higher acid resistance of L.
onella Typhimurium under various pH and NaCl stresses applied singly, sequentially or
t1st(h)b b (log·h−1)c dR2
01)e 137.900 BC (105.45) −0.005 A (0.000) 0.990 0.996006) 188.089 BC (133.97) −0.011 A (0.004) 0.981 0.9960) 868.963 A (45.97) −0.048 AB (0.029) 0.949 0.991002) 276.144 BC (83.24) −0.014 A (0.004) 0.976 0.98306) 80.801 C (95.03) −0.007 A (0.000) 0.837 0.9072) 19.068 C (17.27) −0.006 A (0.006) 0.832 0.90109) 11.107 C (7.63) −0.006 A (0.000) 0.878 0.946001) 151.622 BC (5.12) 0.000 A (0.000) 0.828 0.972003) 555.600 AB (649.97) −0.006 A (0.003) 0.888 0.9178) 167.945 BC (67.85) 0.000 A (0.000) 0.797 0.952007) 125.185 BC (128.45) −0.011 A (0.005) 0.914 0.941002) 51.396 C (30.63) −0.128 B (0.140) 0.852 1.0003) 59.442 C (14.58) −0.005 A (0.000) 0.859 0.932) 60.426 C (30.96) −0.002 A (0.001) 0.888 0.904) 108.419 A (94.589) −0.002 A (0.000) 0.967 0.981) 39.431 A (21.82) −0.004 A (0.000) 0.919 0.968) 153.604 A (40.41) −0.005 A (0.000) 0.913 0.981
17.438 A (4.51) −0.055 D (0.009) 0.800 0.9844) 107.939 A (28.20) −0.003 A (0.000) 0.964 0.992) 36.925 A (47.55) −0.020 BC (0.012) 0.779 0.895) 135.263 A (18.45) −0.002 A (0.003) 0.981 0.9915) 125.797 A (150.63) −0.021 C (0.004) 0.721 0.795) 140.947 A (193.29) −0.007 A (0.010) 0.958 0.967) 148.495 A (0.002) −0.006 A (0.001) 0.922 0.935) 4.269 A (0.000) −0.002 A (0.000) 0.954 0.9545) 12.189 A (0.000) −0.090 E (0.000) 0.954 0.979) 110.746 A (139.33) −0.009 AB (0.006) 0.954 0.966) 129.284 A (4.526) −0.002 A (0.000) 0.853 0.937
ter are not significantly different at the Pb0.05 level.
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016
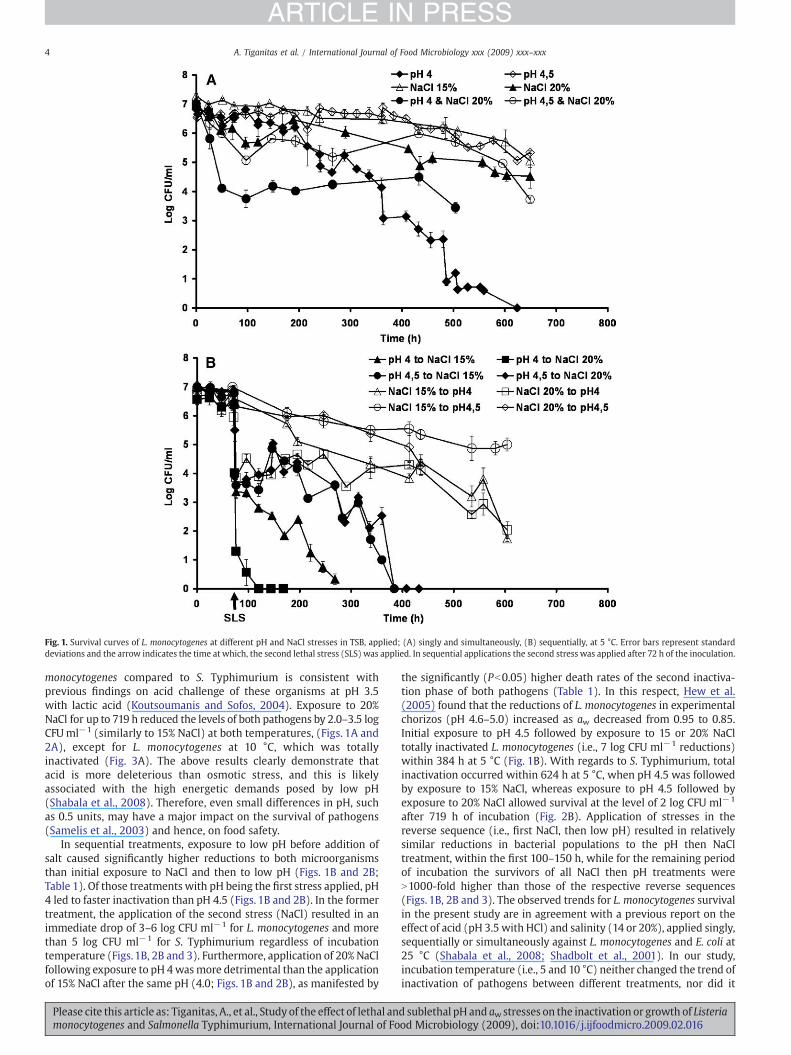
Fig. 1. Survival curves of L. monocytogenes at different pH and NaCl stresses in TSB, applied; (A) singly and simultaneously, (B) sequentially, at 5 °C. Error bars represent standarddeviations and the arrow indicates the time at which, the second lethal stress (SLS) was applied. In sequential applications the second stress was applied after 72 h of the inoculation.
4 A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
monocytogenes compared to S. Typhimurium is consistent withprevious findings on acid challenge of these organisms at pH 3.5with lactic acid (Koutsoumanis and Sofos, 2004). Exposure to 20%NaCl for up to 719 h reduced the levels of both pathogens by 2.0–3.5 logCFUml−1 (similarly to 15% NaCl) at both temperatures, (Figs. 1A and2A), except for L. monocytogenes at 10 °C, which was totallyinactivated (Fig. 3A). The above results clearly demonstrate thatacid is more deleterious than osmotic stress, and this is likelyassociated with the high energetic demands posed by low pH(Shabala et al., 2008). Therefore, even small differences in pH, suchas 0.5 units, may have a major impact on the survival of pathogens(Samelis et al., 2003) and hence, on food safety.
In sequential treatments, exposure to low pH before addition ofsalt caused significantly higher reductions to both microorganismsthan initial exposure to NaCl and then to low pH (Figs. 1B and 2B;Table 1). Of those treatments with pH being the first stress applied, pH4 led to faster inactivation than pH 4.5 (Figs. 1B and 2B). In the formertreatment, the application of the second stress (NaCl) resulted in animmediate drop of 3–6 log CFU ml−1 for L. monocytogenes and morethan 5 log CFU ml−1 for S. Typhimurium regardless of incubationtemperature (Figs. 1B, 2B and 3). Furthermore, application of 20% NaClfollowing exposure to pH 4wasmore detrimental than the applicationof 15% NaCl after the same pH (4.0; Figs. 1B and 2B), as manifested by
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
the significantly (Pb0.05) higher death rates of the second inactiva-tion phase of both pathogens (Table 1). In this respect, Hew et al.(2005) found that the reductions of L. monocytogenes in experimentalchorizos (pH 4.6–5.0) increased as aw decreased from 0.95 to 0.85.Initial exposure to pH 4.5 followed by exposure to 15 or 20% NaCltotally inactivated L. monocytogenes (i.e., 7 log CFU ml−1 reductions)within 384 h at 5 °C (Fig. 1B). With regards to S. Typhimurium, totalinactivation occurred within 624 h at 5 °C, when pH 4.5 was followedby exposure to 15% NaCl, whereas exposure to pH 4.5 followed byexposure to 20% NaCl allowed survival at the level of 2 log CFU ml−1
after 719 h of incubation (Fig. 2B). Application of stresses in thereverse sequence (i.e., first NaCl, then low pH) resulted in relativelysimilar reductions in bacterial populations to the pH then NaCltreatment, within the first 100–150 h, while for the remaining periodof incubation the survivors of all NaCl then pH treatments wereN1000-fold higher than those of the respective reverse sequences(Figs. 1B, 2B and 3). The observed trends for L. monocytogenes survivalin the present study are in agreement with a previous report on theeffect of acid (pH 3.5 with HCl) and salinity (14 or 20%), applied singly,sequentially or simultaneously against L. monocytogenes and E. coli at25 °C (Shabala et al., 2008; Shadbolt et al., 2001). In our study,incubation temperature (i.e., 5 and 10 °C) neither changed the trend ofinactivation of pathogens between different treatments, nor did it
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016
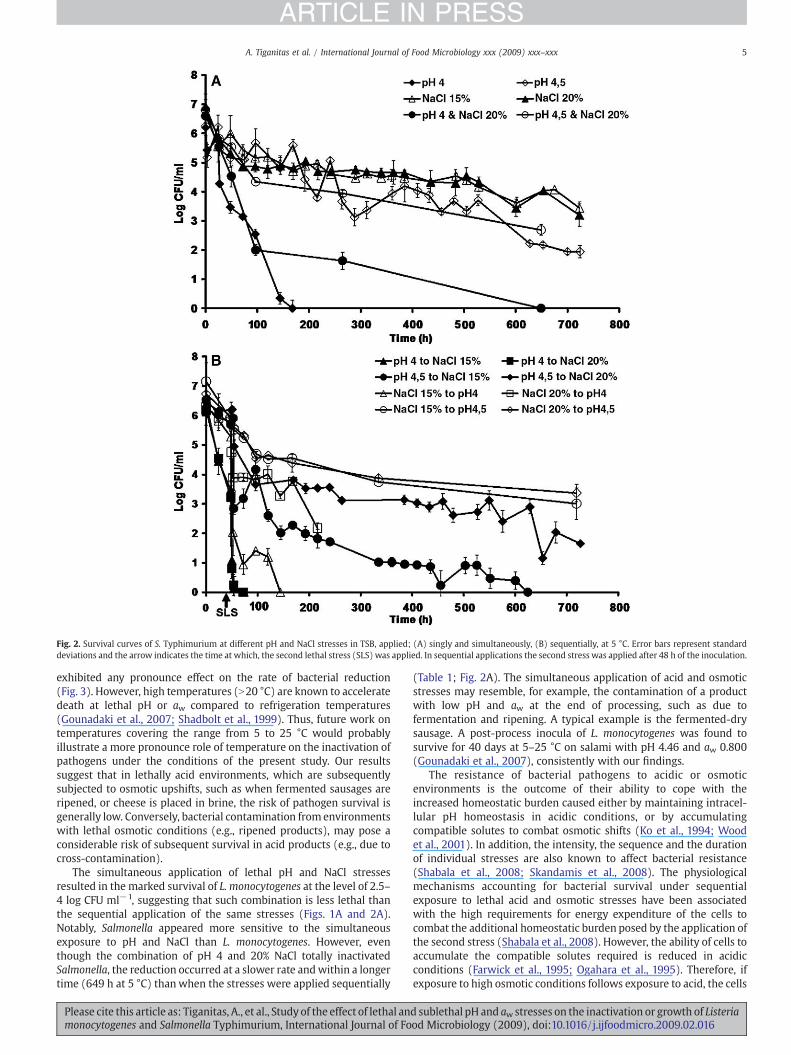
Fig. 2. Survival curves of S. Typhimurium at different pH and NaCl stresses in TSB, applied; (A) singly and simultaneously, (B) sequentially, at 5 °C. Error bars represent standarddeviations and the arrow indicates the time at which, the second lethal stress (SLS) was applied. In sequential applications the second stress was applied after 48 h of the inoculation.
5A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
exhibited any pronounce effect on the rate of bacterial reduction(Fig. 3). However, high temperatures (N20 °C) are known to acceleratedeath at lethal pH or aw compared to refrigeration temperatures(Gounadaki et al., 2007; Shadbolt et al., 1999). Thus, future work ontemperatures covering the range from 5 to 25 °C would probablyillustrate a more pronounce role of temperature on the inactivation ofpathogens under the conditions of the present study. Our resultssuggest that in lethally acid environments, which are subsequentlysubjected to osmotic upshifts, such as when fermented sausages areripened, or cheese is placed in brine, the risk of pathogen survival isgenerally low. Conversely, bacterial contamination from environmentswith lethal osmotic conditions (e.g., ripened products), may pose aconsiderable risk of subsequent survival in acid products (e.g., due tocross-contamination).
The simultaneous application of lethal pH and NaCl stressesresulted in the marked survival of L. monocytogenes at the level of 2.5–4 log CFU ml−1, suggesting that such combination is less lethal thanthe sequential application of the same stresses (Figs. 1A and 2A).Notably, Salmonella appeared more sensitive to the simultaneousexposure to pH and NaCl than L. monocytogenes. However, eventhough the combination of pH 4 and 20% NaCl totally inactivatedSalmonella, the reduction occurred at a slower rate andwithin a longertime (649 h at 5 °C) thanwhen the stresses were applied sequentially
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
(Table 1; Fig. 2A). The simultaneous application of acid and osmoticstresses may resemble, for example, the contamination of a productwith low pH and aw at the end of processing, such as due tofermentation and ripening. A typical example is the fermented-drysausage. A post-process inocula of L. monocytogenes was found tosurvive for 40 days at 5–25 °C on salami with pH 4.46 and aw 0.800(Gounadaki et al., 2007), consistently with our findings.
The resistance of bacterial pathogens to acidic or osmoticenvironments is the outcome of their ability to cope with theincreased homeostatic burden caused either by maintaining intracel-lular pH homeostasis in acidic conditions, or by accumulatingcompatible solutes to combat osmotic shifts (Ko et al., 1994; Woodet al., 2001). In addition, the intensity, the sequence and the durationof individual stresses are also known to affect bacterial resistance(Shabala et al., 2008; Skandamis et al., 2008). The physiologicalmechanisms accounting for bacterial survival under sequentialexposure to lethal acid and osmotic stresses have been associatedwith the high requirements for energy expenditure of the cells tocombat the additional homeostatic burden posed by the application ofthe second stress (Shabala et al., 2008). However, the ability of cells toaccumulate the compatible solutes required is reduced in acidicconditions (Farwick et al., 1995; Ogahara et al., 1995). Therefore, ifexposure to high osmotic conditions follows exposure to acid, the cells
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016
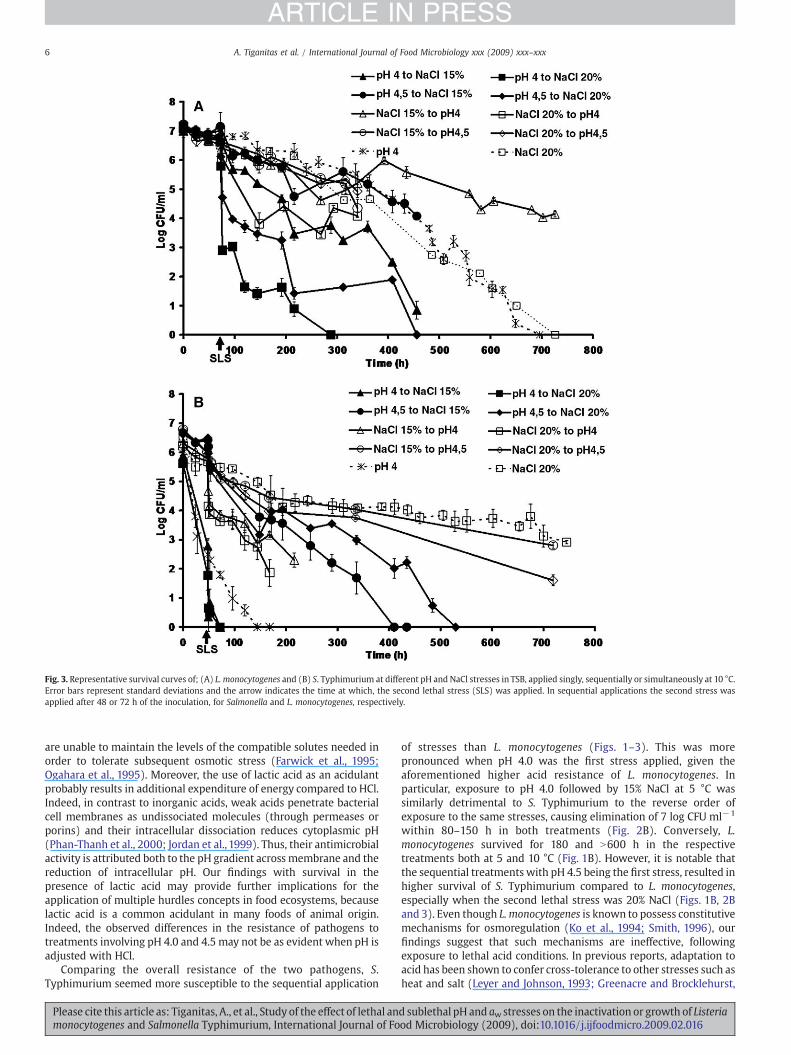
Fig. 3. Representative survival curves of; (A) L. monocytogenes and (B) S. Typhimurium at different pH and NaCl stresses in TSB, applied singly, sequentially or simultaneously at 10 °C.Error bars represent standard deviations and the arrow indicates the time at which, the second lethal stress (SLS) was applied. In sequential applications the second stress wasapplied after 48 or 72 h of the inoculation, for Salmonella and L. monocytogenes, respectively.
6 A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
are unable to maintain the levels of the compatible solutes needed inorder to tolerate subsequent osmotic stress (Farwick et al., 1995;Ogahara et al., 1995). Moreover, the use of lactic acid as an acidulantprobably results in additional expenditure of energy compared to HCl.Indeed, in contrast to inorganic acids, weak acids penetrate bacterialcell membranes as undissociated molecules (through permeases orporins) and their intracellular dissociation reduces cytoplasmic pH(Phan-Thanh et al., 2000; Jordan et al., 1999). Thus, their antimicrobialactivity is attributed both to the pH gradient across membrane and thereduction of intracellular pH. Our findings with survival in thepresence of lactic acid may provide further implications for theapplication of multiple hurdles concepts in food ecosystems, becauselactic acid is a common acidulant in many foods of animal origin.Indeed, the observed differences in the resistance of pathogens totreatments involving pH 4.0 and 4.5 may not be as evident when pH isadjusted with HCl.
Comparing the overall resistance of the two pathogens, S.Typhimurium seemed more susceptible to the sequential application
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
of stresses than L. monocytogenes (Figs. 1–3). This was morepronounced when pH 4.0 was the first stress applied, given theaforementioned higher acid resistance of L. monocytogenes. Inparticular, exposure to pH 4.0 followed by 15% NaCl at 5 °C wassimilarly detrimental to S. Typhimurium to the reverse order ofexposure to the same stresses, causing elimination of 7 log CFU ml−1
within 80–150 h in both treatments (Fig. 2B). Conversely, L.monocytogenes survived for 180 and N600 h in the respectivetreatments both at 5 and 10 °C (Fig. 1B). However, it is notable thatthe sequential treatments with pH 4.5 being the first stress, resulted inhigher survival of S. Typhimurium compared to L. monocytogenes,especially when the second lethal stress was 20% NaCl (Figs. 1B, 2Band 3). Even though L. monocytogenes is known to possess constitutivemechanisms for osmoregulation (Ko et al., 1994; Smith, 1996), ourfindings suggest that such mechanisms are ineffective, followingexposure to lethal acid conditions. In previous reports, adaptation toacid has been shown to confer cross-tolerance to other stresses such asheat and salt (Leyer and Johnson, 1993; Greenacre and Brocklehurst,
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016

Fig. 4. Effect of water activity on; (A) maximum specific growth rate, (B) lag time and (C)ho parameter of L. monocytogenes at 10 °C, pre-incubated in the following conditions of pHand aw in TSBYE at 30 °C: (⁎) aw 0.995 and pH 7.2, (△) aw 0.930 and pH 7.2, (●) aw 0.995and pH 6.0, (○) aw 0.995 and pH 5.0. Data-points represent the average of twoindependent experiments.
7A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
2006) and there is some in vitro and in vivo evidence that it may alsoincrease virulence (Gahan andHill,1999; Conte et al., 2000, 2002; Rishiet al., 2005). However, such cross-protection was rather linked to HCl(pH 5.0 or 5.8) and acetic acid (pH 5.5) adaptation, whereas previousexposure to lactic acid (pH 5.5) was shown to sensitize Salmonellaagainst subsequent salt stress (Greenacre and Brocklehurst, 2006).Probably exposure to as low pH values adjusted with lactic acid, asthose applied in the current study may have resulted in an adaptiveresponse of Salmonella to salt stresses, which was masked in previousstudies with higher adaptation pH. Therefore, environments with pHapproximately 4.5 (i.e., marginally lethal) pose a significant risk ofpathogen survival, especially of Salmonella, even if subsequent osmoticstress occurs, e.g., due to ripening. An additional concern is that saltstress confers cross-tolerance to Salmonella against subsequent heatstress, whereas it may increase the maximum temperature permittinggrowth of this organism (Fletcher and Cosnka, 1998; Wood et al.,2001). Furthermore,many of the sublethal stresses (i.e., acid, salt, heat,etc.) commonly used to control bacterial growth and/or survival infoods (i.e., fermentation and ripening), are known to induce injury tomicro-organisms (Wong andWang, 2004). Injured cells are frequentlysensitive to other environmental stresses (Kalchayananmd et al.,1992;Bogosian et al., 2000) and usually become nonculturable in commonlaboratory growthmedium (Bogosian and Bourneuf, 2001). Therefore,these cells represent a potential food safety risk since; they may havethe ability to repair under less adverse conditions and cause illness(Gnanou Besse et al., 2000). Moreover, it has been suggested that inorder to survive against a combination of stressful conditions, manyfoodborne pathogens enter a viable but nonculturable (VBNC) state(i.e., a dormant state) (Oliver et al., 1991; Wong and Wang, 2004) inwhich they maintain metabolic activity and pathogenicity but cannotbe cultured by common laboratory methods (Nystrom, 2003; Wongand Wang, 2004; del Mar Lleò et al., 2007). Thus, future work on themechanisms of the induction of injured and VBNC cells under differentfood-related stresses will provide practical information on thepotential of such cells to increase health risk.
In addition to the aforementioned results on the responses of L.monocytogenes and S. Typhimurium to lethal osmotic and acidstresses, we evaluated the growth responses of L. monocytogenes atdifferent physiological states (i.e., previous growth at high or low awand pH 5–7 at 30 °C), to osmotic stresses within the permissivegrowth range of 0.940–0.995 (neutral pH) at 10 °C. The growth ratedecreased and the lag phase increased with water activity for allinoculum preparations (Fig. 4A and B). A marked increase of lag phaseduration was observed at awb0.96 (Fig. 4B), showing a biphasicchange of lag time with increasing osmolarity. The maximum specificgrowth rate of L. monocytogenes decreased linearly with increasingosmolarity (Fig. 4A). The above changes in growth kinetics withosmotic stress posed by NaCl are consistent with previous reports(Robinson et al., 1998), suggesting that above a critical concentration,NaCl affects lag more than growth rate. This may be attributed toreduction of ‘adaptation rate’ and/or increase in the work needed(e.g., biosynthetic or homeostatic demands) to adapt to osmoticallyadverse conditions (Robinson et al., 1998; Mellfont et al., 2003). In thisrespect, the different physiological states of the inocula (due toprevious growth at pH 5.0–7.2) did not strongly affect the subsequentgrowth rates at elevated osmolarity but they did affect the lag timeand hence, the parameter ho of the Baranyi model (Fig. 4C). Theparameter ho represents the work that needs to be produced by theorganism to adapt to the new environment and is analogous to therelative lag time (i.e., the ratio of lag by generation time) (Mellfontet al., 2003). In general, ho tended to increase as a result of exposure tohigh levels of NaCl, as well as in combinationwith transition from lowprecultural to neutral growth pH (Fig. 4C). Specifically, inocula grownat aw 0.995 in TSBYE with pH 6.0, showed up to 2 or 3-fold increase ofho during subsequent growth at awb0.96 (pH 7.2), compared to theother inocula. Moreover, cultures grown in 0.93 and subjected to
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
osmotic upshifts (i.e., transferring to higher aw values), indicated aconsiderably higher ho at aw of 0.95–0.97 than that of culturespreincubated at aw 0.995 (Fig. 4C). It has been reported that the Grampositive bacteria, are not as sensitive to osmotic shifts as Gramnegative bacteria, because the former possess endogenous osmoa-daptation mechanisms, which allow them to resist to osmotic shifts(Becker et al., 1998; Gardan et al., 2003; Mellfont et al., 2003). Indeed,previous studies have demonstrated that osmotic upshifts manifestedas reductions of aw by 0.05 to 0.130 units caused a marked increase inthe adaptation work of E. coli and Salmonella, whereas they hadnegligible effect on the essential adaptation work of Staphylococcusaureus and L. monocytogenes (Mellfont et al., 2003, 2004). Further-more, stationary phase inocula such as those used in the presentstudy, are expected to be less affected by osmotic shifts thanexponential phase cells (Mellfont et al., 2004). However, experimentsin the above studies were carried out at 25 °C both at precultural andgrowth stage, in contrast to our study, where inocula were grown at30 °C and then subjected to osmotic shifts at 10 °C. Consequently, ourrelatively high and variable values of adaptationwork at awb0.960 areprimarily associated with additional lag times due to transferring atlower temperature (Delignette-Muller et al., 2005). It is notable,however, that L. monocytogenes grown at pH 5.0 or 6.0 in the presenceof 0.5% NaCl (aw 0.995) initiated subsequent growth at aw of 0.940
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016

8 A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
(pH 7.2) at 10 °C after a lag time of 38 and 55 days, respectively, incontrast to cultures grown at neutral pH, which showed no growth ataw 0.940 after 60 days (Fig. 4). Likewise, the growth rate of thebacterium pre-adapted at pH 6.0 was higher at aw 0.95 than that ofcultures previously grown at pH 7.2 (Fig. 3A). These suggest that pre-adaptation at low pH may have also enhanced osmoadaptation ofcells, thereby enabling them to initiate (faster) growth at lower wateractivity than cells previously grown at neutral pH (Skandamis et al.,2007). Overall, although osmotic shifts alone may not have anypronounced effect on the adaptation effort of L. monocytogenes(Mellfont et al., 2003), when combined with pH and temperatureshifts they may strongly increase the adaptation effort required forgrowth initiation. Such findings may also be useful in modelling L.monocytogenes growth under dynamic pH and aw conditions, byproviding implications on the additional lag time posed by shifts inthe environment. However, more research is needed to comparativelyevaluate the effect of single and combined pH, aw, and temperatureshifts on microbial growth responses, in order to make reliable kineticand stochastic considerations of microbial growth in foods.
In conclusion, the present study illustrated the responses of L.monocytogenes and S. Typhimurium to lethal osmotic and acid stressescommonly occurring during different food manufacturing stages. L.monocytogenes exhibited higher acid tolerance than osmotic tolerancewhile the opposite was observed for Salmonella. In addition, exposureto stresses sequentially resulted in faster inactivation than exposure tosingle stresses whereas; simultaneous application of pH–NaCl stresswas less lethal on both microorganisms than the sequential exposure.For both pathogens, the pH–NaCl order was more detrimental thanthe reverse sequence. These results further suggest that during foodprocessing, the sequence and the severity of the stresses appliedsignificantly affect the inactivation of L. monocytogenes and Salmo-nella. Moreover, osmotic shifts in combination with acid andtemperature shifts had a pronounced effect on the adaptation effortof L. monocytogenes to initiate growth. Overall, the present findingsmay contribute to the review of the critical limits in low pH (withlactic acid) and water activity products, considering the risk of L.monocytogenes and Salmonella survival. Further in situ research withmultiple pathogen strains, as well as focussing on the underlyingmechanisms associated with the resistance of the above organismswill assist in reducing the uncertainty of risk assessment in situationswhere pathogens undergo exposure to multiple hurdles. Moreover,exploration on the effect of environmental shifts on the lag phase andgrowth kinetics of other foodborne pathogens, such as Salmonella orE. coli is also an important subject for further research.
Acknowledgements
This work is funded by the Integrated EU Project “Improved bio-traceability of unintended microorganisms and their substances infood and feed chains” (BIOTRACER), Contract no. 036272.
References
Adams, M.R., Nicolaides, L., 1997. Review of the sensitivity of different foodbornepathogens to fermentation. Food Control 8, 227–239.
Becker, L.A., Çetin, M.S., Hutkins, R.W., Benson, A.K., 1998. Identification of the geneencoding the alternative sigma factor σB from Listeria monocytogenes and its role inosmotolerance. Journal of Bacteriology 180, 4547–4554.
Bogosian, G., Bourneuf, E.V., 2001. A matter of bacterial life and death. EMBO Reports 2,770–774.
Bogosian, G., Aardema, N.D., Bourneuf, E.V., Morris, P.J.L., O'Neil, J.P., 2000. Recovery ofhydrogen peroxide-sensitive culturable cells of Vibrio vulnificus gives the appear-ance of resuscitation from a viable but nonculturable state. Journal of Bacteriology182, 5070–5075.
Bovill, R.A., Bew, J., Baranyi, J., 2001. Measurements and predictions of growth for Lis-teria monocytogenes and Salmonella during fluctuating temperature II. Rapidlychanging temperatures. International Journal of Food Microbiology 67, 131–137.
Conte, M.P., Petrone, G., Di Biase, A.M., Ammendolia, M.G., Superti, F., Seganti, L., 2000.Acid tolerance in Listeria monocytogenes influences invasiveness of enterocyte-likecells and macrophage-like cells. Microbial Pathogenesis 29, 137–144.
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
Conte,M.P., Petrone,G., Di Biase, A.M., Longhi, C., Penta,M., Tinari, A., Superti, F., Fabozzi, G.,Visca, P., Seganti, L., 2002. Effect of acid adaptation on the fate ofListeriamonocytogenesinTHP-1humanmacrophages activatedbygamma interferon. Infection and Immunity70, 4369–4378.
Del Mar Lleò, M., Benedetti, D., Tafi, M.C., Signoretto, C., Canepari, P., 2007. Inhibition ofthe resuscitation from the viable but non-culturable state in Enterococcus faecalis.Environmental Microbiology 9, 2313–2320.
Delignette-Muller, M.L., Baty, F., Cornu, M., Bergis, H., 2005. Modelling the effect of atemperature shift on the lag phase duration of Listeria monocytogenes. InternationalJournal of Food Microbiology 100, 77–84.
Faleiro, M.L., Andrew, P.W., Powder, D., 2003. Stress response of Listeria monocytogenesisolated from cheese and other foods. International Journal of Food Microbiology84, 207–216.
Farwick, M., Siewe, R.M., Krämer, R., 1995. Glycine betaine uptake after hyperosmoticshift in Corynebacterium glutamicum. Journal of Bacteriology 177, 4690–4695.
Fletcher, S.A., Cosnka, L.N., 1998. Characterization of the induction of increasedthermotolerance by high osmolarity in Salmonella. Food Microbiology 15, 307–317.
Francois, K., Valero, A., Geeraerd, A.H., Van Impe, J.F., Debevere, J., Garcıá-Gimeno, R.M.,Zurera, G., Devlieghere, F., 2007. Effect of preincubation temperature and pH on theindividual cell lag phase of Listeria monocytogenes, cultured at refrigerationtemperatures. Food Microbiology 24, 32–43.
Gahan, C.G.M., Hill, C., 1999. The relationship between acid stress responses andvirulence in Salmonella typhimurium and Listeria monocytogenes. InternationalJournal of Food Microbiology 50, 93–100.
Gardan, R., Duché, O., Leroy-Sétrin, S., The European Listeria Genome Consortium,Labadie, J., 2003. Role of ctc from Listeria monocytogenes in osmotolerance. Appliedand Environmental Microbiology 69, 154–161.
Gnanou Besse, N., Dubois Brissonnet, F., Lafarge, V., Leclerc, V., 2000. Effect of variousenvironmental parameters on the recovery of sublethally salt-damaged and acid-damaged Listeria monocytogenes. Journal of Applied Microbiology 89, 944–950.
Gounadaki, A.S., Skandamis, P.N., Drosinos, E.H., Nychas, G-J.E., 2007. Effect of packagingand storage temperature on the survival of Listeria monocytogenes inoculated post-processing on sliced salami. Journal of Food Protection 70, 2313–2320.
Greenacre, E.J., Brocklehurst, T.F., 2006. The acetic acid tolerance response inducescross-protection to salt stress in Salmonella typhimurium. International Journal ofFood Microbiology 112, 62–65.
Guillier, L., Pardon, P., Augustin, J.C., 2005. Influence of stress on individual lag timedistributions of Listeria monocytogenes. Applied and Environmental Microbiology71, 2940–2948.
Hew, C.M., Hajmeer, M.N., Farver, T.B., Glover, J.M., Cliver, D.O., 2005. Survival of Listeriamonocytogenes in experimental chorizos. Journal of Food Protection 68, 324–330.
Jacobsen, T., Koch, A.G., 2006. Influence of different histories of the inoculum on lagphase and growth of Listeria monocytogenes in meat models. Journal of FoodProtection 69, 532–541.
Jordan, S.L., Glover, J., Malcolm, L., Thomson-Carter, F.M., Booth, I.R., Park, S.F., 1999.Augmentation of killing of Escherichia coli O157 by combinations of lactate, ethanol,and low-pH conditions. Applied and Environmental Microbiology 65, 1308–1311.
Kalchayananmd, N., Hanlin, B., Ray, B., 1992. Sublethal injury makes Gram-negative andresistant Gram-positive bacteria sensitive to the bacteriocins, pediocin AcH andnisin. Letters in Applied Microbiology 15, 239–243.
Ko, R., Smith, L.T., Smith, G.M., 1994. Glycine betaine confers enhanced osmotoleranceand cryotolerance on Listeria monocytogenes. Journal of Bacteriology 176, 426–431.
Koutsoumanis, K.P., Sofos, J.N., 2004. Comparative acid stress response of Listeriamonocytogenes, Echerichia coli O157:H7 and Salmonella Typhimurium afterhabituation at different pH conditions. Letters in AppliedMicrobiology 38, 321–326.
Koutsoumanis, K.P., Kendall, P.A., Sofos, J.N., 2003. Effect of food processing-relatedstresses on acid tolerance of Listeria monocytogenes. Applied and EnvironmentalMicrobiology 69, 7514–7516.
Lahti, E., Johansson, T., Honkanen-Buzalski, T., Hill, P., Nurmi, E., 2001. Survival and detectionof Escherichia coli 0157:H7 and Listeria monocytogenes during the manufacture of drysausage using two different starter cultures. Food Microbiology 18, 75–85.
Leistner, L., 2000. Basic aspects of food preservation by hurdle technology. InternationalJournal of Food Microbiology 55, 181–186.
Leyer, G.J., Johnson, E.A., 1993. Acid adaptation induces cross-protection againstenvironmental stresses in Salmonella typhimurium. Applied and EnvironmentalMicrobiology 59, 1842–1847.
Mellfont, L.A., McMeekin, T.A., Ross, T., 2003. The effect of abrupt osmotic shifts on thelag phase duration of foodborne bacteria. International Journal of Food Micro-biology 83, 281–293.
Mellfont, L.A., McMeekin, T.A., Ross, T., 2004. The effect of abrupt osmotic shifts on thelag phase duration of physiologically distinct populations of Salmonella typhimur-ium. International Journal of Food Microbiology 92, 111–120.
Nightingale, K.K., Thippareddi, H., Phebus, R.K., Marsden, J.L., Nutsch, A.L., 2006.Validation of a traditional Italian-style salami manufacturing process for control ofSalmonella and Listeria monocytogenes. Journal of Food Protection 69, 794–800.
Niksic, M., Niebuhr, S.E., Dickson, J.S., Mendonca, A.F., Koziczkowski, J.J., Ellingson, J.L.E.,2005. Survival of Listeria monocytogenes and Escherichia coli O157:H7 duringSauerkraut fermentation. Journal of Food Protection 68, 1367–1374.
Nissen, H., Holck, A., 1998. Survival of Escherichia coli O157:H7, Listeria monocytogenesand Salmonella kentucky in Norwegian fermented, dry sausage. Food Microbiology15, 273–279.
Nystrom, T., 2003. Nonculturable bacteria: programmed survival forms or cells atdeath's door? BioEssays 25, 204–211.
Ogahara, T., Ohno, M., Takayama, M., Igarashi, Kobayashi, H., 1995. Accumulation ofglutamate by osmotically stressed Escherichia coli is dependent on pH. Journal ofBacteriology 177, 5987–5990.
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016

9A. Tiganitas et al. / International Journal of Food Microbiology xxx (2009) xxx–xxx
ARTICLE IN PRESS
Oliver, J.D., Nilsson, L., Kjelleberg, S., 1991. Formation of nonculturable Vibrio vulnificuscells and its relationship to the starvation state. Applied and EnvironmentalMicrobiology 57, 2640–2644.
Phan-Thanh, L., Mahouin, F., Aligé, S., 2000. Acid responses of Listeria monocytogenes.International Journal of Food Microbiology 55, 121–126.
Rishi, P., Pathak, S., Ricke, S.C., 2005. Short chain fatty acids influence virulenceproperties of Salmonella Enterica Serovar Typhimurium. Journal of EnvironmentalScience and Health, Part B 40, 645–657.
Robinson, T.P., Ocio, M.J., Kaloti, A., Mackey, B.M., 1998. The effect of the growthenvironment on the lag phase of Listeria monocytogenes. International Journal ofFood Microbiology 44, 83–92.
Samelis, J., Ikeda, J.S., Sofos, J.N., 2003. Evaluation of the pH-dependent, stationary-phase acid tolerance in Listeria monocytogenes and Salmonella Typhimurium DT104induced by culturing in media with 1% glucose: a comparative study with Escher-ichia coli O157:H7. Journal of Applied Microbiology 95, 563–575.
Shabala, L., Lee, S.H., Cannesson, P., Ross, T., 2008. Acid and NaCl limits of growth ofListeria monocytogenes and influence of sequence of inimical acid and NaCl levelson inactivation kinetics. Journal of Food Protection 71, 1169–1177.
Shadbolt, C., Ross, T., McMeekin, T.A., 1999. Nonthermal death of Escherichia coli.International Journal of Food Microbiology 49, 129–138.
Shadbolt, C., Ross, T., McMeekin, T.A., 2001. Differentiation of the effects of lethal pH andwater activity: food safety implications. Letters in AppliedMicrobiology 32, 99–102.
Skandamis, P.N., Stopforth, J.D., Kendall, P.A., Belk, K.E., Scanga, J.A., Smith, G.C., Sofos, J.N.,2007. Modeling the effect of inoculum size and acid adaptation on growth/no growth
Please cite this article as: Tiganitas, A., et al., Study of the effect of lethal anmonocytogenes and Salmonella Typhimurium, International Journal of Fo
interface of Escherichia coli O157:H7. International Journal of Food Microbiology 120,237–249.
Skandamis, P.N., Yoon, Y., Stopforth, J.D., Kendall, P.A., Sofos, J.N., 2008. Heat and acidtolerance of Listeria monocytogenes after exposure to single and multiple sublethalstresses. Food Microbiology 25, 294–303.
Smith, L.T., 1996. Role of osmolytes in adaptation of osmotically stressed and chill-stressed Listeria monocytogenes grown in liquid media and on processed meatsurfaces. Applied and Environmental Microbiology 62, 3088–3093.
Smith, J.L., Huhtanen, C.N., Kissinger, J.C., Palumbo, S.A., 1975. Survival of Salmonellaeduring pepperoni manufacture. Applied Microbiology 30, 759–763.
Thévenot, D., Delignette-Muller, M.L., Christieans, S., Vernozy-Rozand, C., 2005. Fate ofListeria monocytogenes in experimentally contaminated French sausages. Interna-tional Journal of Food Microbiology 101, 189–200.
Wong, H.C., Wang, P., 2004. Induction of viable but nonculturable state in Vibrioparahaemolyticus and its susceptibility to environmental stresses. Journal ofApplied Microbiology 96, 359–366.
Wood, J.M., Bremer, E., Csonka, L.N., Kraemer, R., Poolman, B., van der Heide, T., Smith, L.T.,2001. Osmosensing and osmoregulatory compatible solute accumulation by bacteria.Comparative Biochemistry and Physiology Part A 130, 437–460.
Yoon, Y., Skandamis, P.N., Kendall, P.A., Smith, G.C., Sofos, J.N., 2006. A predictive modelfor the effect of temperature and predrying treatments in reducing Listeriamonocytogenes populations during drying of beef jerky. Journal of Food Protection69, 62–70.
d sublethal pH and aw stresses on the inactivation or growth of Listeriaod Microbiology (2009), doi:10.1016/j.ijfoodmicro.2009.02.016
Related Documents











