Pharmacological Research 60 (2009) 382–391 Contents lists available at ScienceDirect Pharmacological Research journal homepage: www.elsevier.com/locate/yphrs Studies on the protective effect of green tea against cisplatin induced nephrotoxicity Sara A. Khan, Shubha Priyamvada, Wasim Khan, Sheeba Khan, Neelam Farooq, Ahad N.K. Yusufi ∗ Department of Biochemistry, Faculty of Life Sciences, Aligarh Muslim University, Aligarh 202002, U.P., India article info Article history: Received 6 March 2009 Received in revised form 21 July 2009 Accepted 21 July 2009 Keywords: Cisplatin Nephrotoxicity Oxidative damage Pi transport Carbohydrate metabolism Green tea abstract Cisplatin (CP) an anticancer drug is known to induce nephrotoxicity, which limits its long-term clinical use. Green tea (GT), consumed since ancient times is known for its numerous health benefits. It has been shown to improve kidney functions in animal models of acute renal failure. The present study was under- taken to see whether GT can prevent CP-induced nephrotoxic and other deleterious effects. A nephrotoxic dose of CP was co-administered to control and GT-fed male Wistar rats every fifth day for 25 days. The effect of GT was determined on CP-induced alterations in various serum parameters and on enzymes of carbohydrate metabolism, brush border membrane, and antioxidant defense system in renal cortex and medulla. CP nephrotoxicity was recorded by increased serum creatinine and blood urea nitrogen. CP increased the activities of lactate dehydrogenase and acid phosphatase whereas, the activities of malate dehydrogenase, glucose-6-phosphatase, superoxide dismutase, catalase, and 32 Pi transport significantly decreased. GT consumption increased the activities of the enzymes of carbohydrate metabolism, brush border membrane, oxidative stress, and 32 Pi transport. GT ameliorated CP-induced nephrotoxic and other deleterious effects due to its intrinsic biochemical/antioxidant properties. © 2009 Elsevier Ltd. All rights reserved. 1. Introduction Cisplatin (cis-diamminedichloroplatinum II, CP) is one of the most potent antitumor agent and is a highly effective agent against a diverse spectrum of malignancies [1]. However, the use of this agent in combating cancer is limited by the development of nephro- and ototoxicity [2,3], along with various untoward side effects including nausea, vomiting, diarrhea, and myelosuppression [4,5]. Light and electron microscopy have shown that the CP-induced injury and necrosis in the rat kidney are predominantly localized in S3 subsegments of proximal tubular epithelial cells [6]. Mor- phologically, it is characterized by the loss of microvilli, cellular swelling, and condensation of nuclear chromatin [7]. Mitochondria, lysosomes, and microsomes are critical CP targets [8,9]. Functional alterations are characterized by change in urine volume, increase in blood urea nitrogen, and serum creatinine [10]. There is a continuous search for agents that provide nephro- protection against CP and other platinum drugs [3]. These include antioxidants, modulators of nitric oxide, diuretics, and cytoprotec- tive and apoptotic agents [3]. However, none of these were found to be suitable/safe for clinical use in protecting against CP-induced nephrotoxicity. ∗ Corresponding author. Tel.: +91 571 2700741; fax: +91 571 2706002. E-mail address: yusufi@lycos.com (A.N.K. Yusufi). In the past few years, much interest has been centered on the role of naturally occurring dietary substances for the con- trol and management of various chronic diseases [11,12]. Since ancient times, green tea (GT) consumption has been known to pos- sess health-promoting potential [13]. It has been shown to have cardioprotective, neuroprotective, antidiabetic, and antimicrobial properties [14–16]. GT is an excellent source of polyphenols, par- ticularly of a group known as GT catechins [17,18]. In particular, GT catechins and their derivatives have been characterized as effective antioxidants that scavenge free radicals and demonstrate antimu- tagenic, and anticarcinogenic properties [19,20]. GT has been shown to improve kidney function in diabetic rats [21]. It has been demonstrated to ameliorate cyclosporine A and CP-induced renal dysfunction in experimental rats [22,23] and in LLC-PK1 (Porcine kidney cells, having similar properties to those of renal proximal tubules) cells [24]. GT has been shown to miti- gate CP-induced nephrotoxicity parameters [25]. Recently, we have demonstrated that GT protected against gentamicin (GM) nephro- toxicity by improving antioxidant defense, tissue integrity, energy metabolism, and renal Pi transport [27]. Taking into consideration the potential clinical use of CP and the numerous health benefits of GT, the present work was undertaken to study the detailed mechanisms of CP-induced nephropathy and its protection by GT consumption. We hypothesized that GT would prevent CP-induced nephrotoxicity due to its intrinsic biochemical and antioxidant properties [26]. The present results demonstrate marked reversal of CP-induced nephrotoxicity parameters in serum 1043-6618/$ – see front matter © 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.phrs.2009.07.007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Sn
SD
a
ARRA
KCNOPCG
1
maaai
iipslai
pattn
1d
Pharmacological Research 60 (2009) 382–391
Contents lists available at ScienceDirect
Pharmacological Research
journa l homepage: www.e lsev ier .com/ locate /yphrs
tudies on the protective effect of green tea against cisplatin inducedephrotoxicity
ara A. Khan, Shubha Priyamvada, Wasim Khan, Sheeba Khan, Neelam Farooq, Ahad N.K. Yusufi ∗
epartment of Biochemistry, Faculty of Life Sciences, Aligarh Muslim University, Aligarh 202002, U.P., India
r t i c l e i n f o
rticle history:eceived 6 March 2009eceived in revised form 21 July 2009ccepted 21 July 2009
eywords:isplatin
a b s t r a c t
Cisplatin (CP) an anticancer drug is known to induce nephrotoxicity, which limits its long-term clinicaluse. Green tea (GT), consumed since ancient times is known for its numerous health benefits. It has beenshown to improve kidney functions in animal models of acute renal failure. The present study was under-taken to see whether GT can prevent CP-induced nephrotoxic and other deleterious effects. A nephrotoxicdose of CP was co-administered to control and GT-fed male Wistar rats every fifth day for 25 days. Theeffect of GT was determined on CP-induced alterations in various serum parameters and on enzymes
ephrotoxicityxidative damagei transportarbohydrate metabolismreen tea
of carbohydrate metabolism, brush border membrane, and antioxidant defense system in renal cortexand medulla. CP nephrotoxicity was recorded by increased serum creatinine and blood urea nitrogen. CPincreased the activities of lactate dehydrogenase and acid phosphatase whereas, the activities of malatedehydrogenase, glucose-6-phosphatase, superoxide dismutase, catalase, and 32Pi transport significantlydecreased. GT consumption increased the activities of the enzymes of carbohydrate metabolism, brushborder membrane, oxidative stress, and 32Pi transport. GT ameliorated CP-induced nephrotoxic and other
its in
deleterious effects due to. Introduction
Cisplatin (cis-diamminedichloroplatinum II, CP) is one of theost potent antitumor agent and is a highly effective agent againstdiverse spectrum of malignancies [1]. However, the use of this
gent in combating cancer is limited by the development of nephro-nd ototoxicity [2,3], along with various untoward side effectsncluding nausea, vomiting, diarrhea, and myelosuppression [4,5].
Light and electron microscopy have shown that the CP-inducednjury and necrosis in the rat kidney are predominantly localizedn S3 subsegments of proximal tubular epithelial cells [6]. Mor-hologically, it is characterized by the loss of microvilli, cellularwelling, and condensation of nuclear chromatin [7]. Mitochondria,ysosomes, and microsomes are critical CP targets [8,9]. Functionallterations are characterized by change in urine volume, increasen blood urea nitrogen, and serum creatinine [10].
There is a continuous search for agents that provide nephro-rotection against CP and other platinum drugs [3]. These include
ntioxidants, modulators of nitric oxide, diuretics, and cytoprotec-ive and apoptotic agents [3]. However, none of these were foundo be suitable/safe for clinical use in protecting against CP-inducedephrotoxicity.∗ Corresponding author. Tel.: +91 571 2700741; fax: +91 571 2706002.E-mail address: [email protected] (A.N.K. Yusufi).
043-6618/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.oi:10.1016/j.phrs.2009.07.007
trinsic biochemical/antioxidant properties.© 2009 Elsevier Ltd. All rights reserved.
In the past few years, much interest has been centered onthe role of naturally occurring dietary substances for the con-trol and management of various chronic diseases [11,12]. Sinceancient times, green tea (GT) consumption has been known to pos-sess health-promoting potential [13]. It has been shown to havecardioprotective, neuroprotective, antidiabetic, and antimicrobialproperties [14–16]. GT is an excellent source of polyphenols, par-ticularly of a group known as GT catechins [17,18]. In particular, GTcatechins and their derivatives have been characterized as effectiveantioxidants that scavenge free radicals and demonstrate antimu-tagenic, and anticarcinogenic properties [19,20].
GT has been shown to improve kidney function in diabetic rats[21]. It has been demonstrated to ameliorate cyclosporine A andCP-induced renal dysfunction in experimental rats [22,23] and inLLC-PK1 (Porcine kidney cells, having similar properties to thoseof renal proximal tubules) cells [24]. GT has been shown to miti-gate CP-induced nephrotoxicity parameters [25]. Recently, we havedemonstrated that GT protected against gentamicin (GM) nephro-toxicity by improving antioxidant defense, tissue integrity, energymetabolism, and renal Pi transport [27].
Taking into consideration the potential clinical use of CP and thenumerous health benefits of GT, the present work was undertaken
to study the detailed mechanisms of CP-induced nephropathy andits protection by GT consumption. We hypothesized that GT wouldprevent CP-induced nephrotoxicity due to its intrinsic biochemicaland antioxidant properties [26]. The present results demonstratemarked reversal of CP-induced nephrotoxicity parameters in serum
gical R
abrtaan
2
2
cCocR
2
5cet(l(ci9
2
loaiCw(
Fs
S.A. Khan et al. / Pharmacolo
nd alterations in the enzymes of carbohydrate metabolism, brushorder membrane (BBM), and antioxidant system. The presentesults suggest that GT consumption can be an option for the long-erm clinical use of CP as an anticancer drug without nephrotoxicnd other harmful side effects. The results support a potential ther-peutic use of GT + CP combination in combating cancer withoutephrotoxic and other harmful side effects.
. Materials and methods
.1. Chemicals and drugs
Green tea (Lipton/Kangra-brand) was purchased from commer-ial sources (Jain Pan House, New Delhi, India), Cisplatin (Sigmahemical Co., USA) from the source indicated in parentheses. Allther chemicals used were of analytical grade and were pur-hased either from Sigma Chemical Co. (St Louis, MO, USA) or Siscoesearch Laboratory, Mumbai, India.
.2. Green tea extract
Green tea extract (GTE) was prepared by adding GT (30 g) to00 mL of boiling water, steeped for 15–20 min. The infusion wasooled to room temperature and then filtered. The tea leaves werextracted a second time with 500 mL of boiling water and fil-ered, and the two filtrates were combined to obtain a 3% GTE3 g tea leaves/100 mL H2O). The resulting clear solution is simi-ar to tea brews consumed by humans. A known amount of GTE2 mL × 125 mL = 250 mL/day) was provided in two servings to GTonsuming rats for 25 days and was found to be sufficient. Accord-ng to the manufacturer’s information, the antioxidant content was5 mg/g of GT.
.3. Experimental design
The animal experiments were conducted according to the guide-ines of the Committee for Purpose of Control and Supervisionf Experiments on Animals (CPCSEA), Ministry of Environment
nd Forests, Government of India. Adult male Wistar rats weigh-ng 150–180 g fed with standard rat chow (Aashirwaad Industries,handigarh, India) and water ad libitum were conditioned for 1eek before the start of the experiment. Initially 2 groups of rats16 rats/group) entered the study after acclimatization (Fig. 1).
ig. 1. Experimental design. ND: normal diet, GT: green tea, CP: cisplatin, TCP: green tea +aline injection (↑).
esearch 60 (2009) 382–391 383
They were fed on a normal rat chow diet with one group con-suming water (control) and the other consuming GT extract (3%,w/v) in drinking water as described earlier [26]. After 5 days, therats were subdivided into two groups each (eight rats/group) andcontinued to receive normal rat diet and their respective drink-ing fluids. Nephrotoxicity was induced by administration of CP(3 mg/kg bwt/day), in 0.9% saline intraperitoneally every fifth day(four injections in total) for 25 days to one of the sub-groupdesignated as CP and TCP (green tea + cisplatin). The other sub-group (water-drinking) from each group received an equivalentvolume of normal saline for the same period. The rats were sac-rificed on the fifth day after the last injection under light etheranesthesia.
Blood samples were collected and the kidneys were removedand processed for the preparation of homogenates and brushborder membrane vesicles (BBMVs) as described below. All thepreparations and analyses of various parameters were performedsimultaneously under similar experimental conditions to avoid anyday-to-day variations. Body weights of rats were recorded at thestart and completion of the experimental procedure.
2.4. Preparation of homogenates
After the completion of the experiment, the kidneys wereremoved, decapsulated, and kept in ice-cold buffered saline(154 mM NaCl, 5 mM Tris–HEPES, pH 7.5). The cortex was care-fully separated from medulla as described earlier [28]. A 15% (w/v)homogenate was prepared in 0.1 M Tris–HCl buffer, pH 7.5 usingPotter-Elvehejem homogenizer (Remi motors, Mumbai, India); bypassing five pulses. The homogenate was centrifuged at 3000 × gat 4 ◦C for 15 min to remove cell debris and the supernatant wassaved in aliquots and stored at −20 ◦C for assaying the enzymes ofcarbohydrate metabolism, free radical scavenging enzymes and forthe estimation of total-SH and lipid peroxidation.
2.5. Preparation of brush border membrane
BBMVs were prepared from whole cortex using the MgCl2 pre-
cipitation method as described previously [29]. Briefly, freshlyminced cortical slices were homogenized in 50 mM mannitol and5 mM Tris–HEPES buffer pH 7.0 (20 mL/g), in a glass Teflon homog-enizer with four complete strokes. The homogenate was thensubjected to high speed Ultra-Turrex homogenizer (Type T-25,cisplatin treated, i.p.: intraperitoneal. Green tea (. . . .); water (—); CP injection ( );
384 S.A. Khan et al. / Pharmacological R
Table 1Effect of green tea (GT) consumption on body weight (grams) of rats with/withoutcisplatin (CP) treatment.
Groups Before treatment After treatment % Change
Control 164.3 ± 7.4 162.5 ± 12.5 –CP 165.6 ± 6.6 147.5 ± 8.3 −11GT 164.3 ± 7.4 143.8 ± 7.5 −12.5
R
Jettaispp(1poaff
2
ifiqcpSm[
2
oCadEd(mam
TE
R
TCP 168.8 ± 7.8 141.3 ± 5.9* −16.3
esults are expressed as mean ± SEM.* Significantly different at p < 0.05 from control by one-way ANOVA.
anke & Kunkel GMBH & Co., K.G. Staufen) for three strokes of 15 sach with a 15 s interval between each stroke. MgCl2 was added tohe homogenate to a final concentration of 10 mM, and the mix-ure stirred for 20 min on ice. The homogenate was centrifugedt 2000 × g for 10 min in a Beckman centrifuge (J2 MI, Beckmannstruments Inc Palo Alto, C.A. USA) using JA-17 rotor, and theupernatant was then recentrifuged at 35,000 × g for 30 min. Theellet was resuspended in 300 mM mannitol and 5 mM Tris–HEPES,H 7.4, with four passes by a loose fitting Dounce homogenizerWheaton IL, USA) and centrifuged at 35,000 × g for 20 min in a5 mL cortex tube. The outer white fluffy pellet of BBM was resus-ended in a small volume of buffered 300 mM mannitol. Aliquotsf homogenates and BBM were saved and stored at −20 ◦C for thenalyses of BBM enzymes. Freshly prepared BBM vesicles were usedor Pi transport. Each BBM sample was prepared by pooling tissuesrom 2 to 3 rats.
.6. Serum parameters
Serum samples were deproteinated with 3% trichloroacetic acidn a ratio of 1:3 left for 10 min and then centrifuged at 2000 × gor 10 min. The protein free supernatant was used to determinenorganic phosphate and creatinine. The precipitate was used touantitate total phospholipids. Blood urea nitrogen (BUN) andholesterol concentrations were determined directly in serum sam-les. Glucose was estimated by o-toluidene method using a kit frompan diagnostics, Mumbai, India. These parameters were deter-ined by standard procedures as mentioned in a previous study
27,28].
.7. Assay of carbohydrate metabolism enzymes
The activities of enzymes involved in the oxidation of NADHr reduction of NADP were determined spectrophotometrically onintra 5 fixed for 340 nm using 3 mL of assay in a 1-cm cuvettet room temperature (28–30 ◦C). The enzyme assays of lactateehydrogenase (LDH, E.C.1.1.1.27), malate dehydrogenase (MDH,.C.1.1.1.37), malic enzyme (ME, E.C.1.1.1.40), glucose-6-phosphate
ehydrogenase (G6PDH, E.C.1.1.1.49), and glucose-6-phosphataseG6Pase, E.C.3.1.3.3) activities were studied as described by Khund-iri et al. [30]. Hexokinase was estimated by the method of Cranend Sols [31] and the remaining glucose was measured by theethod of Nelson–Somogyi [32].
able 2ffect of green tea (GT) consumption on several serum parameters with/without cisplatin
Groups Creatinine (mg/dL) BUN (mg/dL) Glucose (mg/dL)
Control 1.07 ± 0.4 22.60 ± 1.89 32.9 ± 1.6CP 1.48 ± 1.11* (+38%) 33.12 ± 0.4* (+47%) 23.8 ± 2.86* (−28%)GT 0.96 ± 0.33 (−10%) 12.6 ± 0.23* (−44%) 26.3 ± 2.47 (−20%)TCP 1.07 ± 0.28 19.1 ± 0.48† (−16%) 20.1 ± 1.25* (−39%)
esults are mean ± SEM of six different samples. Values in parentheses represent percent* Significantly different from control.† Significantly different from CP at p < 0.05 by one-way ANOVA.
esearch 60 (2009) 382–391
2.8. Assay of brush border membrane marker enzymes andlysosomal marker enzymes
The activities of alkaline phosphatase (ALP), leucine aminopeptidase (LAP), �-glutamyl transferase (GGTase), and acid phos-phatase (ACPase) were determined as described by Khundmiri etal. [33].
2.9. Assay of enzymes involved in free radical scavenging
Superoxide dismutase (SOD, E.C.1.15.1.1) was assayed by themethod of Marklund [34]. Catalase (CAT, E.C.1.11.1. 6) activity wasassayed by the method of Giri et al. [35].
2.10. Lipid peroxidation and total–SH group estimation
Total-SH groups were determined by the method of Sedlak andLindsay [36]. Lipid peroxidation (LPO) by the method of Ohkawa etal. [37].
2.11. Transport of 32Pi
Measurement of 32Pi (Bhabha Atomic Research Center, India)uptake in BBMV(s) was performed at 25 ◦C by rapid filtration tech-nique as described by Ref. [38] either in the presence or absenceof Na-gradient. Uptake was initiated by the addition of 30 �L ofthe incubation medium containing 100 mmol/L mannitol, NaCl/KCl100 mmol/L, 5 mmol/L Tris–HEPES, pH 7.5, 0.1 mmol/L K2H32PO4 to15 �L BBM suspension (50–100 �g protein) and incubated for thedesired time intervals (see Section 3). The uptake was stopped bythe addition of 3 mL ice-cold stop solution (containing 135 mmol/LNaCl, 5 mmol/L Tris–HEPES and 10 mM sodium arsenate, pH 7.5)and filtered immediately through 0.45 �m DAWP Millipore (USA)and washed three times with the stop solution using a Cornwalltype syringe (Wheaton IL). Correction for non-specific binding tofilters was made by subtracting from all data the value of cor-responding blank obtained by filtration of the incubation bufferwithout vesicles. The radioactivity of dried filters was measured byliquid scintillation counting (Reckbeta, LKB, Wallac, Sweden) with10 mL scintillation fluid (Cocktail T, SRL, India).
2.12. Statistical analysis
All data are expressed as mean ± SEM for at least 4–5 differ-ent preparations. Statistical evaluation was conducted by one-wayANOVA and by unpaired Student’s t-test using SPSS 7.5 software.
A probability level of p < 0.05 was selected as indicating statisti-cal significance. Most of the changes between various groups werecompared with control values for better understanding and clarity.However, specific differences and statistical significance betweenother groups were evaluated separately, e.g., CP vs. TCP.(CP) treatment.
Cholesterol (mg/dL) Phosphate (�mol/mL) Phospholipid (�g/mL)
119.88 ± 1.3 3.72 ± 0.26 97.2 ± 8.8146.24 ± 3.8* (+22%) 2.95 ± 0.25 (−21%) 132.05 ± 40.05* (+36%)
98.4 ± 0.6 (−18%) 2.6 ± 0.35* (−30%) 142.15 ± 2.35* (+46%)102.12 ± 1.7† (−15%) 2.5 ± 0.2* (−33%) 149 ± 3.3* (+53%)
change from control.
S.A. Khan et al. / Pharmacological Research 60 (2009) 382–391 385
Table 3Effect of green tea (GT) consumption on brush border membrane enzymes in cortical and medullary homogenates with/without cisplatin (CP) treatment.
Tissue ALP (�mol/mg protein/h) GGTase (�mol/mg protein/h) LAP (�mol/mg protein/h) ACPase (�mol/mg protein/h)
CortexControl 2.4 ± 0.01 40.54 ± 0.6 5.42 ± 0.13 4.28 ± 0.32CP 1.84 ± 0.4* (−23%) 25.33 ± 0.89* (−38%) 2.89 ± 0.25* (−47%) 6.13 ± 0.2* (+43%)GT 3.02 ± 0.28* (+26%) 32.4 ± 0.51* (−20%) 3.54 ± 0.08* (−35%) 5.82 ± 0.2* (+36%)TCP 3.20 ± 0.2*,† (+33%) 28.56 ± 1.96* (−30%) 3.23 ± 0.24* (−40%) 6.31 ± 0.25 (+47%)
MedullaTCP 2.0 ± 0.05 7.5 ± 0.16 5.22 ± 0.26 2.25 ± 0.11TCP 1.4 ± 0.1* (−30%) 6.11 ± 0.13* (−19%) 2.3 ± 0.2* (−56%) 2.65 ± 0.17 (+18%)TCP 2.5 ± 0.05 (+25%) 9.3 ± 0.8 (+24%) 3.55 ± 0.1* (−32%) 3.0 ± 0.14* (+33%)TCP 2.45 ± 0.07*,† (+23%) 8.1 ± 0.45† (NS) 2.77 ± 0.09* (−44%) 3.2 ± 0.11* (+42%)
R ent pe
3
p
esults are mean ± SEM of 3–4 different preparations. Values in parentheses repres* Significantly different from control.† Significantly different at p < 0.05 from CP by one-way ANOVA.
. Results
The effect of GT consumption and CP was determined on variousarameters in rat serum and kidney. In general, the rats remained
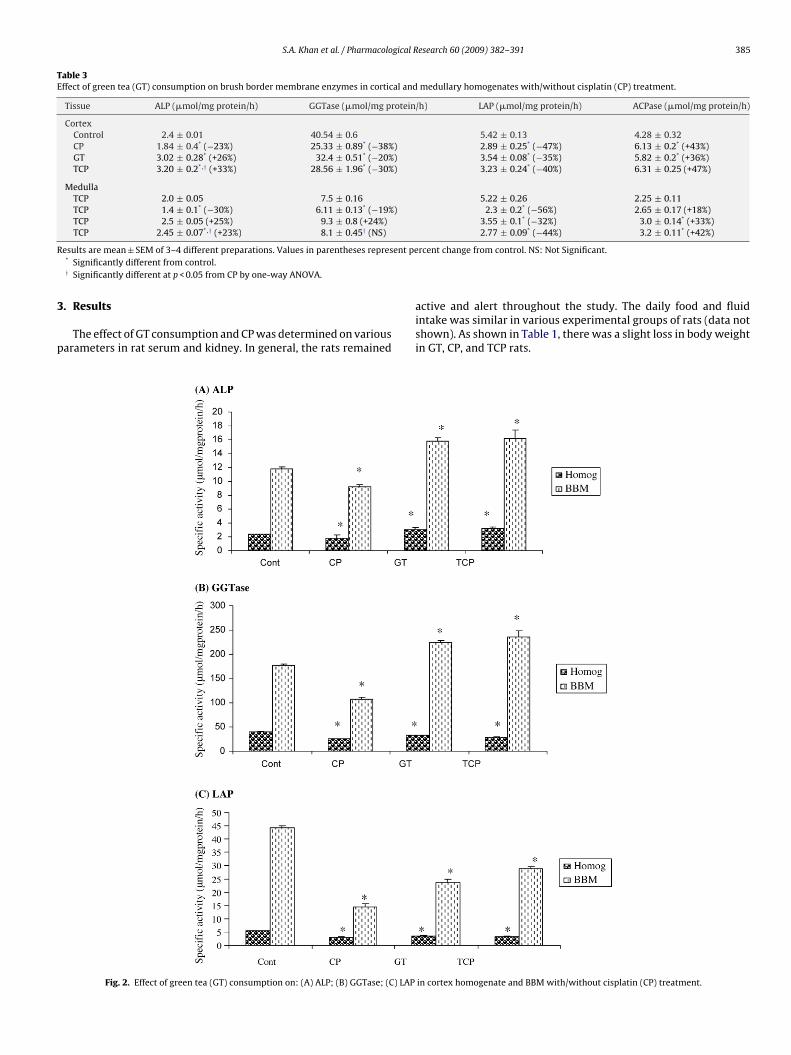
Fig. 2. Effect of green tea (GT) consumption on: (A) ALP; (B) GGTase; (C) LAP
rcent change from control. NS: Not Significant.
active and alert throughout the study. The daily food and fluidintake was similar in various experimental groups of rats (data notshown). As shown in Table 1, there was a slight loss in body weightin GT, CP, and TCP rats.
in cortex homogenate and BBM with/without cisplatin (CP) treatment.

386 S.A. Khan et al. / Pharmacological Research 60 (2009) 382–391
Table 4Effect of green tea (GT) consumption on brush border membrane enzymes in BBMV isolated from renal cortex with/without cisplatin (CP) treatment.
Groups ALP (�mol/mg protein/h) GGTase (�mol/mg protein/h) LAP (�mol/mg protein/h)
Control 11.85 ± 0.24 177.4 ± 2.7 44.4 ± 0.64CP 9.2 ± 0.28* (−22%) 107.1 ± 4.28* (−40%) 14.35 ± 1.27* (−68%)GT 15.8 ± 0 .44* (+33%) 224.1 ± 5.1* (+26%) 23.6 ± 1.33* (−47%)TCP 16.2 ± 1.2*,† (+37%) 236.4 ± 11.8*,† (+33%) 28.9 ± 0.7* (−35%)
R ent pe
3s
i(iGscWaaCHpt
3o
Bmmlaolfd
TE
R
esults are mean ± SEM of 3–4 different preparations. Values in parentheses repres* Significantly different from control.† Significantly different from CP at p < 0.05 by one-way ANOVA.
.1. Effect of GT on CP-induced nephrotoxicity parameters inerum
The administration of CP to control rats caused significantncrease in serum creatinine (+38%), blood urea nitrogen (BUN)+47%), cholesterol (+22%) and phospholipids (+36%) but decreasen serum glucose (−28%) and inorganic phosphate (−21%) (Table 2).T alone had no effect on serum creatinine whereas, BUN con-picuously decreased. GT significantly decreased serum glucose,holesterol, and Pi whereas, phospholipids significantly increased.
hen GT was given to CP rats, CP elicited nephrotoxic alterationsppeared to be ameliorated. CP-induced increase in serum cre-tinine and BUN was prevented by prolonged GT consumption.P-induced increase in serum cholesterol was also lowered by GT.owever, serum glucose and Pi remained depressed whereas, phos-holipids further increased by GT consumption to CP rats comparedo control rats.
.2. Effect of GT on CP-induced alterations on biomarker enzymesf BBM and lysosomes
To assess the structural integrity of various organelles, e.g.,BM and lysosomes, the effect of CP and GT was determined onarker enzymes of BBM in the homogenates of renal cortex andedulla as well as in isolated renal BBMV preparations. Alka-
ine phosphatase (ALP), �-glutamyl transferase (GGTase), leucine
minopeptidase (LAP) are structural and functional componentsf BBM whereas, acid phosphatase (ACPase) is a biomarker forysosomes. The activities of these enzymes demonstrate theunctional status of these organelles under pathophysiologic con-itions.able 5ffect of green tea (GT) consumption on uptake of 32Pi in brush border membrane vesicle
Groups 30 s
(A) Na+-gradient dependent uptake (Nao > Nai) (pmol/mg protein)Control 2246.7 ± 94.8CP 896.2 ± 38.2* (−60%)GT 3024.3 ± 161.5* (+35%)TCP 2507.9 ± 154.4* (+12%)
(B) K+-gradient dependent uptake (Ko > Ki) (pmol/mg protein)Control 152.8 ± 7.5CP 147.8 ± 5.3 (NS)GT 171 ± 6.9 (+12%)TCP 160.7 ± 12.2 (NS)
Groups Na+-gradient dependent uptake
(C) Net Na+-gradient dependent uptake at 30 sControl 2246.7 ± 94.8CP 896.24 ± 38.2*
GT 3024.3 ± 161.5*
TCP 2507.9 ± 154.4*
esults are mean ± SEM for five different preparations. Values in parentheses represent p* Significantly different from control.
rcent change from control.
3.2.1. Effect of CP and GT on biomarkers of BBM and lysosomes inthe homogenates of renal tissues
The activities of ALP, GGTase, LAP, and ACPase were determinedunder different experimental conditions in the homogenates ofrenal tissues (Table 3 and Fig. 2). CP caused a profound decrease inthe activities of ALP, GGTase, and LAP in a similar manner in bothrenal cortex and medulla. However, ACPase activity was markedlyincreased in both renal tissues but to a greater extent in the cor-tex (+43%) than in medulla (+18%). The effect of GT on BBM andlysosomal enzymes was differentially observed in renal cortex andmedulla. The activity of ALP and ACPase significantly increased tovarying magnitudes in cortex whereas, the activity of GGTase andLAP significantly decreased. In contrast to cortex, the activities ofALP, GGTase, and ACPase significantly increased but that of LAPdecreased in the medulla.
The co-administration of GT to CP-treated rats caused a signifi-cant reversal in CP-induced alterations in the activities of BBM andlysosomal enzymes in renal tissues (Table 3 and Fig. 2). Similarly,CP-induced decrease in BBM enzyme activities in both renal cortexand medulla was increased whereas the activity of ACPase was alsoincreased in both renal cortex and medulla by GT consumption.
3.2.2. Effect of GT on CP-induced alterations on brush bordermembrane enzymes in isolated BBM preparations
The effect of CP, GT, and their combination was further ana-lyzed on the specific activities of BBM marker enzymes in BBMV
isolated from renal cortex. The results summarized in Table 4, indi-cated that membrane preparations were several fold purified asreflected by the significant enrichment of their marker enzymes inthe BBM compared to the homogenate (4–10-fold). The data sum-marized in Table 4 (Fig. 2), showed a similar activity pattern ofs (BBMV) from renal cortex with/without cisplatin (CP) treatment.
120 min
395.9 ± 16.4347.3 ± 8.97 (−12%)459.8 ± 28. (+16%)440.2 ± 19.4 (+11%)
393.6 ± 7.4357.8 ± 13.9 (NS)429.3 ± 4.8 (NS)
415 ± 13.2 (NS)
K+-gradient dependent uptake Net uptake (Na+–K+)
152.8 ± 7.5 2094.0147.8 ± 5.3 748.4 (−64%)
171 ± 6.9 2853.3 (+36%)160.7 ± 12.2 2347.2 (+12%)
ercent change from control. NS: Not Significant.
S.A. Khan et al. / Pharmacological Research 60 (2009) 382–391 387
Table 6Effect of green tea (GT) consumption on the activity of metabolic enzymes in cortical and medullary homogenates with/without cisplatin (CP) treatment.
Tissue Hexokinase (�mol/mg protein/h) LDH (�mol/mg protein/h) MDH (�mol/mg protein/h)
CortexControl 123.7 ± 1.6 13.77 ± 0.3 33.05 ± 1.57CP 130.41 ± 2.8 (NS) 23.4 ± 6.91* (+70%) 25.5 ± 1.77* (–23%)GT 106.2 ± 1.7* (−14%) 24.6 ± 0.99* (+79%) 45.2 ± 3.25* (+37%)TCP 118.7 ± 1.3 (NS) 26.0 ± 1.06* (+89%) 52.13 ± 3.99*,† (+58%)
MedullaControl 99.87 ± 2.9 23.14 ± 0.8 41.4 ± 3.3CP 136.01 ± 5.67* (+36%) 29.4 ± 0.89* (+27%) 30.3 ± 1.35* (−27%)GT 130.6 ± 4.43* (+31%) 28.85 ± 1.75* (+25%) 35.7 ± 0.32 (−14%)TCP 141.8 ± 0.5* (+42%) 30.58 ± 0.98* (+32%) 40.7 ± 1.55† (NS)
R ent pe
Bhcahtdpttea
3Nr
ttitfoiBtdeNAw3
c
TE
R
esults are mean ± SEM of 3–4 different preparations. Values in parentheses repres* Significantly different from control.† Significantly different from CP at p < 0.05 by one-way ANOVA.
BM enzymes in BBM preparations as observed in homogenates;owever, the magnitude of effects under different experimentalonditions became more apparent. The increase or decrease in thectivities of ALP, GGTase, and LAP by CP or GT was significantlyigher in BBMVs as compared to homogenates from renal cor-ex. The damaging effect of CP was clearly ameliorated by GT, asecrease in the activity of GGTase and LAP by CP was not onlyrevented but sometimes increased significantly by GT consump-ion as compared to respective control values. Thus, it appears fromhe data that co-administration of GT to CP-treated rats to a largerxtent reduced CP-induced structural alterations in BBM’s integritynd the activities of their components.
.2.3. Effect of GT consumption on CP-induced alterations ina-gradient dependent transport of 32Pi in BBMV(s) isolated from
enal cortexThe bulk of filtered Pi in the kidney is reabsorbed by its proximal
ubule. The Na-gradient dependent {Naoutside (Nao) > Nainside (Nai)}ransport of Pi in renal proximal tubule across its luminal BBM is annitial and regulatory step. The Pi is transported by secondary activeransport mechanism, which requires expenditure of energy in theorm of ATP that is generated by cellular metabolism. The uptakef 32Pi was determined in the presence and absence of Na-gradientn the initial uphill phase (30 s) and after equilibrium (120 min) inBM preparations. The rate of concentrative uphill uptake of 32Pi inhe presence of a Na-gradient (NaCl in the medium) was markedlyecreased in CP-treated compared to control rats (Table 5). How-ver, the uptake of 32Pi at the equilibrium phase (120 min) when
ao = Nai was not significantly different between the two groups.lso, Na-independent uptake (in the absence of a Na-gradient,hen NaCl in the medium was replaced by KCl where Ko > Ki) of2Pi at 30 s and 120 min was also not affected by CP treatment indi-ating specific alteration only when Na-gradient was present. The
able 7ffect of green tea (GT) consumption on the activity of metabolic enzymes in cortical and
Tissue G6Pase (�mol/mg protein/h)
CortexControl 0.29 ± 0.03CP 0.17 ± 0.01* (−41%)GT 0.36 ± 0.01* (+24%)TCP 0.32 ± 0.02† (+10%)
MedullaControl 0.22 ± 0.01CP 0.15 ± 0.01* (−32%)GT 0.34 ± 0.03* (+55%)TCP 0.26 ± 0.01*,† (+18%)
esults are mean ± SEM of 3–4 different preparations. Values in parentheses represent pe* Significantly different from control.† Significantly different from CP at p < 0.05 by one-way ANOVA.
rcent change from control. NS: Not Significant.
net uptake of 32Pi (Table 5c) was also significantly decreased. Incontrast, dietary GT supplementation alone significantly increasedthe uptake of 32Pi in the uphill phase (30 s) in the presence of a Na-gradient. When CP treatment was extended to GT rats, CP-induceddecrease in 32Pi transport was not observed. Moreover, the net rateof uptake of 32Pi remained significantly higher in TCP rats comparedto control rats and significantly much more higher compared to CP-treated rats. Similar to the effect of CP alone, the effect of GT aloneor in combination with CP treatment was specific in the presence ofa Na-gradient. The transport of 32Pi in the absence of Na-gradient(i.e., where Ko > Ki) and after 120 min were not affected by any ofthese treatments.
3.3. Effect of GT on CP-induced alterations on the enzymes ofcarbohydrate metabolism in rat renal tissues
The effect of CP, GT, and TCP was determined on the activi-ties of various enzymes of carbohydrate metabolism in the kidney(Tables 6 and 7). CP treatment to control rats significantly increasedhexokinase activity in the renal cortex (+5%) and medulla (+36%).The activity of LDH (a marker of anaerobic glycolysis), however,profoundly increased in the renal cortex (+70%) and medulla(+27%). CP caused significant decline in MDH activity in the renalcortex (−23%) and medulla (−27%). As previously reported [27],GT consumption significantly enhanced metabolic activities inalmost all tissues studied. The activity of HK, LDH, and MDH sig-nificantly increased in the renal cortex and medulla with theexception of HK activity in the cortex and MDH activity in renal
medulla, which was decreased by GT. When CP treatment wasextended to GT drinking rats, CP-induced decrease in MDH activ-ity was not only prevented but the activity remained significantlyhigher in TCP compared to either control or CP-treated rats in therenal cortex and medulla (Table 6). The activity of LDH was espe-medullary homogenates with/without cisplatin (CP) treatment.
G6PDH (�mol/mg protein/h) ME (�mol/mg protein/h)
0.30 ± 0.03 0.38 ± 0.070.23 ± 0.04 (−23%) 0.48 ± 0.01 (+26%)0.65 ± 0.13* (+117%) 0.3 ± 0.24 (−21%)0.54 ± 0.09*,† (+80%) 0.31 ± 0.14† (−18%)
0.47 ± 0.05 0.96 ± 0.240.21 ± 0.01* (−55%) 1.12 ± 0.03 (+17%)0.34 ± 0.04* (−28%) 0.52 ± 0.07* (−46%)0.23 ± 0.05* (−51%) 0.87 ± 0.04† (NS)
rcent change from control. NS: Not Significant.

3 gical R
cm
(ianmsdGomrGTmab(trhr
3p
e(atnasfeHrdaccenOtt
TE
R
88 S.A. Khan et al. / Pharmacolo
ially found to be profoundly enhanced in the renal cortex andedulla.The effect of CP and GT on the activity of glucose-6-phosphatase
G6Pase), a marker enzyme of gluconeogenesis was also observedn renal tissues. However, CP caused marked reduction in G6Pasectivity in renal cortex and medulla (Table 7). In contrast, GT sig-ificantly enhanced the activity of G6Pase in the renal cortex andedulla. As a result GT consumption by CP-treated rats resulted in
ignificant increase of G6Pase in the renal tissues and CP-inducedecrease in enzyme activity was markedly prevented. The effect ofT extract alone and with CP was also determined on the activitiesf glucose-6-phosphate dehydrogenase (G6PDH, HMP-shunt) andalic enzyme (ME), source of NADPH needed in reductive anabolic
eactions. CP treatment to control rats significantly decreased6PDH activity in the renal cortex (−23%) and medulla (−55%).he activity of ME, however, significantly increased by CP treat-ent in the renal cortex (+26%) and medulla (+17%) (Table 7). The
ctivities of G6PDH and ME, however, were differentially alteredy GT. The activity of G6PDH profoundly increased in renal cortex+117%) whereas, decreased in the medulla (−28%). ME activity onhe other hand was lowered by GT. GT consumption by CP-treatedats resulted in significant increase in G6PDH activity in the cortex,owever, the activity of both G6PDH and ME slightly improved butemained lower in renal tissues compared to control rats.
.4. Effect of GT on CP-induced alterations in antioxidant defensearameters in rat renal tissues
The effect of CP, GT, and TCP was determined on the param-ters of antioxidant defense system such as lipid peroxidationLPO), total-SH, SOD, and catalase (Table 8). Antioxidant status ispotential biomarker to determine the physiological state of a cell,
issue, or organ. The drug including GM or CP and chemical inducedephrotoxicity and tissue injury has been suggested to be medi-ted in part by perturbation in the balance of antioxidant defenseystem [39,40]. CP administration to control rats resulted in a pro-ound increase in the production of malondialdehyde (MDA), annd-product of lipid peroxidation in the renal cortex and medulla.owever, the increase in LPO was much more pronounced in the
enal cortex followed by medulla. Total-SH was found to be slightlyecreased in renal tissues. These changes in LPO and total-SH weressociated with marked decrease in the activity of both SOD andatalase in the cortex and medulla. In contrast, the effect of GTonsumption was differentially observed on the activity of these
nzymes and other parameters in the kidney. GT consumption sig-ificantly increased the activity of SOD in renal cortex and medulla.n the other hand, the activity of catalase significantly increased inhe renal cortex but decreased in the medulla. These alterations inhe antioxidant enzymes were associated with marked decrease of
able 8ffect of green tea (GT) consumption on enzymic and non-enzymic antioxidant paramete
Tissue LPO (nmol/g tissue) Total-SH (�mol/g tissue)
CortexControl 147.2 ± 5.89 8.1 ± 0.1CP 207.3 ± 11.9* (+41%) 8.84 ± 0.46 –GT 107.9 ± 5.8* (−27%) 6.4 ± 0.07* (−21%)TCP 133.3 ± 4.5† (NS) 6.5 ± 0.13* (−20%)
MedullaControl 127 ± 11.2 2.66 ± 0.24CP 143 ± 3.8 (+13%) 3.14 ± 0.24 (+18%)GT 176.75 ± 1.35* (+39%) 3.2 ± 0.15 (+20%)TCP 146.45 ± 3.8 (+15%) 3.2 ± 0.42 (+20%)
esults are mean ± SEM of 3–4 different preparations. Values in parentheses represent pe* Significantly different from control.† Significantly different from CP at p < 0.05 by one-way ANOVA.
esearch 60 (2009) 382–391
LPO in renal cortex. However, LPO was significantly higher in themedulla in GT compared to control rats. The total-SH levels wereincreased in medulla but the levels decreased in the cortex. Con-sumption of GT along with CP treatment significantly preventedCP-induced increase in LPO in the renal tissues. GT consumptionshowed marked protection against CP-induced decrease in SODactivity in the cortex and medulla. CP-induced decrease in cata-lase activity was also prevented by GT consumption in the renalcortex, whereas catalase activity remained lower in the medullaafter GT consumption by CP-treated rats. In general, the presentresults demonstrate marked protection by GT against CP-inducedalterations in oxidative stress parameters.
4. Discussion
Cisplatin (cis-diamminedichloroplatinum II, CP) is a major anti-neoplastic drug for the treatment of various forms of cancers[41,42]. However, CP and its analogs accumulate in the kidney caus-ing nephrotoxicity; thus, limiting its long-term clinical use [43].Several strategies and agents were utilized to prevent CP nephropa-thy but were not found suitable for clinical practice [3].
The concept of cancer prevention using naturally occurring sub-stances that can be included in the diet consumed by the humanpopulation is gaining increasing attention. GT polyphenols namelycatechins help prevent a diverse spectrum of diseases [11,44,45].Experimental studies have shown an inverse association betweenGT consumption with the development of certain types of cancer[46,16].
The present studies were performed to test the hypothesis thatGT consumption would ameliorate CP-induced nephrotoxic andother adverse effects thus maximizing the clinical use of CP in thetreatment of various malignancies without any side effects.
Multiple injections of CP were administered to control andGT consuming rats and the effects on various serum parame-ters and on the activities of various enzymes of carbohydratemetabolism, brush border membrane, mitochondria, and lyso-somes were determined to ascertain potential beneficial effectsof GT. As shown in Section 3, CP administration to control ratsresulted in a marked increase in serum creatinine and BUN, diag-nostic indicators of nephrotoxicity, accompanied by significantdecrease in serum glucose, inorganic phosphate (Pi) and an increasein serum cholesterol and phospholipids. However, GT given toCP-treated rats significantly lowered CP elicited increased con-centrations of both serum creatinine and BUN. Serum glucoseand cholesterol remained depressed whereas serum phospholipids
were profoundly enhanced on CP treatment to GT consuming rats.GT induced reductions in serum glucose, cholesterol along withweight loss can be considered as healthy indicators.The BBM of renal proximal tubules has been shown as the majorsite of CP-induced renal injury [43,47]. Since the BBM contains a
rs in cortical and medullary homogenates with/without cisplatin (CP) treatment.
SOD (Units/mg protein) Catalase (�mol/mg protein/min)
12.8 ± 0.4 133.7 ± 5.29.9 ± 0.63* (−23%) 110.9 ± 4.2* (−17%)
20.9 ± 0.96* (+63%) 176.5 ± 6.3* (+32%)21.1 ± 0.8*,† (+65%) 177.7 ± 2.6*,† (+33%)
9.9 ± 1.3 143.6 ± 6.07.02 ± 1.28 (−29%) 128.9 ± 3.2 (−10%)14.4 ± 0.45* (+46%) 105.7 ± 0.11* (−26%)
11.25 ± 0.6† (+14%) 110.6 ± 4.2* (−23%)
rcent change from control. NS: Not Significant.

gical R
notcesCeosodfdIti[cto
itGprCBcahtAtlethd[
cpodwtipouacicitoiagiree
S.A. Khan et al. / Pharmacolo
umber of hydrolytic enzymes and transport systems, the effectf CP was determined on the activities of BBM enzymes and on Piransport to assess damage caused by CP administration. CP signifi-antly decreased the activities of ALP, GGTase, and LAP, BBM markernzymes, in cortical homogenates and isolated BBM vesicles. Aimilar decrease was observed in medulla, suggesting an overallP-induced damage to the kidney. The CP-induced decrease in BBMnzymes suggests a severe damage to the structural architecturef the BBM affecting its transport functions as these enzymes werehown to be directly or indirectly involved in the transport of vari-us solutes [29,38]. The decrease in BBM enzyme activities is in factue to the loss/release of enzymes and other proteinic componentsrom damaged BBM into the lumen that later appear in the urine asemonstrated previously for CP and other toxicants [27,28,47,48].
n contrast to CP, GT consumption, however, significantly increasedhe activities of BBM enzymes in the homogenate and BBM, indicat-ng an overall improvement in renal BBM integrity as shown earlier26,27]. A variable increase in the activity of ALP/GGTase in the renalortex and medulla can be considered due to their localization inhe thickness of BBM [38] or due to differential access/accumulationf GT in these tissues.
GT consumption in combination with CP treatment resultedn the reversal of CP-induced alterations in the activities of cer-ain BBM enzymes in the renal tissues. The activities of ALP andGTase in renal BBMV remained significantly higher in TCP com-ared to CP-treated renal BBM preparations, indicating a markedeversal of CP-induced effect by GT consumption on these enzymes.P-induced decrease in LAP was also reduced by GT in renalBM preparations. The results convincingly demonstrate that GTonsumption not only prevented the CP elicited decrease in thectivities of certain enzymes, but they remained significantlyigher in TCP compared to control and much higher than CP-reated rats as shown earlier [27]. The activity of lysosomal enzyme,CPase was significantly increased in the cortex and medulla by CP
reatment. Alterations in ACPase activity demonstrate CP-inducedoss of lysosomal function [49,39]. Confirming morphologic andarlier biochemical studies [7,9,47] the present results indicatehat that renal BBM and lysosomes were severely damaged by CP,owever, GT consumption prevented CP effects by lessening theamage caused or by increasing the regeneration process or both27].
The reabsorption of Na+ ions by renal proximal tubular BBM isonsidered to be the major function of the kidney because the trans-ort of other ions and various solutes including Pi depends directlyr indirectly on Na reabsorption [29,38]. Since these transportsepend on structural integrity of BBM and available energy as ATP,hich is supplied by various metabolic pathways, it is imperative
hat any alterations to these metabolic pathways caused by toxicnsult or otherwise modified would determine the rate of trans-ort function [28,30]. To assess the functional aspects, the activitiesf various enzymes of carbohydrate metabolism were determinednder different experimental conditions. As shown in Section 3, thectivities of various enzymes involved in glycolysis, TCA cycle, glu-oneogenesis, and HMP-shunt pathway were differentially alteredn renal tissues by CP treatment and/or by GT consumption. CPaused significant increase in LDH but decrease in MDH activityn the renal cortex and medulla which was associated with simul-aneous increase in hexokinase activity. Although the actual ratesf glycolysis or TCA cycle were not determined, marked decreasen MDH activity in tissues appears to be due to CP-induced dam-ge to mitochondria [8,39]. An impaired oxidative metabolism of
lucose/fatty acid due to mitochondrial dysfunction would resultn lower ATP production by oxidative metabolism and hence lowerenal transport function. A marked increase in LDH and to somextent hexokinase activity with simultaneous decline in TCA cyclenzyme, MDH appears to be an adaptive cellular effect in energyesearch 60 (2009) 382–391 389
metabolism from aerobic metabolism alternatively to anaerobicglycolysis due to CP-induced mitochondrial dysfunction.
CP also caused differential effect on the enzymes of gluco-neogenesis and HMP-shunt pathway in different tissues. Theactivity of G6Pase (gluconeogenesis) and G6PDH (HMP-shunt)significantly decreased but the activity of NADP–malic enzyme(ME), variably increased in the renal tissues. In contrast, GT con-sumption significantly increased G6PDH whereas decreased ME.Increased production of NADPH by G6PDH due to GT consump-tion may support reductive anabolic reactions and also plays a rolein strengthening antioxidative defense mechanism. The presentdata indicate that CP downregulated the enzymes of carbohy-drate metabolism. In contrast to CP, GT consumption significantlyincreased the activity of HK, LDH (glycolysis), MDH (TCA cycle) andG6Pase as reported earlier [26]. Accordingly, GT administration toCP-treated rats significantly improved carbohydrate metabolismas evident by higher activities of LDH, MDH and gluconeogenicenzymes in these tissues in TCP compared to CP alone or controlrats. This can be attributed to the fact that GT might have lowerednumber of damaged mitochondria or increased the number of nor-mally active organelles. The decrease/increase in G6Pase by CP/GT,respectively can be attributed to the availability of oxaloacetateproduced from malate by MDH. Thus decrease/increase in MDHactivity may affect both the TCA cycle and gluconeogenic activity asoxaloacetate is required by both pathways. In turn, ATP productionwould be influenced accordingly.
The Pi required for the production of metabolites and ATP infactis conserved by the kidney. The transport of Pi by renal corticalBBM is an essential and regulatory step. It appears from the datathat CP by lowering oxidative metabolism lowers ATP production,whereas GT consumption increased ATP production by increasingthe metabolic activities to support many cellular functions espe-cially in the kidney. Thus, in both the situations the transportfunctions were affected accordingly. The present results show thatNa-dependent transport of Pi significantly decreased by CP treat-ment but increased by GT consumption. GT given to CP rats furtherenhanced renal BBM Pi transport capacity. Thus it appears thatGT induced Pi conservation by the kidney maintained a positivebalance of the Pi in the serum to support metabolic activities.
It has been reported that some toxicants including certain drugs[27,28,40,48] and heavy metals including CP [47,50,51] exert theirtoxic effects by inducing the generation of reactive oxygen species(ROS). However, oxidative stress can occur as a result of eitherincreased ROS generation and/or decreased antioxidant enzymesystem comprising SOD, catalase and GSH-Px. These antioxidantenzymes protect the cell against cytotoxic ROS. CP has been shownto generate ROS and to inhibit antioxidant enzymes in renal tissues.CP has been shown to enhance lipid peroxidation (LPO), an indi-cator of tissue injury and deplete GSH and protein thiols [47,52].In agreement with previous observations [47,52], CP significantlydecreased the activities of SOD, and catalase and enhanced LPOin the renal tissues indicating CP-induced oxidative damage. Theseverity of the damage appeared to be more prominent in the renalcortex than in the medulla.
The present results show that GT, by virtue of its antioxi-dant properties [16,26,27,47] significantly enhanced antioxidantdefense mechanism. This can be supported by a marked increasein SOD and catalase activities in the cortex and medulla associ-ated with lowering of LPO in the renal cortex. GT consumptionselectively prevented CP-induced increase in LPO and decrease inthe activities of SOD and catalase in the renal cortex. CP-induced
reduction in SOD activity in the medulla was also prevented byGT consumption. The results suggest that GT provide protectionagainst CP-induced oxidative damage either by SOD-mediatedor catalase mediated mechanism or involving both mechanismssimultaneously in the renal tissues as shown earlier [26,27].
3 gical R
uscsbecmdriiaodpiir
5
tieepttcTmwke
A
I(RaApc(ag
R
[
[[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
90 S.A. Khan et al. / Pharmacolo
The mechanism by which CP causes nephrotoxicity remainsnclear. It has been shown that CP induces necrosis of the S3 sub-egment of proximal tubule [6,53]. It has been morphologicallyharacterized by loss of microvilli, alteration in the number andize of lysosomes and mitochondrial vacuolization accompaniedy functional disturbances of various cell organelles [39]. Differ-nt studies have shown that cytotoxicity of CP is probably due toombination of insults including peroxidation of cell membrane,itochondrial dysfunction, inhibition of protein synthesis and DNA
amage in the kidney [54]. It has been suggested that CP inducesenal damage by free radical generation, such as hydroxyl rad-cal and superoxide anion, by altering arginine metabolism, byncreasing activity of calcium independent nitric oxide synthasend more recently by apoptosis [54–56]. However, CP-inducedxidative stress was shown to be strongly involved in acute renalamage [52]. The present findings at least in part support aboveroposed mechanism. Our results indicate an overall improvement
n metabolic activities and augmentation of antioxidant defensesn CP-treated GT-fed rats most likely by GT-polyphenol-mediatededuction of oxidative damage.
. Conclusions
The present results show that CP elicited deleterious nephro-oxic and other adverse effects as indicated by significant increasen serum creatinine, BUN, decrease in the activities of variousnzymes of BBM, mitochondria, lysosomes and metabolism. Theffects appeared to be mediated, at least, in part due to CP-inducederturbation in antioxidant defense mechanism. GT consumptiono a larger extent prevented CP-induced nephrotoxicity parame-ers by enhancing energy metabolism, BBM enzymes, 32Pi transportapacity, and by strengthening antioxidant defense mechanism.aking into account these results, we propose that GT may maxi-ize the clinical use of CP in the treatment of various malignanciesithout nephrotoxic and other side effects as well as due to its
nown anticarcinogenic effect may exhibit a synergistic/additiveffect.
cknowledgements
Council of Scientific and Industrial Research (CSIR), New Delhi,ndia is acknowledged for the award of Junior Research FellowshipJRF)/Senior Research Fellowship to SAK, Indian Council of Medicalesearch (ICMR), New Delhi, India for the award of JRF/SRF to S.P.nd W.K. Authors gratefully acknowledge the help extended by Dr.bbas Abidi, Department of Zoology, Aligarh Muslim University, forroviding the necessary facilities for radioactive counting. Finan-ial support to the department from University Grant CommissionUGC-DRF), Department of Science and Technology (DST-FIST) and
research grant (58/21/2001-BMS) from ICMR to ANKY is alsoratefully acknowledged.
eferences
[1] Cohen SM, Lippard SJ. Cisplatin: from DNA damage to cancer chemotherapy.Prog Nucleic Acid Res Mol Biol 2001;67:93–130.
[2] Ward JM, Fauvie KA. The nephrotoxic effects of cis-diam-mine-dicholroplatinum II in male F344 rats. Toxicol Appl Pharmacol 1976;38:535–47.
[3] Ali BH, Al Moundhri SM. Agents ameliorating or augmenting the nephrotoxicityof cisplatin and other platinum compounds: a review of some recent research.Food Chem Toxicol 2006;44:1173–83.
[4] McKeage MJ. Comparative adverse effect profiles of platinum drugs. Drug Safety
1995;13:228–44.[5] Markman M. Toxicities of the platinum antineoplastic agents. Exp Opin DrugSafety 2003;2:597–607.
[6] Townsend DM, Deng M, Zhang L, Lapus MG, Hanigan MH. Metabolismof cisplatin to a nephrotoxin in proximal tubule cells. J Am Soc Nephrol2003;14:1–10.
[
[
esearch 60 (2009) 382–391
[7] Dobyan DC, Levi J, Jacobs C, Kosek J, Weiner MW. Mechanism of cis-platinum nephrotoxicity: II. Morphologic observations. J Pharmacol Exp Ther1980;213:551–6.
[8] Zhang JG, Lindup WE. Role of mitochondria in cisplatin-induced oxidativedamage exhibited by rat renal cortical slices. Biochem Pharmacol 1993;45:2215–22.
[9] Leibbrandt MEI, Grushenka HIW, Metz AL, Ozobia AA, Haskins JR. Critical sub-cellular targets of cisplatin and related platinum analogs in rat renal proximaltubule cells. Kidney Int 1995;48:761–70.
10] Dauggard G. Cisplatin nephrotoxicity. Experimental and clinical studies. DanMed Bull 1990;37:1–12.
11] Alschuler L. Green tea: healing tonic. Am J Nat Med 1998;5:28–31.12] Connor WE. Importance of n-3 fatty acids in health and disease. Am J Clin Nutr
2000;71:171S–5S.13] Weisburger JH. Tea and health: the underlying mechanisms. Proc Soc Exp Biol
Med 1999;220:271–5.14] Imai K, Mukhtar H. Tea antioxidants in cancer chemoprevention. Cell Biochem
1997;27:S59–67.15] Ahmad N, Mukhtar H. Green tea polyphenols and cancer: biological mecha-
nisms and practical implications. Nutr Rev 1999:78–83.16] Khan N, Mukhtar H. Tea polyphenols for health promotion. Life Sci
2007;81:519–33.17] Zhu QY, Zhang A, Tsang D, Huang Y, Chen ZY. Stability of green tea catechins. J
Agric Food Chem 1997;45:4624–8.18] Dufresne CJ, Farnworth ER. A review of latest research findings on the health
promotion properties of tea. J Nutr Biochem 2001;12:404–21.19] Zloch Z. The role of dietary plant polyphenols in health maintenance. Cas Lek
Cesk 1996;135:84–8.20] Higdon JV, Frei B. Tea catechins and polyphenols: health effects, metabolism
and antioxidant functions. Crit Rev Food Sci Nutr 2003;43:89–143.21] Rhee SJ, Choi JH, Park MR. Green tea catechins improves microsomal phospho-
lipase A2 action in arachidonic acid cascade system in the kidney of diabeticrats. Asia Pac J Clin Nutr 2002;11(3):226–31.
22] Mohamadin AM, El-Beshbishy HA, El-Mahdy MA. Green tea extract attenuatescyclosporine-A induced oxidative stress in rats. Pharmacol Res 2005;51:51–7.
23] Leena P, Balaraman R. Effect of green tea extract on cisplatin induced oxidativedamage on kidney and testes of rats. Ars Pharm 2005;46:5–18.
24] Yokozawa T, Nakagawa T, Kyeoungi M, Eun JC, Shigeya T, Terasawa K. Effect ofgreen tea tannin on cisplatin induced nephropathy in LLC-PK1 cells and rats. JPharm Pharmacol 1995;1:1325–31.
25] Upaganlawar A, Farswan M, Rathod S, Balaraman R. Modification of biochemicalparameters of gentamicin nephrotoxicity by coenzyme Q10 and green tea inrats. Indian J Exp Biol 2006;44:416–8.
26] Khan SA, Priyamvada S, Arivarasu NA, Khan S, Yusufi ANK. Influence of greentea on enzymes of carbohydrate metabolism, antioxidant defense and plasmamembrane in rat tissues. Nutrition 2007;23:687–95.
27] Khan SA, Priyamvada S, Farooq N, Khan S, Khan MW, Yusufi ANK. Protectiveeffect of green tea extract on gentamicin-induced nephrotoxicity and oxidativedamage in rat kidney. Pharm Res 2009;59:254–62.
28] Banday AA, Farooq N, Priyamvada S, Yusufi ANK, Khan F. Time dependent effectsof gentamicin on the enzymes of carbohydrate metabolism, brush border mem-brane and oxidative stress in rat kidney tissues. Life Sci 2008;82(9):450–9.
29] Yusufi ANK, Dousa TP. Studies on rabbit kidney brush border membranes: rela-tionship between phosphate transport, alkaline phosphatase and NAD. MinElect Metab 1987;13:397–404.
30] Khundmiri SJ, Asghar M, Khan F, Salim S, Yusufi ANK. Effect of ischemia andreperfusion on enzymes of carbohydrate metabolism in rat kidney. J Nephrol2004;17:1–7.
31] Crane RK, Sols A. The association of particulate fractions of brain and othertissue homogenates. J Biol Chem 1953;203:273–92.
32] Nelson NA. Photometric adaptation of the Somogyi method for the determina-tion of glucose. J Biol Chem 1944;153:375–81.
33] Khundmiri SJ, Asghar M, Khan F, Salim S, Yusufi ANK. Effect of reversible andirreversible ischemia on marker enzymes of BBM from renal cortical PT sub-populations. Am J Physiol 1997;273:F849–56.
34] Marklund S, Marklund G. Involvement of the superoxide anion radical in theauto oxidation of pyrogallol and a convenient assay for superoxide dismutase.Eur J Biochem 1974;47:469–74.
35] Giri U, Iqbal M, Athar M. Porphyrin-mediated photosensitization has a weaktumor promoting activity in mouse skin: possible role of in-situ generatedreactive oxygen species. Carcinogenesis 1996;17:2023–8.
36] Sedlak J, Lindsay RH. Estimation of total protein bound and non-protein boundSH groups in tissue with Ellman’s reagent. Anal Biochem 1968;25:192–205.
37] Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues bythiobarbituric acid reaction. Anal Biochem 1979;95:351–8.
38] Yusufi ANK, Murayama N, Gapstur SM, Szczepanska-Konkel M, Dousa TP. Dif-ferential properties of brush border membrane vesicles from early and lateproximal tubules of rat kidney. Biochim Biophys Acta 1994;1191:113–7.
39] Kuhlmann MK, Burkhardt G, Kohler H. Insights into potential cellular mech-anisms of cisplatin nephrotoxicity and their clinical application. Nephrol Dial
Transplant 1997;12:2478–80.40] Walker PD, Barri Y, Shah SV. Oxidant mechanisms on gentamicin nephrotoxi-city. Ren Fail 1999;21:433–42.
41] Nakashima T, Maehara Y, Kohnoe S, Hayashi I, Katsuta Y. Histologic differentia-tion and chemosensitivity of human head and neck squamous cell carcinomas.Head Neck 1990;12:406–10.

gical R
[
[[
[
[
[
[
[
[
[
[
[
[
[55] Devipriya S, Shyamala Devi CS. Protective effect of quercetin in cisplatin-induced cell injury in the rat kidney. Indian J Pharmacol 1999;31:
S.A. Khan et al. / Pharmacolo
42] Taguchi T, Nazneen A, Abid MR, Razzaque MS. Cisplatin-associated nephrotox-icity and pathological events. Contrib Nephrol 2005;148:107–21.
43] Arany I, Safirstein RL. Cisplatin nephrotoxicity. Semin Nephrol 2003;23:460–4.44] Graham HN. Green tea composition, consumption and polyphenol chemistry.
Toxicol Sci 1999;52:111–7.45] Liao S. The medicinal action of androgens and green tea epigallocatechin gallate.
Hong Kong Med 2001;7:369–74.46] Mukhtar H, Ahmad N. Tea polyphenols: prevention of cancer and optimizing
health. Am J Clin Nutr 2000;71:1698S–702S.47] Fatima S, Yusufi ANK, Mahmood R. Effect of cisplatin on renal brush
border membrane enzymes and phosphate transport. Hum Exp Toxicol2004;23:547–54.
48] Priyamvada S, Priyadarshini M, Arivarasu NA, Farooq N, Khan S, Khan SA, etal. Studies on the protective effect of dietary fish oil on gentamicin-inducednephrotoxicity and oxidative damage in rat kidney. Prostaglandins LeukotEssent Fatty Acids 2008;78:369–81.
49] Courjault-Gautier F, Le Grimellec C, Giocondi MC, Toutain HJ. Modulation ofsodium coupled uptake and membrane fluidity by cisplatin in renal proximaltubular cells in primary culture and BBM vesicles. Kidney Int 1995;47:1048–56.
50] Baliga R, Zhang Z, Baliga M, Ueda N, Shah SV. In vitro and in vivo evi-dence suggesting a role of iron in cisplatin-induced nephrotoxicity. Kidney Int1998;53:394–401.
[
esearch 60 (2009) 382–391 391
51] Banday AA, Priyamvada S, Farooq N, Yusufi ANK, Khan F. Effect ofuranyl nitrate on enzymes of carbohydrate metabolism and brush bor-der membrane in different kidney tissues. Food Chem Toxicol 2008;46:2080–8.
52] Gonzales R, Romay C, Borrego A, Hernandez F, Zamora Z, Rojas E. Lipid peroxidesand antioxidant enzymes in cispaltin chronic nephrotoxicity in rats. MediatorsInflamm 2005;3:139–43.
53] Portilla D, Li S, Nagothu KK, Megyesi J, Kaissling B, Schnackenberg I,et al. Metabolomic study of cispaltin induced nephrotoxicity. Kidney Int2006;69:2194–204.
54] Santos NA, Catão CS, Martins NM, Curti C, Bianchi ML, Santos AC. Cisplatin-induced nephrotoxicity is associated with oxidative stress, redox stateunbalance, impairment of energetic metabolism and apoptosis in rat kidneymitochondria. Arch Toxicol 2007;81:495–504.
422–6.56] Arany I, Megyesi JK, Kaneto H, Price PM, Safirstein RL. Cisplatin-induced cell
death is EGFR/src/ERK signaling dependent in mouse proximal tubule cells.Am J Physiol Renal Physiol 2004;287:F543–549.
Related Documents











