2013;73:4362-4371. Published OnlineFirst April 10, 2013. Cancer Res Koji Kikuchi, Takeo Narita, Van T. Pham, et al. but Essential Roles in DNA Repair by Homologous Recombination Structure-Specific Endonucleases Xpf and Mus81 Play Overlapping Updated version 10.1158/0008-5472.CAN-12-3154 doi: Access the most recent version of this article at: Material Supplementary http://cancerres.aacrjournals.org/content/suppl/2013/06/13/0008-5472.CAN-12-3154.DC1.html Access the most recent supplemental material at: Cited Articles http://cancerres.aacrjournals.org/content/73/14/4362.full.html#ref-list-1 This article cites by 48 articles, 16 of which you can access for free at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at Permissions . [email protected] To request permission to re-use all or part of this article, contact the AACR Publications Department at on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154 on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

2013;73:4362-4371. Published OnlineFirst April 10, 2013.Cancer Res Koji Kikuchi, Takeo Narita, Van T. Pham, et al. but Essential Roles in DNA Repair by Homologous RecombinationStructure-Specific Endonucleases Xpf and Mus81 Play Overlapping
Updated version
10.1158/0008-5472.CAN-12-3154doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/06/13/0008-5472.CAN-12-3154.DC1.html
Access the most recent supplemental material at:
Cited Articles
http://cancerres.aacrjournals.org/content/73/14/4362.full.html#ref-list-1
This article cites by 48 articles, 16 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications Department at
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154

Therapeutics, Targets, and Chemical Biology
Structure-Specific Endonucleases Xpf and Mus81 PlayOverlapping but Essential Roles in DNA Repair byHomologous Recombination
Koji Kikuchi1, Takeo Narita1, Van T. Pham1, Junko Iijima1, Kouji Hirota1, Islam Shamima Keka1,Mohiuddin 1, Katsuya Okawa2, Tetsuya Hori3, Tatsuo Fukagawa3, Jeroen Essers5,6,7, Roland Kanaar6,7,Matthew C. Whitby8, Kaoru Sugasawa4, Yoshihito Taniguchi1, Katsumi Kitagawa9, and Shunichi Takeda1
AbstractDNA double-strand breaks (DSB) occur frequently during replication in sister chromatids and are dramatically
increased when cells are exposed to chemotherapeutic agents including camptothecin. Such DSBs are efficientlyrepaired specifically by homologous recombination (HR) with the intact sister chromatid. HR, therefore, playspivotal roles in cellular proliferation and cellular tolerance to camptothecin. Mammalian cells carry severalstructure-specific endonucleases, such as Xpf-Ercc1 and Mus81-Eme1, in which Xpf and Mus81 are the essentialsubunits for enzymatic activity. Here, we show the functional overlap between Xpf and Mus81 by conditionallyinactivating Xpf in the chicken DT40 cell line, which has noMus81 ortholog. Althoughmammalian cells deficientin either Xpf or Mus81 are viable, Xpf inactivation in DT40 cells was lethal, resulting in a marked increase in thenumber of spontaneous chromosome breaks. Similarly, inactivation of both Xpf and Mus81 in human HeLacells and murine embryonic stem cells caused numerous spontaneous chromosome breaks. Furthermore, thephenotype of Xpf-deficient DT40 cells was reversed by ectopic expression of human Mus81-Eme1 or humanXpf-Ercc1 heterodimers. These observations indicate the functional overlap of Xpf-Ercc1 andMus81-Eme1 in themaintenance of genomicDNA. BothMus81-Eme1 andXpf-Ercc1 contribute to the completion ofHR, as evidencedby the data that the expression of Mus81-Eme1 or Xpf-Ercc1 diminished the number of camptothecin-inducedchromosome breaks in Xpf-deficient DT40 cells, and to preventing early steps in HR by deleting XRCC3 sup-pressed the nonviability of Xpf-deficient DT40 cells. In summary, Xpf andMus81 have a substantially overlappingfunction in completion of HR. Cancer Res; 73(14); 4362–71. �2013 AACR.
IntroductionHomologous recombination (HR) mediated double-strand
break (DSB) repair is initiated by resection of DSBs andformation of 30-single-strand overhangs, followed by polymer-ization of Rad51 (1, 2). The resulting nucleoprotein filaments,
consisting of the 30-single-strand tail and the polymerizedRad51, undergo homology search and pairing with the intactduplex DNA donor to form a displacement (D)-loop structure.Extensive strand exchange of the D-loop leads to the gener-ation of HR intermediates. HR intermediates are processedinto either crossover products or non-crossover products (3).HR intermediates including Holliday junction (HJ) are here-after called joint molecules.
The processing of joint molecules is carried out by dissolu-tion and resolution pathways. In the dissolution pathway, Sgs1(Blm, an ortholog of Sgs1 in human), topoisomerase III, andRMI1/2 collaboratively catalyze the decatenation of HJs andgenerate non-crossover products (4–6). In the resolution path-way, structure-specific endonucleases cleave the HJs and pro-duce crossover and non-crossover products, depending onthe choice of cleaved strands at the 4-way junction (7). InEscherichia coli (E. coli), the RusA nuclease and the RuvABCcomplex cleave HJs symmetrically and play a key role in theresolution pathway (8–10). Recent work with eukaryotes hasrevealed 2 resolvases, Gen1 (11) and the Slx1–Slx4 complex(12–15), both of which can symmetrically cleave the 4-wayjunction. A third enzyme called Mus81, together with itspartner Eme1/Mms4, has also been implicated in resolvingHJs and the formation of crossover recombinants in both
Authors' Affiliations: 1Department of Radiation Genetics, and 2FrontierTechnology Center, Graduate School of Medicine, Kyoto University,Sakyo-ku, Kyoto; 3Department of Molecular Genetics, National Instituteof Genetics and Sokendai, Shizuoka; 4Biosignal Research Center, Orga-nization of Advanced Science and Technology, Kobe University, Hyogo,Japan; 5Department of Vascular Surgery, Erasmus University MedicalCenter; Departments of 6Genetics and 7RadiationOncology, CancerGeno-mics Center, Erasmus University Medical Center, Rotterdam, the Nether-lands; 8Department of Biochemistry, University of Oxford, Oxford, UnitedKingdom; and 9Center for Childhood Cancer, The Research Institute atNationwide Children's Hospital, Columbus, Ohio
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
K. Kikuchi and T. Narita contributed equally to this work.
CorrespondingAuthor:Shunichi Takeda,Department of RadiationGenet-ics, Graduate School of Medicine, Kyoto University, Yoshida-konoe,Sakyo-ku, Kyoto 606-8501, Japan. Phone: 81-75-753-4410l; Fax: 81-75-753-4419; E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-12-3154
�2013 American Association for Cancer Research.
CancerResearch
Cancer Res; 73(14) July 15, 20134362
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154

mitotic and meiotic recombination in yeast and mammals(16, 17). In vitro data show an ability of Mus81 to incise HJsasymmetrically; however a greater predilection for cleavingD-loops and nicked HJs suggests that, unlike "classical" HJresolvases, Mus81 may process joint molecules before theymature into fully ligated HJs (16).Mus81 is a member of the Xpf family of structure-specific
endonucleases. Human Xpf is best known for its role innucleotide excision repair (NER), together with its partnerErcc1. Moreover, Xpf, but not the other NER factors, is involv-ed in DSB repair including single-strand annealing (18, 19),incision at interstrand crosslinks (20), and gene targeting(21). However, no studies have reported a functional overlapbetween Mus81 and Xpf in any DNA repair or recombinationreactions, or a role of Xpf in HR after the formation of jointmolecules. Intriguingly the Xpf orthologs in both DrosophilaandCaenorhabditis elegans (C. elegans) have been implicated injoint molecules processing and the formation of crossoversduring meiosis. Whether Xpf is similarly involved in jointmolecules processing in vertebrates alongside enzymes likeMus81 is currently unknown.To investigate the role of Xpf in HR, we conditionally
disrupted the XPF gene in the chicken B lymphocyte line DT40(22). Because this line does not possess the MUS81 gene, wepropose that the loss of Xpf in DT40 cells is equivalent tomammalian cells deficient in both Xpf and Mus81. Deletion ofXPF caused extensive chromosomal aberrations and cell death.However, this lethality was substantially reversed by ectopicexpression of human Mus81 together with Eme1 (HsMus81-Eme1), indicating a compensatory relationship between Xpfand Mus81 in the maintenance of chromosomal DNA. Thephenotypic analysis of Xpf-depletedDT40 cells indicated that amarked genomic instability may result from the defectiveprocessing of joint molecules. Our data uncovered that theXpf and Mus81 endonucleases play overlapping and essentialroles in completion of HR.
Materials and MethodsCell culture, plasmid constructs, and siRNAsChicken DT40, mouse embryonic stem [ES; wild-type (IB10)
and MUS81�/�], and HeLa cells were cultured as describedpreviously (23–25). DT40 or ES cells have been maintained byS. Takeda or J. Essers since 1991 or 2004, respectively (22, 24).HeLa cells were obtained from K. Myung (National HumanGenome Research Institute, Bethesda, MD ) in 2007 (25). Allcell lines were tested routinely for various criteria such asmorphology, growth rate, and karyotype. Details of plasmidconstructs and siRNAs are provided in Supplementary Materi-als and Methods.
Cell-cycle analysisAfter bromodeoxyuridine (BrdUrd) pulse-labeling in the
cells exposed to tamoxifen, cell-cycle distribution was mea-sured as described previously (26).
Measurement of chromosomal aberrationsChromosomal aberrations were measured as described
previously (23). Briefly, the cells were exposed to tamoxifen
for 1, 2, or 3 days, with 0.1 mg/mL of colcemid added for thelast 3 hours of incubation before harvesting. To measureionizing radiation (IR)-induced chromosomal aberrations,the cells were exposed to tamoxifen for 24 hours, andcolcemid was added immediately after cells were irradi-ated with 0.3 Gy g-rays. To test the response to camp-tothecin, cells were continuously exposed to tamoxifen for33 hours; these cells were also treated with 100 nmol/Lcamptothecin for the last 9 hours and with colcemid forthe last 3 hours before harvesting of mitotic cells.
Measurement of Rad51 subnuclear fociBefore measurement of Rad51 subnuclear foci, cells were
exposed to tamoxifen for 2 days. Rad51 foci were visualizedwith anti-Rad51 antibody in untreated cells and in cells g-irra-diated with 4 Gy, 3 and 6 hours after treatment as previouslydescribed (26).
Measurement of sister-chromatid exchange levelsSister-chromatid exchange (SCE) levels were measured as
described previously (27). Prior to BrdUrd labeling, cells wereexposed to tamoxifen for 30 hours. Cells were cultured in thepresence of 10mmol/L BrdUrd for 21 hours and treatedwith 0.1mg/mL of colcemid for the last 3 hours before harvesting.Camptothecin (5 or 100 nmol/L) was added 9 hours beforeharvesting.
Measurement of chromosome aberrations in HeLa andmouse ES cells
Chromosomal aberrations were measured as describedpreviously (23). Briefly, transfection of siRNA into HeLa ormouse ES cells using lipofectamine RNAiMAX or lipofecta-mine 2000 was carried out according to the manufacturer'sinstructions, respectively. Forty-five hours after the transfec-tion, HeLa cells were incubated for 1 hour with 0.2 mg/mLcolcemid; after this, metaphase cells were collected by mitoticshake-off. Alternatively, 72 hours after the transfection,mouse ES cells were incubated for 2 hours with 0.1 mg/mLcolcemid, after which metaphase cells were collected bymitotic shake-off.
Statistical analysisWe conducted 3 independent experiments for all of the data
sets. The results were expressed as mean� SD (growth curvesand sensitivity to the genotoxic agents) or mean � SEM(chromosomal aberrations and SCEs). Differences among thedata were tested for statistical significance using the t test.
Additional details are provided in Supplementary Materials.
ResultsMus81 protein is absent in chicken DT40 cells
Chicken XPF cDNA encodes a putative 903 amino acidproteins, compared with the 905 amino acids of the humanXpf (Supplementary Fig. S1). The sequence identity betweenthe 2 proteins is 76.8%. As expected, immunoprecipitants oftagged Xpf included Ercc1 (Supplementary Fig. S2A and S2B),indicating that Xpf associateswith Ercc1 inDT40 cells. As thereis an ortholog of Eme1 but not Mus81 registered in the chicken
Functional Overlap between Xpf and Mus81
www.aacrjournals.org Cancer Res; 73(14) July 15, 2013 4363
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154

genome database (Supplementary Fig. S3A and S3B; ref. 28), weanalyzed proteins that interact with Eme1, which forms aheterodimer with Mus81 in mammalian cells. Immunopreci-pitants of tagged Eme1 included Xpf, but not Mus81 (Supple-mentary Fig. S4A), indicating the absence of a functionalMus81–Eme1 complex in DT40 cells. To verify the absence ofMus81–Eme1, we disrupted the EME1 gene (SupplementaryFig. S5A). EME1�/� DT40 cells were able to proliferate andshowed moderate sensitivity only to cisplatin (SupplementaryFig. S5B and S5C), whose phenotype is in marked contrast withthe hypersensitivity of mouse EME1�/� andMUS81�/� ES cells
to a wide variety of DNA-damaging agents (29, 30). We, there-fore, concluded there was no Mus81 ortholog in DT40 cells.
The data indicate that the relationship among Xpf, Mus81,Eme1, and Ercc1 differs between DT40 and mammalian cells.The moderate sensitivity of EME1�/� DT40 cells to cisplatin(Supplementary Fig. S5C) indicates that Xpf might form afunctional complex with Eme1, whereas mammalian Mus81and Xpf form a heterodimer with Eme1 and Ercc1, respectively.This theory is supported by the data that, consistent with aprevious report (31), interaction of Xpf with Eme1 was con-firmed by co-immunoprecipitation of recombinant proteins
Figure 1. Xpf is essential for the maintenance of genome stability. A, targeted disruption of the chicken XPF gene. The chicken XPF locus, the 2 targetingconstructs, and the resulting targeted locus are shown. The black boxes represent the exons of the XPF gene. The triangles flanking the blastidine-resistance (bsrR) and histidinol-resistance (hisDR) genes represent the loxP sequences, the recognition site of the Cre recombinase. B, growth curveafter addition of tamoxifen to XPF�/�GdXPF-loxP cells at time zero for the excision of the GdXPF-loxP transgene. C, flow cytometric analysis ofcell-cycle distribution after BrdUrd pulse-labeling in XPF�/�GdXPF-loxP cells. D, spontaneous chromosomal aberrations in XPF�/�GdXPF-loxP cells.Top, a representative chromatid-type break (arrowhead) and a chromosome-type break (arrow). Breaks are magnified in the bottom panels. Bottom,measurement of spontaneous chromosomal aberrations. A chromatid-type break indicates a discontinuity in 1 of the 2 sister chromatids, and achromosome-type break indicates discontinuities at the same site of both sisters. Exchange indicates chromosomal translocation. The vertical axisshows the number of aberrations per cell. TAM, tamoxifen.
Kikuchi et al.
Cancer Res; 73(14) July 15, 2013 Cancer Research4364
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154
(Supplementary Fig. S4B and S4C). In addition to the presenceof Xpf–Eme1, it should be noted that we could not formallyexclude the possibility that a functional homolog of Mus81
is present in chicken cells. In conclusion, Xpf–Ercc1, but notXpf–Eme1, plays the major role in DNA damage response inchicken cells.
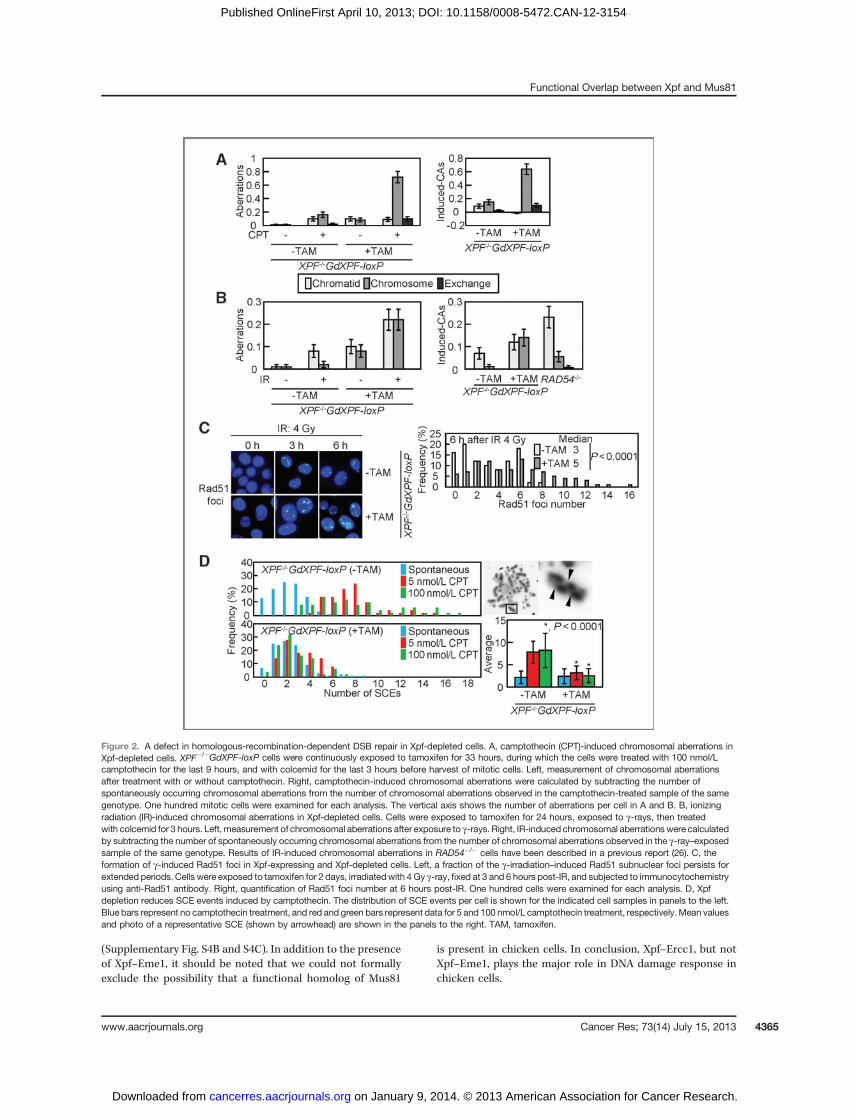
Figure 2. A defect in homologous-recombination-dependent DSB repair in Xpf-depleted cells. A, camptothecin (CPT)-induced chromosomal aberrations inXpf-depleted cells. XPF�/�GdXPF-loxP cells were continuously exposed to tamoxifen for 33 hours, during which the cells were treated with 100 nmol/Lcamptothecin for the last 9 hours, and with colcemid for the last 3 hours before harvest of mitotic cells. Left, measurement of chromosomal aberrationsafter treatment with or without camptothecin. Right, camptothecin-induced chromosomal aberrations were calculated by subtracting the number ofspontaneously occurring chromosomal aberrations from the number of chromosomal aberrations observed in the camptothecin-treated sample of the samegenotype. One hundred mitotic cells were examined for each analysis. The vertical axis shows the number of aberrations per cell in A and B. B, ionizingradiation (IR)-induced chromosomal aberrations in Xpf-depleted cells. Cells were exposed to tamoxifen for 24 hours, exposed to g-rays, then treatedwith colcemid for 3 hours. Left, measurement of chromosomal aberrations after exposure to g-rays. Right, IR-induced chromosomal aberrationswere calculatedby subtracting the number of spontaneously occurring chromosomal aberrations from the number of chromosomal aberrations observed in the g-ray–exposedsample of the same genotype. Results of IR-induced chromosomal aberrations in RAD54�/� cells have been described in a previous report (26). C, theformation of g-induced Rad51 foci in Xpf-expressing and Xpf-depleted cells. Left, a fraction of the g-irradiation–induced Rad51 subnuclear foci persists forextended periods. Cells were exposed to tamoxifen for 2 days, irradiatedwith 4 Gy g-ray, fixed at 3 and 6 hours post-IR, and subjected to immunocytochemistryusing anti-Rad51 antibody. Right, quantification of Rad51 foci number at 6 hours post-IR. One hundred cells were examined for each analysis. D, Xpfdepletion reduces SCE events induced by camptothecin. The distribution of SCE events per cell is shown for the indicated cell samples in panels to the left.Blue bars represent no camptothecin treatment, and red and green bars represent data for 5 and 100 nmol/L camptothecin treatment, respectively.Mean valuesand photo of a representative SCE (shown by arrowhead) are shown in the panels to the right. TAM, tamoxifen.
Functional Overlap between Xpf and Mus81
www.aacrjournals.org Cancer Res; 73(14) July 15, 2013 4365
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154
Deletion of XPF results in an accumulation ofchromosomal aberrations and subsequent cell death
We generated XPF gene-disruption constructs, whichdeleted amino acid coding sequences 1 to 148 together withthe transcriptional promoter sequences (Fig. 1A; Supplemen-tary Fig. S6A). Because we failed to establish XPF�/� cellsfrom XPFþ/� cells, we generated conditional Xpf-deficientcells using a chicken XPF transgene flanked by loxP-signalsequences (GdXPF-loxP), which is excised by the chimericCre recombinase carrying the tamoxifen-binding domain(Supplementary Fig. S6B). As the GdXPF-loxP transgene alsocarries a marker gene encoding the GFP, tamoxifen-medi-ated excision of the GdXPF-loxP transgene can be evaluatedby monitoring the loss of the GFP signal (SupplementaryFig. S6C and S6D). We also confirmed the depletion level ofXPF mRNA by reverse transcription (RT)-PCR (Supplemen-tary Fig. S6E). We generated XPFþ/�GdXPF-loxP clones, and
subsequently generated 2 XPF�/�GdXPF-loxP clones, whichdisplayed indistinguishable phenotypes.
We analyzed the proliferation kinetics of the cells with andwithout treatment with tamoxifen (Fig. 1B andC). TheXPFþ/þ,XPFþ/�GdXPF-loxP, and XPF�/�GdXPF-loxP cells without thetamoxifen treatment divided every 8 hours (SupplementaryFig. S6F). By contrast, at approximately 2 days after treatmentwith tamoxifen, the XPF�/� cells ceased to proliferate (Fig. 1Band C). To explore the cause of this mortality, we examinedspontaneously arising chromosomal aberrations in mitoticcells. It should be noted that chromosomal breaks, hereafter,represent discontinuities, which appear on chromosomes inmetaphase spreads as regions unstained by Giemsa and do notalways result from DSBs. We found a dramatic increase in thenumber of chromosomal aberrations prior to cell death in theXpf-deficient cells (Fig. 1D), an occurrence that is consistentlyobserved in cells that have a severe defect inHR (26, 32–34), but
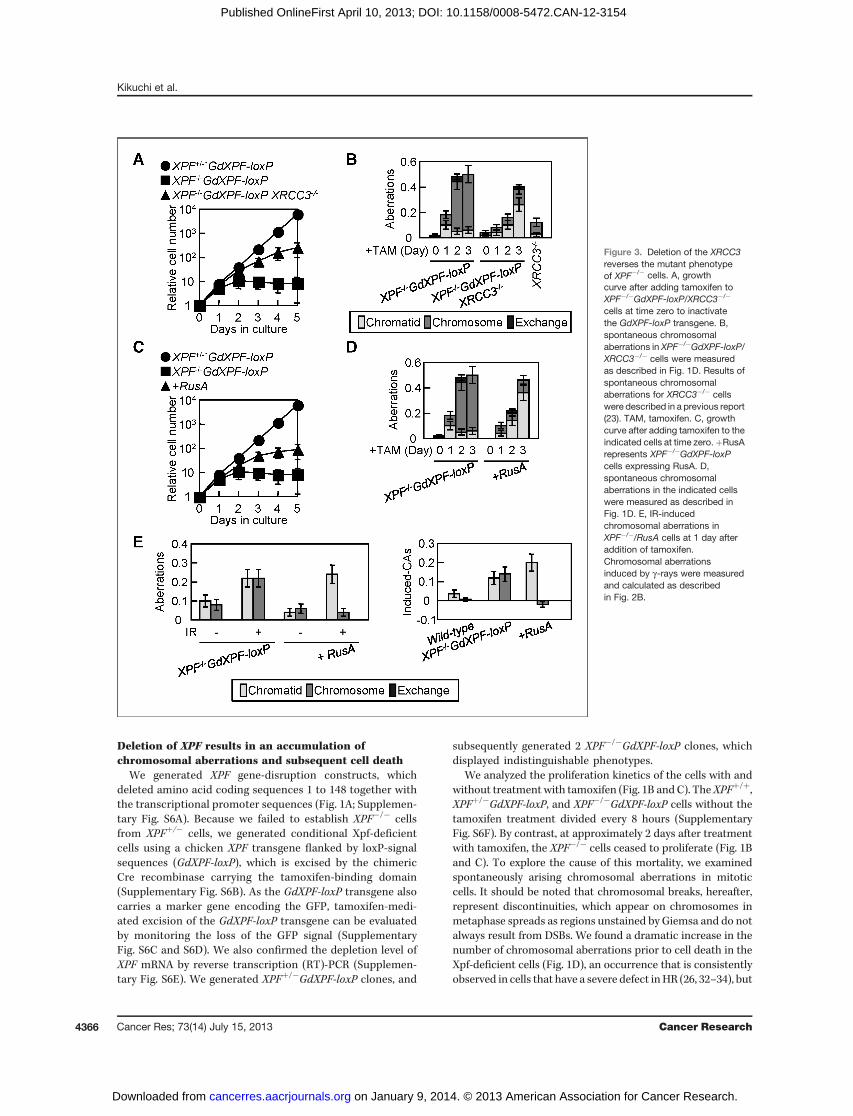
Figure 3. Deletion of the XRCC3reverses the mutant phenotypeof XPF�/� cells. A, growthcurve after adding tamoxifen toXPF�/�GdXPF-loxP/XRCC3�/�
cells at time zero to inactivatethe GdXPF-loxP transgene. B,spontaneous chromosomalaberrations in XPF�/�GdXPF-loxP/XRCC3�/� cells were measuredas described in Fig. 1D. Results ofspontaneous chromosomalaberrations for XRCC3�/� cellswere described in a previous report(23). TAM, tamoxifen. C, growthcurve after adding tamoxifen to theindicated cells at time zero.þRusArepresents XPF�/�GdXPF-loxPcells expressing RusA. D,spontaneous chromosomalaberrations in the indicated cellswere measured as described inFig. 1D. E, IR-inducedchromosomal aberrations inXPF�/�/RusA cells at 1 day afteraddition of tamoxifen.Chromosomal aberrationsinduced by g-rays were measuredand calculated as describedin Fig. 2B.
Kikuchi et al.
Cancer Res; 73(14) July 15, 2013 Cancer Research4366
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154
is not observed in DT40 cells deficient in the XPA or XPGgenes involved in NER (35, 36). To test whether the nucleaseactivity of Xpf is required for cellular viability, we created 2cDNAs mutated at the catalytic center of Xpf, GdXPF(D674A)and GdXPF(D702A) [Supplementary Figs. S1 and S6G]. Incontrast to a wild-type Xpf transgene, thesemutant transgenesyielded no stable clones (Supplementary Fig. S6G), suggestingthat the mutant Xpf proteins may interfere with the endo-genous Xpf by competing for the association with Ercc1.
HR-mediated DSB repair is severely compromised inXpf-deficient cellsTo analyze the role of Xpf inHR,we assessed the capability of
HR-mediated DSB repair at 24 hours after tamoxifen treat-ment, when the cells were still able to proliferate exponentially.To selectively evaluate HR-mediated DSB repair, we measuredthe number of chromosomal aberrations in mitotic cellsfollowing exposure of cells to camptothecin, a DNA topoisom-erase I poison, and to g-rays in the G2 phase. Camptothecininduces single-end breaks during replication, and the restart ofreplication requires HR with the intact sister chromatid (37).Similarly, g-ray–induced chromosomal aberrations arerepaired exclusively by HR in the G2 phase in DT40 cells (38).Xpf-depleted cells had a greater number of camptothecin-
induced chromosomal aberrations than did Xpf-expressingcells (Fig. 2A). We next exposed an asynchronous populationof cells to g-rays and measured the number of chromosomalaberrations in cells that entered theMphasewithin 3 hours afterirradiation. This protocol allows for the evaluation of DSB repair
selectively during the G2 phase, where HR plays a dominantrole in DSB repair (38). Following irradiation, the total numberof chromosomal aberrations in Xpf-deficient cells increasedto 0.26 per cell, whereas the total number in XPFþ/þ cells in-creased only to 0.08 (Fig. 2B). Taken together, these resultsindicate that Xpf plays a key role in HR-mediated DSB repair.
HR-mediated DSB repair in XPF-mutant cells iscompromised at a late step
To differentiate between the early and late steps of HR,we analyzed the formation of g-ray–induced Rad51 foci(Fig. 2C). At 3 hours after IR, there was no change in the rateof Rad51 focus formation for Xpf-expressing or Xpf-depletedcells. However, the Rad51 foci continued to be formed by 6hours in Xpf-depleted cells, whereas the foci were decreasedin Xpf-expressing cells. This suggests that Xpf is required forthe completion of HR after formation of the Rad51 nucleofila-ment. To assess the role of Xpf in the late steps of HR, that is,during jointmolecules formation andprocessing, wemeasuredthe number of SCEs, which represent crossover-type HR (27).The number of camptothecin-induced SCEs in Xpf-depletedcells was only 40% of the number in Xpf-expressing cells(Fig. 2D), indicating that Xpf is required for crossover-type HR.
If the mortality of the XPF�/� cells is caused by impairedcompletion of HR, it might be reversed by blocking theinitiation of HR. To test this hypothesis, we disrupted theXRCC3 gene in XPF�/�GdXPF-loxP cells. Xrcc3 facilitates aninitial step of HR by promoting the polymerization of Rad51at DNA lesions (39). To generate XPF�/�/XRCC3�/� cells, we
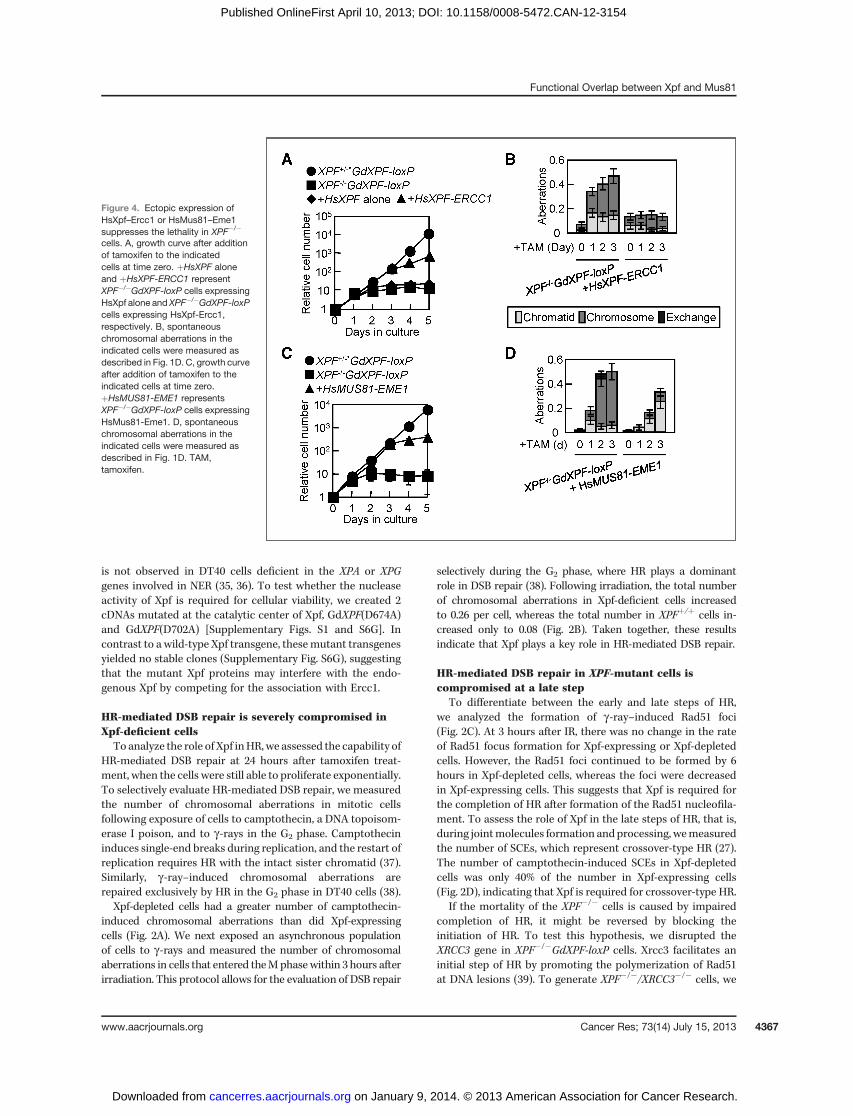
Figure 4. Ectopic expression ofHsXpf–Ercc1 or HsMus81–Eme1suppresses the lethality in XPF�/�
cells. A, growth curve after additionof tamoxifen to the indicatedcells at time zero. þHsXPF aloneand þHsXPF-ERCC1 representXPF�/�GdXPF-loxP cells expressingHsXpf alone andXPF�/�GdXPF-loxPcells expressing HsXpf-Ercc1,respectively. B, spontaneouschromosomal aberrations in theindicated cells were measured asdescribed in Fig. 1D. C, growth curveafter addition of tamoxifen to theindicated cells at time zero.þHsMUS81-EME1 representsXPF�/�GdXPF-loxP cells expressingHsMus81-Eme1. D, spontaneouschromosomal aberrations in theindicated cells were measured asdescribed in Fig. 1D. TAM,tamoxifen.
Functional Overlap between Xpf and Mus81
www.aacrjournals.org Cancer Res; 73(14) July 15, 2013 4367
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154
exposed the resulting XPF�/�GdXPF-loxP/XRCC3�/� cells totamoxifen. Thismutant displayed lower levels of cell death (Fig.3A) and a significant decrease in the number of chromosomalaberrations (Fig. 3B), compared with Xpf-depleted cells,although the XRCC3�/� cells did display moderate genomicinstability (23, 39). These observations support the conclusionthat Xpf plays a critical role in the completion of HR-mediatedrepair after the polymerization of Rad51 at DNA lesions.
We next considered that if Xpf contributes to HR after theformation of joint molecules, the severe phenotype of Xpf-depleted cells might be suppressed by ectopic expression of E.coli RusA resolvase (40). To generate XPF�/� cells stablyexpressing RusA, XPF�/�GdXPF-loxP cells were transfectedwith RusA cDNA, and the resulting XPF�/�GdXPF-loxP/RusAcells were exposed to tamoxifen. The RusA expressionimproved cellular viability (Fig. 3C) and decreased the numberof spontaneous chromosomal aberrations in Xpf-depletedcells (Fig. 3D). Intriguingly, the reduction in chromosome-
type breaks is associated with an increase in chromatid-typebreaks. Similarly RusA expression resulted in a decrease ing-ray–induced chromosome-type breaks without affecting thenumber of chromatid-type breaks (Fig. 3E). These data areconsistent with the idea that chromosome-type breaks resultfrom a failure to complete HR after the formation of jointmolecules in Xpf-deficient cells.
HumanMus81–Eme1 and Xpf–Ercc1 contribute to HR byprocessing joint molecules
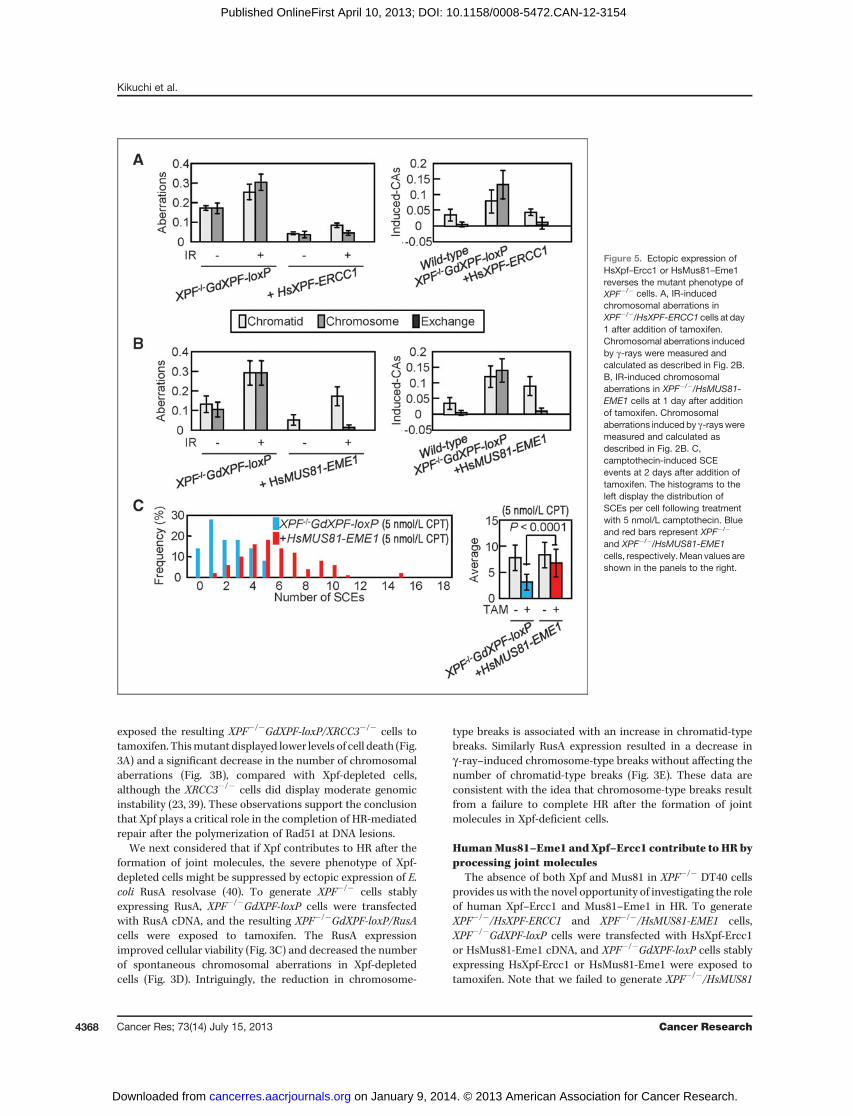
The absence of both Xpf and Mus81 in XPF�/� DT40 cellsprovides us with the novel opportunity of investigating the roleof human Xpf–Ercc1 and Mus81–Eme1 in HR. To generateXPF�/�/HsXPF-ERCC1 and XPF�/�/HsMUS81-EME1 cells,XPF�/�GdXPF-loxP cells were transfected with HsXpf-Ercc1or HsMus81-Eme1 cDNA, and XPF�/�GdXPF-loxP cells stablyexpressing HsXpf-Ercc1 or HsMus81-Eme1 were exposed totamoxifen. Note that we failed to generate XPF�/�/HsMUS81
Figure 5. Ectopic expression ofHsXpf–Ercc1 or HsMus81–Eme1reverses the mutant phenotype ofXPF�/� cells. A, IR-inducedchromosomal aberrations inXPF�/�/HsXPF-ERCC1 cells at day1 after addition of tamoxifen.Chromosomal aberrations inducedby g-rays were measured andcalculated as described in Fig. 2B.B, IR-induced chromosomalaberrations in XPF�/�/HsMUS81-EME1 cells at 1 day after additionof tamoxifen. Chromosomalaberrations induced by g-raysweremeasured and calculated asdescribed in Fig. 2B. C,camptothecin-induced SCEevents at 2 days after addition oftamoxifen. The histograms to theleft display the distribution ofSCEs per cell following treatmentwith 5 nmol/L camptothecin. Blueand red bars represent XPF�/�
and XPF�/�/HsMUS81-EME1cells, respectively.Mean values areshown in the panels to the right.
Kikuchi et al.
Cancer Res; 73(14) July 15, 2013 Cancer Research4368
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154
cells, probably due to instability of HsMus81 as a conse-quence of poor association with GdEme1. HsXpf-Ercc1reversed the mortality in XPF�/�DT40 cells (Fig. 4A). Remark-ably, HsMus81-Eme1 also significantly restored the cellularproliferation of XPF�/� DT40 cells to a level comparablewith the cells complemented with HsXpf-Ercc1 (Fig. 4C),although the amino acid sequence identity betweenGdXpf and HsMus81 is only 9.7%. Next, we analyzedXPF�/�/HsXPF-ERCC1 and XPF�/�/HsMUS81-EME1 cells bycounting both spontaneous (Fig. 4B and D) and g-ray–induced chromosomal aberrations (Fig. 5A and B). In allcases, the ectopic expression of the human enzymes sup-pressed the chromosomal aberrations caused by Xpf defi-ciency and, particularly, chromosome-type breaks. Theseobservations indicate that HsMus81–Eme1 and HsXpf–Ercc1 have very similar functions in HR-mediated DNArepair and genome maintenance.To determinewhetherHsMus81–Eme1 promotes crossover-
type HR, we counted the number of SCEs in XPF�/�/HsMUS81-EME1 cells. The expression of HsMus81–Eme1 restored thenumber of camptothecin-induced SCEs (Fig. 5C) to the num-ber of SCEs in GdXpf-expressing cells (Fig. 2D). We concludethat Mus81 and Xpf have very similar functions in promotingcrossover-type HR.Finally, we tested whether Xpf and Mus81 compensate for
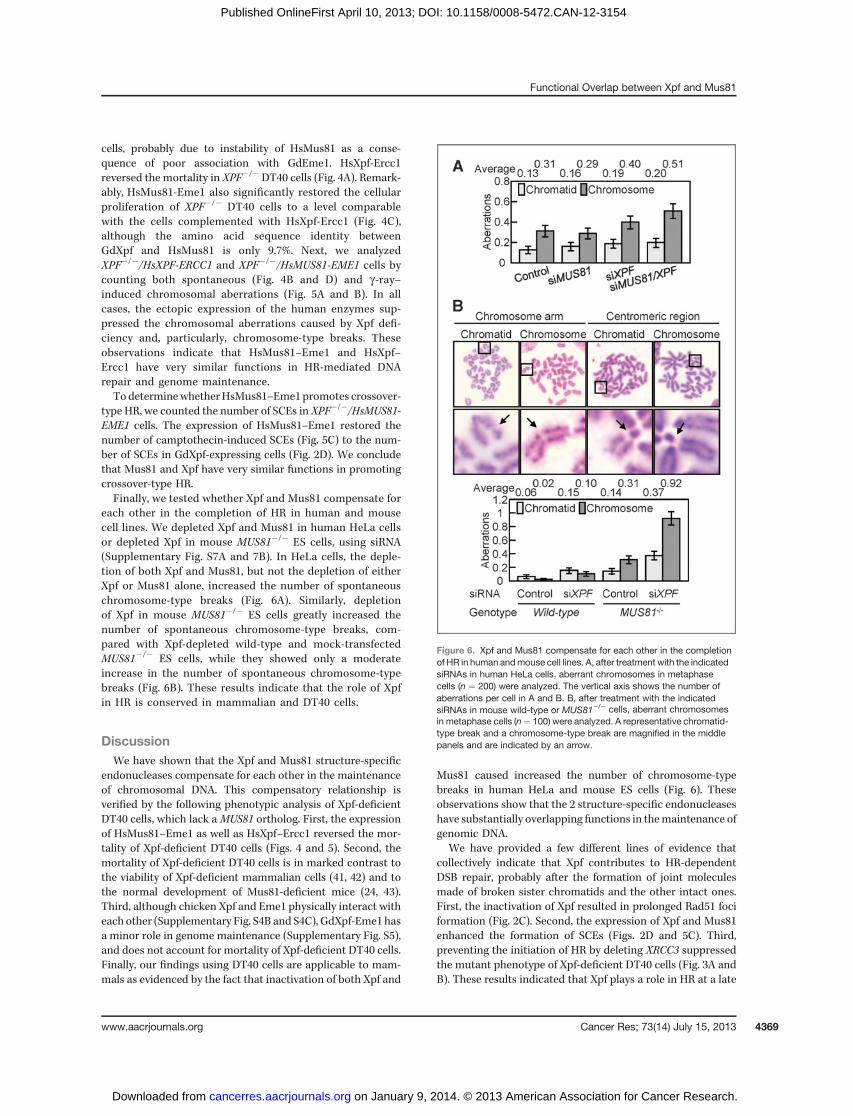
each other in the completion of HR in human and mousecell lines. We depleted Xpf and Mus81 in human HeLa cellsor depleted Xpf in mouse MUS81�/� ES cells, using siRNA(Supplementary Fig. S7A and 7B). In HeLa cells, the deple-tion of both Xpf and Mus81, but not the depletion of eitherXpf or Mus81 alone, increased the number of spontaneouschromosome-type breaks (Fig. 6A). Similarly, depletionof Xpf in mouse MUS81�/� ES cells greatly increased thenumber of spontaneous chromosome-type breaks, com-pared with Xpf-depleted wild-type and mock-transfectedMUS81�/� ES cells, while they showed only a moderateincrease in the number of spontaneous chromosome-typebreaks (Fig. 6B). These results indicate that the role of Xpfin HR is conserved in mammalian and DT40 cells.
DiscussionWe have shown that the Xpf and Mus81 structure-specific
endonucleases compensate for each other in the maintenanceof chromosomal DNA. This compensatory relationship isverified by the following phenotypic analysis of Xpf-deficientDT40 cells, which lack aMUS81 ortholog. First, the expressionof HsMus81–Eme1 as well as HsXpf–Ercc1 reversed the mor-tality of Xpf-deficient DT40 cells (Figs. 4 and 5). Second, themortality of Xpf-deficient DT40 cells is in marked contrast tothe viability of Xpf-deficient mammalian cells (41, 42) and tothe normal development of Mus81-deficient mice (24, 43).Third, although chicken Xpf and Eme1 physically interact witheach other (Supplementary Fig. S4B and S4C), GdXpf-Eme1 hasa minor role in genome maintenance (Supplementary Fig. S5),and does not account for mortality of Xpf-deficient DT40 cells.Finally, our findings using DT40 cells are applicable to mam-mals as evidenced by the fact that inactivation of both Xpf and
Mus81 caused increased the number of chromosome-typebreaks in human HeLa and mouse ES cells (Fig. 6). Theseobservations show that the 2 structure-specific endonucleaseshave substantially overlapping functions in themaintenance ofgenomic DNA.
We have provided a few different lines of evidence thatcollectively indicate that Xpf contributes to HR-dependentDSB repair, probably after the formation of joint moleculesmade of broken sister chromatids and the other intact ones.First, the inactivation of Xpf resulted in prolonged Rad51 fociformation (Fig. 2C). Second, the expression of Xpf and Mus81enhanced the formation of SCEs (Figs. 2D and 5C). Third,preventing the initiation of HR by deleting XRCC3 suppressedthe mutant phenotype of Xpf-deficient DT40 cells (Fig. 3A andB). These results indicated that Xpf plays a role in HR at a late
Figure 6. Xpf and Mus81 compensate for each other in the completionof HR in human andmouse cell lines. A, after treatment with the indicatedsiRNAs in human HeLa cells, aberrant chromosomes in metaphasecells (n ¼ 200) were analyzed. The vertical axis shows the number ofaberrations per cell in A and B. B, after treatment with the indicatedsiRNAs in mouse wild-type or MUS81�/� cells, aberrant chromosomesin metaphase cells (n¼ 100) were analyzed. A representative chromatid-type break and a chromosome-type break are magnified in the middlepanels and are indicated by an arrow.
Functional Overlap between Xpf and Mus81
www.aacrjournals.org Cancer Res; 73(14) July 15, 2013 4369
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154

step. Conceivably, the inactivation of Xpf may cause a defect inthe separation of joint molecules before mitosis that is moretoxic than the defective formation of jointmolecules due to theabsence of XRCC3.
The loss of viability in Xpf-deficient DT40 cells is associatedwith chromosome aberrations, which appear on metaphasespreads as regions unstained by Giemsa. Throughout, we havereferred to these as chomatid-type breaks and chromosome-type breaks depending onwhether the discontinuity in stainingaffects 1 or both sister chromatids, respectively. In the case ofchromatid-type breaks, we suspect that the absence of Giemsastaining actually represents a DSB in the sister chromatid asthese aberrations are also observed in mutants that are defec-tive for early steps of HR (44). However, we think that thechromosome-type breaks represent regions of uncondensedDNA rather than actual broken chromosomes similar to whatwas recently reported (45). It is proposed that regions devoid ofGiemsa staining occur at sites of sister chromatid entangle-ment, which inhibits chromosome condensation. Importantly,the chromosome-type breaks that we observe in Xpf-deficientcells both spontaneously and following g-ray exposure aresuppressed by ectopic expression of the HJ resolvase RusA(Fig. 3D and E), providing strong evidence that they result fromunprocessed joint molecules. The fact that ectopic expressionof HsXpf–Ercc1 and HsMus81–Eme1 also suppress chromo-some-type breaks in Xpf-deficient DT40 cells suggests thatthere is a significant overlap in function between theseenzymes that is most likely related to processing joint mole-cules such as D-loops and nicked single HJs.
Cells deficient in Xpf–Ercc1 are considerably more sensitiveto chemical cross-linking agents than cells deficient in theother NER factors, indicating the critical role for Xpf–Ercc1 ininterstrand cross-link (ICL) repair. Although there is compel-ling evidence that the critical role played by Xpf is carried outby introducing single-strand breaks at cross-links to initiateICL repair (20, 46), Xpf may function in ICL repair also byfacilitating HR. We recently found that Slx4 links the FA-dependent ICL pathway with both Mus81 and Xpf, as Slx4serves as docking sites for the 2 nucleases and the ubiquiti-nated FancD2 protein, which agrees with recent reports show-ing FA patients carrying mutations in the SLX4 gene (47–49).Taking into account the fact that Slx4 also binds to the Slx1 50-
flap endonuclease in addition to the Mus81–Eme1 and Xpf–Ercc1 30-flap endonucleases (Supplementary Fig. S2A and S4D;refs. 12–15), these endonucleases may collaboratively work inICL repair by promoting the completion of HR. In this scenario,it is not surprising that the 2 30-flap endonucleases are com-plementary to each other. Future studies will analyze theinterdependent and complementary relationships of multipleendonucleases when they carry out a variety of DNA repairreactions.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: K. Kikuchi, R. Kanaar, Y. Taniguchi, S. TakedaDevelopment of methodology: K. Kikuchi, M.C. WhitbyAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): K. Kikuchi, T. Narita, J. Iijima, K. Hirota, I.S. KeKa,M. Mohiuddin, K. Okawa, T. Fukagawa, J. Essers, Y. TaniguchiAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): K. Kikuchi, T. Narita, V.T. Pham, I.S. KeKa,M. Mohiuddin, K. Okawa, R. Kanaar, S. TakedaWriting, review, and/or revision of the manuscript: K. Kikuchi, T. Narita,V.T. Pham, R. Kanaar, M.C. Whitby, S. TakedaAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): K. Kikuchi, T. Hori, K. Sugasawa,S. TakedaStudy supervision: K. Kikuchi, R. Kanaar, Y. Taniguchi, K. Kitagawa, S. Takeda
AcknowledgmentsThe authors thank R. Ohta and Y. Satoh for their technical assistance. The
authors also thank Dr. M. Lisby for sharing unpublished data.
Grant SupportFinancial support was provided in part by a grant-in-aid from the Ministry of
Education, Culture, Sports, Science and Technology of the Japanese Government(MEXT; grant no. 20241012 to S. T. and grant no. 22700881 to K. Kikuchi), a grantfrom The Kanae Foundation (K. Kikuchi), grants from the European Commu-nity's Seventh Framework Programme (FP7/2007–2013) under grant agreementno. HEALTH-F2-2010-259893, and from the Netherlands Genomics Initiative/Netherlands Organization for Scientific Research (R. Kanaar), and NIH grantsGM68418 and CA133093 (K. Kitagawa). Funding was also provided by TheWellcome Trust (090767/Z/09/Z; M.C. Whitby).
The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received August 9, 2012; revised February 6, 2013; accepted March 17, 2013;published OnlineFirst April 10, 2013.
References1. Takeda S, Nakamura K, Taniguchi Y, Paull TT. Ctp1/CtIP and theMRN
complex collaborate in the initial steps of homologous recombination.Mol Cell 2007;28:351–2.
2. Bernstein KA, Rothstein R. At loose ends: resecting a double-strandbreak. Cell 2009;137:807–10.
3. Bzymek M, Thayer NH, Oh SD, Kleckner N, Hunter N. Double Hollidayjunctions are intermediates of DNA break repair. Nature 2010;464:937–41.
4. WuLaH ID. The Bloom's syndrome helicase suppresses crossing overduring homologous recombination. Nature 2003;426:870–4.
5. Xu D, Guo R, Sobeck A, Bachrati CZ, Yang J, Enomoto T, et al. RMI, anew OB-fold complex essential for Bloom syndrome protein to main-tain genome stability. Genes Dev 2008;22:2843–55.
6. Chu WK, Hickson ID. RecQ helicases: multifunctional genome care-takers. Nat Rev Cancer 2009;9:644–54.
7. Schwartz E, Heyer WD. Processing of joint molecule intermediates bystructure-selective endonucleases during homologous recombinationin eukaryotes. Chromosoma 2011;120:109–27.
8. Macmaster R, SedelnikovaS, Baker PJ, Bolt EL, LloydRG, Rafferty JB.RusA Holliday junction resolvase: DNA complex structure—insightsinto selectivity and specificity. Nucleic Acids Res 2006;34:5577–84.
9. Shinagawa H, Iwasaki H. Processing the Holliday junction in homol-ogous recombination. Trends Biochem Sci 1996;21:107–11.
10. West SC. Processing of recombination intermediates by the RuvABCproteins. Annu Rev Genet 1997;31:213–44.
11. Ip SC, Rass U, Blanco MG, Flynn HR, Skehel JM, West SC. Identifi-cation of Holliday junction resolvases from humans and yeast. Nature2008;456:357–61.
12. Andersen SL, Bergstralh DT, Kohl KP, LaRocque JR, Moore CB,Sekelsky J. Drosophila MUS312 and the vertebrate ortholog BTBD12
Kikuchi et al.
Cancer Res; 73(14) July 15, 2013 Cancer Research4370
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154

interact with DNA structure-specific endonucleases in DNA repair andrecombination. Mol Cell 2009;35:128–35.
13. Fekairi S, ScaglioneS,ChahwanC, Taylor ER, Tissier A,CoulonS, et al.Human SLX4 is a Holliday junction resolvase subunit that bindsmultiple DNA repair/recombination endonucleases. Cell 2009;138:78–89.
14. Munoz IM,HainK,DeclaisAC,GardinerM, TohGW,Sanchez-Pulido L,et al. Coordination of structure-specific nucleases by human SLX4/BTBD12 is required for DNA repair. Mol Cell 2009;35:116–27.
15. Svendsen JM, Smogorzewska A, Sowa ME, O'Connell BC, Gygi SP,Elledge SJ, et al. Mammalian BTBD12/SLX4 assembles a Hollidayjunction resolvase and is required for DNA repair. Cell 2009;138:63–77.
16. Osman F, Dixon J, Doe CL, Whitby MC. Generating crossovers byresolution of nicked Holliday junctions: a role for Mus81-Eme1 inmeiosis. Mol Cell 2003;12:761–74.
17. Holloway JK, Booth J, EdelmannW,McGowanCH,CohenPE.MUS81generates a subset of MLH1-MLH3-independent crossovers in mam-malian meiosis. PLoS Genet 2008;4:e1000186.
18. Niedernhofer LJ, Odijk H, Budzowska M, van Drunen E, Alex Maas A,Theil AF, et al. The structure-specific endonuclease Ercc1-Xpf isrequired to resolve DNA interstrand cross-link-induced double-strandbreaks. Mol Cell Biol 2004;24:5776–87.
19. Paques F, Haber JE. Multiple pathways of recombination induced bydouble-strand breaks inSaccharomyces cerevisiae.MicrobiolMol BiolRev 1999;63:349–404.
20. Knipscheer P, R€aschle M, Smogorzewska A, Enoiu M, Ho TV, Sch€arerOD, et al. The Fanconi anemia pathway promotes replication-depen-dent DNA interstrand cross-link repair. Science 2009;326:1698–701.
21. Niedernhofer LJ, Essers J,WeedaG, Beverloo B, deWit J,MuijtjensM,et al. The structure-specific endonuclease Ercc1-Xpf is required fortargeted gene replacement in embryonic stem cells. EMBO J 2001;20:6540–9.
22. Buerstedde JM, Takeda S. Increased ratio of targeted to randomintegration after transfection of chicken B cell lines. Cell 1991;67:179–88.
23. Fujimori A, Tachiiri S, Sonoda E, Thompson LH, Dhar PK, Hiraoka M,et al. Rad52 partially substitutes for the Rad51 paralog XRCC3 inmaintaining chromosomal integrity in vertebrate cells. EMBO J 2001;20:5513–20.
24. McPherson JP, LemmersB,ChahwanR, Pamidi A,Migon E,Matysiak-Zablocki E, et al. InvolvementofmammalianMus81 in genome integrityand tumor suppression. Science 2004;304:1822–6.
25. Motegi A, SoodR,MoinovaH,Markowitz SD, Liu PP,MyungK.HumanSHPRH suppresses genomic instability through proliferating cellnuclear antigen polyubiquitination. J Cell Biol 2006;175:703–8.
26. Yamaguchi-Iwai Y, Sonoda E, Sasaki MS, Morrison C, Haraguchi T,Hiraoka Y, et al. Mre11 is essential for the maintenance of chromo-somal DNA in vertebrate cells. EMBO J 1999;18:6619–29.
27. Sonoda E, Sasaki MS, Morrison C, Yamaguchi-Iwai Y, Takata M,Takeda S. Sister chromatid exchanges are mediated by homologousrecombination in vertebrate cells. Mol Cell Biol 1999;19:5166–9.
28. http://genome.ucsc.edu/cgi-bin/hgBlat29. Abraham J, Lemmers B, Hande MP, Moynahan ME, Chahwan C,
Ciccia A, et al. Eme1 is involved in DNA damage processing andmaintenance of genomic stability in mammalian cells. EMBOJ 2003;22:6137–47.
30. Hanada K, Budzowska M, Modesti M, Maas A, Wyman C, Essers J,et al. The structure-specific endonuclease Mus81-Eme1 promotesconversion of interstrand DNA crosslinks into double-strands breaks.EMBO J 2006;25:4921–32.
31. Ciccia A, Ling C, Coulthard R, Yan Z, Xue Y, Meetei AR, et al.Identification of FAAP24, a Fanconi anemia core complex protein thatinteracts with FANCM. Mol Cell 2007;25:331–43.
32. Sonoda E, Sasaki MS, Buerstedde JM, Bezzubova O, Shinohara A,Ogawa H, et al. Rad51-deficient vertebrate cells accumulatechromosomal breaks prior to cell death. EMBO J 1998;17:598–608.
33. NakaharaM,SonodaE,NojimaK,Sale JE, TakenakaK,Kikuchi K, et al.Genetic evidence for single-strand lesions initiating Nbs1-dependenthomologous recombination in diversification of Ig v in chicken Blymphocytes. PLoS Genet 2009;5:e1000356.
34. NakamuraK, KogameT,OshiumiH, ShinoharaA, SumitomoY, AgamaK, et al. Collaborative action of Brca1 and CtIP in elimination ofcovalent modifications from double-strand breaks to facilitate subse-quent break repair. PLoS Genet 2010;6:e1000828.
35. Okada T, Sonoda E, Yamashita YM, Koyoshi S, Tateishi S, YamaizumiM, et al. Involvement of vertebrate polkappa in Rad18-independentpostreplication repair of UV damage. J Biol Chem 2002;277:48690–5.
36. Kikuchi K, Taniguchi Y, Hatanaka A, Sonoda E, Hochegger H, AdachiN, et al. Fen-1 facilitates homologous recombination by removingdivergent sequences at DNA break ends. Mol Cell Biol 2005;25:6948–55.
37. Pommier Y, Leo E, Zhang H, Marchand C. DNA topoisomerases andtheir poisoning by anticancer and antibacterial drugs. Chem Biol2010;17:421–33.
38. Sonoda E, Hochegger H, Saberi A, Taniguchi Y, Takeda S. Differentialusage of non-homologous end-joining and homologous recombina-tion in double strand break repair. DNA Repair 2006;5:1021–9.
39. Takata M, Sasaki MS, Tachiiri S, Fukushima T, Sonoda E, Schild D,et al. Chromosome instability and defective recombinational repair inknockout mutants of the five Rad51 paralog. Mol Cell Biol 2001;21:2858–66.
40. Blais V, GaoH, Elwell CA, BoddyMN,Gaillard PH, Russell P, et al. RNAinterference inhibition of Mus81 reduces mitotic recombination inhuman cells. Mol Biol Cell 2004;15:552–62.
41. Brookman KW, Lamerdin JE, Thelen MP, Hwang M, Reardon JT,Sancar A, et al. ERCC4 (XPF) encodes a human nucleotide excisionrepair protein with eukaryotic recombination homologs. Mol Cell Biol1996;16:6553–62.
42. Tian M, Shinkura R, Shinkura N, Alt FW. Growth retardation, earlydeath, and DNA repair defects in mice deficient for the nucleotideexcision repair enzyme XPF. Mol Cell Biol 2004;24:1200–5.
43. Dendouga N, Gao H,Moechars D, Janicot M, Vialard J, McGowan CH.Disruption of murine Mus81 increases genomic instability and DNAdamage sensitivity but does not promote tumorigenesis. Mol Cell Biol2005;25:7569–79.
44. Sonoda E, Okada T, Zhao GY, Tateishi S, Araki K, Yamaizumi M, et al.Multiple roles of Rev3, the catalytic subunit of polzeta in maintaininggenome stability in vertebrates. EMBO J 2003;22:3188–97.
45. Wechsler T, Newman S, West SC. Aberrant chromosome morphologyin human cells defective for Holliday junction resolution. Nature2011;471:642–6.
46. Wang AT, Sengerov�a B, Cattell E, Inagawa T, Hartley JM, Kiakos K,et al. Human SNM1A and XPF-ERCC1 collaborate to initiate DNAinterstrand cross-link repair. Genes Dev 2011;25:1859–70.
47. Crossan GP, van der Weyden L, Rosado IV, Langevin F, Gaillard PH,McIntyre RE, et al. Disruption of mouse Slx4, a regulator of structure-specific nucleases, phenocopies Fanconi anemia. Nat Genet 2011;43:147–52.
48. Kim Y, Lach FP, Desetty R, Hanenberg H, Auerbach AD, Smogor-zewska A. Mutations of the SLX4 gene in Fanconi anemia. Nat Genet2011;43:142–6.
49. Stoepker C, Hain K, Schuster B, Hilhorst-Hofstee Y, Rooimans MA,Steltenpool J, et al. SLX4, a coordinator of structure-specific endo-nucleases, is mutated in a new Fanconi anemia subtype. Nat Genet2011;43:138–41.
Functional Overlap between Xpf and Mus81
www.aacrjournals.org Cancer Res; 73(14) July 15, 2013 4371
on January 9, 2014. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2013; DOI: 10.1158/0008-5472.CAN-12-3154
Related Documents











