Structure of the Pyrimidine-rich Internal Loop in the Poliovirus 3 0 -UTR: The Importance of Maintaining Pseudo-2-fold Symmetry in RNA Helices Containing Two Adjacent Non-canonical Base-pairs Eveline M. H. P. Lescrinier 1 †, Marco Tessari 1 Frank J. M. van Kuppeveld 2 , Willem J. G. Melchers 2 Cornelis W. Hilbers 1 and Hans A. Heus 1 * 1 NSR Center for Molecular Structure, Design and Synthesis, Laboratory of Biophysical Chemistry University of Nijmegen Toernooiveld 1, 6525 ED Nijmegen, The Netherlands 2 Department of Medical Microbiology, Nijmegen Center for Molecular Life Sciences University Medical Center P.O. Box 9101, 6500 HB Nijmegen, The Netherlands Formation of non-canonical base-pairs in RNA often plays a very import- ant functional role. In addition they frequently serve as factors in stabiliz- ing the secondary structure elements that provide the frame of large compact RNA structures. Here we describe the structure of an internal loop containing a 5 0 CU3 0 /5 0 UU3 0 non-canonical tandem base-pair motif, which is conserved within the 3 0 -UTR of poliovirus-like enteroviruses. Structural details reveal striking regularities of the local helix geometry, resulting from alternating geometrical adjustments, which are important for understanding and predicting stabilities and configurations of tandem non-canonical base-pairs. The C-U and U-U base-pairs severely contract the minor groove of the sugar-phosphate backbone, which might be important for protein recognition or binding to other RNA elements. q 2003 Elsevier Ltd. All rights reserved. Keywords: RNA; RNA structure; RNA folding; NMR; poliovirus *Corresponding author Introduction A unique feature of RNA is its ability to form highly compact structures that accommodate many non-canonical base-pairs. Perhaps the best example is the ubiquitous U-G wobble base-pair, which was first described for the codon–anticodon interaction, where it explained the degeneracy of the genetic code. 1 Many other examples are known, like G-G base-pairs involved in RNA– protein interactions 2 and G-A and A þ -C base- pairs, structurally and functionally important in catalytic RNA. 3–6 Another important aspect of non-canonical base-pairs in RNA is that they can be polymorphic, i.e. they can adopt different base- pair configurations depending on the context. A well known example is the tandem G-A base-pair motif, which forms head to head base-pairs when flanked by G-C base-pairs 7 and sheared base-pairs when flanked by C-G or U-A base-pairs. 8,9 Due to its polymorphism it appears that for a complete understanding of RNA folding and function it is necessary to perform a separate structure determi- nation for every RNA motif containing non- canonical base-pairs in a different context. Polioviruses are small RNA viruses that possess a single-stranded RNA genome consisting of a 5 0 - untranslated region (5 0 -UTR), a single open reading frame (ORF), a 3 0 -untranslated region (3 0 -UTR), also called oriR, and a 3 0 -terminal poly(A) tract. 10 The 3 0 -UTR is a prime example of a multi-domain globular structure. Its secondary structure has been well characterized 11 – 14 and contains two hair- pins, designated domains X and Y and a X–Y loop–loop interaction, designated the K-domain (Figure 1(A)). Like other complex RNA entities of this size its tertiary structure is expected to be highly compact with many intricate interactions to fulfill its structural and functional role. Several reports have provided evidence for the importance of the higher-order structural elements in the 3 0 -UTR in viral RNA replication as manifested by their interaction with viral and cellular proteins. 15 – 17 Recently, a circular model has been 0022-2836/$ - see front matter q 2003 Elsevier Ltd. All rights reserved. † On leave from the University of Leuven, Rega Institute, Belgium. E-mail address of the corresponding author: [email protected] Abbreviations used: NOE, nuclear Overhauser effect. doi:10.1016/S0022-2836(03)00787-3 J. Mol. Biol. (2003) 331, 759–769

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Structure of the Pyrimidine-rich Internal Loop in thePoliovirus 30-UTR: The Importance of MaintainingPseudo-2-fold Symmetry in RNA Helices ContainingTwo Adjacent Non-canonical Base-pairs
Eveline M. H. P. Lescrinier1†, Marco Tessari1
Frank J. M. van Kuppeveld2, Willem J. G. Melchers2
Cornelis W. Hilbers1 and Hans A. Heus1*
1NSR Center for MolecularStructure, Design andSynthesis, Laboratory ofBiophysical ChemistryUniversity of NijmegenToernooiveld 1, 6525 EDNijmegen, The Netherlands
2Department of MedicalMicrobiology, Nijmegen Centerfor Molecular Life SciencesUniversity Medical CenterP.O. Box 9101, 6500 HBNijmegen, The Netherlands
Formation of non-canonical base-pairs in RNA often plays a very import-ant functional role. In addition they frequently serve as factors in stabiliz-ing the secondary structure elements that provide the frame of largecompact RNA structures. Here we describe the structure of an internalloop containing a 50CU30/50UU30 non-canonical tandem base-pair motif,which is conserved within the 30-UTR of poliovirus-like enteroviruses.Structural details reveal striking regularities of the local helix geometry,resulting from alternating geometrical adjustments, which are importantfor understanding and predicting stabilities and configurations of tandemnon-canonical base-pairs. The C-U and U-U base-pairs severely contractthe minor groove of the sugar-phosphate backbone, which might beimportant for protein recognition or binding to other RNA elements.
q 2003 Elsevier Ltd. All rights reserved.
Keywords: RNA; RNA structure; RNA folding; NMR; poliovirus*Corresponding author
Introduction
A unique feature of RNA is its ability to formhighly compact structures that accommodatemany non-canonical base-pairs. Perhaps the bestexample is the ubiquitous U-G wobble base-pair,which was first described for the codon–anticodoninteraction, where it explained the degeneracy ofthe genetic code.1 Many other examples areknown, like G-G base-pairs involved in RNA–protein interactions2 and G-A and Aþ-C base-pairs, structurally and functionally important incatalytic RNA.3 – 6 Another important aspect ofnon-canonical base-pairs in RNA is that they canbe polymorphic, i.e. they can adopt different base-pair configurations depending on the context. Awell known example is the tandem G-A base-pairmotif, which forms head to head base-pairs whenflanked by G-C base-pairs7 and sheared base-pairs
when flanked by C-G or U-A base-pairs.8,9 Due toits polymorphism it appears that for a completeunderstanding of RNA folding and function it isnecessary to perform a separate structure determi-nation for every RNA motif containing non-canonical base-pairs in a different context.
Polioviruses are small RNA viruses that possessa single-stranded RNA genome consisting of a 50-untranslated region (50-UTR), a single open readingframe (ORF), a 30-untranslated region (30-UTR),also called oriR, and a 30-terminal poly(A) tract.10
The 30-UTR is a prime example of a multi-domainglobular structure. Its secondary structure hasbeen well characterized11 –14 and contains two hair-pins, designated domains X and Y and a X–Yloop–loop interaction, designated the K-domain(Figure 1(A)). Like other complex RNA entities ofthis size its tertiary structure is expected to behighly compact with many intricate interactions tofulfill its structural and functional role. Severalreports have provided evidence for the importanceof the higher-order structural elements in the30-UTR in viral RNA replication as manifestedby their interaction with viral and cellularproteins.15 – 17 Recently, a circular model has been
0022-2836/$ - see front matter q 2003 Elsevier Ltd. All rights reserved.
† On leave from the University of Leuven, RegaInstitute, Belgium.
E-mail address of the corresponding author:[email protected]
Abbreviations used: NOE, nuclear Overhauser effect.
doi:10.1016/S0022-2836(03)00787-3 J. Mol. Biol. (2003) 331, 759–769

proposed for the poliovirus RNA, required for theinitiation of negative strand RNA synthesis, inwhich the 50-UTR and 30-UTR come close together,mediated by protein–protein interactions.18,19
Another important feature of large RNA mol-ecules is that the structure of the subdomains areoften maintained in the isolated forms, whichmakes their elucidation relevant for the entireRNA molecule. Alternatively, structural rearrange-ments of subdomains can occur in the sequentialRNA folding process20,21 or upon protein binding.22
Therefore the structure determination of sub-domains is also important for delineatingimportant structural rearrangements.
Here we present the solution structure of thephylogenetically conserved internal loop Y-domainof the poliovirus 30-UTR in an engineered 22-residue RNA hairpin. The internal loop is charac-terized by a C-U and U-U base-pair. The C-Ubase-pair resembles the one observed in thesolution structure of the A loop of 23 S RNA,23 butis different from other C-U base-pair geometriesreported so far. Although the sequence of theinternal loop is asymmetric, compensation of thealternating twist angles and the equal shape ofthe base-pairs convey a pseudo-2-fold symmetryto the tandem structure. The pseudo-2-fold sym-metry observed in the tandem 50CU30/30UU50
base-pair motif has important consequences forunderstanding and predicting the stabilities andstructures of RNA helices containing tandem non-canonical base-pairs in general.
Results
Resonance assignment andstructure determination
The internal loop and parts of the Y-domain
stems were investigated by NMR using the 22-merconstruct shown in Figure 1(B). All aromatic,anomeric, H20 and H30 resonances could beassigned in the non-labeled sample starting from astandard anomeric to aromatic proton walk.24
Heteronuclear NMR experiments on a 13C, 15Nselectively pyrimidine-enriched molecule, con-firmed the assignments and allowed for additionalidentification of all ribose 13C and 1H resonancesof the cytidine and uracyl residues. Residues fromthe GAAA tetraloop showed the same chemicalshifts and nuclear Overhauser effect (NOE) con-tacts as described before,25 indicative of identicalloop structures.
The structure of the RNA hairpin was solvedfrom 299 distance and 163 torsion angle restraints(see Material and Methods). Although no assump-tions or explicit hydrogen bond restraints wereintroduced to impose the C-U base-pair structure(Figure 2(C) and (E)), for which we could notdetect the imino proton signals in the spectrarecorded in H2O, in 36 out of 39 refined structures,its single imino hydrogen is within hydrogen-bonding range (C5O2–U18N3 , 3.0 A). With thedirect NMR evidence for the non-canonical U-Ubase-pair, including a strong mutual NOE betweenthe two uridine imino protons, all structures con-verged with the two U-U imino hydrogen atomswithin hydrogen-bond distance. Figure 2(A)shows the superposition of the 20 lowest energystructures, selected for structural analysis, illustrat-ing well-defined structures of the internal loop andstems.
Phylogeny of the internal loop andcharacterization of mutant virus
The poliovirus is a member of the genus ofenteroviruses, small non-enveloped RNA viruses,which also contains the coxsackie group A and B
Figure 1. Secondary structuremodels of the poliovirus 30-UTR(A) and RNA hairpin used for theNMR studies (B).
760 Structures of Tandem Non-canonical Base-pairs

viruses and ECHO viruses. Within the poliovirusesand polio-like coxsackieviruses the C-U and U-Ubase-pairs in the Y-domain of the 30-UTR arelargely conserved (eight out of ten sequences),with the exception of poliovirus 1 and coxsackieA11 that contain a U-U to U-A base-pair change(Figure 3). In nearly all the non-polio-like coxsackieand ECHO viruses the C-U and U-U base-pairs arereplaced by a U-A and a G-C Watson–Crick base-pair (seven out of nine sequences), respectively,with the exception of the ECHO 6 and 11 viruses,which contain a G-U base-pair. The conservationof the internal 50CU30/50UU30 loop motif (four outof 19 sequences) within these 30-UTRs is surpris-ingly high. For instance, a survey of 79 group Iintrons, 101 small ribosomal subunit RNAs and208 large subunit RNAs revealed only sevenexamples out of a total of 1890 tandemmismatches.26 In vitro mutagenesis was performedto alter the interior loop in the Y-stem of poliovirus,by changing the C-U and U-U base-pair into a C-Gand U-A base-pair, respectively. The viability of themutants was tested by transfecting copy RNA tran-scripts into BGM cells. Viral growth characteristicswere examined by single cycle growth curves at36 8C and 39 8C, both at which the mutant virusexhibited wild-type growth characteristics. Thesedata indicate that replacing the interior loop bytwo Watson–Crick base-pairs had no effect onvirus viability and growth in vitro.
Structure of the Y-domain
The structure of the Y-domain of the poliovirus30-UTR reveals a compact helix with a C-U andU-U base-pair in the internal loop (Figure 2). TheC5-U18 base-pair is stabilized by one hydrogenbond between the C5O2 and U18N3, while theU6-U17 base-pair is stabilized by two hydrogenbonds, U6N3-U17O2 and U6O4-U17N3. Toaccommodate the two pyrimidine–pyrimidinebase-pairs into the A-form helix the minor groovewidth, measured by the interstrand C10 –C10
distance, is significantly reduced from about 11 Ain regular A-form RNA to 8 A in the C5-U18 andU6-U17 base-pairs.
Most residues in the stem, including those in theinternal loop, display backbone torsion anglevalues typical of A-form helices, except for sometorsion angles in the stem below the internal loopthat show crankshaft rotations for a and g. For C4,G19 and U20 the rare gt conformation occurs, com-pensated by a change in a from g2 to trans, and thesugar pucker of G19, is switched from N to S-type.Although we do not have an explanation for thealtered torsion angles at the A3-U20/C4-G19 base-pair step, crankshaft rotations at this position havebeen observed before in RNA helices containingtandem U-U or U-C base-pairs as well.27,28
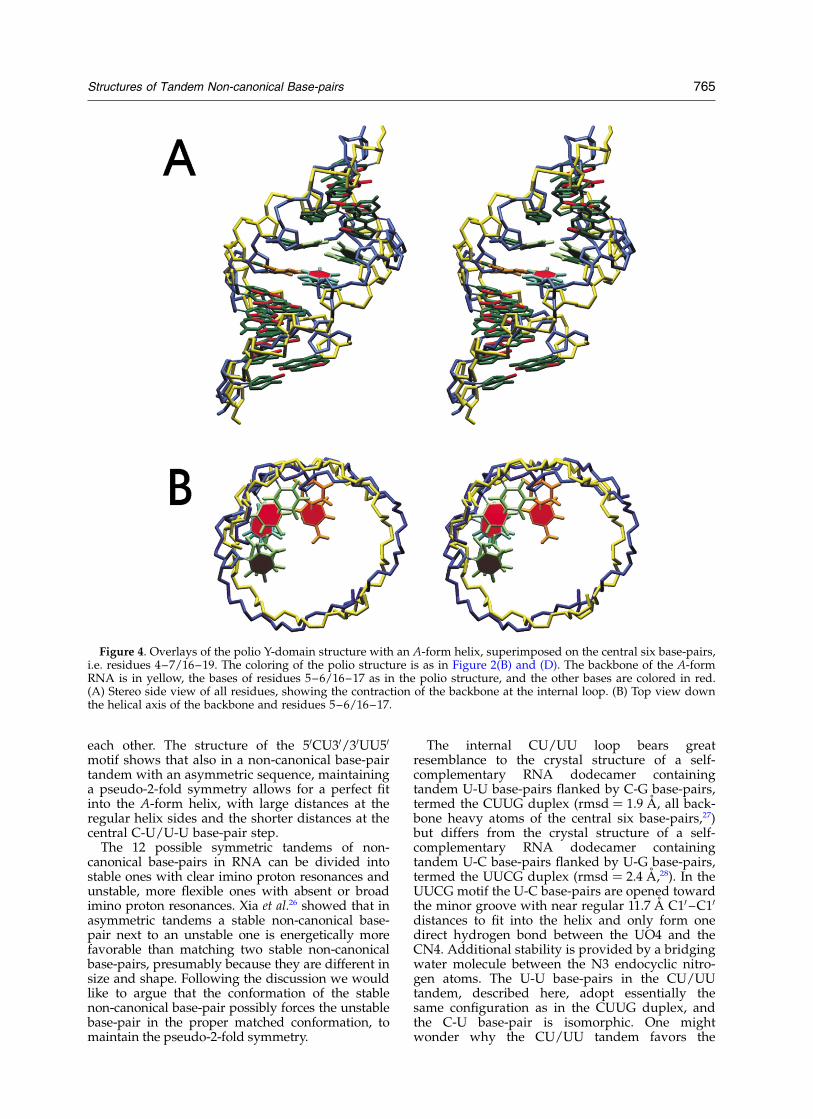
Despite the small backbone torsion angledeviations from A-form, the RNA helix does notsuperimpose well onto an A-form helix(rmsd ¼ 2 A over all heavy atoms of the central
six base-pairs, Figure 4(A)). As a consequence ofincorporating the pyrimidine base-pairs, the sugarphosphate backbones of the opposite strands arelocally moved towards each other, narrowingdown the cross-strand Pn– Pn0 þ 1 and O40n–O40 –n0 þ 2 distances, which are typical measures fordetermining the major groove width,29 to 14 A and8 A, respectively. The shortest distance across themajor groove is the U6O40 –G14O40 (n–n0 þ 1),which amounts to 5.7 A. Although the cross-strandO30 –P50 distances are also narrowed down fromthe regular 17.5 A to 14 A at the C5-U18/U6-U17base-pair step, only the side view (Figure 4(A)) ofthe polio structure superimposed on an A-formhelix reveals the backbone contraction at theinternal loop, while a top view down the helixdoes not (Figure 4(B)). This implies that the con-traction is caused by shifting the sugar-phosphatebackbones towards each other along the cylinderwall of the helix.
Discussion
Local helix geometry
Although the average helical twist over thecentral six base-pairs (338, Table 1)) comes close towhat is found in regular A-form RNA duplexes,the base-pair step between the C-U and U-U base-pair in the internal loop has an unusually highhelical twist (598), which is compensated by anunusually low helical twist of the base-pair stepson either side of the CU/UU tandem (178 and 188for the C-G/C-U and U-U/C-G base-pair step,respectively). The C-U base-pair is isomorphic tothe U-U base-pair. This can be illustrated bymeasuring the virtual bond angle, l, between theC10–C10 line and the glycosyl C10 –N1/N9 bonds.In Watson–Crick base-pairs the l angles are simi-lar, ,55–578, which together with the nearly equalC10–C10 distances makes them isosteric and createsa pseudodyad axis in both of them. This is the ori-gin of the overall regular shape of the A-formhelix with sequentially stacked bases.29 In theinternal loop the 50-residues C5 and U17 display avery large l angle of 93–948, while the 30-residuesU6 and U18 have more regular values of 488 and558, respectively. To accommodate the pyrimidinebase-pairs two major changes occur. The rotationof the bases of the 50-residues orient the hydrogenbond acceptors and donors for proper hydrogenbond formation and the 30-residues are shifted by1.5–3 A to bring the C10-atoms close together.These necessary adjustments for fitting the pyrimi-dine base-pairs into the helix break the pseudo-2-fold symmetry of the individual base-pairs. How-ever, in the tandem the C-U and U-U base-pairsare still related by a pseudo-2-fold symmetry,which maintains the overall symmetry propertiesof the helix. At the central base-pair step in thetandem the combination of the large helical twistand large l angles of the 50 C5 and U17 residues
Structures of Tandem Non-canonical Base-pairs 761
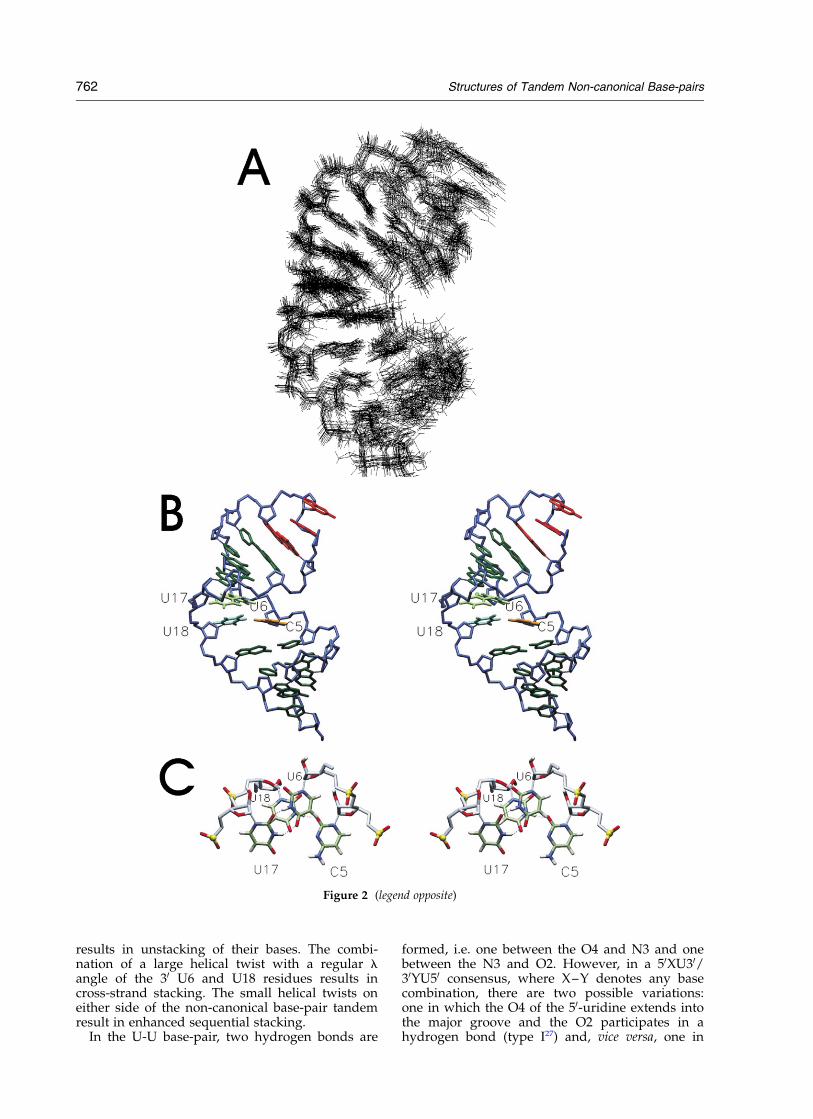
results in unstacking of their bases. The combi-nation of a large helical twist with a regular langle of the 30 U6 and U18 residues results incross-strand stacking. The small helical twists oneither side of the non-canonical base-pair tandemresult in enhanced sequential stacking.
In the U-U base-pair, two hydrogen bonds are
formed, i.e. one between the O4 and N3 and onebetween the N3 and O2. However, in a 50XU30/30YU50 consensus, where X–Y denotes any basecombination, there are two possible variations:one in which the O4 of the 50-uridine extends intothe major groove and the O2 participates in ahydrogen bond (type I27) and, vice versa, one in
Figure 2 (legend opposite)
762 Structures of Tandem Non-canonical Base-pairs
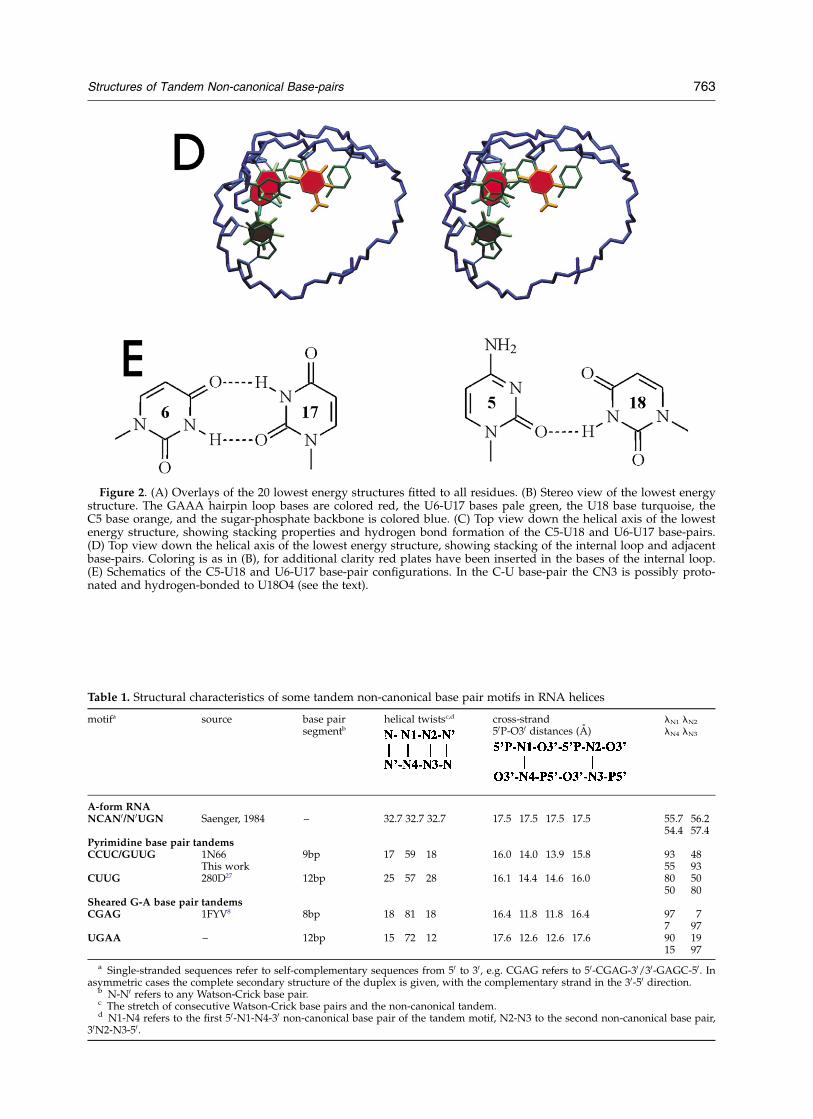
Figure 2. (A) Overlays of the 20 lowest energy structures fitted to all residues. (B) Stereo view of the lowest energystructure. The GAAA hairpin loop bases are colored red, the U6-U17 bases pale green, the U18 base turquoise, theC5 base orange, and the sugar-phosphate backbone is colored blue. (C) Top view down the helical axis of the lowestenergy structure, showing stacking properties and hydrogen bond formation of the C5-U18 and U6-U17 base-pairs.(D) Top view down the helical axis of the lowest energy structure, showing stacking of the internal loop and adjacentbase-pairs. Coloring is as in (B), for additional clarity red plates have been inserted in the bases of the internal loop.(E) Schematics of the C5-U18 and U6-U17 base-pair configurations. In the C-U base-pair the CN3 is possibly proto-nated and hydrogen-bonded to U18O4 (see the text).
Table 1. Structural characteristics of some tandem non-canonical base pair motifs in RNA helices
motifa source base pairsegmentb
helical twistsc,d cross-strand50P-O30 distances (A)
lN1 lN2
lN4 lN3
A-form RNANCAN0/N0UGN Saenger, 1984 – 32.7 32.7 32.7 17.5 17.5 17.5 17.5 55.7 56.2
54.4 57.4Pyrimidine base pair tandemsCCUC/GUUG 1N66
This work9bp 17 59 18 16.0 14.0 13.9 15.8 93 48
55 93CUUG 280D27 12bp 25 57 28 16.1 14.4 14.6 16.0 80 50
50 80Sheared G-A base pair tandemsCGAG 1FYV8 8bp 18 81 18 16.4 11.8 11.8 16.4 97 7
7 97UGAA – 12bp 15 72 12 17.6 12.6 12.6 17.6 90 19
15 97
a Single-stranded sequences refer to self-complementary sequences from 50 to 30, e.g. CGAG refers to 50-CGAG-30/30-GAGC-50. Inasymmetric cases the complete secondary structure of the duplex is given, with the complementary strand in the 30-50 direction.
b N-N0 refers to any Watson-Crick base pair.c The stretch of consecutive Watson-Crick base pairs and the non-canonical tandem.d N1-N4 refers to the first 50-N1-N4-30 non-canonical base pair of the tandem motif, N2-N3 to the second non-canonical base pair,
30N2-N3-50.
Structures of Tandem Non-canonical Base-pairs 763

which the O4 of the opposite 30-uridine extendsinto the major groove and the O4 of the 50-uridineaccepts a hydrogen bond (type II27). The U-Ubase-pair in the internal loop adopts the type IIconfiguration. Close inspection of the base-pairand helix geometry shows that this is a conse-quence of the helix being right-handed. Incorpo-ration of the type I configuration adjacent to aregular A-form helix would require larger twistsand shifts of the uridine bases.
Comparison with other structures
In RNA helices the incorporation of single U-Uor C-U base-pairs commonly destabilizes the
helix.30 For symmetric tandem UU or 50CU30/30UC50 base-pair motifs the situation becomesmore complicated. After the 50GA30/50AG30 and50GU30/50UG30 tandems, the tandem U-U base-pair motif is the most stable and can stabilize thehelix, when flanked by G-C base-pairs, butbecomes destabilizing when flanked by A-U base-pairs.31 The symmetric 50CU30/50UC30 tandembase-pair motif is destabilizing in any context.31
For asymmetric tandems of non-canonical base-pairs the situation is even more complex, becausehere the free energy contribution of the one isdependent on the identity of the other.26 Simplerules for predicting free energies by summation ofthe energy contributions of individual base-pairs,which is the basis of the Tinoco32 or Turner rules,33
do not apply here. The free energy contribution ofintroducing an asymmetric non-canonical tandembase-pair motif is in many cases not equal to theaverage of the free energy contributions of twosymmetric tandems.26 It is interesting to note, how-ever, that the 50CU30/30UU50 tandem is one of therare examples that is energetically more favorablethan the average of the two symmetric C-U andU-U tandem base-pairs.26
Here we describe the structure of an internalloop containing a 50CU30/50UU30 base-pair motif,which reveals pseudo-2-fold symmetry of an asym-metric non-canonical tandem base-pair motif, thatmaintains the overall symmetry of the helix. Wewould like to suggest that maintaining pseudo-2-fold symmetry is a more general phenomenon inRNA helices containing tandem non-canonicalbase-pairs by which stabilities and structures ofthe tandem non-canonical base-pairs can beexplained using geometrical considerations. Amore detailed analysis and discussion is presentedelsewhere.34
Regular A-form RNA can be viewed as a stack ofbase-pair building blocks, n– n0; with fixed cross-strand 50Pn 2 n0O30 distances of 17.5 A on eitherside of the base-pair (Table 1). A single pyrimi-dine–pyrimidine base-pair with a short C10 –C10
cross-strand distance is probably hard to fit intoan A-form helix because the two 50P–O30 distanceson either side of the base-pair are different. Forinstance, in the type II U-U base-pair (vide supra)the 50P–O30 distance is 16 A on one side and 14 Aon the other. Likewise, in a sheared G-A base-pairwith a 9.7 A C10 –C10 distance the 50P–O30
distances are 17 A and 13 A, respectively. In theubiquitous U-G wobble, which is one of the moststable non-canonical base-pairs, the distances onlydeviate less than 0.5 A from A-form and destacking50 to the G is compensated by enhanced stacking 30
to the G.35
As shown and discussed before,8,9,36 in a tandem50GA30/30AG50 base-pair motif the two shearedG-A base-pairs perfectly fit into an A-form helixbecause the asymmetry introduced by the firstG-A base-pair is compensated by the second.Hence the large 50P–O30 distances face the regularhelix, while the two short 50P–O30 distances face
Figure 3. Phylogeny of the central six base-pairs in theY-domain of the enteroviruses. The upper three showvariations within the polio and polio-like coxsackiefamily, the lower six show the variations among thenon-polio-like coxsackie and ECHO viruses.
764 Structures of Tandem Non-canonical Base-pairs
each other. The structure of the 50CU30/30UU50
motif shows that also in a non-canonical base-pairtandem with an asymmetric sequence, maintaininga pseudo-2-fold symmetry allows for a perfect fitinto the A-form helix, with large distances at theregular helix sides and the shorter distances at thecentral C-U/U-U base-pair step.
The 12 possible symmetric tandems of non-canonical base-pairs in RNA can be divided intostable ones with clear imino proton resonances andunstable, more flexible ones with absent or broadimino proton resonances. Xia et al.26 showed that inasymmetric tandems a stable non-canonical base-pair next to an unstable one is energetically morefavorable than matching two stable non-canonicalbase-pairs, presumably because they are different insize and shape. Following the discussion we wouldlike to argue that the conformation of the stablenon-canonical base-pair possibly forces the unstablebase-pair in the proper matched conformation, tomaintain the pseudo-2-fold symmetry.
The internal CU/UU loop bears greatresemblance to the crystal structure of a self-complementary RNA dodecamer containingtandem U-U base-pairs flanked by C-G base-pairs,termed the CUUG duplex (rmsd ¼ 1.9 A, all back-bone heavy atoms of the central six base-pairs,27)but differs from the crystal structure of a self-complementary RNA dodecamer containingtandem U-C base-pairs flanked by U-G base-pairs,termed the UUCG duplex (rmsd ¼ 2.4 A,28). In theUUCG motif the U-C base-pairs are opened towardthe minor groove with near regular 11.7 A C10 –C10
distances to fit into the helix and only form onedirect hydrogen bond between the UO4 and theCN4. Additional stability is provided by a bridgingwater molecule between the N3 endocyclic nitro-gen atoms. The U-U base-pairs in the CU/UUtandem, described here, adopt essentially thesame configuration as in the CUUG duplex, andthe C-U base-pair is isomorphic. One mightwonder why the CU/UU tandem favors the
Figure 4. Overlays of the polio Y-domain structure with an A-form helix, superimposed on the central six base-pairs,i.e. residues 4–7/16–19. The coloring of the polio structure is as in Figure 2(B) and (D). The backbone of the A-formRNA is in yellow, the bases of residues 5–6/16–17 as in the polio structure, and the other bases are colored in red.(A) Stereo side view of all residues, showing the contraction of the backbone at the internal loop. (B) Top view downthe helical axis of the backbone and residues 5–6/16–17.
Structures of Tandem Non-canonical Base-pairs 765

CUUG conformation and rejects the UUCG confor-mation. Again we argue that the answer lies in thetendency of the helix to maintain its overall sym-metry. A comparison of the three structures revealsthat adopting the U-U base-pair configuration asobserved in the CUUG duplex, and maintainingsymmetry allows for a matching C-U base-pairconfiguration with proper hydrogen bonding,while adopting the C-U base-pair configuration asobserved in the UUCG duplex would not. In thiscase maintaining symmetry would force the U-Ubase-pair into a configuration in which the twoO4s and N3Hs would oppose each other, which isthermodynamically unfavorable.
Helices with a sheared tandem G-A base-pairmotif also show a large helical twist (70–808) atthe central G-A/A-G base-pair step, compensatedby small helical twists on either side (,178) againleading to an overall regular helical twist.8,9 Com-pared to the CU/UU tandem, the situation in thetandem sheared G-A base-pair motif is different inthat the 50-guanine bases are also stacked cross-strand as opposed to unstacking of the 50-bases inthe CU/UU loop. However, in this particulartandem G-A base-pair motif the guanosine basesare sheared to narrow down the cross-strandGO3
0–P50A distance to 13 A, which dents the
sugar-phosphate backbone at the 50-guanosinebases, with a 20-endo sugar pucker conformationand changes in several backbone torsion angles.9
In the pyrimidine module no major changes ofbackbone torsion angles are present and the back-bone is not dented (Figure 4).
The configuration of the C-U base-pair observedin our structure comes closest to the one observedin the solution structure of the isolated A-loop ofthe peptidyl transferase center.23 Although theauthors were also not able to detect a cytidineimino proton resonance for the C-U base-pair, thepH-shift dependence of several aromatic reson-ances was in accordance with protonation ofthe cytidine with a pKa of ,6.4. We also sawcomparable ,0.1 ppm chemical shift changes ofC5, H5 and H6 between pH 5.8 and pH 7.0, there-fore the C-U base-pair in our structure might bestabilized by an additional hydrogen bondbetween the CN3 and UO4 as well.
Conclusions and Biological Relevance
Mismatches in RNA helices can fulfill a couple ofroles. They can tune the stability of the RNA helix,which might for instance be important for regulat-ing initiation of unfolding and refolding of RNAmolecules in processes involving strand separationand formation like transcription or translationalcontrol. Also they can direct the position ofhydrogen bond acceptors or donors, which isimportant for the recognition of proteins or otherRNA elements. Understanding the forces that dic-tate the stability and structures of mismatchedRNA helices is therefore of prime importance for
formulating the principles that drive RNA intotheir specific shapes to carry out their function.
The salient structural features of the internalloop are the formation of the two pyrimidinebase-pairs with a typical stacking pattern, contrac-tion of the minor groove and the pseudo-2-foldsymmetry. Although the bases of the CU/UU loopare not expected to be involved in hydrogenbonds, because they are not exposed to solvent,formation of the pyrimidine base-pairs movespossible hydrogen bond donors and acceptors ofthe sugar-phosphate backbone. This makes theinternal loop an interesting site for interactionwith proteins and/or other RNA elements. Thepoliovirus 30-UTR is directly involved in theinitiation of viral replication. The formation of aRNP complex is most probably an essential step inthe regulation of replication. Since no proteins areyet known that specifically interact with thepyrimidine tandem in the Y-loop, the workpresented here provides for an excellent structuralframework to direct further biological studies.Although we were not able to detect an effect inour mutagenesis study upon changing the pyrimi-dine base-pairs in Watson–Crick base-pairs, thisdoes not imply that there is no effect at all, butrather suggests that the in vitro studies are not aptto discover an effect of every mutation in vivo,especially those with subtle effects. We and othershave observed this in vitro– in vivo differencepreviously as well.37
Perhaps the most important feature that emergesfrom this study is the pseudo-2-fold character ofthe asymmetric tandem, which became apparentfrom an analysis of the helical twists and l angles.This has several implications. It allows for predict-ing the conformation of asymmetric tandem mis-matches, containing non-canonical base-pairs,taking the structure of the most stable non-canoni-cal base-pair as a starting point. It also predictsthat in functionally important RNAs changing atandem non-canonical base-pair motif into a singlenon-canonical base-pair probably will have a largerimpact than changing the tandem into twoWatson–Crick base-pairs, since only in the formercase the overall helix symmetry will be lost.Further work is needed to address these interestingaspects and to confirm that retaining symmetry is ageneral phenomenon in A-form helices containingtandem mismatches.
Methods
Sample preparation
An unlabeled and a pyrimidine labeled 13C/15N 22-mer RNA were prepared enzymatically by in vitro tran-scription using T7 RNA polymerase and oligo DNAtemplates.38 The RNA molecules were purified usingpreparative polyacrylamide gel electrophoresis and sub-sequent electroelution. Purified RNA was extensivelydialyzed against water and concentrated using Centriconmicroconcentrators. The final concentration of the
766 Structures of Tandem Non-canonical Base-pairs

unlabeled sample was 3 mM and 1–2 mM for theresidue-specific 13C/15N labeled RNA sample. The pHof the NMR samples was adjusted to 6.8 or 5.5 (meterreading).
NMR spectroscopy
Spectra were recorded on Varian Inova 600 and 750spectrometers. The exchangeable imino proton signalswere assigned from a 750 MHz two-dimensionalNOESY39 spectrum recorded on the unlabeled sample in90% H2O, 10% 2H2O at 0 8C using a jump-returnsequence for water suppression.40 Other spectra wererecorded at 15 8C in 2H2O at 600 MHz. Non-exchangeableprotons were assigned from two-dimensional NOESYspectra (mixing times 50, 100, 200 and 300 ms), DQF-COSY,41 TOCSY42 and a three-dimensional NOESY-HMQC43 spectrum. Complete assignment of the labeledresidues was obtained using a 31P-decoupled HCCH-TOCSY.44
Structural restraints
Distances involving non-exchangeable protons wereextracted from two-dimensional NOESY spectra withmixing times 50, 100 and 200 ms and the three-dimensional NOESY-HMQC with a mixing time of200 ms. To account for spin diffusion effects, distanceswere preferably calculated from the 50 ms and 100 msNOE cross-peak intensities, which were converted intodistance restraints using the modified isolated spin pairapproximation approach.45 The resulting distances wereused with ^20% error bounds. Nineteen NOE distancerestraints involving exchangeable protons were con-servatively estimated from NOESY spectra recorded inH2O. Hydrogen bonding restraints were treated as NOEdistance restraints and imposed on residues involved inWatson–Crick base-pairing, evidenced by the iminoproton spectra and observed NOEs and chemical shifts.Scalar couplings between H10 and H20 were determinedfrom a DQF-COSY, TOCSY and NOESY experimentsand were interpreted as N or S-type sugar puckerswhen they were small (,3 Hz) or large (.8 Hz),respectively. Sugar puckers of G1, A11, A12, G19 andC22 were left unrestrained because of their intermediateH10 to H20 scalar couplings (5–6 Hz). All residues
showed strong H10/H20/H30 to H6/H8 NOE inter-actions, indicative of x-angles in the anti-range. Thesewere restrained to 202(^30)8 for stem residues and202(^60)8 for the others.
A TOCSY-CT-HSQC44 was used to discriminatebetween the gþ and gt rotamers of the 13C-labeled U andC residues. This experiment relies on efficient TOCSYtransfer from H40 to H50/H500 for gt and poor transferfor gþ (and g2 which in practice is not observed.29).Observed gþ and gt rotamers were restrained to54(^30)8 and 180(^30)8, respectively.
The b and 1 dihedral angles in labeled residues werederived from a novel spin-echo difference CT-HCCH-TOCSY experiment (E.M.H.P.L. et al.,unpublishedresults) and restrained to their calculated values witherror bounds of ^208 and ^308, respectively. For theunlabeled residues of the stems the 1 dihedral anglewas restrained to 225(^60)8 to prevent for the “forbid-den” gaucheþ conformation. The dihedral angles a and zwere restricted to 0(^120)8 if the corresponding 31P reso-nated within the 1 ppm range, typical for A-type RNA,and the measured g angles of the involved base-pairstep were in the gaucheþ conformation.
Structure calculations
Starting from an extended strand, a set of 100structures was generated by torsion angle dynamics inX-PLOR,46 largely identical to the protocol proposed fora DNA duplex.47 In the first round of structure calcu-lations, only one imino proton–imino proton NOErestraint was used for the U-U base-pair, which gener-ated a low number of acceptable structures that satisfiedall input data. Convergence could be improved byimposing hydrogen bond restraints compatible with atype II U-U base-pair. Structure calculations for theother possible, type I, U-U base-pair did not result inacceptable structures, due to incompatibility with theinput data as evidenced by a large number of NOEviolations, e.g. between U6H10 and U18H10. Thirty-nineof the obtained structures calculated with the restraintsincluding those for the type II U-U base-pair had nodihedral angle violations larger than 58 and no NOEviolations larger than 0.5 A (Table 2). These 39 acceptedstructures were subjected to refinement and a finalround of energy minimization.
Additional validation of the structures was obtainedby using the program NUChemiCS,48,49 which predictsthe resonance positions of non-exchangeable protons onthe basis of the derived structure. For the stem andinternal loop residues the difference between theobserved and calculated chemical shifts was less than0.1 ppm on average. Color Figures were generatedusing MOLMOL.50 Helical parameters were calculatedusing the program NEWHELIX93 (R. E. Dickerson).
Mutational studies
In vitro mutagenesis and characterization of mutantvirus was performed as described.51 Mutations wereintroduced into the poliovirus type 1 cDNA clone pXPA(a kind gift from Dr R. Andino, University of California,USA) using oligonucleotide p295 (50-AAGAAAAATTTACCCCTACCTCAGTATGACCCAATCCAA-30). Thenucleotide sequence of the mutant construct was verifiedby sequence analysis.
Plasmid DNA was linearized with Mlu I and tran-scribed in vitro by T7 RNA polymerase. Buffalo Green
Table 2. Structural statistics of the final ensemble of 39structures
Distance restraintsIntranucleotide 170Internucleotide 110Hydrogen bonds 19Dihedral angle restraints 160
Rms deviations (A,8)Distance restraints 0.046 ^ 0.003Dihedral restrains 0.475 ^ 0.107
Rms deviation from idealizedBonds (A) 0.0043 ^ 0.0001Angles (8) 1.077 ^ 0.012Impropers (8) 0.818 ^ 0.013
Restraint violationsNumber of distance violations† . 0.2 A 3 ^ 3Number of dihedral violations‡ . 28 3 ^ 3Atomic rms deviations (A) 1.59 ^ 1.04
† None larger than 0.5A.‡ None larger than 58.
Structures of Tandem Non-canonical Base-pairs 767

Monkey kidney cells were transfected with RNA tran-scripts and grown until virus growth was complete.Sequence analysis showed that the introduced mutationwas retained in the viral RNA and that no othermutations had occurred. The obtained viruses werecharacterized by single cycle growth analysis.
Atomic coordinates
Coordinates of the 20 lowest energy structures havebeen deposited in the RCSB Protein Data Bank (accessioncode 1N66).
Acknowledgements
This work was supported by EuropeanCommunities (INTAS 2012) and NWO-CW.
References
1. Crick, F. H. (1966). Codon–anticodon pairing: thewobble hypothesis. J. Mol. Biol. 19, 548–555.
2. Hermann, T. & Westhof, E. (1999). Non-Watson–Crick base pairs in RNA–protein recognition. Chem.Biol. 12, R335–R343.
3. Mckay, D. B. (1996). Structure and function of thehammerhead ribozyme: an unfinished story. RNA, 2,395–403.
4. Cate, J. H., Gooding, A. R., Podell, E., Zhou, K.,Golden, B. L., Kundrot, C. E. et al. (1996). Crystalstructure of a group I ribozyme domain: principlesof RNA packing. Science, 273, 1678–1685.
5. Andersen, A. A. & Collins, R. A. (2000). Rearrange-ment of a stable RNA secondary structure duringVS ribozyme catalysis. Mol. Cell, 5, 469–478.
6. Michiels, P. J., Schouten, C. H., Hilbers, C. W. &Heus, H. A. (2000). Structure of the ribozyme sub-strate hairpin of Neurospora VS RNA: a close lookat the cleavage site. RNA, 6, 1821–1832.
7. Wu, M. & Turner, D. H. (1996). Solution structure of(rGCGGACGC)2 by two-dimensional NMR and theiterative relaxation matrix approach. Biochemistry,35, 9677–9689.
8. SantaLucia, J. & Turner, D. H. (1993). Structure of(rGGCGAGCC)2 in solution from NMR andrestrained molecular dynamics. Biochemistry, 32,12612–12623.
9. Heus, H. A., Wijmenga, S. S., Hoppe, H. & Hilbers,C. W. (1997). The detailed structure of tandem G.Amismatched base-pair motifs in RNA duplexes iscontext dependent. J. Mol. Biol. 271, 147–158.
10. Kitamura, N., Semler, B. L., Rothberg, P. G., Larsen,G. R., Adler, C. J., Dorner, A. J. et al. (1981). Primarystructure, gene organisation and polypeptideexpression of poliovirus RNA. Nature, 291, 547–553.
11. Pilipenko, E. V., Poperechny, K. V., Maslova, S. V.,Melchers, W. J., Bruins Slot, H. J. & Agol, V. I.(1996). Cis-element, oriR, involved in the initiationof (2) strand poliovirus RNA: a quasi-globularmulti-domain RNA structure maintained by tertiary(‘kissing’) interactions. EMBO J. 15, 5428–5436.
12. Melchers, W. J., Hoenderop, J. G., Bruins Slot, H. J.,Pleij, C. W., Pilipenko, E. V., Agol, V. I. & Galama,J. M. (1997). Kissing of the two predominant hairpin
loops in the coxsackie B virus 30 untranslated regionis the essential structural feature of the origin ofreplication required for negative-strand RNA syn-thesis. J. Virol. 71, 686–696.
13. Wang, J., Bakkers, J. M., Galama, J. M., Bruins Slot,H. J., Pilipenko, E. V., Agol, V. I. & Melchers, W. J.(1999). Structural requirements of the higher orderRNA kissing element in the enteroviral 30UTR. Nucl.Acids Res. 27, 485–490.
14. Melchers, W. J., Bakkers, J. M., Bruins Slot, H. J.,Galama, J. M., Agol, V. I. & Pilipenko, E. V. (2000).Cross-talk between orientation-dependent recog-nition determinants of a complex control RNAelement, the enterovirus oriR. RNA, 6, 976–987.
15. Harris, K. S., Xiang, W., Alexander, L., Lane, W. S.,Paul, A. V. & Wimmer, E. (1994). Interaction of polio-virus polypeptide 3CDpro with the 50 and 30 terminiof the poliovirus genome. J. Biol. Chem. 269,27004–27014.
16. Waggoner, S. & Sarnow, P. (1998). Viral ribonucleo-protein complex formation and nucleolar-cyto-plasmic relocalization of nucleolin in poliovirus-infected cells. J. Virol. 72, 6699–6709.
17. Meredith, J. M., Rohll, J. B., Almond, J. W. & Evans,D. J. (1999). Similar interactions of the poliovirusand rhinovirus 3D polymerases with the 30 untrans-lated region of rhinovirus 14. J. Virol. 73, 9952–9958.
18. Herold, J. & Andino, R. (2001). Poliovirus RNAreplication requres genome circularization through aprotein–ptotein bridge. Mol. Cell, 7, 581–591.
19. Barton, D. J., O’Donnell, B. J. & Flanegan, J. B. (2001).50 Cloverleaf in poliovirus RNA is a cis-actingreplication element required for negative strand syn-thesis. EMBO J. 20, 1439–1448.
20. Butcher, S. E., Dieckmann, T. & Feigon, J. (1997).Solution structure of a GAAA tetraloop receptorRNA. EMBO J. 16, 7490–7499.
21. Wu, M. & Tinoco, I., Jr (1998). RNA folding causessecondary structure rearrangement. PNAS, 96,11555–11560.
22. Williamson, J. R. (2000). Induced fit in RNA–proteinrecognition. Nature Struct. Biol. 7, 834–837.
23. Blanchard, S. C. & Puglisi, J. D. (2001). Solutionstructure of the A loop of 23 S ribosomal RNA. Proc.Natl Acad. Sci. USA, 98, 3720–3725.
24. Wijmenga, S. S., Mooren, M. W. & Hilbers, C. W.(1993). NMR of nucleic acids; from spectrum tostructure. In NMR of Macromolecules. A PracticalApproach (Roberts, G. C. K., ed.), pp. 217–288, OxfordUniversity Press, New York.
25. Jucker, F. M., Heus, H. A., Yip, P. F., Moors, E. H. &Pardi, A. (1996). A network of heterogeneous hydro-gen bonds in GNRA tetraloops. J. Mol. Biol. 164,968–980.
26. Xia, T., McDowell, J. A. & Turner, D. H. (1997).Thermodynamics of nonsymmetric tandem mis-matches adjacent to G·C base pairs in RNA.Biochemistry, 36, 12486–12497.
27. Lietzke, S. E., Barnes, C. L., Berglund, J. A. &Kundrot, C. E. (1996). The structure of an RNAdodecamer shows how tandem U–U base pairsincrease the range of stable RNA structures and thediversity of recognition sites. Structure, 4, 917–930.
28. Holbrook, S. R., Cheong, C., Tinoco, I., Jr & Kim, S. H.(1991). Crystal structure of an RNA double helixincorporating a track of non-Watson–Crick basepairs. Nature, 353, 579–581.
29. Saenger, W. (1984). Principles of Nucleic Acid Structure,Springer, New York.
768 Structures of Tandem Non-canonical Base-pairs

30. Kierzek, R., Burkard, M. E. & Turner, D. H. (1999).Thermodynamics of single mismatches in RNAduplexes. Biochemistry, 38, 14214–14223.
31. Wu, M., McDowell, J. A. & Turner, D. H. (1995). Aperiodic table of symmetric tandem mismatches inRNA. Biochemistry, 34, 3204–3211.
32. Tinoco, I., Jr, Borer, P. N., Dengler, B., Levine, M. D.,Uhlenbeck, O. C., Crothers, D. M. & Gralla, J. (1973).Improved estimation of secondary structure in ribo-nucleic acids. Nature New Biol. 246, 40–41.
33. Serra, M. J. & Turner, D. H. (1995). Predictingthermodynamic properties of RNA. MethodsEnzymol. 259, 243–261.
34. Heus, H. A. & Hilbers, C. W. (2003). Structuresof non-canonical base pairs in RNA helices:review. Nucleotides, Nucleosides Nucleic Acids, 22,551–563.
35. Mizuno, H. & Sundaralingam, M. (1978). Stacking ofCrick Wobble pair and Watson–Crick pair: stabilityrules of G–U pairs at ends of helical stems intRNAs and the relation to codon–anticodon Wobbleinteraction. Nucl. Acids Res. 5, 4451–4461.
36. Gautheret, D., Konings, D. & Gutell, R. R. (1994). Amajor family of motifs involving G·A mismatches inribosomal RNA. J. Mol. Biol. 242, 1–8.
37. Merkle, I., van Ooij, M. J., van Kuppeveld, F. J.,Glaudemans, D. H., Galama, J. M., Henke, A. et al.(2002). Biological significance of a human entero-virus B-specific RNA element in the 30 nontranslatedregion. J. Virol. 76, 9900–9909.
38. Milligan, J. F., Groebe, D. R., Witherell, G. W. &Uhlenbeck, O. C. (1987). Oligoribonucleotidesynthesis using T7 RNA polymerase andsynthetic DNA templates. Nucl. Acids Res. 15,8783–8798.
39. Jeener, J., Meier, B. H., Bachmann, P. & Ernst, R. R.(1970). Investigation of exchange processes by twodimensional NMR spectroscopy. J. Chem. Phys. 71,4546–4553.
40. Plateau, P. & Gueron, M. (1982). Exchangeableproton NMR without baseline distortion, using newstrong-pulse sequences. J. Am. Chem. Soc. 104,7310–7311.
41. Shaka, A. J. & Freeman, R. (1983). Simplification of
NMR spectra by filtration through multiple quantumcoherence. J. Magn. Reson. 51, 169–173.
42. Griesinger, C., Otting, G., Wuthrich, K. & Ernst, R. R.(1988). Clean TOCSY for 1H spin system identifi-cation in macromolecules. J. Am. Chem. Soc. 110,7870–7872.
43. Ikura, M., Kay, L. E., Tschudin, R. & Bax, A. (1990).Three-dimensional NOESY-HMQC spectroscopy ofa 13C labeled protein. J. Magn. Reson. 86, 204–209.
44. Kolk, M. H., Wijmenga, S. S., Heus, H. A. & Hilbers,C. W. (1998). On the structure determination of a44n RNA pseudoknot: assignment strategies andderivation of torsion angle restraints. J. Biomol.NMR, 12, 423–433.
45. Barsukov, I. L. & Lian, L. Y. (1993). Structure determi-nations from NMR data I. Analysis of NMR data. InNMR of Macromolecules. A Practical Approach(Roberts, G. C. P., ed.), pp. 315–357, Oxford Univer-sity Press, New York.
46. Brunger, A. T. (1992). X-PLOR, A System for X-rayCrystallography and NMR, Yale University Press,New Haven.
47. Stein, E. G., Rice, L. M. & Brunger, A. T. (1997).Torsion-angle molecular dynamics as a new efficienttool for NMR structure calculation. J. Magn. Reson.124, 154–164.
48. Wijmenga, S. S., Kruithof, M. & Hilbers, C. W. (1997).Analysis 1H chemical shifts in DNA: assessment ofthe reliability of 1H chemical shift calculations foruse in structure refinement. J. Biomol. NMR, 10,337–350.
49. Cromsigt, J. A. M. T. C., Hilbers, C. W. & Wijmenga,S. S. (2001). Prediction of proton chemical shifts inRNA. Their use in structure refinement andvalidation. J. Biomol. NMR, 21, 11–29.
50. Koradi, R., Billeter, M. & Wuthrich, K. (1996).MOLMOL: a program for display and analysis ofmacromolecular structures. J. Mol. Graph. 14, 51–55.
51. van Kuppeveld, F. J., Galama, J. M., Zoll, J. &Melchers, W. J. (1995). Genetic analysis of a hydro-phobic domain of coxsackie B3 virus protein 2B: amoderate degree of hydrophobicity is required for acis-acting function in viral RNA synthesis. J. Virol.69, 7782–7790.
Edited by M. F. Summers
(Received 25 March 2003; received in revised form 16 June 2003; accepted 16 June 2003)
Structures of Tandem Non-canonical Base-pairs 769
Related Documents











