www.landesbioscience.com BioArchitecture 127 BioArchitecture 1:3, 127-133; May/June 2011; © 2011 Landes Bioscience RESEARCH PAPER RESEARCH PAPER *Correspondence to: Chih-Lueh Albert Wang; Email: [email protected] Submitted: 04/07/11; Revised: 05/24/11; Accepted: 05/25/11 DOI:10.4161/bioa.1.3.16714 Introduction Caldesmon (CaD) is an actin-binding protein present in almost all cells from vertebrates. There are two major CaD isoforms: the heavy, or h-CaD, that uniquely exists in smooth muscle cells, and the light, or l-CaD, that exists in all non-muscle cells. 1 The shorter isoform is missing a central repeating region, while the primary actin-binding domains located in the C-terminal region are conserved. Binding of CaD to actin filaments is known to inhibit the actomyosin interaction in vitro and is reversed by Ca 2+ /calmodulin or by ERK-mediated phosphorylation. 2 In con- trast, the effect of CaD on the polymerization of actin is not fully understood. We have shown previously in reference 3, that CaD or its C-terminal fragment (H32K), regardless of phosphorylation, if added while actin was beginning to polymerize, attenuated the characteristic enhancement of pyrene-actin emission that normally accompanies the polymerization. Surprisingly, adding CaD after polymerization accelerated, rather than suppressed, the pyrene- actin fluorescence enhancement. Both ERK-phosphorylated and We have previously reported that actin undergoes a conformational transition (which we named “maturation”) during polymerization, and that the actin-binding protein, caldesmon (CaD), when added at an early phase of polymerization, interferes with this process (Huang et al. J Biol Chem 2010; 285:71). The pre-transition filament is characterized by relatively low pyrene-fluorescence intensity when pyrene-labeled actin is used as a reporter of subunit assembly into filaments, whereas the mature filament emits a characteristic enhanced fluorescence. Previously reported co-sedimentation experiments suggest that filament formation is not inhibited by the presence of CaD, despite blocking the transition associated with filament maturation. In this study we visualized structural effects of CaD on the assembly of actin filaments by TIRF and electron microscopy. CaD-free actin forms “rough” filaments with irregular edges and indistinct subunit organization during the initial phase (~20 min under our conditions) of polymerization as reported previously by others (Steinmetz et al. J Cell Biol 1997; 138:559; Galinska-Rakoczy et al. J Mol Biol 2009; 387:869), which most likely correspond to the pre-transition state preceding the maturation step. Later during the polymerization process “mature” filaments exhibit a smoother F-actin appearance with easily detectible double helically arranged actin subunits. While the inclusion of the actin-binding domain of CaD during actin polymerization does not affect the elongation rate, it is associated with a prolonged pre-transition phase, characterized by a delayed alteration (rough to smooth) of the appearance of filaments, consistent with a later onset of the maturation process. Structural studies on maturing actin filaments Agnieszka Collins, 1,† Renjian Huang, 3 Mikkel Herholdt Jensen, 1,2 Jeffrey R. Moore, 1 William Lehman 1 and Chih-Lueh Albert Wang 3, * 1 Boston University School of Medicine; 2 Boston University Department of Physics; Boston; 3 Boston Biomedical Research Institute; Watertown, MA USA † Current address: Department of Biology; University of Pennsylvania; Philadelphia, PA USA Key words: actin, polymerization, caldesmon, TIRF microscopy, electron microscopy, conformational change, cytoskeleton dynamics Abbreviations: CaD, caldesmon; EM, electron microscopy; ERK, extracellular signal-regulated kinase; CaM, calmodulin; H32K, a C-terminal fragment of CaD; pH32K, ERK-phosphorylated H32K; TIRFM, total internal reflectance fluorescence microscopy unphosphorylated CaD fragments accelerated the pyrene-actin fluorescence increase when added after the initiation of actin polymerization. However, all samples, regardless of whether the CaD fragment was present or not, or of when it was added, yielded the same amount of sedimentable actin filaments, suggesting that what is inhibited by the CaD fragment is not the polymerization per se, but rather a conformational change. We postulate that an irreversible conformational transition (E1 →E2, which we call “maturation” 3 ) occurs during actin polymerization. If CaD is present early in this process, it binds to the low pyrene fluorescence state of actin (E1), thereby prevent- ing maturation. If CaD is present after the transition, it binds primarily to the high pyrene fluorescence state of actin (E2) and promotes maturation. In the current study we visualized the structural impact of CaD by total internal reflectance fluo- rescence microscopy (TIRFM) and electron microscopy (EM). Our results indeed reveal a time-dependent change in the mor- phology of actin filaments corresponding to this transition, and CaD when added at initial stages of polymerization delays such a change.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.landesbioscience.com BioArchitecture 127
BioArchitecture 1:3, 127-133; May/June 2011; © 2011 Landes Bioscience
RESEARCH PAPER RESEARCH PAPER
*Correspondence to: Chih-Lueh Albert Wang; Email: [email protected]: 04/07/11; Revised: 05/24/11; Accepted: 05/25/11DOI:10.4161/bioa.1.3.16714
Introduction
Caldesmon (CaD) is an actin-binding protein present in almost all cells from vertebrates. There are two major CaD isoforms: the heavy, or h-CaD, that uniquely exists in smooth muscle cells, and the light, or l-CaD, that exists in all non-muscle cells.1 The shorter isoform is missing a central repeating region, while the primary actin-binding domains located in the C-terminal region are conserved. Binding of CaD to actin filaments is known to inhibit the actomyosin interaction in vitro and is reversed by Ca2+/calmodulin or by ERK-mediated phosphorylation.2 In con-trast, the effect of CaD on the polymerization of actin is not fully understood.
We have shown previously in reference 3, that CaD or its C-terminal fragment (H32K), regardless of phosphorylation, if added while actin was beginning to polymerize, attenuated the characteristic enhancement of pyrene-actin emission that normally accompanies the polymerization. Surprisingly, adding CaD after polymerization accelerated, rather than suppressed, the pyrene-actin fluorescence enhancement. Both ERK-phosphorylated and
We have previously reported that actin undergoes a conformational transition (which we named “maturation”) during polymerization, and that the actin-binding protein, caldesmon (CaD), when added at an early phase of polymerization, interferes with this process (Huang et al. J Biol Chem 2010; 285:71). The pre-transition filament is characterized by relatively low pyrene-fluorescence intensity when pyrene-labeled actin is used as a reporter of subunit assembly into filaments, whereas the mature filament emits a characteristic enhanced fluorescence. Previously reported co-sedimentation experiments suggest that filament formation is not inhibited by the presence of CaD, despite blocking the transition associated with filament maturation. In this study we visualized structural effects of CaD on the assembly of actin filaments by TIRF and electron microscopy. CaD-free actin forms “rough” filaments with irregular edges and indistinct subunit organization during the initial phase (~20 min under our conditions) of polymerization as reported previously by others (Steinmetz et al. J Cell Biol 1997; 138:559; Galinska-Rakoczy et al. J Mol Biol 2009; 387:869), which most likely correspond to the pre-transition state preceding the maturation step. Later during the polymerization process “mature” filaments exhibit a smoother F-actin appearance with easily detectible double helically arranged actin subunits. While the inclusion of the actin-binding domain of CaD during actin polymerization does not affect the elongation rate, it is associated with a prolonged pre-transition phase, characterized by a delayed alteration (rough to smooth) of the appearance of filaments, consistent with a later onset of the maturation process.
Structural studies on maturing actin filamentsAgnieszka Collins,1,† Renjian Huang,3 Mikkel Herholdt Jensen,1,2 Jeffrey R. Moore,1 William Lehman1
and Chih-Lueh Albert Wang3,*
1Boston University School of Medicine; 2Boston University Department of Physics; Boston; 3Boston Biomedical Research Institute; Watertown, MA USA
†Current address: Department of Biology; University of Pennsylvania; Philadelphia, PA USA
Key words: actin, polymerization, caldesmon, TIRF microscopy, electron microscopy, conformational change, cytoskeleton dynamics
abbreviations: CaD, caldesmon; EM, electron microscopy; ERK, extracellular signal-regulated kinase; CaM, calmodulin; H32K, a C-terminal fragment of CaD; pH32K, ERK-phosphorylated H32K; TIRFM, total internal reflectance fluorescence microscopy
unphosphorylated CaD fragments accelerated the pyrene-actin fluorescence increase when added after the initiation of actin polymerization. However, all samples, regardless of whether the CaD fragment was present or not, or of when it was added, yielded the same amount of sedimentable actin filaments, suggesting that what is inhibited by the CaD fragment is not the polymerization per se, but rather a conformational change.
We postulate that an irreversible conformational transition (E1→E2, which we call “maturation”3) occurs during actin polymerization. If CaD is present early in this process, it binds to the low pyrene fluorescence state of actin (E1), thereby prevent-ing maturation. If CaD is present after the transition, it binds primarily to the high pyrene fluorescence state of actin (E2) and promotes maturation. In the current study we visualized the structural impact of CaD by total internal reflectance fluo-rescence microscopy (TIRFM) and electron microscopy (EM). Our results indeed reveal a time-dependent change in the mor-phology of actin filaments corresponding to this transition, and CaD when added at initial stages of polymerization delays such a change.
128 BioArchitecture Volume 1 Issue 3
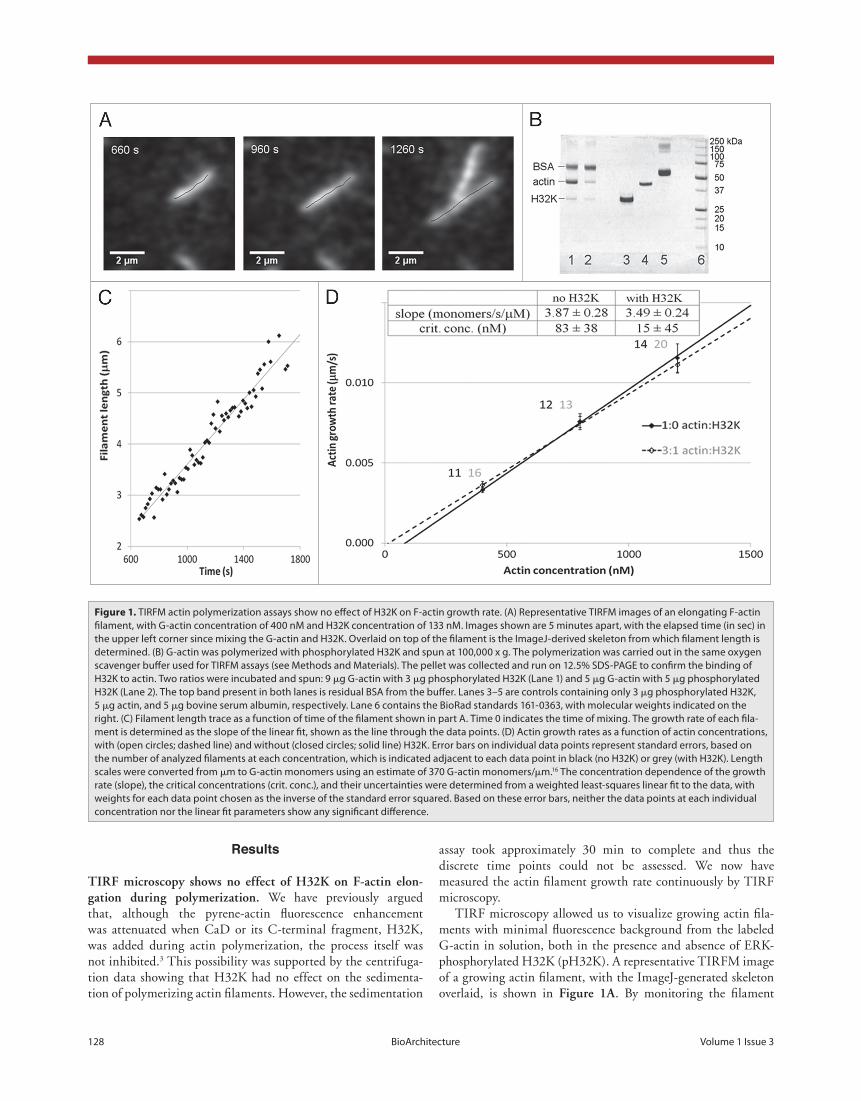
assay took approximately 30 min to complete and thus the discrete time points could not be assessed. We now have measured the actin filament growth rate continuously by TIRF microscopy.
TIRF microscopy allowed us to visualize growing actin fila-ments with minimal fluorescence background from the labeled G-actin in solution, both in the presence and absence of ERK-phosphorylated H32K (pH32K). A representative TIRFM image of a growing actin filament, with the ImageJ-generated skeleton overlaid, is shown in figure 1a. By monitoring the filament
Results
tirf microscopy shows no effect of h32K on f-actin elon-gation during polymerization. We have previously argued that, although the pyrene-actin fluorescence enhancement was attenuated when CaD or its C-terminal fragment, H32K, was added during actin polymerization, the process itself was not inhibited.3 This possibility was supported by the centrifuga-tion data showing that H32K had no effect on the sedimenta-tion of polymerizing actin filaments. However, the sedimentation
Figure 1. TIRFM actin polymerization assays show no effect of H32K on F-actin growth rate. (A) Representative TIRFM images of an elongating F-actin filament, with G-actin concentration of 400 nM and H32K concentration of 133 nM. Images shown are 5 minutes apart, with the elapsed time (in sec) in the upper left corner since mixing the G-actin and H32K. Overlaid on top of the filament is the ImageJ-derived skeleton from which filament length is determined. (B) G-actin was polymerized with phosphorylated H32K and spun at 100,000 x g. The polymerization was carried out in the same oxygen scavenger buffer used for TIRFM assays (see Methods and Materials). The pellet was collected and run on 12.5% SDS-PAGE to confirm the binding of H32K to actin. Two ratios were incubated and spun: 9 μg G-actin with 3 μg phosphorylated H32K (Lane 1) and 5 μg G-actin with 5 μg phosphorylated H32K (Lane 2). The top band present in both lanes is residual BSA from the buffer. Lanes 3–5 are controls containing only 3 μg phosphorylated H32K, 5 μg actin, and 5 μg bovine serum albumin, respectively. Lane 6 contains the BioRad standards 161-0363, with molecular weights indicated on the right. (C) Filament length trace as a function of time of the filament shown in part A. Time 0 indicates the time of mixing. The growth rate of each fila-ment is determined as the slope of the linear fit, shown as the line through the data points. (D) Actin growth rates as a function of actin concentrations, with (open circles; dashed line) and without (closed circles; solid line) H32K. Error bars on individual data points represent standard errors, based on the number of analyzed filaments at each concentration, which is indicated adjacent to each data point in black (no H32K) or grey (with H32K). Length scales were converted from μm to G-actin monomers using an estimate of 370 G-actin monomers/μm.16 The concentration dependence of the growth rate (slope), the critical concentrations (crit. conc.), and their uncertainties were determined from a weighted least-squares linear fit to the data, with weights for each data point chosen as the inverse of the standard error squared. Based on these error bars, neither the data points at each individual concentration nor the linear fit parameters show any significant difference.
www.landesbioscience.com BioArchitecture 129
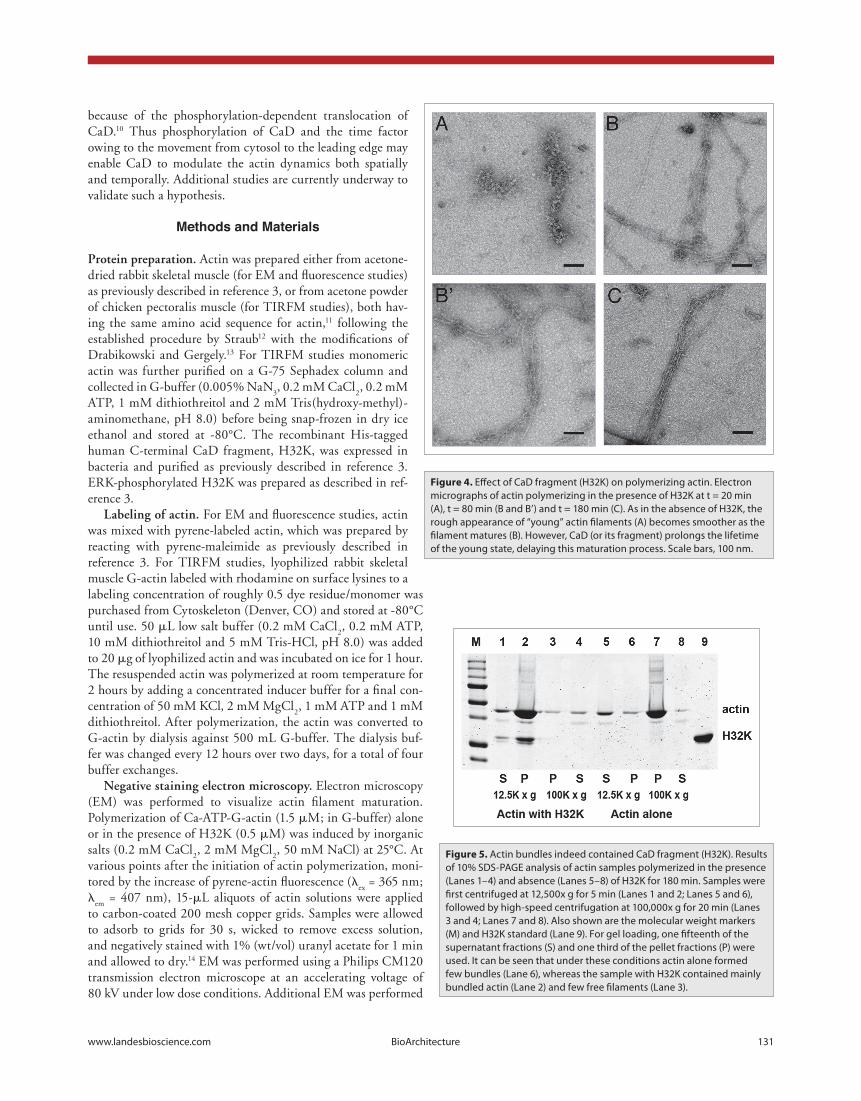
min, when the pyrene emission intensity was approximately the same as that of control actin at the very onset of polymerization, numerous filaments were now observed, many in loosely packed bundles of 3–6 long filaments in lateral association. These were irregular and rough, similar to those filaments of CaD-free actin at the earlier time point (e.g., at 20 min; fig. 4b and b’), even though the amount of polymerized actin was much greater than in the sample of control actin. Still, while a smaller population of the actin filaments was single and regular, their morphology was more like those of control actin filaments at 60 min. Finally, at 180 min the sample of H32K-actin was dominated by actin bundles that were wider and contained many tightly packed fila-ments (fig. 4c). Although some of the unaggregated filaments looked smooth and normal, the bundles still contained filaments that appeared irregular, more similar to that of the control actin sample at 30 min of polymerization than those at later time points.
To ascertain whether the actin bundles contained CaD, we carried out low-speed and high-speed sedimentation experiments. The results showed that actin bundles collected at 12,500x g cen-trifugation indeed contained H32K, while the amount of F-actin pelleted at 100,000x g following the low-speed spin was insignifi-cant (fig. 5). This is consistent with the fact that unbundled fila-ments were rare in the EM field and, therefore, not representative of the sample.
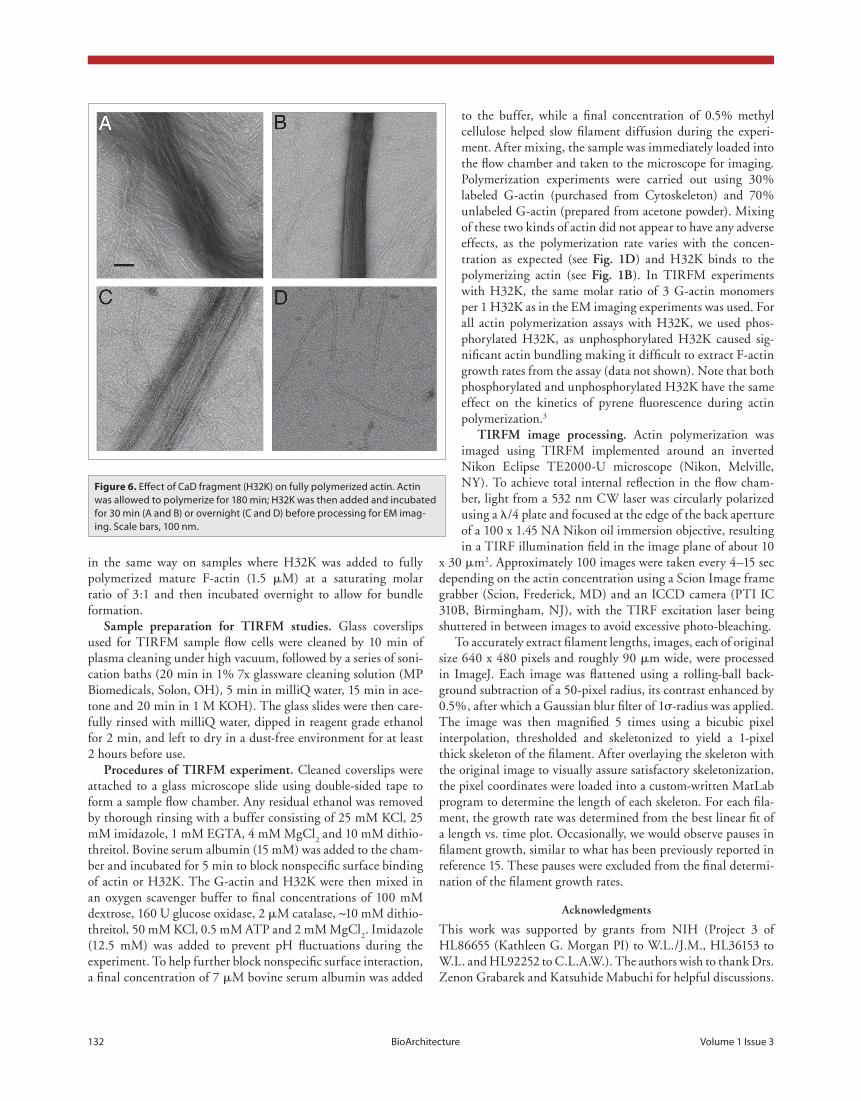
We also prepared a sample by adding H32K to fully polymer-ized actin (at the 180 min time point) and then allowing the mix-ture to incubate for 30 min. The resulting filaments were smooth, but eventually also became bundled (fig. 6). It is noteworthy that after prolonged incubation, the H32K-containing filaments appeared more brittle and easily fragmented (fig. 6c and d).
length over time, the filament growth rate can be determined, as shown in figure 1c. Growth rates of several filaments were measured in this fashion at concentrations of 0.4, 0.8 and 1.2 μM G-actin, both in the presence and absence of pH32K (fig. 1d). Neither the calculated growth rates at each actin concentra-tion, nor the concentration dependence of the growth rate (fig. 1d) showed significant differences between the samples in the presence and absence of pH32K, confirming that the initial addi-tion of H32K to G-actin does not affect the resulting filament growth rate. Moreover, binding of pH32K to actin was ascer-tained by SDS-PAGE after centrifugation of the actin sample polymerized in the presence of pH32K as in the TIRFM assays (fig. 1b; lanes 1 and 2).
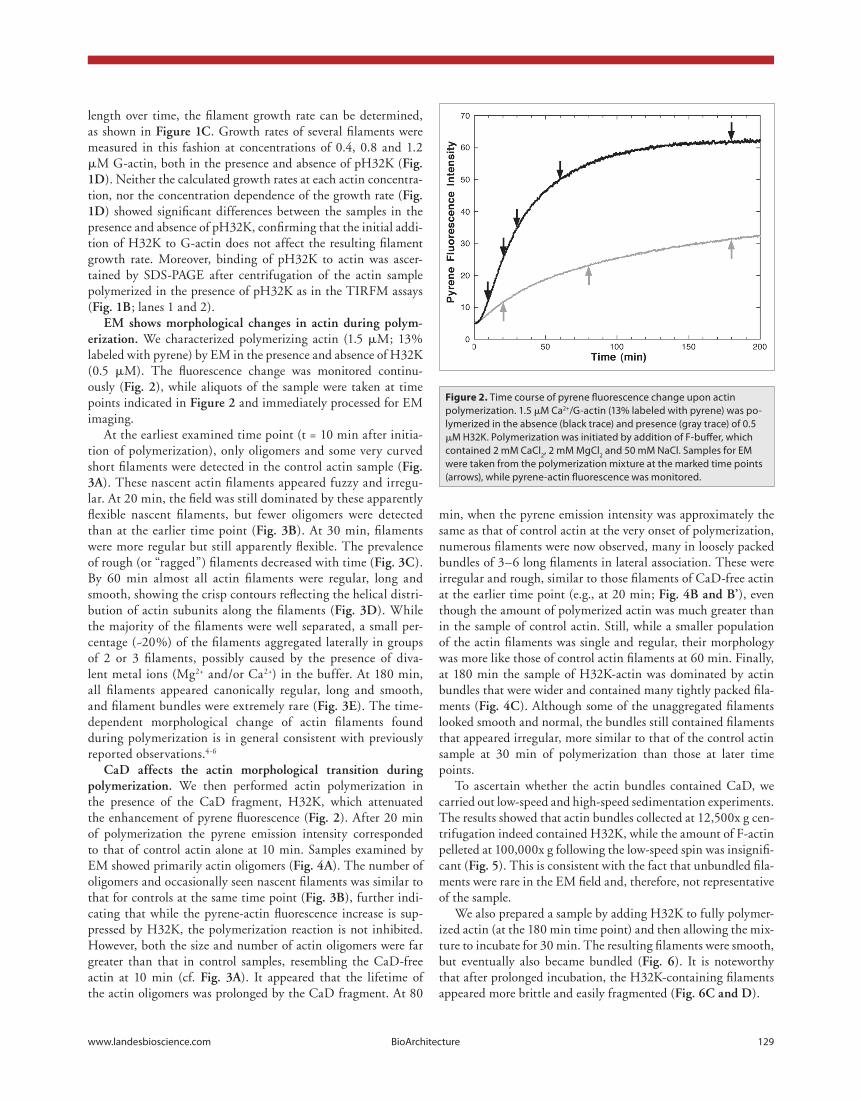
em shows morphological changes in actin during polym-erization. We characterized polymerizing actin (1.5 μM; 13% labeled with pyrene) by EM in the presence and absence of H32K (0.5 μM). The fluorescence change was monitored continu-ously (fig. 2), while aliquots of the sample were taken at time points indicated in figure 2 and immediately processed for EM imaging.
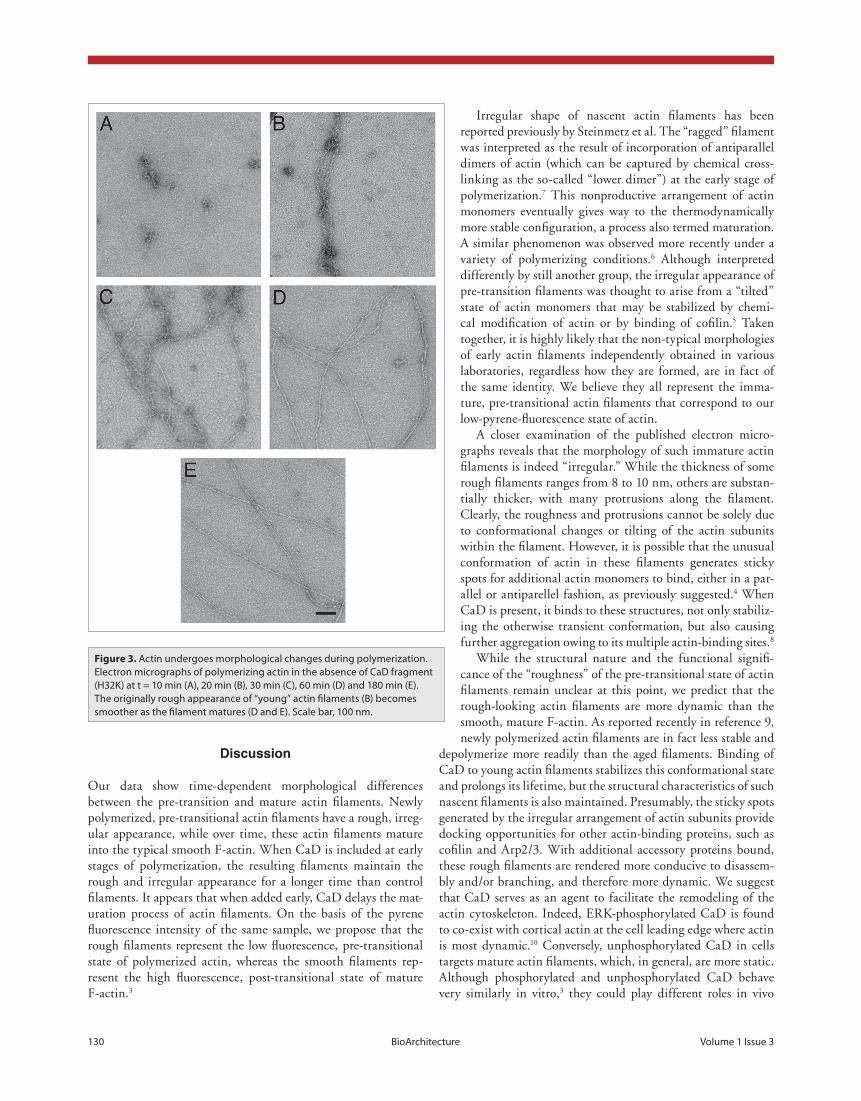
At the earliest examined time point (t = 10 min after initia-tion of polymerization), only oligomers and some very curved short filaments were detected in the control actin sample (fig. 3a). These nascent actin filaments appeared fuzzy and irregu-lar. At 20 min, the field was still dominated by these apparently flexible nascent filaments, but fewer oligomers were detected than at the earlier time point (fig. 3b). At 30 min, filaments were more regular but still apparently flexible. The prevalence of rough (or “ragged”) filaments decreased with time (fig. 3c). By 60 min almost all actin filaments were regular, long and smooth, showing the crisp contours reflecting the helical distri-bution of actin subunits along the filaments (fig. 3d). While the majority of the filaments were well separated, a small per-centage (~20%) of the filaments aggregated laterally in groups of 2 or 3 filaments, possibly caused by the presence of diva-lent metal ions (Mg2+ and/or Ca2+) in the buffer. At 180 min, all filaments appeared canonically regular, long and smooth, and filament bundles were extremely rare (fig. 3e). The time-dependent morphological change of actin filaments found during polymerization is in general consistent with previously reported observations.4-6
cad affects the actin morphological transition during polymerization. We then performed actin polymerization in the presence of the CaD fragment, H32K, which attenuated the enhancement of pyrene fluorescence (fig. 2). After 20 min of polymerization the pyrene emission intensity corresponded to that of control actin alone at 10 min. Samples examined by EM showed primarily actin oligomers (fig. 4a). The number of oligomers and occasionally seen nascent filaments was similar to that for controls at the same time point (fig. 3b), further indi-cating that while the pyrene-actin fluorescence increase is sup-pressed by H32K, the polymerization reaction is not inhibited. However, both the size and number of actin oligomers were far greater than that in control samples, resembling the CaD-free actin at 10 min (cf. fig. 3a). It appeared that the lifetime of the actin oligomers was prolonged by the CaD fragment. At 80
Figure 2. Time course of pyrene fluorescence change upon actin polymerization. 1.5 μM Ca2+/G-actin (13% labeled with pyrene) was po-lymerized in the absence (black trace) and presence (gray trace) of 0.5 μM H32K. Polymerization was initiated by addition of F-buffer, which contained 2 mM CaCl2, 2 mM MgCl2 and 50 mM NaCl. Samples for EM were taken from the polymerization mixture at the marked time points (arrows), while pyrene-actin fluorescence was monitored.
130 BioArchitecture Volume 1 Issue 3
Irregular shape of nascent actin filaments has been reported previously by Steinmetz et al. The “ragged” filament was interpreted as the result of incorporation of antiparallel dimers of actin (which can be captured by chemical cross-linking as the so-called “lower dimer”) at the early stage of polymerization.7 This nonproductive arrangement of actin monomers eventually gives way to the thermodynamically more stable configuration, a process also termed maturation. A similar phenomenon was observed more recently under a variety of polymerizing conditions.6 Although interpreted differently by still another group, the irregular appearance of pre-transition filaments was thought to arise from a “tilted” state of actin monomers that may be stabilized by chemi-cal modification of actin or by binding of cofilin.5 Taken together, it is highly likely that the non-typical morphologies of early actin filaments independently obtained in various laboratories, regardless how they are formed, are in fact of the same identity. We believe they all represent the imma-ture, pre-transitional actin filaments that correspond to our low-pyrene-fluorescence state of actin.
A closer examination of the published electron micro-graphs reveals that the morphology of such immature actin filaments is indeed “irregular.” While the thickness of some rough filaments ranges from 8 to 10 nm, others are substan-tially thicker, with many protrusions along the filament. Clearly, the roughness and protrusions cannot be solely due to conformational changes or tilting of the actin subunits within the filament. However, it is possible that the unusual conformation of actin in these filaments generates sticky spots for additional actin monomers to bind, either in a par-allel or antiparellel fashion, as previously suggested.4 When CaD is present, it binds to these structures, not only stabiliz-ing the otherwise transient conformation, but also causing further aggregation owing to its multiple actin-binding sites.8
While the structural nature and the functional signifi-cance of the “roughness” of the pre-transitional state of actin filaments remain unclear at this point, we predict that the rough-looking actin filaments are more dynamic than the smooth, mature F-actin. As reported recently in reference 9, newly polymerized actin filaments are in fact less stable and
depolymerize more readily than the aged filaments. Binding of CaD to young actin filaments stabilizes this conformational state and prolongs its lifetime, but the structural characteristics of such nascent filaments is also maintained. Presumably, the sticky spots generated by the irregular arrangement of actin subunits provide docking opportunities for other actin-binding proteins, such as cofilin and Arp2/3. With additional accessory proteins bound, these rough filaments are rendered more conducive to disassem-bly and/or branching, and therefore more dynamic. We suggest that CaD serves as an agent to facilitate the remodeling of the actin cytoskeleton. Indeed, ERK-phosphorylated CaD is found to co-exist with cortical actin at the cell leading edge where actin is most dynamic.10 Conversely, unphosphorylated CaD in cells targets mature actin filaments, which, in general, are more static. Although phosphorylated and unphosphorylated CaD behave very similarly in vitro,3 they could play different roles in vivo
Discussion
Our data show time-dependent morphological differences between the pre-transition and mature actin filaments. Newly polymerized, pre-transitional actin filaments have a rough, irreg-ular appearance, while over time, these actin filaments mature into the typical smooth F-actin. When CaD is included at early stages of polymerization, the resulting filaments maintain the rough and irregular appearance for a longer time than control filaments. It appears that when added early, CaD delays the mat-uration process of actin filaments. On the basis of the pyrene fluorescence intensity of the same sample, we propose that the rough filaments represent the low fluorescence, pre-transitional state of polymerized actin, whereas the smooth filaments rep-resent the high fluorescence, post-transitional state of mature F-actin.3
Figure 3. Actin undergoes morphological changes during polymerization. Electron micrographs of polymerizing actin in the absence of CaD fragment (H32K) at t = 10 min (A), 20 min (B), 30 min (C), 60 min (D) and 180 min (E). The originally rough appearance of “young” actin filaments (B) becomes smoother as the filament matures (D and E). Scale bar, 100 nm.
www.landesbioscience.com BioArchitecture 131
because of the phosphorylation-dependent translocation of CaD.10 Thus phosphorylation of CaD and the time factor owing to the movement from cytosol to the leading edge may enable CaD to modulate the actin dynamics both spatially and temporally. Additional studies are currently underway to validate such a hypothesis.
Methods and Materials
protein preparation. Actin was prepared either from acetone-dried rabbit skeletal muscle (for EM and fluorescence studies) as previously described in reference 3, or from acetone powder of chicken pectoralis muscle (for TIRFM studies), both hav-ing the same amino acid sequence for actin,11 following the established procedure by Straub12 with the modifications of Drabikowski and Gergely.13 For TIRFM studies monomeric actin was further purified on a G-75 Sephadex column and collected in G-buffer (0.005% NaN
3, 0.2 mM CaCl
2, 0.2 mM
ATP, 1 mM dithiothreitol and 2 mM Tris(hydroxy-methyl)-aminomethane, pH 8.0) before being snap-frozen in dry ice ethanol and stored at -80°C. The recombinant His-tagged human C-terminal CaD fragment, H32K, was expressed in bacteria and purified as previously described in reference 3. ERK-phosphorylated H32K was prepared as described in ref-erence 3.
labeling of actin. For EM and fluorescence studies, actin was mixed with pyrene-labeled actin, which was prepared by reacting with pyrene-maleimide as previously described in reference 3. For TIRFM studies, lyophilized rabbit skeletal muscle G-actin labeled with rhodamine on surface lysines to a labeling concentration of roughly 0.5 dye residue/monomer was purchased from Cytoskeleton (Denver, CO) and stored at -80°C until use. 50 μL low salt buffer (0.2 mM CaCl
2, 0.2 mM ATP,
10 mM dithiothreitol and 5 mM Tris-HCl, pH 8.0) was added to 20 μg of lyophilized actin and was incubated on ice for 1 hour. The resuspended actin was polymerized at room temperature for 2 hours by adding a concentrated inducer buffer for a final con-centration of 50 mM KCl, 2 mM MgCl
2, 1 mM ATP and 1 mM
dithiothreitol. After polymerization, the actin was converted to G-actin by dialysis against 500 mL G-buffer. The dialysis buf-fer was changed every 12 hours over two days, for a total of four buffer exchanges.
negative staining electron microscopy. Electron microscopy (EM) was performed to visualize actin filament maturation. Polymerization of Ca-ATP-G-actin (1.5 μM; in G-buffer) alone or in the presence of H32K (0.5 μM) was induced by inorganic salts (0.2 mM CaCl
2, 2 mM MgCl
2, 50 mM NaCl) at 25°C. At
various points after the initiation of actin polymerization, moni-tored by the increase of pyrene-actin fluorescence (λ
ex = 365 nm;
λem
= 407 nm), 15-μL aliquots of actin solutions were applied to carbon-coated 200 mesh copper grids. Samples were allowed to adsorb to grids for 30 s, wicked to remove excess solution, and negatively stained with 1% (wt/vol) uranyl acetate for 1 min and allowed to dry.14 EM was performed using a Philips CM120 transmission electron microscope at an accelerating voltage of 80 kV under low dose conditions. Additional EM was performed
Figure 4. Effect of CaD fragment (H32K) on polymerizing actin. Electron micrographs of actin polymerizing in the presence of H32K at t = 20 min (A), t = 80 min (B and B’) and t = 180 min (C). As in the absence of H32K, the rough appearance of “young” actin filaments (A) becomes smoother as the filament matures (B). However, CaD (or its fragment) prolongs the lifetime of the young state, delaying this maturation process. Scale bars, 100 nm.
Figure 5. Actin bundles indeed contained CaD fragment (H32K). Results of 10% SDS-PAGE analysis of actin samples polymerized in the presence (Lanes 1–4) and absence (Lanes 5–8) of H32K for 180 min. Samples were first centrifuged at 12,500x g for 5 min (Lanes 1 and 2; Lanes 5 and 6), followed by high-speed centrifugation at 100,000x g for 20 min (Lanes 3 and 4; Lanes 7 and 8). Also shown are the molecular weight markers (M) and H32K standard (Lane 9). For gel loading, one fifteenth of the supernatant fractions (S) and one third of the pellet fractions (P) were used. It can be seen that under these conditions actin alone formed few bundles (Lane 6), whereas the sample with H32K contained mainly bundled actin (Lane 2) and few free filaments (Lane 3).
132 BioArchitecture Volume 1 Issue 3
to the buffer, while a final concentration of 0.5% methyl cellulose helped slow filament diffusion during the experi-ment. After mixing, the sample was immediately loaded into the flow chamber and taken to the microscope for imaging. Polymerization experiments were carried out using 30% labeled G-actin (purchased from Cytoskeleton) and 70% unlabeled G-actin (prepared from acetone powder). Mixing of these two kinds of actin did not appear to have any adverse effects, as the polymerization rate varies with the concen-tration as expected (see fig. 1d) and H32K binds to the polymerizing actin (see fig. 1b). In TIRFM experiments with H32K, the same molar ratio of 3 G-actin monomers per 1 H32K as in the EM imaging experiments was used. For all actin polymerization assays with H32K, we used phos-phorylated H32K, as unphosphorylated H32K caused sig-nificant actin bundling making it difficult to extract F-actin growth rates from the assay (data not shown). Note that both phosphorylated and unphosphorylated H32K have the same effect on the kinetics of pyrene fluorescence during actin polymerization.3
tirfm image processing. Actin polymerization was imaged using TIRFM implemented around an inverted Nikon Eclipse TE2000-U microscope (Nikon, Melville, NY). To achieve total internal reflection in the flow cham-ber, light from a 532 nm CW laser was circularly polarized using a λ/4 plate and focused at the edge of the back aperture of a 100 x 1.45 NA Nikon oil immersion objective, resulting in a TIRF illumination field in the image plane of about 10
x 30 μm2. Approximately 100 images were taken every 4–15 sec depending on the actin concentration using a Scion Image frame grabber (Scion, Frederick, MD) and an ICCD camera (PTI IC 310B, Birmingham, NJ), with the TIRF excitation laser being shuttered in between images to avoid excessive photo-bleaching.
To accurately extract filament lengths, images, each of original size 640 x 480 pixels and roughly 90 μm wide, were processed in ImageJ. Each image was flattened using a rolling-ball back-ground subtraction of a 50-pixel radius, its contrast enhanced by 0.5%, after which a Gaussian blur filter of 1σ-radius was applied. The image was then magnified 5 times using a bicubic pixel interpolation, thresholded and skeletonized to yield a 1-pixel thick skeleton of the filament. After overlaying the skeleton with the original image to visually assure satisfactory skeletonization, the pixel coordinates were loaded into a custom-written MatLab program to determine the length of each skeleton. For each fila-ment, the growth rate was determined from the best linear fit of a length vs. time plot. Occasionally, we would observe pauses in filament growth, similar to what has been previously reported in reference 15. These pauses were excluded from the final determi-nation of the filament growth rates.
acknowledgments
This work was supported by grants from NIH (Project 3 of HL86655 (Kathleen G. Morgan PI) to W.L./J.M., HL36153 to W.L. and HL92252 to C.L.A.W.). The authors wish to thank Drs. Zenon Grabarek and Katsuhide Mabuchi for helpful discussions.
in the same way on samples where H32K was added to fully polymerized mature F-actin (1.5 μM) at a saturating molar ratio of 3:1 and then incubated overnight to allow for bundle formation.
sample preparation for tirfm studies. Glass coverslips used for TIRFM sample flow cells were cleaned by 10 min of plasma cleaning under high vacuum, followed by a series of soni-cation baths (20 min in 1% 7x glassware cleaning solution (MP Biomedicals, Solon, OH), 5 min in milliQ water, 15 min in ace-tone and 20 min in 1 M KOH). The glass slides were then care-fully rinsed with milliQ water, dipped in reagent grade ethanol for 2 min, and left to dry in a dust-free environment for at least 2 hours before use.
procedures of tirfm experiment. Cleaned coverslips were attached to a glass microscope slide using double-sided tape to form a sample flow chamber. Any residual ethanol was removed by thorough rinsing with a buffer consisting of 25 mM KCl, 25 mM imidazole, 1 mM EGTA, 4 mM MgCl
2 and 10 mM dithio-
threitol. Bovine serum albumin (15 mM) was added to the cham-ber and incubated for 5 min to block nonspecific surface binding of actin or H32K. The G-actin and H32K were then mixed in an oxygen scavenger buffer to final concentrations of 100 mM dextrose, 160 U glucose oxidase, 2 μM catalase, ~10 mM dithio-threitol, 50 mM KCl, 0.5 mM ATP and 2 mM MgCl
2. Imidazole
(12.5 mM) was added to prevent pH fluctuations during the experiment. To help further block nonspecific surface interaction, a final concentration of 7 μM bovine serum albumin was added
Figure 6. Effect of CaD fragment (H32K) on fully polymerized actin. Actin was allowed to polymerize for 180 min; H32K was then added and incubated for 30 min (A and B) or overnight (C and D) before processing for EM imag-ing. Scale bars, 100 nm.

www.landesbioscience.com BioArchitecture 133
12. Straub FB. Actin Stud Ins Med Chem Univ Szeged 1942; 2:3-4.
13. Drabikowski W, Gergely J. The effect of the rempera-ture of extraction and of tropomyosin on the viscosity of actin. In: Gergely J, Ed. Biochemistry of Muscle Contraction: Little Brown 1964; 125-31.
14. Moody C, Lehman W, Craig R. Caldesmon and the structure of smooth muscle thin filaments: electron microscopy of isolated thin filaments. J Muscle Res Cell Motil 1990; 11:176-85.
15. Kuhn JR, Pollard TD. Real-time measurements of actin filament polymerization by total internal reflection fluorescence microscopy. Biophys J 2005; 88:1387-402.
16. Huxley HE, Brown W. The low-angle x-ray diagram of vertebrate striated muscle and its behaviour during contraction and rigor. J Mol Biol 1967; 30:383-434.
7. Millonig R, Salvo H, Aebi U. Probing actin polym-erization by intermolecular cross-linking. J Cell Biol 1988; 106:785-96.
8. Huang R, Li L, Guo H, Wang CLA. Caldesmon bind-ing to actin is regulated by calmodulin and phosphory-lation via different mechanisms. Biochemistry 2003; 42:2513-23.
9. Kueh HY, Brieher WM, Mitchison TJ. Dynamic sta-bilization of actin filaments. Proc Natl Acad Sci USA 2008; 105:16531-6.
10. Kordowska J, Hetrick T, Adam LP, Wang CLA. Phosphorylated l-caldesmon is involved in disassembly of actin stress fibers and postmitotic spreading. Exp Cell Res 2006; 312:95-110.
11. Vandekerckhove J, Weber K. The amino acid sequence of actin from chicken skeletal muscle actin and chick-en gizzard smooth muscle actin. FEBS Lett 1979; 102:219-22.
references1. Wang CLA. Caldesmon and the Regulation of
Cytoskeletal Functions. Adv Exp Med Biol 2008; 644: 250-72.
2. Marston SB, Huber PAJ. Caldesmon. In: Bárány M, Ed. Biochemistry of Smooth Muscle Contraction. San Diego, CA: Academic Press, Inc. 1996; 77-90.
3. Huang R, Grabarek Z, Wang CLA. Differential effects of caldesmon on the intermediate conformational states of polymerizing actin. J Biol Chem 2010; 285:71-9.
4. Steinmetz MO, Goldie KN, Aebi U. A correlative anal-ysis of actin filament assembly, structure and dynamics. J Cell Biol 1997; 138:559-74.
5. Orlova A, Shvetsov A, Galkin VE, et al. Actin-destabilizing factors disrupt filaments by means of a time reversal of polymerization. Proc Natl Acad Sci USA 2004; 101:17664-8.
6. Galinska-Rakoczy A, Wawro B, Strzelecka-Golaszewska H. New aspects of the spontaneous polymerization of actin in the presence of salts. J Mol Biol 2009; 387:869-82.
Related Documents











