THE ANATOMICAL RECORD 217:256-262 (1987) Structural Organization of the Vertebral Artery in the Giraffe (GiraHa camelopardalis) JAMES KIRUMBI KIMANI Department of Human Anatomy, University of Nairobi, Nairobi, Kenya ABSTRACT Vertebral arteries in the giraffe were removed and their structural organization was studied with light microscopy. It has been shown that the caudal segment of the vertebral artery extending to the level of the seventh cervical vertebra has a largely elastic structure, while the cranial segment has a muscular structure. The transition of the arterial wall normally occurs between the seventh and fifth cervical vertebral levels, and involves diminution of elastic tissue in the luminal portion of the tunica media and simultaneous increase of the smooth muscle content. The smooth muscle cells in the cranial portion show a clustered arrange- ment consisting of several smooth muscle fibres. The individual clusters are sepa- rated by connective tissue septa from which finer strands extend to surround individual muscle fibres. It is suggested that the vertebral and carotid arteries though found in the neck have a different structural organization, which, in turn, may imply that the two blood vessels are subjected to different haemodynamic demands. The findings of this study therefore render further support to the state- ment that the largely muscular structure of the common carotid artery in the giraffe may be related to the unique cerebral haemodynamics of this animal, rather than being a common feature of the blood vessels situated in the neck. A corollary to this suggestion is the fact that, in the giraffe, as in most other Artiodactyls, the vertebral blood does not participate in the supply of cephalic structures because it is confined to the cervical region by the pressure barrier in the carotid-vertebral anastomosis. The gross anatomical organization of the carotid arte- rial system of the giraffe has been studied with a view to establishing the mechanisms that underlie the ability of this animal to lower or raise its head without damag- ing the cerebral blood vessels (Lawrence and Rewell, 1984; Goetz and Keen, 1957; Goetz, 1962). These earlier studies have provided limited informa- tion as the pattern of distribution of the carotid arteries in the giraffe is remarkably similar to that of the other Artwdactyls (Daniel et al., 1953; Appleton and Waites, 1957; Waites, 1960; Baldwin and Bell, 1963a-c; Bald- win, 1964, Du Boulay and Verity, 1973). Consequently, neither the fact that the external carotid artery is the main source of blood supply to the brain, nor the pres- ence of an intracranial rete mirabile, or the occurrence of an extracranial carotid-vertebral anastomosis is a unique anatomical feature of the giraffe cerebral vascu- lar system. The structural organization of the carotid arterial sys- tem of the giraffe has been investigated in recent stud- ies, which show that the tunica media has a largely muscular structure (Kimani, 1979, 1981, 1983; Kimani and Mungai, 1983). It has also been demonstrated that in the arteries of hybrid type (Bloom and Fawcett, 1968) such as the cranial portion of the bicarotid trunk [a short trunk measuring about 10 cm which divides into the right and left common carotid arteries (Sisson and Grossman, 197511 and the proximal 20 cm or so of the common carotid artery, the tunica media has the struc- ture of an elastic artery in the luminal third and of a muscular artery in the outer two-thirds (Kimani, 1979, 1983). In the arteries of the hybrid type of most other animals the muscular tissue occupies the luminal por- tion and elastic tissue the outer portion (Katzberg, 1966; Bloom and Fawcett, 1968;Rees, 1968; Somlyo and Som- lyo, 1968). Little is known, however, as to what extent the unique mode of arterial transition in the carotid arterial system of the giraffe is related to the haemodynamic stresses imposed by its long neck. This is partly due to the fact that previous studies on the structure of the common carotid arteries of other long-neckedanimals such as the camel are relatively scanty, and not much is known about other blood vessels of the giraffe in the neck such as the vertebral artery. This communication describes the structural organization of the giraffe vertebral ar- tery which hitherto has not been studied. MATERIALS AND METHODS Materials used in this study were obtained from five adult giraffes which were shot during routine culls un- der a permit issued by the Ministry of Tourism and Wildlife of the Government of Kenya. The neck was separated from the trunk between the seventh cervical Received September 19, 1985; accepted September 19, 1986. This work is dedicated to our second born, the late Miss Nyawira Kirumbi. 0 1987 ALAN R. LISS, INC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE ANATOMICAL RECORD 217:256-262 (1987)
Structural Organization of the Vertebral Artery in the Giraffe (GiraHa camelopardalis)
JAMES KIRUMBI KIMANI Department of Human Anatomy, University of Nairobi, Nairobi, Kenya
ABSTRACT Vertebral arteries in the giraffe were removed and their structural organization was studied with light microscopy. It has been shown that the caudal segment of the vertebral artery extending to the level of the seventh cervical vertebra has a largely elastic structure, while the cranial segment has a muscular structure. The transition of the arterial wall normally occurs between the seventh and fifth cervical vertebral levels, and involves diminution of elastic tissue in the luminal portion of the tunica media and simultaneous increase of the smooth muscle content. The smooth muscle cells in the cranial portion show a clustered arrange- ment consisting of several smooth muscle fibres. The individual clusters are sepa- rated by connective tissue septa from which finer strands extend to surround individual muscle fibres. It is suggested that the vertebral and carotid arteries though found in the neck have a different structural organization, which, in turn, may imply that the two blood vessels are subjected to different haemodynamic demands. The findings of this study therefore render further support to the state- ment that the largely muscular structure of the common carotid artery in the giraffe may be related to the unique cerebral haemodynamics of this animal, rather than being a common feature of the blood vessels situated in the neck. A corollary to this suggestion is the fact that, in the giraffe, as in most other Artiodactyls, the vertebral blood does not participate in the supply of cephalic structures because it is confined to the cervical region by the pressure barrier in the carotid-vertebral anastomosis.
The gross anatomical organization of the carotid arte- rial system of the giraffe has been studied with a view to establishing the mechanisms that underlie the ability of this animal to lower or raise its head without damag- ing the cerebral blood vessels (Lawrence and Rewell, 1984; Goetz and Keen, 1957; Goetz, 1962).
These earlier studies have provided limited informa- tion as the pattern of distribution of the carotid arteries in the giraffe is remarkably similar to that of the other Artwdactyls (Daniel et al., 1953; Appleton and Waites, 1957; Waites, 1960; Baldwin and Bell, 1963a-c; Bald- win, 1964, Du Boulay and Verity, 1973). Consequently, neither the fact that the external carotid artery is the main source of blood supply to the brain, nor the pres- ence of an intracranial rete mirabile, or the occurrence of an extracranial carotid-vertebral anastomosis is a unique anatomical feature of the giraffe cerebral vascu- lar system.
The structural organization of the carotid arterial sys- tem of the giraffe has been investigated in recent stud- ies, which show that the tunica media has a largely muscular structure (Kimani, 1979, 1981, 1983; Kimani and Mungai, 1983). It has also been demonstrated that in the arteries of hybrid type (Bloom and Fawcett, 1968) such as the cranial portion of the bicarotid trunk [a short trunk measuring about 10 cm which divides into the right and left common carotid arteries (Sisson and Grossman, 197511 and the proximal 20 cm or so of the common carotid artery, the tunica media has the struc-
ture of an elastic artery in the luminal third and of a muscular artery in the outer two-thirds (Kimani, 1979, 1983). In the arteries of the hybrid type of most other animals the muscular tissue occupies the luminal por- tion and elastic tissue the outer portion (Katzberg, 1966; Bloom and Fawcett, 1968; Rees, 1968; Somlyo and Som- lyo, 1968).
Little is known, however, as to what extent the unique mode of arterial transition in the carotid arterial system of the giraffe is related to the haemodynamic stresses imposed by its long neck. This is partly due to the fact that previous studies on the structure of the common carotid arteries of other long-necked animals such as the camel are relatively scanty, and not much is known about other blood vessels of the giraffe in the neck such as the vertebral artery. This communication describes the structural organization of the giraffe vertebral ar- tery which hitherto has not been studied.
MATERIALS AND METHODS Materials used in this study were obtained from five
adult giraffes which were shot during routine culls un- der a permit issued by the Ministry of Tourism and Wildlife of the Government of Kenya. The neck was separated from the trunk between the seventh cervical
Received September 19, 1985; accepted September 19, 1986. This work is dedicated to our second born, the late Miss Nyawira
Kirumbi.
0 1987 ALAN R. LISS, INC.
VERTEBRAL ARTERY IN THE GIRAFFE 257
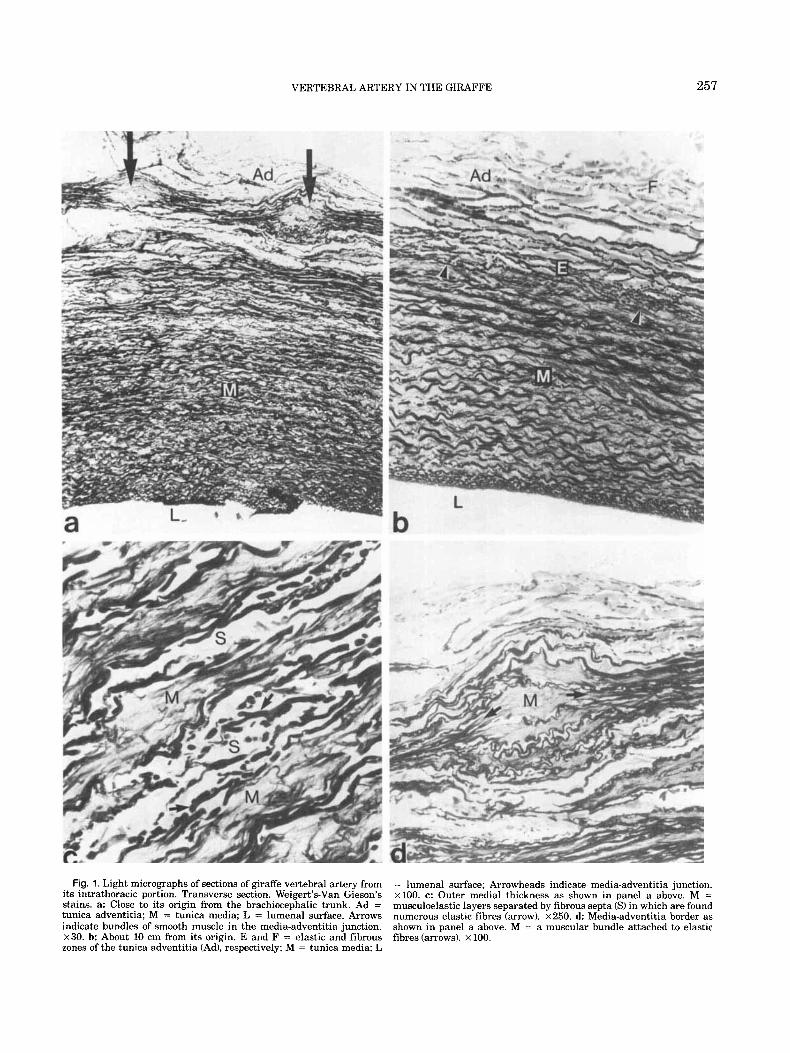
Fig. 1 . Light micrographs of sections of giraffe vertebral artery from its intrathoracic portion. Transverse section. Weigert’s-Van Gieson’s stains. a: Close to its origin from the brachiocephalic trunk. Ad = tunica adventitia; M = tunica media; L = lumenal surface. Arrows indicate bundles of smooth muscle in the media-adventitia junction. x30. b: About 10 cm from its origin. E and F = elastic and fibrous zones of the tunica adventitia (Ad), respectively; M = tunica media; L
= lumenal surface; Arrowheads indicate media-adventitia junction. x100. c: Outer medial thickness as shown in panel a above. M = musculoelastic layers separated by fibrous septa (S) in which are found numerous elastic fibres (arrow). x250. d: Media-adventitia border as shown in panel a above. M = a muscular bundle attached to elastic fibres (arrows). x 100.
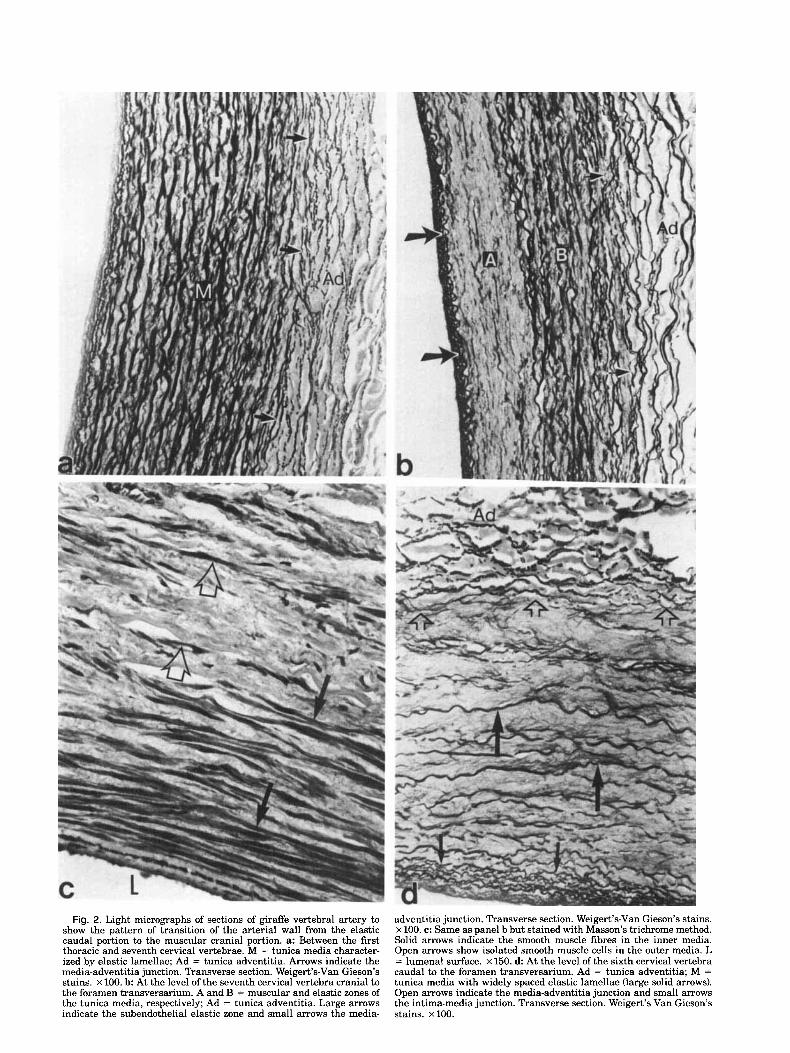
Fig. 2. Light micrographs of sections of giraffe vertebral artery to show the pattern of transition of the arterial wall from the elastic caudal portion to the muscular cranial portion. a: Between the first thoracic and seventh cervical vertebrae. M = tunica media character- ized by elastic lamellae; Ad = tunica adventitia. Arrows indicate the media-adventitia junction. Transverse section. Weigert’s-Van Gieson’s stains. x 100. b At the level of the seventh cervical vertebra cranial to the foramen transversarium. A and B = muscular and elastic zones of the tunica media, respectively; Ad = tunica adventitia. Large arrows indicate the subendothelial elastic zone and small arrows the media-
adventitia junction. Transverse section. Weigert’s-Van Gieson’s stains. x 100. c: Same as panel b but stained with Masson’s trichrome method. Solid arrows indicate the smooth muscle fibres in the inner media. Open arrows show isolated smooth muscle cells in the outer media. L = lumenal surface. X 150. d At the level of the sixth cervical vertebra caudal to the foramen transversarium. Ad = tunica adventitia; M = tunica media with widely spaced elastic lamellae (large solid arrows). Open arrows indicate the media-adventitia junction and small arrows the intima-media junction. Transverse section. Weigert’s-Van Gieson’s stains. x 100.

VERTEBRAL ARTERY IN THE GIRAFFE 259
and first thoracic vertebrae and later fixed by intraca- rotid and intravertebral perfusion with 10% formal-sa- line solution. The heart and the major blood vessels arising from it were removed in the field and immedi- ately fixed by immersion in a solution of 10% formal- saline.
Material for light microscopy was obtained serially at regular intervals of 5 cm caudocranially up to the ter- mination of the vertebral artery. Histological specimens were also removed from the carotid-vertebral anasto- motic branch and then processed for paraffin-embedding sectioning and staining with (a) Weigert’s resorcin fuch- sin counterstained with Van Gieson’s stain and (b) Mas- son’s trichrome stain.
RESULTS The vertebral arteries in the giraffe, one on each side,
arise from the brachiocephalic trunk and ascend in the neck through the foramina in the transverse processes of the cervical vertebrae. Each vertebral artery termi- nates at the junction of the second and third cervical vertebrae by dividing into a superficial branch and a deep branch. The latter enters the vertebral canal through the intervertebral foramen and gives off an ascending branch which anastomoses with the descend- ing occipital artery (Goetz and Keen, 1957; Kimani, 1979).
The proximal (caudal) portion of the vertebral artery extending from its origin to about the level of the sev- enth cervical vertebra has a largely elastic structure (Fig. 1). The tunica media consists of elastic tissue ar- ranged in form of elastic lamellae (Fig. lb). In the prox- imal segment, however, the outer medial thickness has a fibromuscular structure consisting of musculoelastic layers which are separated by septa of circular and longitudinal elastic and collagen fibers (Fig. lc). A num- ber of muscular bundles or nests which are intercon- nected by elastic layers are found in the media- adventitia junction (Fig. la,d).
The outer fibromuscular zone diminishes as the artery approaches the foramen transversarium of the seventh cervical vertebra. At this level, the tunica media con- sists of closely arranged elastic lamellae which are sep- arated by interlamellar spaces containing smooth muscle cells and finer elastic fibers (Fig. 2a). The tunica adven- titia, on the other hand, consists of interconnecting bun- dles of elastic fibers with a preferred longitudinal alignment between which are bundles of collagen fibers (Fig. 2a). The tunica intima shows a subendothelial zone containing numerous elastic fibers (Fig. 2a).
The transition of the arterial wall from the caudal elastic to the cranial muscular portion is gradual and involves diminution of elastic tissue mainly on the lu- mina1 side of the tunica media (Fig. 2b,c). This trans- mural zonation is, however, lost in the cranial segment of the transition zone in which the tunica media consists of widely scattered elastic lamellae (Fig. 2d). The inti- mal subendothelial zone of elastic fibers is also clearly visible and consists of longitudinal bundles of elastic fibers forming several layers on the luminal side of the internal elastic lamina (Fig. 2b). The tunica adventitia consists of an outer fibrous zone with widely scattered elastic fibers and an inner zone which is largely elastic (Fig. 2b,d).
The cranial portion of the vertebral artery extending from about the level of the fifth cervical vertebra is
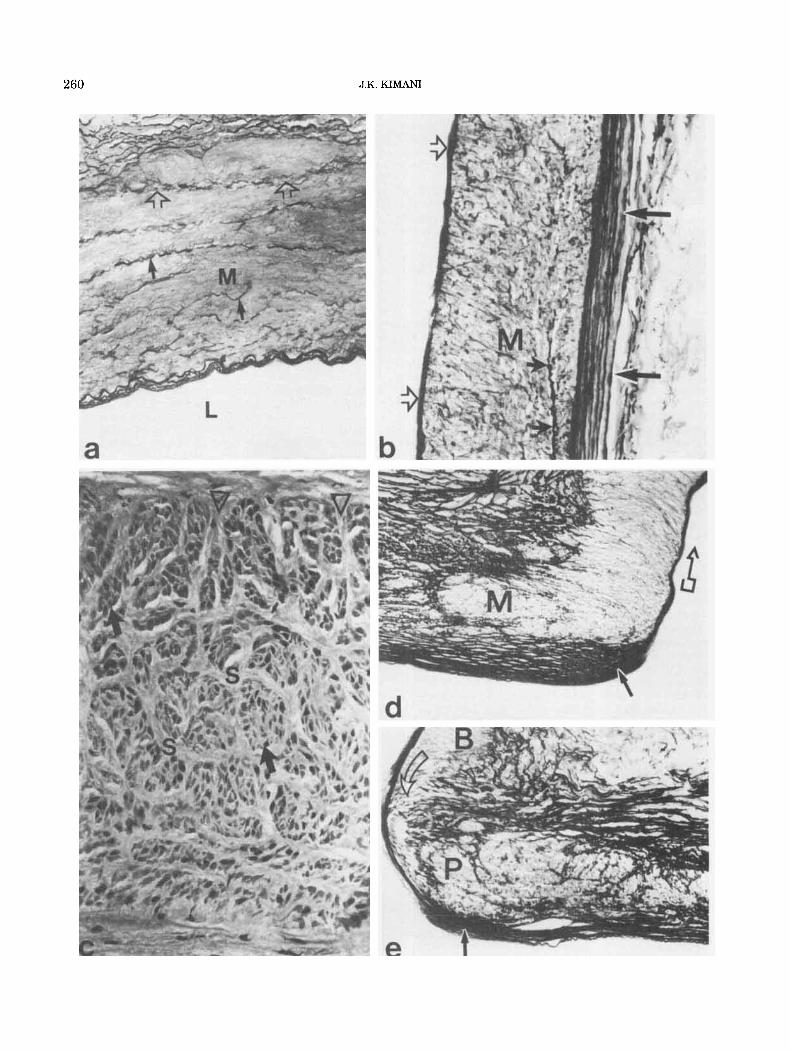
characterized by a largely muscular tunica media in which only a few strands of elastic fibers are found (Fig. 3a). In longitudinally cut sections, the vessel wall is seen to consist of a luminal elastic zone of internal elastic lamina, a medial muscular zone and an outer adventi- tial coat (Fig. 3b). Occasionally isolated smooth muscle cells which are typically longitudinal in their orienta- tion may be found in the deeper adventitial coat.
The muscular tissue of the tunica media is organized in the form of small bundles especially in the outer media (Fig. 34. These muscular bundles are separated by connective tissue septa which are largely elastic, and from which smaller strands arise and permeate between the smooth muscle bundles (Fig. 3c). The fibrous septa of the outer medial thickness are continuous with the fibroelastic network of the deep adventitia (Fig. 3a,c).
Modifications of the arterial wall structure were found at branching sites of both small and the large segmental arteries. The tunica intima is thickened to form what has previously been called an “intimal cushion” (Fig. 3d,e). The intimal thickening is characterized in the main by numerous elastic laminae which interconnect extensively to form a latticework showing a preferential longitudinal orientation. The elastic layers terminate abruptly at the mouth of the branching artery by con- verging to form a single internal elastic lamina which continues into the branching artery (Fig. 3d).
At the branching sites of the small muscular arteries from the elastic portion of the vertebral artery, the me- dial elastic tissue of the parent artery is replaced by smooth muscle (Fig. 3d). The muscular process appears to be encapsulated by a condensation of elastic fibers and extends into the branching artery (Fig. 3d). On the cranial aspect of the osteum, the tunica media may or may not show a similar process (Fig. 3e). The tunica intima shows a thickening, but, on the average, such a thickening is smaller than the one seen on the caudal side of the osteum. Histologically, however, the cushion shows a similar elastic structure in which the elastic layers converge to form a single internal elastic lamina which extends into the branching artery (Fig. 3e).
Another structural feature commonly associated with the cranial aspect of the osteum is a medial defect which is seen between the parent artery and the branching artery (Fig. 3el. In such areas of medial defect the in- ternal elastic lamina is the main structure connecting the branching artery to the parent artery (Fig. 3e). An adventitial fibroelastic tissue fills the gap between the media of the two arteries and reaches the internal elastic lamina onto which the elastic fibers are at- tached (Fig. 3e). A number of branching elastic fibers are also seen on the adventitial side running across the me- dial gap between the parent and the branching artery (Fig. 3e).
DISCUSSION The findings reported in this study on the giraffe ver-
tebral artery have shown that the proximal portion which extends up to the level of the seventh cervical vertebra has a largely elastic structure, while the cra- nial portion extending from the fifth cervical vertebral level has a largely muscular structure. The transition wne from the elastic to the muscular parts of the verte- bral artery extends from the upper part of the seventh cervical vertebra to the level of the fifth cervical verte- bra. Whereas the caudal portion of the transition zone
260 J.K. KIMANI

VERTEBRAL ARTERY IN THE GIRAFFE 261
shows a transmural differentiation into a luminal mus- cular zone and an adventitial elastic zone, its cranial portion has widely scattered elastic lamellae in the tun- ica media which are separated by layers of smooth mus- cle. This mode of arterial transition is in contrast to the pattern observed in the carotid system in which the muscular tissue occupies the outer media and elastic tissue the inner media (Franklin and Haynes, 1927; Kimani, 1979,1983). It resembles, however, the mode of arterial transition from elastic to muscular arteries which has been described in previous studies on the short-necked animals (Katzberg, 1966; Bloom and Faw- cett, 1968; Rees, 1968), as well as in the common carotid artery of the camel and the porcupine (personal unpub- lished data).
Observations made in this study have also revealed that the smooth muscle of the cranial portion of the vertebral artery is organized in form of small clusters which are permeated by an extensive network of connec- tive tissue. This fascicular organization of smooth mus- cle is particularly evident in the media-adventitia junction where the connective tissue septa are also con- tinuous with the adventitial fibroelastic tissue. Such clusters, however, are much smaller than the muscular bundles described in the outer media of the carotid ar- tery (Kimani, 1979,1983).
In the vertebral artery, the smooth muscle appears to be uniformly organized throughout the medial thick- ness, with the exception of the occasional bundles in the media-adventitia junction. This is in contrast to the or- ganizational heterogeneity observed in the carotid ar- tery in which the luminal zone consists of isolated smooth muscle fibers, whereas the outer zone has large bundles of smooth muscle cells (Kimani, 1981, 1983). The significance of this finding, it appears, lies in the statement in previous studies that a transmural zona- tion may indicate differences in function between the luminal and outer muscular layers (Somlyo and Somlyo, 1968; Feigl et al., 1976). The largely uniform organiza- tional pattern of smooth muscle in the media of the
Fig. 3. Light micrographs of sections of giraffe vertebral artery at the level of the fourth cervical vertebra, and at branching sites to illustrate modifications of the arterial wall. a: Tunica media (M) show- ing a largely muscular structure and widely scattered elastic fibres (small arrows). Open arrows indicate a fibrous septum in the outer media and its continuity with the adventitial fibroelastic tissue. L = lumenal surface. Transverse section. Weigert’s-Van Gieson’s stains. x 100. b: Longitudinal section to illustrate the mural tissue compo- nents. M = tunica media. Large arrows indicate the adventitial elastic layer and small arrows a fibrous septum in the outer media. Open arrows show the internal elastic lamina. Weigert’s-Van Gieson’s stains. ~100. c: Same as in panel b, but stained with Masson’s trichrome method. S = fibrous septa in the tunica media surrounding clusters of smooth muscle (arrows). Open arrowheads indicate the continuity of the medial fibrous septa with the adventitial fibrous network. x 150. d Proximal portion of a branching point of a small muscular artery from the elastic segment of the vertebral artery. Medial elastic tissue is replaced by smooth muscle (M) that extends into the branching artery (open arrow). Solid arrow indicates an intimal cushion. Longi- tudinal section. Weigert’s-Van Gieson’s stains. x 75. e: Distal portion of a branching point. Note an area of medial defect (open arrow) between the parent (PI and branching (B) arteries. Small arrow indi- cates an intimal cushion. Longitudinal section. Weigert’s-Van Gieson’s stains. ~ 7 5 .
vertebral artery may on the other hand indicate similar- ity of function, although subtle differences may exist such as in their responses to circulating catecholamines andlor neuronal catecholamines. Keatinge and Torrie (1976) have shown that in the sheep carotid artery the luminal muscle cells are more responsive to circulating catecholamines than the outer medial smooth muscle cells. Mekata (1984) has also demonstrated different electrical responses between the outer and inner muscle of rabbit carotid artery to noradrenaline and nerves.
According to Kimani (1979, 1983) the structural orga- nization of the carotid arterial system of the giraffe, characterized, in the main, by a largely muscular struc- ture, may constitute part of the anatomical basis for the remarkable ability of this animal to autoregulate its cranial circulation. In support of this suggestion has been the demonstration that its muscular structure, and especially that of the outer medial thickness appears during the postnatal growth of the animal (Kimani, 1983). This has led to the suggestion that whereas the elastic tissue on the luminal side of the tunica media has a cushioning effect and modulates the distension of the arterial wall in systole, the tone that may be gener- ated andlor maintained in the smooth muscle cells on the adventitial side prevents overdistension of the arte- rial wall (Kimani, 1979). Such a mechanism, if it occurs, may then sustain a relatively high rate of blood flow in diastole as has been shown to be the case in this animal (Van Citters et al., 1968, 1969). Furthermore, evidence does exist that a characteristic feature in the behaviour of the bovine carotid arteries is a contraction which occurs when the internal pressure is raised above 130 cm of water (Jeager, 1962).
It has been pointed out, however, that the systolic pressure generated by the heart of a giraffe is in favor of an unassisted cerebral circulation such as a peristaltic wave along the carotid artery andJor a siphon-like effect of venous blood rushing down the jugular vein (Goetz and Keen, 1957; Goetz et al., 1960; Goetz, 1962). Yet when the head of a recumbent anaesthetized giraffe was raised for a vertical distance of 150 cm above the heart in one experiment, a rise in pressure of only 50 mm Hg could not suffice to maintain blood pressure at the brain level within normal limit against a calculated hydro- static pressure of 100 mmHg (Goetz et al., 1960). It is conceivable, therefore, that the ability of a giraffe to maintain an adequate cerebral perfusion pressure de- pends as much on the pressure developed by the heart as by the morphology of the carotid arterial system and appropriate cardiovascular regulatory mechanisms.
Thus, the fact that the vertebral artery of the giraffe has a different structural organization from the carotid artery (at least in its transition zone) may indicate that the two blood vessels, though found in the neck, are subjected to different haemodynamic demands. In the sheep and goat, for example, vertebral blood does not participate in the supply of cephalic structures because it is confined to the cervical region by the pressure barrier between carotid and vertebral blood in the occip- ito-vertebral anastomosis (Baldwin and Bell, 1963c; Edelman et al., 1972). The cranial arterial system of the giraffe resembles that of the other Artwdactyls, such as the sheep, except for the fact that in the giraffe the anastomotic artery between the carotid and vertebral arterial systems is a direct branch of the carotid artery

262 J.K. KIMANI
(Lawrence and Rewell, 1948; Daniel et al., 1953; Goetz and Keen, 1957; Du Boulay and Verity, 1973; Kimani, 1979). It is conceivable that in the giraffe, as in the sheep, the entire brain and other cephalic structures are supplied from the carotid system, while vertebral blood is confined to the cervical region. Thus whereas the pressure developed by the heart may be adequate for perfusion of the areas supplied by the vertebral arteries, a peripheral auxiliary mechanism may be necessary in the case of the carotid system and this may be reflected in its mural remodelling process during the postnatal growth and development which is characterized by ap- pearance of smooth muscle in the outer medial thickness (Kimani, 1983).
ACKNOWLEDGMENTS The author is greatly indebted to Prof. Joseph Maina
Mungai for guidance, support, and assistance during this study. The author is also grateful to the Department of Wildlife Conservation and Management in the Min- istry of Tourism and Wildlife of the Government of Kenya for granting permission to cull the animals used in this study. My thanks also go to Messrs. L.K. Wai- naina, Moses, N. Kibiru, Christopher N. Kamwaro, and Daniel Ngugi for their technical help, and lastly to Mrs. Lydia Mwombe, Mrs. Mary Kiarie, Miss Lydia Kinyua, and Mrs. Erna Kelley for typing the manuscript.
LITERATURE CITED
Appleton, A.B., and G.M.H. Waites (1957) A surgical approach to the superior cervical ganglion and related structures in the sheep. J . Physiol., (Lond.), 135.52-57.
Baldwin, B.A. (1964) The anatomy of the arterial supply to the cranial regions of the sheep and ox. Am. J. Anat., 115t101-118.
Baldwin, B.A., and F.R. Bell (1963a) Blood-flow in the carotid and vertebral arteries of the sheep and calf. J. Physiol., (Lond.), 167t448- 462.
Baldwin, B.A., and F.R. Bell (1963b) The effect on blood pressure in the sheep and calf of clamping some of the arteries contributing to the cephalic circulation. J. Physiol., (Lond.), 167t463-479.
Baldwin, B.A., and F.R. Bell (1963~) The anatomy of the cerebral circulation of the sheep and ox. The dynamic distribution of the blood supplied by the carotid and vertebral arteries to the cranial regions. J . Anat., 97t203-215.
Bloom, W., and D.W. Fawcett (1968) A Textbook of Histology. W.B. Saunders, Philadelphia, London, Toronto, p. 370.
Daniel, P.M., J.D.K. Dawes, and M.M.L. Prichard (1953) Studies on the carotid rete and its associated arteries. Philos. Tram R. Soc. Lond., [Biol.], 2372% 173-208.
Du Boulay, G.H., and P.M. Verity (1973) The Cranial Arteries of Mam- mals. Williams Heinemann William) Medical Books, London and Tonbridge.
Edelman, N.H., P. Epstein, N.S. Cherniack, and A.P. Fishman (1972)
Control of cerebral blood flow in the goat; a role of the carotid rete. Am. J. Physiol., 223:615-619.
Feigl, W., H. Sinzinger, L. Howanietz, and C.H. Leithner (1976) A morphologically different type of smooth muscle cell in the inner media of splenic artery. Acta Anat., 94t617-625.
Franklin, K.J., and F. Haynes (1927) The histology of the giraffe’s carotid, functionally considered. J . Anat., 62t115-117.
Goetz, R.H. (1962) A study in the adaptations of the circulation to gravitational stresses. Proc. Rudolf Virchow Med. Soc. New York, 21t110-127.
Goetz, R.H., and E.N. Keen (1957) Some aspects of the cardiovascular system in the giraffe. Angiology, 8542-564.
Goetz, R.H., J.V. Warren, O.H. Gauer, J.L. Patterson Jr., J.T. Doyle, E.N. Keen, and M. McGregor (1960) Circulation of the giraffe. Circ. Res. 8t1049-1058.
Jeager, M. (1962) Study of the elasticity and tension of the carotid artery of the cow in comparison with the aorta and coronary ves- sels. Helv. Physiol. Pharmacol. Acta, 20:7-24.
Katzberg, A.A. (1966) The attenuation of elastic tissue in the aorta of baboon, Papio akguera. Anat. Rec., 154t213-220.
Keatinge, W.R., and C. Torrie (1976) Action of Sympathetic nerves on inner and outer muscle of sheep carotid artery and effect of pres- sure on nerve distribution. J . Physiol., (Lond.), 257t699-712.
Kimani, J.K. (1979) Some Aspects of the Structural Organization of the Carotid Arterial System of the Giraffe (Giraffu carnelopardalis) With Special Reference to the Carotid Sinus Baroreceptor Equiva- lent. Ph.D. Thesis, University of Nairobi, 417 pp.
Kimani, J.K. (1981) Structural evidence for insertion of collagen fibres to smooth muscle cells in the carotid arterial system of the giraffe (Giraffa carnelopardalzs). Cell Tissue Res., 214t219-224.
Rimani, J.K. (1983) The structural organization of the carotid arterial system of the giraffe (Giraffa carnelopardalzs). Afr. J. Ecol., 21t317- 324.
Kimani, J.K., and J.M. Mungai (1983) Observations on the structure and innervation of the presumptive carotid sinus area in the giraffe (Gzraffa carnelopardalis). Acta Anat., 115117-133.
Lawrence, W.E., and R.E. Rewell (1948) The cerebral blood supply in the giraffidae. Proc. Zool. Soc. Lond., 118t202-212.
Mekata, F. (1984) Different electrical responses of outer and inner muscle of rabbit carotid artery to noradrenaline and nerves. J. Physiol. (Lond.), 346589-598.
Rees, P.M. (1968) Electron microscopical observations on the architec- ture of the carotid arterial walls, with reference to the sinus por- tion. J. Anat., 103t35-47.
Sisson, S., and J.D. Grossman (1975) The Anatomy of the Domestic Animals. R. Getty, ed. The Macmillan Company of India, Delhi, Bombay, Calcutta, Madras, p. 964.
Somlyo, A.P., and A.V. Somlyo (1968) Vascular smooth muscle. 1. Normal structure, pathology, biochemistry and biophysics. Phar- macol. Rev., 2Ot197-272.
Van Citters, R.L., W.S. Kemper, and D.L. Franklin (1968) Blood flow and pressure in the giraffe carotid artery. Comp. Biochem. Physiol., 24t1035-1042.
Van Citters, R.L., D.L. Franklin, D.F. Vatner, T. Patrick, and J.V. Warren (1969) Cerebral haemodynamics in the giraffe. Trans. As- SOC. Am. Phys., 82t293-303.
Waites, G.M.H. (1960) The influence of the occipito-vertebral anasto- moses on the carotid sinus reflex of the sheep. J . Exp. Physiol., 45t243-251.
Related Documents











