Structural features and anticoagulant activities of a novel natural low molecular weight heparin from the shrimp Penaeus brasiliensis Carl P. Dietrich a ; *, Jose ¤ F. Paiva b , Ricardo A.B. Castro a , Suely F. Chavante b , Walter Jeske c , Jawed Fareed c , Philip A.J. Gorin d , Aline Mendes a , Helena B. Nader a a Departamento de Bioqu| ¤mica, Escola Paulista de Medicine, Universidade Federal de Sa ‹o Paulo, Rua 3 de Maio 100, 4‡ andar, CEP 04044-020, Sa ‹o Paulo, S.P., Brazil b Departamento de Bioqu| ¤mica, Universidade Federal do Rio Grande do Norte, Natal, R.N., Brazil c Department of Pathology, Loyola Medical School, Maywood, IL, USA d Departamento de Bioqu| ¤mica, Universidade Federal do Parana, Parana, Pr., Brazil Received 9 March 1999; accepted 17 May 1999 Abstract A natural low molecular weight heparin (8.5 kDa), with an anticoagulant activity of 95 IU/mg by the USP assay, was isolated from the shrimp Penaeus brasiliensis. The crustacean heparin was susceptible to both heparinase and heparitinase II from Flavobacterium heparinum forming tri- and di-sulfated disaccharides as the mammalian heparins. 13 C and 1 H NMR spectroscopy revealed that the shrimp heparin was enriched in both glucuronic and non-sulfated iduronic acid residues. The in vitro anticlotting activities in different steps of the coagulation cascade have shown that its anticoagulant action is mainly exerted through the inhibition of factor Xa and heparin cofactor II-mediated inhibition of thrombin. The shrimp heparin has also a potent in vivo antithrombotic activity comparable to the mammalian low molecular weight heparins. ß 1999 Elsevier Science B.V. All rights reserved. Keywords : Shrimp heparin structure; Shrimp heparin NMR; Invertebrate heparin; Natural low molecular weight shrimp heparin; Shrimp heparin, anticlotting activity ; Shrimp heparin, antithrombotic activity ; Crustacean heparin 1. Introduction Heparin, a sulfated glycosaminoglycan present in several mammalian and other vertebrate tissues [1^3], has been widely used in medicine for more than 40 years on account of its anticoagulant, antithrombotic and antilipemic activities [4]. Previous studies have shown that heparin-like compounds are present in some invertebrates [5,6]. A substance denoted mactin, with anticoagulant ac- tivity and structural similarities to mammalian hep- arins, was isolated from the mollusks Cyprinia island- ica and Mactrus pussula [5,7]. Another compound 0304-4165 / 99 / $ ^ see front matter ß 1999 Elsevier Science B.V. All rights reserved. PII:S0304-4165(99)00087-2 Abbreviations : vUA,2S-GlcNS,6S, O-(4-deoxy-hex-4-enopyra- nosyluronic acid 2-sulfate)-(1C4)-2-sulfamino-D-glucose 6-sul- fate; vUA,2S-GlcNS, O-(4-deoxy-hex-4-enopyranosyluronic acid 2-sulfate)-(1C4)-2-sulfamino-D-glucose ; vUA-GlcNS,6S, O-(4- deoxy-hex-4-enopyranosyluronic acid)-(1C4)-2-sulfamino-D-glu- cose 6-sulfate ; vUA-GlcNS, O-(4-deoxy-hex-4-enopyranosyl- uronic acid)-(1C4)-2-sulfamino-D-glucose ; vUA-GlcNAc,6S, O- (4-deoxy-hex-4-enopyranosyluronic acid)-(1C4)-2-acetamido-D- glucose 6-sulfate; Hep, heparin; HS, heparan sulfate; CS, chon- droitin sulfate; DS, dermatan sulfate; NMR, nuclear magnetic resonance * Corresponding author. Fax: +55-11-573-6407; E-mail : [email protected] Biochimica et Biophysica Acta 1428 (1999) 273^283 www.elsevier.com/locate/bba

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Structural features and anticoagulant activities of a novel natural lowmolecular weight heparin from the shrimp Penaeus brasiliensis
Carl P. Dietrich a;*, Jose F. Paiva b, Ricardo A.B. Castro a, Suely F. Chavante b,Walter Jeske c, Jawed Fareed c, Philip A.J. Gorin d, Aline Mendes a, Helena B. Nader a
a Departamento de Bioqu|mica, Escola Paulista de Medicine, Universidade Federal de Sa¬o Paulo, Rua 3 de Maio 100, 4³ andar,CEP 04044-020, Sa¬o Paulo, S.P., Brazil
b Departamento de Bioqu|mica, Universidade Federal do Rio Grande do Norte, Natal, R.N., Brazilc Department of Pathology, Loyola Medical School, Maywood, IL, USA
d Departamento de Bioqu|mica, Universidade Federal do Parana, Parana, Pr., Brazil
Received 9 March 1999; accepted 17 May 1999
Abstract
A natural low molecular weight heparin (8.5 kDa), with an anticoagulant activity of 95 IU/mg by the USP assay, wasisolated from the shrimp Penaeus brasiliensis. The crustacean heparin was susceptible to both heparinase and heparitinase IIfrom Flavobacterium heparinum forming tri- and di-sulfated disaccharides as the mammalian heparins. 13C and 1H NMRspectroscopy revealed that the shrimp heparin was enriched in both glucuronic and non-sulfated iduronic acid residues. Thein vitro anticlotting activities in different steps of the coagulation cascade have shown that its anticoagulant action is mainlyexerted through the inhibition of factor Xa and heparin cofactor II-mediated inhibition of thrombin. The shrimp heparin hasalso a potent in vivo antithrombotic activity comparable to the mammalian low molecular weight heparins. ß 1999 ElsevierScience B.V. All rights reserved.
Keywords: Shrimp heparin structure; Shrimp heparin NMR; Invertebrate heparin; Natural low molecular weight shrimp heparin;Shrimp heparin, anticlotting activity; Shrimp heparin, antithrombotic activity; Crustacean heparin
1. Introduction
Heparin, a sulfated glycosaminoglycan present inseveral mammalian and other vertebrate tissues [1^3],has been widely used in medicine for more than 40years on account of its anticoagulant, antithromboticand antilipemic activities [4].
Previous studies have shown that heparin-likecompounds are present in some invertebrates [5,6].A substance denoted mactin, with anticoagulant ac-tivity and structural similarities to mammalian hep-arins, was isolated from the mollusks Cyprinia island-ica and Mactrus pussula [5,7]. Another compound
0304-4165 / 99 / $ ^ see front matter ß 1999 Elsevier Science B.V. All rights reserved.PII: S 0 3 0 4 - 4 1 6 5 ( 9 9 ) 0 0 0 8 7 - 2
Abbreviations: vUA,2S-GlcNS,6S, O-(4-deoxy-hex-4-enopyra-nosyluronic acid 2-sulfate)-(1C4)-2-sulfamino-D-glucose 6-sul-fate; vUA,2S-GlcNS, O-(4-deoxy-hex-4-enopyranosyluronic acid2-sulfate)-(1C4)-2-sulfamino-D-glucose; vUA-GlcNS,6S, O-(4-deoxy-hex-4-enopyranosyluronic acid)-(1C4)-2-sulfamino-D-glu-cose 6-sulfate; vUA-GlcNS, O-(4-deoxy-hex-4-enopyranosyl-uronic acid)-(1C4)-2-sulfamino-D-glucose; vUA-GlcNAc,6S, O-(4-deoxy-hex-4-enopyranosyluronic acid)-(1C4)-2-acetamido-D-glucose 6-sulfate; Hep, heparin; HS, heparan sulfate; CS, chon-droitin sulfate; DS, dermatan sulfate; NMR, nuclear magneticresonance
* Corresponding author. Fax: +55-11-573-6407;E-mail : [email protected]
BBAGEN 24841 26-7-99
Biochimica et Biophysica Acta 1428 (1999) 273^283
www.elsevier.com/locate/bba

from the clam Mercenaria mercenaria [8] also exhib-ited several structural similarities to heparin.
A heparin-like compound with high anticoagulantactivity has been isolated as well from the mollusksAnomalocardia brasiliana, Donnax striatus and Tivelamactroides. Enzymatic and nitrous acid degrada-tion as well as 13C and 1H nuclear magnetic reso-nance spectroscopy have shown that the molluskheparins were similar to those of mammalian origin[9^11].
We now report the presence of a heparin in thecrustacean Penaeus brasiliensis. Of particular signi¢-cance were the ¢ndings that this heparin has a lowmolecular weight, is enriched with non-sulfated ur-onic acid residues and exhibits a potent antithrom-botic activity.
2. Materials and methods
2.1. Materials
Chondroitin 4-sulfate and dermatan sulfate werepurchased from Miles Laboratories (Elkhart, IN,USA). Heparan sulfate from bovine pancreas, hepa-rin from bovine mucosa, low molecular weight(LMW) and ultra low molecular weight (ULMW)heparins were gifts from Dr. P. Bianchini (OpocrinResearch Laboratories, Modena, Italy). Heparinfrom bovine lung was a gift from the late L.L. Cole-man (Upjohn, Kalamazoo, MI, USA). HeparitinasesI and II, and heparinase (heparinase I, EC 4.2.2.7)were prepared as previously described [11]. Propyle-nediamine (1,3-diaminopropane) was purchased fromAldrich Co. (Milwaukee, WI, USA). Agarose lowMR was purchased from Bio-Rad (Richmond, CA,USA).
2.2. Extraction and puri¢cation of shrimp heparin
Immediately after collection, 20 kg of shrimpheads (which contain the viscera) were added to 1 lof 0.8 M of NaCl in a 40 l capacity pan and main-tained for 30 min at 100³C in order to inactivate theglycosidases present in the tissues. The mixture wasthen ground in a Waring blender. The pH of themixture was adjusted to 8.0 with NaOH and 2 g ofSuperase (P¢zer Laboratory, New York, NY, USA)
was added. After 24 h incubation with agitation andperiodic adjustments of pH at 60³C the mixture was¢ltered through cheesecloth. To the ¢ltrate 200 g ofAmberlyte IRA 900, ion-exchange resin (Rohm andHaas, Sa¬o Paulo, Brazil) was added and the resultingmixture was agitated for 24 h at room temperatureunder a layer of toluene. The suspension was then¢ltered again through the same cheesecloth. The res-in retained in the cloth was washed with 2 l of waterat 50³C and subsequently washed with 2 l of 0.8 MNaCl at room temperature. The washed resin wassuspended in 100 ml of 3 M NaCl and agitated for4 h and ¢ltered. This operation was repeated threetimes. The ¢ltrates were maintained for 48 h at 5³Cafter addition of 2 volumes of methanol. The precip-itate formed was collected by centrifugation at5000Ug for 30 min, washed with 80% methanoland dried. The glycosaminoglycan mixture was fur-ther puri¢ed by acetone fractionation as follows: 1 gof the crude material was dissolved in 20 ml of0.15 M of NaCl and 0.4 volumes of acetone wereadded under agitation. The resulting solution wasmaintained at 5³C for 24 h. The precipitate formedwas collected by centrifugation and dried. The oper-ation was repeated successively with 0.5, 0.6, 0.7 and1.0 volume of acetone calculated from the initial so-lution. The resulting precipitates were dried and ana-lyzed. The fractions precipitated between 0.5 and0.6 volumes of acetone contained the heparin. Theyield of these fractions was 650 mg.
2.3. Enzymatic degradation of shrimp heparin
The products formed from the shrimp heparin byaction of heparinase and heparitinases were preparedand identi¢ed as follows: 100 Wg of heparin or hep-arin fragments (see below) was incubated with 0.1 Uof enzymes, in 0.05 M ethylenediamine-acetate bu¡-er, pH 7.0 for 8 h at 30³C in a ¢nal volume of 20 Wl.The incubation mixtures were then spotted in What-man No. 1 paper and subjected to chromatographyin isobutyric acid: 1 M NH3, 5/3, v/v or isobutyricacid:1.25 M NH3, 5/3.6, v/v for 48 h. The unsatu-rated products formed were detected by short waveUV lamp and silver nitrate staining. The incubationmixtures were also analyzed by HPLC in a 0.95U25cm SAX ion exchange resin (Dupont) and the di-saccharides formed were eluted from the column us-
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283274

ing an NaCl gradient (0.01^2 M) with a £ow of 1 ml/min and monitored at 230 nm. For large scale prep-aration of the fragments, 5 mg of heparin was incu-bated with 5 U of heparinase in 0.05 M ethylenedi-amine-acetate bu¡er, pH 7.0 for 8 h at 30³C in a ¢nalvolume of 500 Wl. The mixture was then fractionatedby molecular sieving in a GS-320 HPLC column(1U20 cm, exclusive MW 40 kDa) coupled to aGS-220 HPLC column (1U20 cm, exclusive MW3 kDa) (Asahipak GS series, Asahi Chemical Indus-try Co., Yakoo, Japan) previously equilibrated in0.8 M CaCl2. The products formed were elutedfrom the column with a £ow of 1 ml/min using0.8 M CaCl2. The unsaturated products were moni-tored at 230 nm whereas the undegraded heparin wasmonitored at 200 nm (carboxyl groups). The frac-tions containing each peak were combined, driedand suspended in 10 ml of acetone to remove theCaCl2. The precipitates were washed twice with ace-tone and dried. Markers of heparin (15 kDa) LMWheparin (6.0 kDa), ULMW heparin (3.5 kDa) hexa-,tetra- and di-saccharides (prepared from heparinwith bacterial heparinase) were used for calibrationof the HPLC columns.
2.4. NMR spectroscopy
Samples (100 mg) were dissolved in D2O andthe 13C NMR spectra were obtained in BrukerAM-400WB and AC300P spectrometers at 35³C.1H NMR spectra of the samples (15^30 mg) werealso obtained in a Bruker AM-400WB spectrometer.
2.5. Pharmacological activities
In vitro anticoagulant activity was determined ac-cording to the United States Pharmacopoeia assay.Thrombin and FXa generation inhibition assayswere performed according to a method previouslydescribed [12]. These assays were performed on afast kinetic centrifugal analyzer (ACL-300, Lexing-ton, MA, USA). The heparins studied were dissolvedin physiological saline to the appropriate concentra-tions and placed in the sample cup carrousel. Re-agent position 1 contains 60 Wl of the heparin solu-tion and 60 Wl of ¢brinogen de¢cient plasma (GeorgeKing Biomedical, Overland Park, KS, USA) diluted1:8 in 100 mM Tris-HCl bu¡er (pH 8.5). Reagent
position 2 contains 60 Wl of heparin solution andactin (Dade, Miami, FL, USA) diluted 1:1 withSpectrozyme TH or Spectrozyme FXa (American Di-agnostica, Greenwich, CT, USA) for measuring in-trinsic IIa or Xa generation. Thromboplastin C(Dade, Miami, FL, USA) diluted 1:6 with Spectro-zyme TH or Spectrozyme FXa was used for measur-ing the inhibition of thrombin and FXa extrinsicgeneration. Optical density was recorded at 405 nm.The results are expressed as % inhibition of an un-supplemented saline control.
Potentiation of AT-III and HC-II activity wasmeasured using chromogenic substrate assays [13].In these assays, heparin was diluted in saline andincubated with puri¢ed HC-II (Diagnostica Stago,Gennevierres, France) or AT-III (Kabi, Stock-holm, Sweden) and either thrombin or factor Xa.After 1 min incubation at 37³C, residual enzymaticactivity was measured with a chromogenic sub-strate.
Antithrombotic activity was determined using therat laser induced thrombosis model as described indetail by Weichert et al. [14]. Brie£y, the rat is anes-thetized with sodium pentobarbital (nembutal, 60 mg/kg, intraperitoneally). An intestinal loop is exposedthrough a hypogastric incision and continuously irri-gated with saline. Vascular lesions are induced withan argon laser mounted on a microscope with inter-ference contrast optics. The e¡ective constant energyis 0.12 W and the exposure time of the laser lesions is1/15 s. The time interval between laser shots is 1 min.Results are evaluated by direct observation of thevessel through the microscope. The number of laserinjuries required to induce a thrombus which is atleast as long and as broad as the vessel diameter isthe endpoint.
2.6. Other methods
Hexosamine, uronic acid, sulfate and acetyl resi-dues were determined as previously described [15].Molecular weight was estimated by polyacrylamidegel electrophoresis calibrated with heparin (15 kDa)and LMW heparin standards [15]. Agarose gel elec-trophoresis of the sulfated glycosaminoglycans in thediscontinuous bu¡er barium acetate/1,3-diaminopro-pane acetate was performed as previously described[16].
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283 275
3. Results
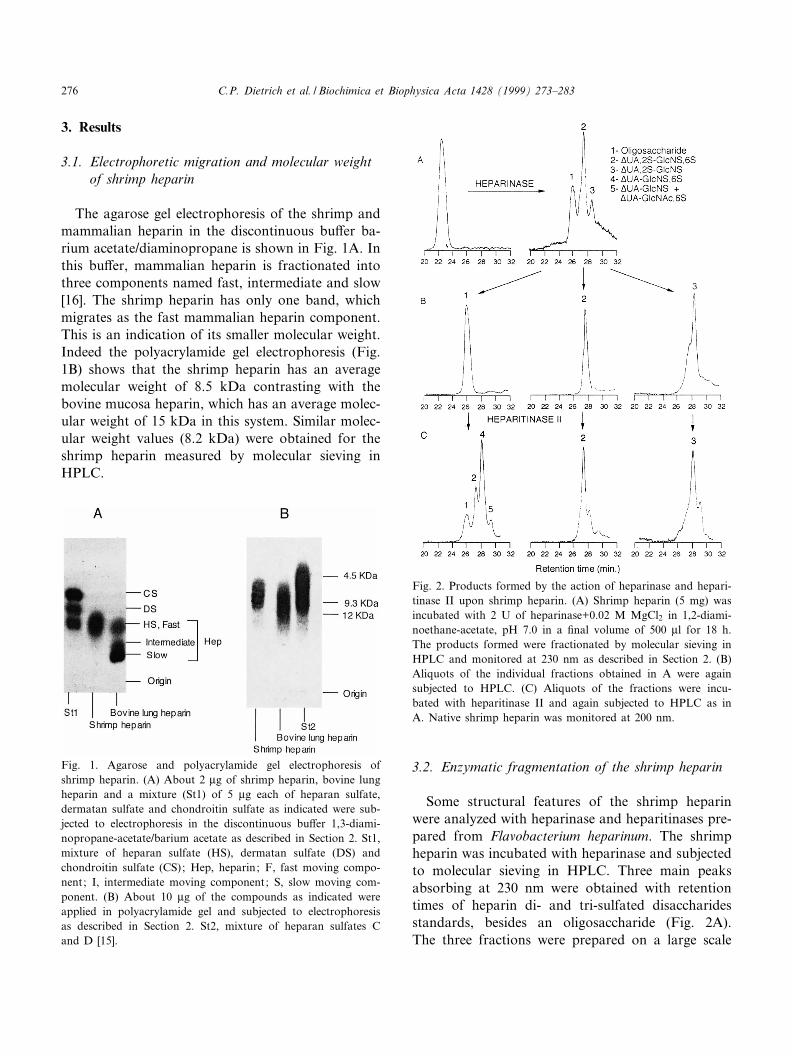
3.1. Electrophoretic migration and molecular weightof shrimp heparin
The agarose gel electrophoresis of the shrimp andmammalian heparin in the discontinuous bu¡er ba-rium acetate/diaminopropane is shown in Fig. 1A. Inthis bu¡er, mammalian heparin is fractionated intothree components named fast, intermediate and slow[16]. The shrimp heparin has only one band, whichmigrates as the fast mammalian heparin component.This is an indication of its smaller molecular weight.Indeed the polyacrylamide gel electrophoresis (Fig.1B) shows that the shrimp heparin has an averagemolecular weight of 8.5 kDa contrasting with thebovine mucosa heparin, which has an average molec-ular weight of 15 kDa in this system. Similar molec-ular weight values (8.2 kDa) were obtained for theshrimp heparin measured by molecular sieving inHPLC.
3.2. Enzymatic fragmentation of the shrimp heparin
Some structural features of the shrimp heparinwere analyzed with heparinase and heparitinases pre-pared from Flavobacterium heparinum. The shrimpheparin was incubated with heparinase and subjectedto molecular sieving in HPLC. Three main peaksabsorbing at 230 nm were obtained with retentiontimes of heparin di- and tri-sulfated disaccharidesstandards, besides an oligosaccharide (Fig. 2A).The three fractions were prepared on a large scale
Fig. 1. Agarose and polyacrylamide gel electrophoresis ofshrimp heparin. (A) About 2 Wg of shrimp heparin, bovine lungheparin and a mixture (St1) of 5 Wg each of heparan sulfate,dermatan sulfate and chondroitin sulfate as indicated were sub-jected to electrophoresis in the discontinuous bu¡er 1,3-diami-nopropane-acetate/barium acetate as described in Section 2. St1,mixture of heparan sulfate (HS), dermatan sulfate (DS) andchondroitin sulfate (CS); Hep, heparin; F, fast moving compo-nent; I, intermediate moving component; S, slow moving com-ponent. (B) About 10 Wg of the compounds as indicated wereapplied in polyacrylamide gel and subjected to electrophoresisas described in Section 2. St2, mixture of heparan sulfates Cand D [15].
Fig. 2. Products formed by the action of heparinase and hepari-tinase II upon shrimp heparin. (A) Shrimp heparin (5 mg) wasincubated with 2 U of heparinase+0.02 M MgCl2 in 1,2-diami-noethane-acetate, pH 7.0 in a ¢nal volume of 500 Wl for 18 h.The products formed were fractionated by molecular sieving inHPLC and monitored at 230 nm as described in Section 2. (B)Aliquots of the individual fractions obtained in A were againsubjected to HPLC. (C) Aliquots of the fractions were incu-bated with heparitinase II and again subjected to HPLC as inA. Native shrimp heparin was monitored at 200 nm.
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283276
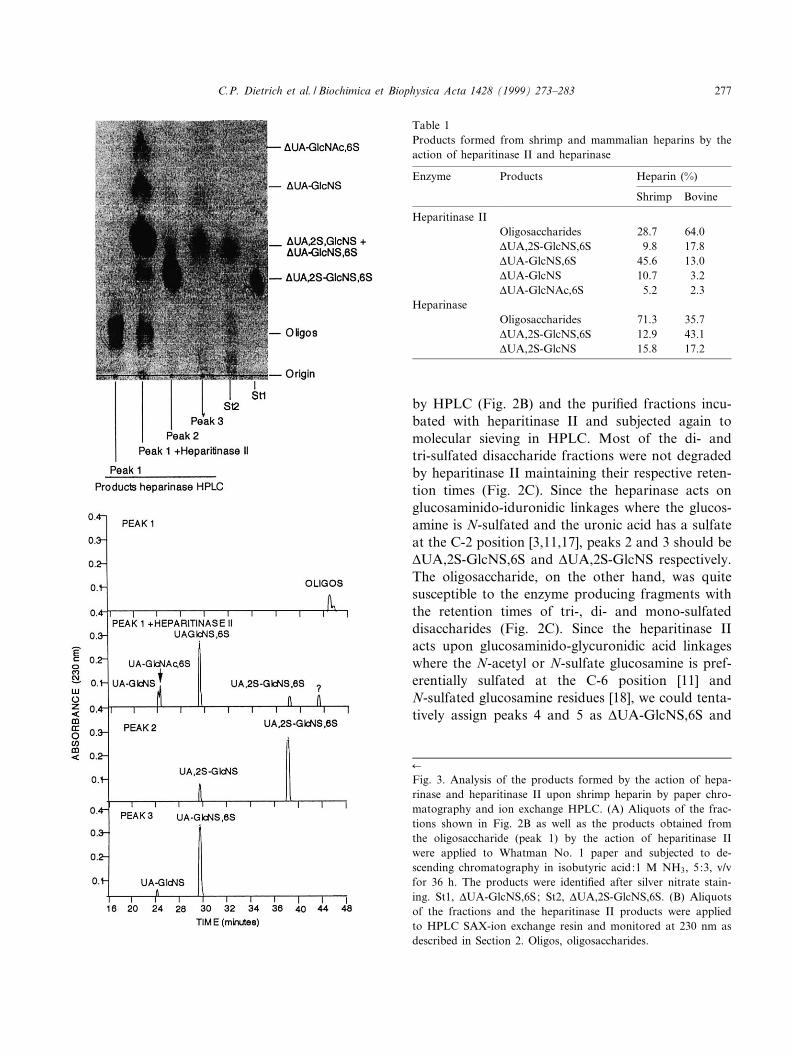
by HPLC (Fig. 2B) and the puri¢ed fractions incu-bated with heparitinase II and subjected again tomolecular sieving in HPLC. Most of the di- andtri-sulfated disaccharide fractions were not degradedby heparitinase II maintaining their respective reten-tion times (Fig. 2C). Since the heparinase acts onglucosaminido-iduronidic linkages where the glucos-amine is N-sulfated and the uronic acid has a sulfateat the C-2 position [3,11,17], peaks 2 and 3 should bevUA,2S-GlcNS,6S and vUA,2S-GlcNS respectively.The oligosaccharide, on the other hand, was quitesusceptible to the enzyme producing fragments withthe retention times of tri-, di- and mono-sulfateddisaccharides (Fig. 2C). Since the heparitinase IIacts upon glucosaminido-glycuronidic acid linkageswhere the N-acetyl or N-sulfate glucosamine is pref-erentially sulfated at the C-6 position [11] andN-sulfated glucosamine residues [18], we could tenta-tively assign peaks 4 and 5 as vUA-GlcNS,6S and
Table 1Products formed from shrimp and mammalian heparins by theaction of heparitinase II and heparinase
Enzyme Products Heparin (%)
Shrimp Bovine
Heparitinase IIOligosaccharides 28.7 64.0vUA,2S-GlcNS,6S 9.8 17.8vUA-GlcNS,6S 45.6 13.0vUA-GlcNS 10.7 3.2vUA-GlcNAc,6S 5.2 2.3
HeparinaseOligosaccharides 71.3 35.7vUA,2S-GlcNS,6S 12.9 43.1vUA,2S-GlcNS 15.8 17.2
6
Fig. 3. Analysis of the products formed by the action of hepa-rinase and heparitinase II upon shrimp heparin by paper chro-matography and ion exchange HPLC. (A) Aliquots of the frac-tions shown in Fig. 2B as well as the products obtained fromthe oligosaccharide (peak 1) by the action of heparitinase IIwere applied to Whatman No. 1 paper and subjected to de-scending chromatography in isobutyric acid:1 M NH3, 5:3, v/vfor 36 h. The products were identi¢ed after silver nitrate stain-ing. St1, vUA-GlcNS,6S; St2, vUA,2S-GlcNS,6S. (B) Aliquotsof the fractions and the heparitinase II products were appliedto HPLC SAX-ion exchange resin and monitored at 230 nm asdescribed in Section 2. Oligos, oligosaccharides.
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283 277
vUA-GlcNAc,6S or vUA-GlcNS. The oligosaccha-ride also forms trisulfated disaccharide by action ofheparitinase II. This disaccharide could either beat the non-reducing end of the oligosaccharide (leftbehind by heparinase) or derived from a sulfatedglucuronic acid disaccharide present in the mole-cule.
The di¡erent fractions and products were also an-alyzed by paper chromatography and ion exchangeHPLC. Peak 2 and peak 3 have the same chromato-graphic migrations and retention times of vUA,2S-GlcNS,6S and vUA,2S-GlcNS, respectively (Fig.3A,B) con¢rming the assignments made above. Thedisaccharide products formed by action of hepariti-nase II upon the oligosaccharide (peak 1) have thesame chromatographic migrations and retentiontimes as vUA,2S-GlcNS,6S, vUA-GlcNS,6S, vUA-GlcNS and vUA-GlcNAc,6S standards.
The shrimp heparin was not degraded by hepariti-nase I (results not shown). Since this enzyme is spe-ci¢c for N-acetyl or N-sulfate glucosaminido-glucur-onic acid linkages [11] one has to conclude that themolecule is devoid of these residues. The ¢nding thatheparitinase II produces vUA-GlcNS as one of theproducts of peak I is suggestive that this disaccharidecontains iduronic acid, rather than glucuronic acid.These results also exclude the possibility that theshrimp heparin could be a special type of heparansulfate since the heparitinase I acts upon all verte-brate and invertebrate heparan sulfates described sofar [6,11,18].
Table 1 show the relative amounts of productsformed from the mammalian and shrimp heparinsby the action of the enzymes. The shrimp heparinis more extensively degraded by heparitinase IIthan by heparinase. Only 13% of vUA,2S-GlcNS,6Sis formed from the shrimp heparin by the action ofheparinase compared to 43% formed from the mam-malian heparin by the action of the enzyme. Thisindicates that the shrimp heparin contains a lowerdegree of sulfation of its uronic acid residues.
3.3. Nuclear magnetic resonance of heparins
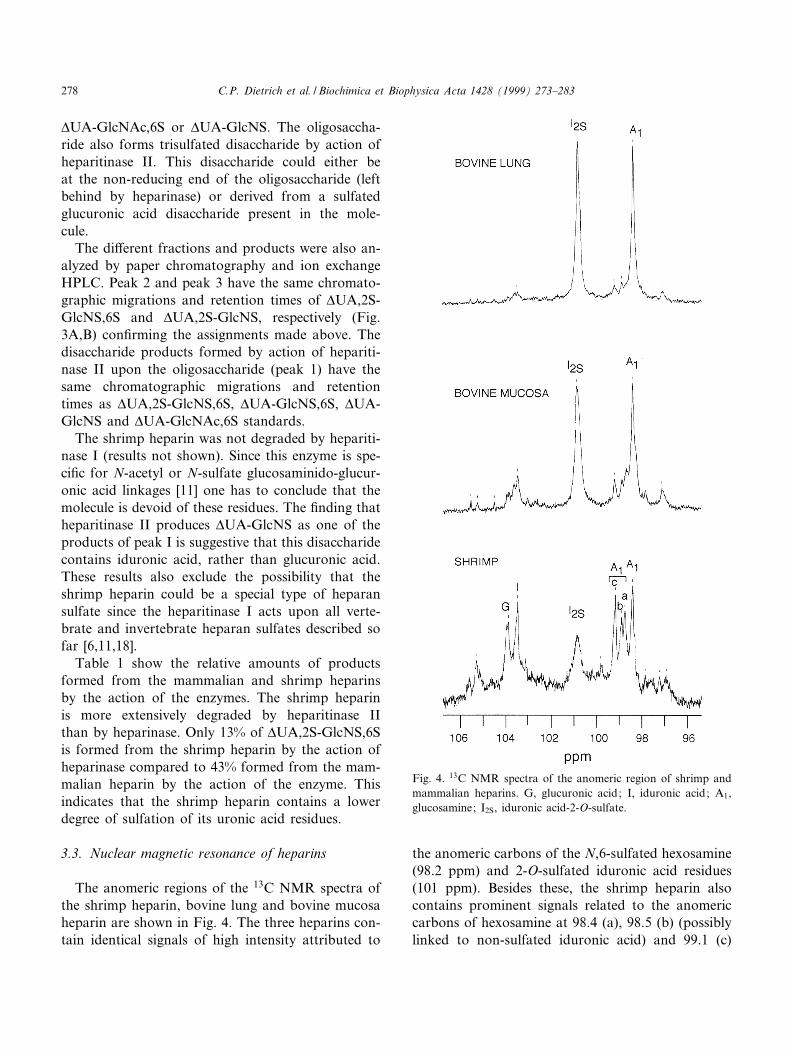
The anomeric regions of the 13C NMR spectra ofthe shrimp heparin, bovine lung and bovine mucosaheparin are shown in Fig. 4. The three heparins con-tain identical signals of high intensity attributed to
the anomeric carbons of the N,6-sulfated hexosamine(98.2 ppm) and 2-O-sulfated iduronic acid residues(101 ppm). Besides these, the shrimp heparin alsocontains prominent signals related to the anomericcarbons of hexosamine at 98.4 (a), 98.5 (b) (possiblylinked to non-sulfated iduronic acid) and 99.1 (c)
Fig. 4. 13C NMR spectra of the anomeric region of shrimp andmammalian heparins. G, glucuronic acid; I, iduronic acid; A1,glucosamine; I2S, iduronic acid-2-O-sulfate.
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283278
ppm (possibly linked to glucuronic acid). The signalsat 103.3 and 103.9 are attributable to non-sulfated K-L-iduronic and L-D-glucuronic acid residues, respec-tively [19^21]. All these signals are also present inbovine mucosa heparin but with lower intensityand are also insinuated in bovine lung heparin butwith still lower intensity when compared to the mu-cosa and shrimp heparins.
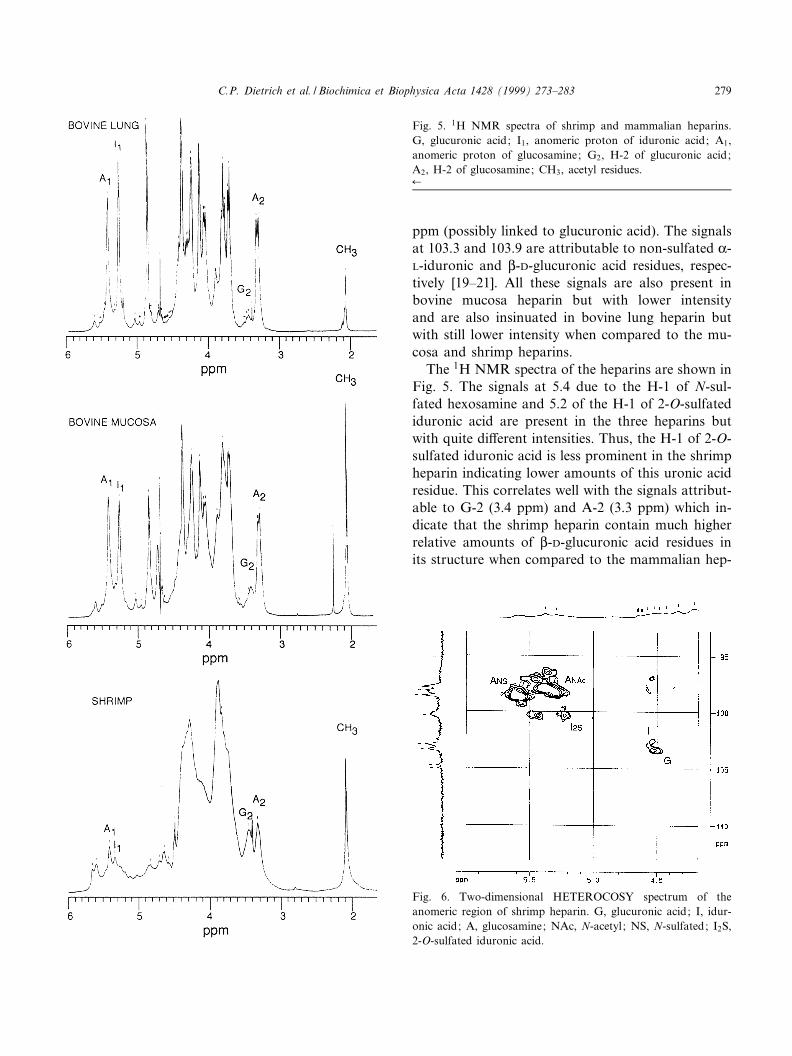
The 1H NMR spectra of the heparins are shown inFig. 5. The signals at 5.4 due to the H-1 of N-sul-fated hexosamine and 5.2 of the H-1 of 2-O-sulfatediduronic acid are present in the three heparins butwith quite di¡erent intensities. Thus, the H-1 of 2-O-sulfated iduronic acid is less prominent in the shrimpheparin indicating lower amounts of this uronic acidresidue. This correlates well with the signals attribut-able to G-2 (3.4 ppm) and A-2 (3.3 ppm) which in-dicate that the shrimp heparin contain much higherrelative amounts of L-D-glucuronic acid residues inits structure when compared to the mammalian hep-
Fig. 6. Two-dimensional HETEROCOSY spectrum of theanomeric region of shrimp heparin. G, glucuronic acid; I, idur-onic acid; A, glucosamine; NAc, N-acetyl ; NS, N-sulfated; I2S,2-O-sulfated iduronic acid.
Fig. 5. 1H NMR spectra of shrimp and mammalian heparins.G, glucuronic acid; I1, anomeric proton of iduronic acid; A1,anomeric proton of glucosamine; G2, H-2 of glucuronic acid;A2, H-2 of glucosamine; CH3, acetyl residues.6
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283 279
arins. The signal at 2.1 due to the acetyl groups ispresent in the three heparins being more prominentin the bovine mucosa and shrimp heparins.
Additional assignments of NMR signals of theshrimp heparin were obtained by heteronuclear 2Dexperiments (HETEROCOSY) (Fig. 6), showing cor-relation of H-1 (4.5 ppm) and C-1 (104 ppm) ofglucuronic acid residues, (H-1, 4.6 ppm; C-1, 102.5ppm) of non-sulfated iduronic acid residues and
those attributed to the residues of the 2-O-sulfatediduronic acid (H-1, 5.24 ppm; C-1, 101.0 ppm). Thesignals of the N-sulfated glucosamine (H-1, 5.6 ppm;C-1, 99 ppm) and the N-acetylated glucosamine(H-1, 5.35 ppm; C-1, 97.5 ppm) are also evident.
The enzymatic and NMR data indicate that theshrimp heparin is richer in non-sulfated iduronicand glucuronic acid residues, when compared tothat of mammalian origin.
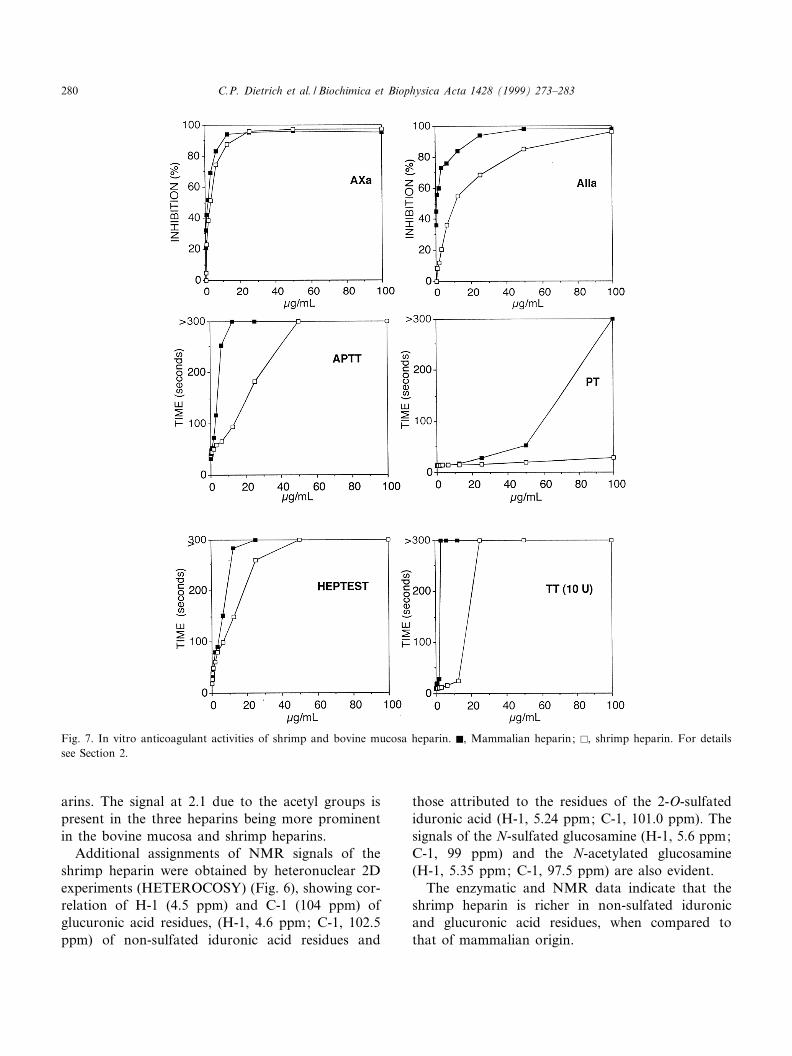
Fig. 7. In vitro anticoagulant activities of shrimp and bovine mucosa heparin. F, Mammalian heparin; E, shrimp heparin. For detailssee Section 2.
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283280
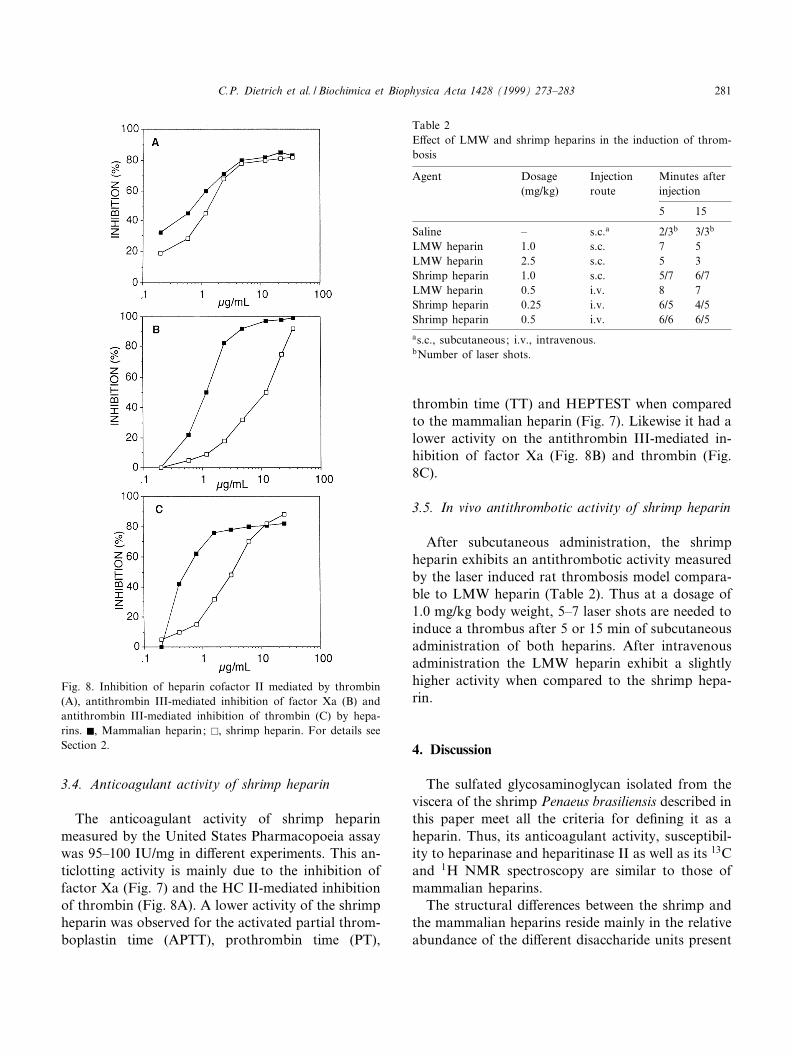
3.4. Anticoagulant activity of shrimp heparin
The anticoagulant activity of shrimp heparinmeasured by the United States Pharmacopoeia assaywas 95^100 IU/mg in di¡erent experiments. This an-ticlotting activity is mainly due to the inhibition offactor Xa (Fig. 7) and the HC II-mediated inhibitionof thrombin (Fig. 8A). A lower activity of the shrimpheparin was observed for the activated partial throm-boplastin time (APTT), prothrombin time (PT),
thrombin time (TT) and HEPTEST when comparedto the mammalian heparin (Fig. 7). Likewise it had alower activity on the antithrombin III-mediated in-hibition of factor Xa (Fig. 8B) and thrombin (Fig.8C).
3.5. In vivo antithrombotic activity of shrimp heparin
After subcutaneous administration, the shrimpheparin exhibits an antithrombotic activity measuredby the laser induced rat thrombosis model compara-ble to LMW heparin (Table 2). Thus at a dosage of1.0 mg/kg body weight, 5^7 laser shots are needed toinduce a thrombus after 5 or 15 min of subcutaneousadministration of both heparins. After intravenousadministration the LMW heparin exhibit a slightlyhigher activity when compared to the shrimp hepa-rin.
4. Discussion
The sulfated glycosaminoglycan isolated from theviscera of the shrimp Penaeus brasiliensis described inthis paper meet all the criteria for de¢ning it as aheparin. Thus, its anticoagulant activity, susceptibil-ity to heparinase and heparitinase II as well as its 13Cand 1H NMR spectroscopy are similar to those ofmammalian heparins.
The structural di¡erences between the shrimp andthe mammalian heparins reside mainly in the relativeabundance of the di¡erent disaccharide units present
Fig. 8. Inhibition of heparin cofactor II mediated by thrombin(A), antithrombin III-mediated inhibition of factor Xa (B) andantithrombin III-mediated inhibition of thrombin (C) by hepa-rins. F, Mammalian heparin; E, shrimp heparin. For details seeSection 2.
Table 2E¡ect of LMW and shrimp heparins in the induction of throm-bosis
Agent Dosage(mg/kg)
Injectionroute
Minutes afterinjection
5 15
Saline ^ s.c.a 2/3b 3/3b
LMW heparin 1.0 s.c. 7 5LMW heparin 2.5 s.c. 5 3Shrimp heparin 1.0 s.c. 5/7 6/7LMW heparin 0.5 i.v. 8 7Shrimp heparin 0.25 i.v. 6/5 4/5Shrimp heparin 0.5 i.v. 6/6 6/5as.c., subcutaneous; i.v., intravenous.bNumber of laser shots.
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283 281

in the heparins. This is clearly observed by the 13CNMR spectroscopy as well as by the amount ofproducts formed by the action of heparinase (whichrecognizes 2-O-sulfated iduronic acid residues) andheparitinase II which act upon the regions containingglucuronic acid and non-sulfated iduronic acid resi-dues. These di¡erences have also been observed re-garding the mollusk heparins which, like the shrimpheparin described here, were more susceptible to hep-aritinase II when compared to the mammalian ones[9,11].
The anticoagulant activity of the shrimp heparin ismainly exerted through the inhibition of factor Xaand the HC II-mediated inhibition of thrombinwhereas the heparin from the mollusk Anomalocardiabrasiliana [9,10,22] as well as the mammalian hepa-rins exert their anticoagulant activity mainly throughthe antithrombin III-mediated inhibition of thrombinand factor Xa. Conceivably these di¡erences of ac-tion are related to the structural diversity of the hep-arins discussed above.
The in vivo antithrombotic activity seems to bequite independent of the structure and molecularweight of the di¡erent heparins. Thus, the shrimpheparin has a similar antithrombotic activity as theLMW heparins. It was also observed that the mol-lusk heparin, in spite of its high anticoagulant activ-ity and high a¤nity for antithrombin III, has an invivo antithrombotic activity comparable to unfrac-tionated and LMW heparins [22].
Except for mollusks, where heparin has been ex-tensively described, this is the ¢rst report of the pres-ence of this compound in another class of inverte-brates. As previously discussed [10] the biologicalactivity of heparins in invertebrates remains enig-matic. The classes Crustacea and Mollusca do notpossess any blood coagulation system similar tothat of mammals and other vertebrates and thusthe presence of compounds that act speci¢callyupon the proteins of the blood coagulation systemis indeed remarkable.
It has been previously speculated that mast cellsand their heparin may serve as modulators of theimmune reactions or other body defence mechanismsand thus the action of heparin upon the coagulationcascade, due to its strong anionic character, may be apharmacological rather than a biological activity[23].
Acknowledgements
Aided in part by grants from FAPESP (Fundac°a¬ode Amparo a Pesquisa do Estado de Sa¬o Paulo),FINEP (Financiadora de Estudos e Projetos) andCNPq (Conselho Nacional de DesenvolvimentoCienti¢co e Tecnologico), Brazil.
References
[1] H.B. Nader, H.K. Takahashi, A.H. Straus, C.P. Dietrich,Biochim. Biophys. Acta 627 (1980) 40^48.
[2] P.B. Gomes, C.P. Dietrich, Comp. Biochem. Physiol. 73B(1982) 857^864.
[3] P. Hovingh, M. Piepkorn, A. Linker, Biochem. J. 237 (1986)573^578.
[4] L.B. Jaques, Pharmacol. Rev. 31 (1980) 99^166.[5] J.A. Cifonelli, M.B. Mathews, Connect. Tissue. Res. 1 (1972)
121^130.[6] C.M.F. Cassaro, C.P. Dietrich, J. Biol. Chem. 252 (1977)
2254^2261.[7] S.L. Burson Jr., M.J. Fahrenbach, L.H. Frommhagen, B.A.
Riccardi, R.A. Brown, J.A. Brockman, H.V. Lowry, E.L.R.Stokstad, J. Am. Chem. Soc. 78 (1956) 5874^5878.
[8] R.E. Jordan, J.A. Marcun, Arch. Biochem. Biophys. 248(1986) 690^696.
[9] C.P. Dietrich, J.F. Paiva, C.T. Moraes, H.K. Takahashi,M.A. Porcionatto, H.B. Nader, Biochim. Biophys. Acta843 (1985) 1^7.
[10] G. Pejler, A. Danielson, I. Bjork, U. Lindahl, H.B. Nader,C.P. Dietrich, J. Biol. Chem. 262 (1987) 1413^1421.
[11] C.P. Dietrich, H.B. Nader, J.F. Paiva, I.L.S Tersariol, E.A.Santos, K.R. Holme, A.S. Perlin, Int. J. Biol. Macromol. 11(1989) 361^366.
[12] B. Kaiser, J. Fareed, D. Hoppensteadt, B. Birdsong, J.M.Walenga, F. Markwardt, Thromb. Res. 65 (1992) 157^164.
[13] W. Jeske, J. Fareed, Semin. Thromb. Hemost. 19 (1993)241^246.
[14] W. Weichert, H.K. Breddin, J. Staubesand, Semin. Thromb.Hemost. 14 (1988) 106^114.
[15] C.P. Dietrich, H.B. Nader, Biochim. Biophys. Acta 345(1974) 34^44.
[16] P. Bianchini, H.B. Nader, H.K. Takahashi, B. Osima, A.H.Straus, C.P. Dietrich, J. Chromatogr. 196 (1980) 455^462.
[17] P. Hovingh, A. Linker, J. Biol. Chem. 257 (1982) 9840^9844.
[18] C.P. Dietrich, I.L.S. Tersariol, L. Toma, C.T. Moraes, M.A.Porcionatto, F,W. Oliveira, H.B. Nader, Cell. Mol. Bio-chem. 44 (1998) 417^429.
[19] M. Ragazzi, D.R. Ferro, A. Provasoli, P. Pumilia, A. Cassi-nari, G. Torri, M. Guerrini, B. Casu, H.B. Nader, C.P. Die-trich, M. Ragazzi, D.R. Ferro, A. Provasoli, P. Pumilia, A.
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283282

Cassinari, G. Torri, H.B. Nader, C.P. Dietrich, J. Carbo-hydr. Chem. 12 (1993) 523^535.
[20] B. Casu, G. Grazioli, N. Razi, M. Guerrini, A. Naggi, G.Torri, P. Oreste, F. Tursi, G. Zoppetti, U. Lindahl, Carbo-hydr. Res. 263 (1994) 271^284.
[21] B. Casu, in: D. Lane, U. Lindahl (Eds.), Heparin, Chemicaland Biological Properties, Clinical Applications, Edward Ar-nold, London, 1989, pp. 25^49.
[22] J.F. Paiva, E.A. Santos, W. Jeske, J. Fareed, H.B. Nader,C.P. Dietrich, Comp. Biochem. Physiol. 111 (1995) 495^499.
[23] H.B. Nader, C.P. Dietrich, in: D.A. Lane, U. Lindahl(Eds.), Heparin: Chemical and Biological Properties, Clini-cal Applications, Edward Arnold, London, 1989, pp. 81^96.
BBAGEN 24841 26-7-99
C.P. Dietrich et al. / Biochimica et Biophysica Acta 1428 (1999) 273^283 283
Related Documents











