Structural differences between the SH3-HOOK-GuK domains of SAP90/PSD-95 and SAP97 Rama Ramesh Vandanapu a,1 , Aditya Kumar Singh a,1 , Marina Mikhaylova b , Pasham Parameshwar Reddy b , Michael R. Kreutz b, * , Yogendra Sharma a, * a Centre for Cellular and Molecular Biology, Council for Scientific and Industrial Research (CSIR), Hyderabad-500 007, India b PG Neuroplasticity, Leibniz Institute for Neurobiology, Brenneckestr. 6, Magdeburg 39118, Germany article info Article history: Received 20 May 2009 and in revised form 14 July 2009 Available online 24 July 2009 Keywords: MAGUK Post synaptic density Thermal unfolding Equilibrium unfolding Hydrophobic interaction chromatography GdmCl, guanidine hydrochloride DSC, differential scanning calorimetry abstract The SH3-HOOK-GUK domains of the postsynaptic scaffolding proteins SAP90/PSD-95 and SAP97 are established targets of synaptic plasticity processes in the brain. A crucial molecular mechanism involved is the transition of this domain to different conformational states. We purified the SH3-HOOK-GUK domain of both proteins to examine variations in protein conformation and stability. As monitored by cir- cular dichroism and differential scanning calorimetry, SAP97 (T m = 64 °C) is significantly more thermal stable than SAP90/PSD-95 (T m = 52 °C) and follows a bimodal phase transition. GdmCl-induced equilib- rium unfolding of both proteins follows the two-state transitions and thus does not involve the accumu- lation of stable intermediate state(s). Equilibrium unfolding of SAP97 is highly cooperative from a native state to an unfolded state. In contrast, SAP90/PSD-95 follows a non-cooperative transition from native to unfolded states. A highly cooperative unfolding reaction in case of SAP97 indicates that the protein existed initially as a compact, well-folded structure, while the gradual, non-cooperative melting reaction in case of SAP90/PSD-95 indicates that the protein is in comparison more flexible. Ó 2009 Elsevier Inc. All rights reserved. Introduction The membrane-associated guanylate kinases (MAGUKs) 2 , a protein family that has been identified at cell–cell contact sites in different organisms including humans [1], are thought to serve important functions in organizing the postsynaptic molecular meshwork. MAGUKs are modular scaffolds that are involved in the clustering of synaptic membrane receptors and cell adhesion molecules, as well as to downstream signaling components of the synapse [1]. The MAGUKs of brain synapses (i.e. SAP90/PSD-95, SAP97, SAP102, PSD-93/Chapsyn110) are multidomain-proteins composed of three PDZ domains, an SH3 domain, a HOOK region and a guanylate kinase-like domain (GUK domain) (Fig. 1). The crystal structure of the SH3-HOOK-GUK region of SAP90/PSD-95 has been solved previously [2,3]. The GUK domain in MAGUKs lacks key amino acid residues required for binding of nucleoside phos- phate and it is assumed that the guanylate kinase-related regions of these multidomain-proteins have adopted a role in protein–pro- tein interactions rather than having an enzymatic role [1]. Accord- ingly, a number of protein–protein interactions have been mapped to this region including those to GKAPs [4,5] and SPAR [6]. Interest- ingly, the SH3 domain in MAGUK proteins interacts intra- and possibly inter-molecularly with the GUK domain [5,7] resulting in oligomerization. It has therefore been proposed that this interac- tion participates in the dynamic formation of the membrane cyto- skeleton [8]. Thus, the transition from closed (intramolecular binding) to open conformations (no intramolecular binding) may influence the accessibility of binding interfaces for protein interac- tions that are crucial for the scaffolding function of MAGUKs. Previous work has also shown that among others, calmodulin binds to the HOOK region localized between the SH3 and the GUK do- mains of SAP97 and SAP102 in a Ca 2+ -dependent manner. It was argued that this binding might be involved in the transition from a closed to an open conformation abrogating the intramolecular interaction [2] and could therefore be an important mechanism for the dynamic organization of the PSD by synaptic Ca 2+ transients [8–10]. Analysis of the amino acid sequence of SH3-HOOK-GUK re- gion of SAP90/PSD-95 and SAP97 reveals that both are quite similar, except that there is an insertion of 37 (I2 motif) or 33 amino acid residues (I3 motif) in SAP97 (Fig. 1a and b). SH3 do- mains in general are intriguing targets for studying signal trans- duction process, drug design and protein folding dynamics [11,12]. In this paper, we describe an efficient method based on hydrophobic interaction chromatography for the preparation 1046-5928/$ - see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.pep.2009.07.007 * Corresponding authors. E-mail addresses: [email protected] (M.R. Kreutz), yogendra@ccmb. res.in (Y. Sharma). 1 These authors contributed equally. 2 Abbreviations used: PSD, post synaptic density; MAGuK, membrane associated guanylate kinase; CD, circular dichroism. Protein Expression and Purification 68 (2009) 201–207 Contents lists available at ScienceDirect Protein Expression and Purification journal homepage: www.elsevier.com/locate/yprep

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Protein Expression and Purification 68 (2009) 201–207
Contents lists available at ScienceDirect
Protein Expression and Purification
journal homepage: www.elsevier .com/ locate /yprep
Structural differences between the SH3-HOOK-GuK domains of SAP90/PSD-95and SAP97
Rama Ramesh Vandanapu a,1, Aditya Kumar Singh a,1, Marina Mikhaylova b, Pasham Parameshwar Reddy b,Michael R. Kreutz b,*, Yogendra Sharma a,*
a Centre for Cellular and Molecular Biology, Council for Scientific and Industrial Research (CSIR), Hyderabad-500 007, Indiab PG Neuroplasticity, Leibniz Institute for Neurobiology, Brenneckestr. 6, Magdeburg 39118, Germany
a r t i c l e i n f o
Article history:Received 20 May 2009and in revised form 14 July 2009Available online 24 July 2009
Keywords:MAGUKPost synaptic densityThermal unfoldingEquilibrium unfoldingHydrophobic interaction chromatographyGdmCl, guanidine hydrochlorideDSC, differential scanning calorimetry
1046-5928/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.pep.2009.07.007
* Corresponding authors.E-mail addresses: [email protected] (M.
res.in (Y. Sharma).1 These authors contributed equally.2 Abbreviations used: PSD, post synaptic density; M
guanylate kinase; CD, circular dichroism.
a b s t r a c t
The SH3-HOOK-GUK domains of the postsynaptic scaffolding proteins SAP90/PSD-95 and SAP97 areestablished targets of synaptic plasticity processes in the brain. A crucial molecular mechanism involvedis the transition of this domain to different conformational states. We purified the SH3-HOOK-GUKdomain of both proteins to examine variations in protein conformation and stability. As monitored by cir-cular dichroism and differential scanning calorimetry, SAP97 (Tm = 64 �C) is significantly more thermalstable than SAP90/PSD-95 (Tm = 52 �C) and follows a bimodal phase transition. GdmCl-induced equilib-rium unfolding of both proteins follows the two-state transitions and thus does not involve the accumu-lation of stable intermediate state(s). Equilibrium unfolding of SAP97 is highly cooperative from a nativestate to an unfolded state. In contrast, SAP90/PSD-95 follows a non-cooperative transition from native tounfolded states. A highly cooperative unfolding reaction in case of SAP97 indicates that the proteinexisted initially as a compact, well-folded structure, while the gradual, non-cooperative melting reactionin case of SAP90/PSD-95 indicates that the protein is in comparison more flexible.
� 2009 Elsevier Inc. All rights reserved.
Introduction ingly, a number of protein–protein interactions have been mapped
The membrane-associated guanylate kinases (MAGUKs)2, aprotein family that has been identified at cell–cell contact sites indifferent organisms including humans [1], are thought to serveimportant functions in organizing the postsynaptic molecularmeshwork. MAGUKs are modular scaffolds that are involved inthe clustering of synaptic membrane receptors and cell adhesionmolecules, as well as to downstream signaling components of thesynapse [1]. The MAGUKs of brain synapses (i.e. SAP90/PSD-95,SAP97, SAP102, PSD-93/Chapsyn110) are multidomain-proteinscomposed of three PDZ domains, an SH3 domain, a HOOK regionand a guanylate kinase-like domain (GUK domain) (Fig. 1). Thecrystal structure of the SH3-HOOK-GUK region of SAP90/PSD-95has been solved previously [2,3]. The GUK domain in MAGUKs lackskey amino acid residues required for binding of nucleoside phos-phate and it is assumed that the guanylate kinase-related regionsof these multidomain-proteins have adopted a role in protein–pro-tein interactions rather than having an enzymatic role [1]. Accord-
ll rights reserved.
R. Kreutz), yogendra@ccmb.
AGuK, membrane associated
to this region including those to GKAPs [4,5] and SPAR [6]. Interest-ingly, the SH3 domain in MAGUK proteins interacts intra- andpossibly inter-molecularly with the GUK domain [5,7] resulting inoligomerization. It has therefore been proposed that this interac-tion participates in the dynamic formation of the membrane cyto-skeleton [8]. Thus, the transition from closed (intramolecularbinding) to open conformations (no intramolecular binding) mayinfluence the accessibility of binding interfaces for protein interac-tions that are crucial for the scaffolding function of MAGUKs.Previous work has also shown that among others, calmodulin bindsto the HOOK region localized between the SH3 and the GUK do-mains of SAP97 and SAP102 in a Ca2+-dependent manner. It wasargued that this binding might be involved in the transition froma closed to an open conformation abrogating the intramolecularinteraction [2] and could therefore be an important mechanismfor the dynamic organization of the PSD by synaptic Ca2+ transients[8–10].
Analysis of the amino acid sequence of SH3-HOOK-GUK re-gion of SAP90/PSD-95 and SAP97 reveals that both are quitesimilar, except that there is an insertion of 37 (I2 motif) or 33amino acid residues (I3 motif) in SAP97 (Fig. 1a and b). SH3 do-mains in general are intriguing targets for studying signal trans-duction process, drug design and protein folding dynamics[11,12]. In this paper, we describe an efficient method basedon hydrophobic interaction chromatography for the preparation
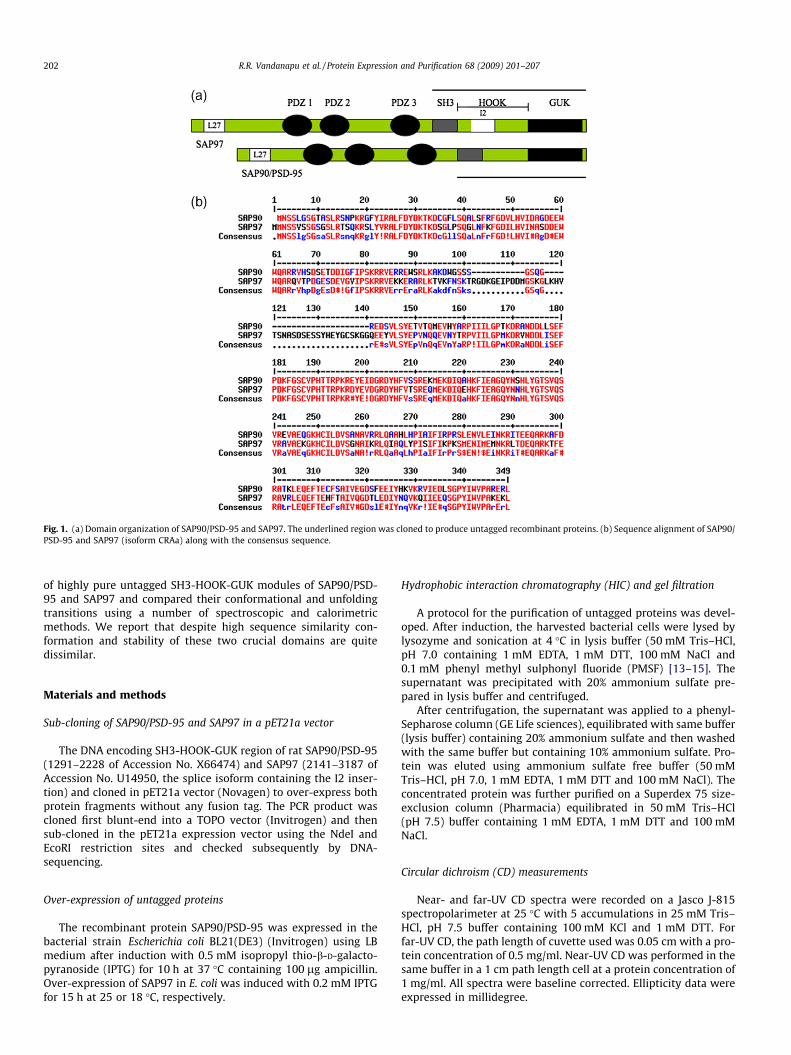
Fig. 1. (a) Domain organization of SAP90/PSD-95 and SAP97. The underlined region was cloned to produce untagged recombinant proteins. (b) Sequence alignment of SAP90/PSD-95 and SAP97 (isoform CRAa) along with the consensus sequence.
202 R.R. Vandanapu et al. / Protein Expression and Purification 68 (2009) 201–207
of highly pure untagged SH3-HOOK-GUK modules of SAP90/PSD-95 and SAP97 and compared their conformational and unfoldingtransitions using a number of spectroscopic and calorimetricmethods. We report that despite high sequence similarity con-formation and stability of these two crucial domains are quitedissimilar.
Materials and methods
Sub-cloning of SAP90/PSD-95 and SAP97 in a pET21a vector
The DNA encoding SH3-HOOK-GUK region of rat SAP90/PSD-95(1291–2228 of Accession No. X66474) and SAP97 (2141–3187 ofAccession No. U14950, the splice isoform containing the I2 inser-tion) and cloned in pET21a vector (Novagen) to over-express bothprotein fragments without any fusion tag. The PCR product wascloned first blunt-end into a TOPO vector (Invitrogen) and thensub-cloned in the pET21a expression vector using the NdeI andEcoRI restriction sites and checked subsequently by DNA-sequencing.
Over-expression of untagged proteins
The recombinant protein SAP90/PSD-95 was expressed in thebacterial strain Escherichia coli BL21(DE3) (Invitrogen) using LBmedium after induction with 0.5 mM isopropyl thio-b-D-galacto-pyranoside (IPTG) for 10 h at 37 �C containing 100 lg ampicillin.Over-expression of SAP97 in E. coli was induced with 0.2 mM IPTGfor 15 h at 25 or 18 �C, respectively.
Hydrophobic interaction chromatography (HIC) and gel filtration
A protocol for the purification of untagged proteins was devel-oped. After induction, the harvested bacterial cells were lysed bylysozyme and sonication at 4 �C in lysis buffer (50 mM Tris–HCl,pH 7.0 containing 1 mM EDTA, 1 mM DTT, 100 mM NaCl and0.1 mM phenyl methyl sulphonyl fluoride (PMSF) [13–15]. Thesupernatant was precipitated with 20% ammonium sulfate pre-pared in lysis buffer and centrifuged.
After centrifugation, the supernatant was applied to a phenyl-Sepharose column (GE Life sciences), equilibrated with same buffer(lysis buffer) containing 20% ammonium sulfate and then washedwith the same buffer but containing 10% ammonium sulfate. Pro-tein was eluted using ammonium sulfate free buffer (50 mMTris–HCl, pH 7.0, 1 mM EDTA, 1 mM DTT and 100 mM NaCl). Theconcentrated protein was further purified on a Superdex 75 size-exclusion column (Pharmacia) equilibrated in 50 mM Tris–HCl(pH 7.5) buffer containing 1 mM EDTA, 1 mM DTT and 100 mMNaCl.
Circular dichroism (CD) measurements
Near- and far-UV CD spectra were recorded on a Jasco J-815spectropolarimeter at 25 �C with 5 accumulations in 25 mM Tris–HCl, pH 7.5 buffer containing 100 mM KCl and 1 mM DTT. Forfar-UV CD, the path length of cuvette used was 0.05 cm with a pro-tein concentration of 0.5 mg/ml. Near-UV CD was performed in thesame buffer in a 1 cm path length cell at a protein concentration of1 mg/ml. All spectra were baseline corrected. Ellipticity data wereexpressed in millidegree.

R.R. Vandanapu et al. / Protein Expression and Purification 68 (2009) 201–207 203
Differential scanning calorimetry measurements
Differential scanning calorimetry measurements were made ona Microcal VP-DSC microcalorimeter (MicroCal, LLC Europe) at aheating rate of 1 �C/min from 10 to 65 �C in case of SAP 90/PSD-95 and from 10 to 75 �C in case of SAP97. For all the measurements,protein was prepared in 50 mM Tris pH 7.5, 100 mM KCl. The con-centration of protein in measuring cell was 0.5 mg/ml. The temper-ature dependence of specific heat capacity (Cp) was analysedaccording to the simple two-state model, assuming the differencebetween heat capacities of the denatured and native protein states(DCp) is independent of temperature (All values were normalizedby protein molecular weight). The experimental data were decon-voluted using ORIGIN software provided by the manufacturer.
Fig. 2. Schematic representation of the protocol for the purification of SH3-HOOK-GUK modules of PSD-95/SAP90 and SAP97 proteins.
Fluorescence spectroscopy
Fluorescence emission spectra were recorded in the correctspectrum mode on a F-4500 Hitachi Fluorescence spectrophotom-eter using an excitation wavelength of 295 nm in 25 mM Tris–HCl,pH 7, containing 100 mM KCl and 1 mM DTT. The excitation andemission band passes were set at 5 nm each. For 8-anilino-1-naph-thalene sulfonic acid (ANS) binding experiments, ANS (final con-centration 100 lM) was mixed with the protein solution in25 mM Tris buffer, pH 7 containing 50 mM KCl, incubated for15 min and spectra were recorded from 400 to 600 nm at the exci-tation of 365 nm in the correct spectrum mode. ANS buffer blankspectra were also recorded under identical conditions withoutany protein.
Equilibrium unfolding studies
Equilibrium unfolding of both SAP90/PSD-95 and SAP97 pro-teins (0.1 mg/ml) was carried out using guanidinium chloride(GdmCl) concentrations in the range of 0–6 M with increments of0.1 M. Trp fluorescence was monitored by exciting the sample at295 nm and fluorescence signal monitored at 340 nm. Correctedspectra at emission/excitation slits of 5/5 nm were recorded witha response time of 2 s. A two-state model was used for fittingSAP90/PSD-95 and SAP97 proteins. Both fits were evaluated by sta-tistical tests available in the software Graph pad prism. The datawere fitted to two or higher state unfolding models described[16] below and best fit parameters were determined.
Y ¼ fsnþ sd� expð�ðg1�m1�DÞ=RTÞgf1þ expð�ðg1�m1�DÞ=RTÞg ð1Þ
where Y is the observed spectroscopic signal, sn and sd representsspectroscopic signal of native and denatured protein, g1 and m1represents the free energy change and slope of the transition, D isthe denaturant concentration, T is the temperature in Kelvin andR is the universal gas constant (value = 1.987 cal K�1 mol�1).
Thermal unfolding by circular dichroism measurements
Circular dichroism measurements were made on a JASCO J-815spectropolarimeter using 1 cm path length cuvette at 220 nm.Temperature was maintained using peltier heating system at a rateof 1 �C/min. Spectra were recorded in buffer containing 50 mMTris–Cl (pH 7.5), 100 mM KCl, 1 mM DTT and protein concentrationof 0.25 mg/ml was used for thermal unfolding studies and the datawere fit using ORIGIN software.
Results
SH3-HOOK-GUK domains of SAP90/PSD-95 and SAP97
To overcome the problem of fusion tag removal, we cloned bothSH3-HOOK-GUK domains (SAP90/PSD-95 and SAP97) in a pET21aexpression vector without tag and over-expressed the constructsin E. coli BL21 (DE3) strain. SAP90/PSD-95 was expressed as a solu-ble protein, when induced at 37 �C with 0.5 mM IPTG. However, incase of SAP97, only a fraction of about 30% expressed as solubleprotein while the rest of the protein incorporated into inclusionbodies. To enhance the protein expression in the soluble fraction,we varied the expression conditions, such as induction at low tem-perature (25 �C) and the use of lower concentrations of IPTG(0.2 mM). At these conditions, the soluble fraction contains morethan 60% of the corresponding protein.
For subsequent biophysical and biochemical studies we de-signed a strategy to standardize the method of purification forthe untagged SH3-HOOK-GUK region of MAGUKs based on theapplication of hydrophobic interaction chromatography (HIC). Inorder to purify SAP90/PSD-95 or SAP97, we loaded the bacterial ly-sate to a phenyl-Sepharose column in the presence of 20% ammo-nium sulfate. Both proteins were bound to the resin efficiently inthe presence of 20% ammonium sulfate and 10% ammonium sul-fate was used for washing the column. Minor impurities were re-moved by adopting gel filtration as a final step of purification(Fig. 2). We used a Superdex 75 preparative column to obtain a bet-ter resolution (Scheme 1). As shown by SDS–PAGE, the proteinsthus purified were highly homogenous and stable after gel-filtra-tion chromatography.
Fluorescence spectroscopy and surface hydrophobicity
SH3-HOOK-GUK domains of SAP90/PSD-95 and SAP 97 have 5and 3 Trp residues respectively. We used Trp fluorescence to assessthe protein tertiary structure and proper folding. Both protein do-mains exhibit wavelength maxima at about 340 and 343 nm, sug-gesting that the Trp is not buried inside the protein core (Fig. 3a).Moreover, both proteins exhibited a similar fluorescence spectro-scopic pattern suggesting that their Trp microenvironments arecomparable as one can expect from their primary structures. Wethen used extrinsic fluorescence to compare the hydrophobicityof the proteins using ANS as a probe. Both proteins exhibited only
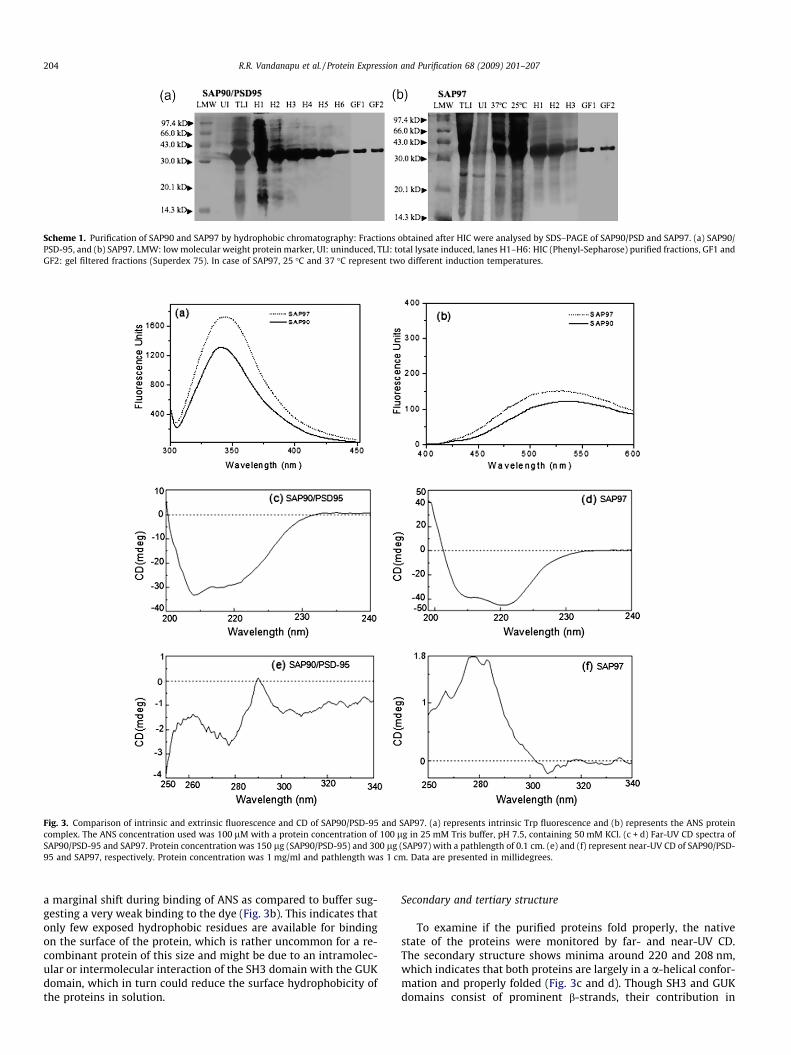
Fig. 3. Comparison of intrinsic and extrinsic fluorescence and CD of SAP90/PSD-95 and SAP97. (a) represents intrinsic Trp fluorescence and (b) represents the ANS proteincomplex. The ANS concentration used was 100 lM with a protein concentration of 100 lg in 25 mM Tris buffer, pH 7.5, containing 50 mM KCl. (c + d) Far-UV CD spectra ofSAP90/PSD-95 and SAP97. Protein concentration was 150 lg (SAP90/PSD-95) and 300 lg (SAP97) with a pathlength of 0.1 cm. (e) and (f) represent near-UV CD of SAP90/PSD-95 and SAP97, respectively. Protein concentration was 1 mg/ml and pathlength was 1 cm. Data are presented in millidegrees.
Scheme 1. Purification of SAP90 and SAP97 by hydrophobic chromatography: Fractions obtained after HIC were analysed by SDS–PAGE of SAP90/PSD and SAP97. (a) SAP90/PSD-95, and (b) SAP97. LMW: low molecular weight protein marker, UI: uninduced, TLI: total lysate induced, lanes H1–H6: HIC (Phenyl-Sepharose) purified fractions, GF1 andGF2: gel filtered fractions (Superdex 75). In case of SAP97, 25 �C and 37 �C represent two different induction temperatures.
204 R.R. Vandanapu et al. / Protein Expression and Purification 68 (2009) 201–207
a marginal shift during binding of ANS as compared to buffer sug-gesting a very weak binding to the dye (Fig. 3b). This indicates thatonly few exposed hydrophobic residues are available for bindingon the surface of the protein, which is rather uncommon for a re-combinant protein of this size and might be due to an intramolec-ular or intermolecular interaction of the SH3 domain with the GUKdomain, which in turn could reduce the surface hydrophobicity ofthe proteins in solution.
Secondary and tertiary structure
To examine if the purified proteins fold properly, the nativestate of the proteins were monitored by far- and near-UV CD.The secondary structure shows minima around 220 and 208 nm,which indicates that both proteins are largely in a a-helical confor-mation and properly folded (Fig. 3c and d). Though SH3 and GUKdomains consist of prominent b-strands, their contribution in
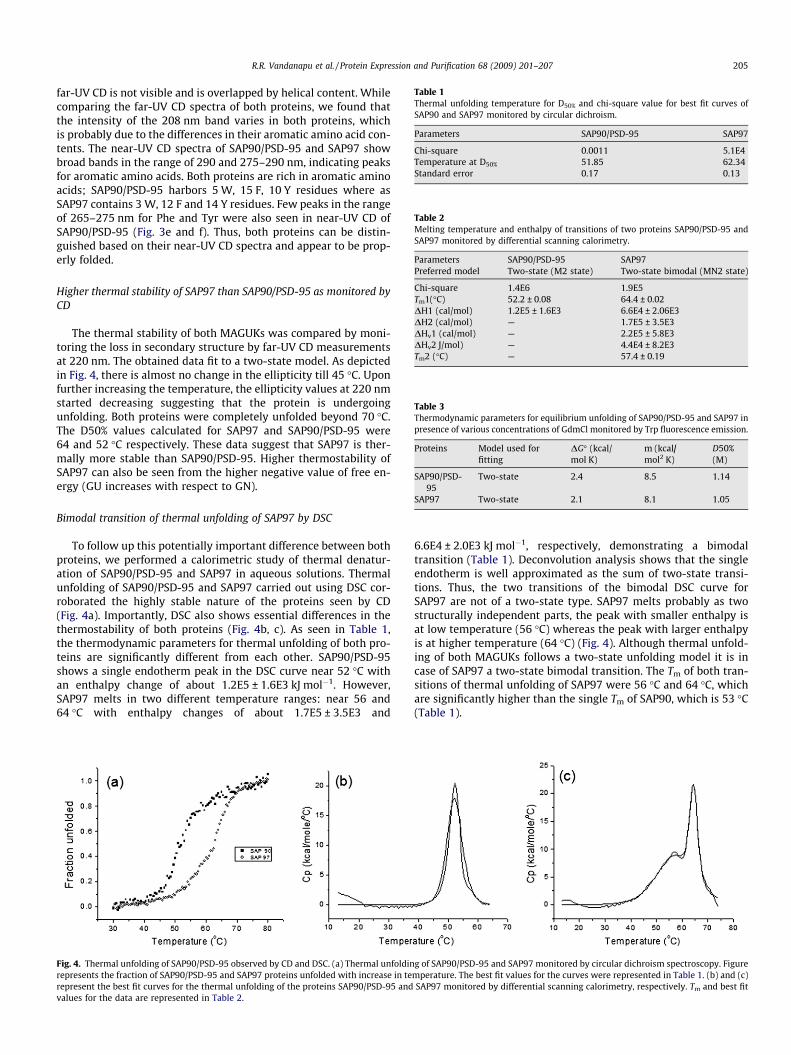
Table 1Thermal unfolding temperature for D50% and chi-square value for best fit curves ofSAP90 and SAP97 monitored by circular dichroism.
Parameters SAP90/PSD-95 SAP97
Chi-square 0.0011 5.1E4Temperature at D50% 51.85 62.34Standard error 0.17 0.13
Table 2Melting temperature and enthalpy of transitions of two proteins SAP90/PSD-95 andSAP97 monitored by differential scanning calorimetry.
Parameters SAP90/PSD-95 SAP97Preferred model Two-state (M2 state) Two-state bimodal (MN2 state)
Chi-square 1.4E6 1.9E5Tm1(�C) 52.2 ± 0.08 64.4 ± 0.02DH1 (cal/mol) 1.2E5 ± 1.6E3 6.6E4 ± 2.06E3DH2 (cal/mol) — 1.7E5 ± 3.5E3DHv1 (cal/mol) — 2.2E5 ± 5.8E3DHv2 J/mol) — 4.4E4 ± 8.2E3Tm2 (�C) — 57.4 ± 0.19
Table 3Thermodynamic parameters for equilibrium unfolding of SAP90/PSD-95 and SAP97 inpresence of various concentrations of GdmCl monitored by Trp fluorescence emission.
Proteins Model used forfitting
DG� (kcal/mol K)
m (kcal/mol2 K)
D50%(M)
SAP90/PSD-95
Two-state 2.4 8.5 1.14
SAP97 Two-state 2.1 8.1 1.05
R.R. Vandanapu et al. / Protein Expression and Purification 68 (2009) 201–207 205
far-UV CD is not visible and is overlapped by helical content. Whilecomparing the far-UV CD spectra of both proteins, we found thatthe intensity of the 208 nm band varies in both proteins, whichis probably due to the differences in their aromatic amino acid con-tents. The near-UV CD spectra of SAP90/PSD-95 and SAP97 showbroad bands in the range of 290 and 275–290 nm, indicating peaksfor aromatic amino acids. Both proteins are rich in aromatic aminoacids; SAP90/PSD-95 harbors 5 W, 15 F, 10 Y residues where asSAP97 contains 3 W, 12 F and 14 Y residues. Few peaks in the rangeof 265–275 nm for Phe and Tyr were also seen in near-UV CD ofSAP90/PSD-95 (Fig. 3e and f). Thus, both proteins can be distin-guished based on their near-UV CD spectra and appear to be prop-erly folded.
Higher thermal stability of SAP97 than SAP90/PSD-95 as monitored byCD
The thermal stability of both MAGUKs was compared by moni-toring the loss in secondary structure by far-UV CD measurementsat 220 nm. The obtained data fit to a two-state model. As depictedin Fig. 4, there is almost no change in the ellipticity till 45 �C. Uponfurther increasing the temperature, the ellipticity values at 220 nmstarted decreasing suggesting that the protein is undergoingunfolding. Both proteins were completely unfolded beyond 70 �C.The D50% values calculated for SAP97 and SAP90/PSD-95 were64 and 52 �C respectively. These data suggest that SAP97 is ther-mally more stable than SAP90/PSD-95. Higher thermostability ofSAP97 can also be seen from the higher negative value of free en-ergy (GU increases with respect to GN).
Bimodal transition of thermal unfolding of SAP97 by DSC
To follow up this potentially important difference between bothproteins, we performed a calorimetric study of thermal denatur-ation of SAP90/PSD-95 and SAP97 in aqueous solutions. Thermalunfolding of SAP90/PSD-95 and SAP97 carried out using DSC cor-roborated the highly stable nature of the proteins seen by CD(Fig. 4a). Importantly, DSC also shows essential differences in thethermostability of both proteins (Fig. 4b, c). As seen in Table 1,the thermodynamic parameters for thermal unfolding of both pro-teins are significantly different from each other. SAP90/PSD-95shows a single endotherm peak in the DSC curve near 52 �C withan enthalpy change of about 1.2E5 ± 1.6E3 kJ mol�1. However,SAP97 melts in two different temperature ranges: near 56 and64 �C with enthalpy changes of about 1.7E5 ± 3.5E3 and
Fig. 4. Thermal unfolding of SAP90/PSD-95 observed by CD and DSC. (a) Thermal unfoldinrepresents the fraction of SAP90/PSD-95 and SAP97 proteins unfolded with increase in terepresent the best fit curves for the thermal unfolding of the proteins SAP90/PSD-95 andvalues for the data are represented in Table 2.
6.6E4 ± 2.0E3 kJ mol�1, respectively, demonstrating a bimodaltransition (Table 1). Deconvolution analysis shows that the singleendotherm is well approximated as the sum of two-state transi-tions. Thus, the two transitions of the bimodal DSC curve forSAP97 are not of a two-state type. SAP97 melts probably as twostructurally independent parts, the peak with smaller enthalpy isat low temperature (56 �C) whereas the peak with larger enthalpyis at higher temperature (64 �C) (Fig. 4). Although thermal unfold-ing of both MAGUKs follows a two-state unfolding model it is incase of SAP97 a two-state bimodal transition. The Tm of both tran-sitions of thermal unfolding of SAP97 were 56 �C and 64 �C, whichare significantly higher than the single Tm of SAP90, which is 53 �C(Table 1).
g of SAP90/PSD-95 and SAP97 monitored by circular dichroism spectroscopy. Figuremperature. The best fit values for the curves were represented in Table 1. (b) and (c)SAP97 monitored by differential scanning calorimetry, respectively. Tm and best fit
206 R.R. Vandanapu et al. / Protein Expression and Purification 68 (2009) 201–207
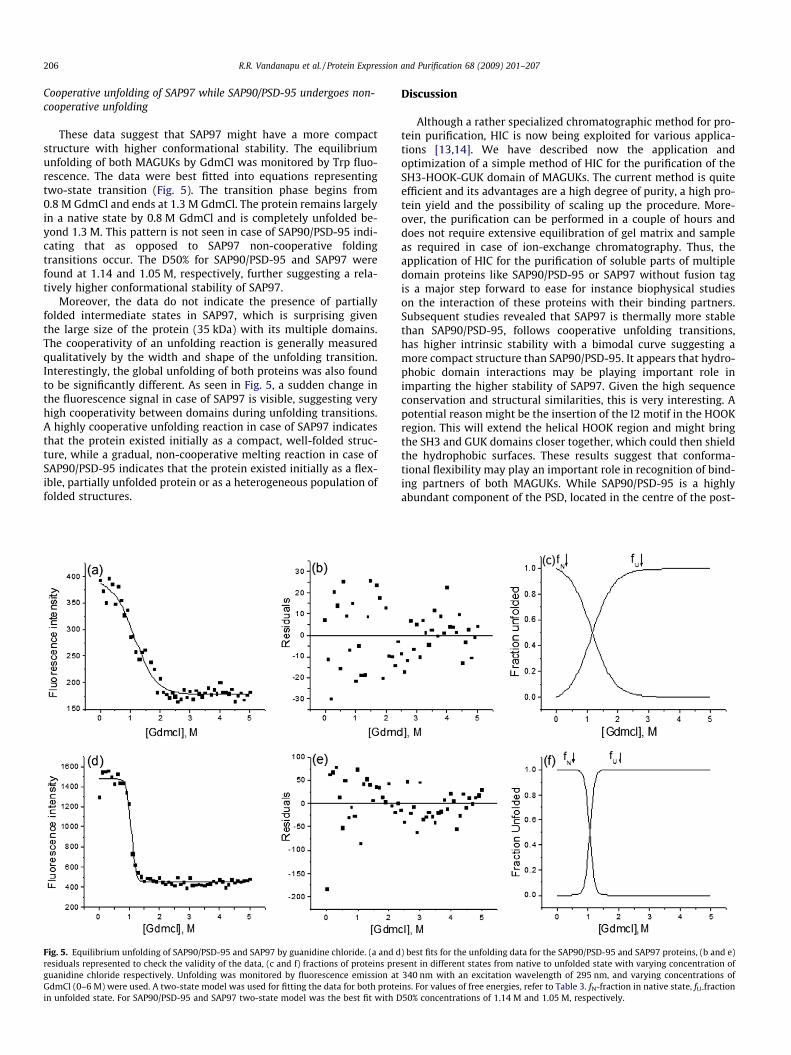
Cooperative unfolding of SAP97 while SAP90/PSD-95 undergoes non-cooperative unfolding
These data suggest that SAP97 might have a more compactstructure with higher conformational stability. The equilibriumunfolding of both MAGUKs by GdmCl was monitored by Trp fluo-rescence. The data were best fitted into equations representingtwo-state transition (Fig. 5). The transition phase begins from0.8 M GdmCl and ends at 1.3 M GdmCl. The protein remains largelyin a native state by 0.8 M GdmCl and is completely unfolded be-yond 1.3 M. This pattern is not seen in case of SAP90/PSD-95 indi-cating that as opposed to SAP97 non-cooperative foldingtransitions occur. The D50% for SAP90/PSD-95 and SAP97 werefound at 1.14 and 1.05 M, respectively, further suggesting a rela-tively higher conformational stability of SAP97.
Moreover, the data do not indicate the presence of partiallyfolded intermediate states in SAP97, which is surprising giventhe large size of the protein (35 kDa) with its multiple domains.The cooperativity of an unfolding reaction is generally measuredqualitatively by the width and shape of the unfolding transition.Interestingly, the global unfolding of both proteins was also foundto be significantly different. As seen in Fig. 5, a sudden change inthe fluorescence signal in case of SAP97 is visible, suggesting veryhigh cooperativity between domains during unfolding transitions.A highly cooperative unfolding reaction in case of SAP97 indicatesthat the protein existed initially as a compact, well-folded struc-ture, while a gradual, non-cooperative melting reaction in case ofSAP90/PSD-95 indicates that the protein existed initially as a flex-ible, partially unfolded protein or as a heterogeneous population offolded structures.
Fig. 5. Equilibrium unfolding of SAP90/PSD-95 and SAP97 by guanidine chloride. (a and dresiduals represented to check the validity of the data, (c and f) fractions of proteins preguanidine chloride respectively. Unfolding was monitored by fluorescence emission atGdmCl (0–6 M) were used. A two-state model was used for fitting the data for both protein unfolded state. For SAP90/PSD-95 and SAP97 two-state model was the best fit with D
Discussion
Although a rather specialized chromatographic method for pro-tein purification, HIC is now being exploited for various applica-tions [13,14]. We have described now the application andoptimization of a simple method of HIC for the purification of theSH3-HOOK-GUK domain of MAGUKs. The current method is quiteefficient and its advantages are a high degree of purity, a high pro-tein yield and the possibility of scaling up the procedure. More-over, the purification can be performed in a couple of hours anddoes not require extensive equilibration of gel matrix and sampleas required in case of ion-exchange chromatography. Thus, theapplication of HIC for the purification of soluble parts of multipledomain proteins like SAP90/PSD-95 or SAP97 without fusion tagis a major step forward to ease for instance biophysical studieson the interaction of these proteins with their binding partners.Subsequent studies revealed that SAP97 is thermally more stablethan SAP90/PSD-95, follows cooperative unfolding transitions,has higher intrinsic stability with a bimodal curve suggesting amore compact structure than SAP90/PSD-95. It appears that hydro-phobic domain interactions may be playing important role inimparting the higher stability of SAP97. Given the high sequenceconservation and structural similarities, this is very interesting. Apotential reason might be the insertion of the I2 motif in the HOOKregion. This will extend the helical HOOK region and might bringthe SH3 and GUK domains closer together, which could then shieldthe hydrophobic surfaces. These results suggest that conforma-tional flexibility may play an important role in recognition of bind-ing partners of both MAGUKs. While SAP90/PSD-95 is a highlyabundant component of the PSD, located in the centre of the post-
) best fits for the unfolding data for the SAP90/PSD-95 and SAP97 proteins, (b and e)sent in different states from native to unfolded state with varying concentration of340 nm with an excitation wavelength of 295 nm, and varying concentrations of
ins. For values of free energies, refer to Table 3. fN-fraction in native state, fU-fraction50% concentrations of 1.14 M and 1.05 M, respectively.

R.R. Vandanapu et al. / Protein Expression and Purification 68 (2009) 201–207 207
synaptic scaffold and thought be involved in its organization [17],SAP97 containing the I2 insertion is to a large degree located extra-synaptic and also seems to be involved in other functions like traf-ficking of AMPA-receptors from Golgi to the synapse [17]. Thesedifferent locations and functions might impose the necessity of amore rigid and compact structure of the SH3-HOOK-GUK domainin case of SAP97 and a more flexible conformation in case ofSAP90/PSD-95. In summary our data suggest that an insertion inthe SH3-HOOK-GUK region can influence the structural organiza-tion and folding dynamics of MAGUK proteins and the presentedproduction and purification scheme for recombinant SH3-HOOK-GUK protein modules will help to elucidate further the moleculardynamics of protein–protein interactions in this region.
Acknowledgments
A.K.S. was supported by the DBT-PDF. The experimental workwas supported by a joint DBT-BMBF and DAAD-DST PPP Grantaward to Y.S. and M.R.K.
References
[1] J.M. Montgomery, P.L. Zamorano, C.C. Garner, MAGUKs in synapse assemblyand function: an emerging view, Cell Mol. Life Sci. 61 (2004) 911–929.
[2] A.W. McGee, S.R. Dakoji, O. Olsen, D.S. Bredt, W.A. Lim, K.E. Prehoda, Structureof the SH3-guanylate kinase module from PSD-95 suggests a mechanism forregulated assembly of MAGUK scaffolding proteins, Mol. Cell 8 (2001) 1291–1301.
[3] G.A. Tavares, E.H. Panepucci, A.T. Brunger, Structural characterization of theintramolecular interaction between the SH3 and guanylate kinase domains ofPSD-95, Mol. Cell 8 (2001) 1313–1325.
[4] E. Kim, S. Naisbitt, Y.P. Hsueh, A. Rao, A. Rothschild, A.M. Craig, M. Sheng, GKAP,a novel synaptic protein that interacts with the guanylate kinase-like domainof the PSD-95/SAP90 family of channel clustering molecules, J. Cell Biol. 136(1997) 669–678.
[5] H. Wu, C. Reissner, S. Kuhlendahl, B. Coblentz, S. Reuver, S. Kindler, E.D.Gundelfinger, C.C. Garner, Intramolecular interactions regulate SAP97 bindingto GKAP, EMBO J. 19 (2000) 5740–5751.
[6] D.T. Pak, S. Yang, S. Rudolph-Correia, E. Kim, M. Sheng, Regulation of dendriticspine morphology by SPAR, a PSD-95-associated RapGAP, Neuron 31 (2001)289–303.
[7] A.W. McGee, D.S. Bredt, Identification of an intramolecular interactionbetween the SH3 and guanylate kinase domains of PSD-95, J. Biol. Chem.274 (1999) 17431–17436.
[8] L. Funke, S. Dakoji, D.S. Bredt, Membrane-associated guanylate kinasesregulate adhesion and plasticity at cell junctions, Annu. Rev. Biochem. 74(2005) 219–245.
[9] I. Paarmann, O. Spangenberg, A. Lavie, M. Konrad, Formation of complexesbetween Ca2+-calmodulin and the synapse-associated protein SAP97 requiresthe SH3 domain-guanylate kinase domain-connecting HOOK region, J. Biol.Chem. 277 (2002) 40832–40838.
[10] N. Masuko, K. Makino, H. Kuwahara, K. Fukunaga, T. Sudo, N. Araki, H.Yamamoto, Y. Yamada, E. Miyamoto, H. Saya, Interaction of NE-dlg/SAP102, aneuronal and endocrine tissue-specific membrane-associated guanylatekinase protein, with calmodulin and PSD-95/SAP90. A possible regulatoryrole in molecular clustering at synaptic sites, J. Biol. Chem. 274 (1999) 5782–5790.
[11] A. Levitzki, Targeting signal transduction for disease therapy, Curr. Opin. CellBiol. 8 (1996) 239–244.
[12] D.C. Dalgarno, M.C. Botfield, R.J. Rickless, SH3 domains and drug design:Ligands, structure and biological function, Biopolymers 43 (1998) 383–400.
[13] B.F. Roettger, M.R. Ladisch, Hydrophobic interaction chromatography,Biotechnol. Adv. 7 (1989) 15–29.
[14] J.R. Fisher, Y. Sharma, S. Iuliano, R.A. Piccioti, D. Krylov, J. Hurley, J. Roder, A.Jeromin, Purification of myristoylated and nonmyristoylated neuronal calciumsensor-1 using single-step hydrophobic interaction chromatography, ProteinExpress. Purif. 20 (2000) 66–72.
[15] M.K. Jobby, Y. Sharma, Rapid purification of recombinant bB2-crystallin usinghydrophobic interaction chromatography, Protein Express. Purif. 28 (2003)158–164.
[16] H.C. Hung, Y.H. Chen, G.Y. Liu, H.J. Lee, G.G. Chang, Equilibrium proteinfolding–unfolding process involving multiple intermediates, Bull. Math. Biol.65 (2003) 553–570.
[17] T.M. Boeckers, The postsynaptic density, Cell Tissue Res. 326 (2006) 409–422.
Related Documents











