Eur. J. Biochem. 242, 608-618 (1996) Q FEBS 1996 Structural characterisation of human recombinant glycohormones follitropin, lutropin and choriogonadotropin expressed in Chinese hamster ovary cells Angela AMORESANO'.', Rosa SICILIAN04, Stefania ORRU Antonino SIRNA' and Piero PUCCI' ' ' Servizio di Spettrometria di Massa del CNR, Naples, Italy ' CEINGE s.c.r.1. Biotecnologie Avanzate, Naples, Italy ' Istituto di Ricerche Cesare Serono, S.p.A., Chemical Department, Ardea, Roma, Italy (Received 31 July/l0 October 1996) - EJB 96 1136/3 Roberta NAPOLEONI', Valter ALTAROCCA5, Eva DE LUCA', Dipartimento di Chimica Organica e Biologica, Universith Federico 11 di Napoli, Italy Istituto Scienze dell' Alimentazione CNR, Avellino, Italy The a and chains from human recombinant gonadotropins follitropin, lutropin and choriogonadotro- pin expressed in CHO cells have been structurally characterised both at the protein and at the carbohydrate level by using advanced mass spectrometric procedures. The three a chains share the same amino acid sequence while they display different glycosylation patterns. The oligosaccharide structures detected belong to the complex-type glycans with different degree of sialylation. Partial proteolytic processing occurred at the N-terminus of the follitropin p chain and at the C-terminus of the lutropin 11 ' chain. The N-linked glycans from the three p chains were found to contain fucosylated and sialylated diantennary and triantennary complex-type structures. The follitropin p chain showed the presence of N-acetyllactos- amine repeats on the antennae. The overall structure of the recombinant glycohormones corresponds to their natural counterparts with the exception that sulphated terminal glycosylation is missing. Keywords: gonadotropin ; recombinant glycoprotein; oligosaccharide; structural characterisation ; mass spectrometry. The gonadotropins constitute a family of closely related gly- coproteins sharing a high degree of structural similarity; they are commonly referred to as glycoprotein hormones (1 1. Follitropin (follicle-stimulating hormone, FSH), lutropin (luteinizing hor- mone, LH) and thyrotropin (thyroid-stimulating hormone, TSH) are secreted by the specialised cells of the anterior pituitary gland, whilst choriogonadotropin (chorionic gonadotropin, CG) is produced by placental cells. These hormones control several physiological functions via individual response pathways, in- volving binding to specific receptors and synthesis of chemical second messengers, such as CAMP, calcium, etc. [2]. Particu- larly, FSH, LH and CG are involved in the regulation of the reproductive functions of the organism. Each hormone is a het- erodimer consisting of non-covalently bound n and 11 subunits. Within a species, the a subunits of all hormones have an iden- tical amino acid sequence, whereas the chain is distinct and confers hormone-specific features to the heterodimer [3]. Both 0: and p subunits are tightly packed proteins containing five and Corresponrlericr to P. Pucci, Servizio di Spettrometria di Massa del Phone: 1-39 81 5465136. Fux: f39 81 5465484. Abhrevi~rtiom. CG, choriogonadotropin; CHO, Chinese hamster ovary : ESMS, electrospray mass spectrometry ; FABMS, fast-atom-botn- bardment mass spectrometry: FSH, follitropin; Fuc, fucose; Hex, hex- ose; HexNAc, N-acetylhexosamine; LH, lutropin; PNGase F. peptide- N-glycosidase F; Sia, sialic acid ; TSH, thyroid stimulating hormonc. Enzymes. Endoproteinase Asp-N (EC 3.4.24.33); endoproteinase Glu-C (EC 3.4.21.19); 0-glycosidase (EC 3.2.1.97); peptide N-glycosi- dase F, PNGase F (EC 3.5.1.52); sialidase (EC 3.2.1.18); trypsin (EC 3.4.21.4). CNR, via S. Pansini 5. 1-80131 Napoli, Italy six disulphide bridges respectively. The chemical assessment of the disulphide pairings has been a long-lasting challenge which has left the S-S pattern still unsolved 14-61. Recently, the crys- tal structure of human CG has been reported showing a folding motif in the a and /I chains similar to that found in the growth factors, thus suggesting that these glyco-hormones might belong to the growth factors superfamily [7]. Two N-glycosylation sites are present in the (x subunit of all the hormones and in the p chains from FSH and CG, whereas /3 LH possesses only one; moreover CG has four 0-linked glycosylation sites within the C-terminal region. Despite the highly similar amino acid sequence, the glycosylation pattern is hormone-specific and is involved in the modulation of biological activities since the glycan chains are directly responsible for rec- ognition and interactions with specific receptors. Recent studies, in fact, reported that specifically deglycosylated FSH mutants displayed different receptor binding capabilities [8], whilst non- glycosylated recombinant LH failed to interact with its receptor [91. Furthermore, the entire set of hormones exhibits variable in vivn activity depending on the extent of glycosylation and the presence of different glycosylated species 11 01. These findings clearly demonstrate the central role of glycans in determining and modulating the biological functions of these hormones. The peculiar biological activity of the gonadotropins in regu- lating reproductive functions of organisms has greatly stimulated the effort to produce active glyco-hormones by DNA recombi- nant methodologies for therapeutic purposes. Human recombi- nant gonadotropins have been expressed in suitable maminalian cell lines having the enzymatic machinery to produce glycosyla- tion patterns similar to the natural counterpart 111 1. However,

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Biochem. 242, 608-618 (1996) Q FEBS 1996
Structural characterisation of human recombinant glycohormones follitropin, lutropin and choriogonadotropin expressed in Chinese hamster ovary cells Angela AMORESANO'.', Rosa SICILIAN04, Stefania ORRU Antonino SIRNA' and Piero PUCCI' ' ' Servizio di Spettrometria di Massa del CNR, Naples, Italy
' CEINGE s.c.r.1. Biotecnologie Avanzate, Naples, Italy
' Istituto di Ricerche Cesare Serono, S.p.A., Chemical Department, Ardea, Roma, Italy
(Received 31 July/ l0 October 1996) - EJB 96 1136/3
Roberta NAPOLEONI', Valter ALTAROCCA5, Eva DE LUCA',
Dipartimento di Chimica Organica e Biologica, Universith Federico 11 di Napoli, Italy
Istituto Scienze dell' Alimentazione CNR, Avellino, Italy
The a and chains from human recombinant gonadotropins follitropin, lutropin and choriogonadotro- pin expressed in CHO cells have been structurally characterised both at the protein and at the carbohydrate level by using advanced mass spectrometric procedures. The three a chains share the same amino acid sequence while they display different glycosylation patterns. The oligosaccharide structures detected belong to the complex-type glycans with different degree of sialylation. Partial proteolytic processing occurred at the N-terminus of the follitropin p chain and at the C-terminus of the lutropin 11' chain. The N-linked glycans from the three p chains were found to contain fucosylated and sialylated diantennary and triantennary complex-type structures. The follitropin p chain showed the presence of N-acetyllactos- amine repeats on the antennae.
The overall structure of the recombinant glycohormones corresponds to their natural counterparts with the exception that sulphated terminal glycosylation is missing.
Keywords: gonadotropin ; recombinant glycoprotein; oligosaccharide; structural characterisation ; mass spectrometry.
The gonadotropins constitute a family of closely related gly- coproteins sharing a high degree of structural similarity; they are commonly referred to as glycoprotein hormones (1 1. Follitropin (follicle-stimulating hormone, FSH), lutropin (luteinizing hor- mone, LH) and thyrotropin (thyroid-stimulating hormone, TSH) are secreted by the specialised cells of the anterior pituitary gland, whilst choriogonadotropin (chorionic gonadotropin, CG) is produced by placental cells. These hormones control several physiological functions via individual response pathways, in- volving binding to specific receptors and synthesis of chemical second messengers, such as CAMP, calcium, etc. [2]. Particu- larly, FSH, LH and CG are involved in the regulation of the reproductive functions of the organism. Each hormone is a het- erodimer consisting of non-covalently bound n and 11 subunits. Within a species, the a subunits of all hormones have an iden- tical amino acid sequence, whereas the chain is distinct and confers hormone-specific features to the heterodimer [3]. Both 0: and p subunits are tightly packed proteins containing five and
Corresponrlericr to P. Pucci, Servizio di Spettrometria di Massa del
Phone: 1-39 81 5465136. Fux: f39 81 5465484. Abhrevi~rtiom. CG, choriogonadotropin; CHO, Chinese hamster
ovary : ESMS, electrospray mass spectrometry ; FABMS, fast-atom-botn- bardment mass spectrometry: FSH, follitropin; Fuc, fucose; Hex, hex- ose; HexNAc, N-acetylhexosamine; LH, lutropin; PNGase F. peptide- N-glycosidase F ; Sia, sialic acid ; TSH, thyroid stimulating hormonc.
Enzymes. Endoproteinase Asp-N (EC 3.4.24.33); endoproteinase Glu-C (EC 3.4.21.19); 0-glycosidase (EC 3.2.1.97); peptide N-glycosi- dase F, PNGase F (EC 3.5.1.52); sialidase (EC 3.2.1.18); trypsin (EC 3.4.21.4).
CNR, via S. Pansini 5. 1-80131 Napoli, Italy
six disulphide bridges respectively. The chemical assessment of the disulphide pairings has been a long-lasting challenge which has left the S-S pattern still unsolved 14-61. Recently, the crys- tal structure of human CG has been reported showing a folding motif in the a and /I chains similar to that found in the growth factors, thus suggesting that these glyco-hormones might belong to the growth factors superfamily [7].
Two N-glycosylation sites are present in the (x subunit of all the hormones and in the p chains from FSH and CG, whereas /3 LH possesses only one; moreover CG has four 0-linked glycosylation sites within the C-terminal region. Despite the highly similar amino acid sequence, the glycosylation pattern is hormone-specific and is involved in the modulation of biological activities since the glycan chains are directly responsible for rec- ognition and interactions with specific receptors. Recent studies, in fact, reported that specifically deglycosylated FSH mutants displayed different receptor binding capabilities [8], whilst non- glycosylated recombinant LH failed to interact with its receptor [91. Furthermore, the entire set of hormones exhibits variable in vivn activity depending on the extent of glycosylation and the presence of different glycosylated species 11 01. These findings clearly demonstrate the central role of glycans in determining and modulating the biological functions of these hormones.
The peculiar biological activity of the gonadotropins in regu- lating reproductive functions of organisms has greatly stimulated the effort to produce active glyco-hormones by DNA recombi- nant methodologies for therapeutic purposes. Human recombi- nant gonadotropins have been expressed in suitable maminalian cell lines having the enzymatic machinery to produce glycosyla- tion patterns similar to the natural counterpart 111 1. However,

Amoresano et al. ( E m J. Biochem. 242) 609
recombinant glycoproteins intended for pharmaceutical use in humans need to be carefully characterised both in terms of their amino acid sequence and the oligosaccharide structure. Protein glycosylation, in fact, modulates biochemical attributes of pro- teins such as folding, bioactivity, and immunogeneity and it is involved in molecular recognition events. This is particularly true in the case of gonadotropins where subtle differences in the glycosylation pattern are reflected in the alteration of their biological activities ; the presence of an ensemble of differenti- ally glycosylated species might constitute a sophisticated mecha- nism of biological control. Therefore, if the recombinant glyco- hormones show a diffuse microheterogeneity, it is extremely im- portant to characterise the entire set of glycoforms to understand their therapeutic consequences.
This paper reports a detailed structural characterisation of the a and p subunits from human recombinant FSH, LH, and CG produced in Chinese hamster ovary (CHO) cell lines both at the protein and carbohydrate level, including the assessment of the glycosylation sites, using advanced mass spectrometric procedures.
MATERIALS AND METHODS
Materials. Trypsin, dithiothreitol, iodoacetic acid, glycerol and thioglycerol were purchased from Sigma. Peptide N-glycosi- dase F (PNGase F), 0-glycosidase, endoproteinase Asp-N, endoproteinase Glu-C and sialidase from Vibrio cholerae were obtained from Boheringer. Pre-packed PD-I 0 gel filtration car- tridges were from Pharmacia. Pre-packed Sep pak C18 car- tridges were purchased from Waters. All other reagents and solvents were of the highest purity available from Carlo Erba.
Purification of recombinant glycoproteins. The a and p chains from human recombinant gonadotropins (FSH, LH and CG, Ares-Serono) were purified by reverse-phase HPLC on a Vydac C4 column (25X0.46 cm, 5 pm) using 0.1 % trifluoro- acetic acid (solvent A) and 0.07% trifluoroacetic acid in 95% acetonitrile (solvent B). The glycoproteins were eluted by means of a linear gradient of 18-43% solvent B over 25 min. Fractions containing a and p chains were collected, dried in a Speed-Vac centrifuge (Savant), lyophilised twice and stored at -20°C.
Chemical and enzymatic hydrolyses. Individual gonadotro- pins chains (300 pg) were reduced and alkylated as previously described [12]. Reduced and carboxymethylated a chains were digested with CNBr (Sigma) in 70% trifluoroacetic acid at room temperature for 18 h in the dark. Samples were then diluted with water and lyophilised. The peptide mixtures were deglycosyl- ated by treatment with PNGase F in 50 mM ammonium bicar- bonate pH 8.5, overnight at 37°C using 0.1 U/300 pg protein. Deglycosylated CNBr digests were subdigested with endopro- teinase Glu-C in 50 mM ammonium acetate pH 4.0 for 18 h at 37 "C using an enzymehbstrate mass ratio of 1 : 50. Samples were then lyophilised.
Reduced and carboxymethylated p chains from FSH, LH and CG were digested with trypsin in 0.4% ammonium bicarbonate pH 8.5 at 37°C overnight using an enzymehbstrate mass ratio of 1 : 50. Tryptic peptides from LH and CG p chains were further hydrolysed with endoproteinase Asp-N in 50 mM ammonium bicarbonate pH 8.5 containing 10% acetonitrile for 18 h at 37 "C using an enzyme/substrate mass ratio of 1 :SO. Peptides mixtures were N-deglycosylated with PNGase F as above described. The enzymatic digest of /z chain from CG was 0-deglycosylated by incubation with 0.03 U sialidase in 50 mM ammonium acetate pH 5.5, at 37°C for 18 h, followed by treatment with 2.5 mU 0-glycosidase in 50 mM sodium phosphate pH 7.0 at 37°C for 18 h.
Isolation of oligosaccharide chains. N-linked oligosaccha- ride chains released by PNGase F were separated from peptides and 0-glycopeptides by reverse-phase chromatography 011 pre- packed Sep-pak cartridges. 0-linked oligosaccharides were re- leased from the reduced and carboxymethylated p CG chain by reductive elimination in 50 mM NaOH containing 3 M sodium borohydryde at 45 "C for 18 h [13].
Chemical derivatisation procedures. Aliquots of peptide mixtures derived from different digestion procedures were meth- yl esterified with 2 M acetyl chloride in methanol for 1 h at room temperature, dried in a Speed-Vac centrifuge and lyophi- lised twice prior to mass spectrometric analysis [14].
N- and 0-linked oligosaccharides were permethylated with methyl iodide in dimethylsulphoxide using the sodium hydrox- ide procedure to increase the sensitivity of fast-atom-bombard- ment (FAB) MS analyses [15]. Derivatised samples were diluted with water, extracted in chloroform and dried in a Speed-Vac centrifuge.
Fractionation of peptide mixtures. Degl ycosylated peptide mixtures from CNBr and endoprotease Glu-C digests were frac- tionated by HPLC on a Vydac C18 column (25X0.46 cm, 5 pm) using 0.1 % trifluoroacetic acid (solvent A) and 0.07% trifluoro- acetic acid in 95% acetonitrile (solvent B) using a linear gradi- ent of 5-129'0 in 8 tnin and to 55 % in 30 min. Individual pep- tide fractions were manually collected, dried in a Speed-Vac centrifuge and lyophilised twice prior to mass spectrometric analysis.
Mass spectrometric analyses. Intact proteins or individual peptide fractions were submitted to electrospray mass spectrom- etry (ESMS) analyses using either a VG Bio-Q triple quadrupole mass spectrometer or a VG Platform instrument (Micromass). Samples were dissolved in 1 % acetic acid in 50% acetonitrile and 2-10 p1 was injected into the mass spectrometer at a flow rate of 10 pl/min. The quadrupole was scanned over mlz 400- 2000 at 10 s/scan and the spectra were acquired using the MASS-LINX software. Calibration was performed by a separate injection of myoglobin (16951.5 Da). All mass values are re- ported as average masses.
FABMS analyses were performed either on a VG ZAB 2SE double-focusing mass spectrometer or on a VG ZAB T tandem four-sector instrument (Micromass) both equipped with caesium guns operating at 25 kV (2 PA). Samples were dissolved in 5 % acetic acid or metanol and 1-2 p1 were loaded onto a glycerol/ thioglycerol coated probe tip. Spectra were either recorded on ultraviolet-sensitive paper and manually counted or acquired on a VAX work station and elaborated using the OPUS program. Signals were assigned to individual peptides within the protein sequences on the basis of their molecular mass with the aid of a suitable computer program developed at our laboratory [16]. All values are reported as monoisotopic masses.
RESULTS
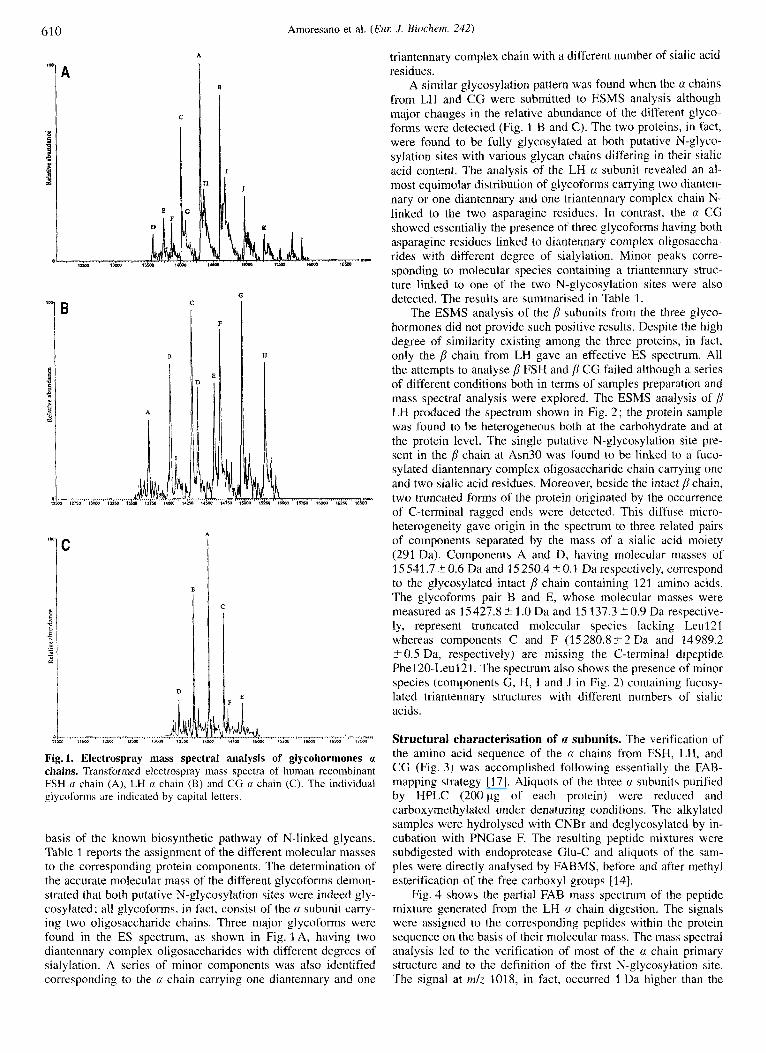
Intact protein analysis. lntact a and chains from human re- combinant gonadotropins (1 00 pg) were purified by HPLC and aliquots of the protein preparations were directly submitted to ESMS analysis. Fig. 1 A shows the electrospray mass spectrum of the FSH a chain transformed on a real mass scale. A series of components were identified in the spectrum demonstrating a high heterogeneity of the protein molecule due to the presence of oligosaccharide chains. The a chain, in fact, is known to pos- sess two N-glycosylation sites at Asn52 and Asn78 [l].
The different glycoforms were identified on the basis of the theoretical molecular mass of the polypeptide portion (10 195.7 Da) and the oligosaccharide structures expected on the
610 Amoresano et al. (Eul: J . Biochenz. 242)
IA A
i s
‘7 B i
G C I
lc
Fig. 1. Electrospray mass spectral analysis of glycohormones a chains. Transformed electrospray mass spectra of human recombinant FSH rx chain (A), LH a chain (B) and CG u: chain (C). The individual glycoforrns are indicated by capital letters.
basis of the known biosynthetic pathway of N-linked glycans. Table 1 reports the assignment of the different molecular masses to the corresponding protein components. The determination of the accurate molecular mass of the different glycoforms demon- strated that both putative N-glycosylation sites were indeed gly- cosylated; all glycoforms, in fact, consist of the a subunit carry- ing two oligosaccharide chains. Three major glycoforms were found in the ES spectrum, as shown in Fig. 1 A, having two diantennary complex oligosaccharides with different degrees of sialylation. A series of minor components was also identified corresponding to the 0: chain carrying one diantennary and one
triantennary complex chain with a different number of sialic acid residues.
A similar glycosylation pattern was found when the a chains from LH and CC were submitted to ESMS analysis although major changes in the relative abundance of the different glyco- forms were detected (Fig. l B and C). The two proteins, in fact, were found to be fully glycosylated at both putative N-glyco- sylation sites with various glycan chains differing in their sialic acid content. The analysis of the LH rx subunit revealed an al- most equimolar distribution of glycoforms carrying two dianten- nary or one diantennary and one triantennary complex chain N- linked to the two asparagine residues. In contrast, the (I CG showed essentially the presence of three glycoforms having both asparagine residues linked to diantennary complex oligosaccha- rides with different degree of sialylation. Minor peaks corre- sponding to molecular species containing a triantennary struc- ture linked to one of the two N-glycosylation sites were also detected. The results are summarised in Table 1.
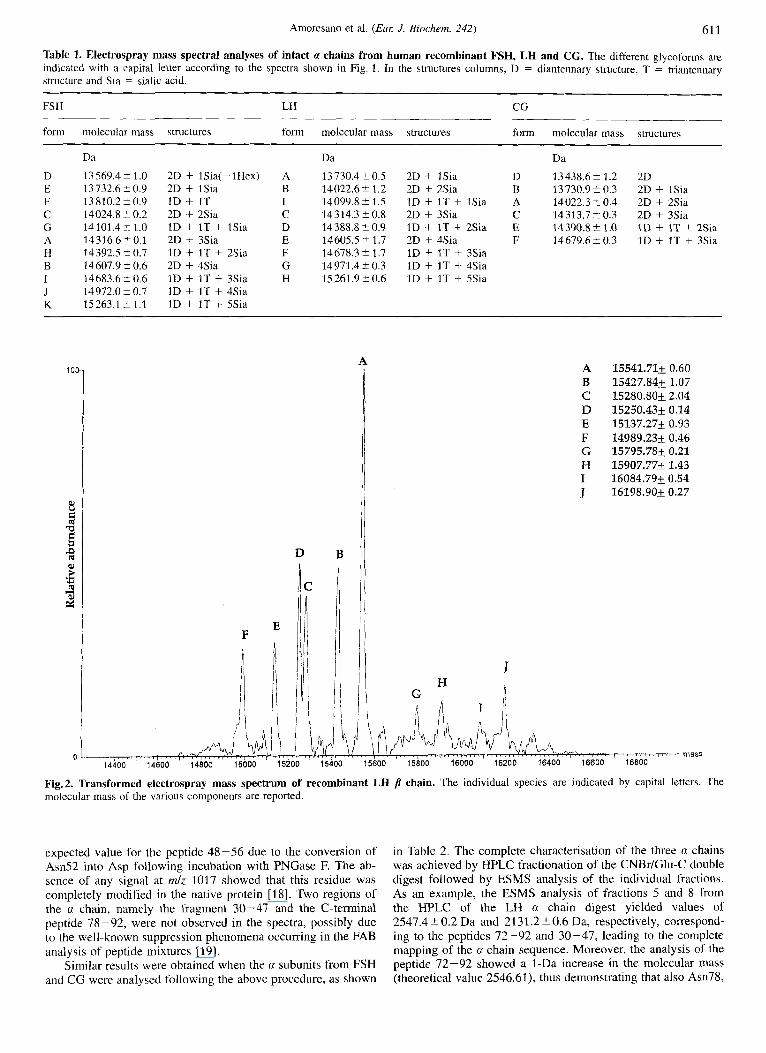
The ESMS analysis of the p subunits from the three glyco- hormones did not provide such positive results. Despite the high degree of similarity existing among the three proteins, in fact, only the p chain from LH gave an effective ES spectrum. All the attempts to analyse p FSH and [I CG failed although a series of different conditions both in terms of samples preparation and mass spectral analysis were explored. The ESMS analysis of p LH produced the spectrum shown in Fig. 2; the protein sample was found to be heterogeneous both at the carbohydrate and at the protein level. The single putative N-glycosylation site pre- sent in the p chain at Am30 was found to be linked to a fuco- sylated diantennary complex oligosaccharide chain carrying one and two sialic acid residues. Moreover, beside the intact /j chain, two truncated forms of the protein originated by the occurrence of C-terminal ragged ends were detected. This diffuse micro- heterogeneity gave origin in the spectrum to three related pairs of components separated by the mass of a sialic acid moiety (291 Da). Components A and D, having molecular masses of 1.5 541.7 t 0.6 Da and IS 250.4 i 0.1 Da respectively, correspond to the glycosylated intact chain containing 121 amino acids. The glycoforms pair B and E, whose molecular masses were measured as 15427.8 2 1 .O Da and 15 137.3 5 0.9 Da respective- ly, represent truncated molecular species lacking Leu121 whereas components C and F (15280.822 Da and 14989.2 2 0.5 Da, respectively) are missing the C-terminal dipeptide Phel20-Leul21. The spectrum also shows the presence of minor species (components G, H, I and J in Fig. 2) containing fucosy- lated triantennary structures with different numbers of sialic acids.
Structural characterisation of a subunits. The verification of the amino acid sequence of the a chains from FSH, LH, and CG (Fig. 3) was accomplished following essentially the FAB- mapping strategy [17]. Aliquots of the three (1 subunits purified by HPLC (200 pg of each protein) were reduced and carboxymethylated under denaturing conditions. The alkylated samples were hydrolysed with CNBr and deglycosylated by in- cubation with PNGase F. The resulting peptide mixtures were subdigested with endoprotease Clu-C and aliquots of the sam- ples were directly analysed by FABMS, before and after methyl esterification of the free carboxyl groups 1141.
Fig. 4 shows the partial FAB mass spectrum of the peptide mixture generated from the LH a chain digestion. The signals were assigned to the corresponding peptides within the protein sequence on the basis of their molecular mass. The mass spectral analysis led to the verification of most of the a chain primary structure and to the definition of the first N-glycosylation site. The signal at m l z 1018, in fact, occurred 1 Da higher than the
Amoresano et al. (Euc J . Biochem. 242) 61 1
Table 1. Electrospray mass spectral analyses of intact a chains from human recombinant FSH, LH and CG. The different glycoforms are indicated with a capital letter according to the spectra shown in Fig. 1 . In the structures columns, D = diantennary structure, T = triantennary structure and Sia = sialic acid.
FS H LH CG
form molecular mass structures form molecular mass structures form molecular mass structures
D E F C G A H B I J K
Da
13569.42 1.0 13 732.6 2 0.9 13810.2-CO.9 14024.8 t 0.2 14 101.4 i- 1.0 14316.6 2 0.1 14 392.5 -C 0.7 14607.9 -C 0.6 14683.62 0.6 14972.0 -C 0.7 15263.1 -C 1.1
2D + lSia(-lHex) A 2D + 1Sia B ID + 1T I 2D + 2Sia C 1D + 1T + ZSia D 2D + 3Sia E 1D + 1T + 2Sia F 2D + 4Sia G 1D + I T + 3Sia H ID + 1T + 4Sia ID + 1T + 5Sia
Da
13 730.4 5 0.5 14022.6 2 1.2 14099.8? 1.5 14314.3 ?0.8 14388.8 20.9 14605.5 '-+ 1.7 14678.3 -C 1.7 14 971.4 t 0.3 15 261.9 2 0.6
2D + 1Sia D 2D + 2Sia B ID + IT + 1Sia A 2D + 3Sia C 1D + IT + 2Sia E 2D + 4Sia F 1D + 1T + 3Sia 1D + 1T + 4Sia 1D + 1T + 5Sia
~~
Da
13438.6-C 1.2 2D 13730.920.3 2D + lSia 14022.3 t 0.4 2D + 2Sia 14313.7 t 0.3 2D + 3Sia 14390.8?1.0 I D + IT t 2Sia 14679,620.3 ID + IT + 3Sia
H G
A B C D E F G H I J
15541.71+ 0.60 15427.84+ 1.07 15280.80+ 2.04 15250.43+ 0.14 15137.27+ 0.93 14989.23+ 0.46 15795.78+ 0.21 15907.77+1.43 16084.795 0.54 16198.90+ 0.27
J ri
Fig. 2. Transformed electrospray mass spectrum of recombinant LH molecular mass of the various components are reported.
chain. The individual species are indicated by capital letters. The
expected value for the peptide 48-56 due to the conversion of Asn52 into Asp following incubation with PNGase F. The ab- sence of any signal at mlz 1017 showed that this residue was completely modified in the native protein [18]. Two regions of the a chain, namely the fragment 30-47 and the C-terminal peptide 78-92, were not observed in the spectra, possibly due to the well-known suppression phenomena occurring in the FAB analysis of peptide mixtures [19].
Similar results were obtained when the a subunits from FSH and CG were analysed following the above procedure, as shown
in Table 2. The complete characterisation of the three a chains was achieved by HPLC fractionation of the CNBdGlu-C double digest followed by ESMS analysis of the individual fractions. As an example, the ESMS analysis of fractions 5 and 8 from the HPLC of the LH a chain digest yielded values of 2547.4 2 0.2 Da and 2131.2 2 0.6 Da, respectively, correspond- ing to the peptides 72-92 and 30-47, leading to the complete mapping of the a chain sequence. Moreover, the analysis of the peptide 72-92 showed a 1-Da increase in the molecular mass (theoretical value 2546.61), thus demonstrating that also Asn78,

61 2 Amoresano et al. ( E m J . Biochem. 242)
a Chains
1 5 10 15 20
Ala-Pro-Asp-Val-Gln-Asp-Cys-Pro-Glu-Cys-Thr~Leu-Gln-Glu~Asn-Pro-Phe-Phe-Ser-Gln 35 40
Pro-Gly-Ala-Pro-Ile-LeuGln-Cys-Met-Gly-Cys-Cys-Phe-Ser-Arg-Ala-~r-Pro~Thr-~ro 50 55 60
Leu-Arg-Ser-Lys-Lys-Thr-Met-Leu-Val-Gln-Lys-Asn-Val-Thr-Ser-Glu-Ser-Thr-Cys~Cys 75 80
val-Ala-Lys-Ser~Tyr-Asn-Arg-Val-Thr-Val-Met-Gly-Gly-Phe-Lys-Val-Glu-Asn-His-Thr
Ala-Cys-His-Cys-Ser-Thr-Cys-Tyr-TyrHis-Lys-Ser
25 30
45
65 70
85 90
FSH p 1 5 10 1 5 20
Asn-Ser-Cys-Glu-Leu-Thr-Asn-Ile-Thr-Ile-Ala-Ile-Glu-Lys-Glu~Glu~Cys-Arg-Phe-Cys 25 30 35 40
Ile-Ser-Ile-Asn-Thr-Thr-Trp-Cys-Ala-Gly-Tyr-Cys-Tyr-Thr-Arg-Asp-Leu-Val-~r-Lys 45 50 55 60
Asp-Pro-Ala-Arg-Pro-Lys-Ile-Gln-Lys-Thr~~s-Thr-Phe-Lys-Glu-Leu-Val-~r-Glu-Thr 65 70 1 5 80
Val-Arg-Val-Pro-Gly-Cys-Ala-H~s-His-Ala-Asp-Ser-Leu-Tyr-Thr-Tyr-Pro-Val-Ala-Thr 85 90 95 100
Gln-Cys-His~Cys-Gly-Lys-~s-Asp-Ser-Asp-Ser-Thr-Asp-Cys-Thr-Val-Arg-Gly-Leu-Gly
Pro-Ser-Tyr-Cys-Ser-Phe-Gly-Glu-Met-Lys-Glu 105 110
LH P 1 5 10 15 20
Ser-Arg-Glu-Prc-Leu-Arg-Pro-Trp-CysHis-Pro-Ile-Asn-Ala~Ile-Leu-Ala-Val-Glu-Lys 25 30 35 do
Glu-Gly-Cys-Pro-Val-Cys-Ile-Thr-Val-Asn-Thr-Thr~Ile-Cys-Asn-Gly-~r-~s-Pro-Thr 45 50 55 60
Met~Met-Arg-Vol-Leu-Gln-Ala-Val-Lcu-Pro-Pro-Leu-Pro~Leu~Val-ValCys-Thr-Thr-Tyr-Arg 65 70 75 80
Asp-Val-Arg-Phe-Glu-Se~-Ile-Arg-Lcu-Pro-Gly-cys-Pro-Arg~Gly~Val~Asp-pro-Val-Val 85 Y O 95 100
Ser-Phe-Pro-Val-Ala-Leu-Ser-Cys~Arg-Cys-Gly-Pro-Cys-~g-Arg-Ser-Thr-Ser-Asp-Cys 1935 110 115 120
Gly-Gly-Pro~Lys~Asp-His-Pro-Leu-Thr-Cys-Asp-His-Pro-Gln-Leu-Ser-Gly-Leu-Leu-Phe
Leu
CG P 1 5 10 15 20
25 30 35 40
45 50 55 60
65 70 75 80
85 911 95 100
Ser-Lys-Glu-Pro-Leu-Arg-Pro-Arg-Cys-Arg-Pro-Ile-Asn-Ala-Thr-Leu-Ala-Val-Glu-Lys
Glu-Gly-~s-Pro-Val-Cys-Ile-Thr-Val-Asn-Thr-Thr-Ile-Cys-Ala-Gly-Tyr-~s-Pro-Thr
Met-Thr -Arg-Va l -Leu-Gln -Gly -Va l -Leu-p ro -Ala -Leu-Pro -Gln -Va l -Va l -Cys -Asn-Tyr -Arg
Asp-Val-Arg Phe-Glu~Ser-Jle~Arg-Leu~Pro-Gly-Cys-Pro-Arg-Gly-Val-Arg-Pro-Val-Val
Ser-Tyr~Ala-Val~Ala-Leu-Ser-Cys-Gln-Cy~-Ala-Leu-Cys-Arg-Arg-Ser-Thr-Thr-As~-~~ 105 110 115 1 ?n
Gly-Gly-Pro LYs-AsP-H~s-Pro-Leu~Thr-Cys~Asp-Asp-Pro-A~g-Phe-Gln-Asp-Ser-Ser-Ser
Ser~Lys-Ala~Pro-Pro~Pro-SerLeuPro-Sfr-Pro-Ser-PIo-Ser-Arg-Leu-Pro-Gly-Pro-Ser-As~-T~~r
Pro-Ile-Leu-Pro-Gln
12s 170 135 140
145
Fig. 3. Amino acid sequences of a and p chains from human recombinant glyco-hormones.
Table 2. Electrospray and FAB mass data obtained from the analysis of CNBr/Glu-C double digest of the reduced and carboxymethylated a chains. CmCys, carboxymethylcysteine.
Mass Peptide Notes
Da
2547.4 i 0.2 72 - 92 CmCys82,84,87 ; Asp78 2308 10-29 Ser29 ; CmCys10,28 2131.2 +0.6 30-47 Ser47; CmCys31,32 1746 57-71 Ser71 ; CmCys59,60 1018 48-56 Asp52
1030 1- 9 CmCys7 635 72-77
the second putative N-glycosylation site, has been changed into Asp by the PNGase Faction and was then effectively glycosylated.
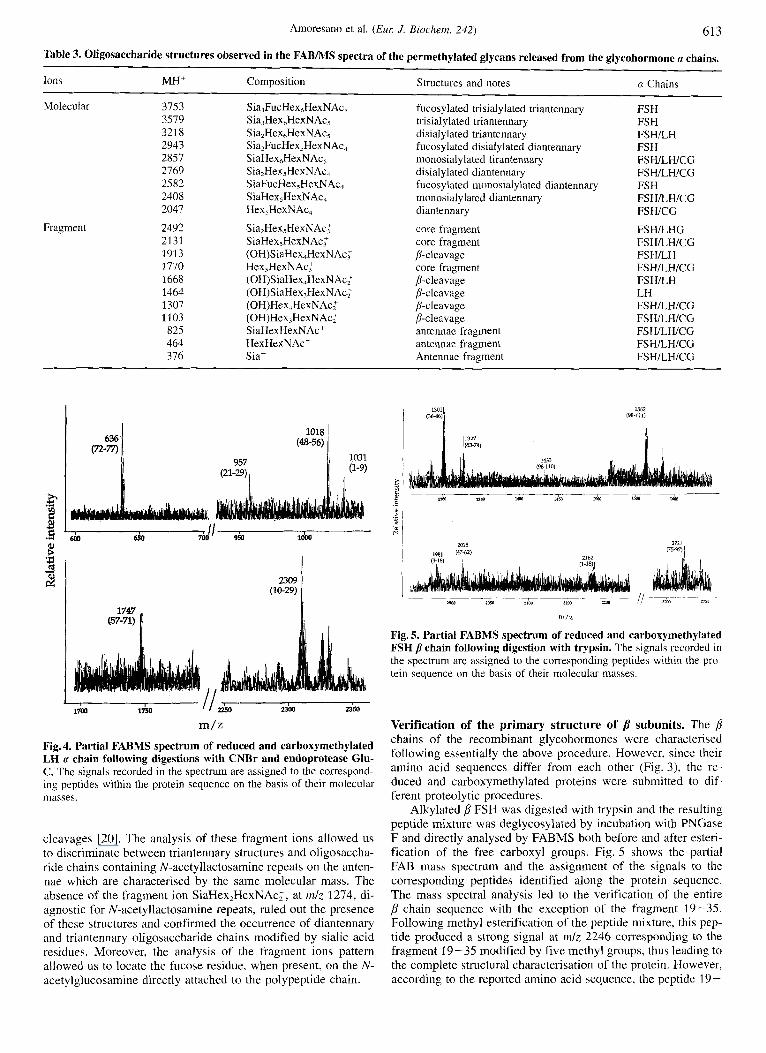
Characterisation of the oligosaccharides a chains. The struc- ture of the glycosidic moieties of the three u chains was investi- gated by direct FABMS analysis of the permethylated oligosac-
charide mixtures. The N-glycans were released from the respec- tive polypeptide chain by tryptic hydrolysis of the cx subunits followed by incubation with PNGase F. The oligosaccharide mixtures were separated from peptides by a Sep-pak reverse- phase chromatographic step, permethylated and submitted to spectral analysis. Table 3 shows the signals recorded in the spectra and the corresponding oligosaccharide structures iden- tified in each a subunit.
As shown in Table 3, the glycans N-linked to the (x chains belong to the complex type and consist essentially of mono- and di-sialylated diantennary and triantennary structures. However, differences exist in the relative amount of these components within the three a chains. Triantennary oligosaccharides consti- tute only minor components in a FSH and a CG whereas they are as abundant as the diantennary structures in the n LH. Fuco- sylated diantennary and triantennary structures with different de- gree of sialylation were identified as minor species in cx FSH whilst they were not detected i n the n chains from LH and CG.
The identification of the different glycans structures was confirmed by their characteristic fragmentation pattern in the FAB spectra mainly due to A-type cleavages occurring at the non-reducing end of N-acetylglucosamine residues and to ring
Amoresano et al. ( E m J. Biochem. 242) 61 3
Table 3. Oligosaccharide structures observed in the FABMS spectra of the permethylated glycans released from the glycohormone a chains.
Ions MH' Composition Structures and notes
Molecular 3753 Sia,FucHex,HexNAc, fucosylated trisialylated triantennary FSH 3579 Sia,HexhHexNAc, trisialylated triantennary FSH 321 8 Sia,HexhHexNAc, disialylated triantennary FSHLH 2943 Sia,FucHex,HexNAc, fucosylated disialylated diantennary FSH 2857 SiaHex,HexNAc, monosialylated tirantennary FSHILHICG 2769 Sia,Hex5HexNAc, disialylated diantennary FSHILHICG 2582 SiaFucHex,HexNAc, fucosylated inonosialylated diantennary FSH
FSWLHICG 2408 SiaHex,HexNAc, monosialylated diantennary 2047 Hex,HexNAc, diantennary FSWCG
a Chains
Fragment 2492 2131 1913 1770 1668 1464 1307 1103 825 464 376
Sia,Hex5HexNAc.F SiaHex,HexNAc; (0H)S i aHex4Hex N Ac: Hex,HexNAc: (OH)SiaHex,HexN Ac; (OH)SiaHex,HexNAc; (OH)Hex,HexNAc: (OH)Hex,HexNAc: SiaHexHexNAc' HexHexNAc' Sia'
core fragment core fragment p-cleavage core fragment ,&cleavage P-cleavage P-cleavage fi-cleavage antennae fragment antennae fragment Antennae fragment
FSWLHG FSHILHICG FSHILH FSHILHICG FSWLH LH FSHILHICG FSHILHICG FSHILWCG FSHILHICG FSHILHICG
636 (T2-77)
957
(21-29) I
- I /& 1700 1750 ww 2350
m/z Fig. 4. Partial FABMS spectrum of reduced and carhoxymethylated LH a chain following digestions with CNBr and endoprotease Glu- C. The signals recorded in the spectrum are assigned to the correspond- ing peptides within the protein sequence on the basis of their molecular masses.
cleavages [20]. The analysis of these fragment ions allowed us to discriminate between triantennary structures and oligosaccha- ride chains containing N-acetyllactosamine repeats on the anten- nae which are characterised by the same molecular mass. The absence of the fragment ion SiaHex,HexNAc:, at m/z 1274, di- agnostic for N-acetyllactosamine repeats, ruled out the presence of these structures and confirmed the occurrence of diantennary and triantennary oligosaccharide chains modified by sialic acid residues. Moreover, the analysis of the fragment ions pattern allowed us to locate the fucose residue, when present, on the N- acetylglucosamine directly attached to the polypeptide chain.
I // -~~ ~
2710 , ________
2 a a zim 2x50 i lw I
m/z
Fig. 5. Partial FABMS spectrum of reduced and carboxymethylated FSH /l chain following digestion with trypsin. The signals recorded in the spectrum are assigned to the corresponding peptides within the pro- tein sequence on the basis of their molecular masses.
Verification of the primary structure of p subunits. The p chains of the recombinant glycohormones were characterised following essentially the above procedure. However, since their amino acid sequences differ from each other (Fig. 3), the re- duced and carboxymethylated proteins were submitted to dif- ferent proteolytic procedures.
Alkylated p FSH was digested with trypsin and the resulting peptide mixture was deglycosylated by incubation with PNGase F and directly analysed by FABMS both before and after esteri- fication of the free carboxyl groups. Fig. 5 shows the partial FAB mass spectrum and the assignment of the signals to the corresponding peptides identified along the protein sequence. The mass spectral analysis led to the verification of the entire p chain sequence with the exception of the fragment 19-35. Following methyl esterification of the peptide mixture, this pep- tide produced a strong signal at mlz 2246 corresponding to the fragment 19-35 modified by five methyl groups, thuv leading to the complete structural characterisation of the protein. However, according to the reported amino acid sequence, the peptide 19-

61 4 Amoresano et al. ( E m J. Biochem. 242)
Table 4. FABMS analysis of the reduced and carboxymethylated LH /I chain digested with trypsin and endoprotease Asp-N before and after methylesterification of the free carboxylic groups. CmCys, carboxymethylcy steine.
MH’ Peptide Esterification Notes
Da Da
2386 1953
1603 1239 1126
979 743 700 65 1
1- 20 2442 1 CmCys, 4 CH, 44- 60 1981 1 CmCys, 2 CH, 30- 43 1734 1 CmCys, 4 CH, 75- 89 1645 1 CmCys, 3 CH,
111-121 1267 2 CH, 111-120 1154 2 CH, 21- 29 1092 2 CmCys, 4 CH,
111-119 1007 2 CH, 105-110 785 1 CmCyr, 3 CH, 69- 74 728 2 CH, 64- 68 679 2 CH,
35 contains only four acidic residues, including the C-terminus. The fifth carboxyl group was originated by the conversion of Am30 into Asp following PNGase F incubation, thus demon- strating that this putative N-glycosylation site was in fact fully modified.
The FAB analysis revealed the presence of an N-terminal truncated form of the /{ chain ; the signals at mlz 21 82 and 1981, in fact, were associated with the N-terminal peptides 1-18 and 3-18 respectively, the latter originated by the loss of the N- terminal dipeptide Asn-Ser. As estimated by peak intensities [21], the two protein forms are present in similar amounts. Moreover, both mass signals occurred 1 Da higher than expected on the basis of the /I’ FSH sequence, thus demonstrating that the second putative N-glycosylation site at Am13 was also fully gl ycosylated.
A sample of reduced and carboxymethylated /j chain from LH (100 pg) was digested with trypsin, deglycosylated by PNGase F treatment and digested with endoprotease Asp-N. The resulting peptide mixture was analysed by FABMS both before and after methyl esterification producing the results reported in Table 4. Most of the protein sequence could be verified by this single experiment, leaving only the regions 61 -63 and 90-104 undetected. In order to control the remaining portion of the p subunit, a second aliquot of the alkylated protein (100 pg) was digested with Asp-N, incubated with PNGase F and submitted to mass spectral analysis. The signals at mlz 2510, 1859 and 634 were assigned to the peptides 77-98, 61 -76 and 99-104 lead- ing to the complete structural characterisation of the ,8 LH.
The N-terminal region of the protein was shown to corre- spond with the expected amino acid sequence; no evidence of proteolytic processing was observed. Conversely, microhetero- geneity of the polypeptide chain was detected at the C-terminus. Mass signals at mlz 979, 1126 and 1239 were detected and as- signed to the fragments 111-119, 111-120 and 111-121 re- spectively, indicating the occurrence of C-terminal ragged ends. These results confirm previous ESMS data on the intact protein showing that the /I LH contains at least three different isoforms differing in the length of the polypeptide chain.
The presence of a fully modified N-glycosylafion site at level of As1130 was demonstrated by the signal at mlz 1734 ob- served in the tryptic1Asp-N double digest following methyl es- terification. This value corresponds to the peptide 30-43 carry- ing four methyl ester groups, one more than those predicted on the basis of the /j chain sequence, thus showing that Am30 had been converted into Asp by PNGase F incubation and hence methyl esterified ( A m = +I5 Da).
Table 5. FABMS analysis of the reduced and carboxymethylated CG /I chain digested with trypsin and endoprotease Asp-N before and after methylesterification of the free carboxylic groups. CmCys, carbox y meth y Icysteine.
Peptide Esterification Notcs MH’
Dd Dd
1926 44- 60 1954 1 CmCys,2 CH, 1648 30- 43 2 CmCys, Asp30 141 1 83- 94 1 CmCy5 1373 9- 20 1415, 1429 1 CmCys, 314 CH,,
Asn/Aspl? A5pl3 1332 64- 74 1374 1 CmCy5, 3 CH,
105-114 1296 1 CmCy5, 5 CH, 1079 95-104 1 CmCys
21- 29 1092 2 CmCys, 4 CH, 982 1- x 1010 2 CH, 834 75- 82 848 1 CH, 743 105-110 7x5 1 CMCys, 3 CH, 700 69- 74 728 2 CH, 65 1 64- 68 67 9 2 CH, 389 61- 63
The heterogeneity of the p LH preparation was further inves- tigated by tryptic digestion of the denatured and alkylated pro- tein followed by HPLC fractionation and mass spectral analysis of the individual fractions. In particular, two fractions yielded signals at mlz 2461 and 1524 which were assigned to the pep- tides 96- 11 7 and 10.5 - 117 respectively. The presence of these fragments demonstrated the occurrence of a further p chain iso- form containing an even shorter polypeptide chain ending at Gly117. This isoform is present as a very minor species judging by the low ultraviolet absorbance of the two fragments as com- pared to the C-terminal peptides derived from the other b chain components. Therefore this isoform was observed neither in the ESMS analysis of the intact protein nor in the FAB mapping of proteolytic digests.
The verification of the primary structure of /I C G was ac- complished by using a slightly different hydrolysis strategy, since this protein is known to contain 0-glycosylation sites in the C-terminal region [22]. The reduced and carboxymethylated p chain was digested with trypsin and N-deglycosylated by PNGase F treatment. The resulting peptides mixture was subdi- gested with Asp-N and analysed by FABMS leading to the veri- fication of about 80% of the entire protein sequence, as shown in Table 5. As expected, the C-terminal portion of the protein escaped the mass spectral analysis. Therefore, an aliquot of the p subunit was submitted to a consecutive sialidase and O-gly- cosidase treatment and then digested with trypsin. The following FAB analysis showed the presence of signals at rnlz 1234 and 1105 corresponding to the C-terminal peptides 134-145 and 123-134. The complete sequence of p CG was then verified with the exception of the fragment 115-122. The FAB mass spectrum of the peptide mixture following methyl esterification showed two signals at mlz 1415 and 1429 corresponding to the fragment 9-20 modified by three and four methyl groups, re- spectively. The fourth carboxyl group originated from the con- version of Asnl3 into Asp following incubation with PNGase F, thus demonstrating that this N-glycosylation site was only par- tially glycosylated. The spectrum also showed the presence of a signal at mlz 1704 corresponding to the fragment 30-43 carry- ing four methyl groups, thus demonstrating that the second N- glycosylation site at Am30 was instead fully glycosylated.
Characterisation of the oligosaccharides from chains. The structural investigation of the glycans N-linked to the chains

Amoresano et al. (Euc J. Biochem. 242) 615
Table 6. Oligosaccharide structures observed in the FAB/MS spectra of the permethylated glycans released from the glycohormone /l chains.
Ions MH + Composition Structures and notes B Chains
Da Molecular 4202
4028 3753 3579 3392 3218 2943 2857 2769 2582
2221
Fragment 3751 2941 2492 2131 1913 1770 1668 2464 1307 1274 1103 913 825 464 376
2408
Sia,FucHex,HexNAc, Sia?Hex,HexNAc, Sia,FucHex,HexNAc, Sia,Hex,HexNAc, SiaZFucHex,HexNAc, Sia,Hex,HexNAc, Sia,FucHex,HexN Ac, SiaHex,HexNAc, Sia,Hex,HexNAc, SiaFucHex,HexNAc, SiaHex,HexNAc, FucHex,HexNAc,
Sia,Hex,HexNAc' Sia,Hex,HexNAch Sia,Hex,HexNAc: SiaHex,HexNAc: (OH)SiaHex,HexNAc: Hex,HexNAc: (OH)SiaHex,HexNAc; (OH)SiaHex,HexNAc; (OH)Hex,HexNAc: SiaHex,HexNAc; (OH)Hex,HexNAc' Hex,HexNAc: SiaHexHexNAc' HexHexNAc' Sia'
fucosylated trisialylated triantennary + N-acetyllactosamine trisialylated triantennary + N-acetyllactosamine repeats fucosylated trisialylated triantennary trisialylated triantennary fucosylated disialylated triantennary disialylated triantennary fucosylated disialylated diantennary monosialylated triantennary disialylated diantennary fucosylated monosialylated diantennary monosialylated diantennary fucosylated diantennary
core fragment core fragment core fragment core fragment ,&cleavage core fragment P-cleavage D-cleavage P-cleavage antennae fragment P-cleavage antennae fragment antennae fragment antennae fragment antennae fragment
repeats FSH FSH FSH/LH FSH FSHLH FSWCG FSWLWCG FSH FSH/CG FSHLWCG FSWCG FS WLH/CG
FSH FSH FSHLWCG FSH/LH/CG FSHLH FSHLWCG FSHLWCG FSH/LH/CG FSWLH/CG FSH FSWLWCG FSH FSWLWCG FSWLWCG FSWLWCG
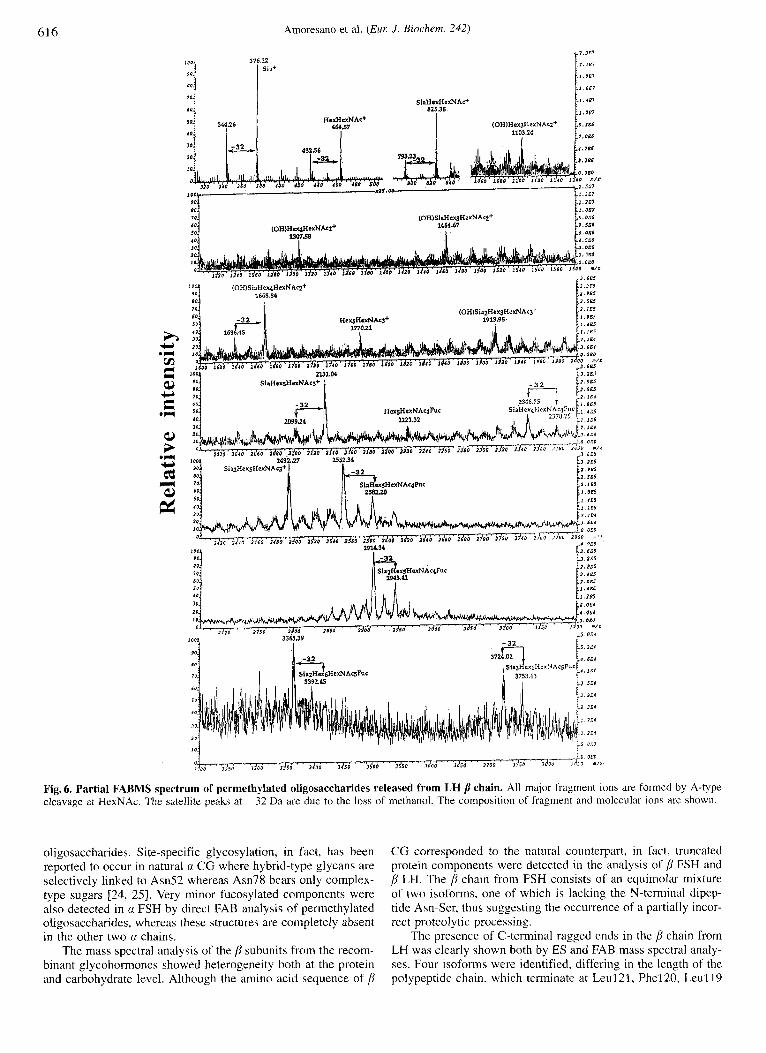
of the recombinant glycohormones was accomplished following the procedure described above for the a subunits. The oligosac- charides were released from the alkylated polypeptide chains following tryptic hydrolysis and PNGase F treatment and sepa- rated from the mixture of peptides by Sep-pak chromatography; the sugars were then permethylated and analysed by FABMS. The mass spectral analysis showed that the glycans N-linked to the j3 chains belong to the complex type consisting essentially of fucosylated and/or sialylated diantennary and triantennary structures, as shown in Table 6.
The FAB spectrum of the p LH oligosaccharides is shown in Fig. 6; the analysis confirmed previous results obtained from the ESMS analysis of the intact protein in that the most abundant components consist of fucosylated mono- and di-sialylated dian- tennary structures, whereas fucosylated and sialylated trianten- nary glycans are present as very minor species.
The p subunit of FSH showed a higher degree of hetero- geneity as compared to the other p chains, possibly due to the presence of two glycosylated Asn residues in this protein. The FAB spectra, in fact, showed the occurrence of fucosylated and/ or sialylated diantennary and triantennary structures in the ratio 3 : 1. Moreover, the signals at m/z 4202 and 4028 might be inter- preted as arising from trisialylated tetraantennary oligosaccha- ride chains carrying one or no fucose residues or, alternatively, as fucosylated trisialylated triantennary structures containing a single N-acetyllactosamine repeat on the antennae. The analysis of the FAB spectrum in the low-mass region showed the pres- ence of strong fragment ions at mlz 1274 and 913, attributed to the fragment ions SiaHex,HexNAcl and Hex,HexNAc: respec- tively, which are diagnostic for N-acetyllactosamine repeats ; this finding suggests the occurrence of triantennary structures con- taining N-acetyllactosamine repeats, although the presence of both structures cannot be ruled out.
In order to characterise the 0-linked oligosaccharides pre- sent in the p chain from CG, the tryptic peptides and 0- glycopeptides eluted from the Sep-pak chromatography were submitted to reductive elimination [13], the free sugars were permethylated and analysed by FABMS. The mass spectra showed the presence of mono- and di-sialylated HexHexNAc structures via signals at mlz 1243 and 873, respectively.
DISCUSSION
The a and p subunits of the human recombinant glycohor- mones FSH, LH and CG produced in CHO cells have been structurally characterised both at the protein and carbohydrate level by using mass spectrometric methodologies.
The amino acid sequence of the a chains was verified both by direct ESMS analysis of the intact proteins and by the FAB mapping procedure [17]. The primary structure of the recombi- nant proteins was shown to be identical to the natural counter- part 1231. Moreover, the presence of N-glycosylation sites at Am52 and Am78 was confirmed, both sites being fully glyco- sylated. Although the a chains from the three glycohormones share the same polypeptide sequence, the investigation of their oligosaccharide content revealed some differences. Sialylated di- antennary structures, in fact, were the major glycans occurring in a FSH and a CG with only 15% and 5%, respectively, of the glycoforms carrying one triantennary structure, as estimated by the ESMS spectra. In contrast, a LH showed an equal amount of components carrying two diantennary or one diantennary and one triantennary structures on the two N-glycosylation sites. These findings might suggest a site-specific glycosylation pro- cess in which triantennary structures are linked to a single spe- cific Asn with the other Asn residue always carrying diantennary
61 6 Amoresano et al. (Eul: J . Biochem. 242)
i . IS7 J.16)
I I 1103.24 7.066
SiaHexHexNAc+ 825.38
1,-32 e::::
/ p n N A @ 294S.41
Fig. 6. Partial FABMS spectrum of permethylated oligosaccharides released from LH p chain. All major fragment ions are formed by A-type cleavage at HexNAc. The satellite peaks at -32 Da are due to the lobs of methanol. The composition of fragment and molecular ionc are shown.
oligosaccharides. Site-specific glycosylation, in fact, has been reported to occur in natural a C G where hybrid-type glycans are selectively linked to Asn52 whereas Asn78 bears only complex- type sugars 124, 2.51. Very minor fucosylated components were also detected in a FSH by direct FAB analysis of permethylated oligosaccharides, whereas these structures are completely absent in the other two a chains.
The mass spectral analysis of the /Y subunits from the recom- binant glycohormones showed heterogeneity both at the protein and carbohydrate level. Although the amino acid sequence of p
C G corresponded to the natural counterpart, in fact, truncated protein components were detected in the analysis of [I FSH and @ LH. The p chain from FSH consists of an equimolar mixture of two isoforms, one of which is lacking the N-terminal dipep- tide Asn-Ser, thus suggesting the occurrence of a partially incor- rect proteolytic processing.
The presence of C-terminal ragged ends in the p chain from LH was clearly shown both by ES and FAB mass spectral analy- ses. Four isoforms were identified, differing in the length of the polypeptide chain, which terminate at Leul21, Phel20, Leu1 19

Amoresano et al. (Eur. J . Biochem. 242) 617
and Glyl17, respectively. The first three proteins are present in almost equal amount whereas the shortest isoform constitutes a very minor component.
Although a and p chains share a high degree of similarity, all the p subunits exhibited essentially fucosylated oligosaccharide structures which were almost absent in the a chains, thus sug- gesting that glycosylation is influenced by the small differences in the protein structures. The p chain from FSH possesses two glycosylated Asn residues in its sequence and shows the occur- rence of fucosylated diantennary and triantennary structures with a molar ratio of about 3: 1, suggesting a situation similar to that observed for the a LH. Again, a site-specific glycosylation pro- cess might have occurred, leading to the presence of triantennary structures always linked to the same Asn residue. It should be noted that /l FSH is the only subunit where the occurrence of N- acetyllactosamine repeats have been detected. The p subunits from LH and CG showed essentially the presence of fucosylated mono- and di-sialylated diantennary structures, being trianten- nary glycans present in a very low amount.
These data are in very good agreement with previous reports on the N-linked glycans from natural CG. The a chain oligosac- charides, in fact, belong to the complex and hybrid types with a very minor amount of fucosylated species 124, 26, 271. The p chain bears only diantennary complex glycans with a large amount of fucosylated structures [26]. The major components in recombinant hCG are identical to their natural counterpart al- though small differences exist since hybrid glycans are absent in the a chain and minor triantennary structure were detected in the chain.
These differences in the oligosaccharide structures within the a and p subunits, which also occur in the natural gonadotropins, confirm that the glycosylation pattern of the various glycohor- mones is dictated by the hormone-specific p chains [lo]. Most likely, the early stages of the glycosylation process take place independently on a and p chains until the GlcNAc,Man,Glc- NAc, key intermediate has been synthesised. This intermediate can then evolve to yield both sialylated and sulphated complex- type structures [28]. Therefore, once the heterodimeric glycohor- mones have been assembled, the final events of the glycosyla- tion process depend on the structure and glycosylation of the individual p subunits as well as on the different cell types where the glycohormones are produced. Pituitary LH and free a sub- units, in fact, bear oligosaccharides terminating in sulphated branches whereas pituitary FSH shows exclusively sialylated oligosaccharides ; placental CG and free a subunits terminate in sialic acid residues. The synthesis of sulphated structures results from the consecutive action of a GalNAc transferase and a sulfo- transferase which catalyses the addition of a sulphate group. The key enzyme GalNAc transferase has been found in the pituitary cells whereas it is absent in the placental cells [29]. This ex- plains the presence of different terminal groups in LH and CG, whereas the absence of sulphate groups in the FSH molecule has to be related to the regulation of the glycosylation process exerted by the three-dimensional structure of the hormone.
Since GalNAc transferase is not expressed in CHO cells, the oligosaccharide structures of the recombinant glycohormones produced in these host cells terminate exclusively in sialic acid residues. However, it has been shown that sulphation of natural human LH glycans seems not to be involved in hormone-recep- tor interactions but in regulating the rate of LH clearance from the circulation [30, 31 1. Therefore, the absence of sulphated structures in the recombinant glycohormones should not affect their biological activities.
The ESMS analysis of the a chains from recombinant glyco- hormones showed the entire pattern of glycoforms present in each protein and allowed us to quantitate their relative abun-
dance, providing a global view of the effective nature of the recombinant product. As an example, the presence of glyco- forms carrying two triantennary structures could be immediately ruled out on the basis of the molecular masses observed. This analysis constitutes an effective fingerprint of both the recombi- nant products and the entire fermentation/expression process. Any alteration in the global process of recombinant protein pro- duction which reflects in changes in the relative abundance of the various glycoforms can, in fact, be detected and identified by this methodology. However, it should be remembered that p FSH and p CG could not be analysed by ESMS despite their structural similarity with p LH and the a chains. This failure seems very likely to be due to peculiar physico-chemical proper- ties of these two glycoproteins as a whole and not simply to the glycosylation itself, as demonstrated by the successful analysis of the heavily glycosilated a subunits.
Direct analysis of permethylated oligosaccharides by FABMS essentially confirmed the results obtained by ESMS of the intact glycoproteins. The same glycan structures and the same relative abundance of the various oligosaccharide chains were detected by direct FABMS analysis of the mixture of permethylated oligosaccharides. However the key role of FABMS in carbohydrate analysis should be underlined in that glycan structures present at a very low level which escape ESMS analysis could be detected. Moreover the presence of N-acetyl- lactosamine repeats on the antennae of FSH triantennary struc- tures could be identified via diagnostic fragment ions occurring in the low-mass range of FAB spectra, thus discriminating these structures from tetraantennary oligosaccharide chains which show the same molecular mass.
Finally, the application of mass spectrometric methodologies in structural glycobiology has to be emphasised. Recently, new NMR-based strategies have been developed to allow simple structural assesment of intact glycoproteins [25, 271. However, the combined mass spectrometric procedure described above is particularly suitable for the rapid definition of the entire pool of glycoforms, including the minor components, and to determine the monomer sequence and the branching pattern through appro- priate fragment ions. This is especially true in the characterisa- tion of recombinant glycoproteins where a rapid and effective comparison with the natural counterpart is required.
REFERENCES 1. Pierce, J. G. & Parson, T. E (1981) Glycoprotein hormones: struc-
ture and function, Annu. Rev. Biochem. 50, 465-495. 2. Stockell Hartree, A. & Renwick, A. G. C. (1992) Molecular struc-
tures of glycoprotein hormones and functions of their carbohy- drate components, Biochem. J . 287, 665-679.
3. Childs, G. V., Ellison, D. G. & Unabia, G. (1990) in Glycoprotein hormones (Chin, W. W. & Boime, I., eds) pp. 1 - 10, Serono Sym- posia, Norwell MA.
4. Cornell, J. S. & Pierce, J. G. (1974) Studies on the disulphide bonds of glycoprotein hormones, J . Biol. Chern. 249, 4266-4174.
5. Bedows, E., Huth, J. R., Suganuma, N., Bartels, C., Boime, I. & Ruddon, R. W. (1993) Disulphide bond mutations affect the fold- ing of the human chorionic gonadotrophin /l subunit in transfected Chinese hamster ovary cells, J. Biol. Chern. 268, 11 655 - 11 662.
6. Lapthorn, A. J., Harris, D. C., Littlejohn, A., Lustbader, J. W., Can- field, R. E., Machin, K. J., Morgan, F. J. & Isaacs, N. W. (1994) Crystal structure of human chorionic gonadotropin, Nature 369,
7. Thotakura, K. & Blithe, D. (1995) Glycoprotein hormones: glyco- biology of gonadotrophins, thyrotrophin and free n subunit, Glycobiology 5, 3-10.
8. Bishop, L. A,, Robertson, D. M., Cahir, N. & Schofield, P. R. (1994) Specific roles for the asparagines-linked carbohydrate residues of
455-461.

61 8 Amoresano et al. (Eur J. Biochem. 242)
recombinant huinan follicle stimulating hormone in receptor bind- ing and signal trasduction, Mol. Endocrinol. 8, 722-731.
9. Zhang, R., Tsai-Morris, C. H., Kitamura, M., Buczko, E. & Dufau, M. L. (1991) Changes in binding activity of lutenizing hormone receptors by site directed mutagenesis of potential glycosylation sites, Biochem. Biophys. Kes. Commun. 181, 804-808.
10. Baenziger, J. U. & Green, E. D. (1988) Pituitary glycoprotein hor- mone oligosaccharides: structure, synthesis and function of the asparagine-linked oligosaccharides on lutropin, follitropin and thyrotropin, Biochim. Biophys. Actu 947, 287- 306.
11. Cummings, D. A. (1991) Glycosylation of recombinaiit protein ther- apeutics: control and functional implication, Glycohiology I ,
12. Nitti, G., Orru, S., Bloch, C., Mohry, L., Marino, G. & Pucci, P. (1995) Amino acid sequence and disulphide bridge pattern of three y-thionins from Sorghum bicolor, EUL J . Biochem. 228,
13. Carlson, D. M. (1968) Structures and immunochemical properties of oligosaccharides isolated from pig submaxillary mucin, J. Biol. Chem. 243, 616-626.
14. Hunt, E. & Morris, H. R. (1973) A mass-spectrometric and 'H- and "C-nuclear magnetic study, Biochem. .I. 135, 833 -843.
15. Dell, A. (1990) Preparation and desorption mass spectrometry of permethyl and peracetyl derivatives of oligosaccharides, Methods Enzimol. 193, 647-660.
16. Pucci, P. & Sepe, C. (1988) Computer program for post-translational modification site assignment in proteins using fast atom bombard- ment mass spectral data, Biomed. Environ. Muss Spectrom. 17,
17. Moms, H. R., Panico, M. & Taylor, G. W. (1983) FAB-mapping of recombinant-DNA protein products, Biochem. Biophys. Res. Commun. 117, 299-306.
18. Carr, S. A. & Roberts, G. L. (1986) Carbohydrate mapping by mass spectrometry: a novel method for identifying attachment sites of Asn-linked sugars in glycoproteins, Anal. Biochem. 157, 396- 402.
19. Naylor, S., Findeis, A,, Gibson, G. W. & Williams, D. H. (1986) An approach toward the complete FAB analysis of enzymic digests of peptides and proteins, J. Am. Chem. Soc. 108, 6359-6363.
20. Dell, A. (1987) FAB inass spectrometry of carbohydrates, Adv. Cur- hohydr: Chem. Hiochem. 45, 19-72.
115-130.
250-256.
287-291.
21. Debiase, D., Barra, D., Bossa, F., Pucci, P. & John, R. A. (1991) Chemistry of the inactivation of 4-aminobutyrate aminotransfer- ase by the antiepilectic drug vigabatrin, J . B i d . Chem. 266,
22. Sairam, M. R. (1983) in Hormonal protein and peptides (Li, C. H., ed.) vol. 11, pp. 1-79, Academic Press, New York.
23. Norman, A. W. & Litwack, G. (1987) Hormones, pp. 171-220. Academic Press, Orlando FL.
24. Blithe, D. L. (1990) Carbohydrate composition of the n-subunit of human choriogonadotropin (hCGn) and the free n molecules pro- duced in pregnancy: most free u and some combined hCG-a mol- ecules are fucosylated, Endocrinology 126, 2788-2799.
25. Van Zuylen, C. W. E. M., de Beer, T., Rademaker, G. J., Haverkamp, J.. Thomas-Oates, J. E., Hard, K., Kamerling, J. P. & Vliegenthart, J. F. G . (1995) Site-specific and complete enzymic deglycosyla- tion of the native huinan chorionic gonadotropin n-subunit, Eul: J . Biochem. 231, 754-760.
26. Mizuochi, T. & Kobata, A. (1980) Different asparagiiie-linked sugar chains on the two polypeptide chains of human chorionic gonado- tropin, Biochem. Biophys. Res. Commun. 97, 772-778.
27. de Beer, T., Van Zuylen, C. W. E. M., Hard, K., Boelens, R., Kap- tein, R., Kamerling, J. P. & Vliegenthart, J. F. G. (1994) Rapid and simple approach for the NMR resonance assignment of the carbohydrate chains of an intact glycoprotein. Application of gra- dient-enhanced natural abundance 'H-"C HSQC and HSQC- TOCSY to the cx-subunit of human chorionic gonadotropin, FEBS Lett. 348, 1-6.
28. Green, E. D.. Boime, I. & Baenziger, J. U. (1986) Biosynthesis of sulphated asparagine-linked oligosaccharides on bovine lutropin, J. B i d . Chem. 261, 16309-16316.
29. Nemansky, M., Schiphorst, W. E., Kolleman, C. A. & Van den Eijn- den, D. H. (1992) Human liver and human placenta both contain CMP-NeuAc: Gal PI-4GlcNAc-R n2-3- as well as u2-6- siulyltransferase activity, FEBS Lett. 312, 31 -36.
30. Fiete, D., Srivastava, V., Hindsgaul, 0. & Baenziger, J . U. (1991) A hepatic reticuloendothelial cell receptor specific for SO,-4Gal- NAcPl, 4GlcNAcP1, 2Mana that mediates rapid clearance of lut- ropin, Cell 67, 1103-1110.
31. Baenziger, J. U., Kumar, S., Brodbeck, R. M., Smith, P. L. & Bera- nek, M. C. (1992) Circulatory half-life but not interaction with the lutropdchorionic gonadotropin receptor is modulated by sul- fation of bovine lutropin oligosaccharides, Proc. Nut1 Arad. Sci.
20056-20061,
USA 89, 334-338.
Related Documents











