Structural Basis for the High Affinity of Amino-Aromatic SH2 Phosphopeptide Ligands Joseph Rahuel 1 , Carlos Garcı ´a-Echeverrı ´a 2 , Pascal Furet 2 Andre ´ Strauss 1 , Giorgio Caravatti 2 , Heinz Fretz 2 , Joseph Schoepfer 2 and Brigitte Gay 2 * 1 Novartis Pharma AG, Core Technology Area and 2 Oncology, Research Department, CH-4002, Basel Switzerland An anthranyl moiety placed at the N terminus of a phosphotyrosine pep- tide potentiates the inhibitory effect of this small peptide on the binding of the Grb2 SH2 domain to the EGF receptor. Using molecular modeling procedures based on the Lck SH2 domain structure, this observation was rationalized in terms of a suitably favorable p-p stacking interaction between the anthranyl moiety and the arginine aA2 (ArgaA2) residue side-chain of Grb2 SH2. The crystal structure of the Grb2 SH2 domain in complex with the inhibitor 2-Abz-EpYINQ-NH2 (IC 50 26 nM) has been solved in two different crystal forms at 2.1 and 1.8 A ˚ resolution. This structure confirms the modeling based on the Lck SH2 domain. The ArgaA2 residue is conserved in most SH2 domains. Thus, as expected, the anthranyl group also confers high affinity to small peptide ligands of other SH2 domains such as Lck-, PLC-g-amino-terminal and p85 amino- terminal SH2 domains as demonstrated by structure affinity relationships (SAR) data. These potent peptides with an amino-terminal surrogate group and the structure of Grb2 SH2 domain in complex with one such peptide represent good starting points for the design and optimization of new inhibitors of many SH2 domains. # 1998 Academic Press Limited Keywords: signal transduction; Grb2; crystal structure; SH2 domain; anthranyl *Corresponding author Introduction Src homology-2 (SH2) domains are phosphotyro- sine-binding modules found in many proteins that are key players in several signal transduction path- ways (Mayer & Baltimore, 1993). SH2 domains have been shown to bind to phosphorylated pro- tein tyrosine kinases or other proteins phosphory- lated in response to extracellular signals. The central roles played by protein tyrosine kinases in mitogenic signaling cascades and the involvement of aberrant or overexpressed protein tyrosine kinases in several cancers and proliferative diseases have made the development of inhibitors that block the SH2 domain binding desirable as poten- tial therapeutic agents (Schlessinger & Bar-Sagi, 1994). SH2 domains are capable of binding phos- photyrosine and recognize the sequence context of the phosphotyrosine. Studies have shown that rela- tively short peptide sequences of less than five resi- dues bind tightly to SH2 domains (Songyang et al., 1993; Zvelebil et al., 1995). The structures of various SH2 domains, both free in solution and complexed with phosphotyrosine peptides, have been deter- mined (Waksman et al., 1992; Eck et al., 1993; Lee et al., 1994; Hatada et al., 1995; Nolte et al, 1996; Rahuel et al, 1996). These structural studies have shown that all SH2 domains consist of a central antiparallel b-sheet with two flanking a-helices, and a second smaller two-stranded b-sheet. The peptide recognition site straddles the exposed edge of the central b-sheet (bD strand). A conserved Abbreviations used: 2-Abz, 2-amino-benzoyl (anthranyl); EGF, epidermal growth factor; EGFR, EGF receptor; GST, glutathione-S-transferase; IRS1, insulin receptor substrate-1; MBP, maltose binding protein; PDGF, platelet derived growth factor; PDGFR, PDGF receptor; PLC-g, phospholipase C-g1; pY, phosphotyrosine, ligand residues are numbered relative to the position of the phosphotyrosine, which is denoted pY 0; r.m.s., root-mean-square; SH2, Src homology-2 domain; TBS, Tris-buffered saline; SAR, structure activity relationship. The nomenclature adopted by Eck et al. (1993) for SH2 domains is used here and explained in Figure 4. Article No. mb981790 J. Mol. Biol. (1998) 279, 1013–1022 0022 – 2836/98/241013–10 $25.00/0 # 1998 Academic Press Limited

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Article No. mb981790 J. Mol. Biol. (1998) 279, 1013±1022
Structural Basis for the High Affinity ofAmino-Aromatic SH2 Phosphopeptide Ligands
Joseph Rahuel1, Carlos GarcõÂa-EcheverrõÂa2, Pascal Furet2
Andre Strauss1, Giorgio Caravatti2, Heinz Fretz2, Joseph Schoepfer2
and Brigitte Gay2*
1Novartis Pharma AG, CoreTechnology Area and2Oncology, ResearchDepartment, CH-4002, BaselSwitzerland
Abbreviations used: 2-Abz, 2-am(anthranyl); EGF, epidermal growtreceptor; GST, glutathione-S-transfreceptor substrate-1; MBP, maltosePDGF, platelet derived growth facreceptor; PLC-g, phospholipase C-gphosphotyrosine, ligand residues ato the position of the phosphotyropY 0; r.m.s., root-mean-square; SHdomain; TBS, Tris-buffered saline;activity relationship. The nomenclaet al. (1993) for SH2 domains is usein Figure 4.
0022±2836/98/241013±10 $25.00/0
An anthranyl moiety placed at the N terminus of a phosphotyrosine pep-tide potentiates the inhibitory effect of this small peptide on the bindingof the Grb2 SH2 domain to the EGF receptor. Using molecular modelingprocedures based on the Lck SH2 domain structure, this observation wasrationalized in terms of a suitably favorable p-p stacking interactionbetween the anthranyl moiety and the arginine aA2 (ArgaA2) residueside-chain of Grb2 SH2. The crystal structure of the Grb2 SH2 domain incomplex with the inhibitor 2-Abz-EpYINQ-NH2 (IC50 26 nM) has beensolved in two different crystal forms at 2.1 and 1.8 AÊ resolution. Thisstructure con®rms the modeling based on the Lck SH2 domain. TheArgaA2 residue is conserved in most SH2 domains. Thus, as expected,the anthranyl group also confers high af®nity to small peptide ligands ofother SH2 domains such as Lck-, PLC-g-amino-terminal and p85 amino-terminal SH2 domains as demonstrated by structure af®nity relationships(SAR) data. These potent peptides with an amino-terminal surrogategroup and the structure of Grb2 SH2 domain in complex with one suchpeptide represent good starting points for the design and optimization ofnew inhibitors of many SH2 domains.
# 1998 Academic Press Limited
Keywords: signal transduction; Grb2; crystal structure; SH2 domain;anthranyl
*Corresponding authorIntroduction
Src homology-2 (SH2) domains are phosphotyro-sine-binding modules found in many proteins thatare key players in several signal transduction path-ways (Mayer & Baltimore, 1993). SH2 domainshave been shown to bind to phosphorylated pro-tein tyrosine kinases or other proteins phosphory-lated in response to extracellular signals. The
ino-benzoylh factor; EGFR, EGFerase; IRS1, insulinbinding protein;
tor; PDGFR, PDGF1; pY,re numbered relativesine, which is denoted2, Src homology-2SAR, structureture adopted by Eckd here and explained
central roles played by protein tyrosine kinases inmitogenic signaling cascades and the involvementof aberrant or overexpressed protein tyrosinekinases in several cancers and proliferative diseaseshave made the development of inhibitors thatblock the SH2 domain binding desirable as poten-tial therapeutic agents (Schlessinger & Bar-Sagi,1994). SH2 domains are capable of binding phos-photyrosine and recognize the sequence context ofthe phosphotyrosine. Studies have shown that rela-tively short peptide sequences of less than ®ve resi-dues bind tightly to SH2 domains (Songyang et al.,1993; Zvelebil et al., 1995). The structures of variousSH2 domains, both free in solution and complexedwith phosphotyrosine peptides, have been deter-mined (Waksman et al., 1992; Eck et al., 1993; Leeet al., 1994; Hatada et al., 1995; Nolte et al, 1996;Rahuel et al, 1996). These structural studies haveshown that all SH2 domains consist of a centralantiparallel b-sheet with two ¯anking a-helices,and a second smaller two-stranded b-sheet. Thepeptide recognition site straddles the exposed edgeof the central b-sheet (bD strand). A conserved
# 1998 Academic Press Limited

1014 Structural Basis for Amino-Aromatic SH2 Ligands
arginine ArgbB5 forms an ion pair with the phos-phate moiety of the tyrosine. Another chargedgroup, ArgaA2, simultaneously interacts via itsside-chain nitrogen atoms with the phosphategroup of the tyrosine residue and with the back-bone carbonyl group of the pY-1 phosphopeptideresidue. This two-residue phosphotyrosine recog-nition element, ArgaA2 and ArgbB5, is a commonstructural feature of nearly all SH2 domainsknown so far. Another charged residue, HisbD4recognizes the ring of the phosphotyrosine and isalso conserved in most SH2 domains.
The amino-terminal binding regions and thephosphotyrosine-binding pockets are not criticalfor determining speci®city. Speci®city is deter-mined by the residues to the carboxy side of thephosphotyrosine residue. For example, while forthe Grb2 SH2 domain an asparagine residue atposition pY � 2 is the dominant feature, the LckSH2 binds preferentially peptides possessing thepYEEI motif. p85 SH2 recognizes the pYMXM orpYVXM motif (Songyang et al., 1993). As a corol-lary, the three-dimensional structures of SH2domains present signi®cant differences in theregions involved in binding the C-terminal part oftheir phosphopeptide ligands.
Grb2 is an ``adaptor'' protein made of one SH2domain ¯anked by two SH3 domains (for a review,see Downward et al., 1994). The SH3 domains bindto proline-rich motifs in the C-terminal part of theRas exchange factor Sos. Since the initial discoverythat the Grb2/Sos complex mediates signal trans-duction from the EGF receptor to Ras (Rozakis-Adcock et al., 1993), it has been observed that thiscomplex is also implicated in signal transductionfrom many other receptor tyrosine kinases to Ras(e.g. see Borrello et al., 1994; Liu et al., 1994).Design of molecules that block the interactionbetween the phosphotyrosine-containing activatedreceptors and the SH2 domain of Grb2 shouldinterrupt the Ras signaling pathway and maypromise therapeutic leads for diseases like cancerin which the Ras signaling pathway plays a majorrole (Sastry et al., 1997). Starting from the shortpeptide sequence Glu-pTyr-Ile-Asn-Gln recognizedby the Grb2 SH2 domain on the EGF receptor tyro-sine kinase, we have initiated the design of lowmolecular mass compounds that can ef®ciently dis-rupt these protein-protein interactions. The obser-vation that anthranyl as N-terminal groupproduces a dramatic increase of the binding af®-nity of the short phosphopeptide Glu-pTyr-Ile-Asn-Gln for Grb2 SH2 has been rationalized bymolecular modeling based on the X-ray structureof the Lck SH2 domain (Furet et al., 1997). Thecrystal structure of the Grb2 SH2 domain in com-plex with the high-af®nity peptide 2-Abz-Glu-pTyr-Ile-Asn-Gln-NH2 has been determined and ispresented here. The binding mode of the 2-Abzmoiety in Grb2 validates the model proposed byFuret et al. (1997). Because this model invoked astacking interaction between the N-terminal groupand the highly conserved SH2 domain residue
ArgaA2, the ``Abz effect'' was anticipated to begenerally applicable to most SH2 domain peptides.The relative binding af®nities for other SH2domains of peptides containing an acetyl or a2-Abz N-terminal capping group, but speci®cC-terminal sequences, have also been measuredin the current study.
Results and Discussion
Crystal structures
The observation that an anthranyl group addedat the N-terminal group of the Grb2 SH2 phospho-peptide ligand Glu-pTyr-Ile-Asn-Gln (EpYINQ)produced a dramatic increase in its binding af®nityhas been rationalized by molecular modeling(Furet et al., 1997) based on published Lck crystalstructures (Eck et al. 1993).
The potent peptide ligand 2-Abz-EpYINQ-NH2(IC50 26 nM) has now been cocrystallized in com-plex with the Grb2-SH2 domain. Two crystal formshave been obtained and the two structures havebeen solved at 2.1 and 1.8 AÊ . They are essentiallyidentical (r.m.s. difference after superposition of Ca
atoms � 0.300).The overall structure of the SH2 domain is con-
sistent with all previous descriptions of SH2domains and similar to that reported earlier at2.1 AÊ resolution (Rahuel et al., 1996; BrookhavenProtein Data Base code 1tze), in complex with theseven-residue phosphopeptide KPFpYVNV-NH2from BCR-abl.
Figure 1 presents a general view of the inhibitorin the binding site of the Grb2 SH2 domain. Asseen for the BCR-abl peptide, the 2-Abz-EpYINQ-NH2 peptide forms a b-turn that anchors at pYand pY � 2. The asparagine side-chain at pY � 2forms three hydrogen bonds with main-chainatoms of residues bD6 and bE4. In addition, theasparagine Ca and Cb atoms face the indolyl ringof TrpEF1, which forces the inhibitor to adopt atype I b-turn conformation.
As observed in the other structures of peptidecomplexes of SH2 domains, the phosphate groupof the 2-Abz-EpYINQ-NH2 peptide is situated in ahighly conserved hydrophilic binding pocket andis involved in an intricate network of hydrogen-bonding and charge-charge interactions. In particu-lar, the amino acid residue ArgaA2 makes adouble hydrogen bond contact with the phosphatemoiety. The various interactions are represented inFigure 2a. They are identical with those publishedfor structures of Src (Waksman et al., 1993) and Lck(Eck et al., 1993) SH2 domains in complex withshort phosphopeptide sequences such as Ac-pTyr-Glu-Glu-Ile (Tong et al., 1996). All structuresreported to date also show the interaction of thecarbonyl oxygen atom of the pY-1 with the side-chain of ArgaA2. Also, as in the structure of theLck SH2 domain with the phosphopeptide pYEEI,a strong hydrogen bond is made between the

Figure 1. Stereoview of the binding site of the Grb2 SH2 domain in the presence of the peptide Abz-EpYINQ-NH2.The surface of the protein is represented by different colors: in blue, the residue ArgaA2 responsible for the stackinginteraction with the Abz moiety; in green, part of the residues PhebD5 and GlnbD3 interacting with the Ile�1 side-chain; in violet, the residue TrpEF1, responsible for the b-turn conformation of the ligand. The inhibitor is shown inorange. Figures 1 to 3 were made with the program GRASP (Nicholls et al., 1991).
Structural Basis for Amino-Aromatic SH2 Ligands 1015
amide proton of the ligand pY � 1 residue and thebackbone carbonyl of HisbD4 (Figure 3).
These results con®rm that the Lck SH2 X-raystructure (and other SH2 domains such as PLC-gand p85) can indeed provide an appropriate frame-work to model the interactions between the Grb2SH2 domain and the phosphotyrosine and theN-terminal residues of its ligand. In the model ofthe complex based on the Lck crystal structure, theanthranyl moiety attached at the N terminus of thepeptide is stacked on the guanidinium group ofArgaA2. This is con®rmed in the Grb2 SH2 crystalstructure with the 2-Abz-EpYINQ-NH2 peptide(Figure 2). In addition to binding the guanidiniumgroup with the phosphate moiety and the carbonylgroup of residue ÿ1 of the ligand, the importance
Figure 2. Stacking interaction of the anthranyl moiety wiresentation of the surrounding hydrogen bond network. Theing the hydrogen bonding network involved in the bindininhibitor in orange. Hydrogen bonds are indicated; distance
of ArgaA2 is now reinforced by a stacking inter-action with an aromatic moiety at the N-terminalend of the phosphopeptide that confers high af®-nity for the SH2 domain. The 2-amino moiety ofAbz is also within hydrogen bonding distance ofan oxygen atom of the phosphate moiety (Figure 2).
Our work provides an additional example ofstacking interaction between the guanidiniumgroup of an arginine side-chain and an aromatic orheteroaromatic moiety, a type of interaction fre-quently observed in structural biology (for areview, see Flocco & Mowbray, 1994). As modeled(Furet et al., 1997), the part of the anthranyl moietyin contact with the electron-de®cient guanidiniumgroup is centered in the middle of the bicyclic elec-tron-rich region formed by the phenyl ring and the
th the residue ArgaA2 side-chain of Grb2 SH2 with rep-Grb2 SH2 backbone and side-chains of residues compos-
g of the phosphate moiety are shown in green and thes are in AÊ . a, Side-view of the stacking; b, front view.

Figure 3. Stereoview of the superposition of the ligand N-terminal binding region of the Grb2 and Lck SH2 domains.The same color code as in Figure 2 is used for the Grb2 SH2 complex. The Lck SH2 domain (Eck et al., 1993) is rep-resented in violet and its inhibitor in pink.
1016 Structural Basis for Amino-Aromatic SH2 Ligands
pseudo six-membered ring resulting from the intra-molecular hydrogen bond between the 2-aminosubstituent and the carbonyl group, which con-®rms the electrostatic origin of the stabilization(Figure 2b).
Competition experiments withsynthetic phosphopeptides
Grb2 and PLC-g bind with high af®nity to theEGFR (at Tyr1068 and Tyr992 of the EGFR,respectively; Batzer et al., 1994; Rotin et al., 1992).Studies using small pY-containing peptides haveshown that molecules containing only ®ve residues(one residue N-terminal and three residues C-term-inal to the critical pY) can effectively block SH2-mediated binding to activated receptors (Fantl et al.,1992). We made use of the synthetic phosphopep-tides EpYINQ (where pY corresponds to Tyr1068of EGFR) and DpYIIP (where pY corresponds toTyr1021 of PDGFR, another binding site of PLC-g)to outcompete Grb2 and PLC-g SH2 domains,respectively, in in vitro association experimentswith a phosphorylated recombinant EGFR. Thesephosphopeptides were capped at the N terminusby an acetyl or an anthranyl moiety and, for Grb2,
Table 1. EGFR ELISA competition binding assay forGST/Grb2 SH2 phosphopeptide inhibitors
No. Sequence IC50 (m)
1 Ac-EpYINQ-NH2 1.25 � 0.262 2-Abz-EpYINQ-NH2 0.026 � 0.0053 Bz-EpYINQ-NH2 0.8 � 0.124 3-Abz-EpYINQ-NH2 0.034 � 0.035 4-Abz-EpYINQ-NH2 0.232 � 0.0156 H-bA-EpYINQ-NH2 2.55 � 0.4
by various analogs of the anthranyl group. Table 1shows the results obtained for Grb2.
Some structure-activity relationships (Table 1)analyzed in the light of the X-ray structure allow abetter understanding of the ``anthranyl effect''. It is®rst apparent that the 2-amino substituent of Abz(compound 2 in Table 1) is essential to produce thelarge gain in af®nity, since the non-substituted ana-log 3 has only similar af®nity for the Grb2 SH2domain as the acetyl-protected reference phospho-peptide 1. One can check whether the role of theamino substituent consists only in ®xing the con-formation of the N-terminal part of the ligand inthe stacking conformation by formation of theinternal hydrogen bond with one of the phosphateoxygen atoms as seen in the structure or whether italso has an electronic effect due to its electron-donating capacity that strengthens the stackinginteraction with the electron-de®cient guanidiniumgroup of ArgaA2. These questions can beanswered by considering the two analogs 4 and 5in which the amino substituent has been moved tothe 3 and 4 positions, respectively. Modelingshows that in the 3 position (compound 4), theamino substituent can still be within hydrogenbonding distance from one of the phosphate oxy-gen atoms, while in the 4 position (compound 5) itis no longer possible. Considering that, as for the2-Abz-protected peptide, almost two orders ofmagnitude in binding af®nity are gained in com-pound 4 against only one with compound 5, it canbe concluded that both factors (internal hydrogenbond and electronic factors) contribute aboutequally to the anthranyl effect. The cooperativity ofthe stacking interaction and the intramolecularamino-phosphate hydrogen bond can be furtherillustrated by comparing compounds 3 and 6. Inthe former, the unsubstituted benzoyl moiety can,

Table 4. Phosphopeptide assay: relative af®nities ofphosphopeptides for GST/Lck SH2 domain
Sequence IC50 (mM)
Ac-QpYEEI-NH2 3.2 � 0.79Abz-QpYEEI-NH2 0.030 � 0.003
Table 2. EGFR ELISA competition binding assay forGST/PLC-g phosphopeptide
Sequence IC50 (mM)
Ac-DpYIIP-NH2 2.47 � 0.27Abz-DpYIIP-NH2 0.058 � 0.006
Structural Basis for Amino-Aromatic SH2 Ligands 1017
in principle, give rise to a stacking interaction withArgaA2, while modeling suggests that in the latter,which lacks a stacking moiety, the N-terminal alky-lamino chain can form an intramolecular hydrogenbond with the phosphate group. Despite posses-sing one of the two important structural features ofthe anthranyl moiety, these compounds do notshow any improvement in potency compared toreference compound 1. Thus, as modeled, the largeincrease in potency induced by the anthranylgroup seems to depend on the synergy between anenthalpic (p-p stacking strengthened by an elec-tron-donating substituent) and an entropic (confor-mational restriction by an internal hydrogen bond)favorable effect.
Since the anthranyl effect was shown to originatein an interaction with the well-conserved ArgaA2residue, it should be observed for SH2 domainsother than Grb2. We applied the same N-terminalmodi®cations to phosphopeptide sequencesspeci®c to PLC-g, p85 and Lck SH2 domains, all ofwhich contain ArgaA2. The results in Table 2show that for PLC-g, as for Grb2, the presence ofthe anthranyl group leads to a dramatic increase inpotency. For Lck and p85 SH2 domains (Grb2 SH2domain as control), biotinylated truncated versionsof the polyoma Middle T Ag (biotin-EEEPQ-pY324EEIPIYL-NH2, Src/Lck SH2 binding site onthe polyoma Middle T Ag), of the PDGF receptor(biotin-DMSKDESVDpY751VPMLDMK-NH2, p85-N-SH2 binding site on the PDGF-R) and of theEGF receptor (biotin-DDTFLPVPEpY1068INQSVPK-NH2, Grb2 SH2 binding site on the EGF-R), weremade to assay binding to the SH2 domains of Lck,p85 and Grb2, respectively. QpYEEI, DpYVPMand EpYINQ peptides with an acetyl or an anthra-nyl N-terminal capping group were then assayedfor binding with different af®nities to Lck, p85 andGrb2 SH2 domains, respectively. The results of thecompetition experiments are summarized inTables 3, 4 and 5, respectively. The results for Grb2are con®rmed in this new assay (Table 3). As fore-seen for Lck and p85, the addition of the anthranylmoiety also remarkably enhanced phosphopeptidebinding af®nity.
Table 3. Phosphopeptide assay: relative af®nities ofphosphopeptides for GST/Grb2 SH2 domain
Sequence IC50 (mM)
Ac-EpYINQ-NH2 2.27 � 0.24Abz-EpYINQ-NH2 0.015 � 0.001
Conclusions
The large increase in af®nity of anthranyl cappedphosphopeptides for different SH2 domains,including Grb2, Lck, PLC-g and p85, has beendemonstrated in in vitro binding assays. The struc-tural analysis of such a powerful ligand in complexwith the Grb2 SH2 domain provides the con®r-mation that p-p stacking interactions take placebetween the N-terminal group and the guanidi-nium group of ArgaA2, the amino substituent of2-Abz stabilizing the conformation of the ligand byan internal hydrogen bond. Sequence analysisshows that ArgaA2 is conserved among almost allSH2 domains. Therefore, it appears possible toboost the general potency of small phosphopep-tides towards SH2 domains just by adding anamino-aromatic moiety to the residue N-terminalto the phosphotyrosine residue. The small size andthe high potency of the phosphopeptides thusobtained represent a major advance in the searchfor low molecular mass compounds that can blockcrucial protein-protein interactions in the signaltransduction pathways of tyrosine kinase growthfactor receptors.
Materials and Methods
Peptide synthesis
Peptides were synthesized on solid phase according toalready published protocols (GarcõÂa-EcheverrõÂa et al.,1997). The purity of the peptides was veri®ed byreversed-phase analytical HPLC, and the identity of the®nal products was assessed by amino acid and massspectral analyses.
EGFR assay
The EGFR assay has been described in detail (Rahuelet al., 1996). Brie¯y, phosphorylated MBP-EGFR immobi-lized on a solid phase (polystyrene microtiter plates,NUNC MAXYSORB) was incubated with a GST/Grb2-SH2 or GST/PLC-g-SH2 fusion protein capable of bind-ing to it, in the presence of phosphopeptide in buffer orbuffer alone. Bound SH2 was detected with polyclonalrabbit anti-GST antibody. Following washing, horse-
Table 5. Phosphopeptide assay: relative af®nities ofphosphopeptides for GST/p85 SH2 domain
Sequence IC50 (mM)
Ac-DpYVPM-NH2 1.15 � 0.099Abz-DpYVPM-NH2 0.037 � 0.001

1018 Structural Basis for Amino-Aromatic SH2 Ligands
radish peroxidase-conjugated mouse anti-rabbit antibodywas added. Peroxidase activity is monitored at 655 nmon a plate reader by adding 100 ml/well of a solution oftetramethylbenzidine as substrate.
Phosphopeptide assays
This assay has been described (GarcõÂa-EcheverrõÂa et al.,1997). A GST/SH2 domain fusion protein was paired,after buffer or varying concentrations of unlabeled phos-phopeptides (as competitors) were added, with anappropriate high-af®nity biotinylated phosphopeptidelinked to streptavidin-coated microtiter plates. These bio-tinylated phosphopeptides are a subset of phosphotyro-syl peptides corresponding to SH2 targets on the EGFR,PDGFR and Polyoma Middle T Ag. Biotinylated phos-phopeptide (100 ml of 10 ng/ml in 50 mM Tris, pH 7.5)was added to wells of streptavidin-coated plates (Boer-hinger Mannheim) and incubated overnight at 4�C, thenrinsed with TBS. Selected peptide concentrations, or buf-fer alone, and GST/Grb2-SH2 (3.2 ng/ml), GST/Lck-SH2 (3.2 ng/ml) or GST/p85-N-SH2 (0.7 ng/ml) werethen added to a 100 ml/well total volume of TBS buffercontaining 0.1% (v/v) Tween. The assay proceeded asdescribed above for the MBP-EGFR assay with primaryanti-GST antibody and secondary peroxidase-conjugatedgoat-antirabbit IgG.
Data Analysis
Peptide inhibition was calculated as a percentage ofthe reduction in absorbance in the presence of each pep-tide inhibitor concentration compared to the absorbanceobtained with GST/SH2 in the absence of peptide inhibi-tor. Dose-response relationships were constructed bynon-linear regression of the competition curves withGra®t (Erithacus Software, London, UK: 50% inhibitoryconcentrations (IC50) were calculated from the regressionlines.
Cloning and expression of recombinant proteins
Glutathione-S-transferase (GST) fusion proteins of p85N-terminal (GST/p85 SH2) and Lck (GST/Lck SH2) SH2domains were obtained from Santa Cruz Biotech. GST/PLC-g-1 (N-terminal domain) was a kind gift from MikeMoran. GST/Grb2 SH2 was expressed as described(Braunwalder et al. 1996). The pMAL-c2 expression vec-tor (New England Biolabs) was used to express theC-terminal domain of the epidermal growth factorreceptor intracellular domain (residues 976 to 1210) as amaltose-binding fusion protein (MBP-EGFR) in Escheri-chia coli (Rahuel et al. 1997).
Purification of the Grb2 SH2 domain
The cell pellet (120 g) from a 10 l culture was soni-cated and the supernatant after centrifugation wasapplied to a 50 ml column of glutathione attached tocross-linked agarose (Sigma G-4510). The column waswashed with a buffer containing 50 mM Tris-HCl(pH 7.5) and the protein eluted with 10 mM glutathionein the same buffer. A 42 ml fraction containing 8 mg/mlof the pure fusion protein was obtained, and stored atÿ80�C. In a second step, 10.5 ml of the pure fusion pro-tein solution was digested in the presence of 250 mg ofFactor Xa (Biolabs 800-10L; ratio of substrate to enzyme�300, w/w) and dialyzed at the same time against
20 mM Hepes buffer (pH 7.5), overnight at 4�C. The sol-ution was applied to a 30 ml column of SP SepharoseFast Flow (Pharmacia). The GST was eliminated in the¯owthrough and the column washed with the Hepesbuffer. The pure SH2 domain was eluted with the Hepesbuffer containing 0.4 M sodium chloride. The 8 mleluted fraction contained 2 mg/ml of the SH2 domain.
The material obtained was analyzed by SDS-PAGE.Two bands of molecular mass differing by less than 1000Da could be seen. Analysis by reverse phase HPLC alsoindicated the presence of two major peaks. Further anal-ysis of the fractions corresponding to each of the twopeaks in an experiment of N-terminal sequencing com-bined with mass determination showed a total of sixdifferent species with various lengths at either the N orthe C-terminal end of the protein.
The protein was also checked by size-exclusion chro-matography: two peaks were observed when analyticalamounts were applied onto the column (Biorad, Bio-SilSEC 125, 300�7.8 mm). From the correlation betweenretention time and molecular mass, it could be inferredthat the solution was a mixture of the monomer with adimer. When the mixture was heated to 50�C for tenminutes, the ``faster'' peak corresponding to the dimerwas almost totally removed, while the area of the``slower'' peak corresponding to the monomer wasincreased by the same amount.
Crystallization and data collection
In spite of the observed heterogeneity, crystallizationtrials were performed. All samples were heated in theabove-described fashion before being concentrated to10 to 20 mg/ml prior to crystallization experiments.The protein was submitted to a screening for crystalli-zation conditions (Jancarik & Kim, 1991) in the pre-sence of the ligand (2-Abz-EpYINQ-NH2) in a proteinto ligand molar ratio of 1:5. Two precipitants werefound for growing crystals, ammonium phosphate andammonium sulfate, which provided two different crys-tal forms. Conditions for crystallization were re®ned inorder to obtain X-ray quality crystals. Type I crystalsgrew within a few days in 0.7 M ammonium phos-phate, 10% PEG 400 in 50 mM sodium cacodylate(pH 6.5). They appeared as thin plates of about0.2 mm�0.2 mm and diffracted to 2.8 AÊ . Macroseedingmethods were successfully applied to get larger crys-tals: in this way a crystal with a size of0.30 mm�0.25 mm�0.1 mm diffracting to 2.1 AÊ couldbe obtained. Type II crystals grew within a few weeksin 2.0 M ammonium sulfate, 2% PEG 400 in 0.1 MHepes (pH 7.5). Crystals grew up to a size of0.7 mm�0.3 mm�0.1 mm and diffracted to 1.8 AÊ .
X-ray diffraction data were collected using a FASTarea detector (Enraf-Nonius, Delft, The Netherlands)positioned on a FR571 rotating anode X-ray generatoroperated at 40 kV and 70 mA producing CuKa graphite-monochromated radiation and having an apparent focalspot of 0.3 mm�0.3 mm. Frames of 0.1� were collectedwith an exposure time of 90 seconds at a 40 mm crystal-to-detector distance for both type I and type II crystals.The frames were processed with the program MADNES(Messerschmidt & Plugrath, 1987) and the data weremerged using the CCP4 program package (1994). Datawere collected from a seeded crystal of the type I crystalto 2.1 AÊ resolution and from a type II crystal to 1.8 AÊ
resolution. The results for data collection from both crys-tals are summarized in Table 6.
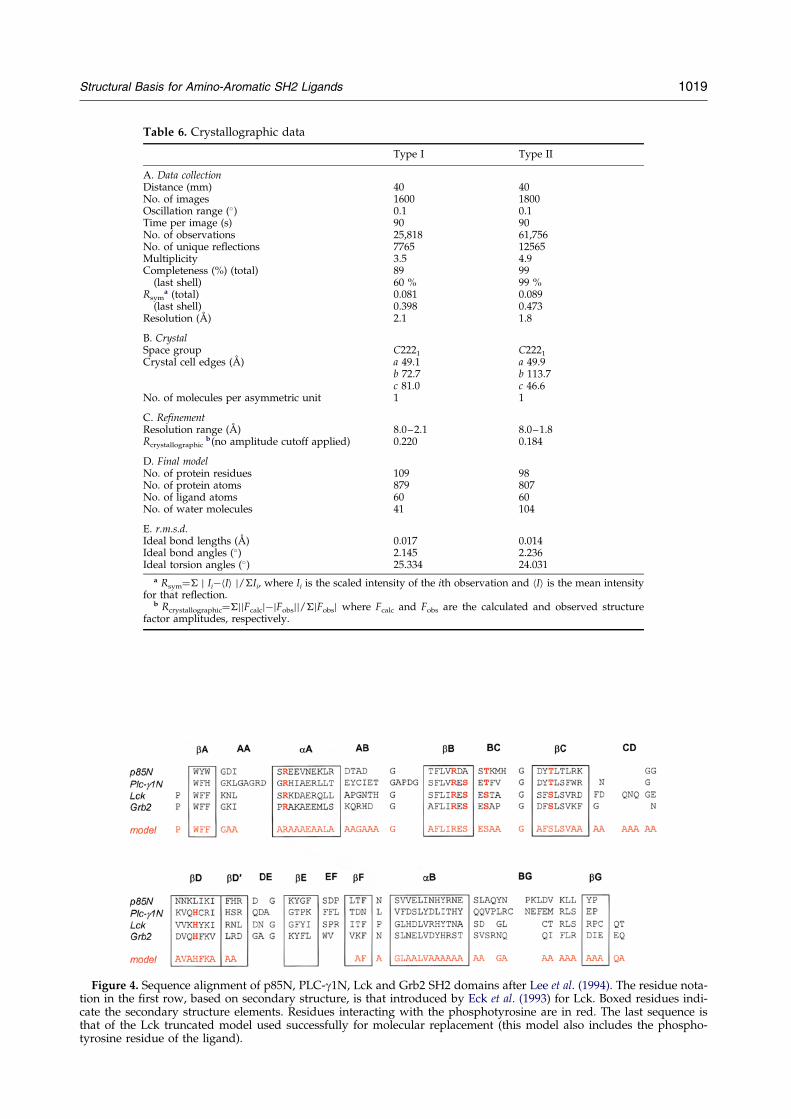
Figure 4. Sequence alignment of p85N, PLC-g1N, Lck and Grb2 SH2 domains after Lee et al. (1994). The residue nota-tion in the ®rst row, based on secondary structure, is that introduced by Eck et al. (1993) for Lck. Boxed residues indi-cate the secondary structure elements. Residues interacting with the phosphotyrosine are in red. The last sequence isthat of the Lck truncated model used successfully for molecular replacement (this model also includes the phospho-tyrosine residue of the ligand).
Table 6. Crystallographic data
Type I Type II
A. Data collectionDistance (mm) 40 40No. of images 1600 1800Oscillation range (�) 0.1 0.1Time per image (s) 90 90No. of observations 25,818 61,756No. of unique reflections 7765 12565Multiplicity 3.5 4.9Completeness (%) (total) 89 99
(last shell) 60 % 99 %Rsym
a (total) 0.081 0.089(last shell) 0.398 0.473
Resolution (AÊ ) 2.1 1.8
B. CrystalSpace group C2221 C2221
Crystal cell edges (AÊ ) a 49.1 a 49.9b 72.7 b 113.7c 81.0 c 46.6
No. of molecules per asymmetric unit 1 1
C. RefinementResolution range (AÊ ) 8.0±2.1 8.0±1.8Rcrystallographic
b(no amplitude cutoff applied) 0.220 0.184
D. Final modelNo. of protein residues 109 98No. of protein atoms 879 807No. of ligand atoms 60 60No. of water molecules 41 104
E. r.m.s.d.Ideal bond lengths (AÊ ) 0.017 0.014Ideal bond angles (�) 2.145 2.236Ideal torsion angles (�) 25.334 24.031
a Rsym�� j IiÿhIi j/�Ii, where Ii is the scaled intensity of the ith observation and hIi is the mean intensityfor that re¯ection.
b Rcrystallographic��jjFcalcjÿjFobsjj/�jFobsj where Fcalc and Fobs are the calculated and observed structurefactor amplitudes, respectively.
Structural Basis for Amino-Aromatic SH2 Ligands 1019

Figure 5. Stereoview of an FobsÿFcalc electron density map from which the inhibitor has been omitted from Fcalc of thebinding site of Grb2-SH2 in the type II crystal after re®nement. The electron density is contoured at 3s (s is the stan-dard deviation of the electron density value in the map covering the asymmetric unit). The inhibitor is represented asa ball-and-stick model.
1020 Structural Basis for Amino-Aromatic SH2 Ligands
Structure solution by molecular replacement
Molecular replacement was performed with the 2.1 AÊ
resolution data set of the type I crystal. Trials were doneusing structures of SH2 domains of Src and Lck: twostructures of the Src SH2 domain in complex with twopeptidic inhibitors elucidated by Waksman et al. (1992)and the structure of the Lck SH2 domain, also in com-plex with a peptidic inhibitor, elucidated by Eck et al.(1993). The sequence identity of the Src and the Lckdomain with the Grb2 SH2 domain is 35% and 42%,respectively.
Models were built with each of the structures takinginto account the actual Grb2 sequence and the knowl-edge of the secondary structure of SH2 domains. Resi-dues that were not identical in the alignment of Grb2and the probe protein were truncated to alanine resi-dues. Parts of the model that belonged to neithera-helices nor b-sheets were removed. Many rotationfunction calculations with models from both Src andLck were carried out by removing loops in differentcombinations, with or without truncation of the side-chains, even with incorporation of the main chain andphosphotyrosine residue of inhibitors. Different pro-grams were used for the calculation, including ALMN(CCP4, 1994), MERLOT (Fitzgerald, 1988) and X-PLOR(BruÈ nger, 1992). None of these attempts provided clearstructural solutions.
These ®rst attempts were followed by a more empiri-cal approach. Starting from the model of Lck, in whichresidues non-identical with those of Grb2 were truncatedto alanine, groups of 20 consecutive residues wereomitted systematically and alternatively along thesequence. Every truncated model was used for rotationfunction calculation with the program MERLOT. Afteromission of residues 61 to 80, a peak signi®cantly higherthan the others was observed. This result was optimizedand the exact sequence of residues to leave out to get thehighest peak was determined. Further optimization waspursued by deletion of residues in other parts of themodel. The sequence of the resulting model is rep-resented in Figure 4 together with the sequence align-ment of Grb2 and other SH2 domains. The highest peakvalue for the rotation function was obtained by includinga phosphotyrosine residue in the model. The peak in therotation function, using data between 10.0 and 4.0 AÊ ,
corresponding to the best model, is 4.97 times the r.m.s.deviation above the mean and 1.35 times higher than the®rst noise peak. The suggested orientation of the modelwas applied and a peak of 7.5 times the r.m.s. deviationabove the mean and 1.57 times higher than the ®rstnoise peak was found in the translation function (pro-gram TFFC, CCP4, 1994) using data in the range of 10.0to 2.2 AÊ . The R-factor between observed and calculateddata between 8.0 and 4.0 AÊ was then 0.540.
When a 1.8 AÊ resolution data set of the type II crystalbecame available, molecular replacement was performedon this new crystal form with a partially re®ned modelfrom crystal type I (R-factor 0.28). After positioning of themodel in the type II crystal cell in agreement with theresults of rotation and translation functions the R-factorwas 0.43.
Crystallographic Refinement
Re®nement of both crystal structures was performedwith the programs X-PLOR (BruÈ nger, 1992) and TNT(Tronrud et al., 1987). Rebuilding of the model was doneusing the program O (Jones et al., 1991) on a SiliconGraphics Indigo Work Station. Re®nement results arelisted in Table 6.
The re®nement was started with X-PLOR using the2.1 AÊ data set of the type I crystal. The crystal packingbased on the results of the molecular replacement wasreasonable (no severe overlapping of symmetry-relatedmolecules), although the R-factor was rather high (0.54).Nevertheless, difference electron density maps revealedelectron density for loops and side-chains that were notincluded in the search model and electron density wasalso observable in the region of the active site where theinhibitor was expected to bind. Missing loops and side-chains were successively built into the model, which wasalso progressively extended at the N and C-terminalends. The model was progressively built, including theinhibitor and solvent molecules. Final re®nement cycleswere performed with TNT.
After re®nement the model consists of residues 53 to161 of Grb2, extending slightly beyond the SH2 domainat both termini: 41 solvent molecules have been added tothe model. All atoms of the ligand were located.

Structural Basis for Amino-Aromatic SH2 Ligands 1021
Re®nement of the type II crystal form followed theprocedure described above for the type I crystal form.The ®nal model consists of residues 56 to 153 of Grb2and 101 solvent molecules have been included. Allatoms of the ligand were located. A difference electrondensity map of the binding site of the Grb2 SH2 domainin crystal type II is shown in Figure 5. All the atoms ofthe ligand are well de®ned with the exception of theside-chain carboxylic group of Glu ÿ1 and the side-chainof Gln � 3.
Acknowledgments
The authors are grateful to C. Stamm, R. Wille, D. Arzfor their technical assistance, to S. E. Shoelson for provid-ing the coordinates of the Lck SH2 domain and to J.Priestle for critical reading of the manuscript.
References
Batzer, A. G., Rotin, D., Urena, J. M., Skolnik, E. Y. &Schlessinger, J. (1994). Hierarchy of binding sites forGrb2 and Shc on the epidermal growth factor recep-tor. Mol. Cell. Biol. 14, 5192±201.
Borrello, M. G., Pelicci, G., Arighi, E., De Filipis, L.,Greco, A., Bongarzone, I., Rizzetti, M. G., Pelicci,P. G. & Pierotti, M. (1994). The oncogenic versionsof the Ret and Trk tyrosine kinases bind SHC andGrb2 adaptor proteins. Oncogene, 9, 1661±1668.
Braunwalder, A. F., Weennogle, L., Gay, B., Lipson, K. E.& Sills, M. (1996). Application of scintillating micro-titer plates to measure phosphopeptide interactionswith the Grb2 SH2 binding domain. J. Biomol.Screening, 1, 23±26.
BruÈ nger, A. T. (1992). A system for X-ray crystallographyand NMR. X-PLOR Version 3.1., Yale UniversityPress, New Haven and London.
Collaborative Computational Project Number 4 (1994).The CCP4 suite, programs for protein crystallogra-phy. Acta Crystallog sect. D, 50, 760±763.
Downward, J. (1994). The GRB2/Sem-5 adaptor protein.FEBS Letters, 338, 113±117.
Eck, M. J., Schoelson, S. E. & Harrison, S. C. (1993). Rec-ognition of a high-af®nity phosphotyrosyl peptideby the Src homology-2 domain of p56lck. Nature,362, 87±91.
Fantl, W. J., Escobedo, J. A., Martin, G. A., Turck, C. W.,del Rosario, M., McCormick, F. & Williams, L. T.(1992). Distinct phosphotyrosines on a growth fac-tor receptor bind to speci®c molecules that mediatedifferent signaling pathways. Cell, 69, 413±423.
Fitzgerald, P. M. D. (1988). MERLOT, an integratedpackage of computer programs for the determi-nation of crystal structures by molecular replace-ment. J. Appl. Crystallog. 21, 273±278.
Flocco, M. M. & Mowbray, S. L. (1994). Planar stackinginteractions of arginine and aromatic side-chains.J. Mol. Biol. 235, 709±717.
Furet, P., Gay, B., GarcõÂa-EcheverrõÂa, C., Rahuel, J.,Fretz, H., Schoepfer, J. & Caravatti, G. (1997). Dis-covery of 3-amino-benzyloxycarbonyl as N-terminalgroup conferring high af®nity to the minimal phos-phopeptide sequence recognized by the Grb2-SH2domain. J. Med. Chem. 40, 3551±3556.
GarcõÂa-EcheverrõÂa, C., Stamm, C., Wille, R., Arz, D. &Gay, B. (1997). Biotinylated phosphotyrosine pep-
tides: a valuable tool for studies on phosphopeptideinteractions with SH2 and PTB domains. LIPS, 4,49±53.
Hatada, M. H., Lu, X., Laird, E. R., Green, J.,Morgenstern, J. P., Lou, M., Marr, C. S., Phillips,T. B., Ram, M. K., Therault, K., Zoller, M. J. &Karas, J. L. (1995). Molecular basis for interaction ofthe protein tyrosine kinase ZAP-70 with the T-cellreceptor. Nature, 377, 32±38.
Jancarik, J. & Kim, S. H. (1991). Sparse matrix sampling:a screening method for crystallization of protein.J. Appl. Crystallog. 24, 409±411.
Jones, A. T., Zou, J.-Y., Cowan, S. W. & Kjeldgaard, M.(1991). Improved methods for building proteinmodels in electron density maps and the location oferrors in these models. Acta Crystallog. sect. A, 47,110±119.
Lee, C.-H., Kominos, D., Jacques, S., Margolis, B.,Schlessinger, J., Shoelson, S. E. & Kuriyan, J. (1994).Crystal structures of peptide complexes of theamino-terminal SH2 domain of the Syp tyrosinephosphatase. Structure, 2, 423±438.
Liu, L., Damen, J. E., Cutler, R. L. & Krystal, G. (1994).Multiple cytokines stimulate the binding of a com-mon 145-kilodalton protein to Shc at the Grb2 rec-ognition site of Shc. Mol. Cell. Biol. 14, 6926±6935.
Mayer, B. J. & Baltimore, D. (1993). Signalling throughSH2 and SH3 domains. Trends Cell Biol. 3, 8±13.
Messerschmidt, A. & P¯ugrath, J. W. (1987). Crystalorientation and X-ray pattern prediction for area-detector diffractometer systems in macromolecularcrystallography. J. Appl. Crystallog. 20, 306±315.
Nicholls, A., Sharp, K. & Honig, B. H. (1991). Proteinfolding and association; insights from the interfacialand thermodynamic properties of hydrocarbons.Proteins: Struct. Funct. Genet. 11, 281±296.
Nolte, R. T., Eck, M. J., Schlessinger, J., Shoelson, S. E. &Harrison, S. C. (1996). Crystal structure of the PI3-kinase p85 amino-terminal SH2 domain and itsphosphopeptide complexes. Nature Struct. Biol. 3,364±373.
Rahuel, J., Gay, B., Erdmann, D., Strauss, A., GarcõÂa-EcheverrõÂa, C., Furet, P., Caravatti, G., Fretz, H.,Schoepfer, J. & GruÈ tter, M. G. (1996). Structuralbasis for speci®city of Grb2±SH2 revealed by anovel ligand binding mode. Nature Struct. Biol. 3,586±589.
Rotin, D., Margolis, B., Mohammadi, M., Daly, R. J.,Daum, G., Li, N., Fischer, E. H., Burgess, W. H.,Ullrich, A. & Schlessinger, J. (1992). SH2 domainsprevent tyrosine dephosphorylation of the EGFreceptor: identi®cation of Tyr992 as the high-af®nitybinding site for SH2 domains of phospholipase Cgamma. EMBO J. 11, 559±567.
Rozakis-Adcock, M., Fernley, R., Wade, J., Pawson, T. &Bowtell, D. (1993). The SH2 and SH3 domains ofmammalian Grb2 couple the EGF receptor to theRas activator mSos1. Nature, 363, 82±85.
Sastry, L., Cao, T. & King, R. (1997). Multiple Grb2-pro-tein complexes in human cancer cells. Int. J. Cancer,70, 208±213.
Schlessinger, J. & Bar-Sagi, D. (1994). Activation of Rasand other signalling pathways by receptor tyrosinekinases. Cold Spring Harbor Symp. Quant. Biol. 59,173±179.
Songyang, Z., Shoelson, S. E., McGlade, J., Olivier, P.,Pawson, T., Bustelo, X. R., Barbacid, M., Sabe, H.,Hanafusa, H., Yi, T., Ren, R., Baltimore, D.,Ratnofsky, S., Feldman, R. A. & Cantley, L. C.

1022 Structural Basis for Amino-Aromatic SH2 Ligands
(1993). Speci®c motifs recognized by the SH2domains of Csk, 3BP2, fps/fes, GRB-2 HCP, SHC,Syk, and Vav. Mol. Cell. Biol. 14, 2777±2785.
Tong, L., Warren, T. C., King, J., Betageri, R., Rose, J. &Jakes, S. (1996). Crystal structures of the humanP56lck SH2 domain in complex with two shortphosphotyrosyl peptides at 1.0 AÊ and 1.8 AÊ resol-ution. J. Mol. Biol. 256, 601±610.
Tronrud, D. E., Ten, Eyck L. F. & Matthews, B. W.(1987). An ef®cient general-purpose least-squarere®nement program for macromolecular structures.Acta Crystallog. sect. A, 43, 489±501.
Waksman, G., Kominos, D., Robertson, S. C., Pant, N.,Baltimore, D., Birge, R. B., Cowburn, D., Hanafusa,
H., Mayer, B. J., Overduin, M., Resh, M. D., Rios,C. B., Silverman, L. & Kuriyan, J. (1992). Crystalstructure of the phosphotyrosine recognitiondomain SH2 of v-src complexed with tyrosine-phos-phorylated peptides. Nature, 358, 646±653.
Waksman, G., Schoelson, S. E., Pant, N., Cowburn, D. &Kuriyan, J. (1993). Binding of a high af®nity phos-photyrosyl peptide to the Src SH2 domain: crystalstructures of the complexed and peptide-free forms.Cell, 72, 779±790.
Zvelebil, M. J. J. M., Panatoyou, G., Linacre, J. &Water®eld, M. D. (1995). Prediction and analysis ofSH2 domain-phosphopeptide interactions. ProteinEng. 6, 527±533.
Edited by D. Rees
(Received 8 December 1997; received in revised form 6 March 1998; accepted 9 March 1998)
Related Documents











