Structural Basis for Syk Tyrosine Kinase Ubiquity in Signal Transduction Pathways Revealed by the Crystal Structure of its Regulatory SH2 Domains Bound to a Dually Phosphorylated ITAM Peptide Klaus Fu ¨ tterer 1 , Jane Wong 2 , Richard A. Grucza 1 , Andrew C. Chan 2,3 and Gabriel Waksman 1 * 1 Department of Biochemistry and Molecular Biophysics 2 Departments of Medicine and Pathology and 3 Howard Hughes Medical Institute Washington University School of Medicine, Campus Box 8231 660 South Euclid Av. Saint Louis, MO 63110, USA The Syk family of kinases, consisting of ZAP-70 and Syk, play essential roles in a variety of immune and non-immune cells. This family of kinases is characterized by the presence of two adjacent SH2 domains which mediate their localization to the membrane through receptor encoded tyrosine phosphorylated motifs. While these two kinases share many structural and functional features, the more ubiquitous nature of Syk has suggested that this kinase may accommodate a greater variety of motifs to mediate its function. We present the crystal structure of the tan- dem SH2 domain of Syk complexed with a dually phosphorylated ITAM peptide. The structure was solved by multiple isomorphous replacement at 3.0 A ˚ resolution. The asymmetric unit comprises six copies of the liganded protein, revealing a surprising flexibility in the relative orien- tation of the two SH2 domains. The C-terminal phosphotyrosine-binding site is very different from the equivalent region of ZAP-70, suggesting that in contrast to ZAP-70, the two SH2 domains of Syk can function as independent units. The conformational flexibility and structural indepen- dence of the SH2 modules of Syk likely provides the molecular basis for the more ubiquitous involvement of Syk in a variety of signal transduc- tion pathways. # 1998 Academic Press Keywords: X-ray crystal structure; Syk kinase; SH2 domains; ITAM *Corresponding author Introduction ZAP-70 and Syk comprise a family of hemato- poietic cell specific protein tyrosine kinases (PTKs) that are required for antigen and antibody receptor function (reviewed by Chan & Shaw, 1995; Wange & Samelson, 1996; Reth & Wienands, 1997; Kurosaki, 1997). While ZAP-70 is expressed in T and natural killer cells, Syk is expressed in B cells, mast cells, polymorphonuclear leukocytes, plate- lets, macrophages, and immature T cells. The absence of either ZAP-70 or Syk results in arrested T and B cell development and in functional defects of a variety of immune receptors including the T cell antigen receptor (TCR), the B cell antigen receptor (BCR), and the receptors for IgG and IgE (FcgRI and FceRI; Costello et al., 1996; Zhang et al., 1996; Crowley et al., 1997; and reviewed by Cheng & Chan, 1997 and by Kurosaki, 1997). In addition, this family of PTKs has been implicated in activat- ing NK cells (Brumbaugh et al., 1997; Lanier et al., 1998) and signaling by non-immune receptors such as G-protein coupled and integrin receptors (Wan et al., 1996; Gao et al., 1997; Poole et al., 1997). The signaling components of the antigen and Ig recep- tors contain sequence motifs known as ITAMs (for Immunoreceptor Tyrosine-based Activation Motifs) which have the consensus sequence YxxL/I-x 7/8 - YxxL/I (Reth, 1989). Phosphorylation of both tyro- sine residues within the ITAM by the Src-family PTKs is required for efficient interaction of ZAP-70 and Syk with the receptor subunits and for recep- tor function (Iwashima et al., 1994; Isakov et al., E-mail address of the corresponding author: [email protected] Abbreviations used: ITAM, immunoreceptor tyrosine- based activation motif; PTK, protein tyrosine kinase; TCR, T cell antigen receptor; SH2, Src homology 2; BCR, B cell antigen receptor; r.m.s., root mean square; MIR, multiple isomorphous replacement; NCS, non- crystallographic symmetry. Article No. mb981964 J. Mol. Biol. (1998) 281, 523–537 0022 – 2836/98/330523–15 $30.00/0 # 1998 Academic Press

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Article No. mb981964 J. Mol. Biol. (1998) 281, 523±537
Structural Basis for Syk Tyrosine Kinase Ubiquity inSignal Transduction Pathways Revealed by theCrystal Structure of its Regulatory SH2 DomainsBound to a Dually Phosphorylated ITAM Peptide
Klaus FuÈ tterer1, Jane Wong2, Richard A. Grucza1, Andrew C. Chan2,3
and Gabriel Waksman1*
1Department of Biochemistryand Molecular Biophysics2Departments of Medicine andPathology and 3HowardHughes Medical InstituteWashington University Schoolof Medicine, Campus Box 8231660 South Euclid Av.Saint Louis, MO 63110, USA
E-mail address of the [email protected]
Abbreviations used: ITAM, immbased activation motif; PTK, proteTCR, T cell antigen receptor; SH2,BCR, B cell antigen receptor; r.m.sMIR, multiple isomorphous replaccrystallographic symmetry.
0022±2836/98/330523±15 $30.00/0
The Syk family of kinases, consisting of ZAP-70 and Syk, play essentialroles in a variety of immune and non-immune cells. This family ofkinases is characterized by the presence of two adjacent SH2 domainswhich mediate their localization to the membrane through receptorencoded tyrosine phosphorylated motifs. While these two kinases sharemany structural and functional features, the more ubiquitous nature ofSyk has suggested that this kinase may accommodate a greater variety ofmotifs to mediate its function. We present the crystal structure of the tan-dem SH2 domain of Syk complexed with a dually phosphorylated ITAMpeptide. The structure was solved by multiple isomorphous replacementat 3.0 AÊ resolution. The asymmetric unit comprises six copies of theliganded protein, revealing a surprising ¯exibility in the relative orien-tation of the two SH2 domains. The C-terminal phosphotyrosine-bindingsite is very different from the equivalent region of ZAP-70, suggestingthat in contrast to ZAP-70, the two SH2 domains of Syk can function asindependent units. The conformational ¯exibility and structural indepen-dence of the SH2 modules of Syk likely provides the molecular basis forthe more ubiquitous involvement of Syk in a variety of signal transduc-tion pathways.
# 1998 Academic Press
Keywords: X-ray crystal structure; Syk kinase; SH2 domains; ITAM
*Corresponding authorIntroduction
ZAP-70 and Syk comprise a family of hemato-poietic cell speci®c protein tyrosine kinases (PTKs)that are required for antigen and antibody receptorfunction (reviewed by Chan & Shaw, 1995; Wange& Samelson, 1996; Reth & Wienands, 1997;Kurosaki, 1997). While ZAP-70 is expressed in Tand natural killer cells, Syk is expressed in B cells,mast cells, polymorphonuclear leukocytes, plate-lets, macrophages, and immature T cells. Theabsence of either ZAP-70 or Syk results in arrested
ding author:
unoreceptor tyrosine-in tyrosine kinase;Src homology 2;
., root mean square;ement; NCS, non-
T and B cell development and in functional defectsof a variety of immune receptors including the Tcell antigen receptor (TCR), the B cell antigenreceptor (BCR), and the receptors for IgG and IgE(FcgRI and FceRI; Costello et al., 1996; Zhang et al.,1996; Crowley et al., 1997; and reviewed by Cheng& Chan, 1997 and by Kurosaki, 1997). In addition,this family of PTKs has been implicated in activat-ing NK cells (Brumbaugh et al., 1997; Lanier et al.,1998) and signaling by non-immune receptors suchas G-protein coupled and integrin receptors (Wanet al., 1996; Gao et al., 1997; Poole et al., 1997). Thesignaling components of the antigen and Ig recep-tors contain sequence motifs known as ITAMs (forImmunoreceptor Tyrosine-based Activation Motifs)which have the consensus sequence YxxL/I-x7/8-YxxL/I (Reth, 1989). Phosphorylation of both tyro-sine residues within the ITAM by the Src-familyPTKs is required for ef®cient interaction of ZAP-70and Syk with the receptor subunits and for recep-tor function (Iwashima et al., 1994; Isakov et al.,
# 1998 Academic Press

524 Crystal Structure of the ITAM-bound SH2 Domains of Syk
1995; Bu et al., 1995; Chen et al., 1996; Kurosakiet al., 1995).
The Syk family of PTKs is characterized by adomain structure consisting of two N-terminal Src-homology 2 (SH2) domains and a C-terminalkinase domain separated from the SH2 domains bya linker or hinge region (reviewed by Chan &Shaw, 1995). The localization of ZAP-70 and Syk tothe receptor is mediated through the high af®nityinteraction between the SH2 domains of ZAP-70and Syk with the ITAM phosphorylated on bothtyrosine residues (Isakov et al., 1995; Bu et al., 1995;Chen et al., 1996). The inability of either tyrosineresidue within the ITAM to be phosphorylated ormutation of either SH2 domain within ZAP-70 orSyk decreases the avidity of this interaction by>100-fold and results in a non-functional antigenreceptor (Kong et al., 1995; Kurosaki et al., 1995;Isakov et al., 1995; Bu et al., 1995). The co-localiz-ation of ZAP-70 and Syk to the receptor complex,in part, permits the membrane-localized Src-PTKsto contribute to the phosphorylation and enzy-matic activation of ZAP-70 and Syk (Chan et al.,1995; Kurosaki et al., 1995; El-Hillal et al., 1997).
While ZAP-70 and Syk have similar structuresand play overlapping roles in T cell biology, thesetwo PTKs also exhibit many different character-istics. First, the Syk kinase domain is �100-foldmore active than ZAP-70 (Latour et al., 1996).Second, the enzymatic activation of ZAP-70 is pri-marily dependent upon the trans-phosphorylationby Src-PTKs (reviewed by Chan & Shaw, 1995). Incontrast, the enzymatic activity of Syk can be aug-mented both by Src-PTKs and by the binding ofdually phosphorylated ITAM peptides (Rivera &Brugge, 1995; Rowley et al., 1995; Kurosaki et al.,1995; El-Hillal et al., 1997). Third, expression of Sykis more ubiquitous amongst hematopoietic cellsand Syk has been shown to be activated by notonly a large variety of immune response receptors,but also by cytokines, integrins, thrombin and Gprotein-coupled receptors (Minami et al., 1995;Corey et al., 1994; Wan et al., 1996; Taniguchi et al.,1993; Clark et al., 1994). Fourth, recent evidencehas indicated that activation of Syk by non-immune response receptors such as integrin and Gprotein-coupled receptors may require only onefunctional SH2 domain for activity and thereforemay not involve a classical ITAM/Syk interaction(Gao et al., 1997; Wan et al., 1996).
Although the distinct patterns of expression ofZAP-70 and Syk may explain their selective invol-vement in signal transduction pathways, differ-ences in ITAM or phosphotyrosine-recognition andbinding speci®city by the SH2 domains of theseproteins may also account for their differences infunction. Recently, the X-ray crystal structure ofthe liganded tandem SH2 domain of ZAP-70 wasdetermined, providing the structural basis for theobligatory requirement of ZAP-70 for dually phos-phorylated ITAMs and insights into ITAM recog-nition by this protein (Hatada et al., 1995). Wepresent here the structure of the tandem SH2
domain of Syk bound to a dually phosphorylatedITAM. In contrast to ZAP-70, where one of the twophosphotyrosine binding pockets is shared by bothSH2 domains, this binding pocket in Syk is self-contained within the N-terminal SH2 domain,suggesting that the N and C-terminal SH2 domainscan function independently. In addition, the twoSH2 domains of Syk display a remarkable ¯exi-bility in their relative orientation, suggesting thatSyk may accommodate a greater variety of spacingsequences between the ITAM phosphotyrosines aswell as singly phosphorylated non-classical ITAMligands. Hence, this structure provides a structuralbasis for the broader spectrum of recognition pro-cesses mediated by the Syk-PTK and addresses theissue of speci®city in the recruitment of Syk toimmune and non-immune receptors.
Results and Discussion
Structure determination
The crystal structure of the Syk-tandem-SH2domain complexed with a dually phosphorylatedITAM peptide, derived from the CD3e chain of theT cell receptor, was determined by multiple iso-morphous replacement and anomalous scatteringto a resolution of 3.0 AÊ (Figure 1 and Materialsand Methods). The asymmetric unit contains sixcopies of the bound tandem-SH2 domain (Figure 2),which were built independently (details inMaterials and Methods). Therefore, this structurereports six independent views of the proteinbound to its ligand. Re®nement of the structureresulted in a ®nal R value of 22.6%, and a free Rvalue of 31.7% at 3.0 AÊ resolution (Table 1).
Conformational flexibility in the architecture ofthe tandem SH2 domain of Syk
The overall topology of the Syk tandem-SH2domain is similar to its counterpart in the ZAP-70kinase (Figures 3 and 4; Hatada et al., 1995) andvery different from that of the tyrosine phospha-tase SHP-2, a protein which also contains tandemSH2 domains (Eck et al., 1996; Hof et al., 1998; anddetails in `The SH2-SH2 interface' section). It con-sists of two canonical SH2 domains, the N-terminalSH2 domain (Syk-N, residues 15 to 109), and theC-terminal SH2 domain (Syk-C, residues 168 to261), connected by an intervening region, whichtogether with the SH2 domains forms a Y-shapedmolecule. Both SH2 domains are very similar infold to other SH2 domains and consist of a largeb-sheet ¯anked by two a-helices (Kuriyan &Cowburn, 1993). Both Syk-N and Syk-C superim-pose with the Src-SH2 domain (Waksman et al.,1993) with a root-mean-square (r.m.s.) deviation inCa atoms of 1.8 AÊ , or with the N or C-terminalSH2 domains of ZAP-70 (Hatada et al., 1995) withr.m.s. deviations of 0.6 and 0.9 AÊ , respectively. Theinter-SH2 region in Syk is similar in topology andstructure to that of ZAP-70 (r.m.s. deviation in Ca

Figure 1. Representative regions of the averaged exper-imental electron density contoured at 1.2s. A, Theregion encompasses the C-terminal phosphotyrosine-binding site and shows continuous and unambiguouselectron density for all elements of the phosphotyrosine-binding site of Syk-N. Helix aA, the peptide's C-term-inal phosphotyrosine (pYc), as well as residues Arg-bB5,Arg-aA2, His-bD3, and His-bD4 in Syk-N are indicated.B, Same region as in A, rotated by 90� about the axis ofthe aA helix, illustrating the proximity of Syk-C. Resi-dues in the C and N-terminal SH2 domains are distin-guished by pre®xes C- and N-, respectively. Cleardensity is observed for the elements of the phosphotyro-sine-binding site contributed by Syk-N, while Lys-bF1,the only residue of Syk-C participating in the coordi-nation of the phosphotyrosine, is poorly de®ned. Unam-biguous density for this side-chain was only observed inomit maps and in only three of the six molecules. TheFigure was generated using the program O (Jones et al.,1991).
Crystal Structure of the ITAM-bound SH2 Domains of Syk 525
atoms of 1.4 AÊ ) and consists of three a-helices, twoof them (aC and aD0) forming a helical coiled coil(Hatada et al., 1995).
The presence of six molecules in the crystallo-graphic asymmetric unit provided six different``snapshots'' of the structure that revealed a sub-stantial variability in the relative orientation of the
two SH2 domains. Figure 5 presents the superposi-tion of molecules 1 through 6 with respect to Syk-N. In this view, it is evident that Syk-C can under-go major conformational re-arrangements in itsposition relative to Syk-N. The conformationalchange can be described by a translation of Syk-Calong the direction indicated by the arrow inFigure 5, accompanied by a rotation about the axisof the Syk-C aB helix. The two extreme orien-tations of Syk-C (represented in green and blue inFigure 5) are related by a rotation of 18� and atranslation of 2.0 AÊ , with the other four orien-tations representing intermediate states. The con-formational change does not affect the total buriedsolvent-accessible area at the interface betweenSyk-N and Syk-C which remains constant (about910 AÊ 2) across the six molecules.
Changes in peptide conformation mirror thevariability in the relative orientation of the twoSH2 domains. These correspond to an adjustmentof the dihedral angle de®ned by the Ca atoms ofresidues pYN � 6 through pYN � 9 from 111.9� to136.5� (with pYN indicating the N-terminal phos-photyrosine in the peptide). The phosphorousatoms in the N-terminal phosphotyrosines of thetwo extreme conformations of Figure 5 are 7.2 AÊ
apart, illustrating the magnitude of the re-arrange-ment undergone by the SH2 domains.
Finally, the change in the relative orientation ofthe SH2 domains extends into the inter-SH2domain region (Figure 5). Moreover, electron den-sity for this region was poorly de®ned and inter-pretable only where crystal packing contacts werepresent, suggesting an inherent structural ¯exibilityof this part of the structure.
General features of ITAM binding
The peptide ligand stretches across the C and N-terminal SH2 domains along the surface oppositeto the inter-SH2 region, or, to use the Y-metaphor,across the branches of the fork (Figure 4). The pro-tein's total solvent accessible surface area buriedupon binding is 1035 AÊ 2 averaged across the sixmolecules.
The peptide binds to the tandem SH2 domainhead-to-tail, i.e. the N-terminal pYxxL/I-motifbinds to the C-terminal SH2 domain and theC-terminal pYxxL/I motif binds to the N-terminalSH2 domain. The peptide conformation in the twomotifs is extended while residues in the inter-motifregion of the peptide form a nearly complete turnof an a-helix. These features are similar to thosereported for ZAP-70 (Hatada et al., 1995) and there-fore establish these as general properties for ITAMbinding and recognition by tandem SH2 domainsof the Syk family of kinases.
Each SH2 domain of Syk contains two pocketswhich bind a pYxxL/I motif of the ITAM peptide.The mode of binding in each SH2 domainresembles closely that of the SH2 domains of theSrc family of proteins: one pocket, positivelycharged, accommodates the phosphotyrosine,
Figure 2. Molecular content of theunit cell. The view is approximatelyalong the 3-fold axis. Each of thesix molecules are color-coded dif-ferently. For clarity, the peptide hasnot been included. This ®gure wasgenerated using O (Jones & Thirup,1986; Jones et al., 1991).
526 Crystal Structure of the ITAM-bound SH2 Domains of Syk
while the other, hydrophobic in nature, buries thenon-polar Ile/Leu residue at the �3 positionC-terminal to the phosphotyrosine (Waksman et al.,1992, 1993; Eck et al., 1993). However, while inZAP-70, the C-terminal phosphotyrosine binding
Table 1. Data collection and re®nement statistics
Data collectionNumber ofreflections
Data set Radiation Resolution Total Uniqu
Native1.08 AÊ , BL7-1,
SSRL 30-3.0 AÊ 202,021 40,67K2PtCl4-1 CuKa, Raxis 30-3.8 AÊ 77,547 18,19K2PtCl4-2 CuKa, Raxis 30-4.0 AÊ 31,079 13,65
RefinementResolution Number of Complete
(AÊ ) reflectionsf (%)f
30.0-3.0 40,397 85.6 (72Number of non hydrogen atoms 12,225Number of residues 1530Number of waters 0Bond length (AÊ )h 0.016Bond angles (�) 1.9RMS deviations between NCS-related atoms
protein main ch. 0.10 AÊ -0.38 AÊ
peptide main ch. 0.19 AÊ -0.47 AÊ
RMSD in B factor for bonded atoms (AÊ 2)main chain 1.9side chain 2.7
Average B (AÊ 2)main chain 43side chain 46
a Completeness for I/s(I) > 2.0, high resolution shell (3.1-3.0 AÊ ) ib Rsym � �jI ÿ hIij/�I, where I � observed intensity, and hIi � a
re¯ections.c Riso � �jjFPHj ÿ jFPjj/�jFPj, where FP � protein structure facto
tude.d Phasing power � RMS (jFHj/e), where FH � heavy-atom structue FOM � Figure of Mert.f Numbers re¯ect the ``Working Set'' of re¯ections at F/s(F) > 2.g Rfree was calculated on the basis of 5% of the total number of rh Bond length and angle parameters from Engh & Huber (1991).
site is formed by elements of both the C andN-terminal SH2 domains, this site in Syk is self-contained within the N-terminal SH2 domain. InZAP-70, the solvent accessible surface area buriedupon binding of the phosphotyrosine amounts to
PhasingCompletenessa Phasingd
e (%) Rsym (%)b Riso (%)c N Sites power
0 89.8 (74.1) 5.102 84.7 (66.1) 8.00 23.8 6 2.29 72.0 (48.0) 6.40 20.8 6 2.5
FOM (SIRAS) 30-4 AÊ e
0.49
ness R-Factor Rfree
(%) (%)g
.2) 22.6 31.7
n parentheses.verage intensity from multiple observations of symmetry-related
r amplitude, FPH � heavy-atom derivative structure factor ampli-
re factor amplitude, e � residual lack of closure.
0, high resolution shell in parentheses (3.14 to 3.0 AÊ ).e¯ections omitted from the re®nement.

Figure 3. SH2 domain sequences and de®nition of the residue notation. A, Alignment of the sequences of the N andC-terminal SH2 domains of Syk, ZAP-70, and the SH2 domain of Src, based on the secondary structure de®nitionsused by Eck et al. (1993), Waksman et al. (1993) and Hatada et al. (1995). B, Alignment of the sequences of the inter-SH2 domains of Syk and ZAP-70, based on the secondary structure de®nitions used by Hatada et al. (1995). Second-ary structural elements are boxed and shown schematically as arrows (b-strands) and cylinders (a-helices) below theboxes. The notation for these elements is shown at the bottom. Functionally important residues are highlighted bythick vertical bars above the sequence and their relative position in the secondary structural elements is indicated.Residue sequence numbers for Syk are indicated above the sequences, with ®lled and open numbers referring to Syk-N and Syk-C, respectively.
Crystal Structure of the ITAM-bound SH2 Domains of Syk 527
285 AÊ 2, of which 50 AÊ 2 (17.5%) is contributed byresidues of the C-terminal SH2 domain (Hatadaet al., 1995). In Syk, the corresponding solventaccessible surface area is on average smaller(245 AÊ 2). More remarkably, in three of the six mol-ecules in the asymmetric unit, only 5% of the totalburied surface area involved in phosphotyrosinebinding is contributed by the C-terminal SH2domain. In the other three molecules, this contri-bution is substantially increased (to 17.9%) by newcontacts formed by a single side-chain, that of Lys-bF1. However, electron density for this side-chainwas consistently missing in maps based oneither experimental or model-combined phases(Figure 1B). Thus, building relied solely on simu-lated annealing omit maps (Hodel et al., 1992) ofthe corresponding regions, which displayed elec-tron density for Lys-bF1 in only three of the sixcomplexes. In contrast, all other elements of thephosphotyrosine binding site were clearly de®nedin all maps (experimental included) and in all mol-
ecules (Figure 1, A and B). These observationsstrongly suggest that Lys-bF1 is at least partiallydisordered and therefore may not contribute sig-ni®cantly to binding. Thus, the molecular basis forITAM recognition by Syk differs distinctly fromZAP-70, and suggests that the two SH2 domains ofSyk may function as independent units.
Binding of the N-terminal pYNEPI motif by theC-terminal SH2 domain
All contacts with the amino-terminal portion thepeptide (positions pYN-2 to pYN � 3) are providedby Syk-C. Four arginine residues are in close proxi-mity at the phosphotyrosine-binding site. Three ofthese residues, bB7, aA2 and bB5, contribute to thenetwork of hydrogen bonds coordinating the phos-phate group of the phosphotyrosyl residue(Figures 6C and 8A). Arg-aA2 also makes a weakamino-aromatic interaction with the phosphotyro-sine ring, and is within hydrogen bonding distance

Figure 4. Stereo ribbon diagram of the tandem SH2 domain of Syk in a complex with the CD3e-ITAM peptide. Syk-N is depicted in blue, Syk-C in green and the inter-SH2 region in brown. The ITAM peptide is shown in ball-and-stick representation. Secondary structural elements are labeled according to the notation adopted in Figure 3. TheFigure was generated using the programs MOLSCRIPT (Kraulis, 1991) and RASTER3D (Merritt & Murphy, 1994).
528 Crystal Structure of the ITAM-bound SH2 Domains of Syk
of Asp(pYN-1 ) of the peptide. A fourth arginineresidue (bD6) closes up the phosphotyrosine bind-ing site by forming a hydrophobic platform for thering of the phosphotyrosine. Most of these struc-tural features have been observed in other SH2domains, including the C-terminal SH2 domain ofZAP-70 (Figure 6D; Waksman et al., 1993; Eck et al.,1993; Hatada et al., 1995). However, the C-terminalSH2 domain of Syk differs from that of ZAP-70 intwo respects: (1) a larger number of positivecharges is observed at the phosphotyrosine bindingsite of Syk; (2) Arg-bB7 in ZAP-70 does not form asalt bridge with the phosphate, but instead neutral-izes Glu-BC2 (Figure 6D). As a result, the net posi-tive charge in the phosphotyrosine binding site islarger in Syk than in ZAP-70. These structural fea-tures strongly suggest that the C-terminal SH2domains of Syk and ZAP-70 may differ substan-tially in their binding properties. Such a suggestionis consistent with the reported observation that, in
Figure 5. Conformational ¯exibility of the Syk tandem SHSyk tandem SH2 domain superimposed with respect todown the axis of the C-terminal SH2 aB helix. The arrowthe text). The Figure was generated using the programsMurphy, 1994).
contrast to ZAP-70, the C-terminal SH2 domain ofSyk retains substantial binding af®nity for tyrosyl-phosphorylated targets (Shiue et al., 1995).
The side-chain of Glu(pYN � 1) in the peptidemakes extensive hydrophobic contacts, observedbetween the aliphatic carbon atoms of its side-chain and the protein, while the terminal oxygenatoms are within 5.3 AÊ of the positively chargedtip of Lys-bD1. Pro(pYN � 2) is mostly solventexposed. Finally, the side-chain of Ile(pYN � 3) isalmost completely buried in the hydrophobic bind-ing pocket of the C-terminal SH2 domain andcontacts are typical of those observed in otherSH2 domain-peptide complexes (see details inFigure 8A; Waksman et al., 1993; Eck et al., 1993).
Binding of the C-terminal pYCSGL motif to theN-terminal SH2 domain
As mentioned above, a striking feature of theC-terminal phosphotyrosine (pYC) binding pocket
2 domain. Stereo ribbon diagram of molecules 1 to 6 of thethe N-terminal SH2 domain. The view is approximatelyindicates the direction of the translational component (seeMOLSCRIPT (Kraulis, 1991) and RASTER3D (Merritt &
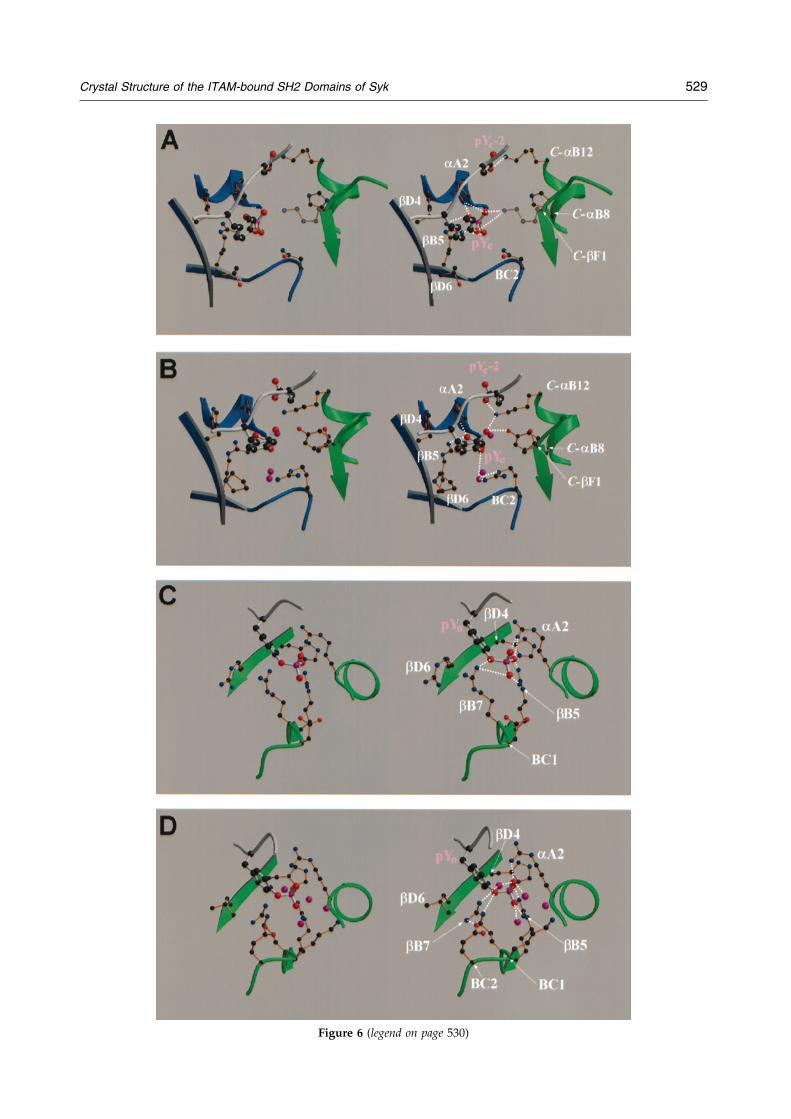
Figure 6 (legend on page 530)
Crystal Structure of the ITAM-bound SH2 Domains of Syk 529

Figure 7. Stereo diagram of the interface between the SH2 domains of Syk. Secondary structural elements in the Nand C-terminal SH2 domain are shown in blue and green, respectively. Side-chains are shown in ball-and-stick rep-resentation. Broken lines indicate hydrogen bonds. The peptide is shown in transparent rendering for reference. TheFigure was generated using the programs MOLSCRIPT (Kraulis, 1991) and RASTER3D (Merritt & Murphy, 1994).
530 Crystal Structure of the ITAM-bound SH2 Domains of Syk
of Syk is the absence of signi®cant interactions con-tributed by the C-terminal SH2 domain in three ofthe six complexes in the asymmetric unit. In theother three molecules, only one residue in thisdomain, Lys-bF1, seems to participate in thecoordination of the phosphotyrosine. However,Lys-bF1 is partially disordered and, as discussedabove, may not contribute signi®cantly to binding.
The C-terminal phosphotyrosine forms hydrogenbonding contacts with arginine residues aA2 andbB5 through its phosphate group (Figures 6A and8C). Arg-aA2 in addition makes a weak amino-aro-matic interaction with the phenyl ring and formsa hydrogen bond with the carbonyl oxygen ofLeu(pYC-1). Both features occur similarly at theN-terminal pYNEPI motif. Ser(pYC � 1) interactsprimarily with the side-chain of Tyr-bD5 and themain-chain atoms of His-bD4, while Gly(pYC � 2)only makes sparse contacts. Leu(pYC � 3) insertsdeeply into the hydrophobic binding pocket ofSyk-N. Finally, Asn(pYC � 4) is the last ligand resi-due that appears to be stabilized by interactionswith the protein (Figure 8C).
It is interesting to note that the residue of Syk-Nfacing the �2 position of the peptide is Glu-bD01.The equivalent residue in Src kinases is Arg-bD01which is responsible for a strong selectivity for Gluat the �2 position of the peptide (Waksman et al.,1993; Eck et al., 1993; Songyang et al., 1993). Byanalogy, one would predict a strong selectivity of
Figure 6. Stereo diagrams of the interactions at the phopocket of Syk-N (A), ZAP-N (B), Syk-C (C), and ZAP-C (D)big spheres for peptide atoms and small spheres for proteand green ribbons representing the N and C-terminal SHbonding interactions. In B and D, water molecules are reprin transparent rendering to indicate its disordered state in tated using the programs MOLSCRIPT (Kraulis, 1991) and R
Syk-N for a basic residue at the �2 position in thepeptide.
Binding of theinter-motif-(pYN�4)RKGQRDL(pYCÿ1)-
Residues of the inter-motif account for only22.1% (on average) of the total surface area of thepeptide buried upon binding. Very few contactsare observed between the protein and peptide resi-dues between Arg(pYN � 4) and Arg(pYC ÿ 3) (seeFigure 8B for peptide residue notation). In contrast,Asp(pYC ÿ 2) makes extensive contacts with theprotein surface and is the only residue in the inter-motif that interacts with residues in both SH2domains, namely Arg-aA2 of Syk-N and Lys-aB12of Syk-C. The side-chain of Leu(pYC ÿ 1) is com-pletely solvent exposed.
The SH2-SH2 interface
The SH2-SH2 interface is polar in nature. Thereare two main regions of inter-domain contacts; inthe ®rst region, the BC-loop in Syk-N faces thebF-strand, the FB-loop and the N-terminal tip ofthe aB helix of Syk-C; in the second region resi-dues in the N-terminal end of the aA helix andin the AA loop of Syk-N make contact with resi-dues in the C-terminal end of the aB helix ofSyk-C (Figure 7).
sphotyrosine binding sites. The phosphotyrosine binding. Residues are shown in ball-and-stick representation within atoms. Secondary structural elements are shown blue2 domains, respectively. Broken lines indicate hydrogenesented by spheres in magenta. In A, Lys-bF1 is depictedhree of the six molecules (see text). The Figure was gener-ASTER3D (Merritt & Murphy, 1994).
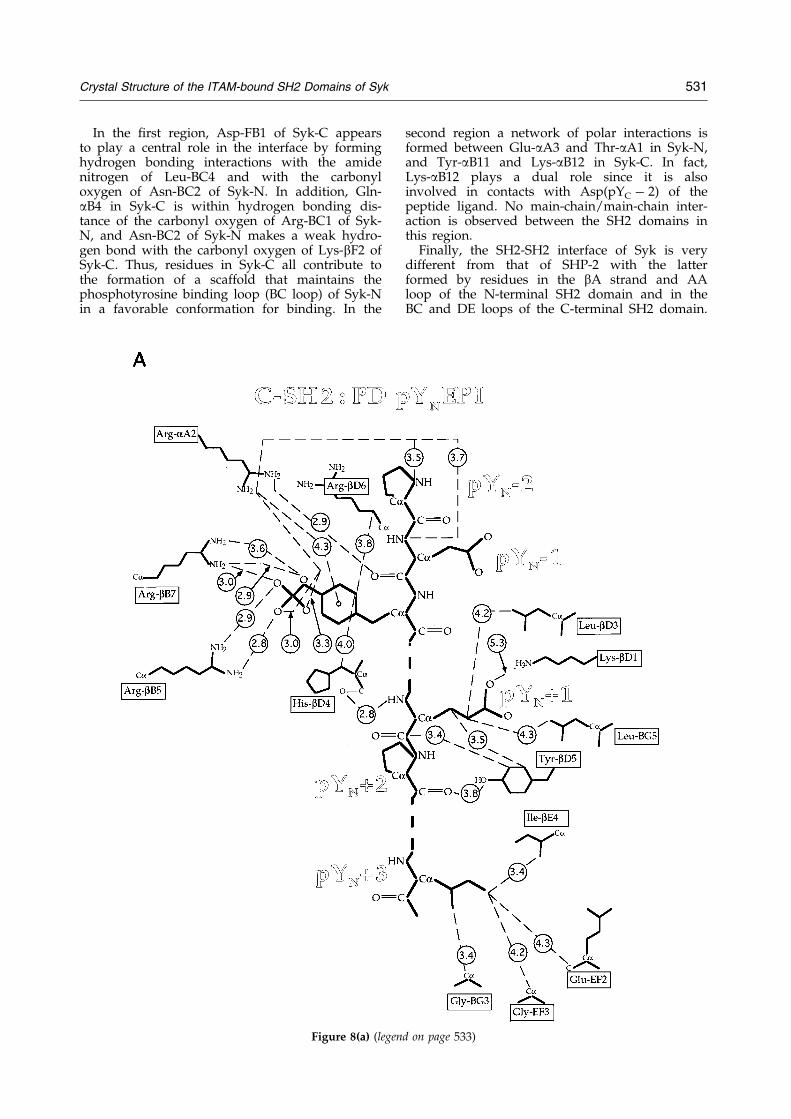
Crystal Structure of the ITAM-bound SH2 Domains of Syk 531
In the ®rst region, Asp-FB1 of Syk-C appearsto play a central role in the interface by forminghydrogen bonding interactions with the amidenitrogen of Leu-BC4 and with the carbonyloxygen of Asn-BC2 of Syk-N. In addition, Gln-aB4 in Syk-C is within hydrogen bonding dis-tance of the carbonyl oxygen of Arg-BC1 of Syk-N, and Asn-BC2 of Syk-N makes a weak hydro-gen bond with the carbonyl oxygen of Lys-bF2 ofSyk-C. Thus, residues in Syk-C all contribute tothe formation of a scaffold that maintains thephosphotyrosine binding loop (BC loop) of Syk-Nin a favorable conformation for binding. In the
Figure 8(a) (lege
second region a network of polar interactions isformed between Glu-aA3 and Thr-aA1 in Syk-N,and Tyr-aB11 and Lys-aB12 in Syk-C. In fact,Lys-aB12 plays a dual role since it is alsoinvolved in contacts with Asp(pYC ÿ 2) of thepeptide ligand. No main-chain/main-chain inter-action is observed between the SH2 domains inthis region.
Finally, the SH2-SH2 interface of Syk is verydifferent from that of SHP-2 with the latterformed by residues in the bA strand and AAloop of the N-terminal SH2 domain and in theBC and DE loops of the C-terminal SH2 domain.
nd on page 533)
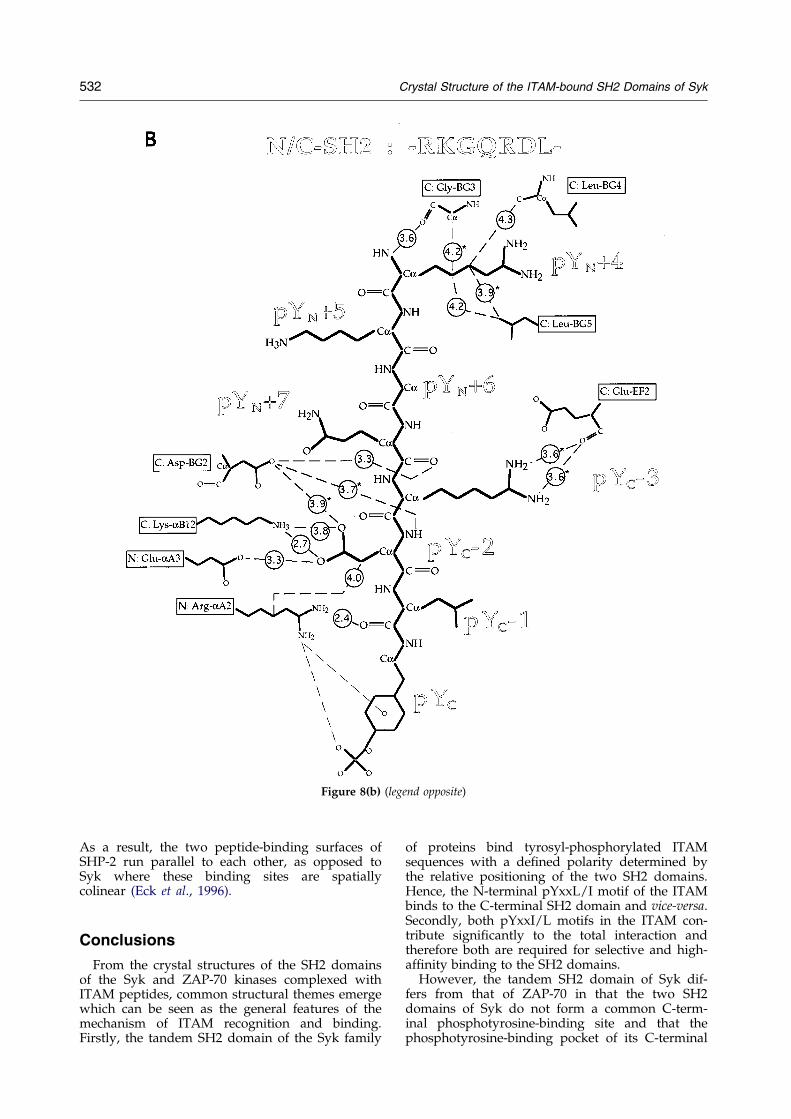
Figure 8(b) (legend opposite)
532 Crystal Structure of the ITAM-bound SH2 Domains of Syk
As a result, the two peptide-binding surfaces ofSHP-2 run parallel to each other, as opposed toSyk where these binding sites are spatiallycolinear (Eck et al., 1996).
Conclusions
From the crystal structures of the SH2 domainsof the Syk and ZAP-70 kinases complexed withITAM peptides, common structural themes emergewhich can be seen as the general features of themechanism of ITAM recognition and binding.Firstly, the tandem SH2 domain of the Syk family
of proteins bind tyrosyl-phosphorylated ITAMsequences with a de®ned polarity determined bythe relative positioning of the two SH2 domains.Hence, the N-terminal pYxxL/I motif of the ITAMbinds to the C-terminal SH2 domain and vice-versa.Secondly, both pYxxI/L motifs in the ITAM con-tribute signi®cantly to the total interaction andtherefore both are required for selective and high-af®nity binding to the SH2 domains.
However, the tandem SH2 domain of Syk dif-fers from that of ZAP-70 in that the two SH2domains of Syk do not form a common C-term-inal phosphotyrosine-binding site and that thephosphotyrosine-binding pocket of its C-terminal

Figure 8. Schematic representation of peptide-protein contacts. Distances are given in AÊ and are averaged across thesix molecules in the asymmetric unit, except when marked with an asterisk. Peptide residues are labeled according totheir position relative to the phosphotyrosine residues, with pYN and pYC denoting the N and C-terminal phospho-tyrosines, respectively. Protein residues are labeled according to the notation of Eck et al. (1993) and Waksman et al.(1993), preceded by an N: or C: where distinction between N and C-terminal SH2 domains is necessary. The peptideis depicted in three portions A, pYN ÿ 2 through pYN � 3; B, pYN � 4 through pYC ÿ 1; and C, pYC through pYC � 4.
Crystal Structure of the ITAM-bound SH2 Domains of Syk 533
SH2 domain is characterized by a more positiveelectrostatic surface potential than its ZAP-70counterpart. These observations may relate to thefact that at least in two signal transduction path-ways, those mediated by integrins and G protein-coupled receptors, recruitment of Syk into thepathways may not necessitate the involvement ofboth SH2 domains (Gao et al., 1997; Wan et al.,1996). Whereas the C-terminal SH2 domain is notrequired for Syk activation by the aIIbb3 integrin
receptor (Gao et al., 1997), Syk-N is not requiredfor activation of the MAP kinase pathway by theGq-coupled muscarinic acetylcholine receptor(Wan et al., 1996). These observations are consist-ent with the interpretation that each individualSH2 domain can function as an independent unit,thus supporting the structural information pro-vided by the present study.
The six different conformations of the tandemSH2 domain of Syk demonstrate a remarkable

534 Crystal Structure of the ITAM-bound SH2 Domains of Syk
¯exibility in domain structure, which may enableSyk to tolerate changes in the size of the ITAM'sinter-motif sequence. Consistent with this hypoth-esis, Syk binds ITAMs derived from the FcgRIIand colony-stimulating factor receptors whichboth have inter-motif regions signi®cantly longer(by four amino acids) than the canonical ITAM(Corey et al., 1994; Chacko et al., 1996). Interest-ingly, Syk binds with higher af®nity to the FcgRIIITAM than does ZAP-70, while both Syk andZAP-70 have comparable af®nity for the CD3eITAM (Bu et al., 1995; J.W. & A.C.C., unpub-lished), suggesting that the tandem SH2 domainof ZAP-70 may not be as ¯exible as that of Syk.Due to the presence of only one molecule in theasymmetric unit, the crystal structure of theITAM-bound SH2 domains of ZAP-70 could notprovide evidence for possible variations in therelative orientation of the SH2 domains (Hatadaet al., 1995).
Together, these studies indicate that discrimi-nation in the function of Syk and ZAP-70 may beachieved independently by two mechanisms. First,tissue selective expression undoubtedly dictatesthe potential function of these two kinases. Second,the selective involvement of Syk or ZAP-70 in sig-nal transduction pathways may reside in differ-ences in binding speci®city and mode determinedby their SH2 domains. Src-homology 2 domains, assingle binding units, have evolved binding selectiv-ity, which when linked into a two-unit protein pro-vides for a uniquely selective macromoleculartemplate. Together with selectivity in expression,recruitment of SH2-containing proteins can then bemodulated in an exquisitely precise way resultingin their selective involvement in signal transduc-tion pathways.
Materials and Methods
Purification, crystallization and data collection
The tandem SH2 domain fragment of Syk (residues 7to 269) was expressed and puri®ed as described by Buet al. (1995). The pure protein was complexed withthe CD3e-chain ITAM peptide (sequence PDpYE-PIRKGQRDLpYSGLNQR, from Quality Controlled Bio-chemicals) at a ratio of 1:1.1 protein to ligand, andconcentrated to typically 35 mg/ml. Crystals weregrown at room temperature using the hanging dropvapor diffusion method (Mc Pherson, 1990). Drops of2 ml protein were mixed with 2 ml of 9 to 12% (w/v)PEG 8000, 10% (v/v) PEG 200, 0.1 M Tris-HCl pH 8.5,(Sigma) and placed over a reservoir of the same compo-sition. Crystals (0.4 mm � 0.3 mm � 0.1 mm) appearedwithin one week. Flash-frozen crystals, protected in 15%glycerol, diffracted to 2.9 AÊ in a laboratory setting (R-axis IIc image plate detector mounted on an RU-200rotating anode generator, Rigaku) and to 2.5 AÊ at thesynchrotron (beamline 7.1, Stanford Synchrotron Radi-ation Laboratory). However, scattering was strongly ani-sotropic, thus limiting the effective maximum resolutionto 3.0 AÊ at a I/s(I) ratio greater than 2.0 (see Table 1).The crystals belonged to space group P21 witha � 85.5 AÊ , b � 146.9 AÊ , c � 91.5 AÊ , and b � 97.6�, and
contained six tandem SH2/ITAM complexes per asym-metric unit. All data were processed with the DENZOand SCALEPACK software packages (Otwinowski,1993).
Structure determination
The structure was determined by multiple isomor-phous replacement with anomalous scattering. Platinumderivatized crystals were obtained by soaking nativecrystals in 2 mM K2PtCl4 for 12 and 24 hours, respect-ively. Complete diffraction data on derivatized frozencrystals could be collected to 4.0 AÊ resolution in the lab-oratory. The isomorphous difference Patterson analysisrevealed distinct peaks in the Harker section (v � 1/2)and ®ve platinum positions could be readily determinedusing the program HASSP (Terwilliger et al., 1987). Asixth position could be found by difference Fouriermethods. The Pt-derivative data showed an anomaloussignal that could be exploited for phasing to 4 AÊ in con-junction with the isomorphous differences using the pro-gram MLPHARE (CCP4 package (CCP4, 1994); seeTable I for relevant statistics). The electron density mapthat resulted from subsequent solvent ¯attening usingDM (CCP4, 1994) showed weak secondary structural fea-tures together with a well de®ned molecular envelope.
Non-crystallographic symmetry (NCS) averaging (pro-gram DM) was used to further improve the initialphases. The NCS-matrices were derived from the coordi-nates of the platinum atoms and found in agreementwith the peaks in the self-rotation function (programPOLARRFN, CCP4, 1994). In an iterative procedure ofbuilding recognizable secondary structural elements,rede®ning the molecular envelope and re®ning the NCS-matrices, the density could be improved to a level thatallowed us to manually dock SH2 domains into the elec-tron density, thus unequivocally identifying the locationof the six molecules of the asymmetric unit. NCS aver-aging was also exploited to extend experimental phasesto 3.0 AÊ (Table 1).
At a later stage of this project the program SHARP(De La Fortelle & Bricogne, 1997) became available andwas used to recalculate the experimental phases. NCS-averaging of these phases led to a much improved elec-tron density map that was very bene®cial for modelbuilding.
Model building and refinement
A poly(Ala) model for the SH2 domains of the mol-ecule best de®ned in the averaged electron density wasbuilt using the program O (Jones & Thirup, 1986; Joneset al., 1991), and subsequently re®ned, subjected to strictNCS constraints (program XPLOR, BruÈ nger, 1992a).However, as the variable orientation between the twoSH2 domains became apparent, all six pairs of SH2domains were built independently and re®ned by apply-ing NCS restraints to Syk-N and Syk-C separately. Theinter-SH2 regions (residues 118 to 163) were only poorlyde®ned and the density deteriorated upon NCS-aver-aging. Interpretable density for these regions wasobtained, but only in three of the six molecules, by com-bining the phases calculated from the partial model withthe unaveraged experimental phases (program SIGMAA,CCP4 package, 1994). The peptide could be built in allsix molecules based on both experimental and phase-combined maps.

Crystal Structure of the ITAM-bound SH2 Domains of Syk 535
Progress in the re®nement that included conjugategradient minimization as well as simulated annealing inboth cartesian and torsional angle spaces (BruÈ nger et al.,1990; Rice & BruÈ nger, 1994) was assessed by monitoringthe free R-factor (BruÈ nger, 1992b; see Table 1). NCS-matrix sets for nine different structural groups werede®ned, i.e. residues 9 to 117 (Syk-N), 167 to 262 (Syk-C), 118 to 136 (helix aC in the inter-SH2 region), 137 to140 (the loop connecting the aC and aD helices), 141 to151 (helix aD in the inter-SH2 region), 152 to 166 (helixaD0 in the inter-SH2 region), residues pYN ÿ 2 topYN � 3, residues pYC ÿ 2 to pYC � 4, and residuespYN � 4 to pYC ÿ 3 in the peptide ligand (see notation inFigure 8). The NCS restraints were such that the r.m.s.deviations between main-chain atoms in equivalentrestrained regions were between 0.10 and 0.38 AÊ in theprotein and between 0.19 and 0.47 AÊ in the peptideligand. Individual B-factor re®nement was used, result-ing in average temperature factors of 43 AÊ 2 for main-chain atoms and 46 AÊ 2 for side-chain atoms, and r.m.s.deviations for bonded main-chain or side-chain atoms of1.9 and 2.7 AÊ 2, respectively. After bulk solvent correc-tion, the re®nement converged to a ®nal R-factor of 22.6% with an R-free of 31.7 % (30 to 3.0 AÊ resolution range;jFj/s(jFj) > 2.0) with good stereochemistry (Table 1). Themodel includes residues 9 to 262 in all six moleculeswith an interruption in the inter-SH2 region in three mol-ecules between residues 118 and 151. Electron densitywas not clearly interpretable for a few residues (Ser9,Lys124, Lys133, Lys222 and Lys261) which were built asalanine. The model does not contain water molecules.All residues are in the allowed region of the Ramachan-dran plot (Ramachandran & Sasisekharan, 1968). Rel-evant statistics for the re®nement are summarized inTable 1. The coordinates of the structure of the tandemSH2 domain of Syk bound to the dually phosphorylatedCD3e-ITAM peptide have been deposited (PDB entrycode 1A81).
Acknowledgments
We thank Ian Wilson and Jack Johnson (ScrippsResearch Institute) for advice on NCS averaging, C.S.Ricard for advice on protein puri®cation, S. Korolev andA.B. Herr for help in data collection, M. Hatada (ARIADPharmaceuticals) for providing the ZAP-70 tandem SH2domain coordinates, the staff of Beamline 7.1 at SSRL forassistance during data collection, and F.S. Mathews forcomments on the manuscript. This work was supportedby P®zer Inc. (G.W.), by funds from the WashingtonUniversity School of Medicine (G.W.), and by NationalInstitute of Health grant CA71516 (C.C.A.). J.W. wassupported by National Institute of Health training grant5T32DK07126.
References
Brumbaugh, K. M., Binstadt, B. A., Billadau, D. D.,Schoon, R. A., Dick, C. J., Ten, R. M. & Leibson, P. J.(1997). Functional role for Syk tyrosine kinase innatural killer cell-mediated natural cytotoxicity.J. Exp. Med. 186, 1965±1974.
BruÈ nger, A. T. (1992a). X-PLOR (Version 3. 1) Manual,The Howard Hughes Medical Institute and Depart-ment of Molecular Biophysics and Biochemistry,
Yale University, New Haven, 260 Whitney Avenue,CT 06511.
BruÈ nger, A. T. (1992b). The free R value: a novel statisti-cal quantity for assessing the accuracy of crystalstructures. Nature, 355, 472±474.
BruÈ nger, A. T., Krukowski, A. & Erickson, J. W. (1990).Slow-cooling protocols for crystallographic re®ne-ment by simulated annealing. Acta Crystallog. sect.A, 46, 585±593.
Bu, J.-Y., Shaw, A. S. & Chan, A. C. (1995). Analysis ofthe interaction of ZAP-70 and syk protein-tyrosinekinases with the T-cell antigen receptor by plasmonresonance. Proc. Natl Acad. Sci. USA, 92, 5106±5110.
CCP4, (1994). Collaborative computational project num-ber 4. The CCP4 suite of programs for protein crys-tallography. Acta Crystallog. sect. D, 50, 760±763.
Chacko, G. W., Brandt, J. T., Coggeshall, K. M. &Anderson, C. L. (1996). Phosphoinositide 3-kinaseand p72syk noncovalently associate with the lowaf®nity Fcg receptor on human platelets through animmunoreceptor tyrosine-based activation motif.J. Biol. Chem. 271, 10775±10781.
Chan, A. C. & Shaw, A. S. (1995). Regulation of antigenreceptor signal transduction by protein tyrosinekinases. Curr. Opin. Immunol. 8, 394±401.
Chan, A. C., Dalton, M., Johnson, R., Kong, G.-H.,Wang, T., Thoma, R. & Kurosaki, T. (1995). Acti-vation of ZAP-70 kinase activity by phosphoryl-ation of tyrosine 493 is required for lymphocyteantigen receptor function. EMBO J. 14, 2499±2508.
Chen, T., Repetto, B., Chizzonite, R., Pullar, C.,Burghardt, C., Dharm, E., Zhao, Z., Carroll, R.,Nunes, P., Basu, M., Danho, W., Visnick, M.,Kochan, J., Waugh, D. & Gil®llan, A. M. (1996).Interaction of phosphorylated FceRIg immunoglo-bulin receptor tyrosine activation motif-based pep-tides with dual and single SH2 domains of p72syk.Assessment of binding parameters and real timebinding kinetics. J. Biol. Chem. 271, 25308±25315.
Cheng, A. M. & Chan, A. C. (1997). PTKs in thymocytedevelopment. Curr. Opin. Immunol. 8, 394±401.
Clark, E. A., Shatiff, S. J., Ginsberg, M. H., Bolen, J. &Brugge, J. S. (1994). Regulation of the protein tyro-sine kinase pp72syk by platelet agonists and theintegrin aIIbb3. J. Biol. Chem. 269, 28859±28864.
Corey, S. J., Burkhardt, A. L., Bolen, J. B., Geahlen, R. B.,Tkatch, L. S. & Tweardy, D. J. (1994). Granulocytecolony-stimulating factor receptor signalinginvolves the formation of a three-component com-plex with Lyn and Syk protein-tyrosine kinases.Proc. Natl Acad. Sci. USA, 91, 4683±4687.
Costello, P. S., Turner, M., Walters, A. E., Cunningham,C. N., Bauer, P. H., Downward, J. & Tybulewicz,V. L. J. (1996). Critical role for the tyrosine kinaseSyk in signalling through the high af®nity IgEreceptor of mast cell. Oncogene, 13, 2595±2605.
Crowley, M. T., Costello, P. S., Fitzer-Attas, C. J.,Turner, M., Meng, F., Lowell, C., Tybulewicz, V. L. J.& DeFranco, A. L. (1997). A critical role for Syk insignal transduction and phagocytosis mediated byFcg receptors on macrophages. J. Exp. Med. 186,1027±1039.
De La Fortelle, E. & Bricogne, G. (1997). Maximum-like-lihood heavy-atom parameter re®nement for mul-tiple isomorphous replacement and multiwave-length anomalous diffraction methods. MethodsEnzymol. 276, 472±493.
Eck, M. J., Shoelson, S. E. & Harrison, S. C. (1993). Rec-ognition of a high af®nity phosphotyrosyl peptide

536 Crystal Structure of the ITAM-bound SH2 Domains of Syk
by the Src homology 2 domain of p56lck. Nature,362, 87±91.
Eck, M. J., Pluskey, S., Trub, T., Harrison, S. C. &Shoelson, S. E. (1996). Spatial constraints on the rec-ognition of phosphoproteins by the tandem SH2domains of the phosphatase SH-PTP2. Nature, 379,277±280.
El-Hillal, O., Kurosaki, T., Yamamura, H., Kinet, J.-P. &Scharenberg, A. M. (1997). Syk kinase activation bya src kinase-initiated activation loop phosphoryl-ation chain reaction. Proc. Natl Acad. Sci. USA, 94,1919±1924.
Engh, R. A. & Huber, R. (1991). Accurate bond andangle parameters for X-ray protein structure re®ne-ment. Acta Crystallog. sect. A, 47, 392±400.
Gao, J., Zoller, K. E., Ginsberg, M. H., Brugge, J. S. &Shattil, S. J. (1997). Regulation of the pp72syk proteintyrosine kinase by platelet integrin aIIbb3. EMBO J.16, 6414±6425.
Hatada, M. H., Lu, X., Laird, E. R., Green, J.,Morgenstern, J. P., Lou, M., Marr, C. S., Phillips,T. B., Ram, M. K., Theriault, K., Zoller, M. J. &Karas, J. L. (1995). Molecular basis for interaction ofthe protein tyrosine kinase ZAP-70 with the T-cellreceptor. Nature, 377, 32±38.
Hodel, A., Kim, S.-H. & BruÈ nger, A. T. (1992). Modelbias in crystal structures. Acta Crystallog. sect. A, 48,851±858.
Hof, P., Pluskey, S., Dhe-Paganon, S., Eck, M. J. &Shoelson, S. E. (1998). Crystal structure of the tyro-sine phosphatase SHP-2. Cell, 92, 441±450.
Isakov, N., Wange, R. L., Burgess, W. H., Watts, J. D.,Aebersold, R. & Samelson, L. E. (1995). ZAP-70binding speci®city to T cell receptor tyrosine-basedactivation motifs: the tandem SH2 domains of ZAP-70 bind distinct tyrosine-based activation motifswith varying af®nity. J. Exp. Med. 181, 375±380.
Iwashima, M., Irving, B. A., van Oers, N. S. C., Chan,A. C. & Weiss, A. (1994). Sequential interactions ofthe TCR with two distinct cytoplasmic tyrosinekinases. Science, 263, 1136±1139.
Jones, T. A. & Thirup, S. (1986). Using known substruc-tures in protein model building and crystallogra-phy. EMBO J. 5, 819±822.
Jones, T. A., Zou, J. Y., Cowan, S. W. & Kjeldgaard, M.(1991). Improved methods for building proteinmodels in electron density maps and the location oferrors in these models. Acta Crystallog. sect. A, 47,110±119.
Kong, G.-H., Bu, J.-Y., Kurosaki, T., Shaw, A. S. & Chan,A. C. (1995). Reconstitution of Syk function by theZAP-70 protein tyrosine kinase. Immunity, 2, 485±492.
Kraulis, P. J. (1991). MOLSCRIPT: a program to repro-duce both detailed and schematic plots of proteinstructure. J. Appl. Crystallog. 24, 946±950.
Kuriyan, J. & Cowburn, D. (1993). Structures of the SH2and SH3 domains. Curr. Opin. Struct. Biol. 3, 828±837.
Kurosaki, T. (1997). Molecular mechanisms in B cellantigen receptor signaling. Curr. Opin. Immunol. 9,309±318.
Kurosaki, T., Johnson, S. A., Pao, L., Sada, K.,Yamamura, H. & Cambier, J. C. (1995). Role of theSyk autophosphorylation site and SH2 domain in Bcell antigen receptor signaling. J. Exp. Med. 182,1815±1823.
Lanier, L. L., Corliss, B. C., Wu, J., Leong, C. & Phillips,J. H. (1998). Immunoreceptor DAP12 bearing a tyro-
sine-based activation motif is involved in activatingNK cells. Nature, 391, 703±707.
Latour, S., Chow, L. M. L. & Veillette, A. (1996). Differ-ential intrinsic enzymatic activity of Syk and Zap-70protein-tyrosine kinases. J. Biol. Chem. 271, 22782±22790.
McPherson, A. (1990). Current approaches to macromol-ecular crystallization. Eur. J. Biochem. 189, 1±23.
Merritt, E. A. & Murphy, M. E. P. (1994). Raster3D Ver-sion 2.0 ± A program for photorealistic moleculargraphics. Acta Crystallog. sect. D, 50, 869±873.
Minami, Y., Nakagawa, Y., Kawahara, A., Miyazaki,T., Sada, K., Yamamura, H. & Taniguchi, T.(1995). Protein tyrosine kinase Syk is associatedwith and activated by the IL-2 receptor: possiblelink with the c-myc induction pathway. Immunity,2, 89±100.
Otwinowski, Z. (1993). Oscillation data reduction pro-gram. In Proceedings of the CCP4 Study Weekend:Data Collection and Processing (Sawyers, L., Isaacs,N. & Bailey, S., eds), pp. 56±62, , Warrington,SERC Daresbury Laboratory.
Poole, A., Gibbins, J. M., Turner, M., van Vugt, M. J.,van de Winkel, J. G. J., Saito, T., Tybulewicz, V. L. J.& Watson, S. P. (1997). The Fc receptor g-chain andthe tyrosine kinase Syk are essential for activationof mouse platelets by collagen. EMBO J. 16, 2333±2341.
Ramachandran, G. N. & Sasisekharan, V. (1968). Confor-mations of polypeptides and proteins. Advan. Pro-tein Chem. 23, 283±437.
Reth, M. (1989). Antigen receptor tail clue. Nature, 338,383±385.
Reth, M. & Wienands, J. (1997). Initiation and processingof signals from the B cell antigen receptor. Annu.Rev. Immunol. 15, 453±479.
Rice, L. M. & BruÈ nger, A. T. (1994). Torsion angledynamics: reduced variable conformationalsampling enhances crystallographic structure re®ne-ment. Proteins: Struct. Funct. Genet. 19, 277±290.
Rivera, V. M. & Brugge, J. S. (1995). Clustering of Syk issuf®cient to induce tyrosine phosphorylation andrelease of allergic mediators from rat basophilic leu-kemia cells. Mol. Cell. Biol. 15, 1582±1590.
Rowley, R. B., Burkhardt, A. L., Chao, H.-G., Matsueda,G. R. & Bolen, J. B. (1995). Syk protein tyrosinekinase is required by tyrosine-phosphorylated Iga/Igb immunoreceptor tyrosine activation motif bind-ing and autophosphorylation. J. Biol. Chem. 270,11590±11594.
Shiue, L., Green, J., Green, O. M., Karas, J. L.,Morgenstern, J. P., Ram, M. K., Taylor, M. K.,Zoller, M. J., Zydowsky, L. D., Bolen, J. B. &Brugge, J. S. (1995). Interaction of p72syk with the gand b subunits of the high-af®nity receptor forimmunoglobulin E, FceRI. Mol. Cell. Biol. 15, 272±281.
Songyang, Z., Shoelson, S. E., Chaudhuri, M., Gish, G.,Pawson, T., Haser, W. G., King, F., Roberts, T.,Ratnofski, S., Lechleider, R. J., Neel, B. G., Birge,R. B., Fajardo, J. E., Chou, M. M., Hanafusa, H.,Schaffhausen, B. & Cantley, L. C. (1993). SH2domains recognize speci®c phosphopeptidesequences. Cell, 72, 767±778.
Taniguchi, T., Kitagawa, H., Yasue, S., Yanagi, S., Sakai,K., Asahi, M., Ohta, S., Takeuchi, F., Nakamura, S.-I. & Yamamura, H. (1993). Protein tyrosine kinasep72syk is activated by thrombin and is negatively

Crystal Structure of the ITAM-bound SH2 Domains of Syk 537
regulated through Ca2� mobilization in platelets.J. Biol. Chem. 268, 2277±2279.
Terwilliger, T. C., Kim, S.-H. & Eisenberg, D. (1987).Generalized method of determining heavy-atompositions using the difference Patterson function.Acta Crystallog. sect. A, 43, 1±5.
Waksman, G., Kominos, D., Robertson, S. C., Pant, N.,Baltimore, D., Birge, R. B., Cowburn, D., Hanafusa,H., Mayer, B. J., Overduing, M., Resh, M. D., Rios,C. B., Silverman, L. & Kuriyan, J. (1992). Nature,358, 646±653.
Waksman, G., Shoelson, S. E., Pant, N., Cowburn, D. &Kuriyan, J. (1993). Binding of a high af®nity phos-photyrosyl peptide to the src SH2 domain: crystal
structures of the complexed and peptide-free forms.Cell, 72, 779±790.
Wan, Y., Kurosaki, T. & Huang, X.-Y. (1996). Tyrosinekinases in activation of the MAP kinase cascade byG-protein-coupled receptors. Nature, 380, 541±544.
Wange, R. L. & Samelson, L. E. (1996). Complex com-plexes: signaling at the TCR. Immunity, 5, 197±205.
Zhang, J., Berenstein, E. H., Evans, R. L. & Siraganian,R. P. (1996). Transfection of Syk protein tyrosinekinase reconstitutes high af®nity IgE receptor-mediated degranulation in a Syk-negative variant ofrat basophilic leukemia RBL-2H3 cells. J. Exp. Med.184, 71±79.
Edited by D. Rees
(Received 6 April 1998; received in revised form 22 May 1998; accepted 22 May 1998)
Related Documents











